
A large marine eosauropterygian reptile with affinities to nothosauroid diapsids from the Early Triassic of British Columbia, Canada
TORSTEN M. SCHEYER, ANDREW G. NEUMAN, and DONALD B. BRINKMAN
Scheyer, T.M., Neuman, A.G., and Brinkman, D.B. 2019. A large marine eosauropterygian reptile with affinities to nothosauroid diapsids from the Early Triassic of British Columbia, Canada. Acta Palaeontologica Polonica 64 (4): 745–755.
Sauropterygia, one of the main clades of Mesozoic marine reptiles, diversified shortly after the Permo-Triassic biotic crisis and afterwards remained one of the major components of Early Triassic and later Mesozoic marine ecosystems. On the other hand, actual specimens of marine reptiles of Olenekian age are still rare in the fossil record, coming only from a few localities worldwide. Here we describe associated remains of a larger marine reptile of around 4 m body length, with nothosauroidean affinities from the Sulphur Mountain Formation exposed at the L cirque locality of Wapiti Lake area in British Columbia. Although the specimen records only scattered parts of the posterior vertebral column, some gastral ribs and most notably, the proximal portion of one hind limb together with a fan-shaped ischium, it represents one of the oldest records of Sauropterygia and larger representatives of aff. Nothosauroidea specifically, as well as the northernmost occurrence of such animals in the Triassic. As such, the new specimen is important for understanding the biogeography and early evolution of the group and that of Sauropterygia, in general.
Key words: Reptilia, Eosauropterygia, paleobiogeography, biotic recovery, apex predator, Olenekian, British Columbia, Ganoid Ridge.
Torsten M. Scheyer [tscheyer@pim.uzh.ch], Universität Zürich, Paläontologisches Institut und Museum, Karl Schmid-Strasse 4, CH-8006 Zürich, Switzerland.
Andrew G. Neuman [Andrew.Neuman@gov.ab.ca] and Donald B. Brinkman [don.brinkman@gov.ab.ca], Royal Tyrrell Museum of Palaeontology, Box 7500, Drumheller, Alberta, T0J 0Y0, Canada.
Received 17 January 2019, accepted 3 May 2019, available online 3 September 2019.
Copyright © 2019 T.M. Scheyer et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Early and early Middle Triassic are key periods for understanding the recovery phase of global ecosystems following the largest mass extinction in Earth’s history, the End-Permian Extinction phase. The late Early Triassic records a turnover of ocean vertebrate communities, induced by the invasion and habitation of oceanic environments by several lineages of reptiles, including the Ichthyosauromorpha (the fish-shaped ichthyosaurs and hupehsuchians), the Thalattosauriformes and the Sauropterygia (Scheyer et al. 2014). Of those, Sauropterygia is the most long-lived, speciose, and perhaps morphologically most diverse clade (Bardet et al. 2014). Among sauropterygians, the different clades have specialized on a variety of feeding strategies, with Nothosauroidea including some of the largest predatory forms that roamed the Triassic oceans. Together with Pachypleurosauroidea and Pistosauroidea they form the clade Eosauropterygia, which is the sister taxon to Placodontiformes, the latter including the durophagous placodonts (Rieppel 2000; Neenan et al. 2013). Nothosauroidea ingroup relationships are still far from resolved, although there is indication that Simosaurus Meyer, 1842 is sister to all remaining nothosaurs, followed by Germanosaurus Nopcsa, 1928, and the remaining genera Nothosaurus Münster, 1834 and Lariosaurus Curioni, 1847 are sister taxa (e.g., Münster 1834; Meyer 1842; Curioni 1847; Nopcsa 1928; Neenan et al. 2013; Jiang et al. 2014; Li et al. 2014). In a recent analysis focusing on Nothosauria only, Liu et al. (2014) found a less clear separation of the latter two highly nested nothosaurs leading to a mixture between Nothosaurus and Lariosaurus species, because many new Chinese taxa were indicated to share morphological characters intermediate between the two genera. Other sauropterygian taxa such as Corosaurus Case, 1936 from the Spathian (late Early Triassic) Alcova Sandstone, Wyoming, USA, and Cymatosaurus Fritsch, 1894 from the Anisian (Middle Triassic) of Europe and ?Israel also constitute large marine predators (Fritsch 1894; Case 1936; Rieppel 1997, 1998). These taxa are not considered as part of Nothosauria sensu stricto herein, because their relationship to nothosaurs and pistosaurs remains unclear (e.g., Storrs 1991; Rieppel 1998; Neenan et al. 2013; Sato et al. 2014; Scheyer et al. 2017; Jiang et al. 2019).
Biogeographically, nothosaurs are now well known both from the western (present-day Europe) and eastern Paleotethys (present-day China) with more than 20 described species in total (e.g., Rieppel and Wild 1996; Rieppel and Hagdorn 1997; Hagdorn and Rieppel 1998; Li et al. 2002; Li and Rieppel 2004; Jiang et al. 2006; Klein and Albers 2009; Liu et al. 2014). Other non-plesiosaur Triassic sauropterygians also show a Paleotethys-wide distribution, whereas, with the exception of the aforementioned Early Triassic Corosaurus and the Middle Triassic Augustasaurus from Nevada, USA (east Panthalassa, modern-day east Pacific), the record of Triassic Sauropterygia in western North America is extremely sparse (Neenan et al. 2013; Scheyer et al. 2014; Bardet et al. 2014; Kelley et al. 2014).
Here we report on fragmentary postcranial remains of a large reptile taxon with affinities to Nothosauroidea from the Triassic Sulphur Mountain Formation of the Wapiti Lake Area, British Columbia, Canada. Although the specimen was recovered as float, there are compelling indications for an Early Triassic age of the specimen. Irrespective of its geological age, this is the first recognition of Sauropterygia from the northern parts of the Panthalassa province and the northernmost occurrence of the group during the Early–Middle Triassic. We briefly discuss the paleobiogeography of the find and its implications for the timing of the recovery phase following the End-Permian mass extinction.
Institutional abbreviations.—PIMUZ, Palaeontological Institute and Museum, University of Zurich, Switzerland; TMP, Royal Tyrrell Museum of Palaeontology, Drumheller, Canada; SMF, Senckenberg Naturmuseum Frankfurt, Frankfurt am Main, Germany.
Material and methods
The specimen TMP 1995.116.0023 was found in the Sulphur Mountain Formation at Ganoid Ridge, Wapiti Lake area, British Columbia (Fig. 1) and is curated at the Royal Tyrrell Museum of Palaeontology. It was studied with a stereomicroscope and a 10 × hand lens. Images of the specimen were taken with a Nikon D90 SLR digital camera and processed using Adobe Creative Suite. Measurements were taken with steel calipers and, due to the weathered state of the bones, rounded to the nearest mm. All bone (or imprint) measurements are compiled in Table 1.
