
Environmental distribution of post-Palaeozoic crinoids from the Iberian and south-Pyrenean basins, NE Spain
SAMUEL ZAMORA, MARCOS AURELL, MARGARET VEITCH, JAMES SAULSBURY, MIKEL A. LÓPEZ-HORGUE, FERNANDO A. FERRATGES, JOSÉ ANTONIO ARZ, and TOMASZ K. BAUMILLER
Zamora, S., Aurell, M., Veitch, M., Saulsbury, J., López-Horgue, M.A., Ferratges F.A., Arz, J.A., and Baumiller, T.K. 2018. Environmental distribution of post-Palaeozoic crinoids from the Iberian and south-Pyrenean basins, NE Spain. Acta Palaeontologica Polonica 63 (4): 779–794.
Post-Palaeozoic crinoids from northeast Spain ranging from the Ladinian (Middle Triassic) to the Ilerdian (lower Ypresian, early Eocene) are documented. Here we provide the first attempt to reconstruct the environmental distribution of these crinoids based on relatively complete material (mostly cups). Triassic forms are dominated by encrinids from outer carbonate ramps. Late Jurassic crinoids are dominated by cyrtocrinids, comatulids, millericrinids, and isocrinids, occurring either on sponge mounds and meadows or on soft substrates within middle to outer carbonate ramps. Aptian (Early Cretaceous) forms include nearly complete isocrinids which are found in extremely shallow environments represented by bioclastic carbonates and interspersed oyster-rich layers. Other Aptian occurrences come from more distal and deep environments and are composed solely of comatulids. Albian forms are dominated by cyrtocrinids and isocrinids associated with coral reefs. Late Cretaceous and Eocene crinoids include mostly bourgueticrinids (Comatulida) that are found either in outer ramp facies or associated with mid-ramp reef complexes. The later corresponds to one of the shallowest occurrence of bourgueticrinids in the Cenozoic. The palaeoecological data for fossil crinoids of northeast Spain contributes to reconstructing the history of the bathymetric distribution of articulate crinoids, supporting the idea that stalked crinoids were able to inhabit a wide range of shallow marine environments in the late Mesozoic and early Cenozoic.
Key words: Crinoidea, Comatulida, Cyrtocrinida, Isocrinida, Millericrinida, Encrinida, Mesozoic, Cenozoic, Iberia.
Samuel Zamora [s.zamora@igme.es], Instituto Geológico y Minero de España, C/Manuel Lasala, 44, 9B, Zaragoza 50006, Spain.
Marcos Aurell [maurell@unizar.es] and Fernando A. Ferratges [nautilo_fer_1987@hotmail.com], Área de Estratigrafía, Departamento de Ciencias de la Tierra, Universidad de Zaragoza, E-50009 Zaragoza, Spain.
Margaret Veitch [maveitch@umich.edu], James Saulsbury [jgsauls@umich.edu], and Tomasz K. Baumiller [tomaszb@umich.edu], Museum of Paleontology and Department of Earth and Environmental Sciences, University of Michigan, Ann Arbor, MI 48109, USA.
Mikel A. López-Horgue [mikel.lopezhorgue@ehu.eus], Estratigrafia eta Paleontologia Saila, Faculty of Science and Technology, University of the Basque Country UPV/EHU,48940 Leioa (Biscay), Basque Country, Spain.
José Antonio Arz [josearz@unizar.es], Departamento de Ciencias de la Tierra, and Instituto Universitario de Investigación en Ciencias Ambientales de Aragón, Universidad de Zaragoza, E-50009 Zaragoza, Spain.
Received 29 June 2018, accepted 21 August 2018, available online 17 September 2018.
Copyright © 2018 S. Zamora et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Crinoids represent an important component of modern ecosystems and have a rich fossil record that extends back to the Early Ordovician (Guensburg and Sprinkle 2001). Among the diverse and vastly disparate crinoids of the Palaeozoic, only the Articulata, likely derived from within the Eucladida (Wright et al. 2017), survived the P–T extinction and rapidly diversified to occupy a wide range of environments (Simms 1988; Foote 1999; Twitchett and Oji 2005; Hess and Messing 2011).
The recent Treatise on Invertebrate Paleontology (Hess and Messing 2011) recognizes four orders of crinoids with extant members: the Isocrinida, Hyocrinida, Comatulida, and Cyrtocrinida. While the relationships of these groups are still debated (e.g., Simms 1988; Simms and Sevastopulo 1993; Hemery 2011; Hemery et al. 2013; Rouse et al. 2013; Roux et al. 2013; Cohen and Pisera 2017), there is enough evidence that most groups have roots in the Mesozoic; thus understanding the phylogeny of the Articulata requires investigation of fossil species from the Mesozoic and Cenozoic. Moreover, the fossil record of articulate crinoids suggests a complex history of both the diversity of various groups as well as their ecology (Gorzelak et al. 2016). For example, cyrtocrinids are represented by only four extant genera, all living in relatively deep water (Donovan and Pawson 2008: table 1), but their Mesozoic record is quite diverse, consisting of over 40 genera found in a wide range of environments from deep water sponge meadows (Charbonnier et al. 2007; Zatoń et al. 2008; Hess et al. 2011) to shallower environments (Salamon and Gorzelak 2007; Salamon 2008; Hess and Gale 2010). The Mesozoic expansion of potential predators of crinoids, including echinoids and teleost fishes, is thought to have restricted all modern stalked crinoids to deep water; only the stalkless featherstars have greatly increased their diversity and remained in shallow waters (Meyer and Macurda 1977; Oji 1996; Baumiller et al. 2010; Gorzelak et al. 2012; Stevenson et al. 2017). The bathymetric history of stalked crinoids appears to be quite complex with some studies suggesting their restriction to deeper water by the end of the Mesozoic (e.g., Bottjer and Jablonski 1988; but see exceptions in Salamon et al. 2009; Salamon and Gorzelak 2010), whereas newer studies indicate that they remained in shallow water well into the Cenozoic, at least in the southern hemisphere (Whittle et al. 2018).
Despite the limitations of the fossil record, it is the only direct source of information about crinoid ecology, including facies distribution and relationships with other organisms, therefore any reasonably complete account of their ecology requires a thorough treatment of their fossil history. Whereas post-Palaeozoic crinoids are common components of marine rocks from Spain and have been cited in several classic stratigraphic works (i.e., Fezer 1988; Ramajo and Aurell 2008), complete specimens or cups revealing diagnostic features are rare (e.g., Bataller 1930; Roux and Plaziat 1978; Mora-Morote 2009), with information on the distribution and palaeoecology of post-Palaeozoic crinoids from Spain sparse and scattered. In an attempt to supplement the ecological history of post-Palaeozoic crinoids, we summarize previously published information and describe morphology and palaeoecology of newly discovered Spanish crinoids. Five years of intermittent investigations of Mesozoic and Cenozoic rocks in north-east Spain have resulted in a substantial collection of fossil crinoids from numerous localities (Fig. 1), ranging from the Triassic to the Eocene, and including representatives of two extinct orders (Millericrinida, Encrinida) and three of the four extant orders (Isocrinida, Comatulida, and Cyrtocrinida). Here we summarize the occurrences of post-Palaeozoic crinoids from this region, characterize the main stratigraphic and sedimentological features of the sedimentary units that have yielded complete identifiable crinoids, and discuss the implications of our findings for crinoid palaeoecology.
Institutional abbreviations.—MGSB, Museo Geológico del Seminario de Barcelona, Barcelona, Spain; MPZ, Museo de Ciencias Naturales de la Universidad de Zaragoza, Zaragoza, Spain.
Historical background
Complete crinoids or even their calyces from the Mesozoic and Cenozoic rocks of Spain are rare and only a few papers have reported such specimens. This contrasts with the much richer record of isolated columnals that have been reported in several publications (e.g., Rasmussen 1961, or the most recently Astibia et al. 2016).
Comatulids were among the first crinoids described from Spain. Loriol (1900) described a single specimen of Aptian age consisting of centrodorsal and a radial circlet that he referred to as Antedon almerai Loriol, 1900 from the “Environs de Barcelone”. Another Aptian comatulid, Actinometra batalleri Astre, 1925 was established by Astre (1925) based on several centrodorsals from Benasal (Castellón). Bataller (1930) reported additional complete cups of comatulids from the Mesozoic of Spain and a few years later (Bataller 1941) named two additional species from the Aptian of Barcelona, “Solanocrinus” astrei, based on a single specimen of a centrodorsal, and “S.” estruchi, based on a centrodorsal with an attached radial circlet; both are currently included in the genus Decameros. A brachial plate, likely belonging to Decameros, was also figured by Bataller (1930: figs. 10–12). Decameros is one of the most common crinoids in the Lower Cretaceous, and Casanova (1974) described a new locality in Castellón (East Spain) that yielded several centrodorsals of Decameros astrei. A more recent study (Mora-Morote 2009) focused on a large collection of comatulids from the Aptian of Alicante (Betic Zone) in southeast Spain that included the species Solanocrinus (Actinometra) batalleri, S. (Antedon) almerai, and S. estruchi.
Rasmussen (1961) reviewed Spanish Solanocrinites and also mentioned the presence of Phyllocrinus malbosianus in the Spanish Cretaceous. SZ has seen material likely belonging to the latter species from Cehegin (Murcia), in southeast Spain (personal communication Francisco José Armijo-Fernández 2017).
Roux and Plaziat (1978) studied crinoids collected from the Cenozoic of the Pyrenees of both France and Spain, with a special focus on bourgueticrinids. This work includes descriptions of Conocrinus doncieuxi from the Ilerdian (lower Eocene) of the Isabena valley, Aren (Huesca) and San Roman (Santander), Conocrinus aff. doncieuxi from Can Monrebey (Lleida), as well as a new species, Conocrinus romanensis, based on material from the Ilerdian–Lutetian (lower–middle Eocene) of San Roman (Santander) and Sierra del Cadi (Lleida). It is interesting to note that the associated fauna from the Tremp-Isabena Basin led Roux and Plaziat (1978) to infer that Conocrinus doncieuxi lived in a siliciclastic environment in relatively deep water (almost 150 m). Hagdorn (2011) in his extensive summary of Triassic crinoids cited cirrals and columnals of Holocrinus cisnerosi Schmidt, 1935 from the Ladinian and a new species of Encrinus. The former corresponds to material collected in southeastern Spain and mentioned in López-Gómez et al. (1994), and the latter to Encrinus sp. from Collbató (see below) (Hans Hagdorn personal communication 2018).
