
Camenellan tommotiids from the Cambrian Series 2 of East Antarctica: Biostratigraphy, palaeobiogeography, and systematics
THOMAS M. CLAYBOURN, CHRISTIAN B. SKOVSTED, MARISSA J. BETTS, LARS E. HOLMER, LUCY BASSETT-BUTT, and GLENN A. BROCK
Claybourn, T.M., Skovsted, C.B., Betts, M.J., Holmer, L.E., Bassett-Butt, L., and Brock G.A. 2021. Camenellan tommotiids from the Cambrian Series 2 of East Antarctica: Biostratigraphy, palaeobiogeography, and systematics. Acta Palaeontologica Polonica 66 (1): 207–229.
Cambrian Series 2 shelly fossils from thick carbonate successions in East Antarctica have received limited systematic treatment through the 20th century. Described here are the East Antarctic camenellan tommotiids from the Shackleton Limestone in the Central Transantarctic Mountains and the Schneider Hills limestone in the Argentina Range. This material comes from both newly sampled collections and incompletely described material from older collections. The assemblage supports correlation to the Dailyatia odyssei Zone and Pararaia janeae Trilobite Zone of South Australia, with the newly examined specimens of Dailyatia decobruta from the Shackleton Limestone providing direct correlation to the Mernmerna Formation of the Ikara-Flinders Ranges and White Point Conglomerate of Kangaroo Island. These East Antarctic assemblages include five species referred to Dailyatia, in addition to an undetermined kennardiid species and fragments of the problematic Shetlandia multiplicata. The results further corroborate the notion that fossiliferous carbonate clasts found on King George Island were sourced from the same carbonate shelf as the Shackleton Limestone, with the taxon S. multiplicata found in both units. The Schneider Hills limestone in the Argentina Range has yielded sclerites of Dailyatia icari sp. nov., currently only known from this location.
Key words: Tommotiida, Dailyatia, biostratigraphy, palaeobiogeography, Cambrian, Central Transantarctic Mountains.
Thomas M. Claybourn [thomas.claybourn@hdr.mqu.edu.au], Department of Earth Sciences, Palaeobiology, Uppsala University, Villav. 16, SE-75236, Uppsala, Sweden; Department of Biological Sciences, Macquarie University, North Ryde, Sydney, NSW, 2109, Australia.
Christian B. Skovsted [christian.skovsted@nrm.se], Department of Palaeobiology, Swedish Museum of Natural History, Box 50007, SE 104 05 Stockholm, Sweden; Early Life Institute and Department of Geology, State Key Laboratory for Continental Dynamics, Northwest University, Xi’an 710069, China.
Marissa J. Betts [marissa.betts@une.edu.au], Palaeoscience Research Centre, School of Environmental and Rural Science, University of New England, Armidale, NSW, 2351, Australia; Early Life Institute and Department of Geology, State Key Laboratory for Continental Dynamics, Northwest University, Xi’an 710069, China.
Lars E. Holmer [lars.holmer@pal.uu.se], Department of Earth Sciences, Palaeobiology, Uppsala University, Villav. 16, SE-75236, Uppsala, Sweden; Early Life Institute and Department of Geology, State Key Laboratory for Continental Dynamics, Northwest University, Xi’an 710069, China.
Lucy Bassett-Butt [lbassettbutt@gmail.com], Department of Earth Sciences, Palaeobiology, Uppsala University, Villav. 16, SE-75236, Uppsala, Sweden.
Glenn A. Brock [glenn.brock@mq.edu.au], Department of Biological Sciences, Macquarie University, North Ryde, Sydney, NSW, 2109, Australia; Early Life Institute and Department of Geology, State Key Laboratory for Continental Dynamics, Northwest University, Xi’an 710069, China.
Received 14 April 2020, accepted 26 July 2020, available online 21 January 2021.
Copyright © 2021 T.M. Claybourn et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The distant reaches of Antarctica have proven a difficult to sample, yet important source of information on the palaeobiology and palaeobiogeography of problematic animals from the traditional “lower Cambrian”. Expeditions to sample the lower Cambrian Byrd Group in the Central Transantarctic Mountains during the 20th Century yielded trilobites (Palmer and Gatehouse 1972; Palmer and Rowell 1995), archaeocyaths (Debrenne and Kruse 1986), molluscs (Rowell et al. 1988; Evans 1992) and other shelly fossils (Rowell et al. 1988; Evans and Rowell 1990). These were mostly from spot-samples, hindering detailed analysis of biostratigraphic or palaeobiogeographic context. Following fieldwork in 2011, the first systematically sampled sections intersecting the Byrd Group from the Churchill Mountains and Holyoake Range (Fig. 1) yielded moderately diverse assemblages of molluscs (Claybourn et al. 2019) and brachiopods (Claybourn et al. 2020). The palaeobiogeographical resolution of East Antarctica has been improved on the back of new systematic descriptions, corroborating links to South Australia and demonstrating new similarities with the brachiopod fauna of North China (Pan et al. 2019; Claybourn et al. 2020) and the molluscan fauna of East Laurentia (Landing and Bartowski 1996; Landing et al. 2002; Skovsted 2004; Atkins and Peel 2004; Peel and Skovsted 2005; Claybourn et al. 2019). Additional systematic description of the tommotiids from East Antarctica further advance and refine the biostratigraphic correlation and palaeobiogeography of this region.
The tommotiids have proven a valuable tool for biostratigraphy, but only in recent years. They have been used in a new biostratigraphic scheme for South Australia, which includes the Micrina etheridgei Zone and Dailyatia odyssei Zone (Betts et al. 2016, 2017), the eponyms of which are both tommotiid taxa. Dailyatia ajax Bischoff, 1976, Dailyatia macroptera Skovsted, Betts, Topper, and Brock, 2015, Dailyatia bacata Skovsted, Betts, Topper, and Brock, 2015, and Dailyatia helica Skovsted, Betts, Topper, and Brock, 2015, are all found in South Australia, where they are important accessory taxa for defining the M. etheridgei Zone (Betts et al. 2016) which spans much of Stage 2 of the Terreneuvian Series and the regionally important Parabadiella huoi Trilobite Zone at the base of Series 2, Stage 3 (Betts et al. 2016, 2018). Many of these taxa also range down into the underlying Kulparina rostrata Zone (Betts et al. 2016). Skovsted et al. (2015) recognised that early descriptions of Dailyatia ajax from South Australia (Bischoff 1976; Demidenko in Gravestock et al. 2001) belonged to four different species, demonstrating the importance of thorough systematic treatment of scleritome animals known only from disarticulated sclerites (Bengtson et al. 1990; Bengtson 2004).
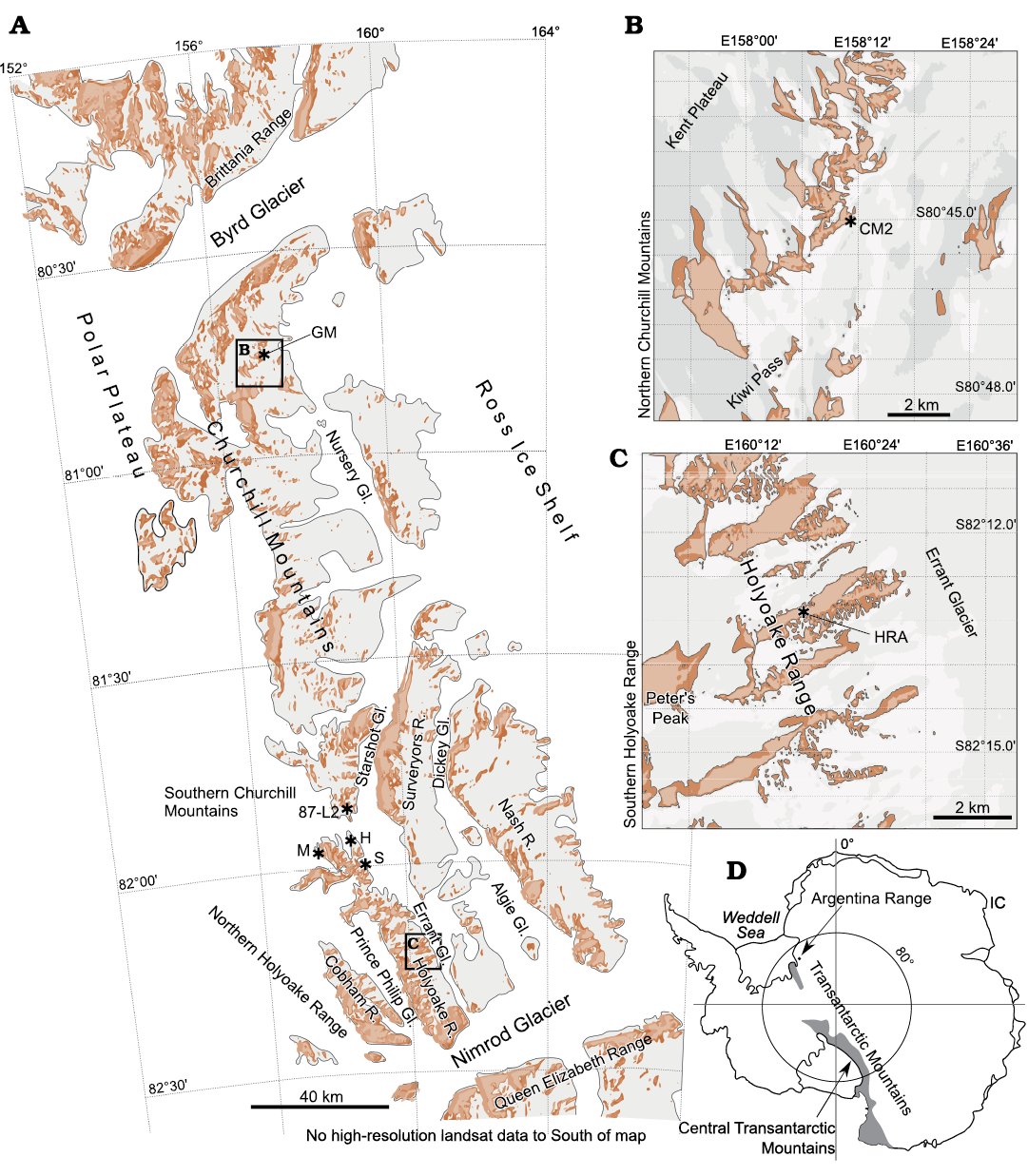
Fig. 1. A.
Topographic map of the Central Transantarctic Mountains (adapted
from the USGS 2008) with the main geographic
features and the sites visited by the Kansas University expeditions
(study areas marked by section names GM, 87-L2, M, H, S). The
locality GM is the type locality for Dailyatia
braddocki, which is close to the location of the CM2
section. The locality M is the type locality of Dailyatia
odyssei). B.
The area of the Churchill Mountains visited by the
Swedish-Australian expedition, showing the location of the CM2
section. C. The area of the
Holyoake Range visited by the Swedish-Australian expedition, showing
the location of the HRA section. D.
Overview of Antarctica showing the extent of the Transantarctic
Mountains, including the the location of the Argentina Range, and IC
(location on sections unknown) in the Schneider Hills (sampled by
the Kansas Expeditions).
Thorough systematic treatment of the morphologically variable tommotiids is also vital for understanding their phylogenetic position. This enigmatic group of armoured bilaterians with an external scleritome forms an assemblage of plesions scattered around the base of the phoronid + brachiopod clade. The relationship of the tommotiids to the lophophorates has been the subject of much recent research, with different members postulated as stems to different crowns. In broad terms, tommotiids can be assigned to two main groups (Skovsted et al. 2009a; Larsson et al. 2014: fig. 22). The first group are the eccentrothecimorphs, some of which occupy a stem position to the linguliform brachiopods, including the tannuolinids (Li and Xiao 2004; Holmer et al. 2008, 2011; Balthasar et al. 2009; Skovsted et al. 2014) and Paterimitra (Skovsted 2009b; Larsson et al. 2014). Another eccentrothecimorph taxon, Eccentrotheca has been interpreted to be in the phoronid stem group (Skovsted et al. 2008, 2011). The second group are the camenellan clade, including families Lapworthellidae, Tommotidae, and Kennardidae (Skovsted et al. 2009a). Camenellans share common features that unite them and distinguish them from those tommotiids that fall within the phoronid + brachiopod crown group. These include cone shaped sclerites (with a variable number of sclerites), a polygonal transverse section and growth series of comarginal ribs (Skovsted et al. 2009a, 2015; Devaere et al. 2014). The reconstruction of these scleritomes also illuminates dramatically different baupläne. The camenellans are reconstructed as having a worm- or slug-like appearance with the scleritome forming a dorsal armour (Evans and Rowell 1990: fig. 5; Skovsted et al. 2015: 100–103, fig. 79) and the sclerites of the eccentrothecimorphs have a variously modified tubular form (Holmer et al. 2008; Skovsted et al. 2011, Murdock et al. 2014).