Table 1. Measurements (in mm) of TMP 1995.116.0023 tentatively identified as Nothosauroidea and PIMUZ T 4829 (= holotype of Paranothosaurus amsleri Peyer, 1939, 385 cm in total body length) of Nothosaurus giganteus for comparison based on measurements from Peyer (1939). For the ratios in the latter, the mean of left and right femur were taken. * Note that the femur of TMP 1995.116.0023 experienced compaction, which likely causes the bone to appear wider and more robust as in during life; we thus refrain from providing ratios for the specimen.
|
Measurement |
TMP 1995.116.0023 (left) |
PIMUZ T 4829 (left / right) |
|
Femur |
||
|
maximum proximodistal length |
238 |
260 / 260 |
|
proximal maximum expansion |
72* |
54 / 57 |
|
distal maximum expansion |
95* |
52 / 54 |
|
minimum shaft midline diameter |
57* |
29 / 29 |
|
ratio total length / prox. expansion |
|
4.68 |
|
ratio total length / dist. expansion |
|
4.91 |
|
Tibia |
||
|
proximal maximum expansion |
61 |
41 / 37 |
|
Width of femoral articular surface |
40 |
31 / 30 |
|
Fibula |
||
|
proximal maximum expansion |
56 |
34 / 30 |
|
Hemal arches (chevron bones) median |
||
|
maximum height as preserved |
82–91 |
|
|
maximum width as preserved |
53 |
|
Geological setting
Many fishes and marine reptile specimens are recovered as float from the Sulphur Mountain Formation at Ganoid Ridge and thus lack good stratigraphic control. It is thus often difficult to elucidate if they are Early Triassic or Middle Triassic in age. In the case of TMP 1995.116.0023, the position of the find at the L cirque locality (54°30’32” N; 120°41’24” W) of Ganoid Ridge, SSE of Wapiti Lake, is certain (Fig. 1; see Orchard and Zonneveld 2009 for an overview of the area and the lithology). In the unpublished field notes of one of us (AN), TMP 1995.116.0023 was recorded to have been found in a talus slope above the “marker bed” and below the Whistler Member (Middle Triassic) of the Sulphur Mountain Formation, together with other slabs containing bivalves (“Posidonia”, now referred to Peribositra; Orchard and Zonneveld 2009), fish remains, and ichthyosaur material assignable to Utatsusaurus. The majority of this talus slope appears to have derived from a big notch in the Vega-Phroso Member; the “marker bed” corresponding to the Meosin Mountain Member, which in certain outcrop areas in western Canada (including the Wapiti Lake area) separates the Phroso Member (Griesbachian to middle Smithian) from the Vega Member (middle Smithian and Spathian) (Orchard and Zonneveld 2009).

Fig. 1. 3D landscape images of Ganoid Ridge, Wapiti Lake area, British Columbia [downloaded from Google Earth Pro on October 3rd 2018; imagery Landsat/Copernicus (09/14/2011), ©2018 DigitalGlobe, ©2018 Province of British Columbia]. A. Overview of the complete Ganoid Ridge (reversed). White letters indicate different cirques at Ganoid Ridge. The stippled white line indicates the trend of the major anticline in the area (adopted from Mutter 2004: fig. 1a and references therein). B. Close-up of the southeastern cirques of Ganoid Ridge seen from the North. The anticline is well visible in the grey sediments of the Permian Mowitch Formation in the E and D cirques, surrounded by the darker brownish colored sediments of the Sulphur Mountain Formation. C. Close-up of the fossil site (54°30’32” N; 120°41’24” W) indicated by the white star in the talus slop of the L cirque, seen from the Northeast. Note that well exposed layers in the E cirque can be traced into the L cirque, indicating a similar stratigraphic succession in both cirques. Abbreviations: MM, Meosin Mountain Member; MO, Mowitch Formation; P, Phroso Member, V, Vega Member.
Abundant and well preserved fish faunas have been described from several layers in the Early Triassic succession from other cirques at Ganoid ridge (e.g., Schaeffer and Mangus 1976; Neuman 1986; Mutter 2004; Mutter and Neuman 2008, 2009; Anderson and Woods 2013; see also Romano et al. 2016), especially from the upper part of the Phroso Member (Orchard and Zonneveld 2009). Other specimens recovered from the L cirque locality include for example the osteichthyan Bobastrania canadensis (e.g., TMP. 1995.116.0038), Boreosomus sp. (e.g., TMP 1995.116.0007), Saurichthys sp. (e.g., TMP 1995.116.0036), and Coelacanthidae indet. (e.g., TMP 1995.116.0004). It is therefore likely that the fishes from the L cirque derive from beds of similar age as TMP 1995.116.0023.
It is further noteworthy that the Anisian Whistler Member of Ganoid Ridge, which occurs for example at the more northwestern D and T cirques, does not contribute to the talus slopes at the L and E cirques in the southeastern part (see also Orchard and Zonneveld 2009; according to John-Paul Zonneveld, personal communication 2018, both, the Whistler Member and the upper Vega Member do not appear to contribute to the L cirque talus slopes). It seems thus likely that TMP 1995.116.0023 might represent a rare find of an Early Triassic marine reptile of Olenekian (Smithian or Spathian) age. Unfortunately, no conodonts could be recovered from the matrix slab that would further constrict the stratigraphic age of the find. In either case, the depositional environment of TMP 1995.116.0023 is similar for both, the Vega and Phroso members and the Whistler Member, as they comprise fine siliciclastic sediments (silts and shales) deposited below the storm wave base on the distal shelf during a first transgressive-regressive cycle (Edwards et al. 1994) that unconformably overlies the Permian Mowitch Formation (Orchard and Zonneveld 2009).
Systematic paleontology
Sauropterygia Owen, 1860
Eusauropterygia Tschanz, 1989
Nothosauroidea Huene, 1956
aff. Nothosauroidea gen. et sp. indet.
Fig. 2.
Material.—TMP 1995.116.0023, consisting of four slabs of siltstone matrix (two large and two small ones numbered 1 to 4 (Fig. 2; SOM 1, Supplementary Online Material available at http://app.pan.pl/SOM/app64-Scheyer_etal_SOM.pdf), on which mostly imprints of partially disarticulated large skeletal remains are present. The remains include the proximal portion of a hind limb and the ischium as part of the pelvic girdle in articulation, on which we mainly base our taxonomic assignment. TMP 1995.116.0023 was recovered on August 9th in 1995 during a National Geographic field trip led by the late Dr. Elisabeth (Betsy) Nicholls, and Donald Henderson, as well as one of the authors (AN) of the Royal Tyrrell Museum of Palaeontology. According to the unpublished field notes of Elisabeth Nicholls, the bones were first identified in the field as the forelimb remains of a large plesiosaur and only later identified as hind limb remains of “Nothosauridae indet.” in the TMP. The material was probably referred to by (Rieppel 1999) when noting unpublished fragmentary Sauropterygia material as personal communication by Elisabeth Nicholls.

Fig. 2. TMP 1995.116.0023 tentatively identified as aff. Nothosauroidea indet. from the Olenekian, L cirque locality, Ganoid Ridge, Wapiti Lake area, British Columbia, Canada. The specimen consists of two associated larger slabs (A), and two small slabs (B, C) of unclear association. The area where the preserved bones and imprints on the large slab derive from the skeleton is indicated by a red rectangle on black outline drawing based roughly on Nothosaurus giganteus (based on PIMUZ T 4829). A. The two larger slabs containing most of the identifiable bones. Gastralia, appendicular, and girdle elements are in light gray, axial elements in darker gray. B. Small slab with the imprint of two bone fragments, the larger of which might pertain to a girdle or appendicular bone. C. A second small slab with three partially preserved bone remains, probably all representing adjacent neural arch fragments. Photographs (A1–C1) and interpretative drawings (A2–C2). Abbreviations: c, centrum; ch, hemal arch or chevron bone; g, gastral elements; na, neural arch.
Description.—TMP 1995.116.0023 comprises a mixture of few bone fragments, as well as imprints clearly indicating bone outlines on four bone slabs. Given the size of the elements and that none of the non-axial elements appears twice, we assume that all pertain to a single large individual. The two larger slabs of siltstone matrix fit easily together, whereas the two smaller slabs did not show continuity with the larger slabs. The skeletal remains on the larger slabs include an isolated cluster of six lateral gastral elements (whether from the right or left side of the body cannot be elucidated), a few strongly weathered Y-shaped elements that most likely represent hemal arches or chevron bones, as well as few smaller (caudal?) vertebrae and neural arches, and most importantly, three hind limb bones (stylopod and zeugopod) and one ischium. On one of the smaller slabs, possibly one caudal transverse process is present.
