
A new rebbachisaurid sauropod from the Aptian–Albian, Lower Cretaceous Rayoso Formation, Neuquén, Argentina
JOSÉ I. CANUDO, JOSÉ L. CARBALLIDO, ALBERTO GARRIDO, and LEONARDO SALGADO
Canudo, J.I., Carballido, J.L., Garrido, A., and Salgado, L. 2018. A new rebbachisaurid sauropod from the Aptian–Albian, Lower Cretaceous Rayoso Formation, Neuquén, Argentina. Acta Palaeontologica Polonica 63 (4): 679–691.
Rebbachisaurids are a group of basal diplodocimorph sauropods that diversified in Gondwana at the end of the Early Cretaceous and the beginning of the Late Cretaceous. It is a group of great palaeobiogeographical interest, for it clearly illustrates various processes of dispersal throughout Gondwana and to Laurasia prior to the breakup of Africa and South America. However, the relationships within the group are still under discussion owing to the scarcity of cranial material that would help clarify them. In the present paper we describe the new rebbachisaurid Lavocatisaurus agrioensis gen. et sp. nov. from the Aptian–Albian (Lower Cretaceous) of Neuquén (Argentina). Remains have been recovered belonging to an adult specimen (holotype) and two immature specimens (paratypes). Taken together, almost all the bones of the taxon are represented, including most of the cranium. Lavocatisaurus agrioensis gen. et sp. nov. is the first rebbachisaurid from Argentina with an almost complete cranium, making it possible to recognize differences with respect to other rebbachisaurids, such as the highly derived Nigersaurus. Among its most notable characters are the presences of a large preantorbital fenestra and maxillary teeth that are significantly larger than those of the dentary. Our phylogenetic study places Lavocatisaurus amongst basal rebbachisaurids, as the sister lineage to Khebbashia (the clade formed by Limaysaurinae + Rebbachisaurinae). This position, which is somewhat more derived than that previously suggested for Comahuesaurus and Zapalasaurus (the Argentinean rebbachisaurids closest in geographical and geological terms), reaffirms the presence of different basal rebbachisaurid lineages in the Early Cretaceous of Patagonia.
Key words: Dinosauria, Rebbachisauridae, phylogeny, Cretaceous, Rayoso Formation, Argentina.
José I. Canudo [jicanudo@unizar.es], Aragosaurus: Recursos geológicos y paleoambientes; IUCA, Facultad de Ciencias, Universidad de Zaragoza 50009, C/ Pedro Cerbuna 12, Zaragoza, Spain.
José L. Carballido [jcarballido@mef.org.ar], CONICET-Museo “Egidio Feruglio”, Fontana 140, 9100 Trelew, Chubut, Argentina.
Alberto C. Garrido [albertocarlosgarrido@gmail.com], Museo Provincial de Ciencias Naturales “Prof. Dr. Juan Olsacher”, Dirección Provincial de Minería, Etcheluz y Ejército Argentino, 8340 Zapala, Neuquén, Argentina; Departamento Geología y Petróleo, Facultad de Ingeniería, Universidad Nacional del Comahue, Buenos Aires 1400, 8300 Neuquén, Neuquén, Argentina.
Leonardo Salgado [lsalgado@unrn.edu.ar], CONICET-Instituto de Investigación en Paleobiología y Geología, Universidad Nacional de Río Negro, Av. Gral. J.A. Roca 1242, 8332 General Roca, Río Negro, Argentina.
Received 4 July 2018, accepted 25 September 2018, available online 29 October 2018.
Copyright © 2018 J.I. Canudo et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Rebbachisaurids are a group of diplodocimorph sauropods that are thought to have diversified around the middle of the Early Cretaceous (Salgado et al. 2006; Sereno et al. 2007; Carballido et al. 2012; Fanti et al. 2013; Wilson and Allain 2015). They are the last diplodocimorphs to go extinct and the only ones to reach the Upper Cretaceous of Gondwana (Ibiricu et al. 2013; Wilson and Allain 2015). Phylogenetic studies place their appearance in the Middle Jurassic as the sister group to the diplodocids and dicraeosaurids (Wilson and Allain 2015; Tschopp et al. 2015), yet they are maintained as a ghost lineage through the Jurassic and part of the Lower Cretaceous, since rebbachisaurids are unrecorded in this period. The oldest rebbachisaurid is Histriasaurus, described from fragmentary remains from the upper Hauterivian–lower Barremian of Croatia (Dalla Vechia 1998). The most modern rebbachisaurids are several taxa recovered from the Cenomanian–Turonian of Argentina (Calvo and Salgado 1995; Garrido 2010; Haluza et al. 2012; Ibiricu et al. 2012).
The identification of the rebbachisaurid lineage is relatively recent. Bonaparte (1997) first proposed its differentiation from other sauropods on the basis of fragmentary postcranial material from the Upper Cretaceous of Neuquén (Patagonia, Argentina), currently identified as Rayososaurus agrioensis (Carballido et al. 2010). Bonaparte (1997) noted the singular morphology of the scapula of R. agrioensis, which was similar to that of Rebbachisaurus garasbae, from the Cenomanian of Morocco (Lavocat 1954; Wilson and Allain 2015). Rebbachisaurus was described on the basis of a scapula and a dorsal vertebra (Lavocat 1954); moreover, there are some bones stored in the Museum National d’Histoire Naturelle of Paris (France) that were not part of the original description (Wilson and Allain 2015). One of the consequences of the presence of rebbachisaurids in Africa and South America is their special palaeobiogeographical interest for the Early–Late Cretaceous transition. This was pointed out initially by Calvo and Salgado (1995) and subsequently used to identify dispersal and vicariance events of terrestrial tetrapods between different parts of Gondwana and between Gondwana and Laurasia (Canudo et al. 2009; Mannion 2009; Torcida Fernandez-Baldor et al. 2011; Carballido et al. 2012; Fanti et al. 2015).
Recent years have seen major advances in what is known of the anatomy and palaeogeographical distribution of the rebbachisaurids, both in South America (Carvalho et al. 2003; Gallina and Apesteguía 2005; Carballido et al. 2012; Ibiricu et al. 2013) and Africa (Sereno et al. 1999; Fanti et al. 2013), as well as their unexpected presence in Europe (Pereda Suberbiola et al. 2003; Mannion 2009; Torcida Fernandez-Baldor et al. 2011). These discoveries have allowed the rebbachisaurids to be placed beyond question at the base of the radiation of the diplodocimorph diplodocoids, and in all the phylogenies they are grouped in a clade that is clearly differentiated from the rest of the diplodocimorphs (e.g., Calvo and Salgado 1995; Wilson and Allain 2015; Tschopp et al. 2015). Recently, a consensus has been reached on the existence of two main lineages of rebbachisaurids (Limaysaurinae and Rebbachisaurinae), currently grouped in Khebbashia. Most species are described using only fragmentary material and, except for Nigersaurus, with little cranial material. This has resulted in a degree of instability in certain taxa, as well as major gaps in what is known about the early evolutionary stages of the group, in particular with respect to its cranial morphology.
In 2009, the Spanish-Argentinean research group that has authored the present paper began to look for the exact site where the bones of the rebbachisaurid Rayososaurus agrioensis had been recovered, our aim being to reopen the excavation in search of more elements from the holotype. In the course of our revision of the Rayoso Formation in the area where R. agrioensis had been collected, we found the remains of another rebbachisaurid, which was excavated between 2010 and 2011. These remains correspond to a partially articulated adult specimen and two juvenile individuals, ascribed provisionally to cf. Zapalasaurus (Salgado et al. 2012). These fieldwork campaigns resulted in the recovery of elements that afford a good idea of the anatomy of this new dinosaur, including cranial remains. Over the last few years, the material has been prepared by mechanical means in the Olsacher Museum (MOZ) and the Trelew Museum (MEF). The aim of the present article is to provide a brief description of this rebbachisaurid, which represents the first dinosaur from the Rayoso Formation, and to discuss its systematics and its phylogenetic importance.
Institutional abbreviations.—MEF, Museo paleontológico Egidio Feruglio, Trelew, Chubut, Argentina; MNN GDF, Musée National Boubou Hama, Niamey, Niger; MOZ, Museo Provincial de Ciencias Naturales “Dr. Prof. Juan A. Olsacher”, Zapala, Argentina.
Other abbreviations.—ITR, isolated tooth rows; MDR, more derived rebbachisaurids.
Geographical and geological setting
The rebbachisaurid bones studied in this paper were recovered near the locality of Agrio del Medio, a small town in the central part of the province of Neuquén (Patagonia, Argentina; Fig. 1). In geological terms, Salgado et al. (2012) situated the fossils in the Rayoso Formation, one of the Lower Cretaceous units that outcrop in the Neuquén Basin (Leanza 2003). The four members of this formation can be clearly identified in the area around Agrio del Medio; from the oldest to youngest these are the Rincón, Quili Malal, Pichi Neuquén, and Cañadón de la Zorra members. Two narrow-gauge sauropod trackways attributed to rebbachisaurids have been found in two distinct outcrops of the Rincón Member (Canudo et al. 2017; Fig. 1). The rebbachisaurid bones studied in this paper were found in the middle part of the Pichi Neuquén Member, approximately 44 metres from its base (Fig. 1). Lithologically, this member is made up of a stratified sequence of detritic levels of quartz-bearing sandstone and reddish clay deposited in coastal environments (Leanza 2003). There is no consensus regarding the age of this member, for no direct datings have been taken. Some authors place it in the Aptian (Musacchio and Vallati 2000), whereas Leanza (2003) considers it to be rather more modern (lower Albian). In this paper, we ascribe an age range from Aptian to lower Albian to the new rebbachisaurid.
