
Bashkirian rugose corals from the Carboniferous Mattson Formation in the Liard Basin, northwest Canada—stratigraphic and paleobiogeographic implications
JERZY FEDOROWSKI, E. WAYNE BAMBER, and BARRY C. RICHARDS
Fedorowski, J., Bamber, E.W., and Richards, B.C. 2019. Bashkirian rugose corals from the Carboniferous Mattson Formation in the Liard Basin, northwest Canada—stratigraphic and paleobiogeographic implications. Acta Palaeontologica Polonica 64 (4): 851–870.
Colonies of the rugose corals Nemistium liardense sp. nov. and Heritschioides simplex sp. nov. were collected from limestone in the upper member of the Mattson Formation in the Liard Range in the Northwest Territories and are the only known identifiable coral species from the Mattson Fm. The Mattson Fm., deposited in the Liard Basin west of the syndepositional Bovie reverse fault, comprises sandstone with subordinate shale and carbonates deposited during several delta cycles. The close morphological similarity and identical mode of offsetting in N. liardense colonies from the Mattson Fm. and the allochthonous Stikine Terrane of British Columbia indicate they belong in the same species. This and the morphological similarity between H. simplex and the late Serpukhovian to early Bashkirian H. columbicum allow assignment of the coral-bearing part of the upper Mattson Fm. to Bashkirian Foraminiferal Biozone 20. Widespread occurrence of the genus Nemistium confirms open communication between the Liard Basin region and the western European and northern African seas.
Key words: Rugosa, Stauriida, taxonomy, paleogeography, Carboniferous, Pennsylvanian, Canada.
Jerzy Fedorowski [jerzy@amu.edu.pl], Institute of Geology, Adam Mickiewicz University, Bogumiła Krygowskiego 12, 61-680 Poznań, Poland.
E. Wayne Bamber [wayne.bamber@canada.ca] and Barry C. Richards [Barry.richards@canada.ca], Geological Survey of Canada (Calgary) Natural Resources Canada/Government of Canada 3303-33rd Street Northwest, Calgary, Alberta T2L 2A7, Canada.
Received 10 May 2019, accepted 3 September 2019, available online 5 December 2019.
Copyright © 2019 J. Fedorowski et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
This paper is devoted to Carboniferous colonial rugose corals collected from the upper member of the late Viséan to Bashkirian Mattson Formation in the Liard Range of the southwest corner of the Northwest Territories, northwestern Canada (Figs. 1–3). That remote area has been extensively mapped and its lithostratigraphy is well established (see Geological setting for details). However, the biostratigraphy, especially that of the upper Viséan to Pennsylvanian deposits (i.e., the Mattson Formation, dealt with in this paper) is not well established because that unit consists mostly of sandstone with subordinate shale and carbonates. Harker (1963) divided the Mattson Fm. into three informal members: lower, middle and upper. Only the upper member contains numerous widely-spaced units of sandy limestone and dolostone, some of which yield fossils. Harker (1963) collected and identified several species of brachiopods from the upper Mattson Fm. but palynomorphs are the best known fossil group from the Mattson Fm. (Braman and Hills 1977; Utting in Richards 1983; and Richards et al. 1993). Braman and Hills interpreted the microflora from the type section to be of late Viséan age. Utting in Richards (1983) and Richards et al. (1993), using palynomorphs collected higher in the section than Braman and Hills, interpreted the type Mattson Fm. to be late Viséan to Serpukhovian.
Rugose corals are rather numerous in the Tournaisian and lower to middle Viséan strata of the Liard Basin and adjacent areas. However, the paper by Sutherland (1958) is the only detailed study of rugose corals from that area and strata, although they are not from the sections measured by BCR. Rugose corals have not previously been found in the Mattson Fm. They were collected by BCR from the middle part of the upper informal member of only one section measured by him, i.e., section 20RAH9 at the headwaters of Blue Bill Creek near Mattson Gap in the Liard Range (see Geological setting for details).
In the present paper we review and systematically describe this coral fauna. Also, we attempt to precisely determine the biostratigraphic position of the upper limestone unit in the upper member of the Mattson Fm. by comparing the colonial rugose corals identified here to those from areas in which the biostratigraphy is well established and to settle the described fauna into a known paleogeographic context.
Institutional abbreviations.—GSC, Geological Survey of Canada.
Other abbreviations.—d, corallite diameter (in mm); Fm., Formation; n, number of major septa; NAD27, North American Datum 1927 map projection; NATMAP, Geological Survey of Canada program name; NTS, National Topographic System; RAH, Geological Survey of Canada officer code for BCR.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in ZooBank: urn:lsid:zoobank.org:pub:CEDBDA7D-F784-4D15-B917-88C628738C35
Material and methods
The coral specimens discussed herein were collected from the Mattson Gap section (20RAH9). The specimens are usually worn and apparently redeposited from their original neritic setting. The colonies are not in growth position at the outcrop and in one of the samples (Fig. 7), colonies of both genera abut closely and the corallites in adjacent colonies are broken. The breakage along the line of contact strongly suggests waves or current action. The matrix is identical in content and grain size within branches of both colonies to the ambient rock, thereby indicating only short transport. Further deformations of particular corallites, such as their compression, silicification and recrystallization resulted from deep burial and diagenesis. That kind of preservation excludes effective use of the peeling method to study blastogeny. Because we could not make series of peels, the blastogeny process is described by using random transverse and longitudinal thin sections of offsetting corallites. Fragments of the corallites preserve the microstructure of the septa and we studied that important character of the corals in both species described. We slabbed and made thin sections from all of the colonies.
All coral specimens from the Mattson Gap section 20RAH9 and resulting thin sections are housed at the Geological Survey of Canada under the collection number C-442050. Studied material has been collected by BCR from a 2.47 m thick limestone unit in the middle part of the upper member of the Mattson Fm. between 1056.7 and 1057.1 m above the base of section 20RAH9 (1021.64 to 1022.04 m above base Mattson Fm.; 401.03 to 401.43 m above base upper member). The locality is at easting 442132, northing 6749414 in zone 10, NAD 27, Etanda Lakes map 95-C-16, scale 1:50 000.
Geological setting
The Mattson Fm., extending from about 58°45’ in northeastern British Columbia to 61°25’N in the southwest corner of the Northwest Territories, was deposited in the Liard Basin (Figs. 1, 2). The basin, a sub-basin of the Prophet Trough and Western Canada Sedimentary Basin, occupies a prominent structural re-entrant in the Cordilleran deformation front near 60°N (Wright et al. 1994). On the east, the Liard Basin is bounded by the deep-seated Bovie reverse fault (MacLean 2002; MacLean and Morrow 2004). On its west side, uplifted Paleozoic strata in the Cordilleran fold and thrust belt define the western margin of the basin. The Bovie reverse fault was periodically active from the Famennian into the Cretaceous and separates thick upper Paleozoic, Triassic, and Cretaceous successions in the Liard Basin from thinner correlative packages of strata to the east (Leckie et al. 1991; Richards et al. 1994; Henderson et al. 1994; MacLean and Morrow 2004; Ferri et al. 2015). The Liard Basin formed above a westerly inclined Famennian carbonate ramp during episodes of Late Devonian to Permian extension along the western margin of Ancestral North America. Several west-facing clinoformal packages of carbonates, shale, and bedded chert prograded across the eastern part of the basin in the Late Devonian to late middle Viséan (Richards et al. 1994). Northerly derived terrigenous clastics terminated carbonate sedimentation in the latest middle Viséan. Subsequently, the basin was infilled with upper Viséan to Serpukhovian prodelta to basinal shale and mudstone of the Besa River and Golata formations, upper Viséan to Bashkirian deltaic deposits of the sandstone-dominated Mattson Fm. (Richards et al. 1993; McMechan et al. 2018), and Serpukhovian to Bashkirian siltstone-dominated slope deposits of the lower Kindle Formation (Chung 1993).
Harker (1963), Douglas (1976), and Douglas and Norris (1976, 1977) divided the Mattson Fm. north of 60° into the informal lower, middle and upper members. Three informal members were also used in more recent regional mapping projects (Currie et al. 2000; Khudoley and Fallas 2006; Lane 2006). The upper member differs from the lower and middle members by containing numerous widely-spaced units of sandy limestone and dolostone. Quartzarenite constitutes most of the sandstone in the lower and middle Mattson Fm. and is common in parts of the upper Mattson Fm. but subchertarenite and chertarenite occur in the middle to upper parts of the upper member in the Mattson Fm. type section and at many other localities in the eastern Liard Basin. The members, most readily recognized north of 60° in the Liard Range and other eastern ranges of the southern Franklin Mountains of the Northwest Territories, become increasingly difficult to recognize toward the southwest and into northeastern British Columbia.
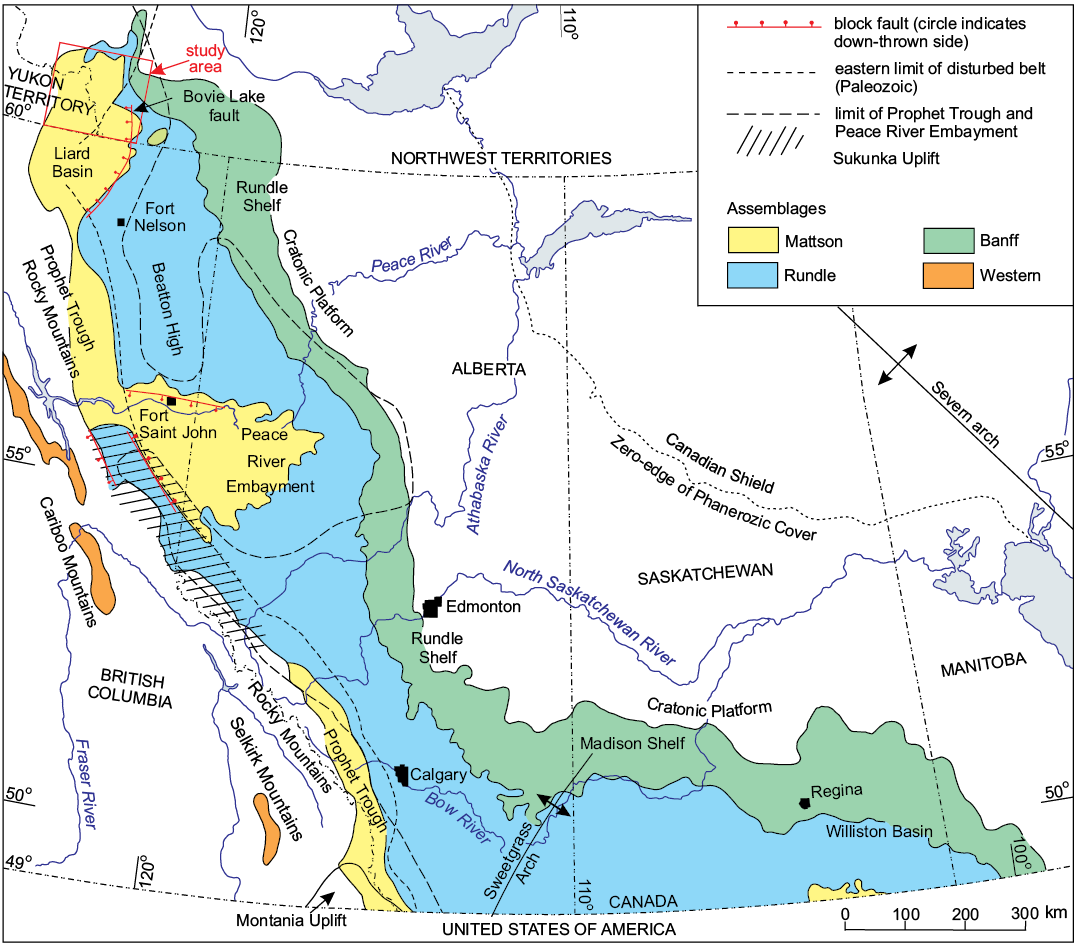
Fig. 1. Map showing location of study area and Carboniferous lithofacies assemblages subcropping beneath Permian and Mesozoic deposits in the Western Canada Sedimentary Basin (modified from Richards et al. 1994). In the Liard Basin, western occurrences of the Rundle and Mattson lithofacies assemblages overlie and pass westward into the Besa River lithofacies assemblage. See Fig. 3 for the formational composition of the Banff, Rundle, and Mattson lithofacies assemblages.