Tommotiids from Cambrian Series 2 rocks of Antarctica were first published by Evans and Rowell (1990) from the Shackleton Limestone in the Central Transantarctic Mountains. They collected considerable numbers of Dailyatia sclerites and assigned them to two species: Dailyatia braddocki Evans and Rowell, 1990, and Dailyatia odyssei Evans and Rowell, 1990. The only other Antarctic tommotiid material discovered prior to the present study was limited to specimens from glacial erratics from the Miocene Cape Melville Formation of King George Island, north of the Antarctic Peninsula of West Antarctica (Wrona 1989, 2004). These include species of Dailyatia (which were assigned to Dailyatia ajax by Wrona 2004), rare sclerites of Dailyatia sp., the lapworthellid Lapworthella fasciculata Conway Morris in Bengtson et al., 1990 and fragments of the problematic Shetlandia multiplicata Wrona, 2004. As earlier stated, the use of abundant tommotiids from South Australia to form a regional chronostratigraphic scheme has demonstrated their utility as biostratigraphic tools after widespread and systematic sampling (Betts et al. 2016, 2017, 2018). However, the sparse data on tommotiids from East Antarctica has hindered their use in providing a biostratigraphic and palaeobiogeographic context to this region. The aim of this work is to rectify this by providing systematic descriptions on a wealth of new tommotiid data from East Antarctica.
Institutional abbreviations.—KUMIP, University of Kansas, Biodiversity Institute and Natural History Museum, Invertebrate Paleontology Collections, Lawrence, USA; NRM, Naturhistoriska riksmuseet (Swedish Museum of Natural History), Stockholm, Sweden.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in ZooBank: urn:lsid:zoobank.org:pub:B4E2734A-4AE6-4389-BEC9-511EDB32D4DA
Geological setting
The upper part of the Shackleton Limestone in the Holyoake Range consists of nodular carbonate, bedded limestone which is often highly bioturbated or oolitic and archoaecyath-microbiolite bioherms (Fig. 2). The Shackleton Limestone is overlain by the Holyoake Formation, an onlapping nodular siltstone, which in turn is overlain by the Starshot Formation. All three units are variably cross-cut by the Douglas Conglomerate. The transition from the Shackleton Limestone to Holyoake Formation probably represents the drowning of a carbonate platform (Rees et al. 1989; Myrow et al. 2002; Boger and Miller 2004) and change to deeper water facies associated with the Holyoake Formation (Goodge et al. 1993, 2004; Myrow et al. 2002). The Starshot Formation and Douglas Conglomerate are likely associated with the Ross Orogeny and mark the start of a collisional tectonic regime between East Antarctica and the palaeo-Pacific Plate (Rowell et al. 1988, 1992b; Myrow et al. 2002; Squire and Wilson 2005; Foden et al. 2006; Paulsen et al. 2007).

Fig. 2. Stratigraphic columns of the sampled sections in the Holyoake Range (HRA) and Churchill Mountains (CM2). Grain size: B, boundstone; G, grainstone; M, mudstone; P, packstone; W, wackestone.
The CM2 Section crosses 130 m of the Shackleton Limestone, with the top part of the section bounded by a severely deformed part of this unit (Fig. 2). The HRA section is 380 m and crosses 321.5 m of the uppermost Shackleton Limestone, although we only report findings from the lowest 150 m of this section, as the upper part of the Shackleton Limestone and younger units of the Byrd Group lack tommotiids. The remainder of this section is reported by Claybourn et al. (2019: fig. 2; 2020: fig. 2). In this unit, four horizons contain tommittid fossils.
The tommotiid-bearing Shackleton Limestone reported by Evans and Rowell (1990) from both the northern margin of the Holyoake Range (sections M, H, S, Fig. 1) and the northern Churchill Mountains (section GM, Fig. 1) are from parts of the Shackleton Limestone that have undergone severe deformation so should be treated as isolated faunas (Evans and Rowell 1990: 692). The Shackleton Limestone from M, H, S sections were described by Rees et al. (1989). The S and H sections contain a shallow-subtidal shelf association containing variable carbonates with biohermal reefs and bioturbation. Archaeocyathan-microbial reefs were reported from the H section (Rees et al. 1989: fig. 14). These sedimentological observations indicate a similar depositional environment between those sampled in the northern Holyoake Range and the new HRA section of the southern Holyoake Range.
A lesser known Cambrian Series 2 unit of East Antarctica, the informally named Schneider Hills limestone, crops out in the Schneider Hills of the southern Argentina Range (Fig. 1). The Schneider Hill limestone has an inadequately constrained biostratigraphy and may have been part of the same carbonate shelf that the Shackleton Limestone was deposited on (Rowell et al. 1992b). Rowell et al. (1992b) considered the Schneider Hills limestone as potentially a shallow-water equivalent of the deeper-water Hannah Ridge Formation (Rowell et al. 2001), which underlies the Drumian-aged Nelson Limestone (Rowell et al. 2001; Liebermann 2004; Bassett-Butt 2016), indicating a Series 2 or early Wuliuan age for these isolated outcrops. The archaeocyath fauna of the Schneider Hills limestone can be correlated with the Botoman Stage in the Siberian timescale (Debrenne and Kruse 1989).
The limited research that has been done on the outcrops in the Schneider Hills as well as other areas of the Argentina Range can give some clues to the age of this unit and relationship to other sedimentary packages of East Antarctica. The trilobite fauna collected from moraine boulders of unknown provenance at Mount Spann in the northern Argentina Range was described by Palmer and Gatehouse (1972). In-situ samples were not collected from Mount Spann, but an age can be estimated based on the rare non-endemic trilobites including Xystridura and Redlichia. Xystridura is known from Australia with two species collected at Mount Spann: X. glacia and X. multilina (Palmer and Gatehouse 1972). In Central and Northern Australia, the Xystridura negrina/Redlichia forresti Biozone covers the regional Ordian Stage (Laurie 2006), which lies within the upper part of Cambrian Series 2, Stage 4 (Sundberg et al. 2016), allowing for an approximate correlation based on this taxon. The upper range of Xystridura in the Northern Territory of Australia is not well constrained and has been recovered from the Tindall Limestone (Kruse 1990), Thorntonia Limestone and Arthur Creek Formation (Laurie 2012), where it ranges through the regional upper Ordian Stage into the Templetonian Stage (Laurie 2012; Smith et al. 2013), indicating Xystridura ranges into the Wuliuan Stage (Smith et al. 2013; Hally and Paterson 2014). Solenopleura pruina Palmer and Gatehouse, 1972, was also collected at Mount Spann, which has also been described from the Wuliuan-aged Nelson Limestone in the nearby Pensacola Mountains (Bassett-Butt 2016). These suggest that the succession in the Argentina Range spans Cambrian Stage 4 to the Wuliuan Stage, but it has not yet been thoroughly investigated biostratigraphically. Further systematic sampling of fossils from the Schneider Hills limestone may constrain this, as previously the only systematically described fossil from this unit is the bradoriid arthropod Bicarinella evansi Rode, Liebermann, and Rowell, 2003 (Rode et al. 2003), known only from this location. The presence of tommotiid sclerites belonging to Dailyatia in the Schneider Hills limestone has been noted previously, but they were not described (Rowell et al. 1992a; Rode et al. 2003).
Material and methods
The fossils described in this paper come from two different collections. New material from the Holyoake Range and Churchill Mountains of the Central Transantarctic Mountains (Figs. 1, 2, sections HRA, CM2) was collected in the Austral summer of 2011 (by GAB, LEH, CBS). Bulk carbonate was processed by dilute 5–10% acetic acid maceration at Uppsala University and Macquarie University to extract phosphatic specimens following protocols outlined in Jeppsson et al. (1999).
The remaining material was sampled during field expeditions organised by personnel at Kansas University who collected material in the austral summers of 1984, 1985, 1987, and 1989. These samples were collected from unmeasured sections in the Holyoake Range (localities prefixed H, M, S), Churchill Mountains (GM and 87-L2) and Argentina Range (IC), shown in Fig. 1. Some tommotiid material has been described from these locations and are not re-described here (Evans and Rowell 1990).
Scanning electron microscope imaging was prepared using a Zeiss Supra 35 SEM at Uppsala University; a Hitachi S-4300 SEM at the Natural History Museum, Stockholm; and a Phenom XL benchtop SEM at Macquarie University. Material collected from the HRA and CM2 sections are deposited at the Swedish Museum of Natural History in Stockholm, Sweden (Naturhistoriska riksmuseet, NRM). Material collected by the Kansas University expeditions are deposited in the Kansas University Museum of Invertebrate Paleontology (KUMIP).
We follow the terminology of Skovsted et al. (2015: 11–16, figs. 5–7). For specimens in open nomenclature more general terminology is used when possible to avoid confusion over any implications of homology.
Results
In the HRA section, in the southern Holyoake Range (Figs. 1, 2), the tommotiids are represented by Dailyatia cf. odyssei, Dailyatia sp. 1, and Shetlandia multiplicata. They are clustered at four horizons in archaeocyath-rich biohermal limestone of the upper Shackleton Limestone, in the lower parts of the section (samples HRA/14, 22, 24, 25). In the CM2 section, in the northern Churchill Mountains (Figs. 1, 2), Dailyatia braddocki and Dailyatia sp. 1 are present only at the collection locality marking the base of the section (samples CM2/130). The results are summarised in Table 1 and Fig. 2.
The H section (Fig. 1), in the northern Holyoake Range (Table 2) yielded predominantly sclerites of Dailyatia decobruta Betts in Betts et al., 2019 with rare fragments of Dailyatia sp. 1. The M section (Fig. 1) yielded almost exclusively sclerites from Kennardiidae indet., in addition to a single D. decobruta sclerite. The samples from the Argentina Range (section IC, Fig. 1) yield exclusively sclerites of Dailyatia icari sp. nov., which is known only from this area. The findings from the northern Holyoake Range and Argentina Range are summarised in Table 2.
Table 2. Sample localities for taxa described from previously collected samples described in Evans and Rowell (1990) from the Holyoake Range, Churchill Mountains, and Argentina Range. Abbreviations: f, fragments.
| |
Dailyatia
icari |
Dailyatia sp. 1 |
Dailyatia decobruta |
Kennardiidae indet. |
|||||||
|
Sclerite type |
f |
A |
C1 |
C2 |
f |
f |
C |
f |
1 |
2 |
3 |
|
Section H, northern Holyoake Range |
|||||||||||
|
H84.2 |
|
|
|
|
|
|
1 |
|
|
|
|
|
H84.6 |
|
|
|
|
|
1 |
|
|
|
|
|
|
H84.17 |
|
|
|
|
|
1 |
|
|
|
|
|
|
H84.20 |
|
|
|
|
1 |
7 |
|
|
|
|
|
|
H84.25 |
|
|
|
|
|
9 |
|
|
|
|
|
|
H84.26 |
|
|
|
|
1 |
15 |
2 |
|
|
|
|
|
H85.25 |
|
|
|
|
|
2 |
|
|
|
|
|
|
Section M, northern Holyoake Range |
|||||||||||
|
M84.1 |
|
|
|
|
|
|
1 |
4 |
|
|
1 |
|
M84.2 |
|
|
|
|
|
|
|
3 |
1 |
1 |
5 |
|
M87.1 |
|
|
|
|
|
|
|
|
|
|
1 |
|
M87.3 |
|
|
|
|
|
|
|
1 |
|
|
|
|
M87.4 |
|
|
|
|
|
|
|
5 |
|
|
|
|
Section IC, Argentina Range |
|||||||||||
|
IC84.2 |
2 |
|
|
|
|
|
|
|
|
|
|
|
IC89.1 |
11 |
|
1 |
|
|
|
|
|
|
|
|
|
IC89.2 |
50+ |
5 |
4 |
1 |
|
|
|
|
|
|
|
|
IC3B |
12 |
|
2 |
2 |
|
|
|
|
|
|
|
Biostratigraphy
Cambrian Series 2 strata of East Antarctica are well exposed in the Churchill Mountains and Holyoake Range in the Central Transantarctic Mountains (Fig. 1), where the fossiliferous units of the Byrd Group are exposed. Combined data from newly described molluscs (Claybourn et al. 2019) and brachiopods (Claybourn et al. 2020) indicate a Cambrian Epoch 2, Age 4 for the upper part of the Byrd Group (upper Shackleton Limestone, Holyoake and Starshot formations), correlatable with the uppermost part of the Dailyatia odyssei Zone in South Australia (Betts et al. 2017). Previously collected D. odyssei from the northern Holyoake Range (sections M, H, S, Fig. 1; Evans and Rowell 1990) directly correlates this part of the Shackleton Limestone to the D. odyssei Zone of South Australia. Re-examined sclerites assigned in this work to Dailyatia decobruta from the northern Holyoake Range also provide correlation to the carbonate clasts of the White Point Conglomerate recovered from Kangaroo Island, South Australia (Betts et al. 2019) where it also occurs with D. odyssei. The assemblage of small shelly fossils recovered from the White Point Conglomerate clasts indicated an Epoch 2, Age 4 age, correlating to the Pararaia janeae Trilobite Zone and upper D. odyssei Zone (Jell in Bengtson et al. 1990; Betts et al. 2017). These fossils corroborated the original age designation of Jell in Bengtson et al. (1990) who recognised the trilobite assemblage of the White Point Conglomerate as part of their P. janeae Zone. Dailyatia odyssei from the Shackleton Limestone has only been recovered from the Churchill Mountains and northern Holyoake Range (Evans and Rowell 1990). Abundant D. odyssei sclerites reported by Evans and Rowell (1990) co-occurring with rarer D. decobruta enables correlation to the upper D. odyssei Zone and P. janeae Zone of the Mernmerna Formation (section NB; Betts et al. 2017: fig. 10). Species of Dailyatia which bear strong similarity to Dailyatia decobruta have also been recovered from the glacial erratics of King George Island (Wrona 2004), although reanalysis of this material is required to understand the taxonomic affinity of these specimens.