The hind limb remains are represented by the clear impression of a left femur (whose preaxial border is partially not preserved proximally) and the proximal parts of a tibia and fibula in ventral view. The femur has a slightly sigmoidally curved shaft with a postaxial concave border and preaxial straight to slightly convex margin. It also has an overall massive appearance as indicated by its length and width measurements (Table 1). Because it is mostly preserved as imprint, there are only few additional anatomical features visible, such as the internal trochanter forming a prominent crista postaxially. The zeugopodial elements meet proximally and form a larger spatium interosseum, the dimensions of which cannot be measured. Proximally the tibia has an angled (somewhat rounded) facet, whereas the fibula shows a single straight facet for articulation with the femur. The well-developed distal facets of the femur left behind distinct imprints, forming a broad angle of 128°.
In addition, the partial impression of one large pelvic girdle element, identified herein as an ischium, is present. The bone impression as preserved shows a strong constriction with a steep angle between the proximal acetabular region and the flaring fan-shaped ventral blade.
The gastral rib elements all lie closely spaced to each other and resemble straight elongated and slightly conical bones, representing the lateral-most elements of six gastralia. Unfortunately, none of them is complete enough to allow total length measurements, but the longest imprint of a single element extends over 148 mm.
Several Y-shaped bones, interpreted herein as hemal arches or chevron bones of proximal caudal vertebrae, lie scattered on the stone slabs in various states of preservation and orientation. All bones show different stages of breakage, with most consisting of a mixture of weathered bluish bone matrix flaking off to leave behind only imprints in the sediment matrix. Alas, the imprints are usually incomplete, which makes it difficult to trace their shape accurately. The length of the more complete bones ranges between 75 and 90 mm. These bones are not identified as neural arches and spines however, which in some nothosaurs such as N. haasi (Rieppel et al. 1999) can also be quite high, because of a complete lack of ventral structures that would articulate with the caudal centra forming the lateral walls of the neural canal. Instead the bones show laterally projected bases which would articulate with the ventral surface of the caudal centra.
Four bone imprints are tentatively identified as belonging to axial elements, including one isolated neural arch, which is embedded close to the gastral elements and three vertebrae associated with remnants of neural arches. Other smaller incomplete imprints and patches of bone fragments are scattered among the Y-shaped hemal arches/chevron bones, but they are too poorly preserved to be positively identified.
The smaller slabs of rock also show bone impressions with little remains of bone tissue still attached. Most of the imprints cannot be associated with any recognizable bone, with the possible exception of one imprint. This imprint shows a weak striation of the bone surface and a slight constriction separable from an expanded area, thus resembling the distal part of a caudal transverse process (compare to Peyer 1939: pl. 70).
Remarks.—TMP 1995.116.0023 derives from a large Triassic marine reptile that has the following character combination: femur robust, with proximal head expanded and distal epiphysis distinctly expanded; distal condylar facets are well developed and defined, forming a broad angle; ischium hatchet-shaped with strong constriction between acetabular region and wide, fan-shaped ventral blade; tibia and fibula form distal spatium interosseum; tibia with angled proximal facet; fibula with straight proximal facet; both tibia and fibula proximally similarly expanded; lateral gastral elements straight.
Given the enormous femoral size of more than 230 mm in length, there are only a few Triassic marine reptiles that reach similar dimensions, those being representatives of Ichthyopterygia among Ichthyosauromorpha, as well as Nothosauria and Pistosauroidea among Sauropterygia. This would already preclude an assignment of the new form to small-sized sauropterygian pachypleurosaurs or heavily armed forms such as placodonts or saurosphargids (e.g., Sinosaurophargis). However, size and age of the fossil alone are not deemed valid characters on their own to separate taxa. Although we identified TMP 1995.116.0023 as pertaining to Sauropterygia with affinities to Nothosauroidea based on the above mentioned characteristics, we performed an osteological comparison (see Appendix 1) also with other marine reptiles from the Triassic, including (i) protorosaurs, (ii) saurosphargids, (iii) thalattosaurs, (iv) ichthyosauromorphs, (v) placodonts and Helveticosaurus.
Although TMP 1995.116.0023 does not show any of the synapomorphies of Nothosauridae as noted by Sues (1987; three pertaining to the cranium; one to the humerus, and one to the intermedium in the forelimb), it nevertheless closely resembles the hind limb and ischium anatomy (and size) described for one of the largest species of Nothosaurus, N. giganteus (which includes the holotype PIMUZ T 4829 of the so far largest and most complete, articulated nothosaur “Paranothosaurus amsleri”, from the Besano Formation, Monte San Giorgio, Ticino, southern Switzerland; Peyer 1931; Rieppel 2000). The ratios of femoral epiphyseal expansion, however, differ between the new specimen and the Middle Triassic N. giganteus. The lateral gastral elements and the axial elements preserved in TMP 1995.116.0023 are morphologically in accordance with those described for Nothosaurus (Peyer 1939; Rieppel and Wild 1996), with the former bones being densely packed and straight, rod-like elements tapering into a single tip laterally, and the latter bones having moderately elongated and not strongly anteroposteriorly expanded neural arches and Y-shaped hemal arches.
Furthermore, larger specimens of Ceresiosaurus–Lariosaurus (e.g., C. calcagnii PIMUZ T 4836, C. lanzi PIMUZ T 2464; Hänni 2004; see also Rieppel 2000 and Liu et al. 2014 for discussion on the taxonomic status of Ceresiosaurus and Lariosaurus) show also similar ischium, femur, tibia, and fibula shapes (SOM 2).
TMP 1995.116.0023 resembles to some degree also other sauropterygians, but it also shows distinctive features. As such, the new specimen differs from the non-nothosauroid eosauropterygian Majiashanosaurus discocoracoidis from the Olenekian (Spathian) of China in overall body size (the partial skeleton of the holotype of M. discocoracoidis is less than 50 cm long) and in the shape of the ischium and proximal hind limb shape (Jiang et al. 2014). The ischium of M. discocoracoidis is of overall plate-like shape with a weakly concave anterior margin; its femur impression indicates a straight bone, which was expanded proximally and distally, and straight tibia and fibula (Jiang et al. 2014).
TMP 1995.116.0023 differs from Simosaurus gaillardoti in that the femora of the latter are all proximally slightly more expanded than distally (Rieppel 1994), and Sues (1987) noted that the bones of S. gaillardoti are slender. The shapes of the articulation facets of the femur, tibia and fibula of Simosaurus gaillardoti do not match those of TMP 1995.116.0023, but the ischium does exhibit the strong constriction seen in the latter.
Furthermore, TMP 1995.116.0023 differs from Corosaurus alcovensis, the so far oldest and only eosauropterygian taxon from the Early Triassic of the North American continent described, in having a much more robust but less sigmoidally curved femur (although that could be assigned to taphonomic alteration of the bone), as well as a more acute constriction in the ischium (Zangerl 1963; Storrs 1991; Rieppel 1998). Storrs (1991: 36–38) further noted that the “distal articular face of the femur is roughly semicircular [in which] the two equisized tibial condyles are reduced relative to the primitive condition but remain distinct. There is, however, no intercondylar fossa, although ventrally a shallowly depressed popliteal space exists. No clear fibular facet is present” in Corosaurus. The lack of a fibular facet is in clear contrast to the morphology of TMP 1995.116.0023.