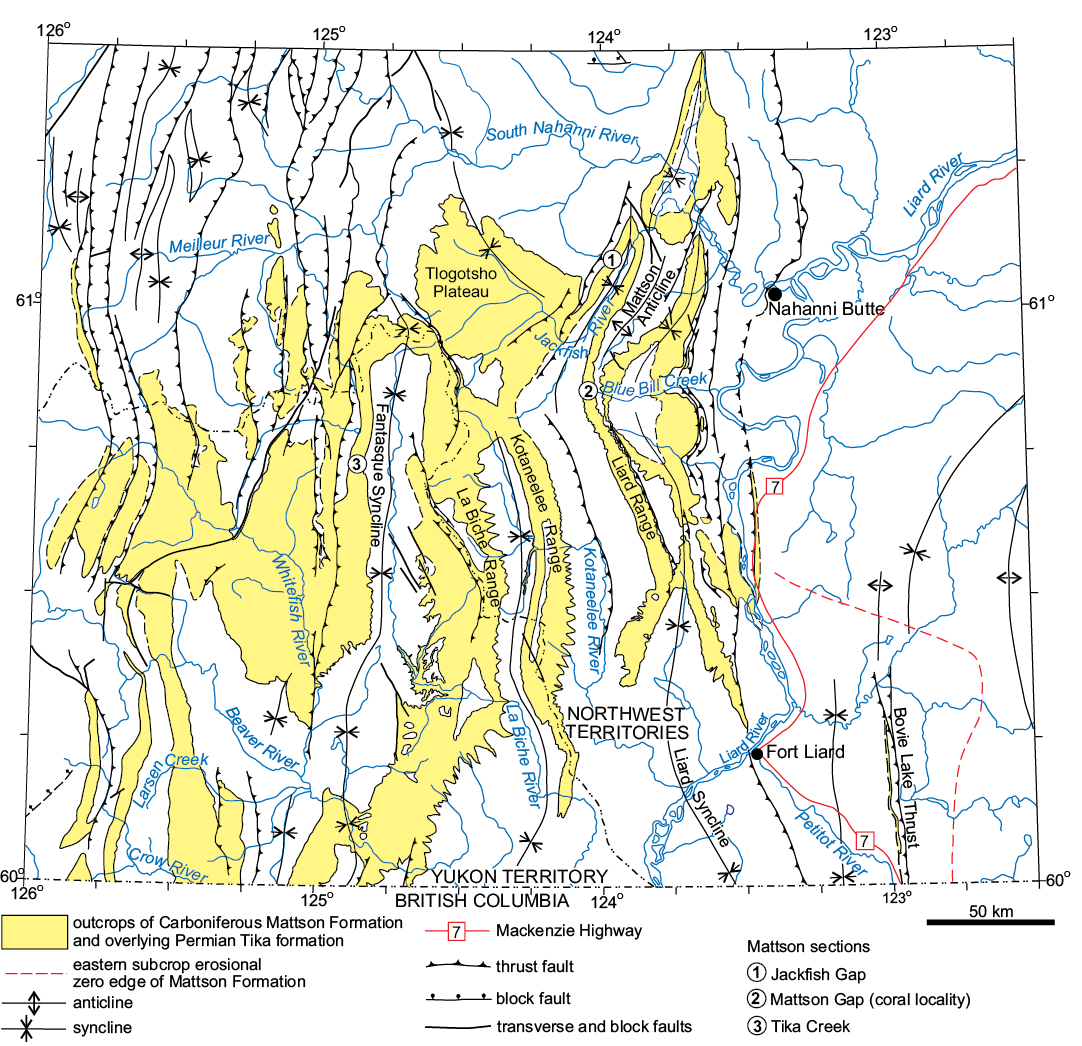
Fig. 2. Simplified geological map of study region showing distribution of outcrops of Carboniferous Mattson Formation in southwest corner of the Northwest Territories and southeastern Yukon Territory (modified from Harker 1963 and an unpublished Geological Survey of Canada 1:1 000 000 scale map prepared by Andrew Okulitch in 2005).
The Mattson Fm. conformably overlies the shale-dominant Viséan Golata Fm. in the northeast and the shale-dominated Middle Devonian to Serpukhovian Besa River Fm. to the southwest (Fig. 3). Toward the southwest (basinward), sandstone and shale of the lower and middle Mattson Fm. pass into the Golata and Besa River formations. In northeastern British Columbia, much of the upper member of the Mattson Fm. passes basinward into Serpukhovian and Bashkirian siltstone and silty carbonates of the Kindle Fm. (Chung 1993; McMechan et al. 2018).
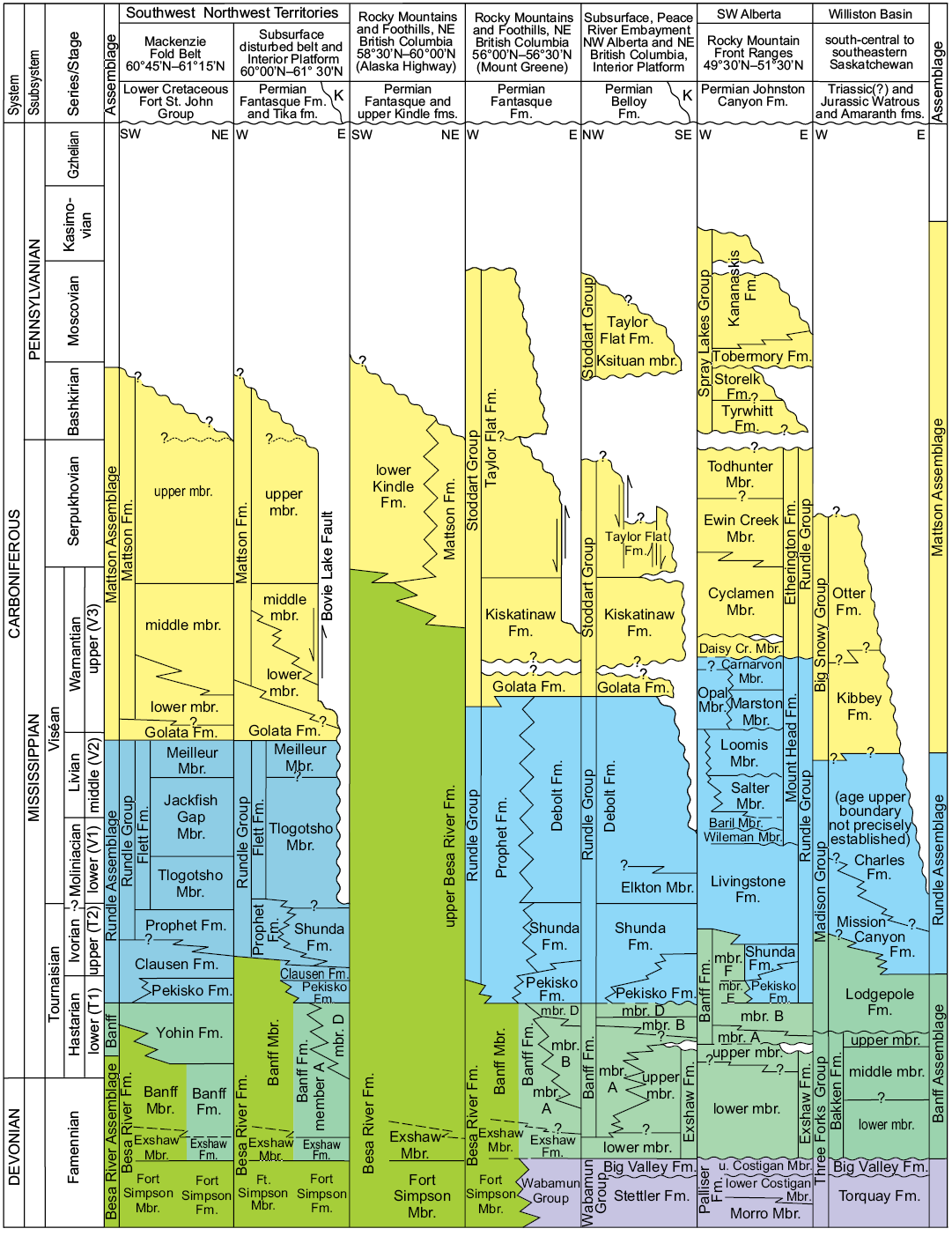
Fig. 3. Correlation of Carboniferous lithostratigraphic units from study area in southwest corner of the Northwest Territories to the Williston Basin in southern Saskatchewan. Shows relationship of late Viséan to Bashkirian Mattson Formation to other sandstone dominated Carboniferous formations in the Western Canada Sedimentary Basin and to underlying and overlying formations (modified from Richards et al. 1994). Figure also shows formational composition of the assemblages mapped on Fig. 1; colours correlate with those on Fig. 1. Abbreviations: Cr., Creek; Fm., Formation (formal unit); fm., formation (informal unit); K, Cretaceous; Mbr., Member (formal unit); mbr., member (informal unit); u., upper.
The Mattson Fm. attains a thickness of 1008.7 m at its type section on Yohin Ridge in the Northwest Territories and thickens southeastward to 1293 m in section 20RAH9 at the head of Blue Bill Creek in the Liard Range (Fig. 2: section 2) and is up to 1400 m thick at Tika Creek (Fig. 2: section 3) in eastern Yukon Territory (Richards et al. 1989). In northeast British Columbia, the Mattson Fm. is thickest in the northeastern part of the Liard Basin and thins southwestward to less than 10 m in the fold and thrust belt near the Alaska Highway. The marked thinning results from the lower Mattson Fm. passing into the Besa River and the upper Mattson formations passing basinward into the Kindle Fm. The Mattson Fm. thins abruptly at the Bovie fault occurring on the east side of the fault as an erosional remnant that is generally less than 15 m thick.
In most of the Liard Basin, lower Permian strata of the upper Kindle Fm. and correlative informal Tika formation unconformably overlie the Mattson Fm. and lower Kindle Fm. (Bamber et al. 1968; Richards et al. 1994; Henderson et al. 1994). The upper Kindle Fm. comprises sandstone and subordinate sandy limestone passing upward and southwestward into siltstone. In the Liard Basin north of 60°, the upper Kindle Fm. has been informally named the Tika formation on recent maps (Currie et al. 2000; Khudoley and Fallas 2006; Khudoley 2003). North of the erosional zero edge of the Tika formation in southeast Yukon and southwest District of Mackenzie, Lower Cretaceous strata of the Fort Saint John Group unconformably overlie the Mattson Fm. and cut down section toward the north.
One of us (BCR) collected the coral specimens described in this paper when he measured a stratigraphic section 3.3 km south of Mattson Gap at the headwaters of Blue Bill Creek in the Etanda Lakes map area 95-C-16 during the 2000 to 2003 field seasons as part of the Geological Survey of Canada Central Forelands NATMAP project. Following the GSC-Calgary curatorial procedure, Richards named the Mattson Gap section containing the coral locality 20RAH9, indicating he started measuring it in 2000 and that it was the ninth locality he sampled in 2000. The section starts in middle Viséan limestone of the uppermost Flett Fm. at the entrance to a steep narrow canyon on the east side of Jackfish Ridge in the Liard Range at 60º52’36.38”N, 124º02”02.94”W (NTS location: easting 443850, northing 6749300, zone 10v, NAD 27). It continues upward through the lower Mattson Fm. and lower part of the middle member of the Mattson Fm. to the top of the Liard Range. From there, it continues down and westward through the remainder of the middle Mattson Fm. and the upper member along a steep-sided canyon containing a tributary to Chinkeh Creek on the west side of the range. The section terminates in the lower beds of the Lower Cretaceous Chinkeh Fm. at 60°52’35.26”N, 124°04’25.46”W (NTS location: easting 441700, northing 6749300, zone10v, NAD 27). Fallas et al. (2003) and Lane and Hynes (2005) provide detailed 1:50 000 scale bedrock geology maps of the area.