In the southern Holyoake Range (section HRA, Figs. 1, 2) an assemblage including D. cf. odyssei, Dailyatia sp. 1, and Shetlandia multiplicata is present. Biostratigraphic correlation is less clear based on these fossils alone, with no temporally constrained tommotiid fossils found in this section. The presence of S. multiplicata is the first example of this enigmatic taxon from autochthonous carbonates; it was previously only known from the glacial erratics of King George Island where it occurs with Dailyatia (Wrona 2004). The lack of D. odyssei may also indicate these fossils occur above the last appearance datum for this taxon (although specimens of D. cf. odyssei do occur here).
The tommotiid fauna from the Argentina Range consists exclusively of Dailyatia icari sp. nov., which is currently only known from this region. The Schneider Hills limestone remains biostratigraphically unconstrained with the discovery of this species.
Palaeobiogeography
Within Antarctica.—Original descriptions of Dailyatia from Antarctica were made by Evans and Rowell (1990) from collection sites in the northern Holyoake Range and southern and northern areas of the Churchill Mountains (Fig. 1; Evans and Rowell 1990: fig. 1). The endemic Dailyatia braddocki was only recovered from the northern Churchill Mountains (locality GM87.1 of the GM section, Fig. 1; Evans and Rowell 1990: fig. 1, table 1). In our investigation, this species was also recovered from the CM2 section (Fig. 2), in the same region of the Churchill Mountains (Fig. 1). Within Antarctica, previously described Dailyatia odyssei is also known from the northern and southern parts of the Churchill Mountains and the northern Holyoake Range in the Central Transantarctic Mountains (Fig. 1, localities GM, 87-L2, H, M, S; Evans and Rowell 1990: fig. 1, table 1). New material comparable to D. odyssei from the Holyoake Range, include only poorly preserved C sclerites of Dailyatia cf. odyssei in the southern Holyoake Range (section HRA, Figs. 1, 2).
Table 1. Sample localities for taxa described from new collections, taken from systematically sampled sections in the Southern Holyoake Range and Northern Churchill Mountains. Abbreviations: f, fragments.
| |
Dailyatia
|
Dailyatia braddocki |
Dailyatia cf. odyssei |
Shetlandia |
||||
|
Sclerite type |
f |
A |
C |
f |
C |
f |
C |
f |
|
Section HRA, southern Holyoake Range |
||||||||
|
HRA/14 |
16 |
1 |
3 |
|
|
|
|
6 |
|
HRA/25 |
|
|
|
|
|
|
|
9 |
|
HRA/24 |
3 |
|
|
|
|
|
|
7 |
|
HRA/22 |
3 |
|
|
|
|
2 |
2 |
|
|
Section CM2, northern Churchill Mountains |
||||||||
|
CM2/130 |
33 |
|
|
9 |
8 |
|
|
|
The Shackleton Limestone of the southern Holyoake Range (section HRA, Figs. 1, 2) contains a faunule including three tommotiids: Dailyatia sp. 1, Dailyatia cf. odyssey, and Shetlandia multiplicata. The Shackleton Limestone in the northern Holyoake Range has a different assemblage of tommotiids, including Dailyatia sp. 1, Dailyatia decobruta, and Kennardiidae indet. originally described by Evans and Rowell (1990) and identified here as Dailyatia odyssei. The northern Holyoake Range also hosts the type locality for Dailyatia odyssei (locality M84.2; Evans and Rowell 1990). As such Dailyatia sp. 1 is the only tommotiid species that occurs in both the northern and southern regions in the Holyoake Range, although it is known only from two sclerite fragments in the northern Holyoake Range (Table 1). In the northern Churchill Mountains (sections CM2 and GM, Figs. 1, 2), D. odyssei co-occurs with Dailyatia braddocki (Evans and Rowell 1990: table 1), making D. odyssei the only faunal link between the northern Holyoake Range and the northern Churchill Mountains.
These taxa also provide evidence for the provenance of glacial erratics recovered from the Miocene glaciomarine Cape Melville Formation of King George Island. These erratics yielded Shetlandia multiplicata which also occurs in the autochthonous carbonates of East Antarctica (Fig. 3; Wrona 2004). Shetlandia multiplicata is known from the Shackleton Limestone of the southern part of the Holyoake Range (section HRA, Figs. 1, 2, Table 1). The Shackleton Limestone is a likely source for these erratics, as suggested by Wrona (2004), as they also contain the camenellan tommotiid Lapworthella fasciculata (Wrona 2004) and similar assemblages of brachiopods (Holmer et al. 1996) and bradoriids (Wrona 2009) not currently known from the autochthonous rocks of the Shackleton Limestone. The Argentina Range yields sclerites of Dailyatia icari sp. nov. (Fig. 3). This species is currently only known from disarticulated sclerites of the temporally unconstrained Schneider Hills limestone.
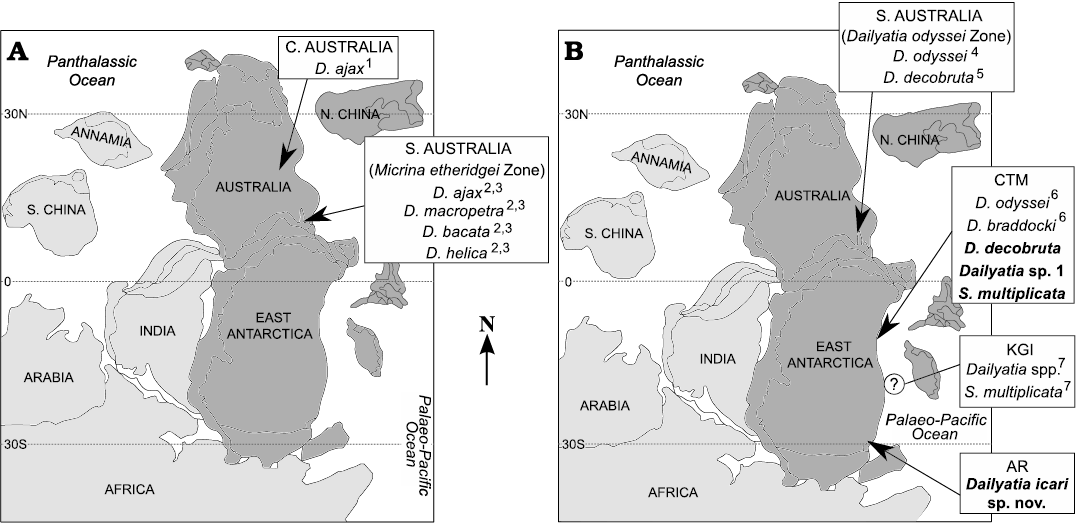
Fig. 3. Palaeobiogeographic maps of the distribution of Dailyatia and other camenellan tommotiids from East Gondwana (dark grey). New occurrences described in this paper in bold. Maps show the Cambrian (A) Terreneuvian Series to basal Series 2, Stage 3 (i.e., upper range of the Micrina etheridgei Zone for South Australia, Betts et al. 2016, 2017) and (B) Series 2 (from the base of the Dailyatia odyssei Zone for South Australia, Betts et al. 2016, 2017). Data for previous occurrences: 1 Laurie 1986; 2 Skovsted et al. 2015; 3 Betts et al. 2016; 4 Betts et al. 2017; 5 Betts et al. 2019; 6 Evans and Rowell 1990; 7 Wrona 2004. Abbreviations: CTM, Central Transantarctic Mountains; KGI, King George Island; AR, Argentina Range. Map adapted from Torsvik and Cocks (2013) and Yang et al. (2015).
East Gondwana.—Of the seven previously named species of Dailyatia, which all occur in the Cambrian Series 2 of East Gondwana, only Dailyatia odyssei and Dailyatia decobruta occur in both East Antarctica and South Australia (Fig. 3). Dailyatia braddocki (Shackleton Limestone, Churchill Mountains), the newly described Dailyatia icari sp. nov. (Argentina Range), Dailyatia sp. 1, and Kennardiidae indet. (Shackleton Limestone, Holyoake Range), and the problematic Shetlandia multiplicata (Shackleton Limestone, Holyoake Range and the King George Island erratics) are apparently endemic to East Antarctica (Fig. 3). In East Antarctica, Dailyatia decobruta has been recovered from the Shackleton Limestone, with the majority of South Australian specimens collected from the allochthonous limestone clasts of the White Point Conglomerate of Kangaroo Island. The carbonate clasts of the White Point Conglomerate have uncertain provenance but bear faunal similarities to shelly fossil assemblages from the Koolywurtie Limestone Member of the Parara Limestone of the Yorke Peninsula, South Australia (Paterson et al. 2007; Betts et al. 2019). Rare sclerites of D. decobruta are also found in the Mernmerna Formation of the Arrowie Basin (“Dailyatia sp. A” of Skovsted et al. 2015; Betts et al. 2017).
Dailyatia odyssei is also present in both East Antarctica and South Australia. Sclerites were reported from the Shackleton Limestone by Evans and Rowell (1990), where they were found widespread in the rocks sampled in the northern Churchill Mountains and northern Holyoake Range (Fig. 2). In the southern Holyoake Range, the new HRA section yielded no unambiguous D. odyssei sclerites, with only a few poorly preserved sclerites tentatively referred to D. cf. odyssei. In South Australia, D. odyssei is widespread in its eponymous D. odyssei Zone of both the Arrowie and Stansbury basins (Gravestock et al. 2001; Skovsted et al. 2015; Betts et al. 2017).
Systematic palaeontology
Lophophorata Hyman, 1959
Class Incertae sedis
Order Tommotiida Missarzhevsky, 1970
Family Kennardiidae Laurie, 1986
Dailyatia Bischoff, 1976
Type species: Dailyatia ajax Bischoff, 1976, lower Cambrian, Ajax Limestone, Mt. Scott Range, northern Flinders Ranges, South Australia.
Dailyatia icari sp. nov.
Figs. 4–6.
ZooBank LSID: urn:lsid:zoobank.org:act:33673DE4-7750-48D8-BC 9C-7DCBE8198C7A
Etymology: From Latin Icarus (latinized genitive derivation of the Greek Ikaros), the legendary Greek character who flew to close to the sun; in reference to the radial plicae resembling rays of the sun.
Type material: Holotype: KUMIP 585059, C1 sclerite from locality IC3B (Fig. 5A). Paratypes: KUMIP 585054, A sclerite from locality IC89.2 (Fig. 4A); KUMIP 585070, C2 sclerite from locality IC3B (Fig. 6C).
Type locality: Sample locality IC3B of the Schneider Hills limestone, Argentina Range, East Antarctica.
Type horizon: Unknown horizon in Cambrian Series 2.
Material.—Six A sclerites from sample IC89.2 (KUMIP 585054–585058, 5 figured), six C1 sclerites from sample IC3B (KUMIP 585059, 585060, 2 figured), one C1 sclerite from sample IC89.1 (KUMIP 585061), four C1 sclerites from IC89.2 (KUMIP 585062, 585064, 585066, 3 figured), one C2 sclerite from sample IC89.2 (KUMIP 585068), two C2 sclerites from sample IC3B (KUMIP 585069, 585070). Numerous fragments from sample IC84.2, IC89.1, IC89.2 (KUMIP 585063, 585065, 585067, 3 figured) and IC3B, listed in Table 2. Cambrian Series 2, Schneider Hills limestone, Argentina Range, East Antarctica.