The new specimen differs also from the non-plesiosaur pistosauroid eosauropterygians Pistosaurus, Yunguisaurus, and Wangosaurus in the general shape of the femur and the ischium (Geissler 1895; Sues 1987; Sato et al. 2014; Ma et al. 2015). For the Pistosaurus sp. femur (SMF R4041c), Sues (1987: 117) noted that the “proximal articular end is much more robust than the distal one”, whereas the ischium (SMF R76) was shown to be less constricted than in TMP 1995.116.0023. The tibia and fibula are only insufficiently known on block SMF R4041b of Pistosaurus. Yunguisaurus liae and Wangosaurus brevirostris, on the other hand, have similarly robust zeugopodial and stylopodial elements (Sato et al. 2010, 2014; Ma et al. 2015), and in Yunguisaurus, also a fan-shaped ischium is present (not known in Wangosaurus). The shaft of the ischium, however, is wide and the blade less flaring than in TMP 1995.116.0023. The distal articulation of the femora and the proximal articulation facets of the tibiae and fibulae in Yunguisaurus are distinctly different from those seen in TMP 1995.116.0023. The Canadian specimen lacks the proximal broadening of the tibia visible in Yunguisaurus liae, resembling more the condition seen in nothosaurs and in Wangosaurus. The femur of Wangosaurus is distinctly sigmoidally curved, with a slender shaft and it appears to be more expanded proximally than at the distal end (Ma et al. 2015; TMS personal observation), thus showing a different shape to TMP 1995.116.0023. Finally, in Y. liae, the left and right hemapophyseal elements do not fuse to form Y-shaped chevron bones as are present in TMP 1995.116.0023 (Sato et al. 2014; the condition is not known in Pistosaurus and Wangosaurus).
The upper Anisian Augustasaurus from Nevada, USA, does not have hind limbs or the posterior axial skeleton preserved (Sander et al. 1997). TMP 1995.116.0023 differs from Bobosaurus forojuliensis, potential sister taxon to all remaining plesiosaurs (Dalla Vecchia 2006; Fabbri et al. 2014), in having lower neural arches in the sacro-caudal region and a more constricted ischium; a femur is not present for comparison in Bobosaurus. TMP 1995.116.0023 further differs from the oldest known plesiosaur Rhaeticosaurus mertensi from the Late Triassic of Germany (Wintrich et al. 2017) and more highly nested plesiosaurs such as Tetanectes laramiensis, Peloneustes philarchus, and Meyerasaurus victor by having more and thinner gastralia, a slightly sigmoidal femur with both proximal and distal epiphyses being expanded (e.g., Smith and Vincent 2010; Ketchum and Benson 2011; O’Keefe et al. 2011). Plesiosaur femora are usually straight and the distal epiphyses much broader than the proximal articular heads, the latter often not being expanded at all. Furthermore, the zeugopodial elements in TMP 1995.116.0023 appear to be much longer and not indicative of a plesiosaur-type paddle.
Discussion
Based on the comparisons described above, all observable anatomical details of the bones of TMP 1995.116.0023 are conform with non-pistosauroid nothosauroid eosauropterygians; although some disparity in proportions and the ratios of femoral epiphyseal expansions are present due to the robustness of the new specimen. In contrast, similarities to only some bones of certain pistosauroid taxa do exist (as nothosaurs and pistosaurs share some characteristics in their postcrania), but disparity among these taxa and TMP 1995.116.0023 is larger in comparison to the nothosauroids. We therefore tentatively assign TMP 1995.116.0023 to Sauropterygia aff. Nothosauroidea indet. We cannot completely rule out, however, the possibility that the Canadian taxon is a yet unknown form of non-nothosauroid eosauropterygian. In either case, the new taxon indicates that eosauropterygians were more widely spread in North America than previously known.
The femur of the specimen is slightly shorter than that of PIMUZ T 4829, one of the largest and almost complete specimens of Nothosaurus giganteus (see Table 1), a similar total body size of around 4 m, but much more robust body shape (based on the femoral proximal and distal expansion in comparison to the femur length) is proposed for the Canadian taxon. Assuming a predatory lifestyle as in all other known nothosauroids (e.g., Liu et al. 2014) it is reasonable to assume that the new Canadian taxon could have been an apex predator in the ancient ecosystem. A possible late Early Triassic age of the specimen TMP 1995.116.0023 would further indicate one of the oldest records of nothosauroid eosauropterygians, which was so far represented by material identifiable as N. marchicus from the early Anisian Jena Formation of Winterswijk, The Netherlands and Rüdersdorf in Germany (Albers 2005; Voeten et al. 2015), as well as by indeterminate remains of Nothosaurus sp. from the Röt Formation, lowermost Anisian, of Central Europe (Hagdorn and Rieppel 1998; Rieppel et al. 1999).
Among the largest representatives of nothosauroids, N. giganteus ranges between the upper Anisian (Illyrian) and Carnian (Julian) (Dalla Vecchia 1994; Rieppel and Wild 1996; Vickers-Rich et al. 1999; Kear et al. 2008; Kelley et al. 2014), whereas the largest Chinese nothosauroid material, Nothosaurus zhangi, is from the middle Anisian (Pelsonian; Liu et al. 2014) and thus slightly older. Furthermore, the so far oldest described eosauropterygian, Majiashanosaurus discocoracoidis, together with some of the basal-most members of ichthyosauriforms (Motani et al. 2014a), was recovered from the Upper Member of the Nanlinghu Formation (Spathian, Early Triassic) of Majiashan, Chaohu Province (Jiang et al. 2014), indicating the importance of that locality in southern China for marine reptile evolution. Accordingly, it was so far assumed that the center of origination of Nothosauroidea, and of Sauropterygia in general, was situated in the Paleotethys (Rieppel 1999); either in the western or in the eastern part of that ocean, although somewhere between these two borders remained an option as well. The presence of a potential nothosauroid of Olenekian age in Canada (Fig. 3), on the other hand, underscores the rapid dispersal of sauropterygians already during that time also along the western margin of Pangaea. This happened potentially shortly after the origination of the group, but based on the available data, the center of origination cannot be pinpointed with confidence. The current lack of a better phylogenetic understanding of the relationships of TMP 1995.116.0023 and its fragmentary nature render any further conclusions speculative. Its recovery nevertheless fits with the assessment of Scheyer et al. (2014) that late Early Triassic (and earliest Middle Triassic) marine tetrapod faunas are still undersampled, because localities are usually restricted to remote areas that are difficult to reach and are neither of the concentration nor the conservation deposit type (Konzentrat- and Konservat-Lagerstätten of Seilacher 1970; see also Allison 1988).