Fezer (1988) cited Balanocrinus sp. and Apiocrinus sp. in the Upper Jurassic oolitic marl of Griegos (Teruel) but did not describe or figure any specimens. More recently, Astibia et al. (2016) figured some stem fragments of the isocrinid Isselicrinus sp. from the Bartonian (Eocene) Pamplona Marls Formation. According to Astibia et al. (2005), they represent some of the deepest crinoid occurrences found in Spain, coming from outer shelf and slope environments.
Material and methods
Most of the specimens described in this study were surface collected directly from the outcrops as macrofossils but a few kilograms of sediment were bulk collected from Jurassic of Aguilón and Tosos, Cretaceous of Fuentes de Rubielos and Irañeta, and Eocene of Puebla de Roda. The latter were placed in buckets containing water and hydrogen peroxide (H2O2) to dissolve the rock and after two days of soaking, the material was washed for ca. 16 hours until clean. Crinoid remains larger than 1 mm were separated for analyses, whereas the finer fraction waits further processing. From the 1 mm and larger fraction, only specimens represented by at least partially articulated calyxes were examined in detail; all other material was set aside for future research.
The Lower Cretaceous Oliete locality (see below) is a classic Echinoderm-Lagerstätte, where specimens appear well articulated on the surface of large slabs. In this instance, large slabs were excavated and cleaned with water in the laboratory. Most specimens were prepared using potassium hydroxide (KOH), and later neutralized with hydrochloric acid (10%).
Specimen were photographed using a Nikon D7100 equipped with AF-S Micro NIKKOR 60 mm objective after coating with ammonium chloride to increase contrast.
Ages of samples from different localities were estimated based on key fossil indicators, either collected by us from the new localities or reported in the literature. In dating the Triassic samples, we relied on conodonts and foraminifera (Calvet and Marzo 1994). Jurassic and Lower Cretaceous samples were dated using ammonoid biozonation schemes (Meléndez and Fontana 1993; López-Horgue et al. 1999; Moreno-Bedmar et al. 2010; Bover-Arnal et al. 2016). To date Upper Cretaceous samples we relied on planktic foraminifera, and Eocene samples on macroforaminifera (Serra-Kiel et al. 1994).
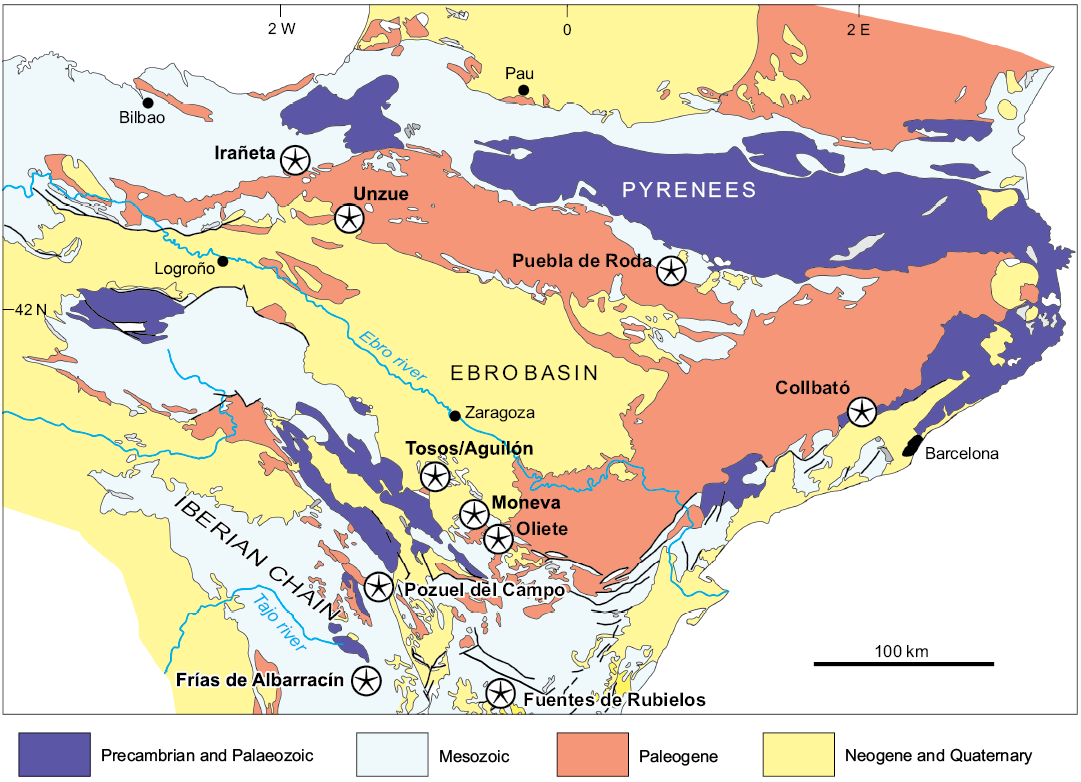
Fig. 1. Map of northeast Spain showing post-Palaeeozoic crinoid localities (stars) discussed in the text.
Geological setting
Field work in northeast Spain and a review of previous collections resulted in the discovery of new localities some of which produced articulated crinoids and/or diagnostic cups (Fig. 1). Here we provide description of the localities with special emphasis on associated fauna and their age.
Collbató unit (Triassic).—An assemblage of articulated echinoderms was found in an abandoned quarry, La Pedrera d’en Rogent, close to the village of Collbató (Barcelona Province). These sedimentary rocks are part of the Catalan Coastal Ranges and are situated within the Gaià-Montseny Domain (sensu Calvet and Marzo 1994; Fig. 2). The strata were included within the M3 unit of the Muschelkalk facies by Smith and Gallemí (1991). Specimens were collected from the Collbató unit which is a lateral equivalent of La Riba reefs unit, and is assigned to the Ladinian (Rosell et al. 1975; Calvet et al. 1987; Calvet and Marzo 1994). The sedimentary rocks at La Pedrera d’en Rogent consist of an interval 16–20 m thick of micritic, finely laminated, almost lithographic limestone with marly intercalations (Smith and Gallemí 1991). This locality contains articulated ophiuroids, holothurians and a single crinoid described here. Associated fauna includes abundant mollusks and isolated vertebrate remains (Calzada and Gutiérrez 1987; Smith and Gallemí 1991; Fortuny et al. 2011). Smith and Gallemí (1991) assigned the single crinoid they reported to the genus Encrinus, and recognized its similarities to E. aculeatus from the Lower Muschelkalk of Germany and Poland. Hagdorn (2011) referred to the Spanish crinoid material as representing a new species of Encrinus pending formal description.
There are no studies that provide a well-constrained age for the Collbató unit. Nevertheless, the lateral equivalents, La Riba reefs and La Rasquera units, have been dated by means of foraminifera (Márquez and Trifonova in Calvet and Marzo 1994). They fall within the Turriglomina mesotriasica Zone, suggesting a late Ladinian age. March, in Calvet and Marzo (1994), analyzed the conodonts of the laterally equivalent Rasquera unit; the presence of Pseudofurnishius murcianus and Sephardiella mungoensis suggests a Longobardian (late Ladinian) age.
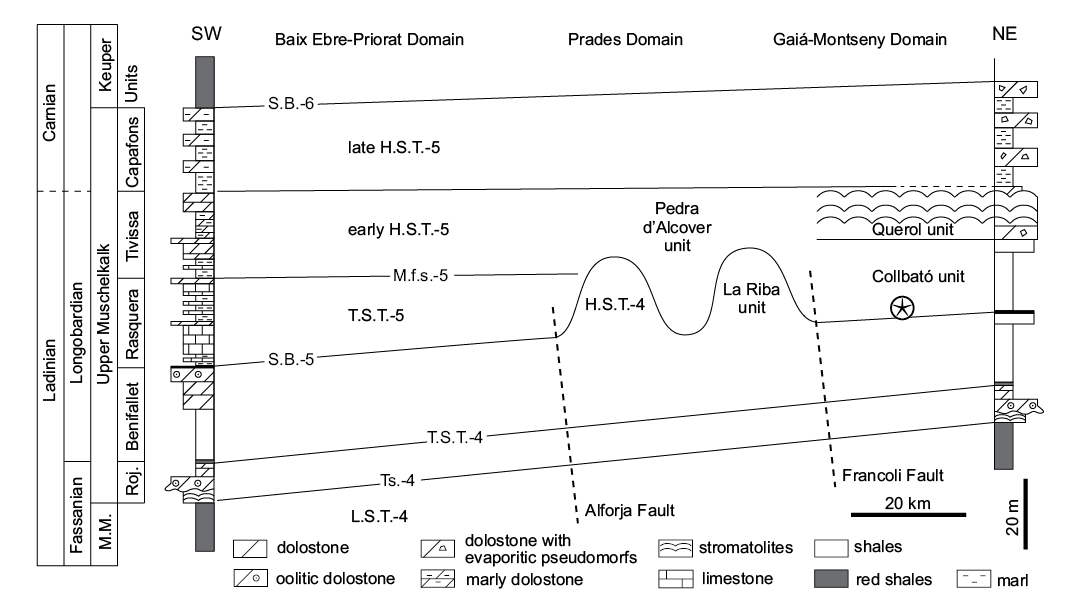
Fig. 2. Stratigraphic framework of the Upper Muschelkalk of the Catalan Ranges showing the crinoid-bearing Collbató unit (after Calbet and Marzo 1994). The horizon containing crinoids (star) coincides with the maximum flooding surface and is laterally equivalent to La Riba reef complex. Abbreviations: M.M., Middle Muschelkalk; Roj., Rojals; H.S.T., Highstand system track; L.S.T., Lowstand system track; M.f.s., Maximun flooding surface; S.B., Sequence boundary; T.S., Transgressive surface; T.S.T., Transgressive system track.