Diagnosis.—Species of Dailyatia with distinctive ornament of pseudoplicae formed by pustules in a single or double-row. Ornament of densely set, narrow concentric ribs. Sclerite subtypes C1 and C2 recognised, no subtypes recognised for A sclerites. A sclerites pyramidal, triangular or pentagonal in transverse section. No plicae present on concave anterior field. Apex slightly coiled anteriorly. C1 sclerites slightly coiled over ventral side with array of well-developed and regular radial plicae on dorsal side. Dorsal and proximal edges with pseudoplicae, ventral side concave with comarginal ribs and lacking pseudoplicae. C2 sclerites strongly torted, triangular in transverse section and coiled ventrally. Ventral field concave, with weakly developed pseudoplicae. Dorsal field with broad central surface bounded by broad furrows separating the central plicae from the distal and marginal edges.
Description.—Sclerites pyramidal with two sclerite types identifiable (A and C) and two C sclerite subtypes identifiable: C1 and C2. C1 sclerites dorso-ventrally compressed (Fig. 5A1, A4, B1, C2, D1, H2, H3), C2 sclerites conical and torted (Fig. 6A1, C1).
A sclerites conical and bilaterally symmetrical. Comarginal ribs are dense and cover sclerites with larger specimens with closely packed rows of pseudoplicae formed by rounded pustules (Fig. 4A). Pseudoplicae are only weakly developed on smaller specimens (Fig. 4D, E). Anterior field is concave and lacking plicae (Fig. 4A1, A3, B, C, D1, D2 E1, E2). Growth disturbances are present on the anterior surface in the form of comarginal depressions (Fig. 4A1, B, C, D1). The anterior field is separated from the lateral fields by a deep furrow and an anterolateral plication (Fig. 4A1, A3, A4, C, D1–D3, E1, E2). The lateral field is concave, with dense pseudoplicae (Fig. 4A3) or lacking pseudoplicae (Fig. 4E2). The posterior field is partially preserved in part on two sclerites where they are separated from the lateral field by a weakly developed posterolateral plication (Fig. 4D2, E2). A deltoid cannot be clearly delineated on the posterior field.

Fig. 4. The camenellan tommotiid Dailyatia icari sp. nov. A sclerites from the lower Cambrian Schneider Hills limestone, Schneider Hills, Argentina Range, Antarctica. A. KUMIP 585054, view of anterior field (A1), enlarged view of the anterior apical area (A2), oblique view of the lateral field (A3), apical view with anterior to the top (A4). B. KUMIP 585055, anterior field of broken sclerite. C. KUMIP 585056 anterior field of broken sclerite. D. KUMIP 585057, view of anterior field (D1), apical view with anterior to the top (D2), lateral view (D3). E. KUMIP 585058, view of anterior field (E1), apical view with anterior to the top (E2). Scale bars 200 µm.
C1 sclerites all have dorsoventral compression and a broad V-shape when viewed apically (Fig. 5A1, B1, C2, D1). The central part of the ventral side lacks plicae and pseudoplicae but has 1–3 weakly developed pseudoplicae at the proximal edge (Fig. 5A3, B1, D3). Pseudoplicae are lacking on the central surface of the dorsal side but are present on the proximal edge (2–5 pseudoplicae, Fig. 5A1, A5, B2, C2, D1) and distal edge (1–3 pseudoplicae, Fig. 5A1, A4, B2, C2, D1). The central surface of the dorsal side is dominated by 7–8 strongly developed radiating plicae which dominate the entire field forming an evenly curved central surface (Fig. 5A1, A2, B1, B2, C1, C2, D1, C2, F1, F2). The plications start at the smooth, unornamented apex and radiate towards the aperture of the sclerite (Fig. 5A1, B2, D2). One sinistral C1 sclerite is slightly torted, with the apex overhanging the proximal edge (Fig. 5B).
C2 sclerites are strongly torted, pyramidal and triangular in transverse section (Fig. 6A1, B1, C1). The ventral field is concave and lacks plicae but has 2–7 weakly developed pseudoplicae and is separated from the proximal and distal edges by furrows (Fig. 6A3, B3, C3). The dorsal field has a broad central surface, covered by densely set pseudoplicae formed by rounded pustules (Fig. 6A4, A5, B1, C2, C4, C5). The central area is bounded proximally by a plication, with a broad furrow separating the central surface from the proximal edge (Fig. 6A2, C1, C4). A narrow furrow with three plicae separating the central surface from the distal edge is present in one specimen (Fig. 6C1, C2, C5).
The comarginal ribs are ubiquitously distributed across all persevered sclerites, except at the smooth apex of the C1 sclerite (Fig. 5A1, B2, D2) and some A sclerites (Fig. 5E) and are separated by narrow inter-rib grooves. The pseudoplicae are typically unevenly spaced and are delineated by a series of broad pustules (Fig. 5E, G, I), separated by furrows (Fig. 5G). On one C2 sclerite the pseudoplicae are variably developed on the dorsal fields with well-developed pustules developed in a central band, with only comarginal ribs towards the apex and weakly developed, intermittent bands of pustules towards the base (Fig. 6C). Some sclerite fragments have pseudoplicae formed by paired radial rows of pustules (Figs. 5E, G, I, 6A6). This arrangement develops out of a single radial row of pustules and splits into a paired radial row towards the base, the pairs forming single pseudoplicae (Fig. 5E–G). These paired pustules are arranged along comarginal ribs in the same manner as other specimens’ pseudoplicae with single columns of pustules (Fig. 5E–G, compare Dailyatia bacata, Skovsted et al. 2015: fig. 34). Reticulate micro-ornament seen in other Dailyatia species is not present on most specimens, but a poorly preserved example may be seen on one (Fig. 6A6).
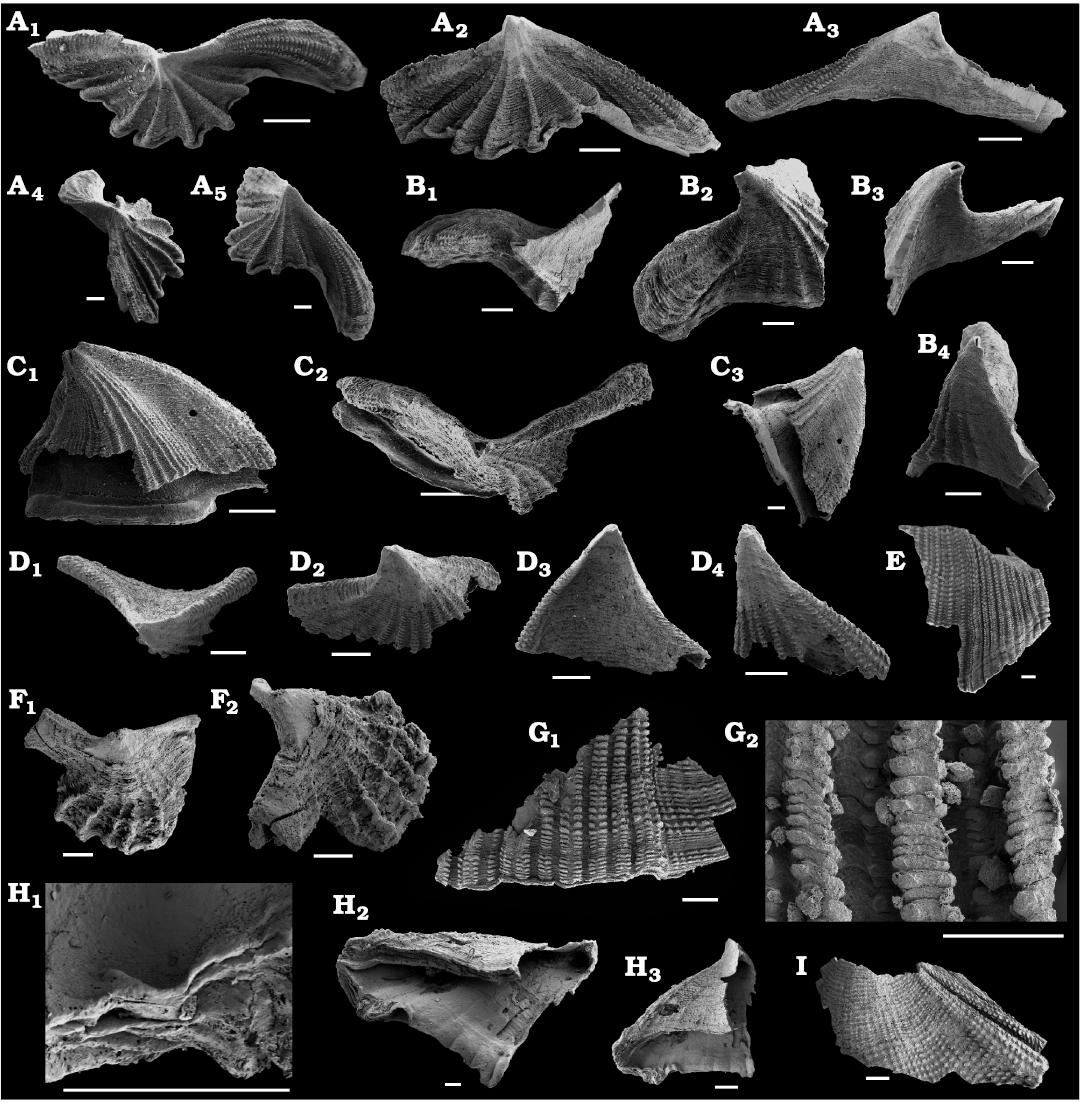
Fig. 5. The camenellan tommotiid Dailyatia icari sp. nov. C1 sclerites and sclerite fragments from the lower Cambrian Schneider Hills limestone, Schneider Hills, Argentina Range, East Antarctica. A. Dextral sclerite, KUMIP 585059, apical view (A1), view of dorsal (A2) and ventral (A3) surfaces, view of distal (A4) and proximal (A5) edges. B. Sinistral sclerite, KUMIP 585060, apical view (B1), oblique view of proximal edge and dorsal surface (B2), oblique view of distal edge and ventral surface (B3), view of distal edge (B4). C. Dextral sclerite, KUMIP 585061, dorsal (C1), apical (C2), and oblique dextral (C3) views. D. Dextral sclerite, KUMIP 585062, apical (D1), apical-dorsal (D2), ventral (D3), and lateral (D4) views. E. Large fragment from unknown sclerite type, KUMIP 585063. F. Sclerite of unknown chirality, KUMIP 585064, apical (F1) and lateral (F2) views. G. Sclerite fragment from unknown sclerite type, KUMIP 585065, detail showing distinctive pseudoplicae (G1), general view (G2). H. Sclerite of unknown chirality, KUMIP 585066, detail of apertural margin (H1), oblique views of the aperture (H2, H3). I. Fragment of unknown sclerite type showing pseudoplicae, KUMIP 585067. Scale bars 200 µm.F

Fig. 6. The camenellan tommotiid Dailyatia icari sp. nov. C2 sclerites from the lower Cambrian Schneider Hills limestone, Schneider Hills, Argentina Range, East Antarctica. A. KUMIP 585068, apical (A1), oblique lateral (A2), ventral (A3), lateral (A4), and dorsal (A5) views, detail of pustulose ornament on the central surface of A1 (A6). B. KUMIP 585069, apical (B1), lateral (B2), and ventral (B3) views. C. KUMIP 585070, apical (C1), lateral-dorsal (C2), ventral (C3), lateral (C4), and dorsal (C5) views. Scale bars 200 µm, except A6, 100 µm.
Remarks.—The few available specimens of Dailyatia icari sp. nov. from the Schneider Hills limestone in the Argentina Range have a distinct morphology and ornament clearly delineating them from other species of Dailyatia. The A sclerites of D. icari are not well preserved, with the posterior part of the sclerite typically broken. A distinguishing feature of the A sclerites of D. icari is the well-developed furrow at the lateral edges of the anterior field bounding the well-developed anterolateral plication, which is not found on other species of Dailyatia. The distinctive radially plicate dorsal side of the C1 sclerites is also unique amongst species of Dailyatia. Other species, like Dailyatia ajax (Skovsted et al. 2015: figs. 16C, L, N, P, 17Q) and D. helica (Skovsted et al. 2015: fig. 39A–F, K–T, W, X) share a well-developed series of plicae on the dorsal side of C1 sclerites. However, the C1 sclerites of D. ajax are conical and taller than they are wide (Skovsted et al. 2015: figs. 15–17), unlike the squat C1 sclerites of D. icari, which are typically wider than they are tall (Fig. 5A1, A2, C1, C2). The C sclerites of D. helica have a dorsally elongated central surface which forms a deep recession between the central surface and the distal edge (Skovsted et al. 2015: fig. 39), a feature not present in D. icari sp. nov. The C1 sclerites of D. icari sp. nov. are dorsoventrally flattened and have a broad V-shape when viewed apically (Fig. 5A1, C2, D1, F1). This is similar to the C sclerites of Dailyatia braddocki (Evans and Rowell 1990: figs. 6.8, 6.14). Dailyatia icari is easily distinguished from D. braddocki as the C sclerites of D. braddocki lacks radial plicae and pseudoplicae (Evans and Rowell 1990: figs. 6.7–6.14).