The fossils of the new sauropod were found in a thin stratum of sandstone of reddish-brown colour. This sandstone displayed slight lamination, with levels of current ripples and mudcracks. However, the fossil bones did not coincide with a surface of current ripples. Furthermore, the sandstone was slightly bioturbated, with full-relief burrows both vertical (Skolithos-type) and random in distribution and unbranching (Planolites-type). The geometry of the sandstone and the structures they present suggest that it was deposited in non-confined deposits formed by decantation in areas with high sedimentation and low energy (Salgado et al. 2012).
The nomenclature for the vertebral laminae follows Wilson (1999), with modifications (accessory posterior centrodiapophyseal lamina) from Salgado et al. (2005) and Wilson et al. (2011). The nomenclature for the vertebral pneumatic structures follows Wedel (2003) and Wilson et al. (2011).
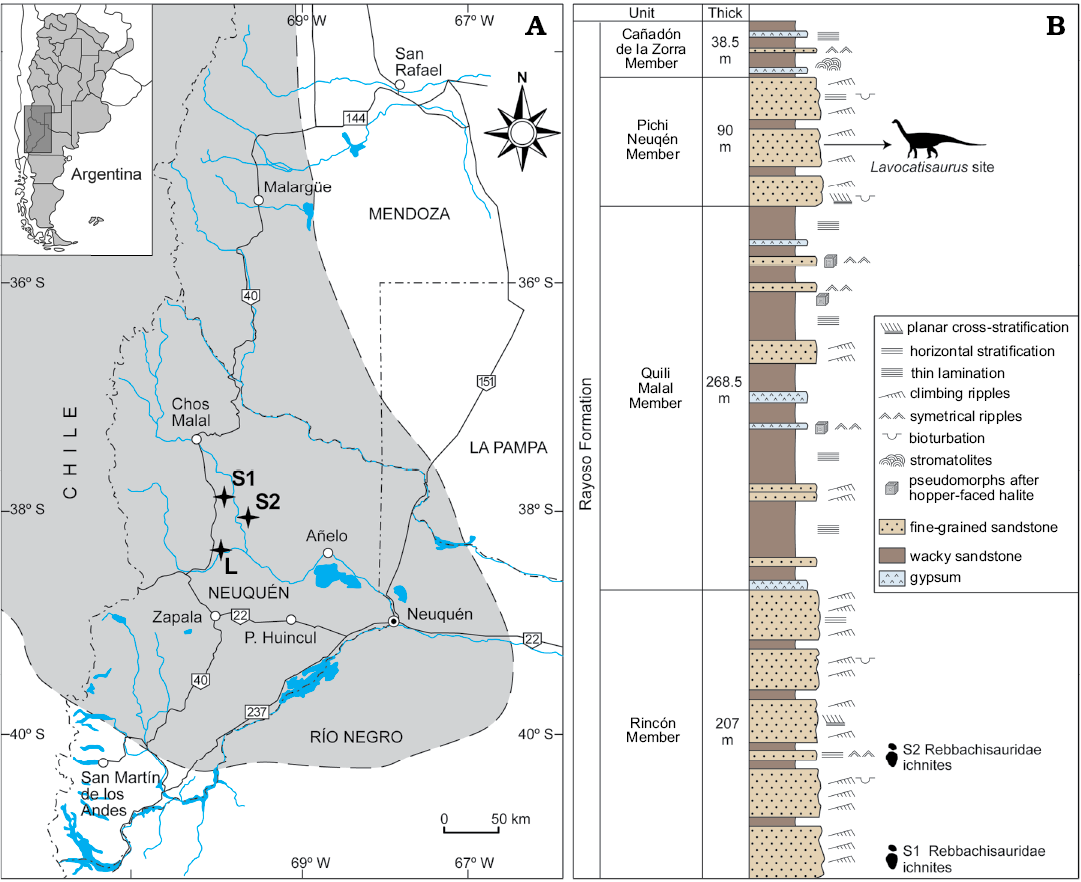
Fig. 1. A. Map of location and geological outcrops of the region of Agrio del Medio (Neuquén, Argentina), where the material was collected; gray area represents the Neuquén Basin. B. Lithostratigraphic column of the Rayoso Formation in the study area. L, Lavocatisaurus site; S1 and S2, sites of the Rebbachisauridae ichnites (Canudo et al. 2017).
Systematic palaeontology
Saurischia Seeley, 1887
Sauropoda Marsh, 1878
Diplodocoidea Marsh, 1878 sensu Upchurch, 1995
Diplodocimorpha Calvo and Salgado, 1995
Rebbachisauridae Bonaparte, 1997
Genus Lavocatisaurus nov.
Type species: Lavocatisaurus agrioensis sp. nov., see below.
Etymology: In honour of the French researcher René Lavocat (1909–2007), who described Rebbachisaurus, the first known representative of Rebbachisauridae.
Diagnosis.—As for the type and only known species.
Lavocatisaurus agrioensis sp. nov.
Figs. 2, 3.
Etymology: In reference to the locality of Agrio del Medio, from which the new species was found.
Type material: Holotype: MOZ-Pv1232, partially articulated specimen: dentaries, left surangular, premaxillae and maxillae, left jugal, right squamosal, quadrates, 23 isolated teeth and two series of 8 and 9 maxillary teeth, hyoid bone, 11 cervical vertebrae (including atlas and axis), 28 caudal vertebrae, cervical ribs, 2 dorsal ribs, humerus, fragment of ?radius. Paratypes, all from type locality: the juvenile specimens (Salgado et al. 2012): MOZ-Pv 1248, posterior cervical centrum; MOZ-Pv 1249, cervical neural arch; MOZ-Pv 1251, dorsal neural arch; MOZ-Pv 1252, 1253, 1254, anterior caudal centra; MOZ-Pv 1255, scapula; MOZ-Pv 1267, left radius; MOZ-Pv 1256, left ulna; MOZ-Pv 1257, right metatarsal I; MOZ-Pv 1258, ?metatarsal V. An association of MOZ-Pv 1233, 1250, cervical centra; MOZ-Pv 1236, 1237, incomplete cervical neural arches; MOZ-Pv 1238, 1239, fragmentary neural arches; MOZ-Pv 1240, dorsal centrum; MOZ-Pv 1241, rib fragments; MOZ-Pv 1242, haemal arch; MOZ-Pv 1243, right ulna; MOZ-Pv 1244, left tibia; MOZ-Pv 1245, left fibula; MOZ-Pv 1246, end of metatarsal; MOZ-Pv 1247, indeterminate flat fragment.
Type locality: Agrio del Medio site, Neuquén Province, Argentina.
Type horizon: Aptian–lower Albian, Lower Cretaceous.
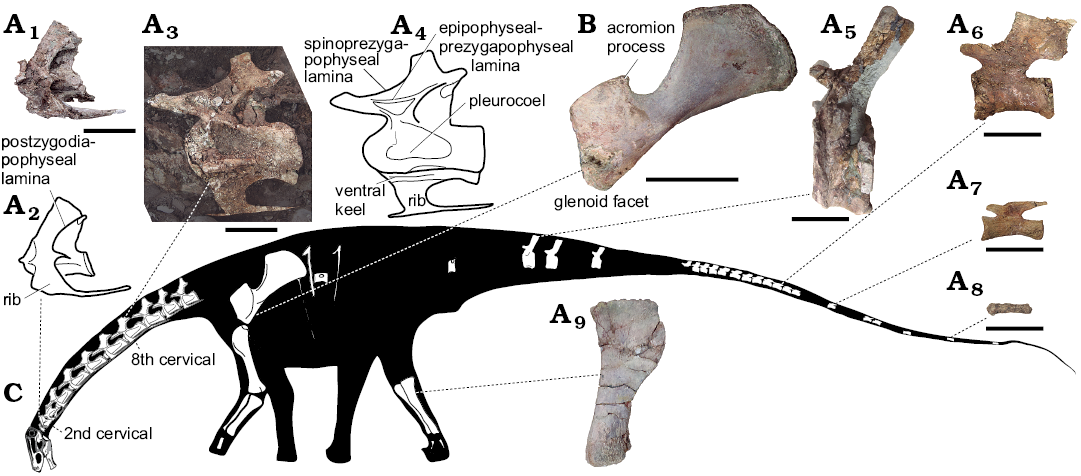
Fig. 2. Rebbachisaurid sauropod Lavocatisaurus agrioensis gen. et sp. nov. from Agrio del Medio (Argentina), Aptian–lower Albian. A. MOZ-Pv1232, axis in lateral view (A1, photograph; A2, drawing); eight cervical vertebrae in lateral view view (A3, photograph; A4, drawing, ); anterior caudal vertebra in lateral view (A5); middle caudal vertebra in lateral view (A6); posterior caudal vertebra in lateral view (A7); posteriormost caudal vertebra in lateral view (A8); left tibia in lateral view (A9). B. MOZ-Pv 1255, left scapula from a juvenile specimen, in lateral view. C. Skeletal reconstruction based on the holotype and paratype specimens. Scale bars 10 cm.
