
The oldest diving anseriform bird from the late Eocene of Kazakhstan and the evolution of aquatic adaptations in the intertarsal joint of waterfowl
Nikita Zelenkov
Zelenkov, N. 2020. The oldest diving anseriform bird from the late Eocene of Kazakhstan and the evolution of aquatic adaptations in the intertarsal joint of waterfowl. Acta Palaeontologica Polonica 65 (4): 733–742.
A specialized diving lifestyle has repeatedly evolved in several lineages of modern and fossil waterfowl (Anseriformes), with the oldest previously known representative being the late Oligocene Australian oxyurine ducks Pinpanetta. However, diving specializations have never been previously documented for any of the primitive Paleogene anseriforms (“stem-anatids”), and thus may be associated with the origin of modern anatid-like body plan. Here I describe a tarsometatarsus of a new duck-sized diving anseriform bird from the latest Eocene (late Priabonian) Kusto Svita in Eastern Kazakhstan, which predates the previously reported occurrence of diving specialization in Anseriformes by at least 6 MA. The new taxon Cousteauvia kustovia gen. et sp. nov. has an unusual and previously undocumented morphology, but partly resembles the stem-anatids Paranyrocidae and Romainvilliidae, thus representing the first known occurrence of diving adaptations in primitive non-anatid anseriforms. The evolutionary appearance of specialized waterfowl taxa in the late Eocene of Central Asia supports a view that this region might have played an important role in the evolution of morphologically derived Anseriformes. The structure of the intertarsal joint in basal and modern anseriforms is here further discussed in relation with adaptations for aquatic locomotion. The presence of elongate and evenly narrow condyles of the tibiotarsus in Anatidae and other swimming/diving birds allows a firm contact with the hyperprotracted tarsometatarsus at the initial phase of the propulsion. This morphology contrasts with the restricted condyles of Presbyornithidae, which indicate a different, strictly wading locomotory specialization. Cousteauvia obviously evolved diving specializations on the basis of a more primitive structure of the intertarsal joint.
Key words: Aves, Anseriformes, diving, evolution, intertarsal joint, Eocene, Kazakhstan, Central Asia.
Nikita Zelenkov [nzelen@paleo.ru], Borissiak Paleontological institute of Russian Academy of Sciences, Profsoyuznaya 123, Moscow, Russia.
Received 2 May 2020, accepted 13 June 2020, available online 8 October 2020.
Copyright © 2020 N. Zelenkov. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Waterfowl (order Anseriformes) are characteristic elements of modern avian faunas and are among the most common birds in the late Cenozoic fossil record. The group comprises three commonly recognized living families, which may be classified within two suborders Anhimae (with one family Anhimidae) and Anseres (with families Anseranatidae and Anatidae; Worthy et al. 2017). The early (pre-Late Oligocene) fossil record of Anseriformes is rather scanty, and thus the early diversity and disparity of the group remains poorly known (reviewed in Mayr 2009, 2017). Most early-diverged Paleocene to early Eocene anseriforms (Presbyornis isoni, Anatalavis oxfordi, Conflicto antarcticus, Naranbulagornis khun) were relatively large-bodied birds (Olson 1994, 1999; Dyke 2001; Tambussi et al. 2019; Zelenkov 2019), and the exceptionally well-studied smaller Presbyornithidae are characterized by highly mosaic and apparently somewhat aberrant postcranial morphology, in which they rather resemble waders and flamingoes (e.g., Olson and Feduccia 1980; Ericson 2000; Mayr 2009, 2017; De Pietri et al. 2016).
Derived anseriforms (family Anatidae sensu Dickinson and Remsen 2013; hereafter Anatidae sensu lato) constitute the overwhelming majority of the modern taxonomic anseriforms diversity and display an array of feeding and locomotor specializations, including repeatedly occurring diving adaptations (Carboneras 1992; McCracken et al. 1999; Worthy and Lee 2008). Anatomically modern Anatidae sensu lato first appear in the fossil record in the early Oligocene of Belgium (Mayr and Smith 2001), but already in the late Oligocene they have a broad geographical distribution and are represented by several phylogenetical lineages (Agnolin 2004; Mourer-Chauviré et al. 2004; Worthy 2009). Many diving anatids, including flightless forms, have been described from the late Oligocene to Pleistocene (Howard 1964; Cheneval 1987; Worthy et al. 2007, 2008; Noriega et al. 2008; Worthy and Lee 2008; Worthy 2009; Zelenkov 2011, 2012; Stidham 2015; Watanabe and Matsuoka 2015; Stidham and Hilton 2016; Stidham and Zelenkov 2017; Watanabe 2017; De Mendoza 2019), clearly showing that such adaptations represent a common evolutionary pathway of the group. However, no specialized diving forms have been previously known among early-diverging Anseriforms, as the relationships between the specialized divers Vegaviidae and Anseriformes (and even Galloanseres) remain unclear (Agnolin et al. 2017; Worthy et al. 2017; Mayr et al. 2018). The oldest known duck-like anseriforms (late Eocene to Oligocene Romainvilliidae) still resemble Presbyornithidae morphologically, thus showing that Anatidae sensu lato may be evolutionary linked with these aberrant long-legged Paleogene anseriforms (Mayr 2008; Zelenkov 2018). The derived elaborate ability to filter-feed on small floating objects could have been the driving force that led to the origin of short-legged anatid-like morphological type on the basis of plesiomorphic wading morphological type of Presbyornithidae (Zelenkov and Stidham 2018). This scenario implies a possibility that the evolutionary occurrence of diving adaptations may only be implemented on the basis of the derived morphological type of modern Anatidae sensu lato, which is supported by the current fossil record of diving anseriforms. However, the Eocene diversity of Anseriformes, and especially that of small-bodied forms, remains very poorly known.
Here I describe a tarsometatarsus from the late Eocene of Eastern Kazakhstan, which belongs to a new primitive diving anseriform bird, representing the first evidence of the evolutionary occurrence of diving adaptations in early-diverged Anseriformes. The new form displays a previously undocumented mosaic of plesiomorphic and autapomorphic features, showing partial similarities with modern Anseranatidae and fossil Paranyrocidae, thus adding significantly to the known diversity of fossil Anseriformes. This is further only the second Eocene anseriform bird from Asia, which supports previously expressed views that shallow epicontinental seas of Central Asia might have played an important role in the evolution of Anseriformes (Zelenkov 2018). The derived morphology of the intertarsal joint in anatids is further explained here in terms of the characteristic aquatic locomotion of these birds.
Institutional abbreviations.—PIN, Borissiak Paleontological Institute of Russian Academy of Sciences, Moscow, Russia; USNM, Smithsonian National Museum of Natural History, Washington D.C., USA.
Other abbreviations.—M, musculus.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:2E8462E0-3D8E-4155-8619-8AAB6836DA90.