The corals were collected by BCR from the middle part of the Mattson’s Fm. upper member in a 2.47 m thick unit of cherty, skeletal lime wackestone and packstone lying between 400.46 and 402.93 m above the base of the upper member in section 20RAH9. The coral-bearing unit lies between 1021.07 and 1023.49 m above the base of the Mattson Fm. and 269.66 m below its top at easting 442132, northing 6749414 in zone 10, NAD 27, 95-C-16. The upper member in section 20RAH9 is a cyclic succession dominated by sandstone deposited in offshore-marine, shoreline, coastal- dune, and fluvial settings. In its lower part (between 655.67 m and 736.96 m above base of section), the member contains several limestone units and fossiliferous, calcareous to dolomitic sandstone units that grade into sandy limestone and dolostone of neritic to shallow-neritic aspect. A similar interval occurs in the middle part of the upper member between 994.77 and 1058.55 m above the base of the section and includes the coral-bearing unit at its top. In the interval from 994.77 to 1058.55 m, the deposits become more calcareous and fossiliferous upward and from 1038.52 to 1058.55 m skeletal lime wackestone to packstone predominate. The 2.74 m limestone unit containing the corals abruptly overlies a 1.55 m thick unit of skeletal lime grainstone and is the highest known limestone unit in the section. Carboniferous marine sandstone, which has not been precisely dated, abruptly overlies the unit containing the corals.
Although both coral species collected in the Liard Range section are new, they are important for indicating the age of the youngest carbonate beds in the upper member in that section as Bashkirian. Most previous workers reported the Mattson Fm. to be of late Viséan to Serpukhovian age (Braman and Hills 1977; Bamber et al. 1984; Richards et al. 1989, 1993, 1994; Potter et al. 1993) and that the youngest deposits in the upper member are either Viséan or Serpukhovian. Harker (1963), however, stated that the upper member contains widely preserved Pennsylvanian deposits. Palynological data indicate the upper member is of late Viséan to Serpukhovian age in the Mattson Fm. type section at Jackfish Gap (Utting in Richards 1983; Richards et al. 1993). Elsewhere, the age of the upper member is poorly constrained because of a lack of published biostratigraphic data. Conodont data from the Kindle Fm. in northeastern British Columbia (Chung 1993) provide indirect evidence that Bashkirian deposits may occur in the upper Mattson Fm. at some locations. In northeastern British Columbia, the Kindle Fm. contains Bashkirian strata and passes northeastward into the upper Mattson Fm.
Systematic palaeontology
Subclass Rugosa Milne Edwards and Haime, 1850
Order Stauriida Verrill, 1865
Suborder Lithostrotionina Spasskiy and Kachanov, 1971
Family Lithostrotionidae d’Orbigny, 1852
Subfamily Diphyphyllinae Dybowski, 1873
Genus Nemistium Smith, 1928
Type species: Nemistium edmondsi Smith, 1928; Eskett Quarry near Winder Station, West Cumberland, UK; late Viséan, Subzone D2 (= Brigantian); by original designation.
Emended diagnosis.—After Hill (1981: F383), supplemented, original diagnosis in quotation marks. “Phaceloid, with peripheral increase”; “axial ends of major septa abutting axial structure”, but median lamella derived from counter (?) septum and/or septal lamellae form interrupted axial structure; “arched axial tabulae, each resting on the arch below” form continuous columnotheca; “periaxial tabulae abut axial structure”; dissepiments interseptal; microstructure finely trabecular (?).
Remarks.—Question marks in the proposed diagnosis resulted from: (i) absence of definite data concerning the septum from which pseudocolumella is separated, (ii) lack of data concerning the microstructure of septa in the type species. This feature is suggested on the basis of the Canadian material studied in this paper.
Smith (1928: 114) diagnosed his new genus as “agreeing with Diphyphyllum β in growth habit, manner of gemmation and in most internal structure, but differing from these in having a loosely formed and irregular axial structure”. Thus, he pointed to the most important characters of that genus, indicating its relationships at the same time. He did not describe the microstructure of the septa, the insertion of the septa in offsets, the way the medial lamella appeared or its relationship to the protosepta. Despite the value of those details in terms of the comprehensive characteristic of that genus, nothing has been added concerning those terms since his study. Publications by Hill (1956, 1981) and Nudds (1981) only repeated Smith’s (1928) data. Moreover, Nudds (1981) synonymized Nemistium with Lithostrotion, a suggestion that cannot be followed.
The generic name Nemistium is seldom mentioned in the rugose coral literature (e.g., Poty 1984, 2002; Rodríguez 1984; Semenoff-Tian-Chansky 1985; Fedorowski 2009; Denayer 2014), but some species assigned to Lithostrotion Fleming, 1828 or Diphyphyllum Lonsdale, 1845 may either belong to Nemistium or represent a new, related genus. Poty (1984) suggested the transfer to Nemistium of taxa assigned by earlier authors to other genera. These are: (i) Caryophyllea affinis Fleming, 1828, listed by Poty (1984: 465) as Nemistium affine (Fleming, 1828). That species, redescribed by Kato (1971), is not accepted here as a member of Nemistium (see below). (ii) Three species described by Dobrolyubova (1958) from the Moscow Basin, i.e., Diphyphyllum fasciculatum (Fleming, 1828), Diphyphyllum gracile McCoy, 1851 and Lithostrotion scoticum Hill, 1940. All those species are here considered to belong to a new, as yet unnamed genus. (iii) Lithostrotion (Siphonodendron) dobroljubovae Khoa, 1977 from the Lublin Basin in Poland belonging to the same genus as the Russian species Diphyphyllum fasciculatum, Diphyphyllum gracile, and Lithostrotion scoticum described by Dobroljubova (1958). In addition to those four species, two other European species, not mentioned by Poty (1984), may belong to that new genus. These are Lithostrotion caespitosum (Martin, 1809) sensu Dobrolyubova (1958) from the Moscow Basin and the British specimens of Diphyphyllum fasciculatum. Poty’s (1984) suggestions published in a short symposium paper cannot be accepted as final for the evolutionary pattern within the family Lithostrotionidae. However, Poty’s (1984) paper is important in drawing attention to the species outside Great Britain that potentially belong to Nemistium or resemble it in morphology. The brief analysis below documents our attitude to those questions.
Caryophyllea fasciculata Fleming, 1828 was assigned by Hill (1938–1941) to Diphyphyllum. Two characters legitimize its relationship with Nemistium (all citations from Hill 1938–1941: 183): “Axial tabulae flattened, domes, their axial parts turning steeply downward either to abut on lower tabulae or to proceed into the outer tabularium”, “the columella is occasionally present, seen as thin lenticle”. Two other characters, i.e., the incompleteness of the columnotheca described above as axial parts of some tabulae extending to the dissepimentarium and the “cardinal septum frequently shorter than others” constitute the most important difference from N. edmondsi; at least as far as the existing data suggest. The “prevalence of axial increase with 2–4 offsets arising simultaneously” will be an argument against the assignment to Nemistium and the inclusion in the new unnamed genus mentioned above, if the mode of axial increase is confirmed. However, Hill (1938–1941) did not illustrate the lectotype chosen by Smith and Lang (1930) and characterized by Kato (1971: 4) in his redescription of Fleming’s (1828) species as follows: “Axial structure is completely absent. Increase is by ‘fission’”. The latter expression can perhaps be understood as the axial increase mentioned by Hill (1938–1941). Although the existing illustrations do not confirm that increase, two independent opinions allow the exclusion of the peripheral increase typical for Nemistium. The differences prevail over the similarities and therefore Caryophyllea fasciculata is not assigned by us to Nemistium.
All four species described by Dobrolyubova (1958) and listed above resemble Nemistium in having major septa slightly amplexoid, equal in length (including the protosepta) and the columnotheca either continuous (D. fasciculatum) or only slightly broken by rare, sinuous tabulae extended to the dissepimentarium (Lithostrotion cespitosum, L. scoticum, Diphyphyllum gracile). Offsetting specimens were not illustrated by Dobrolyubova (1958) in D. gracile, but the offsetting is axial-like in all remaining species, being closely comparable to the offsets in Lithostrotion (Siphonodendron) dobroljubovae Khoa, 1977. Most Russian species were derived from upper Viséan deposits (Aleksinskiy to Venevskiy horizons). Rare specimens of D. gracile and D. fasciculatum were collected from the Taruskiy and Steshevskiy horizons (lower Serpukhovian, Pendleian). The mode of offsetting and the incomplete columnotheca make the Russian species of D. fasciculatum comparable to the British Caryophyllea fasciculata (= Diphyphyllum fasciculatum) to an extent which makes their conspecific position acceptable.
Kato (1971: 5, pl. 1: 1, 2) redescribed Caryophyllea affinis Fleming, 1828, chose its lectotype, illustrated part of that colony and suggested its position within the genus Siphonodendron. He did not illustrate an offsetting corallite, but described the increase as follows: “New buds appear in the dissepimentarium as a vesicular portion for the first time”. Two facts make it possible to contest his conclusion “So the increase is peripheral.”, and to suppose lateral increase in the topotype colony: (i) The parent corallite skeleton disappears from the axial part of peripherally offsetting corallites, whereas offset in C. affine “appear in the dissepimentarium” as Kato (1971: 5) mentioned, i.e., following exactly the lateral increase of dissepimented corals. (ii) Groups of young corallites are invariably observed above the level of offsetting in peripheral increase whereas a single young corallite accompanies a mature one in lateral increase. Such a single young corallite attached to a mature one is illustrated by Kato (1971: pl. 1: 1) in the lectotype of C. affine. Thus, the increase in that species is here accepted as lateral, allowing one to accept its position within the genus Siphonodendron, as suggested by Kato (1971), but not in Nemistium, as suggested by Poty (1984). The lateral increase documented by Khoa (1977: fig. 17) in the Polish specimen assigned by that author to Lithostrotion (Siphonodendron) affine supports that conclusion to some extent.
Khoa (1977: 341, figs. 19–22) introduced the new species Lithostrotion (Siphonodendron) dobroljubovae from the Brigantian (upper Viséan) of Poland and included Lithostrotion scoticum Hill, 1940 of Dobrolyubova (1958) and, questionably, Diphyphyllum lateseptatum McCoy, 1851 of Vassilyuk (1960) in its synonymy. The slightly incomplete columnotheca with the cardinal septum shortened, the axial-like increase and the occurrence of partition but not dividing wall between the offsets in their early growth stage, are the most important characters of that thoroughly investigated species. Those characters combined with the detailed description and illustration of the remaining features, would allow the acceptance of that species as the type for the new, unnamed genus mentioned above.
Semenoff-Tian-Chansky (1985: table 9, pl. 15: 6a, b) listed and illustrated one colony from the upper Serpukhovian or lower Bashkirian strata in Algeria (North Africa) and identified it as Nemistium sp. The specimen illustrated has a continuous columnotheca, the major septa equal in length and an axial structure consisting of median lamella and short septal lamellae. All those characters correspond to the type species of Nemistium. However, the only offsetting specimen illustrated by Semenoff-Tian-Chansky (1985) does not allow us to establish the mode of its increase. It may be either lateral, if the largest corallite is parental or axial-like if that corallite is ontogenetically more advanced than the two remaining offsets. Thus, the taxonomic position of that colony and the occurrence of Nemistium in North Africa, although probable, still remains to be proven.
Poty (2002: pl. 1: 7) illustrated and named Nemistium sp. from the upper Viséan of Nova Scotia (Canada). He gave no description for this very small, restricted fragment of a colony, represented by a single transverse section. Probable peripheral offsetting, the presence of a median lamella in the mature corallite and its absence from the offset point towards Nemistium, but lack of a longitudinal section and the restriction in size of the specimen preclude a firm identification of that colony. Nevertheless, its position within the genus Nemistium is very probable.
Fan (in Fan et al. 2003) described Diphyphyllum fasciculatum (Fleming, 1828) and Tschussovskenia gerzeensis Fan, 2003 from the Viséan strata of Tibet, which may either belong or be related to Nemistium. The almost complete columnotheca and the inconsistent median lamella suggest that possibility. Unfortunately, offsetting corallites are not illustrated and the mode of increase is not mentioned in the descriptions. These inadequately studied corals are the only Nemistium-like species found by us in the Chinese literature. They are included in this analysis as potentially important for rugose coral palaeogeography when studied in detail.