Diagnosis.—A middle-sized rebbachisaurid sauropod diagnosed by the following combination of characters (unique characters are marked with an *): extremely well-developed preantorbital fenestra (shared with Nigersaurus); marked laterodorsal fossa in the dentary (shared with Demandasaurus and Nigersaurus); ventrally expanded squamosal (shared with Limaysaurus); dentary with pronounced ventral projection in the mesio-ventral corner; jugal long and contacting the squamosal (shared with Nigersaurus) but without foramina as present in Nigersaurus; *maxillary teeth significantly larger than the mandibular teeth; middle caudal vertebrae with anteriorly (nearly horizontally) projecting prezygapophysis
Remarks.—Lavocatisaurus agrioensis gen. et sp. nov. is represented by almost all the anatomical elements, with the exception of the neural arches of the dorsal vertebrae (there is only one dorsal centrum, corresponding to one of the paratypes). Salgado et al. (2012) described in detail the bones of the juvenile specimens such as the radius, fibula, ulna, metacarpals, etc.; that description will not be repeated in the present work. Rather, we shall briefly describe the bones of the holotype, and any bones of the paratypes not described in the work cited above. Most of the adult material here presented is still under preparation, therefore a detailed description of it is out of the scope of the present work and will be presented once all the materials have been prepared. At the moment, and given the low number of elements in common between the juvenile specimens and the adult specimen, we couldn’t find any reliable ontogenetic difference.
Rebbachisaurids skull anatomy is mainly based on the reconstructions of Nigersaurus, the single taxon of this clade that preserved enough bones for allowing such reconstruction (Sereno et al. 2007). The new taxon here described allowed us to present an almost complete skull reconstruction, which was through comparisons with Nigersaurus (Sereno et al. 2007) and diplodocids (e.g., Diplodocus; Whitlock et al. 2010). As reconstructed (Fig. 3), the cranium of Lavocatisaurus was elongated, as in diplodocids. Indeed, the general shape of the skull resembles that of Diplodocus more than that of Nigersaurus (although sharing some new characters with the latter). The premaxilla is elongated, with space for 4 teeth, as in other sauropods. The maxilla is anteroposteriorly elongated, with a long ascending process, and pierced by a very large oval preantorbital fenestra (Fig. 3A, B). A completely opened preantorbital fenestra is present in derived titanosaurs such as Tapuiasaurus and diplodocids such as Diplodocus (Marsh 1884; Zaher et al. 2011). A similar opening was initially labelled in Nigersaurus as the antorbital fenestra (Sereno et al. 1999: fig. 2D). Based on our personal observation on the skull elements of Nigersaurus we reinterpret this opening as the preantorbital fenestra, which is not reconstructed in following reconstructions published for this taxon (Sereno and Wilson 2006; Sereno et al. 2007). There are only 12 dental positions in the preserved maxillae, differing from the 25 dental positions of Nigersaurus (Sereno et al. 2007) but having a similar number of maxillary teeth presents in diplodocids (Whitlock et al. 2010; Tschopp and Mateus 2017). The maxillary teeth are large, decreasing in size laterally, as evidenced by the shape of the lateral plate of the maxillae (Fig. 3B2) and the two series of isolated functional teeth (see below). The nutritional foramina of the premaxilla and maxilla are small and circular as in most sauropods and differing from the extremely elongated nutritional foramina of Nigersaurus (Sereno et al. 1999: fig. 2D). The jugal is elongated and fragile bone, lacking the foramina present in Nigersaurus. A novel articulation of the jugal with the squamosal excludes the postorbital from the infratemporal fenestra, resulting in an uncommon morphology not described for other sauropods. Nevertheless, a similar articulation and the resulting exclusion of the postorbital from the infratemporal fenestra can be detected in Limaysaurus (Calvo and Salgado 1995: fig. 4) and Nigersaurus (Sereno et al. 2007: fig. 1), indicating that this is a relatively widespread morphology amongst rebbachisaurids. The squamosal is ventrally expanded as in Limaysaurus, although in the latter the broad ventral expansion of the squamosal was interpreted as the quadratojugal by Calvo and Salgado (1995: fig. 4). The mandible has the typical rectangular outline, as in other diplodocids and derived titanosaurs, and presents 22 dental positions. The elevated number of dentary teeth in L. agrioensis gen. et sp. nov. is an uncommon character amongst Neosauropoda, differing from the 9 tooth positions of Demandasaurus (Torcida Fernández-Baldor et al. 2011) but also from the 34 dentary teeth of Nigersaurus (Sereno et al. 2007). Although no functional teeth are preserved in any of the dentaries, several replacement teeth are observed, which seem to correspond to the first line of replacement teeth. These are much smaller and more fragile than those of the maxillae, an unusual discrepancy in size interpreted as an autapomorphy of the new taxon. A laterodorsal fossa is present in the dentary, as is a pronounced ventral projection in the mesio-ventral corner. The presence of an anteroventrally directed triangular process is a widespread morphology amongst Diplodocimorpha, with a reversion in more derived rebbachisaurids (Demandasaurus and Nigersaurus). Teeth are located exclusively in the anteriormost part of the maxilla and the mandibles (Fig. 3). The teeth are elongated, with an slenderness index (sensu Upchurch 1998) of around 7–8, fine and pencil-like, as in all other diplodocimorphs known up to date (e.g., Nigersaurus, Diplodocus, Galeamopus; Sereno et al. 2007; Whitlock et al. 2010; Tschoopp and Mateus 2017). The compression index (sensu Díez Díaz et al. 2013) cannot be evaluated at the moment as most of the teeth are still embedded in the matrix (Fig. 3G1, H1). The enamel is asymmetrical, being around 5 times thicker on the labial surface than on the lingual one. Three series of functional teeth were recovered in what seems to be their natural position. One is composed by 9 teeth from the left maxilla (Fig. 3G1), the second has 10 teeth from the right maxilla (Fig. 3H1), and the third is composed by 4 teeth from the premaxilla. Given their size and in comparison with the alveoli of the upper and lower jaws and preserved dentary teeth, they are interpreted as upper teeth. As noted by Wiersma and Sander (2016), isolated tooth rows (ITRs) are not uncommon and have been reported for other sauropods such as Shunosaurus, Apatosaurus, Europasaurus, Giraffatitan, and Phuwiangosaurus (see Wiersma and Sander 2016, and references therein). As mentioned above, the dentary teeth are much smaller than the maxillary teeth and lack wear facets (at least those preserved), whereas the maxillary teeth have a single, low-angle lingual wear facet (Fig. 3H1).
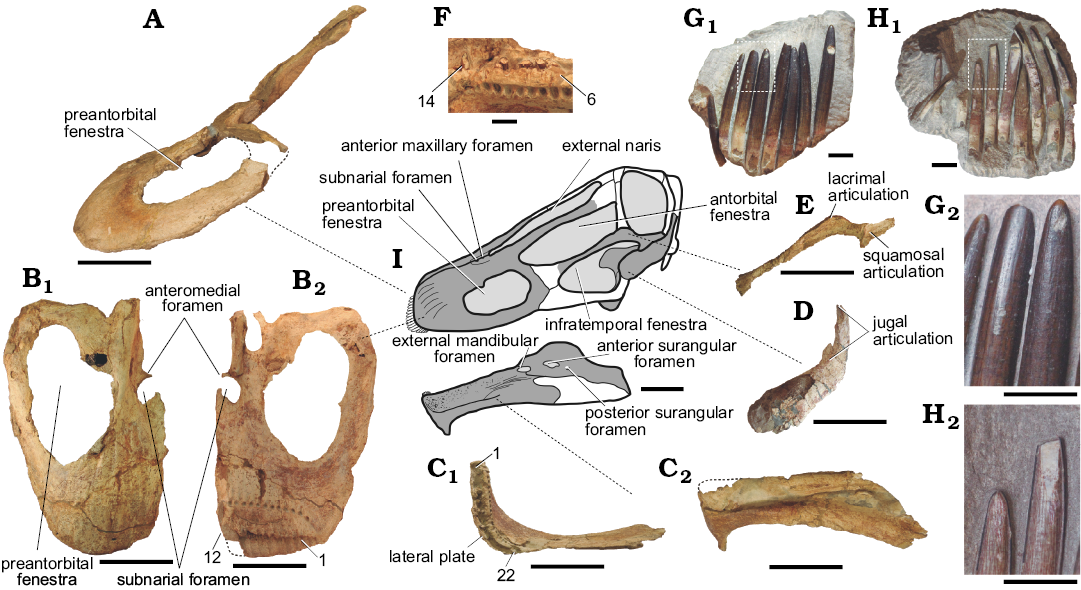
Fig. 3. Rebbachisaurid sauropod Lavocatisaurus agrioensis gen. et sp. nov. from Agrio del Medio (Argentina), Aptian–lower Albian. A. Left maxilla in lateral view. B. Right maxilla in anterodorsal (B1) and posteroventral (B2) views, numbers indicate the tooth positions. C. Left dentary in dorsal (C1) and lateral (C2) views, numbers indicate the tooth positions. D. Right squamosal (inverted) in lateral view. E. Right jugal (inverted) in lateral view. F. Dentary teeth, numbers indicate the tooth positions. G. Eight associated teeth in labial view (G1), close up of two teeth in labial view (G2), showing the absence of wear facets. H. Nine associated teeth in lingual view (H1), close up of the teeth in labial view (H2), showing the wear facets. I. Skeletal reconstruction, based on the holotype MOZ-Pv1232. Scale bars: A–E, 10 cm; F–H, 1 cm.