Material and methods
The specimen described in this work comes from the Kusto-Kyzylkain (“Plesh”) locality in the Eastern Kazakhstan (Fig. 1) and was collected by Nina S. Shevyreva and Vyacheslav M. Chkhikvadze of the Paleontological Institute of Soviet Academy of Sciences in 1977. The locality is situated in the Zaysan Basin between the rivers Kusto and Kyzylkain some 20 km to the South of Zaysan lake and 60 km to the West of the Zaysan city (see Tleuberdina 2017). A short list of amphibia and reptilia has been reported previously from this locality (reviewed in Tleuberdina 2017). Fossil remains comes from the Kusto Svita (“Formation”), which is broadly exposed in the Zaysan Basin and is dated to the Ergilian (late Priabonian; latest Eocene, Emry et al. 1998). No bird remains have been previously described from the Kusto-Kyzylkain locality, but Ergilornis sp. and Romainvillia kazakhstanensis Zelenkov, 2018 are known from other localities of the Kusto Svita (Kurochkin 1981; Zelenkov and Kurochkin 2015; Zelenkov 2018).
Fig. 1. General outline map showing the geographical position of Kazakhstan (A) and Kusto-Kyzylkain locality (asterisk) at Zaysan Basin (B).
Comparative material on fossil and modern Anseriformes were examined primarily at the collections of the Paleontological Institute of Russian Academy of Sciences and Smithsonian National Museum of Natural History.
Anatidae of earlier authors (e.g., Carboneras 1992) often included the morphologically distinct Anseranatidae and hence distinct fossil forms (e.g., Romainvilliinae, Paranyrocinae) could also be easily included within broad-sensed Anatidae as subfamilies. However, recent authors classified Anseranatidae as a separate family of Anseriformes (e.g., Dickinson and Remsen 2013). The recently proposed exclusion of the phylogenetically basal and morphologically easily distinguishable (e.g., Woolfenden 1961) whistling ducks Dendrocygninae from Anatidae and their treatment as a separate family (Sun et al. 2017) allows recognition of fossil Romainvilliinae and Paranyrocinae as separate families as well (Zelenkov 2018). This taxonomic treatment, adopted in this work, corresponds with the distinctive osteology of these fossil groups as compared to modern Anatidae sensu stricte and Dendrocygnidae (see Mayr 2008; Mayr and Smith 2017).
Systematic paleontology
Class Aves Linnaeus, 1758
Order Anseriformes Wagler, 1831
Family incertae sedis
Remarks.—The new taxon (Fig. 2) is referred to Anseriformes based on the following combination of characters: the medial angle of the proximal end is narrow and prominent, the shaft is mediolaterally compressed, the hypotarsus with one closed canal (for the tendon of m. flexor digitorum longus), the crista medialis hypotarsi is dorsoplantarly high and proximodistally elongate, the cotyla lateralis forms a characteristic distally extending “lip” on the dorsal surface of the bone, the plantar opening of the medial proximal vascular foramen is shifted proximally towards the proximal part of the hypotarsus. In particular, these features (except the structure of the hypotarsus) distinguish the new fossil from diving Alcidae. The structure of the hypotarsus is also more complicated in Alcidae. Yet another character of the new fossil that is shared with Anseriformes is the presence of a tubercle on the lateral surface of the bone just distal to the level of the hypotarsus. The hypotarsus of the new form is somewhat similar to that of some Galliformes, but aquatic forms with thick bone walls are not known for these ground and usually forest-dwelling birds. Galliformes further have well-developed fossae parahypotarsales.
The new taxon represents stem-Anatidae sensu lato (Anatidae+Dendrocygnidae) because it does not possess an apomorphic dorsoplantarly low hypotarsus with four crest and three well-defined sulci, which invariantly characterizes crown anatids (Mayr 2016). Instead, the tarsometatarsus of the new form is high and has only two pronounced crests bordering one medial canal (for the tendon of m. flexor digitorum longus) together with a plantarly located sulcus. This structure of the hypotarsus is definitively more derived than in Anhimidae and fossil Presbyornithidae, which have only one medial sulcus (Mayr and Smith 2017). The presence of closed medial hypotarsal canal distinguishes the new taxon from fossil Paranyroca Miller and Compton, 1939 (Paranyrocidae, Mayr and Smith 2017), which otherwise have similar outlines of the hypotarsus, though a similarly-built hypotarsus with a canal might have been present in Saintandrea, a large-sized representative of Romainvilliidae, but the referred specimen is not sufficiently preserved to confirm it (Mayr and De Pietri 2013). Although affinities of the new taxon with Romainvilliidae seem probable, the type genus Romainvillia Lebedinsky, 1927 has a morphologically rather different tarsometatarsus, more similar to that of Anatidae (Mayr 2008). Anseranatidae also have one closed medial hypotarsal canal, but otherwise are characterized by a more complex and an autapomorphically strongly laterally shifted hypotarsus, which might have evolved independently from Anatidae (Mayr and Smith 2017). Thus, the familiar assignment of the new taxon is unclear; it may represent a separate undescribed family of belong to either stem Anseranatidae, specialized Romainvilliidae or Paranyrocidae. The latter assignment appears to be most probable. The presence of a distinct distally extending lip of the cotyla lateralis (see below) indicates that the new genus occupies a more derived position than Anseranatidae.
Genus Cousteauvia nov.
ZooBank LSID: urn:lsid:zoobank.org:act:8C421BB2-4B4D-47F1-BA F0-74E3D2400964
Type species: Cousteauvia kustovia gen. et sp. nov., monotypic; see below.
Etymology: In honor of ocean explorer and prominent diver Jacques-Yves Cousteau (1910–1997), appealing to the fact that the new taxon is the oldest known (and presumably the first evolved) diving crown-anseriform bird. The gender is feminine.
Diagnosis.—As for the type species, by monotypy.
Remarks.—Cousteauvia differs from all known anseriforms by having a highly prominent dorsomedial crest, running distally from the tip of the cotyla medialis, and further by the presence of a crista plantaris medialis, representing a continuation of the medial hypotarsal crest. Cousteauvia further differs from crown-group ducks (Anatidae and Dendocygnidae) by the hypotarsus having only two (instead of four) crests and a common broad lateral sulcus.
Cousteauvia kustovia sp. nov.
Fig. 2A.
ZooBank LSID: urn:lsid:zoobank.org:act:45AFA67E-25E5-4A42-A066- 204CFCA2CBDF
Etymology: In reference to the geological origin of the specimen (Kusto Svita), which is itself named after the Kusto river.
Holotype: PIN 2612/4, proximal half of left tarsometatarsus.
Type locality: Kusto-Kyzylkain (“Plesh”), Kusto Svita, Zaysan Basin, Eastern Kazakhstan (Fig. 1).
Type horizon: Late Priabonian (MP 19–MP 20); latest Eocene (see Emry et al. 1998; Tleuberdina 2017).
Diagnosis.—Medium sized anseriform, roughly similar in size to modern wild specimens of Anas platyrhynchos. Proximal tarsometatarsus with bulbous and proximally prominent eminentia intercotylaris; cotyla medialis oval-shaped, narrow and dorsally protruding; articular surface of the cotyla medialis does not extend on the medial surface of eminentia intercotylaris; cotyla lateralis wider and with cut dorsolateral angle; dorsomedial margin of the shaft forming prominent ridge; dorsolateral ridge vestigial; tuberositas m. tibialis cranialis short; hypotarsus moderately low, with closed medial canal for m. flexor digitorum longus and two plantar grooves, sulcus for m. fibularis longus absent; crista medialis hypotarsi is the best developed and extends moderately distally as a low crista medialis plantaris; caudal surface of the bone distal to hypotarsus slightly inclined medially (not flat); bone walls very thick.