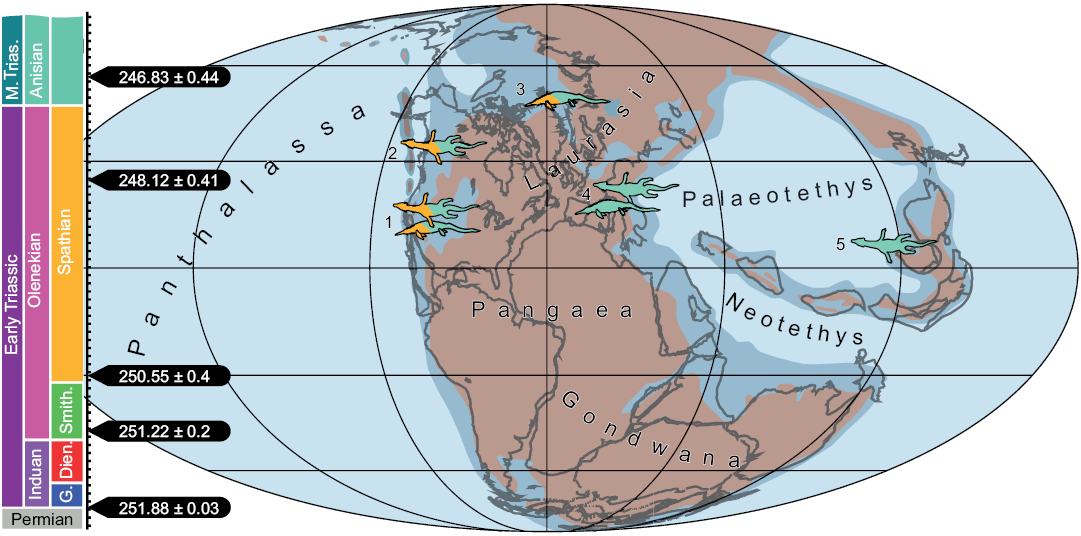
Fig. 3. Early Triassic map showing occurrences of Early Triassic (Olenekian) and Middle Triassic (Anisian) with potential apex predators in the respective marine ecosystems. Larger Eosauropterygia are represented by a nothosaur-shape and larger Ichthyopterygia by an ichthyosaur-shape. 1, Nevada and Idaho, USA: ichthyopterygian Thalattoarchon (Anisian), “Cymbospondylus”-like, shastasaur-like ichthyopterygians and Cymbospondylus spp. (Smithian/Spathian–Anisian); 2, British Columbia, Canada: Sauropterygia cf. Nothosauroidea indet. (TMP 1995.116.0023) (Smithian/Spathian–Anisian); 3, Svalbard: “Cymbospondylus” like ichthyopterygians or Pessopteryx? (Smithian/Spathian–Anisian); 4, Central Europe: Cymbospondylus buchseri; Nothosaurus giganteus (Anisian); 5, Luoping, southern China: Nothosaurus zhangi (Anisian). Please note that, although there are Spathian age eosauropterygians and ichthyosaurs known from Anhui Province, eastern China, these are not considered large predators herein, although they could still be apex-predators in their respective ecosystems (compare to Hansen and Galetti 2009). It is further noteworthy that the reptile record from the Smithian is virtually inexistent, with maybe the exception of a few isolated bones (e.g., Massare and Callaway 1994), which so far supports an early rapid dispersal of reptiles mainly during the Spathian. Data and map modified from Scheyer et al. 2014 and Romano et al. 2017, with additions by Fröbisch et al. 2013; Maxwell and Kear 2013; Liu et al. 2014. Abbreviations: Dien., Dienerian; G., Griesbachian; M. Trias., Middle Triassic; Smith., Smithian.
Large apex predators such as the macrophagous ichthyosaur Thalattoarchon saurophagis from Nevada, USA (Fröbisch et al. 2013) and the nothosaurid Nothosaurus zhangi from Luoping, southern China (Liu et al. 2014), have been used as indicators of recovery and ecosystem stabilization following the aftermath of the Permo-Triassic mass extinction event by the Anisian (Middle Triassic). A definition for gigantic apex predators was proposed by Liu et al. (2014), with the criterion being a minimum body length of 5 m. This categorization, which would include also very large individuals of Nothosaurus giganteus and N. zhangi, has merits as important indicators of ecosystem recovery and food web complexity on a global scale. Following a context-dependent approach similar to assessing missing megafaunal elements in Pleistocene-modern ecosystems (Hansen and Galetti 2009) on different-sized land masses, distinguishing between “normal-sized”, “large-sized”, and “gigantic” apex predators on local or regional scales might seem less important when analyzing global recovery dynamics. Accordingly, apex predators (also smaller ones with <5 m body length) were present in marine ecosystems throughout the Early Triassic, although with different faunal composition and community structures (Scheyer et al. 2014) and as such, the smaller sized eosauropterygian Majiashanosaurus might still have played the role of apex predator in its environment in which other faunal elements were also of smaller size.
Scheyer et al. (2014) indicated a transition from fish-amphibian to fish-reptile dominated faunas in marine ecosystems around the Smithian/Spathian boundary, about 2 Ma after the Permo-Triassic mass extinction. The appearance of a Canadian large reptile with nothosauroid affinities in the late Early Triassic thus adds another piece to the puzzle of the timing of the recovery phase following the extinction event. Similar to the case of Thalattoarchon saurophagis and Nothosaurus zhangi, TMP 1995.116.0023 could act as further indication for globally reestablishing complex marine ecosystems towards the end of the Early Triassic.
Rieppel (1999) noted that many of the smaller-sized Triassic sauropterygians probably lacked the capacity to cross open oceans; instead, dispersal occurred likely along the coastlines of Pangaea in these taxa. It is doubtful whether this would have been a hindrance for a larger animal of four or more meters in body length. However, as has been proposed for Early Triassic ichthyopterygians (Cuthbertson et al. 2013), pre-Olenekian intermittent shallow epi-continental seaways spanning Laurasia, thus connecting the western Paleotethys with the eastern Panthalassa faunal province, might have facilitated the dispersal of other marine reptile groups such as the nothosauroids as well.
Conclusions
The description of a new, large fossil with nothosauroid affinities from Canada adds data to our understanding of the early evolution of this important group of sauropterygian reptiles. It also serves as a reminder that the Early Triassic remains the key period to (i) understand the origins and paleogeography of Mesozoic marine reptiles (see also Jiang et al. 2014) in general, and (ii) reconstruct the timing of the oceanic recovery phase following the largest mass extinction event in Earth history. As such, the Early Triassic sites at Ganoid Ridge in British Columbia, Canada, deserve renewed attention in future studies pertaining to these research areas.
Acknowledgements
TMS and DBB devised the study and performed the anatomical analysis, AN provided data of the geological setting. TMS wrote a draft of the manuscript and prepared the figures, and all co-authors participated in the finalization of the manuscript. Brandon Strilisky and Jim Gardner (both TMP) are thanked for access to specimens and various help in the collections. We thank Jean-Paul Zonnefeld (University of Alberta, Edmonton, Canada), as well as Carlo Romano, Hugo Bucher, and Marcelo Sánchez (all PIMUZ) for discussions. Mike Orchard (Geological Survey of Canada, Vancouver, Canada) is thanked for analysing the matrix of the find for conodonts. We thank the editor and two anonymous reviewers for the constructive comments. The study was partly funded by the Swiss National Science Foundation (grant no. 205321_162775 to TMS).
References
Albers, P.C.H. 2005. A new specimen of Nothosaurus marchicus with features that relate the taxon to Nothosaurus winterswijkensis. PalArch Papers (Vertebrate Palaeontology) 3: 1–7.
Allison, P.A. 1988. Konservat-Lagerstätten: cause and classification. Paleobiology 14: 331–344. Crossref
Anderson, K. and Woods, A.D. 2013. Taphonomy of Early Triassic fish fossils of the Vega-Phroso Siltstone Member of the Sulphur Mountain Formation near Wapiti Lake, British Columbia, Canada. Journal of Palaeogeography 2: 321–343.
Bardet, N., Falconnet, J., Fischer, V., Houssaye, A., Jouve, S., Pereda Suberbiola, X., Pérez-García, A., Rage, J.-C., and Vincent, P. 2014. Mesozoic marine reptile palaeobiogeography in response to drifting plates. Gondwana Research 26: 869–887. Crossref
Callaway, J.M. 1997. A new look at Mixosaurus. In: J.M. Callaway, and E.L. Nicholls (eds.), Ancient Marine Reptiles, 45–59. Academic Press, San Diego. Crossref
Case, E.C. 1936. A nothosaur from the Triassic of Wyoming. Contributions from the Museum of Paleontology, University of Michigan 5: 1–36.