Yátova Formation (Upper Jurassic).—In the surroundings of Aguilón, Tosos and Moneva (Zaragoza, Teruel, Iberian Range), a sponge-limestone dominated unit, the Yátova Formation (middle–upper Oxfordian), contains abundant echinoderm remains, including a diverse crinoid fauna. Here, the Yátova Formation (Fig. 3) consists of tabular to nodular limestone with marly interbeds, locally forming decimeter to meter-thick, upward-thickening sequences (Ramajo and Aurell 2008). The main components are siliceous sponges (Dictyida, Lychniskida, and Lithistida, in descending order of abundance; e.g., Deusch et al. 1990) of variable morphologies (dish, cup, and tubular), typically broken and found in graded beds. They are associated with a microbial crust and encrusting organisms such as annelids (serpulids, Terebella), bryozoans, benthic foraminifera (nubecullarids, Bullopora) and Tubiphytes. Also common are tuberoids and encrusted fragments of sponges. Locally, ammonites, belemnites, bivalves, brachiopods, echinoids, asterozoans, foraminifera, ostracodes, ahermatypic corals, and the aforementioned crinoids can be common.
The maximum age range of the Yátova Formation is middle Oxfordian (i.e., lower Gregoryceras transversarium Biozone) to upper Oxfordian (i.e., lower Subnebrodites planula Biozone, Subnebrodites planula Subzone).
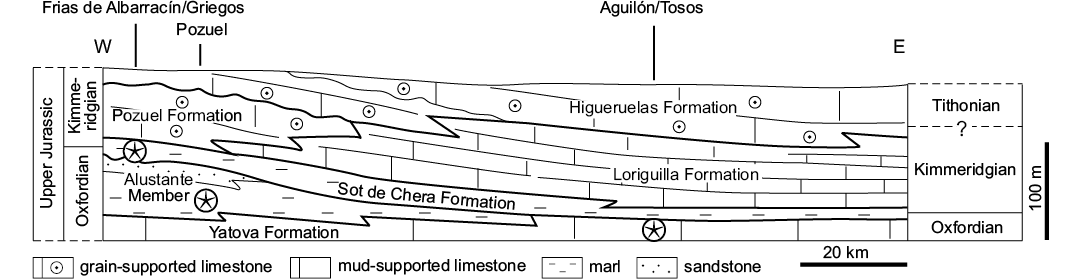
Fig. 3. Stratigraphy of Upper Jurassic crinoid localities (on the top) from the Iberian Ranges. Stars indicate crinoid-bearing formations.
Sot de Chera Formation (Upper Jurassic).—The uppermost Oxfordian outcrops located around Pozuel del Campo, Griegos and Frías de Albarracín (Teruel, Iberian Range), consist of thick marly dominated successions, including decimeter-thick siliciclastic and skeletal-rich limestone intercalations, called Sot de Chera Formation. The Pozuel del Campo locality corresponds to the Alustante Member and contains abundant ammonites of Subnebrodites planula Subzone (Subnebrodites planula Zone). The Griegos and Frías de Albarracín localities are found higher up within the Sot de Chera Formation, in a skeletal-rich horizon which includes ammonites of Sutneria galar Subzone (Subnebrodites planula Zone). This horizon also contains resedimented ooids from more proximal and shallow carbonate shoal environments (Pomar et al. 2015). The studied bioclastic and marly intervals contain abundant echinoderm remains, including a diverse crinoid fauna. Here, the Sot de Chera Formation (Fig. 3) consists of a marly progradational wedge-shaped deposits with siliciclastics derived from the emergent areas located to the west, including abundant graded skeletal accumulations interpreted as tempestites (Aurell et al. 2003, 2010). The main skeletal components found in these mid-ramp deposits are bivalves, echinoderms, gastropods, brachiopods, siliceous sponges, serpulids, benthic foraminifera, ahermatypic colonial and solitary corals, belemnites, and ammonites.
Forcall Formation (Lower Cretaceous).—Specimens were collected in the Forcall Formation of the Penyagolosa Sub-basin (Fig. 4), from outcrops on the south of the road connecting the villages of Fuentes de Rubielos (Teruel Province) with Cortes de Arenoso. This section of the Forcall Formation contains mostly marl and marly limestone and preserves a small lens (less than three meters in length) that was excavated by amateurs, and contains well-preserved specimens of the crinoid Decameros and associated fauna, including echinoids, bivalves, and ammonites.
The Forcall Formation in the Penyagolosa Sub-basin has been dated by means of ammonites (Moreno-Bedmar et al. 2010). The horizon with crinoids contains the ammonites Deshayesites cf. multicostatus, which indicates the upper part of the Deshayesites deshayesi Biozone, and is of lower Aptian age.
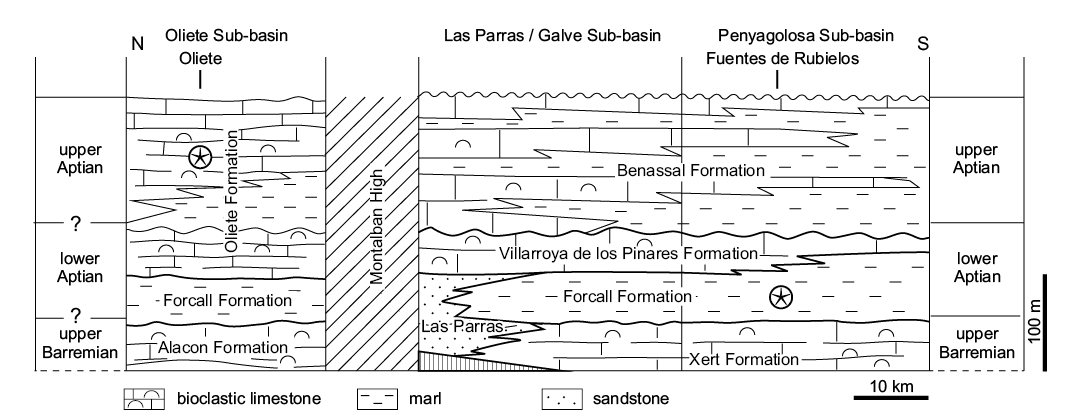
Fig. 4. Stratigraphy of Lower Cretaceous crinoid localities (on the top) from the Iberian Ranges. Stars indicate crinoid-bearing formations.
Oliete Formation (Lower Cretaceous).—The exposures found north of the Oliete village (Teruel, Oliete Sub-basin) include a complete Lower Cretaceous (i.e., upper Barremian–Aptian) shallow marine succession of the Alacón, Forcall, and Oliete formations (Fig. 4), with fossiliferous beds containing crinoids in the middle part of the Oliete Formation. The horizons containing crinoids are tabular bioclastic limestone with marly interbeds. Surfaces of the limestone units contain abundant and well-preserved echinoderm assemblages that could be described as Echinoderm-Lagerstätte. They include nearly complete isocrinids, echinoids with spines attached, and goniasterid asteroids, in addition to other faunal components, such as molluscan fragments and fish teeth.
The age of the Forcall and Oliete formations in the area has been constrained by means of ammonite biostratigraphy. The Barremian/Aptian boundary was recently placed within the lower part of the Forcall Formation in the Oliete Sub-basin (Bover-Arnal et al. 2016; Aurell et al. 2018). There are two significant Aptian transgressive events indicated by marl-rich intervals bearing scarce ammonites. The lower transgressive event is recorded in the marls of the Forcall Formation, which contains ammonites of the lower Aptian Deshayesites forbesi Zone (Moreno-Bedmar et al. 2010). The second transgressive unit is represented by a marl-dominated interval located in the middle part of the Oliete Formation (the Dehesa Marls Member; Canérot et al. 1982), which contains scarce ammonites from the upper Aptian Parahoplites melchioris and Acanthohoplites nolani zones (García et al. 2014).
Albeniz unit (Lower Cretaceous).—Limestone to the north of the Irañeta locality crops out on the southern flank of the Aralar mountains folded area, which is part of the inverted Basque-Cantabrian Basin (Fig. 1). The limestone represents the part of shallow marine carbonate platforms of Aptian–Albian age that form the maximum relief of the Aralar mountains. During the late Aptian and Albian, the siliciclastic input increased and carbonate platforms recorded a transition from ramps to rimmed platforms. This change is also influenced by differential subsidence associated with a basinwide tectonism (e.g., García-Mondéjar et al. 1996). Carbonate platform sedimentation developed on shallow marine highs bounded by active faults, especially during the late Albian. Consequently, downthrown blocks formed deeper marine troughs with siliciclastic sedimentation dominant.
Figure 5 shows the main stages of carbonate platform development in the early Albian to late Albian transition recorded to the north of the Irañeta locality in the Aitzondo peak area (e.g., López-Horgue et al. 1997). Stage 4 is characterized by a drastic reduction in carbonate sedimentation relative to the previous stages and the development of a pinnacle reef geometry (Lertxundi and García-Mondéjar 1998) facing deeper troughs with calcareous mudstone and bioclastic calcarenite and breccia. The age of stage 4 containing crinoid material is early late Albian, based on stratigraphical correlation with coeval successions to the west (López-Horgue et al. 1996, 1997) and the occurrence of ammonites of the Hysteroceras varicosum Zone age (sensu López-Horgue et al. 1999) in the succession above the Aitzondo pinnacle reef (Wiedmann 1962). Both carbonates and siliciclastics of stage 4 could be included in the Albeniz unit (López-Horgue et al. 1996 erected the Albeniz unit to group time-equivalent carbonates and their coeval siliciclastic sediments in the Alsasua area to the west).