Description.—The holotype represents a well-preserved proximal half of the left tarsometatarsus with complete proximal articular articular area and hypotarsus (Fig. 2). The shaft is notably narrowed relative to the proximal end, being roughly similar to the condition in Presbyornithidae and Anseranatidae but unlike Anhimidae, Romainvillia and Anatidae sensu lato. A similarly widened proximal end might have also been present in large romainvilliid Saintandrea Mayr and De Pietri, 2013 (Mayr and De Pietri 2013). The cross-sectional area of the mid-shaft is sub-trapezoid. The bone walls are very thick as in modern diving Anatidae and in contrast to thin bony walls of dabbling ducks (Fig. 2).
The eminentia intercotylaris is roughly symmetrical and bulbous, and it protrudes further proximally than the medial border of the cotyla medialis, as in Saintandrea and some non-anseriform taxa (e.g., Palaelodidae), but unlike all other fossil and modern Anseriformes. In Anatidae and Anseranatidae, the eminentia is asymmetrical and its more pointed tip is inclined medially. Anhimidae have an even bigger, but also symmetrical eminentia intercotylaris. In proximal view, the eminentia intercotylar is also prominent and extends plantarly approximately up to the mid-point of the cotylae, whereas it is restricted to the dorsal part of the articular area in Anatidae. The articular surface of the cotyla medialis does not expand on the medial wall of the eminentia, in contrast to the condition in Anatidae.
The cotyla medialis is oval-shaped and rather narrow; it protrudes more dorsally than the eminentia intercotylaris unlike the condition in all other Anseriformes except the supposed late presbyornithid Wilaru tedfordi Boles, Finch, Hofheins, Vickers-Rich, Walters, and Rich, 2013, in which this cotyla is, however, unusually mediolaterally wide (De Pietri et al. 2016). The dorsal margin of the cotyla medialis is distinctly convex, which is different from nearly straight margin in all other anseriforms except Anhimidae. The cotyla lateralis is wider than the cotyla medialis and is sub-circular in shape as in Paranyroca but unlike all other Anseriformes. The dorsomedial angle of the cotyla lateralis is prominent (as in Paranyroca) because of the well-developed “lip” (Fig. 2: l), extending to the dorsal surface of the bone. This lip is positioned entirely on the dorsolateral edge of the bone in Anatidae, whereas in Cousteauvia it is strongly shifted medially. The condition in Romainvillia is unclear due to poor preservation. Such a “lip” (distal extension of the cotyla lateralis) is not present in Anhimidae, Presbyornithidae and Anseranatidae and hence may represent an apomorphy of Anatidae sensu lato + Cousteauvia. The exact state of this character in Romainvilliidae is unclear, but Paranyroca may have a similar morphology. Plantar parts of both cotylae medialis et lateralis are bordered by not high but distinct beads, which are present in Presbyornithidae, but are absent or at least indistinct in Anatidae and other Anseriformes. The condition in Romainvilliidae is unclear.
Dorsally, there are two distinct concavities in the proximal margin of the bone, formed by the cotylae. The lateral concavity protrudes distinctly more distally than the medial one, as in Presbyornithidae and many Anatidae, but unlike Paranyroca. The condition of this feature in Romainvilliidae is unclear, but Saintandrea appears to be similar to Paranyroca in this respect (Mayr and De Pietri 2013). The fossa infracotylaris is deep proximally as in most Anseriformes except Anhimidae (Anseranatidae have a moderately excavated fossa). The foramina vascularia proximalia are small and relatively widely separated, as in Anatidae. The tuberositas m. tibialis cranialis are short and proximally located, as in Anhimidae, Presbyornithidae, and Anseranatidae. In the more derived anseriforms, the tuberositas are notably elongate and shifted distally.
The most unusual feature of Cousteauvia kustovia gen. et sp. nov. is the relative development of the dorsal lateral and medial crests, which border the fossa infracotylaris. In all known Anseriformes except Anseranatidae, the lateral crest is more prominent (both in robustness and the degree of dorsal protrusion) than the medial one, and this asymmetry is especially well-pronounced in diving taxa (e.g., Oxyurini, Mergini). Cousteauvia kustovia gen. et sp. nov. displays an alternative condition, with the medial crest being higher and more dorsally prominent than the lateral one. As a result, in medial view only the dorsomedial crest is visible in Cousteauvia kustovia gen. et sp. nov. (Fig. 2A2: cdm) and Anseranatidae, whereas in other anseriforms the dorsolateral crest is also visible as it protrudes dorsally well above the medial one (Fig. 2B2: cdl). This medial crest continues distally, forming a low medial wall of the dorsal sulcus (sulcus extensorius), as in Presbyornithidae and Anseranatidae, but not other Anseriformes. An apparently more poorly defined sulcus extensorius is visible in Cygnopterus alphonsi Cheneval, 1984 and Paranyroca magna Miller and Compton, 1939 (Mayr and Smith 2017: fig. 1). Modern Anserini also have a well-developed (although less prominent) dorsomedial crest.
The medial surface of the bone is slightly concave, forming a rather shallow but still distinct fossa parahypotarsalis medialis, which is totally absent in Anatidae, but a relatively-well defined fossa is present in Anseranatidae and Presbyornithidae. A large vascular foramen is present in the middle area of the fossa. The fossa parahypotarsalis lateralis does not form a concavity, being nearly vestigial. However, the lateral side of the shaft is not flat proximally, but have a slightly sloping plantar part. Two muscular/ligamental scars are present in this area, one is located dorsoproximally, and the other one is shifted distally and plantarly. Distally the lateral surface of the tarsometatarsus is flat as in modern Anatidae.
The hypotarsus is low and in general proportions is most similar to that of Paranyroca (see Mayr and Smith 2017), from which it still differs in having a closed canal for the tendon of m. digitorum longus, as in Anseranatidae, Dendrocygnidae and most Anatidae. The medial hypotarsal crest, bordering this canal and a plantarly located sulcus, is prominent and continues distally as a gradually lowering crista medialis plantaris, a feature which is not present in any other Anseriformes (the medial hypotasal crest is still elongate in Anserini). This crest distally borders a distinct sulcus for the tendon of m. digitorum longus, which is also lacking in all other Anseriformes. Lateral of the abovementioned canal, the hypotarsus forms one broad sulcus, as in Paranyroca. Unlike Paranyroca and Anatidae, the hypotarsus extends as much laterally as the lateral margin of the cotyla lateralis. In medial view, the straight proximal wall of the hypotarsus is oriented sub-perpendicular to the long axis of the bone, unlike Anatidae, where this wall is angled and sometimes concave. Distally to the hypotarsus, the plantar surface of the shaft slopes laterally more notably than in Anatidae due to the plantar prominence of the medial part of the shaft (as in modern Cereopsini).