Denayer (2014) identified as Nemistium cf. affine (Fleming, 1828) a single, poorly preserved colony derived from a loose block in the Kisla section (Turkey) and modified Hill’s (1981: F383) diagnosis of the genus. Some characters introduced by him in that new diagnosis are absent from the type species. These are: “major septa long, reaching the axis or not”. The septa in N. edmondsi are slightly amplexoid, but they do not reach the corallite axis. Dissepiments “V-shaped and herringbone”. Rectangular dissepiments are prevalent in N. edmondsi whereas the two kinds noted by Denayer (2014) are absent from the illustrations of Smith (1928: pl. 5: 1, 2). “Increase axial, parricidal, bipartite, tripartite or quadri-partite”, if followed, would eliminate at least one paratype colony of N. edmondsi from the genus Nemistium. That colony produced five offsets (Smith 1928: pl. 5: 2). Also, the term “axial”, introduced by Denayer (2014) is incorrect for N. edmondsi. Its increase is peripheral as correctly indicated by Hill (1981) in her diagnosis of Nemistium. The substantial difference between those two modes of offsetting was previously diagnosed by Fedorowski and Jull (1976: 40, 41). “Minor septa long” is one of the correct items introduced by Denayer (2014) in his emended diagnosis. However, minor septa are commonly variable within the framework of particular lithostrotionid genera and cannot be considered as diagnostic at the genus level. Their length bears some diagnostic value for species, but only when compared to the length of the major septa or the width of the dissepimentarium. Denayer’s (2014) diagnosis is rendered doubtful by the above statements and by the absence of a character so important as the occurrence of the continuous columnotheca of Fedorowski (2009). Also, the latter feature was described by Hill (1981: F383) as “strongly arched axial tabulae each resting on the arch below”. Thus, we follow Hill’s (1981) diagnosis, supplemented by the data from both the type species and Nemistium liardense sp. nov.
Denayer (2014: fig. 16: Ca–c) appears to be correct in considering the increase as “axial” in his poorly preserved colony, although that increase should rather be referred to as axial-like when compared to that in Stauria (see Smith and Ryder 1927: pl. 9). The walls of the offset in Denayer’s (2014) colony, which are most probably partitions, are directly joined at the parent corallite‘s axis. The mode of increase, the occurrence of a cardinal fossula with the cardinal septum shortened, and the lack of an unquestionably continuous columnotheca in longitudinal section makes the Turkish species closely comparable with Lithostrotion (Siphonodendron) dobroljubovae Khoa, 1977. It is also comparable in most characters to the Russian species described by Dobroljubova (1958), except for the shortened cardinal septum, but those characters exclude it from Nemistium. Thus, the phrase “All the generic characters of Nemistium Smith, 1928 are present in that [i.e., Turkish] specimen.” (Denayer 2014: 262) does not find support in the morphology of that specimen.
The following should be pointed out to sum up this discussion: (i) Nemistium sp. of Semenoff-Tian-Chansky (1985) and Nemistium sp. of Poty (2002) may belong to that genus if the peripheral offsetting in the former and the presence of a columnotheca in the latter species are confirmed. Both are included here in the paleogeographical considerations below. (ii) The following taxa should be grouped together in a new genus closely related to Nemistium: Lithostrotion (Siphonodendron) dobroljubovae Khoa, 1977 [with Lithostrotion caespitosum of Dobrolyubova (1958) and D. lateseptatum of Vassilyuk (1960) as synonyms], Diphyphyllum fasciculatum of Dobrolyubova (1958), Nemistium cf. affine of Denayer (2014) and Diphyphyllum fasciculatum of Hill (1938–1941), all of which have an incomplete columnotheca and axial-like increase. The main differentiating features with this group are the presence or absence of shortening of the cardinal septum and the presence or absence of a cardinal fossula. Those differences may be sufficient to subdivide them into two subgenera, but this question is beyond the scope of the present paper. The occurrence of a partition in the very early stage (Khoa 1977), but the lack of a dividing wall (Fedorowski and Jull 1976) is of special significance in that group of corals. (iii) Caryophyllea affinis Fleming, 1828, transferred to Nemistium by Poty (1984) is here accepted within Siphonodendron as suggested by Kato (1971).
Colonies described here from both the Liard Basin and the Stikine Accreted Terrane bear all of the main diagnostic characters of Nemistium, except in their axial structures. The main axial features differentiating them from N. edmonsi are the presence of a simple, interrupted median lamella, very seldom accompanied by short septal lamellae, and a cardinal fossulae (?) developed temporarily in rare corallites. We do not consider those differences important enough to assign the North American specimens to a separate genus or subgenus. However, the trabecular septal microstructure established in our specimens (see description) may constitute a substantial generic difference if that character is absent from N. edmondsi. However, we conditionally included that character in the emended diagnosis of Nemistium.
Nemistium liardense sp. nov.
Figs. 4–7.
ZooBank LSID: urn:lsid:zoobank.org:act:8AA4EF27-438D-4449-8B 20-5EF589E63209
Etymology: Named for its occurrence in the Liard Basin.
Holotype: GSC 140520, fragments of four colonies.
Type locality: GSC locality C-442050, Mattson Gap section 20RAH9, Northwestern Territories, Canada.
Type horizon: Upper member Mattson Fm. between 1056.7 and 1057.1 m above base (1021.64 to 1022.04 m above base Mattson Fm.; 401.03 to 401.43 m above base upper member); Foraminiferal Biozone 20 of Mamet and Ross in Bamber and Waterhouse (1971), Mamet (1976).
Material.—Ten thin sections and five peels studied from Liard Basin type locality (GSC 140520–140523); also, several thin sections (GSC 140524, 140525) from Stikine Accreted Terrane (GSC locality C-442050, Foraminiferal Biozone 20 of Mamet 1976, lower Bashkirian) examined and some illustrated in part for comparison. Colonies ranging in approximate size from 7 × 9 × 13 cm to 1 × 14 × 20 cm. Corallites silicified in part. Preservation inadequate for serial sectioning with acetate peels clear enough for blastogenetic study. Slightly altered septal microstructure locally preserved in parts of some corallites. Some or many corallites in each colony crushed by compaction.
Diagnosis.—Nemistium with n:d value of 23–25 × 2 (rarely 27 × 2), septa at 6.2–7.5 mm (rarely 10 mm) corallite diameter; median lamella thin, interrupted, absent from early growth stage of offsets; 1–3 short, thin septal lamellae may rarely occur; minor septa ranging in length from short, thin plates at periphery to penetrating tabularium; 1–3 rows of rectangular, interseptal dissepiments.
Description.—Corallites slender, densely packed in sections cut just above level of offsetting; distance between corallites increases during colony growth; very large corallites rare (Figs. 4A1, B2, D2, C2, 7A2). Major septa slightly amplexoid, radially arranged, slightly thickened at tabularium border, generally equal in length. Cardinal fossula-like structure in very few corallites, marked by reduction of dissepiments (Fig. 6C). Minor septa differentiated in length both within individual transverse sections of a given corallite and between corallites; may penetrate tabularium or be restricted to peripheral parts of corallites within sections of a single colony. Median lamella thin, mostly short, interrupted, in many corallites reduced to few short plates either attached to upper surfaces of tabulae or intersecting two to several tabulae, long lasting median lamellae rare, present mainly in colonies from Stikine Terrane (Fig. 4A2, B1 vs. 4C1, D1), extension of median lamella above tabulae (i.e., calice floor), allows it to be called a pseudocolumella; in rare instances it may protrude so as to be present in middle of offsetting specimen, from which the polyp’s body is absent at that level (Fig. 5C); derivation of pseudocolumella from septa observed in rare young corallites (Fig. 6B1), but its relationship to protosepta not established. Very weak axial structure, consisting of thin pseudocolumella, one to three septal lamellae and axial tabellae exceptional (Fig. 6B2, B3). Dissepiments interseptal, regular in 1–3 rows; peripheral row largest, vertical in longitudinal section.

Fig. 4. Thin sections of lithostrotionid rugose coral Nemistium liardense sp. nov. from Stikine Terrane, Northwestern Territories, Canada, Mattson Formation, lower Bashkirian (A, B), and Liard Basin, Northwestern Territories, Canada, Mattson Formation, perhaps lower Bashkirian (C–E). A. GSC 140524, transverse (A1) and longitudinal (A2) section. B. GSC 140525, transverse (B1) and longitudinal (B2) section. C. GSC 140522, longitudinal section of two corallites (C1) and transverse section (C2). D. GSC 140523, longitudinal section of offsetting corallite with strong pseudocolumella (D1) and transverse section of part of colony (D2). E. GSC 140520, holotype, oblique section of two young corallites.
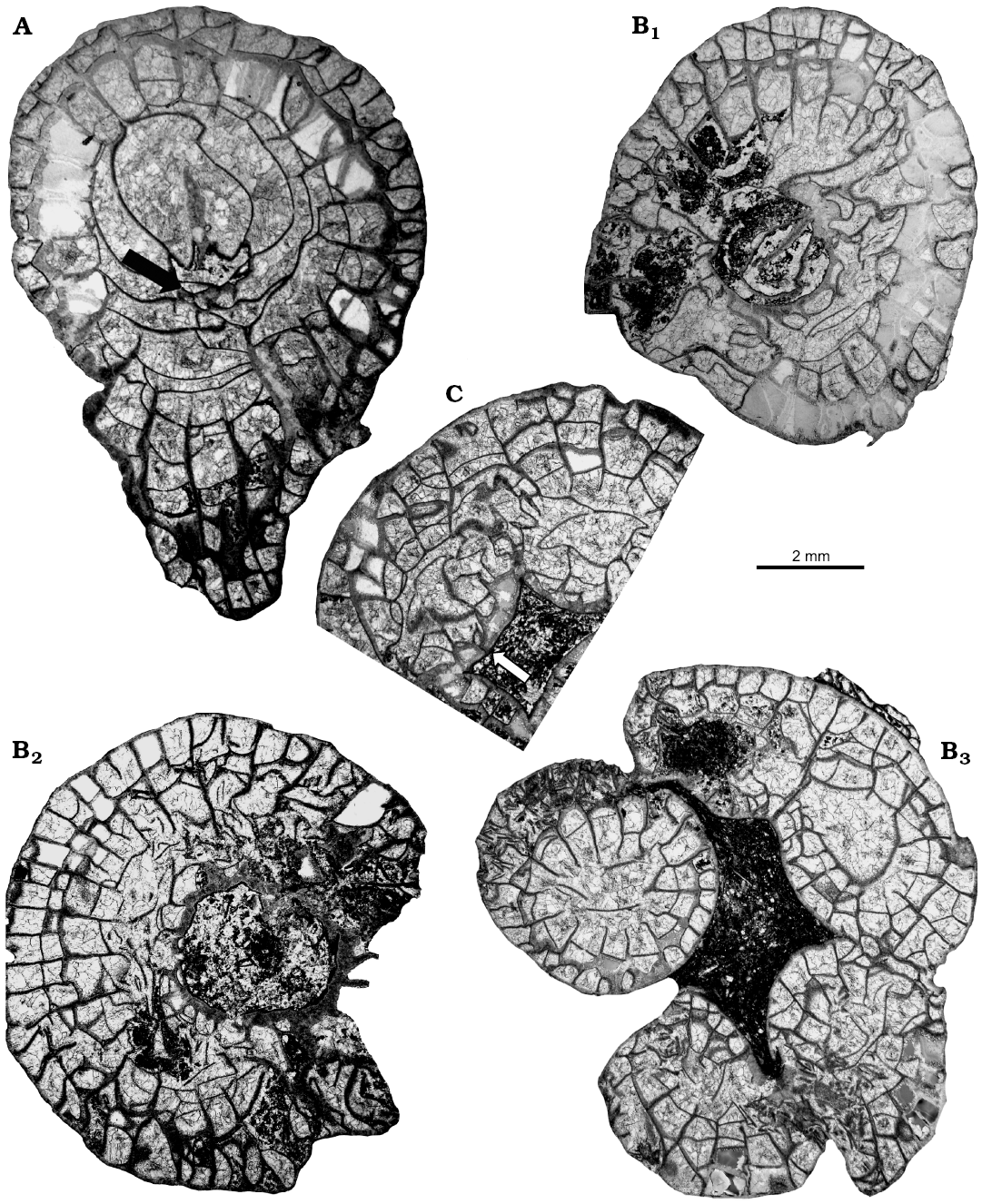
Fig. 5. Transverse thin sections of lithostrotionid rugose coral Nemistium liardense sp. nov. from Liard Basin, Northwestern Territories, Canada, Mattson Formation, perhaps lower Bashkirian. A. GSC 140520, holotype, beginning of offsetting (arrow). B. GSC 140523, early (B1, B2) and advanced (B3) development of offsets. C. GSC 140522, offset with elongated septum at early growth stage (arrow).