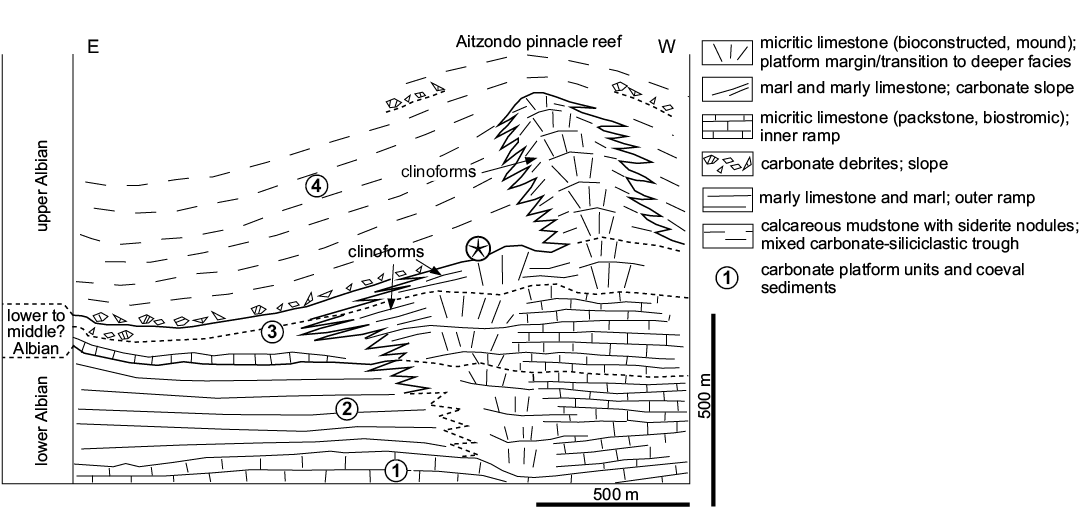
Fig. 5. Albian carbonate platform of Aralar with crinoid-bearing horizon (star) at the base of unit 4 (based on López-Horgue et al. 1997; Lertxundi and García-Mondéjar 1998; and new data from this paper).
Unnamed Formation in Alaiz (Upper Cretaceous).—The locality of Unzue is situated in the western part of the Jaca-Pamplona Basin, within the Sierra de Alaiz. The studied section is mostly composed of marl and contains abundant crinoid material in the lower horizons. Specimens of crinoids appear associated with echinoid, asterozoan, mollusk, and fish remains.
In order to constrain the age of the studied samples, a micropaleontological analyses was conducted on the marly sample (F0) collected in the Unzué section and corresponding to the horizon yielding crinoids. The preservation of the planktic foraminifera is poor, with the planktic/benthic ratio of 48% indicating middle to outer shelf environment (Murray 1991). Planktic foraminifera are scarce and the assemblage is composed of Archaeoglobigerina cretacea, Contusotruncana fornicata, Globigerinelloides praeriehillensis, Globigerinelloides volutus, Globigerinelloides yaucoensis, Globotruncana aegyptiaca, Globotruncana arca, Globotruncana bulloides, Globotruncana linneiana, Hedbergella holmdelensis, Hedbergella monmouthensis, Heterohelix globulosa, Heterohelix labellosa, Heterohelix pulchra, Pseudoguembelina costulata, Pseudotextularia nuttalli, Gublerina cuvillieri, and Rugoglobigerina rugosa. The estimated age of this assemblage is between late Campanian and early Maastrichtian. Index-species for the Campanian–Maastrichtian boundary (as Rugoglobigerina scotti or Contusotruncana contusa; see Odin 2001) are absent in this sample.
Serraduy Formation (Eocene).—The Graus-Tremp Basin in the south Pyrenees records marine sediments from the Thanetian–lower Cuisian (lower Ypresian, lowermost Cenozoic). In the northern part of Puebla de Roda (Huesca), reef complexes and associated facies record a rich benthic fauna including crinoids, mostly bourgueticrinids. They occur in the Serraduy Formation (Fig. 6), which has been divided into three members by Cuevas-Gozalo et al. (1985): a basal Alveolina Limestone, an intermediate Reef Limestone, and an upper Riguala Member (sensu Tosquella 1988). Crinoids and an associated fauna composed of benthic and planktic foraminifera, bivalves, gastropods, nautiloids, terebratulid brachiopods, corals, echinoderms (regular and irregular echinoids and asteroids), decapod crustaceans, bryozoans, serpulids, and fishes (selaceans and teleosts) occur in the Riguala Member. The lithology consists of marl and subsidiary marly limestone associated with the reef complexes composed of coral-algal domes.
Serra-Kiel et al. (1994) used various approaches, including biostratigraphic indicators (micro and macroforaminifera, nanoplankton) and magnetostratigraphy to estimate the age of the Serraduy Formation. They concluded that these horizons correspond to the middle Ilerdian (lower Ypresian, lower Eocene).
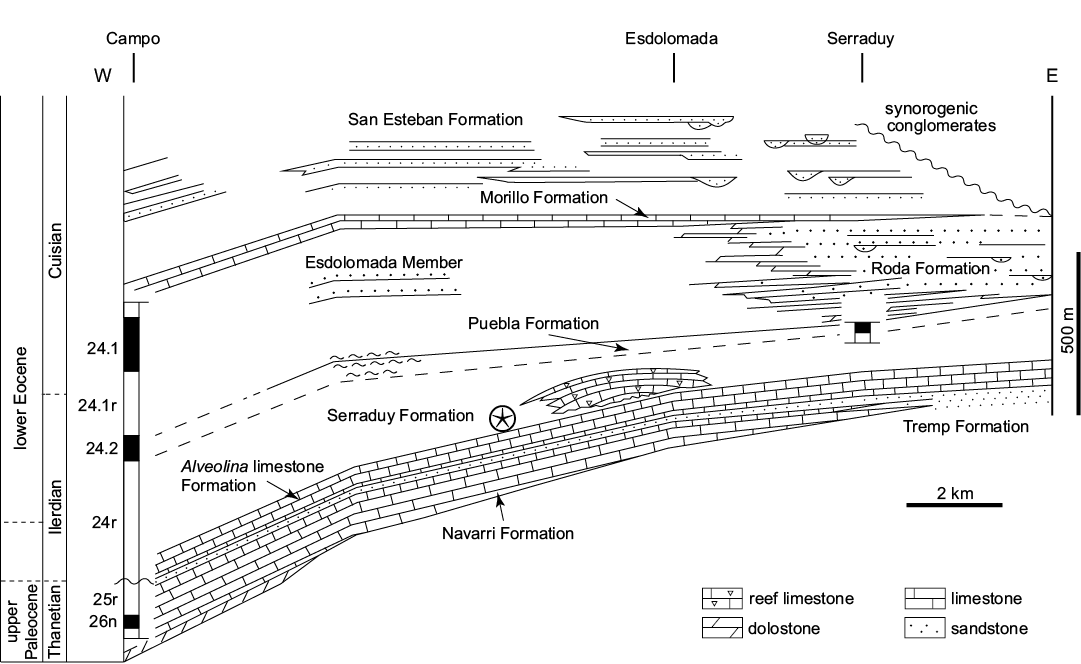
Fig. 6. Eocene stratigraphic units of the Graus-Tremp basin (Central Pyrenees) with crinoid-bearing horizon within the Serraduy Formation indicated (after Serra-Kiel et al. 1994). 24–26 correspond to magnetozones; n, normal; r, reverse.
Results
Crinoids found in Spanish post-Palaeozoic rocks are members of the Encrinida (Fig. 7), Millericrinida (Fig. 8), Cyrtocrinida (Figs. 9, 10), Isocrinida (Fig. 10), and Comatulida (including Comatulidina, Figs. 8, 10 and Bourgueticrinina, Fig. 10) (sensu Hess et al. 2011).
Encrinids are rare and articulated specimens, including Encrinus sp. (Fig 7A), have only been recovered from a single Ladinian locality of Collbató (Barcelona). At that locality, encrinids are also represented by abundant columnals (Fig. 7B), and the associated fauna includes well-preserved ophiuroids and holothurians.
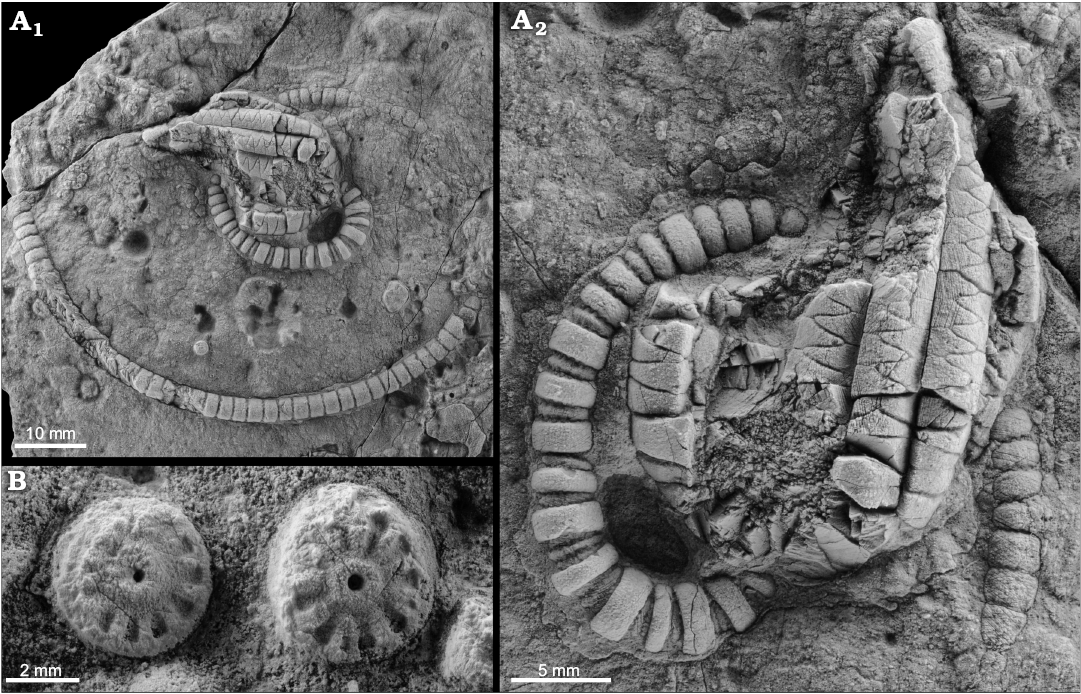
Fig. 7. Crinoid Encrinus sp. from Triassic of NE Spain, the Collbató unit (Ladinian). A. MGSB/45007, well-preserved specimens showing a coiled stem (A1), detail of the crown and proximal stem (A2). B. MGSB/45006, isolated columnals.