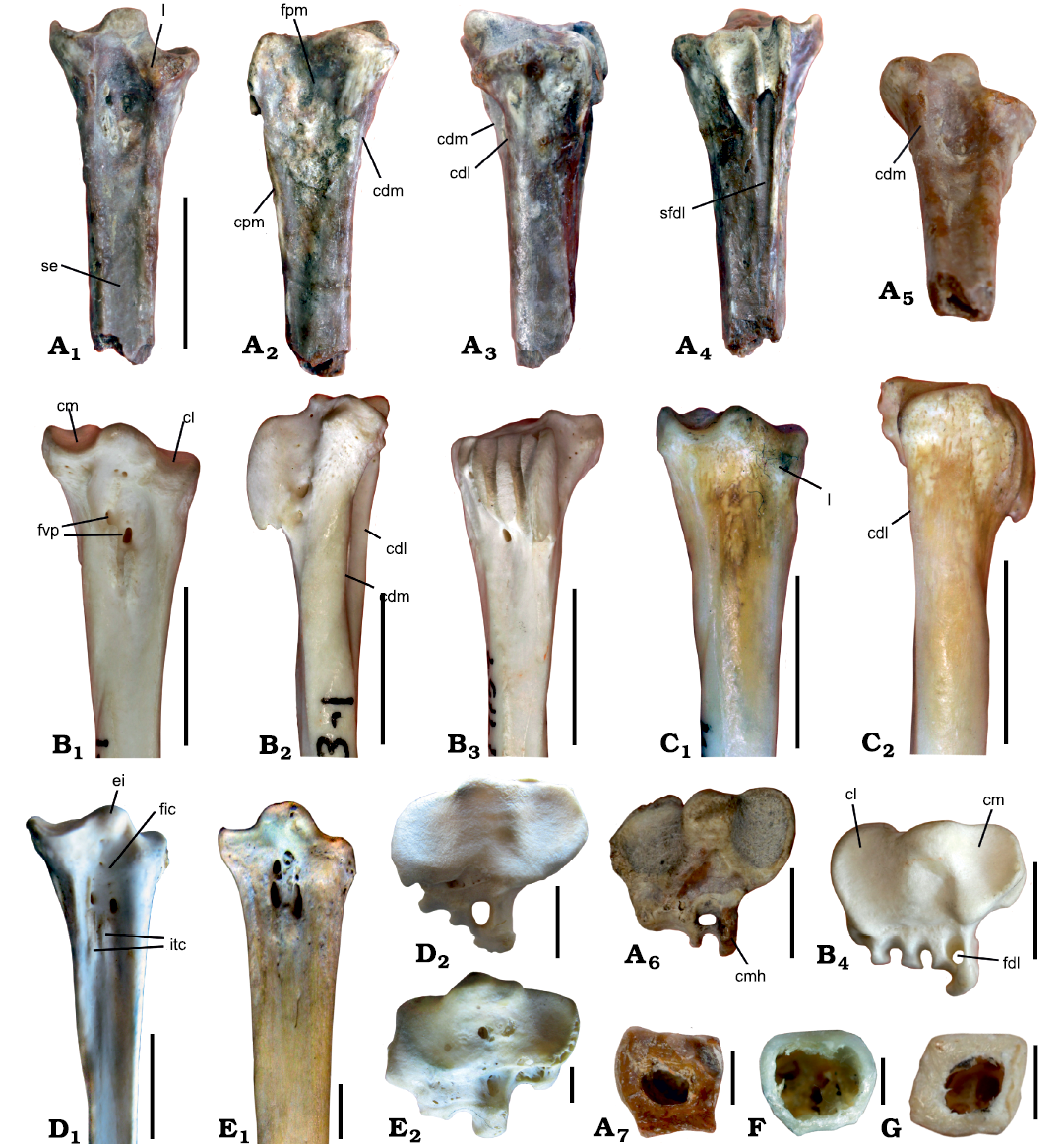
Fig. 2. Tarsometatarsi of anseriform bird Cousteauvia kustovia gen. et sp. nov. and selected modern Anseriformes. A. Cousteauvia kustovia gen. et sp. nov., holotype PIN 2612/4, latest Eocene of Kusto-Kyzylkain, Eastern Kazakhstan, in dorsal (A1), medial (A2), lateral (A3), plantar (A4), angled disto-dorsal (A5), and proximal (A6) views, and distal view on the cross-section (A7). B. Melanitta perspicillata Linnaeus, 1758 (Anatidae), PIN 41-9-1, Recent, in dorsal (B1), medial (B2), plantar (B3), and proximal (B4) views. C. Anas undulata Dubois, 1839 (Anatidae), PIN 40-32-2, Recent, in dorsal (C1) and lateral (C2) views. D. Anseranas semipalmata (Latham, 1798) (Anseranatidae), USNM 621019, Recent, in dorsal (D1) and proximal (D2) views. E. Anhima cornuta (Linnaeus, 1766) (Anhimidae), USNM 345208, Recent, in dorsal (E1) and proximal (E2) views. F. Anas platyrhynchos Linnaeus, 1758 (Anatidae), PIN 40-30-3, Recent, in distal view on the cross-section. G. Clangula hyemalis (Linnaeus, 1758) (Anatidae), PIN 41-7-8, Recent, in distal view on the cross-section. Abbreviations: cdl, dorsolateral crest of the shaft; cdm, dorsomedial crest of the shaft; cl, cotyla lateralis; cm, cotyla medialis; cmh, crista medialis hypotarsi; cpm, crista plantaris medialis; ei, eminentia intercotylaris; fdl, canal for tendon of m. flexor digitorum longus; fic, fossa infracotylaris; fpm, fossa parahypotarsalis medialis; fvp, foramina vascularia proximalia; itc, impressio m. tibialis cranialis; se, sulcus extensorius; sfdl, sulcus for tendon of m. flexor digitorum longus; l, lip-like distal extension of the cotyla lateralis. Scale bars: A1–A5, B1–B3, C, D1, 10 mm; A6, B4, D2, E2, 5 mm; A7, F, G, 2 mm.
Remarks.—The Kusto Svita has yielded a stem-anatid taxon Romainvillia kazakhstanesis (Zelenkov 2018), which must be compared with Cousteauvia kustovia gen. et sp. nov. Romainvillia kazakhstanesis is known from a coracoid that is very similar to the coracoid of Romainvillia stehlini Lebedinsky, 1927 from the contemporary latest Eocene of France (Mayr 2008). Given such a close similarity of the coracoids in two species, it would be reasonable to assume the same for their tarsometatarsi. However, the tarsometatarsus of R. stehlini significantly differs from that of Cousteauvia kustovia gen. et sp. nov., preventing any possible association of the new Kazakh specimen with Romainvillia. In R. stehlini, the tarsometatarsus is of typical anatid proportions and the shaft is not restricted relative to the proximal end, the dorsolateral margin of the proximal part of the shaft is thick and dorsally protruding, at least one visible tuberculum m. tibialis cranialis is elongate, the sulcus extensorius is poorly pronounced due to the absence of the dorsomedial ridge of the shaft, and the medial plantar ridge and a distally extending sulcus for the tendon of m. flexor digitorum longus are absent. These features thus clearly distinguish R. stehlini from Cousteauvia kustovia gen. et sp. nov. and strongly indicate closer affinities of Romainvillia with Anatidae. Additionally, in overall size, the tarsometatarsus of R. stehlini is comparable with modern small-sized teals (Mayr 2008), whereas Cousteauvia kustovia gen. et sp. nov. was a larger bird approximating wild specimens of A. platyrhynchos, judging from the size of the articular surface.