Chen, X.-H., Motani, R., Cheng, L., Jiang, D.-Y., and Rieppel, O. 2014. A carapace-like bony “body tube” in an Early Triassic marine reptile and the onset of marine tetrapod predation. PLoS ONE 9(4): e94396. Crossref
Curioni, G. 1847. Cenni sopra un nuovo saurio fossile dei monti di Perledo sul Lario e sul terreno che lo racchiude. Giornale dell’ I. R. Istituto Lombardo di Scienze, Lettre ed Arti 16: 159–170.
Cuthbertson, R.S., Russell, A.P., and Anderson, J.S. 2013. Cranial morphology and relationships of a new grippidian (Ichthyopterygia) from the Vega-Phroso Siltstone Member (Lower Triassic) of British Columbia, Canada. Journal of Vertebrate Paleontology 33: 831–847. Crossref
Dalla Vecchia, F.M. 1994. Reptile remains from the Middle–Upper Triassic of the Carnic and Julian alps (Friuli-Venezia Giulia, Northeastern Italy). Gortania. Atti del Museo Friulano di Storia Naturale 15 (for 1993): 46–66.
Dalla Vecchia, F.M. 2006. A new sauropterygian reptile with plesiosaurian affinity from the Late Triassic of Italy. Rivista Italiana di Paleontologia e Stratigrafia 112: 207–225.
Drevermann, F. 1933. Die Placodontier. 3. Das Skelett von Placodus gigas Agassiz im Senckenberg-Museum. Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft 38: 321–364.
Edwards, D.E., Barclay, J.E., Gibson, D.W., Kvill, G.E., and Halton, E. 1994. Triassic strata of the Western Canadian Sedimentary Basin. Geological Atlas of the Western Canada Sedimentary Basin: Canadian Society of Petroleum Geologists and Alberta Research Council, Special Report 4: 259–275.
Fabbri, M., Dalla Vecchia, F.M., and Cau, A. 2014. New information on Bobosaurus forojuliensis (Reptilia: Sauropterygia): implications for plesiosaurian evolution. Historical Biology 26: 661–669. Crossref
Fritsch, K. von 1894. Beitrag zur Kenntnis der Saurier des Halle’schen unteren Muschelkalkes. Abhandlungen der Naturforschenden Gesellschaft zu Halle 20: 273–302.
Fröbisch, N.B., Fröbisch, J., Sander, P.M., Schmitz, L., and Rieppel, O. 2013. A macropredatory ichthyosaur from the Middle Triassic and the origin of modern trophic networks. PNAS 110: 1393–1397. Crossref
Geissler, G. 1895. Ueber neue Saurier-Funde aus dem Muschelkalk von Bayreuth. Zeitschrift der Deutschen Geologischen Gesellschaft 47: 331–355.
Hagdorn, H. and Rieppel, O. 1998. Stratigraphy of marine reptiles in the Triassic of Central Europe. Zentralblatt für Geologie und Paläontologie, Teil I 1998 (7–8): 651–678.
Hänni, K. 2004. Die Gattung Ceresiosaurus. Ceresiosaurus calgagnii Peyer und Ceresiosaurus lanzi n. sp. (Lariosauridae, Sauropterygia). 146 pp. Vdf Hochschulverlag AG, Zürich.
Hansen, D.M. and Galetti, M. 2009. The forgotten megafauna. Science 324: 42–43. Crossref
Huene, F. von 1956. Paläontologie und Phylogenie der Niederen Tetrapoden. 716 pp. Gustav Fisher Verlag, Jena.
Jiang, D.-Y., Lin, W.-B., Rieppel, O., Motani, R., and Sun, Z.-Y. 2019. A new Anisian (Middle Triassic) eosauropterygian (Reptilia, Sauropterygia) from Panzhou, Guizhou Province, China. Journal of Vertebrate Paleontology 38: 4 [published online] Crossref
Jiang, D.-Y., Maisch, M.W., Hao, W.-C., Sun, Y.-L., and Sun, Z.-Y. 2006. Nothosaurus yangjuanensis n. sp. (Reptilia, Sauropterygia, Nothosauridae) from the middle Anisian (Middle Triassic) of Guizhou, southwestern China. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 2006 (5): 257–276. Crossref
Jiang, D.-Y., Motani, R., Tintori, A., Rieppel, O., Chen, G.-B., Huang, J.-D., Zhang, R., Sun, Z.-Y., and Ji, C. 2014. The Early Triassic eosauropterygian Majiashanosaurus discocoracoidis, gen. et sp. nov. (Reptilia, Sauropterygia), from Chaohu, Anhui Province, People’s Republic of China. Journal of Vertebrate Paleontology 34: 1044–1052.Crossref
Kear, B.P., Rich, T.H., Ali, M.A., Al-Mifarrih, Y.A., Matiri, A.H., Masary, A.M., and Attia, Y. 2008. Late Cretaceous (Campanian–Maastrichtian) marine reptiles from the Adaffa Formation, NW Saudi Arabia. Geological Magazine: 1–7. Crossref
Kelley, N.P., Motani, R., Jiang, D.-Y., Rieppel, O., and Schmitz, A. 2014. Selective extinction of Triassic marine reptiles during long-term sea-level changes illuminated by seawater strontium isotopes. Palaeogeography, Palaeoclimatology, Palaeoecology 400: 9–16. Crossref
Ketchum, H.F. and Benson, R.B.J. 2011. A new pliosaurid (Sauropterygia, Plesiosauria) from the Oxford Clay Formation (Middle Jurassic, Callovian) of England: evidence for a gracile longirostrine grade of Early–Middle Jurassic pliosaurids. Special Papers in Palaeontology 86: 109–129.
Klein, N. and Albers, P.C.H. 2009. A new species of the sauropsid reptile Nothosaurus from the Lower Muschelkalk of the western Germanic Basin, Winterswijk, The Netherlands. Acta Palaeontologica Polonica 54: 589–598. Crossref
Klein, N. and Scheyer, T.M. 2014. A new placodont sauropterygian from the Middle Triassic of the Netherlands. Acta Palaeontologica Polonica 59: 887–902. Crossref
Li, C., Jiang, D.-Y., Cheng, L., Wu, X.-C., and Rieppel, O. 2014. A new species of Largocephalosaurus (Diapsida: Saurosphargidae), with implications for the morphological diversity and phylogeny of the group. Geological Magazine 151: 100–120. Crossref
Li, C., Wu, X.-C., Cheng, Y.-N., Sato, T., and Wang, L. 2006. An unusual archosaurian from the marine Triassic of China. Naturwissenschaften 93: 200–206. Crossref
Li, J.-L. and Rieppel, O. 2004. A new nothosaur from Middle Triassic of Guizhou, China. Vertebrata PalAsiatica 42: 1–12.
Li, J.-L., Liu, J., and Rieppel, O. 2002. A new species of Lariosaurus (Sauropterygia: Nothosauridae) from Triassic of Guizhou, southwest China. Vertebrata PalAsiatica 40: 114–126.
Liu, J., Hu, S.-X., Rieppel, O., Jiang, D.-Y., Benton, M.J., Kelley, N.P., Aitchison, J.C., Zhou, C.-Y., Wen, W., Huang, J.-Y., Xie, T., and Lv, T. 2014. A gigantic nothosaur (Reptilia: Sauropterygia) from the Middle Triassic of SW China and its implication for the Triassic biotic recovery. Scientific Reports 4: 7142 . Crossref
Liu, J., Zhao, L.-J., Li, C., and He, T. 2013. Osteology of Concavispina biseridens (Reptilia, Thalattosauria) from the Xiaowa Formation (Carnian), Guanling, Guizhou, China. Journal of Paleontology 87: 341–350. Crossref
Ma, L.-T., Jiang, D.-Y., Rieppel, O., Motani, R., and Tintori, A. 2015. A new pistosauroid (Reptilia, Sauropterygia) from the late Ladinian Xingyi marine reptile level, southwestern China. Journal of Vertebrate Paleontology 35 (1): e881832. Crossref
Massare, J.A. and Callaway, J.M. 1994. Cymbospondylus (Ichthyosauria: Shastasauridae) from the lower Triassic Thaynes Formation of southeastern Idaho. Journal of Vertebrate Paleontology 14: 139–141. Crossref
Maxwell, E.E. and Kear, B.P. 2013. Triassic ichthyopterygian assemblages of the Svalbard Archipelago: a reassessment of taxonomy and distribution. GFF 135 (1): 85–94. Crossref
McGowan, C. and Motani, R. 2003. Ichthyopterygia. Handbuch der Paläoherpetologie / Handbook of Paleoherpetology Part 8: 1–173.