Millericrinids have been found only in Upper Jurassic rocks (Yátova and Sot de Chera formations). The Yátova Formation contains two millericrinids, Millericrinida indet. A and an undescribed taxon (Fig. 8C), whereas the Sot de Chera Formation is rich in crinoids, especially millericrinids and isocrinids. Sot de Chera millericrinids are represented by abundant material including large fragments of stems, roots and calyx material. Three taxa have been identified based on cups including Millericrinida indet. B (Fig. 8D), Pomatocrinus cf. mespiliformis (Fig. 8E) and one species of Apiocrinites (Fig. 8A).

Fig. 8. Millericrinids (A–E) and comatulid (F) from the Upper Jurassic of NE Spain, Frías de Albarracín (A, B, D), Pozuel del Campo (E), and Griegos (F), latest Oxfordian, Sot de Chera Formation; Aguilón, early late Oxfordian, Yatova Formation (C). A. Apiocrinites sp., MPZ-2018/470 in oral (A1), aboral (A2), and lateral (A3) views. Note multiple epibionts on the lateral and oral sides suggesting a complex processes of disarticulation and exposure prior to final burial. B. Millericrinida indet., MPZ-2018/471, distal root. C. Millericrinida indet. A, MPZ-2018/472 in oral (C1), aboral (C2), and lateral (C3) views. D. Millericrinida indet. B, MPZ-2018/473 in oral (A1), aboral (A2), and lateral (A3) views. E. Pomatocrinus cf. mespiliformis, MPZ-2018/474 cup lacking radial plates in aboral (E1) and lateral (E2) views. F. Solanocrinites sp., MPZ-2018/475 single centrodorsal in oral (F1), aboral (F2), and lateral (F3) views.
Cyrtocrinids are by far the most diverse, especially in the Yátova Formation. Our preliminary analysis suggests that this rich assemblage includes the genera Eugeniacrinites (Fig. 9A), Pilocrinus (Fig. 9B), Gammarocrinites (Fig. 9C, D), and Tetracrinidae indet. (Fig. 9E), all represented by well-preserved specimens of near complete calyxes. The locality of Irañeta, which is Albian in age, contains a less diverse assemblage of indeterminate isocrinids and a new cyrtocrinid. The remains of the cyrtocrinid Proholopus are abundant and consist of cups, stem fragments and brachial plates (Fig. 10D).
Fragmentary isocrinid remains are common in the Jurassic but articulated specimens have been found only in the Lower Cretaceous Oliete Formation in Oliete (Teruel), where numerous Isocrinus cf. nicoleti occur on surfaces of slabs (Fig. 10A, B).
Comatulids (excluding bourgueticrinids) are generally very rare except for the genus Decameros (Fig. 10C), which is relatively abundant at various Aptian localities (Loriol 1900; Astre 1925; Bataller 1930, 1941; Casanova 1974). The new material presented in this study consists of well-preserved specimens assigned to Decameros ricordeanus d’Orbigny, 1850, which include centrodorsals and fragments of arms and cirri. This comatulid is distinguished by the presence of only five arms. Another occurrence of a single cup of Solanocrinites sp. (Fig. 8F) was collected from the Upper Jurassic of Griegos (Teruel), where it is associated with more common millericrinids (see above) and rare isocrinids.
Remains of bourgueticrinids are relatively common in the Upper Cretaceous and Eocene units. Their diversity is likely to be underestimated, as some bourgueticrinid-bearing localities have yet to be thoroughly investigated (Roux and Plaziat 1978). Complete specimens of Democrinus sp. (Fig. 10E, F, I), including complete cups (Fig. 10I), stem fragments (Fig. 10F), roots (Fig. 10E), and isolated brachials have been collected from Campanian–Maastrichtian strata of Unzue. Bourgueticrinids of Ilerdian (lower Eocene) age have also been recovered from the Serraduy Formation in Puebla de Roda, including dense accumulations of Bourgueticrinus sp. (Fig. 10G, H). B. hagenowii (Goldfuss, 1840), B. constrictus (von Hagenow in Quenstedt, 1876), and B. danicus (Brunnich 1913) all have similarities with Bourgueticrinus sp., but only B. danicus is of Cenozoic age, with B. hagenowii, B. constrictus being found no later than the Maastrichtian in other localities (Rasmussen 1961; Kjaer and Thomsen 1999; Jagt 1999; Jagt and Salamon 2007). Nearly half collected specimens of Bourgueticrinus sp. are greater than 6 mm, the max height of B. danicus (Rasmussen 1961); keeping placement for species undecided until further investigation.

Fig. 9. Cyrtocrinids from the Upper Jurassic Yátova Formation of NE Spain. A. Eugeniacrinites sp. from Aguilón (early late Oxfordian), MPZ-2018/476 in oral (A1) and (A2) lateral views. B. Pilocrinus sp. from Moneva (early late Oxfordian), MPZ-2018/477 in oblique-oral (B1), aboral (B2), and lateral (B3) views. C. Gammarocrinites sp. A from Aguilón (early late Oxfordian), MPZ-2018/478 in oral (C1), aboral (C2), and lateral (C3) views. D. Gammarocrinites sp. B from Tosos (early late Oxfordian), MPZ-2018/479 in oral (D1) and lateral (D2) views. E. Tetracrinidae indet. from Aguilón (early late Oxfordian), MPZ-2018/480, basal circlet in oral (E1), aboral (E2), and lateral (E3) views.

Fig. 10. Isocrinids (A, B), comatulids (C, E–I), and cyrtocrinids (D) from the Cretaceous and Eocene of NE Spain. A, B. Isocrinus cf. nicoleti from Oliete (Aptian) Oliete Formation. A. MPZ-2018/481, complete crown. B. MPZ-2018/482, calyx and proximal arms in aboral view. C. Decameros ricordeanus from Rubielos de Mora (Aptian) Forcall Formation, MPZ-2018/483, centrodorsal in oral (C1), aboral (C2), and lateral (C3) views. D. Proholopus sp. from Irañeta (Albian) Albeniz unit, MPZ-2018/484 in lateral (D1) and oral (D2) views. E, F, I, J. Democrinus sp. from Unzue (Campanian). E. MPZ-2018/485, distal root. F. MPZ-2018/486, isolated columnal. I. MPZ-2018/487, calyx in lateral view. J. MPZ-2018/488, calyx in oral view. G, H. Bourgueticrinus sp. from Puebla de Roda (Ilerdian) Serraduy Formation. G. MPZ-2018/489, calyx in lateral view (G1), specimen in oral view (G2). H. MPZ-2018/490, distal stem with cirri.
Palaeoenvironmental distribution
While recent phylogenies of extant crinoids provide an important framework, to fully understand the evolutionary history of articulates requires a thorough assessment of their diverse and rich fossil record. Likewise, their environmental history can be understood more fully if it includes critical information on the distribution of modern as well as fossil taxa. Table 1 includes all studied crinoid assemblages plus information about environmental distribution and facies.
Table 1. List of lithostratigraphic units, age and depositional setting and major crinoid groups recorded in each unit.
|
Lithostratigraphic unit |
Age |
Recorded groups |
Facies |
Depositional setting |
Localities |
|
Collbató unit |
Ladinian (Triassic) |
Encrinida |
laminated carbonates |
outer ramp |
Collbató |
|
Yátova Formation |
early late Oxfordian (Jurassic) |
Millericrinida, Cyrtocrinida, Isocrinida |
sponges mounds and meadows |
outer to middle ramp |
Aguilón, Tosos, Moneva |
|
Sot de Chera Formation |
latest Oxfordian (Jurassic) |
Millericrinida, Isocrinida, Comatulida |
marl with resedimented ooids |
middle to outer ramp |
Frías de Albarracín, Griegos, Pozuel del Campo |
|
Oliete Formation |
Aptian (Cretaceous) |
Isocrinida |
bioclastic carbonates between oyster horizons |
inner ramp |
Oliete |
|
Forcall Formation |
Aptian (Cretaceous) |
Comatulida |
marl |
outer ramp |
Fuentes de Rubielos |
|
Albeniz unit |
Albian (Cretaceous) |
Cyrtocrinida, Isocrinida |
clay with small coral-sponges patches |
reef slope; graben below wave base |
Irañeta |
|
unnamed unit |
late Campanian to early Maastrichtian (Cretaceous) |
Comatulida |
mud |
outer ramp |
Unzue |
|
Serraduy Formation |
Ilerdian (Eocene) |
Comatulida (Bourgueticrinina) |
bioclastic marl |
middle ramp |
Puebla de Roda |
Triassic encrinids appear exquisitely preserved with other echinoderms in sediments of the Collbató unit that were deposited along a carbonate ramp, in a relatively quiet environment below fair weather wave-base (Calvet et al. 1987; Calvet and Tucker 1988). Calvet and Marzo (1994) suggested that the horizon containing the abundant echinoderm fauna represents the maximum flooding surface. Previously described encrinids have been found attached to hard substrates, such as terquemiid shells and stem fragments of other encrinids (Hagdorn 1978; Hagdorn et al. 1996), and the Spanish specimens provide further evidence for this mode of attachment.
Upper Jurassic cyrtocrinids, millericrinids, and isocrinids from the Yátova Formation are associated with sponge meadows. Ramajo and Aurell (2008) considered the spongiolithic facies to have been deposited in relatively shallow and open platform areas; depth of these environments is likely to have fluctuated from near storm wave base, as evidenced by the presence of graded beds interpreted as tempestites, to below storm wave base, as indicated by the interspersed horizons bearing autochthonous or parautochthonous ammonoids and microbial-sponge mounds (Ramajo et al. 1999); but it probably never exceeded a depth of 60 m. The presence of a rich and diverse crinoid fauna suggests abundant nutrients and temporary decreases in sedimentation rate, perhaps due to sea level fluctuations that would have allowed for larval attachment and colonization. There is no further information on attachment of millericrinids and isocrinids from such units, but based on what we know about cyrtocrinids, they probably attached to hard-substrates. Similar crinoid associations have been described from the Oxfordian of the Swiss and Swabian Jura (Hess 1975).