Offsetting peripheral; formation and separation of particular offsets sequential in their individual, slightly differentiated timing (Fig. 5A, B); beginning of offsetting results in changes difficult to recognize in transverse sections (Fig. 5A, arrow), but well documented in longitudinal sections (Figs. 4D1, 6A3, 7A1). Each offset inherits peripheral part of parent’s skeleton including peripheral tabularium, dissepimentarium and septa, i.e., “atavotissue” (this and following terms in inverted comas after Smith and Ryder [1927], followed in part by Hill [1935]). Thus, all septa on peripheral margins of offsets are “atavosepta”. Walls between offsets absent in early stage of offsetting (Fig. 5B1, B2). Uppermost basal skeleton of parent corallite becomes modified (Figs. 4D1, 6A3, 7A1); particularly the axial part of columnotheca. Those modifications, slightly differentiated in terms of arrangement of tabellae in parent/offset intermediate area of individual offsets, lead to formation of offset’s columnotheca. At very early growth stage (Figs. 6A3, 7A1), last parential tabulae indicated by black arrows and offset’s “neotheca” marked by white arrow; peripheral tabularium absent from “neotheca” in offsets (Fig. 7A1). It appears as “neotissue” during further growth of offsets. All septa inserted in “neotheca” are “neosepta”. Median lamella or pseudocolumella almost invariably absent from offsets during their early growth stage (Figs. 4E, 5B3, 6B4); it appears at various growth levels of offset, but not until complete isolation of offset in overwhelming number of instances observed, except for possibly one (Fig. 5C, lower offset); one major septum in that offset, which is lengthened to reach the axis, can be interpreted as forming median lamella. Median lamella arose from inner margin of major septum in another, ontogenetically more advanced offset (Fig. 6B1); formula of that septum (cardinal or counter) not established. Connection of median lamella to septum rarely observed; thus, it may last a very short time; median lamella free from all septa in almost all corallites studied. Formation of dividing wall of Fedorowski and Jull (1976) begins from depressions in “neotheca” on both sides of partition (Fig. 5C, B3). Those depressions separate middle line of dividing wall, step by step up to its complete division into two external walls of young corallites. Trabeculae present in partition are replaced by cryptocrystalline peripheral line of external offset’s wall, which is typical for every basal skeletal element in corals, as documented independently by Wells (1969) and Sorauf (1970). Remnants of parent’s skeleton and tissue may persist in some corallites until they accompany separated offsets (Fig. 6B4, arrow). Offsetting in Nemistium liardense sp. nov. resembles that in Tizraia? sp. from Bashkirian strata of Canadian Arctic Archipelago, described in detail by Fedorowski et al. (2012). Unfortunately, sequence of septal insertion in offsets of N. liardense sp. nov. could not be established. Microstructure of septa is trabecular; trabeculae approximately 0.02 mm thick, long (as documented in oblique sections of septa), and rather loosely arranged (Fig. 6A1, A2). One peculiar character in the relationship between the septa, dissepiments or tabulae, and the distribution of sclerenchymal sheets should be pointed out. Some basal skeletal structures join the sclerenchymal covering of septa directly (Fig. 6A2, black arrow), or appear to be a continuation of those coverings (Fig. 6A1, black arrows). Sclerenchymal coverings in other instances may be locally isolated from a primary septum (Fig. 6A1, white arrow). Also, sclerenchymal coverings of septa and basal skeletal structures may meet without a clear border (Fig. 6A2, white arrow). Reasons for apparent differences in distribution of sclerenchyme unknown.
Remarks.—There are two main characters of Nemistium liardense sp. nov. that distinguish it from the type species: (i) the minor septa are strongly differentiated in length, with some restricted to the most peripheral parts of corallites and (ii) the axial structure is reduced to a single, thin pseudocolumella, which appears only sporadically in most corallites. The rare incompleteness of the columnotheca (Fig. 4C1, left corallite) may have resulted either from the obliqueness of the longitudinal section or from changes leading to the formation of offsets. This especially concerns the colonies from the Mattson Fm. In colonies from the Stikine Terrane both those characters are better developed, forming the main difference between those two groups of colonies of N. liardense sp. nov. We do not consider those features adequate for either distinguishing between those two groups as different species or distinguishing them from N. edmondsi as a different genus or subgenus. Specimens derived from the Stikine Terrane are more numerous and much more diversified than those from the Mattson Fm. (JF and EWB preliminary identifications, unpublished material), but their description is beyond the scope of the present paper.
Stratigraphic and geographic range.—Type locality and horizon only.
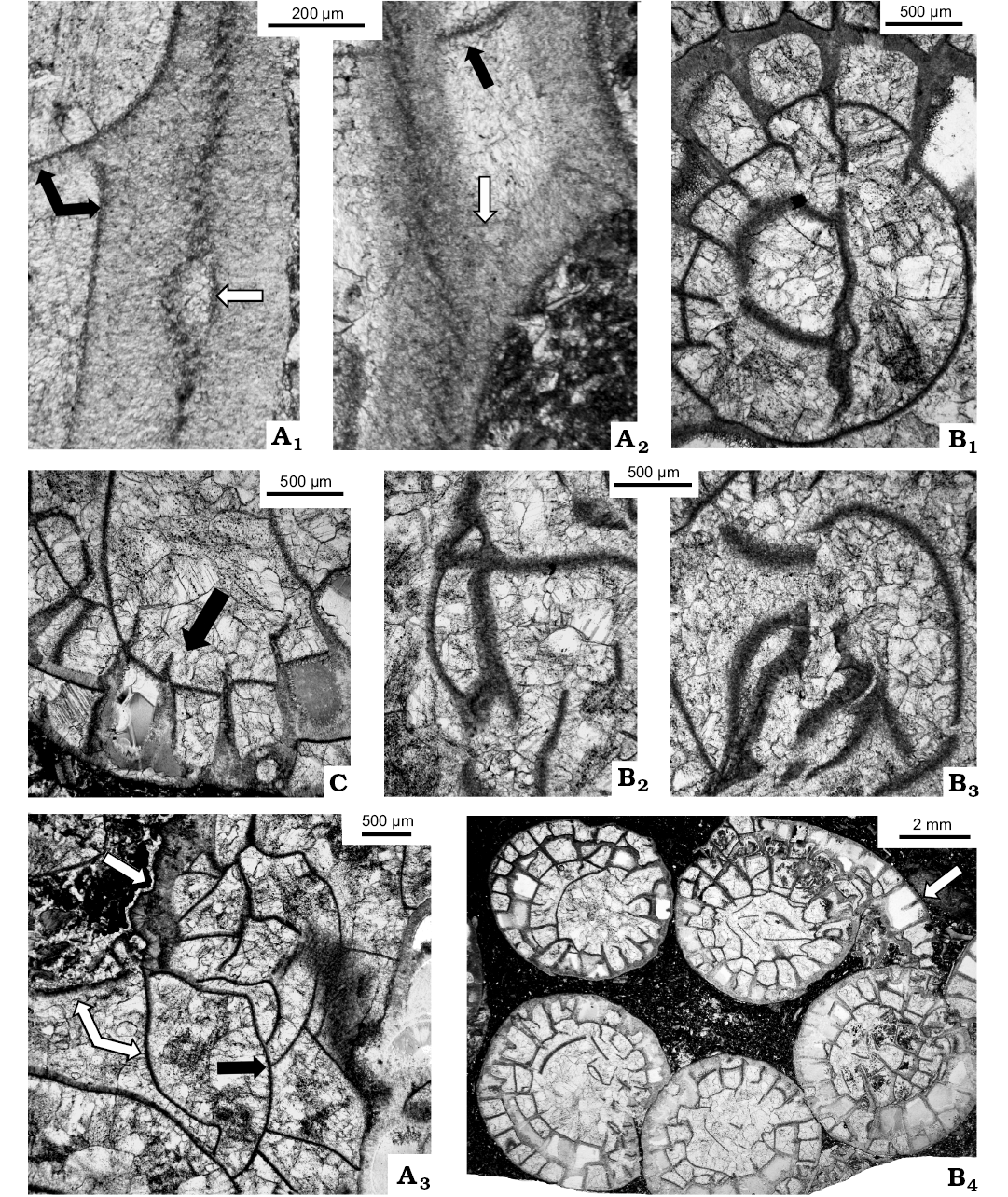
Fig. 6. Transverse thin sections (except A3, which is longitudinal section) of lithostrotionid rugose coral Nemistium liardense sp. nov. from Liard Basin, Northwestern Territories, Canada, Mattson Formation, perhaps lower Bashkirian. A. GSC 140520, holotype. Microstructure in transverse (A1) and oblique (A2) sections; in A1: black arrows, tabula apparently continued sclerenchymal cover of septum; white arrow, sclerenchymal cover isolated from primary septum; in A2: black arrow, tabula joins sclerenchymal cover directly; white arrow, lack of border between sclerenchymal cover of septum and tabula; A3, longitudinal thin section, beginning of offsetting; two lower white arrows, last parent’s tabulae; black arrow, first offset’s tabula; neotheca indicated by upper white arrow. B. GSC 140523, median lamella united with major septum; slight break caused diagenetically (B1), axial structure consisting of median lamella and irregular tabellae (B2, B3), offsets separated from each other completely, but rudiment of parent’s skeleton continued to occur (arrow) (B4). C. GSC 140521, lack of dissepiments in one loculum (arrow) suggests cardinal fossula.
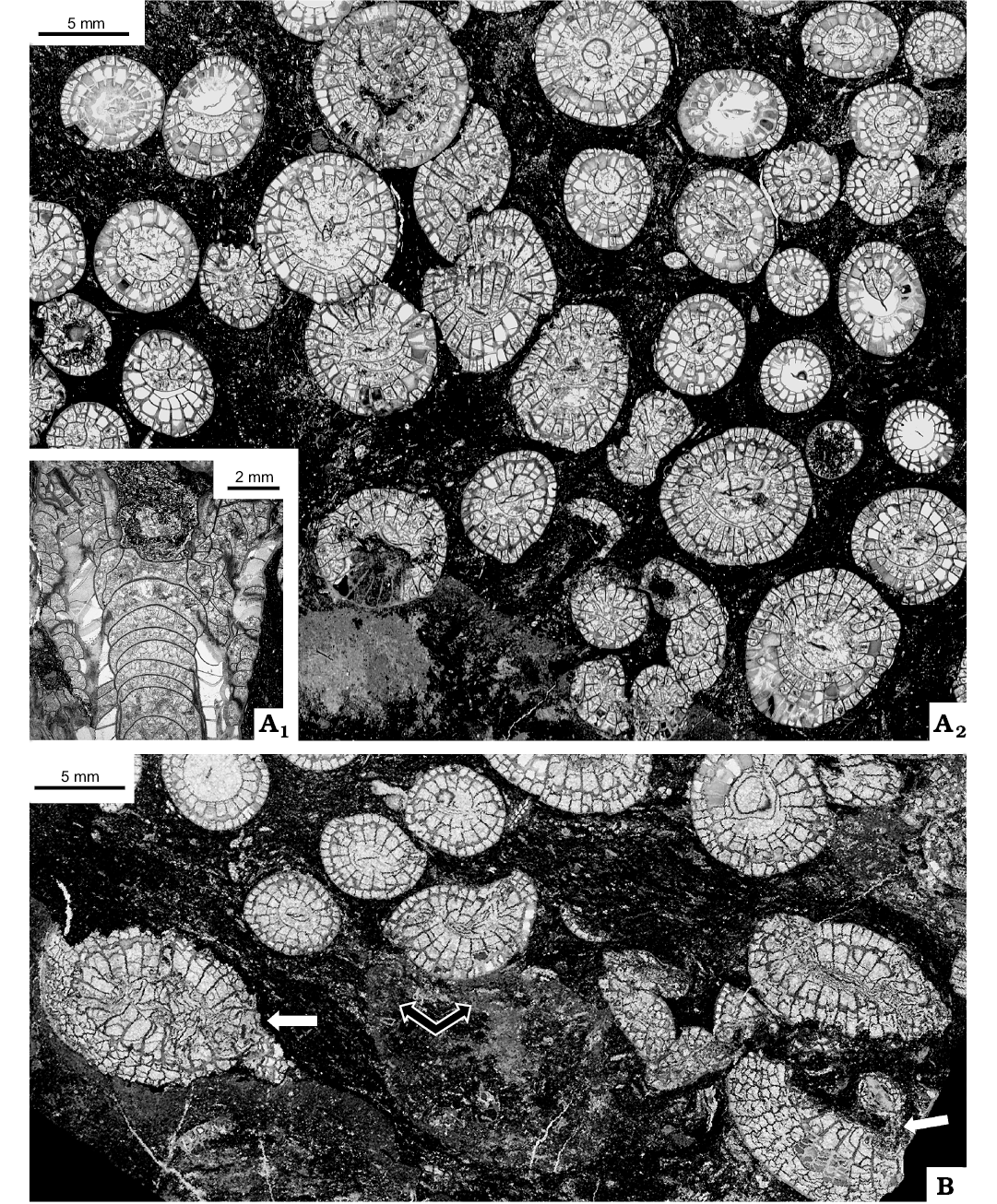
Fig. 7. Lithostrotionid rugose coral Nemistium liardense sp. nov. from Liard Basin, Northwestern Territories, Canada, Mattson Formation, perhaps lower Bashkirian. A. GSC 140520, holotype, longitudinal section of offsetting corallite (A1), transverse section of colony fragment (A2). B. GSC 140526, transverse section showing broken corallites (white arrows) at periphery of colony of Heritschioides simplex sp. nov.; upper part (black arrows) indicates fragment of colony of Nemistium liardense sp. nov.