Discussion
The notably increased thickness of the bony wall of the tarsometatarsus indicates that Cousteauvia kustovia gen. et sp. nov. was a specialized diving bird, thus being the first known Eocene and the oldest diving representative of the order Anseriformes. It displays an unusual combination of characters, but is definitively more derived than Anhimidae and is more primitive than Anatidae and fossil Romainvilliidae (see comparisons above). Some similarities with modern Anseranatidae are notable and may indicate their closer relationships, but assignment of Cousteauvia kustovia gen. et sp. nov. to Anseranatidae is not supported by the structure of the hypotarsus. The new genus represents the first documented case of diving specializations in the early-diverged Anseriformes and shows that pronounced ecological diversification of the group already took place in the Eocene. The occurrence of Cousteauvia kustovia gen. et sp. nov. in the late Eocene of Kazakhstan supports a previously expressed view (Zelenkov 2018) that a retreat of large epicontinental Central Asian seas, which took place in the Priabonian epoch and resulted in dramatic changes in marine ecosystems in this geographical area (Nevesskaya 1999), might have played a significant role in the evolution of derived anseriforms. Currently, three taxa of primitive anseriforms are known from the late Eocene of this regions: teal-sized Romainvillia kazakhstanensis, medium-sized Cousteauvia kustovia gen. et sp. nov., and larger swan-sized “Cygnavus” formosus Kurochkin, 1968 (Zelenkov and Kurochkin 2015; Zelenkov 2018).
Although the tarsometatarsus of Cousteauvia kustovia gen. et sp. nov. is similar to that of some modern marine ducks (Mergini) in the general outlines (including the lateral flatness, which reduces profile drag; Lovvorn 1991), it nevertheless displays a series of profound structural differences, clearly indicating a different morpho-functional pattern. In particular, the proximally located tuberositas m. tibialis cranialis of Cousteauvia kustovia gen. et sp. nov. show that speed was more important than power in the “dorsiflexion” (anatomical hyperextension) of the foot. The position of the tuberositas m. tibialis cranialis relative to the intertarsal joint vary depending on different ecological specializations and hence different leg usage in modern birds (Zeffer and Norberg 2003). Norberg (1979) has calculated that even a slight (1–2 mm) increase in lever arm (due to distal shift of the insertion point of the muscle) results in a significant increase of the resultant force. Zeffer and Norberg (2003) revealed a difference in the mechanical properties of the leg geometry in fast-swimming versus slow-smimming birds. In the former group, the speed is favored over force, because fast-swimming birds catch their prey underwater and hence a high stroke frequency is important for them. This explains a shorter moment arms of the tibialis cranialis relative in this group relative to the condition in slow-swimmers (Zeffer and Norberg 2003). A similar mechanical requirement could explain the proximally positioned scars for m. tibialis cranialis in Cousteauvia kustovia gen. et sp. nov. as well.
The most specialized modern diving ducks Oxyura and Biziura have a dorsoplantarly extended hypotarsus (especially, the medial hypotarsal ridge), which provides a mechanical advantage for m. gastrocnemius, the main plantar flexor of the feet, playing the key role in paddling (Raikow 1970). A similar posteriorly expanded hypotarsus is also characteristic of other specialized diving birds (Mayr 2016; Clifton et al. 2018). Cousteauvia kustovia gen. et sp. nov. has a moderately protruding medial ridge of the hypotarsus, more comparable with that of Anas ducks, and thus apparently was specialized in a different way than Oxyura. However, the presence of the plantar medial ridge, which represents an ossified continuation of the Achilles tendon of m. gastrocnemius, may still imply an increased mechanical role of this muscle in Cousteauvia kustovia gen. et sp. nov. The well-visible groove for the tendon of the m. flexor digitorum longus, which characterizes Cousteauvia kustovia gen. et sp. nov., likely indicates a relative importance of the corresponding muscle. The proximal toe flexors are enlarged in diving birds (Clifton et al. 2018) and during propulsive phase these muscle can effectively control unnecessarily hyperextension of toes, which would reduce effectiveness of the foot as a paddle (Raikow 1970).
The dorsal facies of the tarsometatarsus (sulcus extensorius) is the origin site of m. extensor brevis digiti IV, a short extensor of the fourth pedal digit, which is a relatively well-developed muscle with a long belly in ducks (Raikow 1970). The deepened sulcus extensorius in Cousteauvia kustovia gen. et sp. nov. probably accounts for a better development of this muscle, which in turn implies a relatively greater functional role of the fourth toe in locomotion. The biological meaning of such adaptation, which might also be present in Anseranatidae, requires an explanation.
A suite of peculiar modifications of the tarsometatarsus in Cousteauvia kustovia gen. et sp. nov. is likely associated with diving specialization of this taxon. Modern diving anatids convergently evolved a set of shared morphological traits (e.g., McCracken et al. 1999; Worthy and Lee 2008), and so the unusual nature of adaptations in Cousteauvia kustovia gen. et sp. nov. may be explained by the fact that they are realized on a different structural background, i.e. a more plesiomorphic osteology and function of the hindlimb. Indeed, Cousteauvia kustovia gen. et sp. nov. share many structural similarities (see above) with modern Anseranatidae and fossil Presbyornithidae, which are likely a sister taxon of Anseres (Worthy et al. 2017). Presbyornithidae were long-legged waders and Anseranatidae also have a relatively long tarsometatarsus as compared with modern Anatidae sensu lato. The tarsometatarsus of Cousteauvia kustovia gen. et sp. nov. thus might have also been longer than in modern diving ducks, and this proportional difference could be one of the factors explaining the overall structural modifications in the fossil taxon.
Yet another important factor could be the different kinematics of the intertarsal joint. The bulbous eminentia intercotylaris of Cousteauvia kustovia gen. et sp. nov. highly contrasts with the smaller, asymmetrical and pointed eminentia of modern anatids and implies a different structure of the intertarsal joint. In anatids, the medial wall of the eminentia represents a continuation of the cotyla medialis and articulates with the lateral surface of the condylus medialis of the tibiotarsus, which is evenly narrow in these birds (Fig. 3A1: cm2). The eminentia does not act as a stop in anatids and allows a considerable dorsiflexion of the tarsometatarsus, staying in full contact with the tibiotarsus at all possible angles of the dorsiflexion (Fig. 3A2, A3). The lack of the articular surface on the medial wall of the eminentia in Cousteauvia kustovia gen. et sp. nov. indicates that in this taxon it did not significantly articulate with the condyle, but the great proximal protrusal and wide apex of the eminentia intercotylaris in Cousteauvia kustovia gen. et sp. nov. show that it might have acted as a stop limiting dorsiflexion. However, the main function of the enlarged eminentia is likely to prevent significant rotation of the tarsometatarsus, as exemplified by the complete reduction of this structure in parrots (see Zelenkov and Dyke 2008). Rotation of the tarsometatarsus relative to the long axis of the tibiotarsus is also present in modern ducks (e.g., Provini et al. 2012) and is correlated with the presence of small and asymmetrical eminentia intercotylaris. Such a rotation hence was obviously limited in Cousteauvia kustovia gen. et sp. nov.