The Sot de Chera Formation in the Albarracin area (Frías de Albarracín, Griegos and Pozuel del Campo) is a rich fossiliferous marly unit with resedimented ooids. Specimens of Apiocrinites and Pomatocrinus were accumulated within the middle to outer ramp and show evidences of high degree of incrustation by epibionts and resedimentation processes that indicate a complex taphonomic history. These taxa probably lived in muddy substrates where they attached by means of robust holdfast to tests of other organisms (Fig. 8B). This agrees with previous reports of millericrinids attached to shell fragments (Ausich and Wilson 2012) or hardgrounds (Palmer and Fürsich 1974).
Aptian isocrinids and comatulids appear in two different environments and formations. The Lower Cretaceous Alacón, Forcall, and Oliete formations represent a mixed carbonate-siliciclastic, shallow marine, coastal succession with a variable thickness of 100–300 m across the Oliete Sub-basin (e.g., Vennin et al. 1993). The off-shore facies of the Forcall Formation contain accumulations of the comatulid Decameros, which likely lived in dense aggregations on muddy substrates within the outer ramp. Notably, Decameros is one of just a few groups of five-armed featherstars (Atopocrinus, Eudiocrinus, Pentametrocrinidae, other Decameridae; Hess and Messing 2011); most living featherstars have 10 arms or more.
The remains of Isocrinus cf. nicoleti are quite common in the Oliete Formation, where it coexisted with other echinoderms, such as goniasterid asterozoans and echinoids. The Oliete Formation represents a depositional sequence with a lower transgressive interval (the Cabezo Negro limestone and Dehesa marl) followed by an upper regressive interval (the Estancos limestone member), with an increasing proportion of siliciclastic-dominated units towards the top of the succession. The horizons rich in crinoids are found in the lower part of the Estancos member. In this shallow marine environment, Isocrinus cf. nicoleti probably lived with a substantial length of the distal part of the stem recumbent, allowing it to attach to the substrate by means of many, long cirri. Isocrinus cf. nicoleti specimens occur in bioclastic limestone representing very shallow environments, between oyster horizons, deposited in the inner ramp.
Albian cyrtocrinids lived in relatively shallow water and on soft substrates, lateral equivalents of carbonate reefs in areas below the storm wave base. Specimens appear with complete cups and articulated stems, but with arms disarticulated, and are associated with a rich fauna that included siliceous sponges and corals. The reef core facies is estimated to be very shallow (less than a few dozen m depth) but siliciclastic material containing crinoids was deposited deeper in marly calcarenite and bioclastic breccia of the toe of slope of the Aitzondo pinnacle reef. Some fossils in this part of the carbonate system (including crinoids) could have been transported from shallower coeval sediments on the reef.
Bourgueticrinids occur in Upper Cretaceous (Campanian–Maastrichtian) and Eocene rocks representing a range of environments from outer ramp siliciclastics in the former to reef complexes formed in the middle ramp in the latter. Democrinus sp. from the Upper Cretaceous have been found with holdfasts in the form of branched roots attached in situ in muddy environments (Fig. 10E), suggesting they lived off-shore in dense colonies. Bourgueticrinus sp. is very common in the Riguala Member of the Serraduy Formation, which was deposited mid-ramp, in open marine conditions (Serra-Kiel et al. 1994). Bourgueticrinus sp. is most commonly found within tempestites associated with fore-reef facies, but is absent from the reef core and rare in the back reef. It probably occupied the soft muddy environments associated with slope facies within the fore-reef, where it attached by its delicate roots that branched from distal columnals (Fig. 10H).
All extant stalked crinoids occur only in deep (> 100 m) water environments (Lane and Webster 1980; Oji 1989) but the Spanish occurrences support the idea that many Mesozoic groups occupied a wide range of relatively shallow marine environments in epicontinental seas (see above). This trend is also observed in some Cenozoic groups that survived in shallow marine environments, such as isocrinids (Eagle 1993; Eagle and Hayward 1993; Meyer and Oji 1993; Stillwell et al. 1994; Baumiller and Gaździcki 1996; Whittle et al. 2018), but also bourgueticrinids that remained associated with coral reefs in the Eocene (this paper).
Conclusions
Mesozoic and Cenozoic crinoids from Northeast Spain (Iberian and South-Pyrenean basins) are reviewed. They include taxa from eight different formations and a wide range of environments, from very shallow marine settings (inner ramp) to relatively deep conditions (outer ramp), but none from bathyal depths where modern stalked crinoids dominate today. Encrinids occur in the Triassic with rare, complete specimens recovered from laminated carbonates. Millericrinids are abundant in the Jurassic, especially in sponge meadows and marly substrates. Cyrtocrinids are extremely abundant and diverse in the Jurassic when they co-occur with sponges; by the Cretaceous they become rarer and are found only in association with coral-sponge patches. Isocrinids are very common in the Lower Jurassic, but complete specimens are known only from the Lower Cretaceous bioclastic carbonates between oyster horizons, representing one of the shallowest environments they occupied in their whole evolutionary history. Comatulids (excluding Bourgueticrinina) first appear in Jurassic rocks, but abundant material has been found only in Lower Cretaceous deposits. In both cases they occurred in muddy environments. Finally, bourgueticrinids are common in the Late Cretaceous, when they are associated with relatively deep (outer ramp) muddy environments; in the Paleogene, they are very abundant in shallow water reef settings. The occurrence and distribution of fossil crinoids in Northern Spain, although probably incomplete, reflect processes associated with major geological cycles that affected the Iberian plate, as well as trends in post-Palaeozoic crinoid evolution. They support the idea that some groups occupied a wide range of environments by the Mesozoic and some stalked taxa persisted in relatively shallow environments in the early Cenozoic.
Acknowledgements
We thank David Meyer (University of Cincinnati, USA) and anonymous reviewer for thorough and constructive comments that helped improve the manuscript. Luis Moliner (ENSAYA Laboratory, Zaragoza, Spain) helped with identification of ammonites from the Upper Jurassic. We offer special thanks to Benjamin Botanz (Navarra, Spain), Pedro Gracia (Navarra, Spain), Luis Lázaro (Zaragoza, Spain), and Joaquín Espílez (Teruel, Spain) for help in the field and finding some of the outcrops described in this paper. Isabel Pérez (University of Zaragoza, Spain) assisted with photography. This work was supported by the Spanish Ministry of Economy and Competitiveness (grants CGL2017-87631 and CGL2015-64422-P), cofinanced by the European Regional Development Fund. SZ was funded by a Ramón y Cajal Grant (RYC-2012-10576). MALH research is a contribution to the Spanish Ministry of Economy and Competitiveness (grant CGL2014-52096-P), to Basque Government Research Group IT-1044-16 and to the University of the Basque Country UPV/EHU Research Group PPG17/05. Funding for field work has been in part provided by project “Aragosaurus: recursos geológicos y paleoambientales” (ref: E18_17R ) funded by the Government of Aragón.
References
Astibia, H., Payros, A., Ortiz, S., Elorza, J., Álvarez-Pérez, G., Badiola, A., Bardet, N., Berreteaga, A., Bitner, M.A., Calzada, S., Corral, J.C., Díaz-Martínez, I., Merle, D., Pacaud, J.-M., Pereda-Suberbiola, X., Pisera, A., Rodríguez-Tovar, J.-M., and Tosquella, J. 2016. Fossil associations from the middle and upper Eocene strata of the Pamplona Basin and surrounding areas (Navarre, western Pyrenees). Journal of Iberian Geology 42: 7–28. Crossref
Astibia, H., Payros, A., Pereda-Suberbiola, X., Elorza, J., Berreteaga, A., Etxebarria, N., Badiola, A., and Tosquella, J. 2005. Sedimentology and taphonomy of sirenian remains from the Middle Eocene of the Pamplona Basin (Navarre, western Pyrenees). Facies 50: 463–475. Crossref
Astre, G. 1925. Une comatule aptienne de la province de Castellón. Butlletí de la Institució Catalana d’Història Natural 2ª sèrie 5: 176–181.
Aurell, M., Robles, S., Bádenas, B., Quesada, S., Rosales, I., Meléndez, G., and García-Ramos, J.C. 2003. Transgressive–regressive cycles and Jurassic palaeogeography of northeast Iberia. Sedimentary Geology 162: 239–271. Crossref
Aurell, M., Bádenas, B., Ipas, J., and Ramajo, J. 2010. Sedimentary evolution of an Upper Jurassic carbonate ramp (Iberian Basin, NE Spain). In: F. van Buchem, K. Gerdes, and M. Esteban (eds.), Reference Models of Mesozoic and Cenozoic Carbonate Systems in Europe and the Middle East—Stratigraphy and Diagenesis. Geological Society of London, Special Publication 329: 87–109. Crossref
Aurell, M., Soria, A.R., Bádenas, B., Liesa, C.L., Canudo, J.I., Gasca, J.M., Moreno-Azanza, M., Medrano-Aguado, E., and Meléndez, A. 2018. Barremian synrift sedimentation in the Oliete sub-basin (Iberian Basin, Spain): Palaeogeographical evolution and distribution of vertebrate remains. Journal of Iberian Geology 44: 285–308. Crossref
Ausich, W.I. and Wilson, M.A. 2012. New Tethyan Apiocrinitidae (Crinoidea; Articulata) from the Jurassic of Israel. Journal of Paleontology 86: 1051–1055. Crossref
Bataller, J.R. 1930. Els comatulits fossils iberics. Butlletí de la Institució Catalana d’Història Natural 2ª sèrie 10: 63–65.
Bataller, J.R. 1941. Equinodermos fósiles nuevos o poco conocidos de España. Las Ciencias 6 (1): 60–73.