The C2 sclerites of D. icari sp. nov. can be distinguished from those of other species by the broad central surface. For example, the C2 sclerites of Dailyatia odyssei have a comparable degree of torsion, but have a well-defined central plication (Skovsted et al. 2015: fig. 49A–H). The C2 sclerites of Dailyatia macroptera are also strongly torted, but these have a dorsal surface with a deep concave surface (Skovsted et al. 2015: fig. 25A–I, M–S), easily distinguishing them from the convex central surface on the dorsal field of C2 sclerites of D. icari.
Dailyatia icari sp. nov. have pseudoplicae formed by radial rows of pustules. Similar pseudoplicae are found in other species, most notably Dailyatia decobruta (Betts et al. 2019: fig. 17G–I) and Dailyatia bacata (Skovsted et al. 2015: fig. 34), however, the pseudoplicae of D. bacata have crenulated walls between them which are not present in D. icari sp. nov. Dailyatia bacata also lacks the pseudoplicae formed by paired pustules that occurs in some specimens of D. icari sp. nov. The pustules of D. bacata also protrude further from the main body of the sclerite than those of D. icari (Skovsted et al. 2015: fig. 34B, H), and have a reticulate ornamentation both on the pustules and the depressions between their pseudoplicae (Skovsted et al. 2015: fig. 34C, H), whereas D. icari sp. nov. lacks such micro-ornamentation.
Dailyatia decobruta, known from both the Cambrian Series 2, Stage 4 carbonate clasts of the White Point Conglomerate, Kangaroo Island, and the Mernmerna Formation in the Flinders Ranges, South Australia (Betts et al. 2019: figs. 15–18) and the Shackleton Limestone of the northern Holyoake Range (Fig. 7C–F) also has dense pustules arranged in pseudoplicae (Betts et al. 2019: fig. 17G–I). The C sclerites of D. decobruta are easily distinguishable from C sclerites of D. icari as they lack radiating central plicae on their dorsal side (Betts et al. 2019: figs. 16D, E, H–J, 17A, B). The C2 sclerites of D. decobruta are similarly torted to the C2 sclerites of D. icari. However, the strong dorsoventral compression of the C2 sclerites of D. decobruta (Betts et al. 2019: fig. 17D–F) clearly distinguish these species, as the C2 sclerites of D. icari are pyramidal in shape (Fig. 6A1, C1).
Another East Antarctic endemic, Dailyatia sp. 1, from the Shackleton Limestone in the Holyoake Range, has an arrangement of chaotically distributed plicae with small rows of crests with a reticulated ornament rather than pustules (Fig. 9F2). These often form oblique rows that join the main sequence of concentric ribs at irregular intervals (Fig. 8A1, 9F, G). This ornamentation distinguishes these fragments from D. icari sp. nov. which exhibits single or paired pustulose pseudoplicae.
A broken D. icari sp. nov. specimen shows the typically continuous growth laminations of Dailyatia (Fig. 5H). Small pits are visible at the apertural margin at the base of the sclerite may be the imprints of epithelial cell moulds (Fig. 5H1).
Stratigraphic and geographic range.—Unknown horizon in Cambrian Series 2, Schneider Hills limestone, Argentina Range, East Antarctica (Table 2).
Dailyatia braddocki Evans and Rowell, 1990
Fig. 7A, B.
1988 Dailyatia spp.; Rowell et al. 1988: figs. J, K, M.
1990 Dailyatia braddocki n. sp.; Evans and Rowell 1990: 696, figs. 6.1–6.17.
2015 Dailyatia braddocki: Skovsted et al. 2015: fig. 9A–F.
Material.—Eight broken C sclerites from the CM2/130 sample (NRM X10001, X10002, 2 figured) (Table 1, Fig. 2). Cambrian Series 2, Stages 3, 4, Shackleton Limestone of the Northern Churchill Mountains, East Antarctica.
Description.—C sclerites with strong dorso-ventral compression. A single well-developed plica is preserved on proximal edge of ventral side, projecting slightly away from the edge, partially covering a small furrow on the ventral side (Fig. 7A2, B1). On the ventral side, the distal edge is separated from a plica by a furrow (Fig. 7A). Sclerites with stepped concentric ribs and ornament of small pustules arranged along the ribs but do not form pseudoplicae (Fig. 7A2, B2).

Fig. 7. The camenellan tommotiids from from the Cambrian Series 2, Stages 3, 4, Shackleton Limestone, Transantarctic Mountains, East Antarctica. Dailyatia braddocki Evans and Rowell, 1990, Churchill Mountains (A, B), Dailyatia decobruta Betts in Betts et al., 2019, Holyoake Range (C–F), and Dailyatia cf. odyssei, Holyoake Range (G–I). A. C sclerite, NRM X10001, ventral view (A1), detail showing fold in the margin of the ventral field (A2). B. Sclerite of unknown chirality, NRM X10002, dorsal view (B1), detail showing apex (B2). C. Dextral C sclerite, KUMIP 585071, details showing micro-ornament on the anterior-lateral (C1) and the posterior (C3) fields, apical view (C2). D. Dextral C sclerite, KUMIP 585072, apical view (D1), view of proximal edge and dorsal field (D2). E. Sclerite of unknown chirality, KUMIP 585073, oblique view of ventral field (E1), view of apical area showing perforated apex (E2). F. Sclerite of unknown chirality, KUMIP 585074, dorsal (F1) and oblique apical (F2) views. G. Dextral C sclerite, NRM X10003, view of broken dorsal field (G1), detail showing fine growth series (G2), apex (G3), oblique basal view showing plicae at distal edge (G4). H. Sinistral C sclerite, NRM X10004, apical view (H1), oblique view of distal edge and part of ventral field (H2). I. Unknown sclerite type, NRM X10005, ventral view (I1), detail showing fine growth series (I2). Scale bars 200 µm, except E2, 50 µm.
Remarks.—The sclerites found in the CM2 section in the Churchill Mountains are both broken but fall within the diagnosis for Dailyatia braddocki (Evans and Rowell 1990: 696). Dailyatia braddocki was originally only found at one locality in the northern Churchill Mountains (GM87.1, Fig. 1; Evans and Rowell 1990: table 1), close to the CM2 locality (Fig. 1) reported herein. The broad-shaped profile of the sclerites with single radial plica on the ventral side at the proximal edge are the same as those from the original collections of Evans and Rowell (1990: figs. 6.7, 7–10, 13, 14). The weakly developed pustules of D. braddocki were not described or figured by Evans and Rowell (1990) but reimaging material from their GM87.1 locality (Fig. 1) shows this ornamentation is present (Fig. 9H, I; Skovsted et al. 2015: fig. 9A–E).
Stratigraphic and geographic range.—Cambrian Series 2, Stages 3, 4, Shackleton Limestone of the Northern Churchill Mountains, Central Transantarctic Mountains, East Antarctica (Table 1, Evans and Rowell 1990).
Dailyatia decobruta Betts in Betts et al., 2019
Fig. 7C–F.
cf. 2004 Dailyatia ajax; Wrona 2004: figs. 9A–D, 11A, B, 12A.
2015 Dailyatia sp. A; Skovsted et al. 2015: fig. 51.
2019 Dailyatia decobruta sp. nov.; Betts et al. 2019: 514–515, figs. 15–18.
Material.—Four C1 sclerites, one from H.84.2, two from H.84.26, and one from M84.1 (KUMIP 585071–585074, all figured) (Fig. 1, Table 2; Evans and Rowell 1990). Fragmentary material found through the H and M sections of Evans and Rowell (1990), listed in Table 2. Cambrian Series 2, Stage 4(?) of the Shackleton Limestone in northern Holyoake Range, East Antarctica.
Description.—The specimens fit within the diagnosis (Betts et al. 2019: 513–515) for C1 sclerites. They are triangular in cross section (Fig. 7C2, D1) with the apex slightly coiled over the concave ventral field. Dense pustulose ornamentation forms pseudoplicae on the convex dorsal field (Fig. 7C1, C3). The pustules and troughs between the pseudoplicae are covered in a fine reticulate micro-ornamentation (Fig. 7C1, C3).
Plicae are generally poorly developed, but a broad central radial plication on the dorsal field is present in two specimens (Fig. 7C2, D1, F) and a single plica is present on the dorsal field adjacent to the proximal edge in one specimen (Fig. 7E1). The ventral field lacks plicae. The distal and proximal edges are variable, with broad rounded edges in two specimens (Fig. 7C2, D1) and narrow edges in two (Fig. 7E1, F2). The apertural margins of the sclerites from the Shackleton Limestone are broken, so information on the complete size and shape of the aperture is missing. A single minute oval perforation is present on the apices of all specimens (Fig. 7C2, D1, E2, F2) with a width of 31–69 µm and a length of 15–30 µm (n = 3). The apical part is smooth in all but one specimen (Fig. 7E).
Remarks.—Ornament consisting of pustules arranged into regular rows (pseudoplicae) and covered by reticulate micro-ornament is a key characteristic of this species. The three specimens from the Shackleton Limestone exhibit this ornamentation and are overall very closely similar to those recently described from the White Point Conglomerate (Betts et al. 2019: 513–515), though they vary in some non-diagnostic characteristics. For example, the Antarctic specimens lack the torsion seen in the C2 and C2a sclerites from the White Point Conglomerate (Betts et al. 2019: fig. 18). The extent of the pustulose ornament also differs slightly, which covers the entire sclerite surface, including the apex in specimens from the White Point Conglomerate (Betts et al. 2019: fig. 17A–C), but is not present on the apical area in specimens from the Shackleton Limestone, which is smooth. The ventral surface of two C1 sclerites from the Shackleton Limestone are also slightly less dorsoventrally compressed than those figured by Betts et al. (2019: fig. 16). The specimens of D. decobruta from the White Point Conglomerate also have an apex that overhangs the ventral part of the aperture when viewed apically (Betts et al. 2019: fig. 16D2, E, I2), whereas the apertural margin in specimens from the Shackleton Limestone extend out beyond the apex, increasing the size of the internal cavity (Fig. 7C2, D1, D2, E1).
Dailyatia decobruta co-occurs with two fragments of Dailyatia sp. 1 in the Shackleton Limestone in the northern Holyoake Range (samples H84.20 and H84.26, Table 2). These taxa can be easily distinguished as Dailyatia sp. 1 lacks the densely set pseudoplicae of D. decobruta, and bears distinctive, strongly developed pseudoplicae formed from aligned crenulated projections. The sclerites of D. decobruta from the Shackleton Limestone bear similarity to specimens assigned to Dailyatia ajax from the glacial erratics from King George Island in West Antarctica (Wrona 2004). As mentioned by Betts et al. (2019) those from King George Island do not develop the densely arranged pseudoplicae characteristic of D. decobruta, although they share similar gross morphology.
Stratigraphic and geographic range.—Cambrian Series 2, Stage 4(?) of the Shackleton Limestone in northern Holyoake Range, Central Transantarctic Mountains, East Antarctica (Table 2); Cambrian Series 2, Stage 4 (upper Dailyatia odyssei Zone, Pararaia janeae Zone) of the Mernmerna Formation (NB section) in the eastern Flinders Ranges (Skovsted et al. 2015; Betts et al. 2017) and carbonate clasts recovered from the White Point Conglomerate of Kangaroo Island, South Australia (Betts et al. 2019).
Dailyatia cf. odyssei (Evans and Rowell, 1990)
Fig. 7G–I.
Material.—Two C sclerites of uncertain subtype, one of unknown type and a single fragment from the sample HRA/22 (NRM X10003–X10005, 3 figured) (Fig. 1, Table 1). Southern Holyoake Range, Cambrian Series 2.
Description.—Two C sclerites and one sclerite of uncertain type, triangular in transverse section with slight compression (Fig. 7H), slightly recurved at the apex, which overhangs the ventral side (Fig. 7H) and lacking torsion. Dorsal field convex, ventral field concave. Sclerite ornament is densely packed with concentric ribs which are lacking pustules (Fig. 7G2, I1). Micro-ornament not preserved. Dorsal side with single preserved plication in the central surface (Fig. 7G1, H1). In one specimen, the ventral side close to both the proximal and distal edges has two weakly developed plicae (Fig 7G).