Among Anseriformes, Anhimidae display a bulbous and symmetrical eminentia intercotylaris (somewhat similar to that of Cousteauvia kustovia gen. et sp. nov.), which in this taxon is associated with proximally restricted (subtriangular in cranial view) condylus medialis of the tibiotarsus, as in wading birds (Fig. 3B: cm1). This proximal restriction of the articular surface results in that the area of contact between the condylus medialis of the tibiotarsus and the cotyla medialis of the tarsometatarsus gradually becomes smaller as the tarsometatarsus progressively dorsiflexes. Thus, a firm contact between the two bones is not possible at greatest angles of dorsiflexion, which itself implies that the main working diapason of such kind of intertarsal joint (as in Anhimidae; hereafter Type 1 intertarsal joint) covers a lower range of angles than in birds with an evenly narrow condylus medialis (as in Anatidae; Type 2 intertarsal joint). The condylus medialis of Cousteauvia kustovia gen. et sp. nov. thus might have been somewhat proximally restricted, as in the coeval fossil taxon “Cygnavus” formosus (see Zelenkov and Kurochkin 2015).
It is hypothesized here that the Type 1 intertarsal joint, which is present in Anhimidae and Presbyornithidae among Anseriformes, but further occurs in waders (Charadriiformes) and Phoenicopteriformes, is likely related with the wading (predominately walking) type of locomotion. A similar structure is also present in some ground birds, e.g., Otidiformes, Gruiformes, and others. The structure of Type 1 joint prevents significant rotation and thus provides a greater stability at standing or during the stance phase, when the foot experiences the greatest load. Morphology of the eminentia intercotylaris and condylus medialis of the tibiotarsus adds to this stability, but nevertheless prevents effective loading at the joint during considerable degree of dorsiflexion. The type 2 joint, as in advanced Anseriformes (Anatidae sensu lato) and other swimming birds, represents an adaptation for transmitting the force at increased dorsiflexion angles, as it allows a full contact between the tibiotarsus and tarsometatarsus.
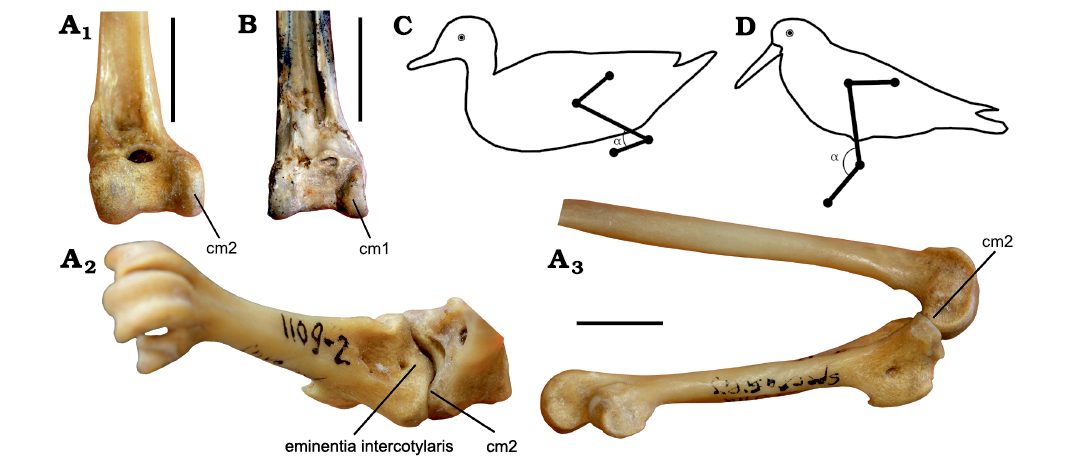
Fig. 3. The intertarsal joint of Recent Anatidae (exemplified by Somateria spectabilis (Linnaeus, 1758), PIN 41-2-2; A) and fossil wading bird (PIN 3104-65; early Eocene of Tsagaan-Khushu locality; southern Mongolia; B) as related to the swimming locomotion. Two types of the general structure of the distal tibiotarsus (A1, B), illustrating the difference in the shape of the condylus medialis (cm1, cm2). Maximal anatomically possible dorsiflexion of the tarsometatarsus in Anatidae, in craniomedial view (A2, showing full contact between the condylus medialis type 2 and the articular surface of the tarsometatarsus), in medial view (A3). Position of the tarsometatarsus relative to the tibiotarsus at the beginning of the propulsive phase of the stride in swimming duck (C) and walking wader (D); note the strongly dorsiflexed tarsometatarsus in the former (modified after Provini et al. 2012; Killbourne et al. 2016). Abbreviations: cm1, cm2; condylus medialis in type 1 and type 2 intertarsal joints (see text). Scale bars 10 mm.
The above proposed functional difference is in agreement with overall leg kinematics in waders and ducks. During walking, the long-legged Tringa waders (Scolopacidae) dorsiflex their intertarsal joint only to 90° (e.g., Reichert et al. 2017). A similar 70–90° flexion was recorded for walking/running in Vanellus and Recurvirostra, whereas this value may reach about 50° in Haematopus (Kilbourne et al. 2016). However, in waders the maximal dorsiflexion of the intertarsal joint occurs during the middle of the swing phase (Reichert et al. 2017), i.e. long before the contact between the leg and the substrate. The mechanical stress transmitted between the tibiotarsus and tarsometatarsus is higher at the beginning of the stance phase, when the intertarsal joint is extended (Fig. 3D). In swimming ducks, the dorsiflexion at the intertarsal joint brings the foot to an appropriate protracted position required for the propulsion, which is being initiated when the dorsiflexion is maximal (Provini et al. 2012). Thus, the maximal dorsiflexion at the intertarsal joint in swimming bird occurs during the moment of active interaction between the leg and the substrate (Fig. 3C), and hence the transmitted stress must be obviously much greater than during the mid-swing phase in waders.
The presence of large eminentia intercotylaris without articular surface in Cousteauvia kustovia gen. et sp. nov. indicates the presence of generally Type 1 intertarsal joint in this diving bird, and thus implies imperfect swimming adaptations as compared to modern anatids.
Acknowledgements
I am greatly indebted to Gerald Mayr (Senckenberg Research Institute and Natural History Museum Frankfurt, Frankfurt-am-Main, Germany) and Ricardo De Mendoza (Museo de la Plata, la Plata, Argentina) for valuable reviews. I am further thankful to Anna Melikhova (Lyceum No. 1535, Moscow, Russia) for the nomenclature suggestion, and Andrey V. Zinoviev (Tver State Univerity, Tver, Russia) for help with literature. This study was supported by the grant RFBR 20-04-00975.
References
Agnolín, F. 2004. Revisión sistemática de algunas aves deseadenses (Oligocene Medio) descriptas por Ameghino en 1899. Revista del Museo Argentino de Ciencias Naturales, Nueva Series 6: 239–244. Crossref
Agnolín, F.L., Egli, F.B., Chatterjee, S., Marsà, J.A.G., and Novas F.E. 2017. Vegaviidae, a new clade of southern diving birds that survived the K/T boundary. Science of Nature 104: 87. Crossref
Boles, W.E., Finch, M.A., Hofheins, R.H., Vickers-Rich, P., Walters M., and Rich, T.H. 2013. A fossil stone-curlew (Aves: Burhinidae) from the Late Oligocene/Early Miocene of South Australia. In: U.B. Göhlich and A. Kroh (eds.), Paleornithological Research 2013. Proceedings of the 8th International Meeting of the Society of Avian Paleontology and Evolution, Vienna, 2012, 43–61, Naturhistorisches Museum Wien, Vienna.