Suborder Aulophyllina Hill, 1981
Family Aulophyllidae Dybowski, 1873
Subfamily Heritschioidinae Sando, 1985
Genus Heritschioides Yabe, 1950
Type species: Waagenophyllum columbicum Smith, 1935; Blind Creek Limestone near Keremeos, southern British Columbia, Canada; late Serpukhovian–early Bashkirian.
Heritschioides simplex sp. nov.
Figs. 7B, 8, 9.
ZooBank LSID: urn:lsid:zoobank.org:act:3E992402-6358-49F2-8644-6ED18C91AC1D
Etymology: From Latin simplex, simple; after simplified morphology.
Holotype: GSC 140526, one fragment of worn colony.
Type locality: GSC locality C-442050, Mattson Gap section 20RAH9, Northwestern Territories, Canada.
Type horizon: Upper member Mattson Fm. between 1056.7 and 1057.1 m above base Mattson Gap section 20RAH9 (1021.64 to 1022.04 m above base Mattson Fm. and 401.03 to 401.43 m above base upper member); Foraminiferal Biozone 20 of Mamet and Ross in Bamber and Waterhouse (1971).
Material.—Holotype, colony measuring approximately 9 × 11 × 15 cm. Corallites partly silicified, some compressed to a variable extent. Six thin sections available for study.
Diagnosis.—Heritschioides with n:d value variable; maximum septal number 29 × 2, maximum corallite diameter 23 × 18.5 mm; major septa stop short of simple axial structure with long thin median lamella; extra septal lamellae either absent or very restricted in number; most minor septa restricted to dissepimentarium, which occupies approximately ¼ corallite radius; cardinal fossula distinct, bordered by two parallel major septa.
Description.—Corallites loosely and irregularly arranged; twins occur (Fig. 8A8, arrow) external walls of corallites thin. Septa attached to wall rather than penetrate it (Fig. 8A4, A5). Major septa thin in dissepimentarium, strongly wavy, rarely interrupted by small lonsdaleoid dissepiments (Fig. 8A4), their tabularial sectors slightly dilated, mostly next to slightly thickened inner wall; they thin gently toward axis, most major septa withdrawn from axial structure, rarely temporarily united with corresponding lamellae, in part of one corallite major septa “doubled” (Fig. 9A2, A5), with additional septal blades present in tabularium not corresponding to minor septa, their occurrence and function doubtful. Minor septa closely resemble peripheral sectors of major septa in morphology, most restricted to dissepimentarium, rarely cross it slightly. Cardinal septum shortened, but may be met or closely approached by elongated, thin extension of median lamella (Fig. 8A3, A2, respectively). Counter septum equal in length and thickness to adjacent major septa in most corallites, may be slightly elongated (Fig. 8A1, A2). Cardinal fossula shallow, but distinct; bordered by two major septa commonly equal to remaining ones in length and thickness (Figs. 8A1–A3, 9A2). Axial structure variable in morphology, commonly simple, consisting of long, thin median lamella, several septal lamellae and numerous axial tabellae; arrangement, length and number of septal lamellae and sections of axial tabellae on either side of median lamella may differ greatly (Figs. 8A2, 9A2); extra septal lamellae not found, except for problematic one or two in one corallite. Crushing of parts of skeleton precludes firm documentation of axial column in longitudinal section (Figs. 8A6, 9A1). However, obvious difference in arrangement between incomplete peripheral tabulae, almost semispherical, and axial tabellae elevated steeply towards median lamella, suggest occurrence of isolated axial column. Microstructure of septa finely trabecular as documented both in transverse and longitudinal sections (Fig. 9A3, A5, respectively), i.e., matches that in H. columbicum (Smith, 1935) (Fedorowski et al. 2014b: fig. 7: 4, 7); lack of sclerenchymal sheets in part of septum documents original width of primary septa (Fig. 9A3, arrow). Offsetting lateral (Fig. 8A6), but details in septal insertion unknown; pinnacles seen at parent/offset border (Fig. 8A7) suggest general similarity to offsetting documented in detail by Fedorowski et al. (2014a, b).
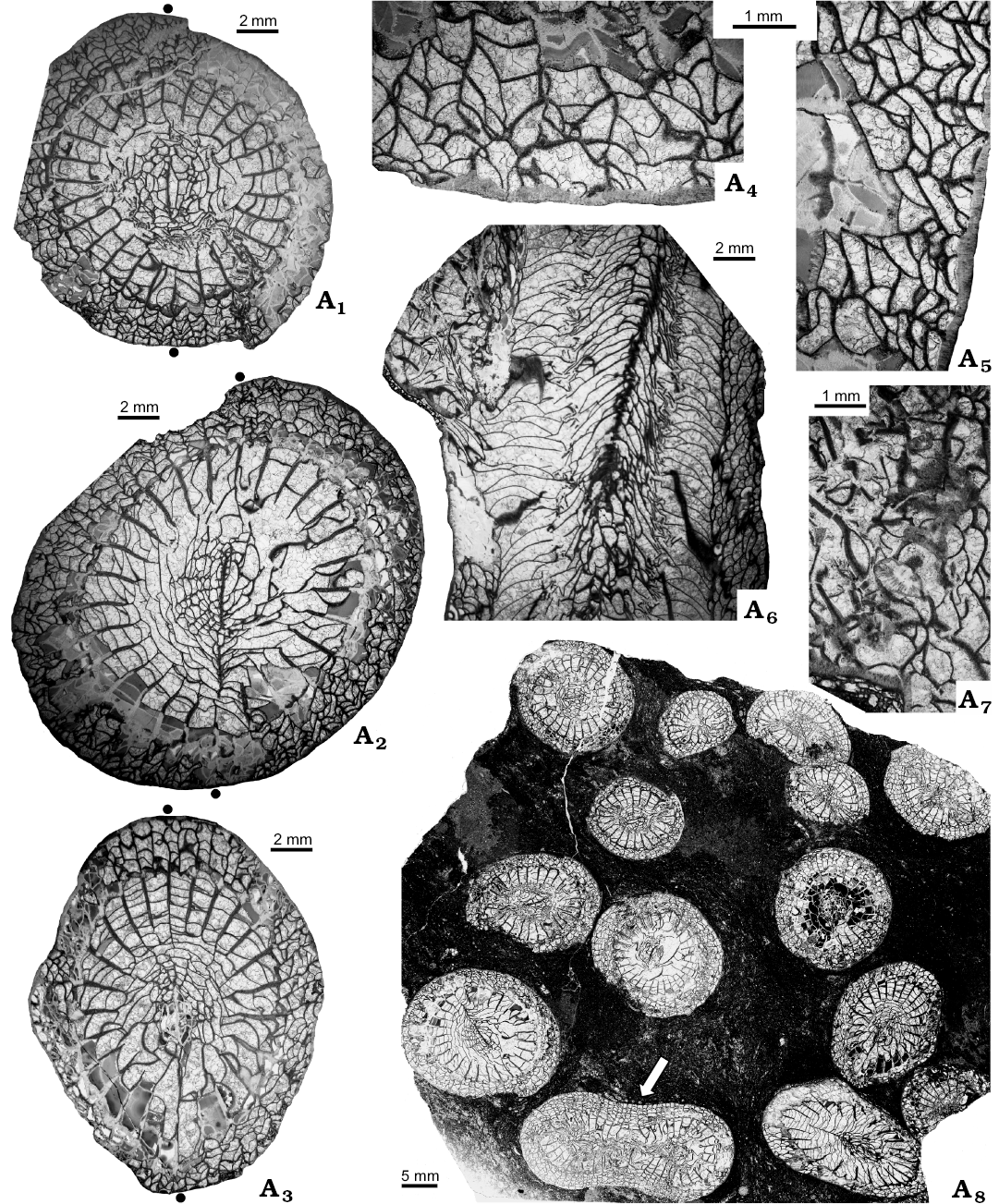
Fig. 8. Transverse (A1–A5, A8) and longitudinal (A6, A7) thin sections of aulophyllid rugose coral Heritschioides simplex sp. nov. (GSC 140526, holotype) from Liard Basin, Northwestern Territories, Canada, Mattson Formation, perhaps lower Bashkirian. Corallites in different growth stages (A1–A3), dissepimentarium and peripheral parts of septa (A4, A5), laterally offsetting corallite (A6), detail of parent/offset common tissue (A7), fragment of colony with twins (arrow) (A8). Black dots correspond to cardinal (lower) and counter (upper) septa.