Remarks.—The few sclerites of this species from the Shackleton Limestone of the Holyoake Range are too few in number and too poorly preserved to make a positive species-level assignment. They bear closest resemblance to Dailyatia odyssei C1 sclerites from the type locality in the Shackleton Limestone (Evans and Rowell 1990: figs. 7.1–7.8) and Series 2 carbonates of the Arrowie and Stansbury Basins, South Australia (Skovsted et al. 2015: figs. 44–50). Material from the type locality in the northern Holyoake Range (M84.2; Fig. 1) have the same densely set comarginal ribs (Fig. 7G2, I1) as those from the Southern Holyoake Range, and overall pyramidal shape and triangular transverse section (Evans and Rowell 1990: figs. 7.4, 7.5).
Weakly developed plicae are present at both the distal and proximal edges of the ventral side of D. cf. odyssei, a feature not present in D. odyssei sensu stricto, which has plicae developed regularly across the ventral surface in some specimens (Skovsted et al. 2015: fig. 48S, U). The gross morphology of D. cf. odyssei from the Shackleton Limestone and the C1 sclerites from South Australia are similar, with overall pyramidal shape, triangular transverse cross section, concave ventral fields and slightly recurved apex (Skovsted et al. 2015: fig. 48B–J, X–AA). Dailyatia cf. odyssei cannot be definitively assigned to D. odyssei as certain important characteristics are not present or have not been preserved. These include the reticulate micro-ornament and pustules on sclerites of D. odyssei (Skovsted et al. 2015: fig. 50). In addition, only a single radial plication is preserved on the dorsal side of D. cf. odyssei, whereas D. odyssei C1 sclerites typically have two. The central surface forms a concave furrow between the central radial plication and distal edge on the dorsal side of the C1 sclerites of D. odyssei (Skovsted et al. 2015: fig. 48) which is not present in D. cf. odyssei. In contrast, D. cf. odyssei from the Shackleton Limestone has a flat central region of the dorsal surface. The sclerites from the Shackleton Limestone are also dissimilar to C2 sclerites of D. odyssei from the Mernmerna Formation in the Flinders Ranges, South Australia, which typically have a much more recurved apex and are torted (Skovsted et al. 2015: fig. 49). The single dorsal radial plication in C2 sclerites of D. odyssei is separated from both the proximal and distal edges by broad concave fields.
The specimens can be easily distinguished from Dailyatia ajax which has tall, conical sclerites with abundant radial plications (Skovsted et al. 2015: figs. 16, 17). C sclerites of D. cf. odyssei are harder to distinguish from the C1 sclerites of Dailyatia macroptera, as they both have plicated distal and proximal edges on the ventral surface (Skovsted et al. 2015: fig. 24O, S) and dense concentric ribs (Skovsted et al. 2015: fig. 26). However, the C1 sclerites of D. macroptera are more dorso-ventrally compressed than in D. cf. odyssei and have a reduced, flat central surface on the dorsal side, separated by two plicae (Skovsted et al. 2015: fig. 24D, G–I, J, Y–X, AC). Dailyatia. cf. odyssei can also be distinguished from Dailyatia bacata as it lacks the diagnostic rounded pustules ornamenting the concentric ribs (Skovsted et al. 2015: fig. 34). Compared to Dailyatia helica, the C1 sclerites of D. cf. odyssei lack the distinctive plicate central surface of C sclerite subtypes. The C2a sclerites of D. helica are strongly coiled, unlike the slightly overhanging apex of D. cf. odyssei.
Dailyatia sp. 1
Figs. 8, 9A–G.
Material.—One A sclerite and numerous A sclerite fragments (NRM X10006–X10012, 7 figured), three C sclerites (NRM X10013–X10015), and numerous indeterminate fragments (NRM X10016–X10019, 4 figured) from sample HRA/14, southern Holyoake Range (Figs. 1, 2, Table 1). Fragmentary material also found in samples H84.20 and H84.26, northern Holyoake Range (Fig. 1, Table 2; Evans and Rowell 1990) and sample CM2/130, northern Churchill Mountains (Figs. 1, 2, Table 1). Cambrian Series 2, Stages 3, 4.
Description.—Two sclerite types identifiable (A and C). Pyramidal sclerites with distinctive irregular arrangement of widely spaced out plicae. Plicae have densely set comarginal ribs with reticulate ornamentation preserved on their crests (Fig. 9E1, E2, F).
A single, large A sclerite fragment partially preserving the anterior, lateral and posterior fields was recovered from the HRA section (Fig. 8A). This fragment is low and uncoiled with a blunt apex. Smaller fragments of the anterior and lateral fields of A sclerites are also identifiable (Fig. 8B–G). Antero-lateral plication is well-developed (Fig. 8A1–A4). Postero-lateral plication present, but truncated by a break in the sclerite close to the apex (Fig. 8A1). The anterior field and anteriormost part of the lateral field are dominated by large plicae, themselves coated by thick and well-developed comarginal ribs with broad, expanded crenulations which obscure the furrows (Fig. 8A1–A5). The plicae on the anterior field are chaotically spaced and developed, and bifurcate towards the apertural margin (Fig. 8A2, B, C). On the antero-lateral region, plicae are more regular, and have sets of small rows of pustules that develop from near the base of the furrows between the plicae and merge with the plicae towards the aperture (Fig. 8A1, A5, B2, G). The lateral trough is concave and smooth close to the apex and have dense comarginal ribs towards the aperture, without any ornamentation (Fig. 8A1, A3, A4, D).
Three C sclerites are preserved. It is not possible to identify subtypes due to the low numbers of sclerites. The sclerites are pyramidal and triangular or rectangular in transverse section lacking torsion (Fig. 9A1, B3, C1), with one sclerite slightly coiled towards the ventral side (Fig. 9A1, A3). The posterior field is concave with a variable number and distribution of plicae (Fig. 9A5, B2). One specimen has three regular plicae bordering or intersecting the proximal edge and ten plicae irregularly developed across the ventral field (Fig. 9A4, A5). Another has two plicae at the distal edge and a tangled arrangement of plicae adjacent to the proximal edge on the ventral field (Fig. 9B2). A single large plication in the centre of the dorsal surface separates two concave fields bordered by the proximal edge and distal edge. (Fig. 9A2, B1, C2). These concave fields generally lack plicae or pseudoplicae except for 1–2 weakly developed plicae close to the distal edge (Fig. 9A2, B1, B3).
The plicae are erratic in their shape and distribution, sometimes degenerating into a chaotic and tangled arrangement (Figs. 8A2, B, C, G, 9F). They are defined by densely set, rounded and broad crenulations (Figs. 8A5, A6, C3, E, F, G2, 9E2, F, G). Between well-developed plicae multiple, irregularly-spaced sets of pustules develop in the furrows forming pseudoplicae that grow towards the crest of each plica and merge with them aperturally (Figs. 8A1, G, 9E1, E2, G). In some fragments, plicae also bifurcate or have whole new plicae develop within the furrows of older growth sets (Figs. 8E–G, 9G). On the anterior field of an A sclerite fragment, the plicae are truncated by thickened comarginal bands which are raised across the furrows, with a new growth series continuing basally but laterally offset from the previous series (Fig. 8C). One fragment from an unknown sclerite type (Fig. 9E) also has three distinct types of ornament: First is a weakly developed field of comarginal ribs and minute pustules. This merges laterally with a second type of ornament which consists of a set of three plicae which are bounded by a furrow lacking any form of ornament preserved (Fig. 9E2). A third type is a dense growth series of pseudoplicae which are crossed by comarginal troughs which interrupt the pseudoplicae (Fig. 9E4). A reticulate micro-ornament is present on the wide, densely set crests of plicae and pseudoplicae (Figs. 8A6, C1, C3, 9F2). The furrows are typically smooth, lacking any micro-ornament (Fig. 9E1, E2, F).
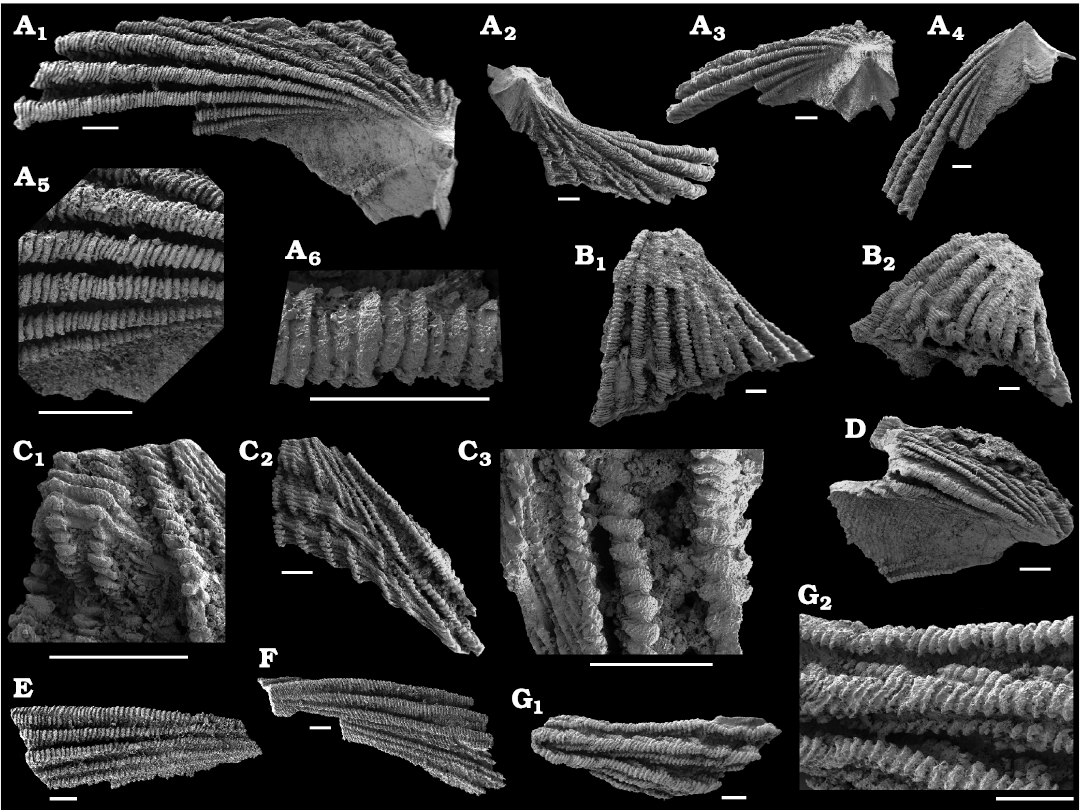
Fig. 8. The camenellan tommotiid Dailyatia sp. 1 A sclerites and fragments from the Cambrian Series 2, Stages 3, 4, Shackleton Limestone, Holyoake Range, Transantarctic Mountains, East Antarctica. A. A sclerite, NRM X10006, apical (A1), anterior (A2), oblique posterior (A3), and posterior (A4) views, magnification of radial plicae on the lateral field (A5), magnification of single plica showing the densely stacked growth series of the radial plicae (A6). B. Fragment of A sclerite anterior field, NRM X10007, general view (B1), oblique view showing cross section of plicae (B2). C. Fragment of A sclerite anterior field, NRM X10008, top of specimen (C1), general view (C2), detail showing plicae (C3). D. Fragment of A sclerite with partial lateral and anterior field, NRM X10009. E. Fragment of sclerite lateral field, NRM X10010. F. Fragment of A sclerite lateral field, NRM X10011. G. Fragment of unknown sclerite type, with chaotically distributed plicae, NRM X10012, general view (G1), detail (G2). Scale bars 200 µm.

Fig. 9. The camenellan tommotiid from the Cambrian Series 2, Stages 3, 4, Shackleton Limestone, Transantarctic Mountains, East Antarctica. Dailyatia sp. 1, C sclerites and indeterminate fragments, Holyoake Range and Churchill Mountains (A–G) and Dailyatia braddocki Evans and Rowell, 1990, Churchill Mountains (H, I). A. Dextral C sclerite, NRM X10013, apical view (A1), dorsal field (A2), proximal edge (A4), distal edge (A3), ventral field (A5), detail showing broken apex (A6). B. Dextral C sclerite, NRM X10014, oblique view of distal edge and dorsal field (B1), oblique view of proximal edge and ventral field (B2), and apical view (B3). C. Sinistral C sclerite, NRM X10015, apical view (C1), oblique view of proximal edge and dorsal field (C2). D. Fragment of unknown sclerite type, NRM X10016. Showing fragment surface (D1) and oblique view (D2) showing plicae cross-section. E. Fragment of unknown sclerite type, NRM X10017, various views of multiple ornament types (E1–E4). F. Fragment of unknown sclerite type, NRM X10018, general view (F1), detail showing pseudoplicae emerging from furrows between the radial plicae (F2). G. Fragment of unknown sclerite type, NRM X10019, general view (G1), detail showing plicae bifurcation (G2). H. Fragment of unknown sclerite type, KUMIP 585075, general view (H1), detail of surface showing raised pustules forming the pseudoplicae (H2). I. Dextral C sclerite with band of disturbed growth, KUMIP 585076, view of ventral field (I1), apical view (I2). Scale bars 200 µm, except F2, 100 µm; H2, 50 µm.