The spongy tissue of the bone is normal in all the bones of the skeleton. The cervical vertebrae are still embedded in the plaster with the right lateral side exposed. The cervical vertebrae are opisthocoelous and short (Fig. 2). The eighth cervical is the longest (26 cm), and otherwise the fourth to the tenth vary between 20–22 cm in length. The cervical centra have pleurocoels, which are apparently undivided as in Nigersaurus, and are teardrop-shaped, with the more excavated side in the anterior part. From the third cervical vertebra on, there is a keel in the ventral part of the centrum. The neural spine of the axis is anteriorly elongated and V-shaped as in Demandasaurus. The neural spine of the rest of the cervical vertebrae is simple and slightly displaced anteriorly. The cervical neural spines of Lavocatisaurus are relatively higher than in Nigersaurus and Demandasaurus (Sereno et al. 2007; Torcida Fernández-Baldor et al. 2011). The parapophysis is situated outside the centrum. The cervical vertebrae possess a well-developed system of laminae, as is characteristic of Eusauropoda (Wilson 2002). These include the anterior centrodiapophyseal, posterior centrodiapophyseal, spinoprezygapophyseal, spinopostzygapophyseal, prezygodiapophyseal and postzygodiapophyseal laminae (Fig. 2). The epipophyseal-prezygapophyseal lamina is well marked, although none of the cervical present a well-developed epipophysis, resembling the cervical vertebrae of other rebbachisaurids (e.g., Zapalsaurus, Limaysaurus; Salgado et al. 2006: fig. 4; Calvo and Salgado 1995: fig. 8B) and differing from Nigersaurus (Sereno et al. 2007: fig. 3B). In the axis, the centropostzygapophyseal laminae are united halfway along by a horizontal lamina, as in the third cervical of Zapalasaurus. The third cervical has postzygodiapophyseal and epipophyseal-prezygapophyseal laminae, as in Zapalasaurus (Salgado et al. 2006). In all the cervical vertebrae, the prezygapophyseal fossa is located between the prezygodiapophyseal and the centroprezygapophyseal laminae. The third cervical of Lavocatisaurus is similar in size to that of Zapalasaurus, but its laminae are more delicate, being in this character more similar to Nigersaurus than to Limaysaurus. The cervical rib has a well-marked anterior projection. The third and fourth cervical preserved their cervical ribs, which are as long as the centrum (Fig. 2), as in other diplodocoids (Wilson and Sereno 1998).
The centra of the caudal vertebrae are generally amphicoelous, although there are variations within the series. The anteriormost preserved vertebrae have centra with plano-convex anterior faces. The posterior face in these vertebrae is more concave than the anterior face. Both articular faces of the middle and posterior caudal vertebrae are concave. The distal caudal vertebrae have centra with convex articular faces. The caudal centra lack pleurocoels, with the exception of the most anterior one preserved, which has a small one. The anterior caudal vertebral centra have a ventral surface that is transversely concave, whereas in the middle caudal centra it is flat.
The neural spine in the caudal vertebrae is well developed, expanded anteroposteriorly, and slightly projecting posteriorly throughout the series. In the anterior ones, it is located in the middle part of the centrum, whereas in the middle and posterior ones it is located in the anterior part of the centrum as in other rebbachisaurids (e.g., Limaysaurus) but not as anteriorly as is in titanosaurs (Salgado et al. 1997; Wilson and Sereno 1998). The neural spine is rectangular in lateral view (Fig. 2). The posteriormost part of the neural spine in the middle caudal vertebrae is in the same plane as the posterior articular face. The transverse processes are dorsally directed in the anteriormost caudal vertebrae, becoming perpendicular to the centrum in the more posterior of the anterior caudals. The pedicles are low beneath the prezygapophyses. In the middle caudal vertebrae, the prezygapophyses are oriented nearly horizontally, projecting anteriorly in such a way that they protrude beyond the anterior articular face of the vertebra. The prespinal lamina is present in the anterior caudal vertebrae, which lack a hyposphene ridge and a triangular process. The distal vertebral centra are distinctively elongated.
The scapula is racket-shaped (Fig. 2). The scapular blade is perpendicular with respect to the orientation of the coracoid and shows a distal expansion. The scapular acromion is broad and well developed, and the dorsal face has a V-shaped concavity. The highest point of the dorsal margin of the scapular blade is higher than the dorsal margin of the proximal end. The ventral edge of the scapula does not show any sign of the ventral process observed in some diplodocids and titanosauriforms (Tschopp et al. 2015; Mannion et al. 2017). The cross section of the scapular blade at its base is D-shaped.
The humerus is crushed and badly preserved, but some anatomical traits can be observed. Compared to other sauropods, the humerus of Lavocatisaurus is relatively gracile (medium sensu Wilson and Upchurch 2003). The lateral margin of the proximal half of the bone is almost straight. Its proximolateral corner is pronounced, with the dorsal surface almost flat. The midshaft cross-section is elliptical. The deltopectoral attachment is relatively narrow and has a low crest throughout its length. The articular surface of the condyles is restricted to the distal portion of the humerus.
Remarks.—The scapula of Lavocatisaurus agrioensis gen. et sp. nov. shows the typical racket shape, which is one of the diagnostic characters of Rebbachisauridae (Bonaparte 1997; Carballido et al. 2010; Wilson and Allain 2015). The bones of the new species are comparable to most of the rebbachisaurid specimens from Neuquén Basin. Among the taxa that are closest in geographical terms, it is notable that the scapula is reasonably similar to that of Rayososaurus (Bonaparte 1997; Carballido et al. 2010), although it is distinguished by the development and orientation of the acromial process.
The deltopectoral crest of the humerus of Lavocatisaurus differs from Comahuesaurus in its position and lesser development. Particularly, the humeral articular head of the new species is more developed than in Comahuesaurus (Carballido et al. 2012).
Lavocatisaurus preserves the third cervical, as does Zapalasaurus. They are similar in size, but the one from Zapalasaurus is significantly more robust. The middle and posterior caudal vertebrae of Zapalasaurus have a neural spine that is anteroposteriorly elongated as in Lavocatisaurus. Nevertheless, the dorso-anterior corner of the neural spines of Zapalasaurus is located on a higher level than the posterior end, unlike in Lavocatisaurus, where the two corners are on the same level. The inclination of the prezygapophysis in the caudals of the new species is different from that of Zapalasaurus and is more similar to the more derived rebbachisaurids.
The holotype presented the cervical vertebrae and the first twenty vertebrae of the caudal series, articulated and arched, all of these together with the rest of the material scattered over an area of some 8 m2 (map of site in Salgado et al. 2012). The paratypic material was differentiated on the basis of its size, as well as by the fact that the neural arch of the only dorsal vertebra preserved was unfused, unlike in the adult specimen. The state of preservation of the bones is good, without signs of abrasion or breakages prior to burial; nor do they show signs of scavenging. Accordingly, the presence of a disarticulated cranium, articulated vertebral series, the low dispersion of the bones, and a low-energy sedimentary environment suggest that the preserved specimens were conserved in an autochthonous mode of concentration (Salgado et al. 2012).
Stratigraphic and geographic range.—Type locality and horizon only.
Phylogenetic analysis
The phylogenetic position of Lavocatisaurus agrioensis gen. et sp. nov. was examined through a cladistic analysis including the new taxon in a modified version of the data matrix recently used by Carballido et al. (2017). In addition to Lavocatisaurus, two recently described rebbachisaurids were included in the data matrix, Katepensaurus (Ibiricu et al. 2013, 2015) and Tataouinea, thus including all valid rebbachisaurid genera described to date, except for the fragmentary taxa Nopcsaspondylus (Apesteguía 2007) and Xenoposeidon (Taylor 2018). The resulting data matrix of 413 characters and 90 taxa was constructed using Mesquite V. 2.74 (Maddison and Maddison 2011) and exported into TNT format (see Supplementary Online Material available at http://app.pan.pl/SOM/app63-Canudo_etal_SOM.pdf). An equally weighted parsimony analysis was carried out using TNT v.1.5 (Goloboff and Catalano 2016). A heuristic tree search was performed starting from 1000 replicates of Wagner trees followed by TBR branch swapping and saving 10 trees per replicate. Under this procedure, we retrieved 180 most parsimonious trees (MPTs) found in 18 of the replicates and with a tree length of 1365 steps. The resulting trees were subjected to an additional round of branch swapping (TBR), obtaining 75 000 trees and collapsing the possible trees in memory. The resulting complete strict consensus tree recovered the rebbachisaurid lineage as completely collapsed (Fig. 4A; SOM), due to the multiple positions that Rayososaurus could take inside this clade (Fig. 4B). The same analysis was repeated excluding Rayososaurus and Lusotitan (which caused a large polytomy at the base of titanosauriforms) from the analysis, obtaining 9900 MPTs of 1363 steps. The strict consensus tree recovered is identical to the reduced consensus tree obtained once Rayososaurus is pruned from the analysis. The general topology recovered is fairly similar to that obtained in previous analyses (Carballido et al. 2012; Wilson and Allain 2015; Fanti et al. 2015). Lavocatisaurus was recovered as a relatively basal rebbachisaurid, being the sister lineage to Khebbashia (Figs. 4B, C, 5). Below we provide a short discussion of the principal nodes obtained in the analysis and the characters supporting them.