Meyer, H. von 1842. Simosaurus, die Stumpfschnauze, ein Saurier aus dem Muschelkalke von Luneville. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde 1842: 184–198.
Motani, R., Jiang, D.-Y., Chen, G.-B., Tintori, A., Rieppel, O., Ji, C., and Huang, J.-D. 2014a. A basal ichthyosauriform with a short snout from the Lower Triassic of China. Nature 517: 483–488. Crossref
Motani, R., Jiang, D.-Y., Tintori, A., Rieppel, O., and Chen, G.-B. 2014b. Terrestrial origin of viviparity in Mesozoic marine reptiles indicated by Early Triassic embryonic fossils. PLoS ONE 9(2): e88640. Crossref
Müller, J. 2005. The anatomy of Askeptosaurus italicus from the Middle Triassic of Monte San Giorgio and the interrelationships of thalattosaurs (Reptilia, Diapsida). Canadian Journal of Earth Sciences 42: 1347–1367. Crossref
Münster, G. 1834. Vorläufige Nachricht über einige neue Reptilien im Muschelkalke von Baiern. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde 1834: 521–527.
Mutter, R.J. 2004. Fossile Fische aus der Trias der kanadischen Rocky Mountains. Vierteljahresschrift der Naturforschenden Gesellschaft in Zürich 149 (2–3): 51–58.
Mutter, R.J. and Neuman, A.G. 2008. New eugeneodontid sharks from the Lower Triassic Sulphur Mountain Formation of Western Canada. In: L. Cavin, A. Longbottom, and M. Richter (eds.), Fishes and the Break-up of Pangaea. Geological Society, London, Special Publications 295: 9 –41. Crossref
Mutter, R.J. and Neuman, A.G. 2009. Recovery from the end-Permian extinction event: Evidence from “Lilliput Listracanthus”. Palaeogeography, Palaeoclimatology, Palaeoecology 284: 22–28. Crossref
Neenan, J.M., Klein, N., and Scheyer, T.M. 2013. European origin of placodont marine reptiles and the evolution of crushing dentition in Placodontia. Nature Communications 4: 1621.
Neuman, A.G. 1986. Fossil Fishes of the Families Perleididae and Parasemionotidae from the Lower Triassic Sulphur Mountain Formation of Western Canada. 142 pp. M.Sc. Thesis, Department of Zoology, University of Alberta, Edmonton.
Nopcsa, F. 1928. Palaeontological notes on reptiles. Geologica Hungarica, Palaeontology Series 1: 1–84.
O’Keefe, F.R., Street, H.P., Wilhelm, B.C., Richards, C.D., and Zhu, H. 2011. A new skeleton of the cryptoclidid plesiosaur Tatenectes laramiensis reveals a novel body shape among plesiosaurs. Journal of Vertebrate Paleontology 31: 330–339. Crossref
Orchard, M.J. and Zonneveld, J.-P. 2009. The Lower Triassic Sulphur Mountain Formation in the Wapiti Lake area: lithostratigraphy, conodont biostratigraphy, and a new biozonation for the lower Olenekian (Smithian) Canadian Journal of Earth Sciences 46: 757–790. Crossref
Owen, R. 1860. Palaeontology, or, a Systematic Summary of Extinct Animals and their Geological Relations. xv + 420 pp. Adam and Charles Black, Edinburgh. Crossref
Peyer, B. 1931. Paraplacodus broilii nov. gen. nov. sp., ein neuer Placodontier aus der Tessiner Trias. Centralblatt für Mineralogie, etc. Abt. B 10: 570–573.
Peyer, B. 1939. Die Triasfauna der Tessiner Kalkalpen XIV. Paranothosaurus amsleri nov. gen. nov. spec. Abhandlungen der schweizerischen Paläontologischen Gesellschaft 62: 1–87.
Rieppel, O. 1989. Helveticosaurus zollingeri Peyer (Reptilia, Diapsida) skeletal paedomorphosis, functional anatomy and systematic affinities. Palaeontographica A 208: 123–152.
Rieppel, O. 1994. Osteology of Simosaurus gaillardoti and the relationships of stem-group Sauropterygia. Fieldiana (Geology), New Series 28: 1–85. Crossref
Rieppel, O. 1997. Revision of the sauropterygian reptile genus Cymatosaurus v. Fritsch, 1894, and the relationships of Germanosaurus Nopcsa, 1928, from the Middle Triassic of Europe. Fieldiana (Geology), New Series 36: 1–38. Crossref
Rieppel, O. 1998. Corosaurus alcovensis Case and the phylogenetic interrelationships of Triassic stem-group Sauropterygia. Zoological Journal of the Linnean Society 124: 1–41. Crossref
Rieppel, O. 1999. Phylogeny and paleobiogeography of Triassic Sauropterygia: problems solved and unresolved. Palaeogeography, Palaeoclimatology, Palaeoecology 153: 1–15. Crossref
Rieppel, O. 2000. Sauropterygia. I. Placodontia, Pachypleurosauria, Nothosauroidea, Pistosauroidea. Handbuch der Paläoherpetologie 12A: 1–134.
Rieppel, O. and Hagdorn, H. 1997. Paleobiogeography of Middle Triassic Sauropterygia in central and western Europe. In: J.M. Callaway, and E.L. Nicholls (eds.), Ancient Marine Reptiles, 121–144. Academic Press, San Diego. Crossref
Rieppel, O. and Wild, R. 1996. A revision of the genus Nothosaurus (Reptilia: Sauropterygia) from the Germanic Triassic, with comments on the status of Conchiosaurus clavatus. Fieldiana (Geology), New Series 34: 1–82. Crossref
Rieppel, O., Li, C., and Fraser, N.C. 2008. The skeletal anatomy of the Triassic protorosaur Dinocephalosaurus orientalis Li, from the Middle Triassic of Guizhou Province, Southern China. Journal of Vertebrate Paleontology 28: 95–110. Crossref
Rieppel, O., Liu, J., and Li, C. 2006. A new species of the thalattosaur genus Anshunsaurus (Reptilia: Thalattosauria) from the Middle Triassic of Guizhou Province, Southwestern China. Vertebrata PalAsiatica 44: 285–296.