Remarks.—The external ornament of Dailyatia sp. 1 is most comparable to the densely plicate Dailyatia ajax. The plicae of Dailyatia sp. 1 are more densely set where present (Fig. 8A5, G2), compared to the more widely distributed but slightly less densely set plicae in D. ajax (Skovsted et al. 2015: figs. 11–17). Overall, the morphology of Dailyatia sp. 1 sclerites are easily distinguished from D. ajax which has distinctly tall and conical sclerites, whereas the sclerites of Dailyatia sp. 1 are low. The A sclerite of Dailyatia sp. 1 has dense and parallel radial plications similar to the A1 sclerites of Dailyatia macroptera (Skovsted et al. 2015: fig. 21), especially the plicate variety (Skovsted et al. 2015: fig. 27A, B). The low, pyramidal sclerites of Dailyatia bacata are also similar in gross form to Dailyatia sp. 1. However, the development of radial plicae on Dailyatia sp. 1 distinguishes it from D. bacata, which lacks the dense plicae on the ventral side of the C sclerites and the irregular, dense plicae on the anterior side of the A sclerite seen in Dailyatia sp. 1. The extreme coiling of Dailyatia helica sclerites is also not observed in any Dailyatia sp. 1 sclerites.
The C sclerites of Dailyatia sp. 1 also have some similarities to C1 sclerites of Dailyatia odyssei in overall pyramidal shape. However, the dorsal radial plications on the C1 sclerites and ventral radial plications on the C2 sclerites of Dailyatia sp. 1 are typically absent from the C1 and C2 sclerites of D. odyssei. Only in larger C1 (Skovsted et al. 2015: fig. 48S, U) and C2 sclerites (Skovsted et al. 2015: fig. 49C) of D. odyssei do plicae form on the ventral field.
The ornamentation of Dailyatia sp. 1 is distinctive and indicate a consistent novel development of plicae. The emergence of new plicae from furrows is known from Dailyatia odyssei, but is generally uncommon (e.g., Skovsted et al. 2015: fig. 50H). The most common example of such irregularity in the development of Dailyatia sclerites occurs in damaged or gerontic specimens. Interruption of the development of growth series is marked by comarginal troughs and is known from most other species, including Dailyatia ajax (Skovsted et al. 2015: figs. 71, 73G, H), Dailyatia macroptera (Skovsted et al. 2015: figs. 72C–F, 73A–C, 74A–E) Dailyatia bacata (Skovsted et al. 2015: fig. 73D–F), Dailyatia helica (Skovsted et al. 2015: fig. 73I, J), Dailyatia odyssei (Skovsted et al. 2015: figs. 50G, 74F, G) and Dailyatia braddocki from the Shackleton Limestone (Fig. 9I). Dailyatia sp. 1 develops similar interruptions in plicae development, with plicae laterally offset from the previous growth series (Fig. 8A2, B, C), a growth pattern is also seen in damaged or gerontic specimens of Dailyatia (Skovsted et al. 2015: figs. 71–73). However, growth interruptions in Dailyatia sp. 1 are unlikely to be gerontic features as the interruptions appear in the earliest stages of the anterior field of A sclerites, differentiating it from typical gerontic growth disturbance in Dailyatia that typically start closer to the aperture (e.g., Skovsted et al. 2015: fig. 73). An example of gerontic disturbance of a Dailyatia braddocki C sclerite from the Shackleton Limestone, Churchill Mountains (sample GM87.1, Fig. 1) is shown in Fig. 9I.
The irregular arrangement of radial plications on the anterior field of Dailyatia sp. 1 is also similar to a large, potentially gerontic D. macroptera from the Mernmerna Formation (Skovsted et al. 2015: figs. 21F–H, 71F, G). Strongly plicate sclerite varieties of Dailyatia macroptera (Skovsted et al. 2015: fig. 27) and Dailyatia helica (Skovsted et al. 2015: fig. 43) are uncommon but do occur in South Australia. Hence, while Dailyatia sp. 1 can be distinguished from D. macroptera and D. odyssei based on gross morphology, there is a possibility that Dailyatia sp. 1 is a “plicate variant” of these or another Dailyatia species.
Kennardiidae indet.
Fig. 10.
1990 Kennardia sp. A; Evans and Rowell 1990: 699, fig. 7.9.
1990 Kennardia sp. B; Evans and Rowell 1990: 699, figs. 7.10–7.12.
Material.—One symmetrical first sclerite type (KUMIP 585077, 1 figured), one symmetrical second sclerite type (KUMIP 585082, 1 figured) and seven asymmetrical third sclerite types (KUMIP 233002, 585078–585081, 585083, 585084, 6 figured) from M section, northern Holyoake Range (Fig. 1, Table 2; Evans and Rowell 1990). Numerous fragments were also recovered from this section (Table 2). Cambrian Series 2, Stages 3–4.
Description.—Three identifiable sclerite types. First sclerite type symmetrical, low, curved over the ventral field and quadrangular in transverse section (Fig. 10A). Second sclerite type coniform, symmetrical, tall, straight and hexagonal in transverse section (Fig. 10G). Third sclerite type coniform, asymmetrical, tall, coiled over the ventral field, and triangular in transverse section (Fig. 10B–F, H, I). Sclerites with a distinctive growth set of wide, wall-like comarginal ribs and inter-rib grooves.
The first sclerite type symmetrical with dorsal field lacking plicae and with no clear distinction from the lateral fields (Fig. 10A1) which have a single, weakly developed plica bound by furrows (Fig. 10A3). Ventral field is flat and straight, lacking plicae (Fig. 10A2). The sclerite is coiled only at the apex where it overhangs the ventral field (Fig. 10A2). Growth series forming wall-like comarginal ribs, well-developed and present on entire sclerite surface, following the transverse profile of the sclerite.
The second sclerite type known from single, poorly preserved sclerite. The anterior field is gently curved laterally with two weakly developed plicae (Fig. 10G1). Lateral fields triangular in shape, flat and straight, lacking plicae (Fig. 10G1, G3). Posterior field side triangular and straight, lacking plicae (Fig. 10G2). Growth sets developed on the entire sclerite surface and follow the transverse profile of the sclerite on the dorsal side and lateral fields. The growth sets on the ventral side curve upwards into broad inverted U-shape (Fig. 10G2).
The third sclerite type is tall, triangular, asymmetrical with a concave ventral surface and convex dorsal surface in smaller specimens, but curves back towards the dorsal side in larger specimens (Fig. 10E1, H1). The ventral side has a series (3–7) of well-developed radial plicae with the outmost bordering the proximal and distal edges (Fig. 10C2, D1, E3, E4, F1, H1). The proximal edge is shorter than the distal edge. The dorsal side has a single, large and broad plication in larger specimens (Fig. 10C1, D2, H2, H3). The fields bounded by the central plication and the proximal edge is flat or slightly concave (Fig. 10D2, H1). The field bounded by the central plication and distal edge is convex (Fig. 10C1, H3). The dorsal side lacks pseudoplicae on all but the largest specimen, where they are formed by lateral irregularly spaced series of weakly developed pustules (Fig. 10E1, E2). In one small specimen, the dorsal side lacks any plication and forms a single evenly convex field (Fig. 10B).

Fig. 10. The tommotiid Kennardiidae indet. from the Cambrian Series 2, Stages 3, 4, Shackleton Limestone, Holyoake Range, Transantarctic Mountains, East Antarctica. A. Symmetrical first sclerite type, KUMIP 585077, apical view (A1), oblique view of posterior field (A2), lateral field (A3). B. Third sclerite type, KUMIP 585078, apical (B1) and lateral (B2) views. C. Third sclerite type, KUMIP 585079, apical view (C1), ventral side (C2), dorsal side (C3). D. Third sclerite type, KUMIP 585080, ventral side (D1), apical view (D2). E. Third sclerite type, KUMIP 233002, apical view (E1), detail of dorsal side (E2), detail of ventral side ( E3), ventral side (E4). F. Third type of sclerite, KUMIP 585081, ventral side (F1), detail showing plicae on ventral surface (F2), distal edge (F3). G. Second type of sclerite, KUMIP 585082, apical view (G1), oblique view of posterior field (G2), lateral field (G3). H. Third type of sclerite, KUMIP 585083, posterior side (H1), distal edge (H2), apical view (H3). I. Third type of sclerite, KUMIP 585084, ventral side (I1), detail showing plicae on the ventral surface (I2). Scale bars 200 µm, except E2, E3, I2, 100 µm.
Remarks.—Specimens of Kennardiidae indet. from the Shackleton Limestone of the Holyoake Range have a mosaic of kennardiid features. They are united by wall-like comarginal ribs on their growth sets similar to those found in Kennardia (Laurie 1986: fig. 7). They also fit the diagnostic criteria for Kennardiidae in having three distinct sclerite types with ubiquitous growth sets, but potentially have two symmetrical forms instead of one. No reticulate micro-ornamentation is present on the sclerites, but this is similar to other specimens recovered from the same samples (e.g., Dailyatia odyssei from samples M84.1, M84.2, recovered by Evans and Rowell 1990, see Fig. 1), and hence may be preservational.
Evans and Rowell (1990: 699, figs. 7.9–7.12) described specimens of Kennardiidae indet. from the same localities and referred them to two species in open nomenclature: “Kennardia sp. A” and “Kennardia sp. B” on the basis of the coarse concentric ribs similar to those in Kennardia. “Kennardia sp. B” was considered by Skovsted et al. (2015: 22, 31) to potentially represent Dailyatia ajax C1 sclerites, as they share a tall, conical shape, triangular transverse section with well-developed radial plicae and comarginal ribs. Evans and Rowell’s (1990) specimens were split into two species based on “Kennardia sp. B” being much larger and with pustules forming pseudoplicae. These criteria are not sufficient to delineate these species based on the limited number of sclerites originally described. Size is unsuitable in this case as it can be a product of different ontogenetic stages and ornamentation of kennardiid sclerites is variable within and across sclerite types.
The symmetrical first sclerite form of Kennardiidae indet. is similar to the symmetrical Form A of the monospecific genus Kennardia Laurie, 1986, which was described from the Todd River Dolostone of the Amadeus Basin, Northern Territory, Australia (Laurie 1986: fig. 8A). They can be clearly delineated by the lack of reticulate micro-ornament on Kennardidae indet., which is a ubiquitous character of Kennardia, along with the more strongly developed comarginal ribs (Laurie 1986: fig. 8F–H). There may be a difference in transverse section, with the single symmetrical Kennardiidae sclerite more triangular at the apex and subcircular towards the base, and Kennardia Form A sclerites described as subrectangular (Laurie 1986: 439), although this is not figured clearly by Laurie (1986) and based on a single sclerite of Kennardiidae indet. An A sclerite of Kennardia sp. from the Mt Scott Range of South Australia (Skovsted et al. 2015: fig. 8) is also similar to the first sclerite form of Kennardiidae indet. The dorsal side of this sclerite is also gently curved laterally with no clear delineation from the lateral fields and the apex overhangs the posterior field (Skovsted et al. 2015: fig. 8A, B). The posterior field is concave in Kennardia sp., whereas it is flat in Kennardiidae indet.
The symmetrical second sclerite type is similar to the symmetrical sclerite of Tommotiid gen. et sp. nov. from the Koolywurtie Limestone Member of the Parara Limestone of the Stansbury Basin, South Australia (Paterson et al. 2007: fig. 4A–C). This sclerite is also tall and conical and has the same hexagonal transverse section. However, the symmetrical sclerite of Tommotiid gen. et sp. nov. is slightly curved over the ventral field, unlike Kennardiidae indet. and the comarginal ribs curve towards the apex on the lateral fields of Tommotiid gen. et sp. nov., whereas they are straight in Kennardiidae indet.