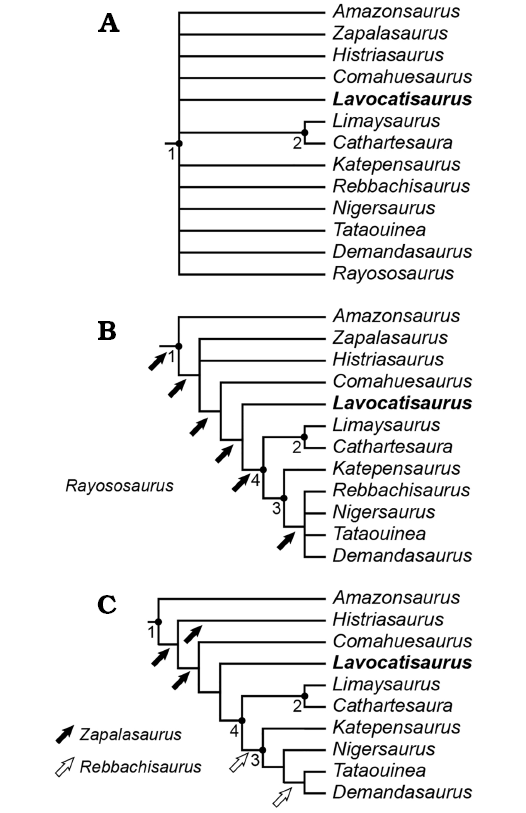
Fig. 4. Results of the phylogenetic analysis. A. Strict consensus tree of the 49 500 most parsimonious trees found, showing a complete polytomy amongst Rebbachisauridae with the sole exception of Limaysaurinae. B. Reduced consensus tree after pruning Rayososaurus, showing all possible positions of this taxon. C. Reduced consensus tree after pruning Rayososaurus, Zapalasaurus, and Rebbachisaurus (whose possible positions are indicated by the arrows). Resolved clades are indicated with numbers: 1, Rebbachisauridae; 2, Limaysaurinae; 3, Rebbachisaurinae; 4, Khebbashia.
Rebbachisauridae.—As in most previous analyses, Amazonsaurus was recovered at the base of Rebbachisauridae (e.g., Mannion et al. 2012; Carballido et al. 2012; Torcida Fernández-Baldor et al. 2011), a position slightly different from that suggested by some previous works (e.g., Carvalho et al. 2003; Salgado et al. 2006; Carballido et al. 2010). Rebbachisauridae are here supported by the same three synapomorphies recently obtained by Wilson and Allain (2015): character 241, absence of the hyposphenal ridge on anterior caudal vertebrae (reversed in Demandasaurus and Tataouinea); character 244, presence of spinodiapophyseal lamina (convergently present in Lognkosauria; Carballido et al. 2017); character 250, middle caudal vertebrae with a flat ventral surface; and character 413, middle caudal vertebrae with anterodorsally oriented prezygapophyses (reverted in Lavocatisaurus and more derived rebbachisaurids (MDR).
Histriasaurus–Zapalasaurus–MDR.—Histriasaurus, Zapalasaurus, and MDR were recovered with unresolved relationships at the base of the rebbachisaurids more derived than Amazonsaurus. This polytomy was caused by the three different positions that Zapalasaurus can take (Fig. 4C): as the sister taxon to Histriasaurus and MDR, as the sister taxon to Histriasaurus, or as a more derived rebbachisaurid (with Histriasaurus as the sister taxon of Zapalasaurus and MDR). A similarly unresolved relationship for these two taxa has been recovered in other recent analyses (Ibiricu et al. 2015; Fanti et al. 2015; Wilson and Allain 2015). Rebbachisauridae except for Amazonsaurus are supported by two derived characters, obtained as ambiguous synapomorphies due to the absence of information in Zapalasaurus or Histriasaurus: 198, festooned spinodiapophyseal laminae (unknown in Zapalasaurus); and character 23, dorsally directed transverse process (unknown in Histriasaurus).
Comahuesaurus + MDR.—As in Carballido et al. (2012), Comahuesaurus was recovered as a basal rebbachisaurid more derived than Histriasaurus and Zapalasaurus. Three characters (180, 231, 338) diagnose this clade, but these characters are unknown in Histriasaurus and/or Zapalasaurus, so they were recovered as ambiguous synapomorphies of the clade. The characters are: the reduced hyposphene in dorsal vertebrae (character 180); the posterior articular surface of anterior caudal vertebrae markedly more concave than the anterior one (character 231); and the ischium acetabular articulation narrower in its central part (character 338).
Lavocatisaurus + MDR.—Two synapomorphic characters support the clade formed by Lavocatisaurus and more derived rebbachisaurids: asymmetrical enamel with the lingual layer thinner than the labial one (character 117); and middle caudal vertebrae with anteriorly (nearly horizontally) projecting prezygapophysis (character 413).
Khebbashia.—This is a clade recently proposed by Fanti et al. (2015), which is formed by Limaysaurinae+Rebbachisaurinae. It is supported by three derived characters: 113, teeth with one high-angle and one low-angle wear facet; 118, teeth with lingual grooves; and 304, gracile humerus with robustness index (sensu Wilson and Upchurch 2003) lower than 0.27.
Limaysaurinae.—The clade Limaysaurinae is composed of two taxa, Limaysaurus and Cathartesaura, and supported by a broad scapular acromion (character 284).
Rebbachisaurinae.—Katepensaurus, the basalmost taxon of this clade, and Demandasaurus, Nigersaurus, Rebbachisaurus, and Tataouinea here constitute Rebbachisaurinae. This clade is supported by the presence of a deep centroprezygapophyseal fossa on middle and posterior dorsal vertebrae (character 409). The position of Katepensaurus within Rebbachisaurinae was recently recovered by Fanti et al. (2015) and contrasted with that originally proposed by Ibiricu et al. (2013) and subsequently recovered by Ibiricu et al. (2015). Indeed, some characters widespread in Rebbachisaurinae are also present in Katepensaurus, as previously noted by Ibiricu et al. (2013). Three further characters present in Katepensaurus are ambiguously optimized in this clade due to the absence of information. These are: the presence of a posterior centroparapophyseal lamina in dorsal vertebrae (character 194, unknown for Tataouinea); anterior caudal vertebrae with a triangular lateral process on the neural spine (character 235, absent in Rebbachisaurus; Wilson and Allain 2015); and the presence of a divided centroprezygapophyseal lamina in middle and posterior dorsal vertebrae (character 411, absent in Rebbachisaurus). The polytomy recovered for Rebbachisaurinae is caused by the two different positions that Rebbachisaurus can take within this group.
Cranial evolution in Rebbachisauridae
The amount of information recovered from the skull of Lavocatisaurus and its position amongst the basal rebbachisaurids provides the first opportunity to investigate the evolution of certain characters in one of the least-known lineages of Neosauropoda. Below, we list and briefly discuss the evolution of certain characters upon which the new taxon described here sheds further light.
Preantorbital fenestra.—The presence of a preantorbital fenestra was recognized as a derived character of Jobaria and more derived sauropods (Wilson and Sereno 1998; Wilson 2002). Amongst Neosauropoda it is a widespread character but with varying degrees of development. This fenestra is poorly developed in basal macronarians (e.g., Camarasaurus, Giraffatitan; Madsen et al. 1995; Janensch 1935), being similar in shape and size to the posterior maxillary foramen of basal sauropodomorphs (e.g., Plateosaurus; Sereno 2007). It differs from the posterior maxillary foramen in its anteroventral direction and in its connection to the antorbital fenestra (Wilson and Sereno 1998). Its presence in derived titanosaurs and diplodocids is much clearer, as the preantorbital fenestra is usually a well-developed opening in the maxilla (e.g., Diplodocus, Tapuiasaurus; Marsh 1884; Zaher et al. 2011). The maxillae here described possess a large opening that can be identified as the preantorbital fenestra. Although it resembles that of diplodocids and derived titanosaurs, in Lavocatisaurus the preantorbital fenestra is extremely developed, occupying a great area of the maxilla. As noted above, a similarly placed and apparently equally large fenestra was first identified as the antorbital fenestra in Nigersaurus (Sereno et al. 1999: fig. 2D; Sereno and Wilson 2005: fig. 5.5). To judge by the published images (Sereno et al. 1999; Sereno and Wilson 2005) and personal observation of the preserved Nigersaurus maxilla (MNN GDF512), the latter fenestra corresponds more probably to the preantorbital fenestra. Indeed, the anterior margin of the preantorbital fenestra is at the height of the subnarial foramen in Nigersaurus (Sereno et al. 1999: fig. 2D), whereas the antorbital fenestra, as reconstructed (Sereno and Wilson 2005; Sereno et al. 2007), is well above the foramen. Therefore, Lavocatisaurus and Nigersaurus share the presence of a large preantorbital fenestra, which occupies around half of the maxillary body. The preantorbital fenestra of both rebbachisaurids is not inset in any kind of fossa, differing from most flagellicaudatans known (e.g., Dicraeosaurus, Apatosaurus; see Tschopp et al. 2015).