Carboneras, C. 1992. Family Anatidae (ducks, geese and swans). In: J. del Hoyo, A. Elliott, and J. Sargatal (eds.), Handbook of the Birds of the World. Vol. 1, 536–573, Lynx Edicions, Barcelona.
Cheneval, J. 1984. Les oiseaux aquatiques (Gaviiformes à Ansériformes) du gisement aquitanien de Saint-Gérandle-Puy (Allier, France): Révision systématique. Palaeovertebrata 14: 33–115.
Cheneval, J. 1987. Les Anatidae (Aves, Anseriformes) du Miocene de France. Revision systematique et evolution. Documents des Laboratoires de Geologie, Lyon 99: 137–156.
Clifton, G.T., Carr, J.A., and Biewener, A.A. 2018. Comparative hindlimb myology of foot-propelled swimming birds. Journal of Anatomy 232: 105–123. Crossref
De Mendoza, R.S. 2019. Phylogenetic relationships of the Early Miocene diving and flightless duck Cayaoa bruneti (Aves, Anatidae) from Patagonia: homology or convergence? Papers in Palaeontology 5: 743–751. Crossref
De Pietri, V.L., Scofield, R.P., Zelenkov, N., Boles, W.E., and Worthy, T.H. 2016. The unexpected survival of an ancient lineage of anseriform birds into the Neogene of Australia: the youngest record of Presbyornithidae. Royal Society Open Science 3: 150635. Crossref
Dickinson, E.C., and Remsen J.V., Jr. (eds.). 2013. The Howard and Moore Complete Checklist of the Birds of the World. 4th Edition, Vol. 1. 461 pp. Aves Press, Eastbourne.
Dubois, C.F. 1839. Ornithologische Gallerie, oder Abbildungen aller bekannten Vögel. 172 pp. J.A. Mayer, Aachen.
Dyke, G.J. 2001. The fossil waterfowl (Aves: Anseriformes) from the Eocene of England, American Museum Novitates 3354: 1–15. Crossref
Emry, R.J., Lucas, S.G., Tyutkova, L., and Wang, B. 1998. The Ergilian– Shandgolian (Eocene-Oligocene) transition in the Zaysan Basin, Kazakhstan. Bulletin of the Carnegie Museum of Natural History 34: 298–312.
Ericson, P.G.P. 2000. Systematic revision, skeletal anatomy, and paleoecology of the New World early Tertiary Presbyornithidae (Aves: Anseriformes). PaleoBios 20: 1–23.
Howard, H. 1964. Fossil anseriformes. In: J. Delacour (ed.), Waterfowl of the World, 233–326, Country Life, London.
Kilbourne, B.M., Andrada, E., Fischer, M.S., and Nyakatura, J.A. 2016. Morphology and motion: hindlimb proportions and swing phase kinematics in terrestrially locomoting charadriiform birds. Journal of Experimental Biology 219: 1405–1416. Crossref
Kurochkin, E.N. [Kuročkin, E.N.] 1968. New Oligocene birds from Kazakhstan [in Russian]. Paleontologičeskij žurnal 1968 (2): 92–101.
Kurochkin, E.N. [Kuročkin, E.N.] 1981. New representatives and evolution of two archaic gruiform families in Eurasia [in Russian]. Transaction of the Joint Soviet-Mongolian Paleontological Expedition 15: 59–86.
Latham, J. 1798. An essay on the tracheæ or windpipes of various kinds of birds. Transactions of the Linnean Society of London 4: 90–128. Crossref
Lebedensky, N.G. 1927. Romainvillia Stehlini n.g. n.sp. canard eocène provenant des marnes blanches du Bassin de Paris. Mémoires de la société paléontologique suisse 17: 1–8.
Linnaeus, C. 1758. Systema natura per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Vol. 1. 824 pp. Laurentii Salvii, Holmiae. Crossref
Linnaeus, C. 1766. Systema naturae per regna tria naturae: secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Ed. 12. Vol. 1. 532 pp. Laurentii Salvii, Holmiae. Crossref
Lovvorn, J.R. 1991. Mechanics of underwater swimming in foot-propelled diving birds. Proceedings of International Ornithological Congress 20: 1868–1874.
Mayr, G. 2008. Phylogenetic affinities and morphology of the late Eocene anseriform bird Romainvillia stehlini Lebedinsky, 1927. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 248: 365–380. Crossref
Mayr, G. 2009. Paleogene Fossil Birds. 262 pp. Springer-Verlag, Berlin. Crossref
Mayr, G. 2016. Variations in the hypotarsus morphology of birds and their evolutionary significance. Acta Zoologica 97: 196–210. Crossref
Mayr, G. 2017. Avian Evolution. The Fossil Record of Birds and its Paleobiological Significance. 293 pp. John Wiley & Sons, Inc, Chichester. Crossref
Mayr, G. and De Pietri, V.L. 2013. A goose-sized anseriform bird from the late Oligocene of France: the youngest record and largest species of Romainvilliinae. Paläontologische Zeitschrift 87: 423–430. Crossref
Mayr, G. and Smith, R. 2001. Ducks, rails, and limicoline waders (Aves: Anseriformes, Gruiformes, Charadriiformes) from the lowermost Oligocene of Belgium. Geobios 34: 547–561. Crossref
Mayr, G. and Smith, T. 2017. First Old-World record of the poorly known, swan-sized anseriform bird Paranyroca from the late Oligocene/early Miocene of France. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 286: 349–354. Crossref
Mayr, G., De Pietri, V.L., Scofield, R.P., and Worthy, T.H. 2018. On the taxonomic composition and phylogenetic affinities of the recently proposed clade Vegaviidae Agnolín et al., 2017—neornithine birds from the Upper Cretaceous of the Southern Hemisphere. Cretaceous Research 86: 178–185. Crossref
McCracken, K.G., Harshman, J., McClellan, D.A., and Afton, A.D. 1999. Data set incongruence and correlated character evolution: an example of functional convergence in the hind-limbs of stifftail diving ducks. Systematic Biology 48: 683–714. Crossref
Miller, A.H. and Compton, L.V. 1939. Two fossil birds from the Lower Miocene of South Dakota. The Condor 41: 153–156. Crossref
Mourer-Chauviré, C., Berthet, D., Hugueney, M., 2004. The late Oligocene birds of the Créchy quarry (Allier, France), with a description of two new genera (Aves: Pelecaniformes: Phalacrocoracidae, and Anseriformes: Anseranatidae). Senckenbergiana Lethaea 84: 303–315. Crossref
Nevesskaya, L.A. [Nevesskaâ, L.A.] 1999. Etapy razvitiâ bentosa anerozojskih Morej (mezozoj i kainozoj) [in Russian]. 503 pp. Nauka, Moskva.