Baumiller, T.K. and Gaździcki, A. 1996. New crinoids from the Eocene la Meseta Formation of Seymour Island Antarctic Peninsula. Palaeontologia Polonica 55: 101–116.
Baumiller, T.K., Salamon, M., Gorzelak, P., Mooi, R., Messing, C.G., and Gahn, F.J. 2010. Post-Paleozoic crinoid radiation in response to benthic predation preceded the Mesozoic marine revolution. Proceedings of the National Academy of Sciences USA 107: 5893–5896. Crossref
Bottjer, D.J., and Jablonski, D. 1988. Paleoenvironmental patterns in the evolution of post-Paleozoic benthic marine invertebrates. Palaios 3: 540–560. Crossref
Bover-Arnal, T., Moreno-Bedmar, J.A., Frijia, G., Pascual-Cebrian, E., and Salas, R. 2016. Chronostratigraphy of the Barremian–Early Albian of the Maestrat Basin (E Iberian Peninsula): Integrating strontium-isotope stratigraphy and ammonoid biostratigraphy. Newsletters on Stratigraphy 49: 41–68. Crossref
Brunnich, N.K. 1913. Crinoideme i Danmarks Kridtaflejringer. Danmarks geologiske Undersogelse 26: 1–120.
Calzada, S. and Gutiérrez, D. 1987. Ofiuras (Echinodermata) del Ladiense catalán. Batalleria 1: 31–38.
Calvet, F. and Marzo, M. 1994. El Triásico de las Cordilleras Costero Catalanas: estratigrafía, sedimentología y análisis secuencial. In: A. Arche (ed.), Excursiones del III Coloquio Estratigráfico y Paleogeográfico del Pérmico y Triásico de España, 1–53. Caja Castilla la Mancha, Cuenca.
Calvet, F. and Tucker, M.E. 1988. Outer ramp cycles in the Upper Muschelkalk of the Catalan Basin, northeast Spain. Sedimentary Geology 57: 185–198. Crossref
Calvet, F., March, M., and Pedrosa, A. 1987. Estratigrafía, sedimentología y diagénesis del Muschelkalk superior de los Catalánides. Cuadernos Geologia Ibérica 11: 171–197.
Canérot, J., Cugny, P., Pardo, G., Salas, R., and Villena, J. 1982. Ibérica Central-Maestrazgo. In: A. García (ed.), El Cretácico de España: 273–344. Universidad Complutense de Madrid, Madrid.
Casanova, S. 1974. Una nueva localidad para Solanocrinites (Decameros) astrei (Bataller, 1940). Acta Geológica Hispánica 5: 151–153.
Charbonnier, S., Vannier, J., Gaillard, C., Bourseau, J.-P., and Hantzpergue, P. 2007. The La Voulte Lagerstätte (Callovian): Evidence for a deep water setting from sponge and crinoid communities. Palaeogeography, Palaeoclimatology, Palaeoecology 250: 216–236. Crossref
Cohen, B.L. and Pisera, A. 2017. Crinoid phylogeny: New interpretation of the main Permo-Triassic divergence, comparisons with echinoids and brachiopods, and EvoDevo interpretations of major morphological variations. Biological Journal of the Linnean Society 120: 38–53.
Cuevas-Gozalo, M., Donselaar, M.E., and Nio, S.D. 1985. Eocene clastic tidal deposits in the Tremp-Graus Basin (Prov. of Lérida and Huesca). In: L.D. Mila and J. Rosell (eds.), Excursion Guidebook, 6th European Regional Meeting: Lérida, Spain, International Association of Sedimentologists, 215–266. Institut d’Estudis Illerdencs, Lérida.
Deusch, M., Friebe, A., Geyer, O.F., and Krautter, M. 1990. Las facies espongiolíticas del Jurásico español y unidades semejantes de Europa Central. Cuadernos de Geología Ibérica 14: 199–214.
Donovan, S.K. and Pawson, D.L. 2008. A new species of the sessile crinoid Holopus d’Orbigny from the tropical western Atlantic, with comments on holopodid ecology (Echinodermata: Crinoidea: Holopodidae). Zootaxa 1717: 31–38.
Eagle, M.K. 1993. A new fossil isocrinid crinoid from the Late Oligocene of Waitete Bay, northern Coromandel. Records of the Auckland Institute and Museum 30: 1–12.
Eagle, M.K. and Hayward, B.W. 1993. Oligocene paleontology and paleoecology of Waitete Bay, northern Coromandel Peninsula. Records of the Auckland Institute and Museum 30: 13–26.
Fezer, R. 1988. Die oberjurassische karbonatische Regressionsfazies im südwestlichen Keltiberikum zwischen Griegos und Aras de Alpuente (Prov. Teruel, Cuenca, Valencia, Spanien). Arbeiten aus dem Institut für Geologie und Paläontologie an der Universität Stuttgart 84: 1–119.
Foote, M. 1999. Morphological diversity in the evolutionary radiation of Paleozoic and Post-Paleozoic crinoids. Paleobiology 25 (S2): 1–115. Crossref
Fortuny, J., Bolet, A., Sellés, A.G., Cartanyà, J., and Galobart, À. 2011. New insights on the Permian and Triassic vertebrates from the Iberian Peninsula with emphasis on the Pyrenean and Catalonian basins. Journal of Iberian Geology 37: 65–86.
García, R., Moreno-Bedmar, J.A., Bover-Arnal, T., Company, M., Salas, R., Latil, J.L., Martín-Martín, J.D., Gómez-Rivas, E., Bulot, L.G., Delanoy, G., Martínez, R., and Grauges, A. 2014. Lower Cretaceous (Hauterivian–Albian) ammonite biostratigraphy in the Maestrat Basin (E Spain). Journal of Iberian Geology 40: 99–112. Crossref
García-Mondéjar, J., Agirrezabala, L.M., Aranburu, A., Fernández-Mendiola, P.A., Gómez-Pérez, I., López-Horgue, M.A., and Rosales, I. 1996. Aptian–Albian tectonic pattern of the Basque-Cantabrian Basin. Geological Journal 31: 13–45.Crossref
Guensburg, T.E. and Sprinkle, J. 2001. Earliest crinoids: New evidence for the origin of the dominant Paleozoic echinoderms. Geology 29: 131–134. Crossref
Gorzelak, P., Salamon, M.A., and
Baumiller, T.K. 2012. Predator-induced macroevolutionary trends in
Mesozoic crinoids. Proceedings of the National
Academy of Sciences USA 109: 7004–7007. Crossref
Gorzelak, P., Salamon, M.A., Trzęsiok, D., Lach, R., and Baumiller, T.K. 2016. Diversity dynamics of post-Palaeozoic crinoids—in quest of the factors affecting crinoid macroevolution. Lethaia 49: 231–244. Crossref
Hagdorn, H. 1978. Muschel/Krinoiden-Bioherme im Oberen Muschelkalk (mo1, Anis) von Crailsheim und Schwäbisch Hall (Südwestdeutschland). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 156: 31–86.
Hagdorn, H. 2011. Triassic: the crucial period of post-Palaeozoic crinoid diversification. Swiss Journal of Palaeontology 130: 91–112. Crossref
Hagdorn, H., Głuchowski, E., and Boczarowski, A. 1996. The crinoid fauna of the Diplopora Dolomite (Middle Muschelkalk, Triassic, Upper Anisian) at Piekary Śląskie in Upper Silesia. Geologisch-Paläontologische Mitteilungen Innsbruck 21: 47–87.
Hemery, L.G. 2011. Diversité moléculaire, phylogéographie et phylogénie des crinoïdes (Echinodermes) dans un environnement extrême: l’océan Austral. 381 pp. Unpublished Ph.D. Thesis, Muséum national d’Histoire naturelle, Paris.
Hemery, L.G., Roux, M., Ameziane, N., and Eleaume, M. 2013. High-resolution crinoid phyletic inter-relationships derived from molecular data. Cahiers de Biologie Marine 54: 511–523.
Hess, H. 1975. Die fossilen Echinodermen des Schweizer Juras. Veröffentlichungen aus dem Naturhistorischen Museum, Basel 8: 1–130.
Hess, H. and Messing, C.G. 2011. In: P. Selden (ed.), Treatise on Invertebrate Paleontology Part T, Echinodermata 2, Revised, Crinoidea 3, 1–261. The University of Kansas Paleontological Institute, Lawrence.
Hess, H. and Gale, A.S. 2010. Crinoids from the Shenley Limestone (Albian) of Leighton Buzzard, Bedfordshire, UK. Journal of Systematic Palaeontology 8: 427–447. Crossref
Hess, H., Salamon, M.A., and Gorzelak, P. 2011. Late Jurassic–Early Cretaceous (Tithonian–Berriasian) cyrtocrinids from southeastern Poland. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 260: 119–128.
Jagt, J.W.M. 1999. Late Cretaceous–Early Palaeogene echinoderms and the K/T boundary in the southeast Netherlands and northeast Belgium—Part 2: Crinoids. Scripta Geologica 116: 59–255.
Jagt, J.W.M. and Salamon, M.A. 2007. Late Cretaceous bourgueticrinid crinoids from southern Poland—preliminary observations. Scripta Geologica 134: 61–76.
Kjaer, C.R. and Thomsen, E. 1999. Heterochrony in bourgueticrinid sea-lilies at the Cretaceous/Tertiary boundary. Paleobiology 25: 29–40.
Lane, N.G. and Webster, G.D. 1980. Crinoidea. In: T.W. Broadhead and J.A. Waters (eds.), Echinoderms: Notes For a Short Course. University of Tennessee, Studies in Geology 3: 144–157.
Lertxundi, D. and García-Mondéjar, J. 1998. Facies and constructional dynamics of an Albian carbonate pinnacle (Aitzondo buildup, Aralar, Basque-Cantabrian Basin). In: J.C. Cañaveras, García M.A., and J. Soria (eds.), Abstracts of the 15th Sedimentological Congress, 500–501. Universidad de Alicante, Alicante.