Infratemporal fenestra. In basal sauropodomorphs, the infratemporal fenestra is posteriorly placed with respect to the orbit, being anteriorly delimited by the postorbital (anterodorsally) and jugal (anteroventrally), and posteriorly by the squamosal (posterodorsally) and the quadratojugal (posteroventrally) (e.g., Plateosaurus). A similar position and configuration of the infratemporal fenestra is observed in Camarasaurus and Europasaurus (Madsen et al. 1995; Marpmann et al. 2015), although in these taxa the fenestra is placed posteroventrally to the orbit, with the anterior margin close to the anterior margin of the orbit. In contrast, in most sauropods the squamosal–quadratojugal contact is missing, with the quadrate forming a small posterior edge of the infratemporal fenestra (see Wilson and Sereno 1998). This is observed in most basal sauropods and neosauropods (e.g., Giraffatitan, Diplodocus). In diplodocimorphs, the infratemporal fenestra projects anteriorly, its anterior margin going beyond that of the orbit (e.g., Diplodocus, Apatosaurus; Berman and McIntosh 1978). This anterior displacement of the infratemporal fenestra seems to be greater in rebbachisaurids, with that of Nigersaurus placed completely anteriorly to the orbit (Sereno et al. 2007). The anterior displacement of the infratemporal fenestra in rebbachisaurids seems to have had its origin early in their evolution, as most of the infratemporal fenestra is positioned anteriorly to the orbit in Lavocatisaurus, with its posterior margin slightly posterior to the anterior margin of the orbit (Fig. 3). Amongst rebbachisaurids, this is not the only morphological change in the configuration of the infratemporal fenestra. In Lavocatisaurus the jugal articulates with the squamosal, a novel articulation not previously described for any sauropodomorph, but that can also be detected in Limaysaurus and Nigersaurus (see description). Therefore, the infratemporal fenestra of rebbachisaurids is not delimited by the postorbital. In Lavocatisaurus this fenestra is certainly bounded by the jugal (anteriorly and posterodorsally) and the squamosal (posteriorly). The wide ventral end of the squamosal shows no signs of articulation with the quadratojugal, making it possible that the squamosal fails to contact the quadratojugal, as in Flagellicaudata (Tschopp et al. 2015). Nevertheless, a badly preserved articulation for the quadratojugal (ventrally) and the jugal (dorsally) can be detected in the preserved quadrate, making it impossible to discern whether the squamosal–quadratojugal contact was present or not. Based on the restoration of Nigersaurus, the ventral margin of the infratemporal fenestra should be bounded by the quadratojugal (not preserved either in Lavocatisaurus or in Nigersaurus).
Teeth and keratinized sheaths.—The well-preserved premaxillae, maxillae, dentaries and teeth of Lavocatisaurus yield an improved understanding of many aspects of the feeding adaptations in Rebbachisauridae. A great number of teeth were described for Nigersaurus, which has 58 upper teeth (4 premaxillary and 25 maxillary per side) and 68 lower teeth (34 in each dentary). In contrast, only 7 alveoli are present in the single dentary recovered for Demandasaurus (Torcida Fernandez-Baldor et al. 2011). In Lavocatisaurus we found 32 teeth in the upper series (4 in the premaxilla and 12 in the maxilla per side), and 44 teeth are present in the lower series (22 per dentary). The number of teeth in Lavocatisaurus is similar to that of non-neosauropod sauropods (e.g., 20 dentary teeth are present in Jobaria; Sereno et al. 1999), but differs from the number of lower tooth positions in other, non-rebbachisaurid diplodocimorphs and camarasauromorphs (Wilson and Sereno 1998). Basal rebbachisaurids thus seem to keep the plesiomorphic state (20 teeth or more per dentary), with a marked increment in tooth positions in Nigersaurus and a decrease in Demandasaurus.
All the preserved teeth in Lavocatisaurus (both upper and lower) have highly asymmetrical enamel, as in derived rebbachisaurids (Nigersaurus, Limaysaurus, Demandasaurus) and differing from the symmetrical enamel of the more basal rebbachisaurid Comahuesaurus (Salgado et al. 2004). This indicates that, whereas symmetrical enamel was present in basal forms, asymmetrical enamel evolved prior to the diversification of the clade into its two major lineages (Rebbachisaurinae and Limaysaurinae). As noted above, the upper teeth of Lavocatisaurus are much larger (both longer and wider) than those of the lower series. Although a similar pattern was mentioned for Nigersaurus (Sereno et al. 2007), in the case of Lavocatisaurus this difference is of an order such that the largest maxillary teeth are almost three times bigger than the dentary teeth (Fig. 3). The upper teeth present a low-angle wear facet on their lingual surface, but none of them has the second, labial, wear facet as described for Nigersaurus, although slightly polished surfaces are recognized on its tips. The lingual low-angle wear facet in Nigersaurus has been interpreted as being a result of tooth-tooth occlusion. The enormous differences in tooth size in Lavocatisaurus, especially close to the mid-point of the tooth series, make it less likely that the smooth and planar low-angle wear facets were the product of tooth-to-tooth occlusion. This is due to the fact that a single lower tooth cannot produce a single wear facet wider than its root. Therefore, the tooth-to-tooth abrasion producing this low-angle wear facet should be revised, at least in Lavocatisaurus, pending further studies on its micro-wear (see below). The second, labial high-angle facet present in Nigersaurus is also present in diplodocids and dicraeosaurids, and has been regarded as being a result of tooth-to-plant abrasion during ground-level browsing (Sereno et al. 2007). The absence of a labial, high-angle wear facet in Lavocatisaurus could indicate a different browsing height in basal rebbachisaurids or different movements when cutting plants, although another possibility is that it results from different types of food in diplodocoids. Whether or not the dentary teeth present similar facets to those of the upper teeth remains an open question calling for further and more complete discoveries.
It is widely accepted that sauropods lacked any masticatory adaptation, such as cheeks (Upchurch et al. 2007) or beaks (Sereno 2007). Nevertheless, the presence of a beak was suggested for Bonitasaura and other related taxa, based on the horizontal edge posterior to the tooth-bearing region (Apesteguía 2004; but see Wilson 2005). The presence of a keratinized sheath was suggested in Nigersaurus (Sereno et al. 2007) on the basis of the presence of numerous neurovascular grooves in the maxilla and dentary. The presence of a keratinous beak was recently proposed by Wiersma and Sander (2016) in Camarasaurus and other sauropods, based on the presence of exposed roots and extensive vascular foramina. For these authors, the presence of such a structure would explain the numerous ITRs reported for several sauropods; it would have served as extra protection from abrasive plants and provided a continuous cutting surface. The presence of ITRs and dentigerous bones in Lavocatisaurus is evidence against the consumption of bones by insects (Britt et al. 2008), reinforcing the taphonomic separation of the teeth as suggested by Wiersma and Sander (2016). As in Nigersaurus, the premaxilla and maxilla of Lavocatisaurus are laterally covered with numerous irregular grooves that are more numerous in the lower part of these bones. A similar pattern, though much more developed, can be observed in the dentaries. The dorsal margin of the dentaries has a depressed area that runs posteriorly from the whole lateral margin of the dentary ramus and ascends dorsally at the anterolateral margin. A similar, but less developed, morphology was described in Demandasaurus and suggested to be equivalent to the vascular canal shown in Nigersaurus by Sereno et al. (2007; see also Torcida Fernandez-Baldor et al. 2011). In Lavocatisaurus the laterodorsal surface of the dentary, above this marked step, is full of irregular, middle-to-large-sized pores and neurovascular foramina. Such morphology is frequently cited as evidence of keratin sheaths (see Sereno 2007). This region of the dentary coincides with the horizontally placed, anterior lateral edge of the maxilla (Fig. 3), which is sharp and covered with irregular grooves, both medially and laterally. Given this morphology, we do not rule out the presence of keratinous sheaths in the anteriormost edentulous region of the maxillae and dentaries. The presence of a keratinous sheath covering the anteriormost region of the snout could explain the ITRs of Lavocatisaurus (see Wiersma and Sander 2016), as well as the polished, single, low-angle wear facet of the upper teeth (see above). If this is the case, the lingual surface of the teeth, with thinner enamel, will have been subject to wear against a keratinous beak. Nevertheless, further analyses are needed in order to test this hypothesis further.