Rieppel, O., Mazin, J.-M., and Tchernov, E. 1999. Sauropterygia from the Middle Triassic of Makhtesh Ramon, Negev, Israel. Fieldiana (Geology), New Series 40: 1–85. Crossref
Romano, C., Jenks, J.F., Jattiot, R., Scheyer, T.M., Bylund, K.G., and Bucher, H. 2017. Marine Early Triassic Actinopterygii from Elko County (Nevada, USA): implications for the Smithian equatorial vertebrate eclipse. Journal of Paleontology 91: 1025–1046. Crossref
Romano, C., Koot, M.B., Kogan, I., Brayard, A., Minikh, A.V., Brinkmann, W., Bucher, H., and Kriwet, J. 2016. Permian–Triassic Osteichthyes (bony fishes): diversity dynamics and body size evolution. Biological Reviews 91: 106–147. Crossref
Sander, P.M., Rieppel, O.C., and Bucher, H. 1997. A new pistosaurid (Reptilia: Sauropterygia) from the Middle Triassic of Nevada and its implications for the origin of the plesiosaurs. Journal of Vertebrate Paleontology 17: 526–533. Crossref
Sato, T., Cheng, Y.-N., Wu, X.-C., and Li, C. 2010. Osteology of Yunguisaurus Cheng et al., 2006 (Reptilia; Sauropterygia), a Triassic pistosauroid from China. Paleontological Research 14: 179–195. Crossref
Sato, T., Zhao, L.-J., Wu, X.-C., and Li, C. 2014. A new specimen of the Triassic pistosauroid Yunguisaurus, with implications for the origin of Plesiosauria (Reptilia, Sauropterygia). Palaeontology 57: 55–76. Crossref
Schaeffer, B. and Mangus, M. 1976. An Early Triassic fish assemblage from British Columbia. Bulletin of the American Museum of Natural History 156: 515–564.
Scheyer, T.M., Neenan, J.M., Bodogan, T., Furrer, H., Obrist, C., and Plamondon, M. 2017. A new, exceptionally preserved juvenile specimen of Eusaurosphargis dalsassoi (Diapsida) and implications for Mesozoic marine diapsid phylogeny. Scientific Reports 7: 4406. Crossref
Scheyer, T.M., Romano, C., Jenks, J., and Bucher, H. 2014. Early Triassic marine biotic recovery: the predators’ perspective. PLoS ONE 9 (3): e88987. Crossref
Seilacher, A. 1970. Begriff and Bedeutung der Fossil-Lagerstätten. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1970 (1): 34–39.
Smith, A.S. and Vincent, P. 2010. A new genus of pliosaur (Reptilia: Sauropterygia) from the Lower Jurassic of Holzmaden, Germany. Palaeontology 53: 1049–1063. Crossref
Storrs, G.W. 1991. Anatomy and relationships of Corosaurus alcovensis (Diapsida: Sauropterygia) and the Triassic Alcova Limestone of Wyoming. Peabody Museum of Natural History Bulletin 44: 1–151.
Sues, H.-D. 1987. The postcranial skeleton of Pistosaurus and the interrelationships of the Sauropterygia (Diapsida). Zoological Journal of the Linnean Society 90: 109–131. Crossref
Tschanz, K. 1989. Lariosaurus buzzii n. sp. from the Middle Triassic of Monte San Giorgio (Switzerland) with comments on the classification of nothosaurs. Palaeontographica Abteilung A 208 (4–6): 153–179.
Vickers-Rich, P., Rich, T.H., Rieppel, O., Thulborn, R.A., and McClure, H.A. 1999. A Middle Triassic vertebrate fauna from the Jilh Formation, Saudi Arabia. Neues Jahrbuch fuer Geologie und Paläontologie, Abhandlungen 213: 201–232. Crossref
Voeten, D.F.A.E., Sander, P.M., and Klein, N. 2015. Skeletal material from larger Eusauropterygia (Reptilia: Eosauropterygia) with nothosaurian and cymatosaurian affinities from the Lower Muschelkalk of Winterswijk, The Netherlands. Paläontologische Zeitschrift 89: 943–960. Crossref
Wild, R. 1973. E. Kuhn-Schnyder und B. Peyer: Die Triasfauna der Tessiner Kalkalpen. XXIII. Tanystropheus longobardicus (Bassani) (Neue Ergebnisse). Schweizerische Paläontologische Abhandlungen 95: 1–162.
Wintrich, T., Hayashi, S., Houssaye, A., Nakajima, Y., and Sander, P.M. 2017. A Triassic plesiosaurian skeleton and bone histology inform on evolution of a unique body plan. Science Advances 3: e1701144 [doi: 10.1126/sciadv.1701144]. Crossref
Wu, X.-C., Cheng, Y.-N., Sato, T., and Shan, H.-Y. 2009. Miodentosaurus brevis Cheng et al., 2007 (Diapsida: Thalattosauria): its postcranial skeleton and phylogenetic relationships. Vertebrata PalAsiatica 47: 1–20.
Zangerl, R. 1963. Preliminary results of a restudy of Corosaurus alcovensis Case, the only known new world nothosaur. Rocky Mountain Geology (Contributions to Geology) 2: 117–123.
Appendix 1
Additional osteological comparison.
(i) TMP 1995.116.0023 differs from larger protorosaurs such as Dinocephalosaurus orientalis (Rieppel et al. 2008) in the absence of a strongly concave posterior margin of the femur and the equal robusticity and proximal expansion of the zeugopodial elements, and from Tanystropheus longobardicus (Wild 1973) in having a distinctly greater expansion and differently shaped femoral epiphyses. A large spatium interossium between the zeugopodial bones is shared with D. orientalis (Rieppel et al. 2008); whereas the ischial shape in TMP 1995.116.0023 resembles only weakly that of some earlier growth stages of T. longobardicus (see Wild 1973: fig. 92g). It further differs from the Chinese marine archosaur Qianosuchus mixtus (see Li et al. 2006) in having a fan-shaped ischiadic ventral blade, overall much more robust stylopod and zeugopodial elements, and a less sigmoidal shaped robust femur.
(ii) It differs from the Chinese saurosphargid Largocephalosaurus qianensis (Li et al. 2014) in the similarly broad expansions of the proximal and distal ends of the femur, the proximal shapes of the tibia and fibula, and the strong constriction of the ischium.
(iii) TMP 1995.116.0023 differs from large thalattosaurs such as Askeptosaurus italicus, Anshunsaurus wushanensis, Concavispina biseridens, and Miodentosaurus brevis (these are the largest thalattosaurs known) in femur shape. Thalattosaur femora lack clearly developed distal condyles of the femur for the articulation with the tibia and fibula. In addition, these femora usually reach only slightly more than half the size of TMP 1995.116.0023, except in Miodentosaurus, whose femur almost reaches the dimensions of the Canadian specimen (Müller 2005; Rieppel et al. 2006; Wu et al. 2009; Liu et al. 2013). Furthermore, thalattosaur taxa lack the strong constriction of the ischiadic shaft region and the broader ventral fan-shaped blade.
(iv) TMP 1995.116.0023 differs from ichthyosauromorphs, including the Chinese hupehsuchians Parahupehsuchus longus and Hupehsuchus nanchangensis (Chen et al. 2014), as well as ichthyosauriforms (e.g., the Triassic forms Cartorhynchus lenticarpus, Chaohusaurus spp., Mixosaurus spp., Utatsusaurus hataii, Shastasaurus spp.) in having a less broadened femur with a long diaphyseal shaft and widely expanded proximal and distal epiphyseal areas and there is further no indication for the formation of paddle-shaped hind limbs with reduced or plate-like/lunate-shaped ischia as are typical for ichthyopterygians (Callaway 1997; McGowan and Motani 2003; Motani et al. 2014a, b).
(v) TMP 1995.116.0023 shares with the enigmatic marine reptile Helveticosaurus from the Middle Triassic of Monte San Giorgio, Switzerland, the massive stylopod and zeugopod, a spatium interossium between tibia and fibula, as well as the similar proximal articulation facets in these elements (Rieppel 1989). However, TMP 1995.116.0023 differs in femoral shape and its distal articulation facets, as well as the distinctly different shape of the ischium. In addition, the new specimen differs also from larger placodonts including Placodus gigas and Pararcus diepenbroeki (Drevermann 1933; Klein and Scheyer 2014) in possessing a constricted rather than plate like ischium.
Acta Palaeontol. Pol. 64 (4): 745–755, 2019
https://doi.org/10.4202/app.00599.2019