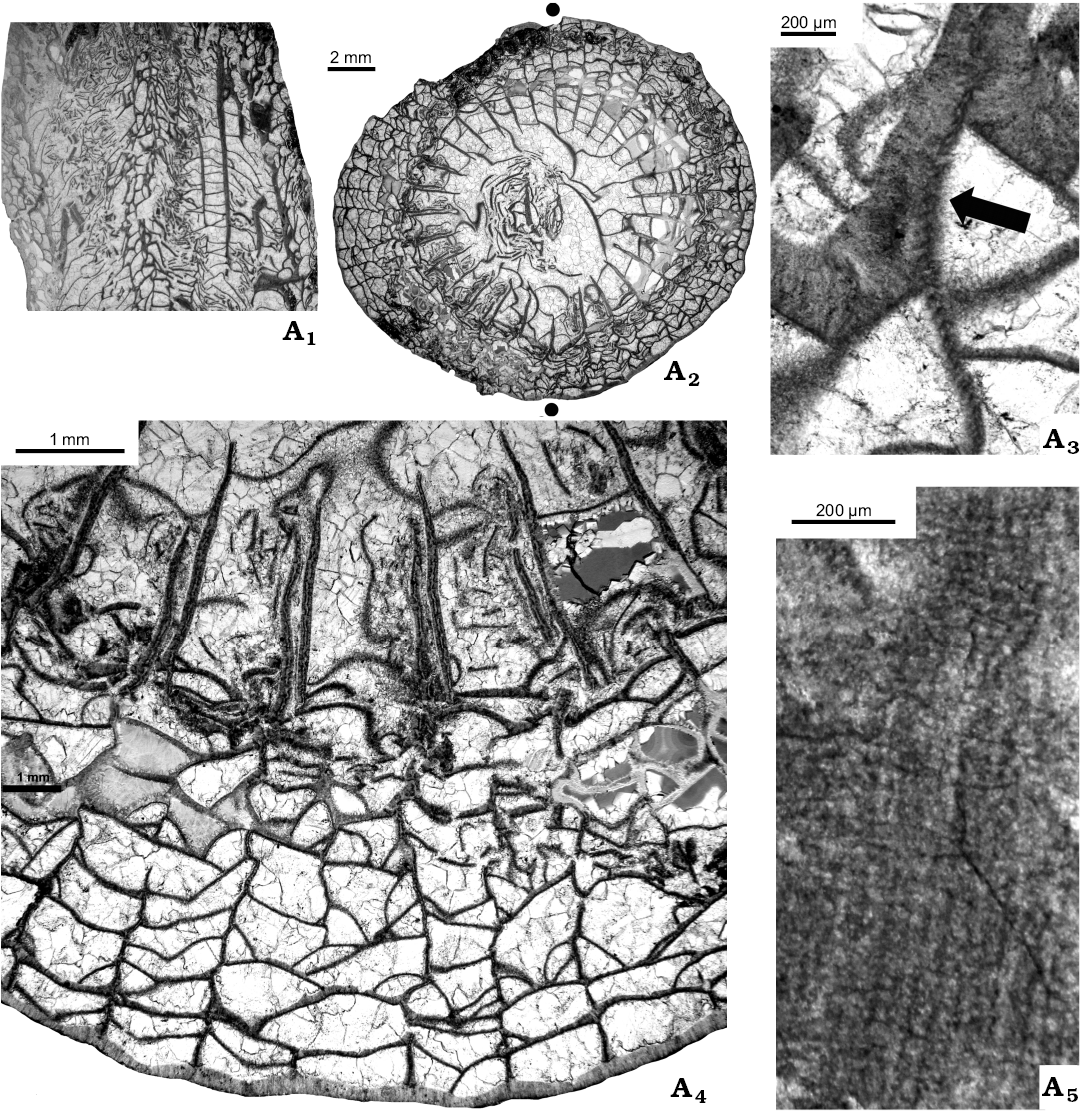
Fig. 9. Transverse (A2–A4) and longitudinal (A1, A5) thin sections of aulophyllid rugose coral Heritschioides simplex sp. nov. (GSC 140526, holotype) from Liard Basin, Northwestern Territories, Canada, Mattson Formation, perhaps lower Bashkirian. Trabecular microstructure of septum and sclerenchymal cover unequally distributed (A1–A3), detail demonstrating doubled major septa (A4), microstructure of septum (A5); arrow in A3 points to primary septum lacking sclerenchymal cover. Black dots correspond to cardinal (lower) and counter (upper) septa.
Remarks.—In view of the fairly recent revision of Waagenophyllum columbicum Smith, 1935, the type species of the genus Heritschioides (Fedorowski et al. 2014a), supplemented by descriptions of species from Alaska (Armstrong 1972; Fedorowski et al. 2014b), no discussion is given here of the diagnostic characters of Heritschioides and its taxonomic relationships. Data from the above papers allow both a close comparison of Heritschioides simplex sp. nov. with the known species of that genus and a suggestion of its probable stratigraphic position. The redescription and reillustration of Smith’s (1935) type collection and the study of topotype material (Fedorowski et al. 2014b: figs. 2: 1–7; 5: 8, 9; 7: 7 and figs. 3: 1–8; 4: 1–5; 5: 1–7, 10, 11; 6: 1–8; 7: 1–6; 8: 1–4) document large intraspecific variability in the type species and thus the possibility of comparison with taxa having both complex and simple morphology. Four? characters in common between the holotype and the paratype of H. columbicum and H. simplex sp. nov. are of special value in the context of the present paper. These are: (i) a very similar trabecular septal microstructure, (ii) a restricted number or absence of extra septal lamellae from the axial structures of mature corallites, and (iii) a comparatively narrow and simple morphology of the dissepimentarium and (iv) similar morphology of the tabularium. Much more complex dissepimentaria and more common occurrence of extra septal lamellae in the topotypes were accepted by Fedorowski et al. (2014a) as intraspecific variants, but they also may be considered as adequate for separation of a new species. The main differences between the simplest specimens of the type species and the specimen described here include different n:d values, (i.e., larger corallite diameter and smaller septal number), much shorter major septa that seldom reach the simple axial structure, and less steeply orientated axial tabellae in the axial column of H. simplex sp. nov.
Heritschioides separatus Fedorowski and Stevens, 2014b, first described by Armstrong (1972) as the paratype of his species Corwenia jagoensis, is another species of simple morphology to consider. That species, which was collected from the Wahoo Limestone (Atokan) in the Brooks Range (Alaska), resembles H. simplex sp. nov. in several features. These include a narrow, simple axial structure with extra septal lamellae absent or very rare, similar morphology of the dissepimentarium and similar n:d values of mature corallites. In comparison with H. simplex sp. nov., H. separatus shows a common reduction of its median lamella, less regularity in the morphology of its axial structure in longitudinal section, a cardinal fossula that is more difficult to recognize in some corallites and a wider dissepimentarium in its largest corallites. Some of those differences are slight, so that these two species are morphologically close to each other.
The similarity of H. simplex sp. nov. to morphologically simplest and stratigraphically lowest species of Heritschioides and its occurrence along with Nemistium liardense (Fig. 7B, black arrows) suggest that it is probably not younger than early Bashkirian.
Stratigraphic and geographic range.—The type locality and horizon only.
Concluding remarks
The chapter about the geological setting of the coral-bearing deposits documents the position of the rugose coral fauna within the Mattson Fm., whereas, the comparison of that fauna to rugose coral faunas elsewhere in North America, both on the craton and in accreted terranes, allows their placement in a more precise chronostratigraphic context and wider paleogeographic framework. As mentioned in the Material and methods, all the colonies from the Mattson Fm. are worn specimens. The colonies are bioclasts reworked penecontemporaneously with deposition of the limestone unit from which they were collected. They were not eroded from an older lithified limestone bed. In one of the samples (Fig. 7), strong fragmentation and pre-burial deformation of the corallites along the contact zone between the Heritschioides and Nemistium colonies provides additional support for the thesis of the pre-burial transport. Both species are, therefore, suitable for indicating the relative age of the deposits from which they were collected.
The presence of Nemistium liardense sp. nov. is of a special value in determining the age of the upper Mattson Fm. because it also occurs in Foraminiferal Biozone 20 (see Mamet 1976) in the allochthonous Stikine Terrane of British Columbia. The coral fauna of that terrane, including Nemistium, comprises many specimens and species (JF and EWB preliminary identifications, unpublished material). One of the three species of Nemistium from the Stikine Terrane resembles the colonies from the Mattson Fm. so closely that it is considered conspecific (see Description). The remaining colonies from the Stikine Terrane belong to two new species of Nemistium that are closely related to N. liardense sp. nov. We cannot estimate the exact stratigraphic range of the species from the Stikine Terrane, but their known occurrence is confined to Bashkirian Foraminiferal Biozone 20. Thus, we consider the specimens from both areas to be Bashkirian.
The morphology of H. simplex sp. nov. allows its comparison to both the type species for the genus, dated uppermost Serpukhovian or lower Bashkirian, and H. separatus from the Brooks Range in northern Alaska. The Moscovian species of that genus from Kuiu Island in Alaska are morphologically much more complex (Fedorowski et al. 2014b). That comparison suggests a comparatively old stratigraphic (pre-Moscovian) position for H. simplex sp. nov., indirectly supporting the Bashkirian age determination based on N. liardense sp. nov. In conclusion, the age of the coral-bearing limestone unit in the upper Mattson Fm. lies within Foraminiferal Biozone 20 and is within the early Bashkirian.
Both of the new species described herein provide important paleobiogeographic information about Carboniferous rugose corals. Heritschioides from the Liard Basin supplements the knowledge on connections between the Carboniferous rugose coral faunas flourishing on the western shelves of Laurussia and those living on the accreted terrains of British Columbia, which originated in Panthalassa west of ancestral North America. The presence of Nemistium is important because it confirms the suggestion of Fedorowski and Bamber (2007) that a marine connection existed between the western shelves of Laurussia and the seas of western to central Europe from the late Viséan into the Bashkirian. In Great Britain, Nemistium appeared during the Brigantian.
Acknowledgements
We thank Sergio Rodríguez (Universidad Complutense de Madrid, Spain), Calvin H. Stevens (San Jose University, California, USA), and Tomasz Wrzołek (University of Silesia in Katowice, Sosnowiec, Poland) for their comments on the manuscript. We are indebted to Marta Bartkowiak (Adam Mickiewicz University, Poznań, Poland) for her excellent technical assistance. This research was funded by the National Science Center grant UMO-2016/21/8/01861.
References
Armstrong, A.K. 1972. Pennsylvanian carbonates, paleoecology and rugose colonial corals, north flank, eastern Brooks Range, Arctic Alaska. USGS Professional Paper 747: 1–21. Crossref
Bamber, E.W. and Waterhouse, J.B. 1971. Carboniferous and Permian stratigraphy and paleontology, northern Yukon Territory, Canada. Bulletin of Canadian Petroleum Geology 19: 29–250.
Bamber, E.W., Macqueen, R.W., and Richards, B.C. 1984. Facies relationships at the Mississippian carbonate platform margin, Western Canada. In: E.S. Belt and R.W. Macqueen (eds.), Sedimentology and Geochemistry, Neuvième Congrès International de Stratigraphie et de Géologie du Carbonifère, 1979. Compte-Rendu 3: 461–478. Crossref
Bamber, E.W., Taylor, G.C., and Procter, R.M. 1968. Carboniferous and Permian stratigraphy of northeastern British Columbia. Geological Survey of Canada Department of Energy, Mines and Resources, Paper 68-15: 1–25. Crossref
Braman, D.R. and Hills, L.V. 1977. Palynology and paleoecology of Mattson Formation, northwest Canada. Bulletin of Canadian Petroleum Geology 25: 582–630.
Chung, P. 1993. Conodont Biostratigraphy of the Carboniferous to Permian Kindle, Fantasque, an Unnamed, and Belloy Formations, Western Canada. 189 pp. Unpublished M.Sc. Thesis, University of Calgary, Calgary.
Currie, L.D., Kubli, T.E., and McDonough, M.R. 2000. Preliminary Geology, Babiche Mountain (95C/9), Yukon and Northwest Territories. Geological Survey of Canada Open File Map 3844, scale 1:50 000. [published online] Crossref
Denayer, J. 2014. Viséan Lithostrotionidae (Rugosa) from Zonguldak and Bartýn (NW Turkey). Bulletin of Geosciences 89 (4): 737–771. Crossref
Dobroljubova, T.A. [Dobrolûbova, T.A.] 1958. Lower Carboniferous colonial tetracorals from the Russian Platform. Akademiâ Nauk SSSR. Trudy Paleontologičeskogo Instituta 70: 1–224.
Douglas, R.J.W. 1976. Geology, La Biche River, District of Mackenzie. Geological Survey of Canada Map 1380A, scale 1:250 000. [published online]. Crossref
Douglas, R.J.W. and Norris, D.K. 1976. Geology, Fort Liard, District of Mackenzie. Geological Survey of Canada Map 1379A, scale 1:250 000. [published online]. Crossref
Douglas, R.J.W. and Norris, D.K. 1977. Geology, Sibbeston Lake, District of Mackenzie. Geological Survey of Canada Map 1377A, scale 1:250 000. [published online]. Crossref
Dybowski, W.N. 1873−1874. Monographie der Zoantharia Sclerodermata Rugosa aus der Silurformation Estlands, Nordlivlands und der Insel Gotland. Archiv für Livlands, Estlands, Kurlands 1: 257−414 (1873), 415−532 (1874).
Fallas, K.M., Hynes, G.F., and Lane, L.S. 2003. A GIS Dataset of Geological Features for the Etanda Lakes Map area (95C/16), Northwest Territories and Yukon Territory. Geological Survey of Canada Open File 1802. [published online]. Crossref
Fan, Y., Yu, X., He, Y., Pan, Y., Li, X., Wang, F., Tang, D., Chen, S., Zhao, P., and Liu, J.. 2003. The Late Palaeozoic Rugose Corals of Xizang (Tibet) and Adjacent Regions and Their Palaeobiogeography. 679 pp. National Science Foundation of China. Hunan Science and Technology Press, Hunan.
Fedorowski, J. 2009. Morphogenesis and taxonomic value of the circum-axial skeleton in Rugosa (Anthozoa). Lethaia 42: 232–247. Crossref
Fedorowski, J. and Bamber, E.W. 2007. Remarks on lithostrotionid phylogeny in western North America and western Europe. Österreichische Akademie der Wissenschaften. Schriftenreihe der Erdwissenschaftlichen Kommissionen 17: 251–273.