Paleobiogeographical remarks
Some interesting information of a palaeobiogeographical nature can be inferred from the calibrated phylogeny (Fig. 5). African forms such as Nigersaurus, Rebbachisaurus, and Tataouinea and the European Demandasaurus are grouped together in the clade Rebbachisaurinae. The presence in South America of Katepensaurus as a sister taxon to this clade could indicate an early evolution in South America and a subsequent dispersal of Rebbachisaurinae in Africa and Europe. However, the initial establishment of the clade in Africa and its dispersion in South America cannot be ruled out either, with subsequent vicariant evolution in both continents fostered by the opening of the South Atlantic and the definitive separation of Gondwana.
The taxa placed outside Khebbashia, such as Comahuesaurus, Zapalasaurus, and Lavocatisaurus (Fig. 5), could either be among the primitive rebbachisaurids prior to the dispersal to Africa or the first representatives that migrated to South America. This possibility cannot be ruled out since Histriasaurus from the Hauterivian of Croatia the oldest rebbachisaurid in the fossil record, is Gondwanan in palaeobiogeographical terms and could indicate an African origin of the rebbachisaurid lineage. However, the fragmentary nature of the Histriasaurus record could be distorting its real phylogenetic position. Another possible scenario, given the presence of phylogenetically more primitive taxa in South America, could be that Histriasaurus is a representative of a dispersion of primitive rebbachisaurids from South America to Africa and beyond at the beginning of the Cretaceous period, a dispersion for which we have no further information.

Fig. 5. Stratigraphically calibrated strict tree after pruning Lavocatisaurus, showing the geographical provenance of the taxa (silhouettes of continents) and ghost lineages (dashed lines). Abbreviations: AF, Africa, AS, Asia; EU, Europe; SA, South America.
Conclusions
Lavocatisaurus agrioensis is the first of the South American rebbachisaurids that preserves the rostral region of the cranium, which has allowed us to undertake a reliable enough reconstruction of the skull. The skull has a combination of morphological characters that justifies its definition as a new taxon within Rebbachisauridae along with the less derived rebbachisaurids from the Early Cretaceous of Argentina such as Zapalasaurus and Comahuesaurus. Our phylogenetic study situates Lavocatisaurus as the sister lineage of Khebbashia (Fig. 5). Lavocatisaurus is the most derived basal rebbachisaurid known to date, providing reliable information on the evolutionary steps that occurred just prior to the diversification of Rebbachisaurinae.
Acknowledgements
Verónica Díez Díaz (Museum für Naturkunde Berlin, Germany) in her review provided helpful comments that improved the manuscript. Rupert Glasgow (Zaragoza, Spain) translated the text into English. This paper forms part of the project CGL2017-85038-P and is subsidized by the Spanish Ministry of Science and Innovation, the European Regional Development Fund, the Museo Provincial de Ciencias Naturales “Prof. Dr. Juan Olsacher”, the Dirección Provincial de Minería de la Provincia del Neuquén, the CONICET (PIP-2014-2016 0683), the FONCyT (PICT 0668), and the Government of Aragón (“Grupos de referencia”).
References
Apesteguía, S. 2004. Bonitasaura salgadoi gen. et sp. nov.: a beaked sauropod from the Late Cretaceous of Patagonia. Naturwissenschaften 91: 493–497. Crossref
Apesteguía, S. 2007. The sauropod diversity of the La Amarga Formation (Barremian), Neuquén (Argentina). Gondwana Research 12: 533–546. Crossref
Berman, D.S. and McIntosh, J.S. 1978. Skull and relationships of the Upper Jurassic Sauropod Apatosaurus (Reptilia, Saurischia). Bulletin of Carnegie Museum of Natural History 8: 1–35.
Bonaparte, J.F. 1997. Rayososaurus agrioensis Bonaparte, 1995. Ameghiniana 34: 116.
Britt, B.B., Scheetz, R.D., and Dangerfield, A. 2008. A suite of dermestid beetle traces on dinosaur bone from the Upper Jurassic Morrison Formation, Wyoming, USA. Ichnos 15: 59–71. Crossref
Calvo, J.O. and Salgado, L. 1995. Rebbachisaurus tessonei sp. nov. a new Sauropoda from the Albian–Cenomanian of Argentina; new evidence on the origin of the Diplodocidae. Gaia 11: 13–33.
Canudo, J.I., Barco, J.L., Pereda-Suberbiola, X., Ruiz-Omeñaca, J.I., Salgado, L., Torcida Fernández-Baldor, F., and Gasulla, J.M. 2009. What Iberian dinosaurs reveal about the bridge said to exist between Gondwana and Laurasia in the Early Cretaceous. Bulletin de la Société Géologique de France 180: 5–11. Crossref
Canudo, J.I., Garrido, A., Carballido, J.L., Castanera, D., and Salgado, L. 2017. Icnitas de dinosaurios saurópodos en la Formación Rayoso (Cuenca Neuquina, Albiense, Argentina). Geogaceta 61: 43–46.
Carballido, J.L., Garrido, A.C., Canudo, J.I., and Salgado, L. 2010. Redescription of Rayososaurus agrioensis Bonaparte (Dinosauria, Diplodocoidea), a rebbachisaurid of the early Upper Cretaceous of Neuquén. Geobios 43: 493–502. Crossref
Carballido, J.L., Pol, D., Otero, A., Cerda, I.A., Salgado, L., Garrido, A.C., Ramezani, J., Cúneo, N.R., and Krause, J.M. 2017. A new giant titanosaur sheds light on body mass evolution among sauropod dinosaurs. Proceedings of the Royal Society B 284: 20171219. Crossref
Carballido, J.L., Salgado, L., Pol, D., Canudo, J.I., and Garrido, A. 2012. A new basal rebbachisaurid (Sauropoda, Diplodocoidea) from the Early Cretaceous of the Neuquén Group; evolution and biogeography of the group. Historical Biology 24: 631–654. Crossref
Carvalho, I.S., Santos Avilla, L., and Salgado, L. 2003. Amazonsaurus maranhensis gen. et sp. nov. (Sauropoda, Diplodocoidea) from the Lower Cretaceous (Aptian–Albian) of Brazil. Cretaceous Research 24: 697–713. Crossref
Dalla Vecchia, F. 1998. Remains of Sauropoda (Reptilia, Saurischia) in the Lower Cretaceous Upper Hauterivian/Lower Barremian) limestones of SW Istria (Croatia). Geologia Croatica 51: 105–134.
Díez Díaz, V., Tortosa, T., and Le Loeuff, J. 2013. Sauropod diversity in the Late Cretaceous of Southwestern Europe: The lessons of odontology. Annales de Paléontologie 99: 119–129. Crossref
Fanti, F., Cau, A., Cantelli, L., Hassine, M., and Auditore, M. 2015. New information on Tataouinea hannibalis from the Early Cretaceous of Tunisia and implications for the tempo and mode of rebbachisaurid sauropod evolution. Plos ONE 10: e0123475. Crossref
Fanti, F., Cau, A., Hassine, M., and Contessi, M. 2013. A new sauropod dinosaur from the Early Cretaceous of Tunisia with extreme avian-like pneumatization. Nature Communications 4: 2080. Crossref
Garrido, A. 2010. Estratigrafía del Grupo Neuquén, Cretácico Superior de la Cuenca Neuquina (Argentina): nueva propuesta de ordenamiento litoestratigráfico. Revista Museo Argentino de Ciencias Naturales, nueva serie 12: 121–177.
Gallina, P.A. and Apesteguía, S. 2005. Cathartesaura anaerobica gen. et sp. nov., a new rebbachisaurid (Dinosauria, Sauropoda) from the Huincul Formation (Upper Cretaceous), Río Negro, Argentina. Revista Museo Argentino de Ciencias Naturales, nueva serie 7: 153–166.
Goloboff, P.A. and Catalano, S.A. 2016. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32: 221–238. Crossref
Haluza, A., Canale, J.I., Otero, A., Pérez, L.M., and Scanferla, C.A. 2012. Changes in vertebral laminae across the cervicodorsal transition of a well-preserved rebbachisaurid (Dinosauria, Sauropoda) from the Cenomanian of Patagonia, Argentina. Journal of Vertebrate Paleontology 32: 219–224. Crossref
Ibiricu, L.M., Casal, G.A., Lamanna, M.C., Martínez, R.D., Harris, J.D., and Lacovara, K.J. 2012. The southermost record of Rebbachisauridae (Sauropoda: Diplodocoidea) from the early Late Cretaceous deposits in central Patagonia. Cretaceous Research 34: 210–232. Crossref
Ibiricu, L.M., Casal, G.A., Martínez, R.D., Lamanna, M.C., Luna, M., and Salgado, L. 2013. Katepensaurus goicoecheai, gen. et sp. nov., a Late Cretaceous rebbachisaurid (Sauropoda, Diplodocoidea) from Central Patagonia, Argentina. Journal of Vertebrate Paleontology 33: 1351–1366. Crossref
Ibiricu, L.M., Casal, G.A., Martínez, R.D., Lamanna, M.C., Luna, M., and Salgado, L. 2015. New material of Katepensaurus goicoecheai (Sauropoda: Diplodocoidea) and its significance for the morphology and evolution of Rebbachisauridae. Ameghiniana 52: 430–446. Crossref
Janensch, W. 1935. Die Schädel der Sauropoden Brachiosaurus, Barosaurus und Dicraeosaurus aus den Tendaguru-Schichten Deutsch-Ostafrikas. Palaeontographica (Supplement 7) 2: 147–298.