López-Gómez, J., Márquez-Aliaga, A., Calvet, F., Márquez, L., and Arche, A. 1994. Aportaciones a la estratigrafía y paleontología del Triásico Medio de los alrededores de Agost, Alicante. Boletín de la Real Sociedad española de Historia Natural (Sección Geológica) 89: 109–120.
López-Horgue, M.A., Hernández-Samaniego, A., Baceta Caballero, J.I., Faci Paricio, E., and Larrañaga Sancena, M.J. 1997. Mapa Geológico de Navarra a Escala 1:25.000, Hoja 114-II (Uharte-Arakil). Gobierno de navarra, servicio de obras públicas, Pamplona.
López-Horgue, M.A., Lertxundi-Manterola, D., and Baceta-Caballero, J.I. 1996. Evolución sedimentaria del episodio mixto carbonatado-terrígeno del Albiense Superior–Cenomaniense Inferior entre Altsasu (Nafarroa) y Asparrena (Araba): la Unidad Albeniz. Príncipe de Viana 14–15: 81–96.
López-Horgue, M.A., Owen, H.G., Rodríguez-Lázaro, J., Orue-Etxebarria, X., Fernández-Mendiola, P.A., and García-Mondéjar, J. 1999. Late Albian–Early Cenomanian stratigraphic succession near Estella-Lizarra (Navarra, central northern Spain) and its regional and interregional correlation. Cretaceous Research 20: 369–402. Crossref
Loriol, P. de 1900. Notes pour servir à l’ étude des échinodermes. 1re Série, 8e article. Revue Suisse de Zoologie 8: 55–96.
Meléndez, G. and Fontana, B. 1993. Biostratigraphic correlation of the Middle Oxfordian sediments in the Iberian Chain, eastern Spain. Acta Geologica Polonica 43: 193–211.
Meyer, D.L. and Macurda, D.B. 1977. Adaptive radiation of the comatulid crinoids. Paleobiology 3: 74–82. Crossref
Meyer, D.L. and Oji, T.1993. Eocene crinoids from Seymour Island, Antarctic Peninsula: paleobiogeographic and paleoecologic implications. Journal of Paleontology 67: 250–257. Crossref
Mora-Morote, P. 2009. Comatúlidos en el Cretácico Inferior de la Alcoraya (Alicante). Cidaris 29: 17–22.
Moreno-Bedmar, J.A., Company, M., Bover-Arnal, T., Salas, R., Maurrasse, F.J., Delanoy, G., Grauges, A., and Martínez, R. 2010. Lower Aptian ammonite biostratigraphy in the Maestrat Basin (Eastern Iberian Chain, Eastern Spain). A Tethyan transgressive record enhanced by synrift subsidence. Geologica Acta 8: 281–299.
Murray, J.W. 1991. Ecology and Palaeoecology of Benthic Foraminifera. 397 pp. Longman Scientific & Technical, New York.
Odin, G.S. (compiler). 2001. The Campanian–Maastrichtian boundary: definition at Tercis (Landes, SW France) principle, procedure, and proposal. In: G.S. Odin (ed.), The Campanian–Maastrichtian Stage Boundary: Characterisation at Tercis les Bains (France) and Correlation with Europe and other Continents. Developments in Palaeontology and Stratigraphy 19: 820–833.
Oji, T. 1989. Distribution of the stalked crinoids from Japanese and nearby waters. In: H. Ohba, I. Hayami, and K. Mochizuki (eds.), Current Aspects of Biogeography in West Pacific and East Asian Regions, 27–43. University Museum, University of Tokyo, Tokyo.
Oji, T. 1996. Is predation intensity reduced with increasing depth? Evidence from the west Atlantic stalked crinoid Endoxocrinus parrae (Gervais) and implications for the Mesozoic marine revolution. Paleobiology 22: 339–351. Crossref
Palmer, T.J. and Fürsich, F.T. 1974. The ecology of a Middle Jurassic hardground and crevice fauna. Palaeontology 17: 507–524.
Pomar, L., Aurell, M., Bádenas, B., Morsilli, M., and Al-Awwad, S.F. 2015. Depositional model for a prograding oolitic wedge, Upper Jurassic, Iberian Basin. Marine and Petroleum Geology 67: 556–582.Crossref
Ramajo, J. and Aurell, M. 2008. Long-term Callovian–Oxfordian sea-level changes and sedimentation in the Iberian carbonate platform (Jurassic, Spain): possible eustatic implications. Basin Research 20: 163–184. Crossref
Ramajo, J., Aurell, M., Bádenas, B., Bello, J., Delvene, G., Meléndez, G., and Pérez-Urresti, I. 1999. Síntesis bioestratigráfica y paleogeográfica del Oxfordiense en la Cuenca Ibérica (Sector Oriental) y correlación con la Cuenca Catalana. Cuadernos de Geología Ibérica 25: 111–138.
Rasmussen, H.W. 1961. A monograph on the Cretaceous Crinoidea. Biologiske Skrifter udgivet af det kongelige Danske Videnskabernes Selskab 12: 1–428.
Rosell, J., Tritlla, J., Obrador, A., Peon, A., Alonso, F., Ramírez, J., and Cabañas, J. 1975. Mapa Geológico de España. E. 1:50 000 Sabadell (392). 31 pp. Instituto Geológico y Minero de España, Madrid.
Rouse, G.W., Jermiin, L.S., Wilson, N.G., Eeckhaut, I., Lanterbecq, D., Oji, T., Young, C.M., Browning, T., Cisternas, P., Helgen, L.E., Stuckey, M., and Messing, C.G. 2013. Fixed, free, and fixed: The fickle phylogeny of extant Crinoidea (Echinodermata) and their Permian–Triassic origin. Molecular Phylogenetics and Evolution 66: 161–181. Crossref
Roux, M. and Plaziat, J.C. 1978. Inventaire des Crinoïdes et interprétation paléobathymétrique de gisements du Paléogêen franco-espagnol. Bulletin de la Société Géologique de France, sér. 7 20 (3): 299–308. Crossref
Roux, M., Eleaume, M., Hemery L.G., and Ameziane, N. 2013. When morphology meets molecular data in crinoid phylogeny: a challenge. Cahiers de Biologie Marine 54: 541–548.
Salamon, M.A. 2008. Jurassic cyrtocrinids (Cyrtocrinida, Crinoidea) from extra-Carpathian Poland. Palaeontographica Abteilung A 285: 77–99. Crossref
Salamon, M.A. and Gorzelak, P. 2007. Evidence of shallow-water cyrtocrinids (Crinoidea) from the Callovian of Poland. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 244: 257–260. Crossref
Salamon, M.A., and Gorzelak, P. 2010. Late Cretaceous crinoids (Crinoidea) from Eastern Poland. Palaeontographica Abteilung A 291: 1–43. Crossref
Salamon, M.A., Gorzelak, P., Borszcz, T., Gajerski, A., and Kaźmierczak, J. 2009. A crinoid concentration Lagerstätte in the Turonian (Late Cretaceous) Conulus Bed (Miechów-Wolbrom area, Poland). Geobios 42: 351–357. Crossref
Serra-Kiel, J., Canudo, J.I., Dinares, J., Molina, E., Ortiz, N., Pascual, J.O., Samsó, J.M., and Tosquella, J. 1994. Cronoestratigrafía de los sedimentos marinos del Terciario inferior de la Cuenca de Graus-Tremp (Zona Central Pirenaica). Revista de la Sociedad Geológica de España 7 (3–4): 273–297.
Simms, M.J. 1988. The phylogeny of post-Palaeozoic crinoids. In: C.R.C. Paul and A.B. Smith (eds.), Echinoderm Phylogeny and Evolutionary Biology, 269–284. Clarendon Press, Oxford.
Simms, M.J. and Sevastopulo, G.D. 1993. The origin of articulate crinoids. Palaeontology 36: 91–109.
Smith, A.B. and Gallemí, J. 1991. Middle Triassic holothurians from northern Spain. Palaeontology 34: 49–76.
Stevenson, A., Gahn, F., Baumiller, T.K., and Sevastopulo, G. 2017. Predation on feather stars by regular echinoids as evidenced by laboratory and field observations and its paleobiological implications. Paleobiology 43: 274–285. Crossref
Stilwell, J.D., Fordyce, R.E., and Rolfe, P.J. 1994. Paleocene isocrinids (Echinodermata: Crinoidea) from the Kauru Formation, South Island, New Zealand. Journal of Paleontology 68: 135–141. Crossref
Tosquella, J. 1988. Estudi sedimentològic i bioestratigràfic de la Formació Gresos de Roda (Eocè, Conca de Tremp-Graus). 540 pp. Unpublished M.Sc. Thesis, University of Barcelona, Barcelona.
Twitchett, R.J. and Oji, T. 2005. Early Triassic recovery of echinoderms. Comptes Rendus Palevol 4: 531–542. Crossref
Vennin, E., Soria, A.R., Meléndez, A., and Preat, A. 1993. Análisis secuencial durante el intervalo Barremiense–Aptiense en la Cubeta de Oliete. Cuadernos de Geología Ibérica 17: 257–283.
Whittle, R.J., Hunter, A.W., Cantrill, D.J., and McNamara, K.J. 2018. Globally discordant Isocrinida (Crinoidea) migration confirms asynchronous Marine Mesozoic Revolution. Communications Biology 1 (46): 1–10.
Wiedmann, J. 1962. Ammoniten aus der Vascogotischen Kreide (Nordspanien), I. Phylloceratina, Lytoceratina. Palaeontographica Abteilung A 118: 119–237.
Wright, D.F., Ausich, W.I., Cole, S. R., Peter, M.E., and Rhenberg, M.E. 2017. Phylogenetic taxonomy and classification of the Crinoidea (Echinodermata). Journal of Paleontology 91: 829–846. Crossref
Zatoń, M., Salamon, M.A., and Kaźmierczak, J. 2008. Cyrtocrinids (Crinoidea) and associated stalked crinoids from the Lower/Middle Oxfordian (Upper Jurassic) shelfal deposits of southern Poland. Geobios 41: 559–569. Crossref
Acta Palaeontol. Pol. 63 (4): 779–794, 2018
https://doi.org/10.4202/app.00520.2018