Fedorowski, J. and Jull, R.K. 1976. Review of blastogeny in Palaeozoic corals and description of lateral increase in some Upper Ordovician rugose corals. Acta Palaeontologica Polonica 21: 37–78.
Fedorowski, J., Bamber, E.W., and Baranova, D.V. 2012. An unusual occurrence of Bashkirian (Pennsylvanian) rugose corals from the Sverdrup Basin, Arctic Canada. Journal of Paleontology 86: 979–995. Crossref
Fedorowski, J., Bamber, E.W., and Stevens, C.H. 2014a. A revision of Heritschioides Yabe, 1950 (Anthozoa, Rugosa), latest Mississippian and earliest Pennsylvanian of western North America. Palaeontologia Electronica 17: 11A.
Fedorowski, J., Stevens C.H., and Katvala, E. 2014b. New Late Carboniferous Heritschioidinae (Rugosa) from the Kuiu Island area and Brooks Range, Alaska. Geologica Acta 12: 29–52.
Ferri, F., McMechan, M., and Creaser, R. 2015. The Besa River Formation in Liard Basin, British Columbia. In: Oil and Gas Geoscience Reports 2015, 1–27. B.C. Ministry of Natural Gas Resources, Victoria.
Fleming, J. 1828. A History of British Animals. XXIII+265 pp. Bell and Bredfute, Edinburgh. Gunning, M.H., Fedorowski, J., and Bamber, E.W. 2006. Paleogeographic significance of mid-Carboniferous limestone, northwestern Stikine terrane, British Columbia. In: J.W. Haggart, R.J. Enkin, and J.W.H. Monger (eds.), Paleogeography of the North American Cordillera: Evidence For and Against Large-scale Displacements. Geological Association of Canada Special Paper 46: 59–69.
Gunning, M.H., Bamber, E.W., Anderson, R.G., Fedorowski, J., Friedman, R., Mamet, B.L., Orchard, M.J., and Rui, L. 2007. The mid-Carboniferous Arctic Lake Formation, northwestern Stikine terrane, British Columbia. Bulletin of Canadian Petroleum Geology 55: 21–50. Crossref
Harker, P. 1963. Carboniferous and Permian rocks, southwestern District of Mackenzie. Geological Survey of Canada, Bulletin 95: 1–102. Crossref
Henderson, C.M., Richards, B.C., and Barclay, J.E. 1994. Permian strata of the Western Canada Sedimentary Basin. In: G.D. Mossop and I. Shetson (eds), Geological Atlas of the Western Canada Sedimentary Basin, 251–258. Canadian Society of Petroleum Geologists and Alberta Research Council, Calgary.
Hill, D. 1935. British terminology for rugose corals. The Geological Magazine 72: 481–519. Crossref
Hill, D. 1938–1941. A monograph of the Carboniferous rugose corals from Scotland. Palaeontographical Society 1938 (91): 1–78; 1939 (92): 79–114; 1940 (93): 115–204; 1941 (94): 205–213.
Hill, D. 1956. Rugosa. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology. Part F: Coelenterata, F233–F324. Geological Society of America, Boulder and University of Kansas Press, Lawrence.
Hill, D. 1981. Coelenterata, Supplement 1. In: C. Teichert (ed.), Treatise on Invertebrate Paleontology, Part F: Rugosa and Tabulata, 1–762. Geological Society of America, Boulder and University of Kansas Press, Lawrence.
Kato, M. 1971. J. Fleming’s species of British Lower Carboniferous corals. Transactions and Proceedings of the Paleontological Society of Japan, New Series 81: 1–10.
Khoa, N.D. 1977. Carboniferous Rugosa and Heterocorallia from boreholes in the Lublin region (Poland). Acta Palaeontologica Polonica 22: 301–404.
Khudoley, A.K. 2003. A GIS dataset of the geological features for the Tika Creek. Map area (95C/10), Yukon Territory and Northwest Territories. Geological Survey of Canada Open File 1777 [published online]. Crossref
Khudoley, A.K. and Fallas, K.M. 2006. Geology of Brown Lake (95C7), Yukon Territory. Geological Survey of Canada A Series Map 2083A, scale 1:50 000 [published online].
Lane, L.S. 2006. Geology Mount Flett, District of Mackenzie, Northwest Territories (95B/12). Geological Survey of Canada Open File Map 5236, scale 1:50 000 [published online].
Lane, L.S. and Hynes, G.F. 2005. Geology Sawmill Mountain (95B/13), District of Mackenzie, Northwest Territories. Geological Survey of Canada Open File 4940, scale 1:50 000 [published online]. Crossref
Leckie, D.A., Potocki, D.J., and Visser, K. 1991. The Lower Cretaceous Chinkeh formation: A frontier-type play in the Liard basin of western Canada. American Association of Petroleum Geologists, Bulletin 75: 1324–1352. Crossref
Lonsdale, W. 1845. Description of some characteristic Palaeozoic corals from Russia. In: R.I. Murchison, E. de Verneuil, and A. von Keyserling (eds.), The Geology of Russia in Europe and Ural Mountains. Volume 1, 591–634. John Murray, London.
MacLean, B.C. 2002. Bovie Structure: A decapitated thrust? Geological Survey of Canada Open File 4274 [published online] Crossref
MacLean, B.C. and Morrow, D.W. 2004. Bovie Structure: its evolution and regional context. Bulletin of Canadian Petroleum Geology 52: 302–324. Crossref
Mamet, B.L. 1976. An atlas of microfacies in Carboniferous carbonates of the Canadian Cordillera. Geological Survey of Canada, Bulletin 255: 1–131. Crossref
McMechan, M., Currie, L., Richards, B.C., Ferri, F., Matthews, W., and O’Sullivan, P. 2018. Zircon provenance of the Carboniferous Mattson delta complex, western Laurentian margin, Canada: record of an eastern Greenland sourced pancontinental river system. Canadian Journal of Earth Science 55: 1129–1138. Crossref
Martin, W. 1809. Petrifacta Derbiensia; or Figures and Descriptions of Petrifactions Collected in Derbyshire. 102 pp. D. Lyon, Wigan. Crossref
McCoy, F. 1851. A description of some Mountain Limestone fossils. The Annals and Magazine of Natural History, Series 2 7: 167–175.
Milne Edwards, H. and Haime, J. 1850–1855. A Monograph of the British Fossil Corals. i–lxxxv + 1–71 (1850), 147–210 (1852), 211–244 (1853), 245–299 (1855). The Palaeontographical Society of London, London.
Nudds, J. 1981. An illustrated key to the British lithostrotionid corals. Acta Palaeontologica Polonica 25: 385–394.
d’Orbigny, A. 1852. Cours élémentaire de paléontologie et de géologie stratigraphique. Volume 2 (1). 382 pp. Victor Masson, Paris.
Potter, J., Richards, B.C., and Cameron, A.R. 1993. The petrology and origin of coals from the Lower Carboniferous Mattson Formation, southwestern District of Mackenzie, Canada. International Journal of Coal Geology 24: 113–140. Crossref
Poty, E. 1984. An evolutionary pattern for the W. European Lithostrotionidae. Palaeontographica Americana 54: 465–469.
Poty, E. 2002. Stratigraphy and paleobiogeography of Carboniferous rugose corals of Nova Scotia. In: L.V. Hills, C.M. Henderson, and E.W. Bamber (eds.), Carboniferous and Permian of the World. Canadian Society of Petroleum Geologists, Memoir 19: 580–587.
Richards, B.C. 1983. Uppermost Devonian, and Lower Carboniferous Stratigraphy, Sedimentation, and Diagenesis Southwestern District of Mackenzie and Southeastern Yukon Territory. 373 pp. Unpublished Ph.D. Thesis, University of Kansas, Lawrence.
Richards, B.C., Bamber, E.W., Higgins, A.C., and Utting, J. 1993. Carboniferous, Subchapter 4E. In: D.F. Stott and J.D. Aitken (eds.), Sedimentary Cover of the Craton in Canada. Geological Survey of Canada, Geology of Canada Series 5: 202–271. Crossref
Richards, B.C., Barclay, J.E., Bryan, D., Hartling, A., Henderson, C.M., and Hinds, R.C. 1994. Carboniferous strata of the Western Canada Sedimentary Basin. In: G.D. Mossop and I. Shetson (eds), Geological Atlas of the Western Canada Sedimentary Basin, 221–250. Canadian Society of Petroleum Geologists and Alberta Research Council, Calgary.
Richards, B.C., Mamet, B.L., Bamber, E.W., Higgins, A.C., McGregor, D.C., and Utting, J. 1989. Uppermost Devonian and Lower Carboniferous stratigraphy, sedimentation, and diagenesis, southwestern District of Mackenzie and southeastern Yukon Territory. Geological Survey of Canada, Bulletin 390: 1–135. Crossref
Rodríguez, S. 1984. Corales rugosos del este des Asturias. 528 pp. Doctoral thesis, Departamento de Paleontologia, Facultad de Ciencias Geológicas, Universidad Complutense de Madrid, Madrid.
Sando, W.J. 1985. Paraheritschioides, a new rugose coral genus from the Upper Pennsylvanian of Idaho. Journal of Paleontology 59: 979–985.
Semenoff-Tian-Chansky, P. 1985. Corals. North Africa. In: C. Martinez Diaz, R.H. Wagner, C.F. Winkler Prins, and L.F Granados (eds), The Carboniferous of the World. Volume 2. Australia, Indian Subcontinent, South Africa, South America and North Africa. Instituto Geologico y Minero de España. International Union of Geological Sciences. Publication 16: 374–381, 430–435.
Smith, S. 1928. The Carboniferous coral Nemistium edmondsi, gen et sp. nov. The Annals and Magazine of the Natural History, Series 10 1: 112–120. Crossref
Smith, S. 1935. Two Anthracolithic corals from British Columbia and related species from the Tethys. Journal of Paleontology 9: 30–42.
Smith, S. and Lang, W.D. 1930. Descriptions of the type specimens of some Carboniferous corals of the genera “Diphyphyllum”, “Stylastraea”, Aulophyllum, and Chaetetes. The Annals and Magazine of Natural History, Series 10 5: 178–194. Crossref
Smith, S. and Ryder, T.A. 1927. On the structure and development of Stauria favosa (Linnaeus). The Annals and Magazine of the Natural History, Series 9 20: 337–343. Crossref
Sorauf, J.E. 1970. Microstructure and formations of dissepiments in the skeletons of the Recent Scleractinia (hexacorals). Biomineralisation Forschungsberichte 2: 1–22.
Spasskiy, N.Y. and Kachanov, E.I. 1971. New primitive Early Carboniferous corals of the Altay and Urals [in Russian]. Zapiski Leningradskogo Gornogo Instituta 59: 48–64.
Vassilyuk, N.P. [Vasil’uk, N.P.] 1960. Lower Carboniferous corals of the Donetsk Basin [in Russian]. Akademiâ Nauk Ukrainskoj SSR, Trudy Instituta Geologičeskih Nauk, Seriâ Stratigrafii i Paleontologii 13: 1–178.
Verrill, A.E. 1865. Classification of polyps. (Extract condensed from a synopsis of the polypi of the North Pacific Exploring Expedition, under captains Ringgold and Rogers, U.S.N.). Proceedings of the Essex Institute 4: 145–149. Crossref
Wells, J.W. 1969. The formation of dissepiments in Zoantharian corals. In: K.S.W. Campbell (ed.), Stratigraphy and Palaeontology. Essays in Honour of Dorothy Hill, 17–26. Australian National University Press, Canberra.
Wright, G.N., McMechan, M.E., Potter, D.E.G., and Holter, M.E. 1994. Structure and Architecture of the Western Canada Sedimentary Basin. In: G.D. Mossop and I. Shetsen (eds.), Geological Atlas of the Western Canada Sedimentary Basin, 25–40. Canadian Society of Petroleum Geologists and Alberta Research Council, Calgary.
Yabe, H. 1950. Permian corals resembling Waagenophyllum and Corwenia. Proceedings of the Japan Academy 26: 74–79. Crossref
Acta Palaeontol. Pol. 64 (4): 851–870, 2019
https://doi.org/10.4202/app.00636.2019