Lavocat, R. 1954. Sur les dinosauriens du continental intercalaire des Kem-Kem de la Daoura. Congrès Géologique International. Comptes Rendus de la Dix-neuvième Session 15: 65–68.
Leanza, H.A. 2003. Las sedimentitas huitrinianas y rayosianas (Cretácico inferior) en el ámbito central y meridional de la cuenca Neuquina, Argentina. Servicio Geológico Minero Argentino, Serie de Contribuciones Técnicas-Geología 2: 1–31.
Maddison, W.P. and Maddison, D.R. 2011. Mesquite: a modular system for evolutionary analysis. Version 2.75. http://mesquiteproject.org.
Madsen, J.H. Jr., McIntosh, J.S., and Berman, D.S. 1995. Skull and atlas-axis complex of the Upper Jurassic sauropod Camarasaurus Cope (Reptilia: Saurischia). Bulletin of the Carnegie Museum of Natural History 31: 1–115.
Mannion, P.D. 2009. A rebbachisaurid sauropod from the Lower Cretaceous of the Isle of Wight, England. Cretaceous Research 30: 521–526. Crossref
Mannion, P.D., Allain, R., and Moine O. 2017. The earliest known titanosauriform dinosaur and the evolution of Brachiosauridae. PeerJ 5: e3217. Crossref
Marsh, O.C. 1884. Principal characters of American Jurassic dinosaurs. Part VII. On the Diplodocidae, a new family of the Sauropoda. American Journal of Science. Third series XXVII 158: 161–167.
Marpmann, J.S., Carballido, J.L., Martin Sander. P., and Knötschke, N. 2015. Cranial anatomy of the Late Jurassic dwarf sauropod Europasaurus holgeri (Dinosauria, Camarasauromorpha): ontogenetic changes and size dimorphism. Journal of Systematic Palaeontology 13: 221–263. Crossref
Musacchio, E. and Vallati, P. 2000. La regresión del Barremiano–Aptiano en Bajada del Agrio, Neuquén (Argentina). IX Congreso Geológico Chileno, Actas, 230–234.
Pereda Suberbiola, X., Torcida, F., Izquierdo, L.A., Huerta, P., Montero, D., and Pérez, G. 2003. First rebbachisaurid dinosaur (Sauropoda, Diplodocoidea) from the early Cretaceous of Spain: palaeobiogeographical implications. Bulletin de la Société Géologique de France 174: 471–479. Crossref
Salgado, L., Coria, R.A., and Calvo, J.O. 1997. Evolution of titanosaurid sauropods. I: Phylogenetic analysis based on the postcranial evidence. Ameghiniana 34, 1, 3–32.
Salgado, L., Apesteguía, S., and Heredia, S.E. 2005. A new specimen of Neuquensaurus australis, a Late Cretaceous saltasaurine titanosaur from North Patagonia. Journal of Vertebrate Paleontology 25: 623–634. Crossref
Salgado, L., Canudo, J.I., Garrido, A.C., and Carballido, J.L. 2012. Evidence of gregrariousness in rebbachisaurids (Dinosauria, Sauropoda, Diplodocoidea) from the Early Cretaceous of Neuquén (Rayoso Formation), Patagonia, Argentina. Journal of Vertebrate Paleontology 32: 603–613. Crossref
Salgado, L., Garrido, A., Cocca, S., and Cocca, J.R. 2004. Lower Cretaceous rebbachisaurid sauropods from Cerro Aguada del León (Lohan Cura Formation), Neuquén Province, Northwestern Patagonia, Argentina. Journal of Vertebrate Paleontology 24: 903–912. Crossref
Salgado, L., Souza Carvalho, I. de, and Garrido, A.C. 2006. Zapalasaurus bonapartei, un nuevo dinosaurio saurópodo de la Formación La Amarga (Cretácico Inferior), noroeste de Patagonia, Provincia de Neuquén, Argentina. Geobios 39: 695–707. Crossref
Sereno, P.C. 2007. Basal Sauropodomorpha: historical and recent phylogenetic hipotheses, with comments on Ammosaurus major (Marsh, 1889). Special Papers in Paleontology 77: 261–289.
Sereno, P.C. and Wilson, J.A. 2005. Structure and evolution of a sauropod tooth batery. In: K.A. Curry Rogers and J.A. Wilson (eds.), The Sauropods: Evolution and Paleobiology, 157–177. University of California Press, Oakland.
Sereno, P.C., Beck, A.L., Dutheil, D.B., Larsson, H.C.E., Lyon, G.H., Moussa, B., Sadleir, R.W., Sidor, C.A., Varricchio, D.J., Wilson, G.P., and Wilson, J.A. 1999. Cretaceous sauropods from the Sahara and the uneven rate of skeletal evolution among dinosaurs. Science 286: 1342–1347. Crossref
Sereno, P.C., Wilson, J.A., Witmer, M., Whitlock, J.A., Maga, A., Ide, O., and Rowe, T.A. 2007. Structural extremes in a Cretaceous dinosaur. Plos ONE 2: e1230. Crossref
Taylor, M.P. 2018. Xenoposeidon is the earliest known rebbachisaurid sauropod dinosaur. PeerJ 6: e5212. Crossref
Torcida Fernández-Baldor, F., Canudo, J.I., Huerta, P., Montero, D., Pereda Suberbiola, X., and Salgado, L. 2011. Demandasaurus darwini, a new rebbachisaurid sauropod from the Early Cretaceous of the Iberian Peninsula. Acta Palaeontologica Polonica 56: 535–552. Crossref
Tschopp, E. and Mateus, O. 2017. Osteology of Galeamopus pabsti sp. nov. (Sauropoda: Diplodocoidea), with implications for neurocentral closure timing, and the cervico-dorsal transition in diplodocids. PeerJ 5: e3179. Crossref
Tschopp, E., Mateus, O., and Benson, R.B.J. 2015. A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda). PeerJ 3: e857. Crossref
Upchurch, P. 1998. The phylogenetic relationships of sauropod dinosaurs. Zoological Journal of the Linnean Society 124: 43–103. Crossref
Upchurch, P., Barrett, P.M., and Galton, P.M. 2007. A phylogenetic analysis of basal sauropodomorph relationships: implications for the origin of sauropod dinosaurs. Special Papers in Palaeontology 77: 57–90.
Wedel, M.J. 2003. Vertebral pneumaticity air sacs, and the physiology of sauropod dinosaurs. Paleobiology 29: 243–255. Crossref
Whitlock, J.A. 2011. A phylogenetic analysis of Diplodocoidea (Saurischia: Sauropoda). Zoological Journal of the Linnean Society 161: 872–915. Crossref
Whitlock, J.A., Wilson, J.A., and Lamanna, M.C. 2010. Description of a Nearly Complete Juvenile Skull of Diplodocus (Sauropoda: Diplodocoidea) from the Late Jurassic of North America. Journal of Vertebrate Paleontology 30: 442–457. Crossref
Wiersma, K. and Sander, M. 2016. The dentition of a well-preserved specimen of Camarasaurus sp.: implications for function, tooth replacement, soft part reconstruction, and food intake. Paläontologische Zeitschrift 91: 145–161. Crossref
Wilson, J.A. 1999. A nomenclature for vertebral laminae in sauropods and other saurischian dinosaurs. Journal of Vertebrate Paleontology 19: 639–653. Crossref
Wilson, J.A. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society 136: 215–275. Crossref
Wilson, J.A. and Allain, R. 2015. Osteology of Rebbachisaurus garasbae Lavocat, 1954, a diplodocoid (Dinosauria, Sauropoda) from the early Late Cretaceous-aged Kem Kem beds of southeastern Morocco. Journal of Vertebrate Paleontology 35 (4): e1000701. Crossref
Wilson, J.A. and Upchurch, P. 2003. A revision of Titanosaurus Lydekker (Dinosauria–Sauropoda), the first dinosaur genus with “gondwanan” distribution. Journal of Systematic Palaeontology 1: 125–160. Crossref
Wilson, J.A. and Sereno, P.C. 1998. Early evolution and higher-level phylogeny of sauropod dinosaurs. Society of Vertebrate Paleontology Memoir 5: i–vi, 1–68; Supplement 2 to Journal of Vertebrate Paleontology 18.
Wilson, J.A., D’Emic, M.D., Ikejiri, T., Moacdieh, E.M., and Whitlock, J.A. 2011. A nomenclature for vertebral fossae in sauropods and other saurischian dinosaurs. Plos ONE 6: e17114. Crossref
Zaher, H., Pol, D., Carvalho, A.B., Nascimento, P.M., Riccomini, C., Larson, P., Juarez-Valieri, R., Pires-Domingues, R., Jorge da Silva, N., and Almeida Campos, D. 2011. A complete skull of an Early Cretaceous sauropod and the evolution of advanced titanosaurians. Plos ONE 6: e16663. Crossref
Acta Palaeontol. Pol. 63 (4): 679–691, 2018
https://doi.org/10.4202/app.00524.2018