Norberg, U.M.L. 1979. Morphology of the wings, legs and tail of three coniferous forest tits, the goldcrest, and the treecreeper in relation to locomotor pattern and feeding station selection. Philosophical Transactions. Royal Society London B 287: 131–165. Crossref
Noriega, J.I., Tambussi, C.P., and Cozzuol, M.A. 2008. New material of Cayaoa bruneti TONNI, an Early Miocene anseriform (Aves) from Patagonia, Argentina. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 249: 271–280. Crossref
Olson, S.L. 1994. A giant Presbyornis (Aves: Anseriformes) and other birds from the Paleocene aquia formation of Maryland and Virginia. Proceedings of the Biological Society of Washington 107: 429–435.
Olson, S.L. 1999. The anseriform relationships of Anatalavis Olson and Parris (Anseranatidae), with a new species from the Lower Eocene London Clay. Smithsonian Contributions to Paleobiology 89: 231–243.
Olson, S.L. and Feduccia, A. 1980. Presbyornis and the origin of the Anseriformes (Aves: Charadriomorphae). Smithsonian Contributions to Zoology 323: 1–24. Crossref
Raikow, R.J. 1970. Evolution of diving adaptations in the stifftail ducks. University of California Publications in Zoology 94: 1–52.
Provini, P., Goupil, P., Hugel, V., and Abourachid, A. 2012. Walking, paddling, waddling: 3D kinematics Anatidae locomotion (Callonetta leucophrys). Journal of Experimental Zoology A 317: 275–282. Crossref
Reichert, J., Mayr, G., Wilke, T., and Peters, W.S. 2017. Waders (Scolopacidae) surviving despite malaligned leg fractures in the wild: kinematics of bipedal locomotion. Avian Research 8: 23. Crossref
Stidham, T.A. 2015. Re-description and phylogenetic assessment of the Late Miocene ducks Aythya shihuibas and Anas sp. (Aves: Anseriformes) from Lufeng, Yunnan, China. Vertebrata PalAsiatica 53: 335–349.
Stidham, T.A. and Hilton, R.P. 2016. New data on stiff-tailed duck evolution and dispersal from a new species of diving duck (Anseriformes: Anatidae: cf. Oxyurinae) from the Miocene High Rock Caldera in north-west Nevada, USA. Papers in Palaeontology 2: 41–58. Crossref
Stidham, T.A. and Zelenkov N.V. 2017. North American-Asian aquatic bird dispersal in the Miocene: evidence from a new species of diving duck (Anseriformes: Anatidae) from North America (Nevada) with affinities to Mongolian taxa. Alcheringa 41: 222–230. Crossref
Sun, Z., Pan ,T., Hu, C., Sun, L., Ding, H., Wang, H., Zhang, C., Jin ,H., Chang, Q., Kan, X., and Zhang, B. 2017. Rapid and recent diversification patterns in Anseriformes birds: Inferred from molecular phylogeny and diversification analyses. PLoS One 12: e0184529. Crossref
Tambussi, C.P., Degrange, F.J., De Mendoza, R.S., Sferco, E., and Santillana, S. 2019. A stem anseriform from the early Palaeocene of Antarctica provides new key evidence in the early evolution of waterfowl. Zoological Journal of Linnean Society 186: 673–700. Crossref
Tleuberdina, P.A. 2017. Sites of Fossil Vertebrates of Kazakhstan Phanerozoic [in Russian]. 300 pp. Ministry of Education and Science of the Republic of Kazakhstan, Almaty.
Wagler, J.G. 1831. Einige Mittheilungen ueber Thiere Mexicos. Isis von Oken 5: 510–535.
Watanabe, J. 2017. Quantitative discrimination of flightlessness in fossil Anatidae from skeletal proportions. Auk 134: 672–695.
Watanabe, J. and Matsuoka, H. 2015. Flightless diving duck (Aves, Anatidae) from the Pleistocene of Shiriya, northeast Japan. Journal of Vertebrate Paleontology 35: e994745. Crossref
Woolfenden, G.E. 1961. Postcranial osteology of the Waterfowl. Bulletin of the Florida State Museum. Biological Sciences 6: 1–129.
Worthy, T.H. 2009. Descriptions and phylogenetic relationships of two new genera and four new species of Oligo-Miocene waterfowl (Aves: Anatidae) from Australia. Zoological Journal of Linnean Society 156: 411–454. Crossref
Worthy, T.H. and Lee, M.S.Y. 2008. Affinities of Miocene waterfowl (Anatidae: Manuherikia, Dunstanetta and Miotadorna) from the St Bathans Fauna, New Zealand. Palaeontology 51: 677–708. Crossref
Worthy, T.H., Degrange, F.J., Handley, W.D., and Lee, M.S.Y. 2017. The evolution of giant flightless birds and novel phylogenetic relationships for extinct fowl (Aves, Galloanseres). Royal Society Open Science 4: 170975. Crossref
Worthy, T.H., Tennyson, A.J.D., Hand, S.J., and Scofield, R.P. 2008. A new species of the diving duck Manuherikia and evidence for geese (Aves: Anatidae: Anserinae) in the St Bathans Fauna (Early Miocene), New Zealand. Journal of Royal Society of New Zealand 38: 97–114. Crossref
Worthy, T.H., Tennyson, A.J.D., Jones, C., McNamara, J.A., and Douglas, B.J. 2007. Miocene waterfowl and other birds from central Otago, New Zealand. Journal of Systematic Palaeontology 5: 1–39. Crossref
Zeffer, A. and Norberg, U.M.L. 2003. Leg morphology and locomotion in birds: requirements for force and speed during ankle flexion. Journal of Experimental Biology 206: 1085–1097. Crossref
Zelenkov, N.V. 2011. Diving ducks from the middle Miocene of Western Mongolia. Paleontological Journal 45: 191–199. Crossref
Zelenkov, N.V. 2012. Neogene geese and ducks (Aves: Anatidae) from localities of the Great Lakes Depression, Western Mongolia. Paleontological Journal 46: 607–619. Crossref
Zelenkov, N.V. 2018. The Earliest Asian Duck (Anseriformes: Romainvillia) and the origin of Anatidae. Doklady Biological Sciences 483: 225–227. Crossref
Zelenkov, N.V. 2019. A swan-sized anseriform bird from the late Paleocene of Mongolia. Journal of Vertebrate Paleontology 38: e1531879. Crossref
Zelenkov, N.V. and Dyke, G.J. 2008. The fossil record and evolution of mousebirds (Aves: Coliiformes). Palaeontology 51: 1403–1418. Crossref
Zelenkov, N.V. and Kurochkin, E.N. [Kuročkin, E.N.] 2015. Class Aves [in Russian]. In: E.N. Kuročkin, A.V. Lopatin, and N.V. Zelenkov (eds.), Fossil Vertebrates of Russia and Adjacent Countries: Fossil Reptiles and Birds: Part 3, 86–290. Geos, Moskva.
Zelenkov, N.V. and Stidham, T.A. 2018. Possible filter-feeding in the extinct Presbyornis and the evolution of Anseriformes (Aves). Zoologičeskij žurnal 97: 943–956. Crossref
Acta Palaeontol. Pol. 65 (4): 733–742, 2020
https://doi.org/10.4202/app.00764.2020