Radial plicae have been considered as a diagnostic character of Dailyatia (Skovsted et al. 2015: 18) and are also present on the ventral surface of the third, symmetrical sclerite type of Kennardiidae indet. These sclerites of Kennardiidae indet. are similar to C sclerites of some forms of Dailyatia, for example the C1 sclerites of Dailyatia ajax. The C1 sclerites of D. ajax in particular have a similar tall and pyramidal form to the third sclerite type of Kennardiidae indet. (Fig. 10C1, C2, F3, H, I1; Skovsted et al. 2015: fig. 16L–Q). However, the dorsal side of the C1 sclerite of D. ajax has plicae (Skovsted et al. 2015: fig. 16L–Q), a feature which is only present on the ventral side of Kennardiidae indet. (Fig. 10C2, E4, F1, H1). Plicae are also present on the dorsal side of C2 sclerites of D. ajax (Skovsted et al. 2015: figs. 10N, 17Q).
Kennardiidae indet. can be excluded from both Kennardia and Dailyatia as it has two symmetrical sclerite types and single asymmetrical type, in contrast to a single symmetrical sclerite type and two asymmetrical types in Kennardia and Dailyatia. They can also be distinguished from Kennardia as Kennardiidae indet. has radial plicae, a feature which is diagnostic of Dailyatia. The wall-like ribs present on Kennardiidae indet. are similar to the wall-like ribs of Canadiella (Skovsted et al. 2020: figs. 5, 6), but lacks the pustulose ornamentation of this genus (Skovsted et al. 2020: figs. 5M, 6M, N)
Family uncertain
Shetlandia Wrona, 2004
Type species: Shetlandia multiplicata Wrona, 2004; Lower Cambrian clasts from the Miocene Cape Melville Formation, King George Island, South Shetland Islands, Antarctica.
Shetlandia multiplicata Wrona, 2004
Fig. 11.
2004 Shetlandia multiplicata sp. nov.; Wrona 2004: 34, figs. 14, 15A–E, H–J, 16.
Material.—Exclusively fragmentary material, with 22 fragments recovered from samples HRA/24, HRA/25, and HRA/14 (NRM X10020–X10024, 5 figured), southern Holyoake Range (Figs. 1, 2, Table 1). Cambrian Series 2, Stages 3, 4.
Description.—Fragments preserve flat to gently curved sclerite fields; no large plications or apexes are preserved. No micro-ornament is preserved. Two distinct arrangements of plicae occur on the fragments. Firstly, rows of closely stacked, overlapping arches delineate tall, prominent radial plicae which extend from the top of the plicae to the groove (Fig. 11A). The arches increase in size towards the base of the sclerite. These plicae are separated by grooves with concentrically oriented ribs at their base. These ribs do not contact the base of the arches (Fig. 11A2) but are part of the same growth series. The second type of ornament (found on only one sclerite fragment) is weakly developed pseudoplicae formed by a series of pustules in concentric rows (Fig. 11D). A markedly different type of ornament also occurs and consists not of plicae but of densely packed concentric, wall-like ribs which are also present on one fragment (Fig. 11B1, C).

Fig. 11. The tommotiid Shetlandia multiplicata Wrona, 2004, unknown type sclerite fragments from the Cambrian Series 2, Stages 3, 4, Shackleton Limestone, Holyoake Range, Transantarctic Mountains, East Antarctica. A. NRM X10020, general view (A1), detail showing arched growth series (A2). B. NRM X10021, general view (B1), detail of broken margin (B2). C. NRM X10022, general view (C1), detail of comarginal ribs and broken edge (C2). D. NRM X10023. E. NRM X10024. Scale bars 200 µm, except B2, 100 µm. A–C taken with backscattered electron detector.
Remarks.—Shetlandia multiplicata was previously known only from glacial erratics recovered from the glaciomarine Miocene Cape Melville Formation, King George Island, South Shetland Islands of West Antarctica and is generally poorly understood (Wrona 1989, 2004). However, despite the fragmentary nature of the holotype, it has unique ornamentation and plication that is unknown from any other tommotiid genus.
The few fragments from the Shackleton Limestone in the Holyoake Range can be confidently assigned to Shetlandia multiplicata Wrona, 2004, due the unique morphology of the dense plicae. The densely packed arches that make up these plicae are identical in morphology to those originally described from King George Island (Wrona 2004: fig. 14M–T). Pustules arranged into pseudoplicae are also found on some sclerite fragments from King George Island (Wrona 2004: fig. 15H, I). The third ornament of concentric wall-like ribs is also found on both sets of fragmentary material.
Stratigraphic and geographic range.—Cambrian Series 2, Stages 3, 4, of the Shackleton Limestone in southern Holyoake Range, Central Transantarctic Mountains, East Antarctica; carbonate clasts (Cambrian Series 2, Stage 4?) from the Miocene Cape Melville Formation, King George Island, West Antarctica (Wrona 2004).
Conclusions
Seven tommotiid taxa from Antarctica are described. From the Argentina Range, the apparently endemic Dailyatia icari sp. nov. is the first tommotiid described from this region of East Antarctica. From new sections through the Shackleton Limestone in the Holyoake Range, Dailyatia cf. D. odyssei, Dailyatia sp. 1, and Shetlandia multiplicata are described. A new section in the Churchill Mountains yielded sclerites of Dailyatia braddocki. From previously collected material in the Holyoake Range, Dailyatia decobruta and Kennardiidae indet. are described.
The tommotiid fauna of East Antarctica provides new data for biostratigraphic correlation and relative dating of the Shackleton Limestone. Dailyatia decobruta, occurring alongside Dailyatia odyssei in the northern Holyoake Range provides a new platform for age designation. These co-occurring species correlate this part of the Shackleton Limestone to the Cambrian Series 2, Stage 4 (South Australian upper Dailyatia odyssei Zone and Pararaia janeae Zone) White Point Conglomerate (Betts et al. 2019) and upper Mernmerna Formation (Betts et al. 2017) in South Australia.
The material described tells a more complex story of endemism in Cambrian Series 2 tommotiids. Discovery of D. decobruta from the Holyoake Range is only the second species of Dailyatia found in autochthonous rocks of East Antarctica and South Australia, after D. odyssei. The two remaining named species: Dailyatia braddocki and Dailyatia icari sp. nov. are known only from their type localities in the Churchill Mountains and Argentina Range, respectively. Unnamed species Dailyatia sp. 1 and tommotiids Shetlandia multiplicata and Kennardiidae indet. from autochthonous rocks are known only from East Antarctica.
Acknowledgements
Paul M. Myrow (Colorado College, Colorado Springs, USA), G. Dylan Taylor (Chamonix, France), and Lars Stemmerik (Geological Survey of Denmark and Greenland, Copenhagen, Denmark) were integral parts of the fieldwork team in Antarctica. The manuscript was improved by reviews from Bing Pan (Chinese Academy of Sciences, Nanjing, China), Léa Devaere (Museum für Naturkunde, Berlin, Germany), and Duncan J.E. Murdock (University of Oxford, UK). Processing of rock samples was performed by David Mathieson (Macquarie University, Sydney, Australia) and Therese Hultgren (Uppsala University, Sweden). Staff at the Macquarie University Facility of Science and Engineering Microscope Facility (MQFoSE MF) are thanked for their assistance with scanning electron microscopy. We also thank Albert J. Rowell (formerly University of Kansas, Lawrence, USA) who collected the material and Bruce S. Lieberman (University of Kansas, Lawrence, USA) for arranging its study. Una C. Farrell, Julien Kimmig, and Natalia López-Carranza (all University of Kansas, Lawrence, USA) are thanked for facilitating access to the material and processing the loan. The work by Lars E. Holmer was supported by grants from the Swedish Research Council (VR 2009-4395, 2012-1658). The field work in Antarctica was supported by operational support for polar research (RFI Polar VR 2010-6176) and the Swedish Polar Secretariat in collaboration with the United States Antarctic Program and US National Science Foundation (NSF). Glenn A. Brock was supported in Antarctica by the Trans-Antarctic Association.
References
Atkins, C.J. and Peel, J.S. 2004. New species of Yochelcionella (Mollusca; Helcionelloida) from the Lower Cambrian of North Greenland. Bulletin of the Geological Society of Denmark 51: 1–9. Crossref
Balthasar, U., Skovsted, C.B., Holmer, L.E., and Brock, G.A. 2009. Homologous skeletal secretion in tommotiids and brachiopods. Geology 37: 1143–1146. Crossref
Bassett-Butt, L. 2016. Systematics, biostratigraphy and biogeography of brachiopods and other fossils from the middle Cambrian Nelson Limestone, Antarctica. GFF 138: 377–392. Crossref
Bengtson, S. 2004. Early Skeletal Fossils. In: J.H. Lipps and B.M. Waggoner (eds.), Neoproterozoic–Cambrian Biological Revolutions. The Paleontological Society Papers 10: 67–77. Crossref
Bengtson, S., Conway Morris, S., Cooper, B.J., Jell, P.A., and Runnegar, B.N. 1990. Early Cambrian fossils from South Australia. Memoirs of the Association of Australasian Palaeontologists 9: 1–364.
Betts, M.J., Claybourn, T.M., Brock, G.A., Jago, J.B., Skovsted, C.B. and Paterson, J.R. 2019. Shelly fossils from the lower White Point Conglomerate, Kangaroo Island, South Australia. Acta Palaeontologica Polonica 64: 489–522. Crossref
Betts, M.J., Paterson, J.R., Jacquet, S.M., Andrew, A.S., Hall, P.A., Jago, J.B., Jagodzinski, E.A., Preiss, W.V., Crowley, J.L., Brougham, T. and Mathewson, C.P., 2018. Early Cambrian chronostratigraphy and geochronology of South Australia. Earth-Science Reviews 185: 498–543. Crossref
Betts, M.J., Paterson, J.R., Jago, J.B., Jacquet, S.M., Skovsted, C.B., Topper, T.P., and Brock, G.A. 2016. A new lower Cambrian shelly fossil biostratigraphy for South Australia. Gondwana Research 36: 176–208. Crossref
Betts, M.J., Paterson, J.R., Jago, J.B., Jacquet, S.M., Skovsted, C.B., Topper, T.P. and Brock, G.A. 2017. Global correlation of the early Cambrian of South Australia: Shelly fauna of the Dailyatia odyssei Zone. Gondwana Research 46: 240–279. Crossref
Bischoff, G.C.O. 1976. Dailyatia, a new genus of the Tommotiidae from Cambrian strata of SE. Australia (Crustacea, Cirripedia). Senckenbergiana Lethaea 57: 1–33.
Boger, S.D. and Miller, J.M.L. 2004. Terminal suturing of Gondwana and the onset of the Ross-Delamerian Orogeny: The cause and effect of an Early Cambrian reconfiguration of plate motions. Earth and Planetary Science Letters 219: 35–48. Crossref
Claybourn, T.M., Jacquet, S.M., Skovsted, C.B., Topper, T.P., Holmer, L.E., and Brock, G.A. 2019. Mollusks from the upper Shackleton Limestone (Cambrian Series 2), Central Transantarctic Mountains, East Antarctica. Journal of Palaeontology 93: 437–459. Crossref
Claybourn, T.M., Skovsted, C.B., Holmer, L.E., Pan, B., Myrow, P.M., Topper, T.P., and Brock, G.A. 2020. Brachiopods from the Byrd Group (Cambrian Series 2, Stage 4) Central Transantarctic Mountains, East Antarctica: biostratigraphy, phylogeny and systematics. Papers in Palaeontology 6: 349–383. Crossref
Debrenne, F. and Kruse, P.D. 1986. Shackleton limestone archaeocyaths. Alcheringa 10: 235–278. Crossref
Debrenne, F. and Kruse, P.D. 1989. Cambrian Antarctic archaeocyaths. Geological Society, London, Special Publications 47: 15–28. Crossref
Devaere, L., Clausen, S., Monceret, E., Vizcaïno, D., Vachard, D., and Genge, M.C. 2014. The tommotiid Kelenella and associated fauna from the early Cambrian of Southern Montaigne Noire (France): Implications for camenellan phylogeny. Palaeontology 57: 979–1002. Crossref
Evans, K.R. 1992. Antarctic specimens of an enigmatic Cambrian animal. Journal of Palaeontology 66: 558–562. Crossref
Evans, K.R. and Rowell, A.J. 1990. Small shelly fossils from Antarctica: An early Cambrian faunal connection with Australia. Journal of Paleontology 64: 692–700. Crossref
Foden, J., Elburg, M.A., Dougherty-Page, J., and Burtt, A. 2006. The timing and duration of the Delamerian Orogeny: Correlation with the Ross Orogen and Implications for Gondwana assembly. The Journal of Geology 114: 189–210. Crossref
Goodge, J.W., Walker, N.W., and Hansen, V.L. 1993. Neoproterozoic–Cambrian basement-involved orogenesis within the Antarctic margin of Gondwana. Geology 21: 37–40.Crossref
Goodge, J.W., Williams, I.S., and Myrow, P.M. 2004. Provenance of Neoproterozoic and lower Paleozoic siliciclastic rocks of the central Ross orogen, Antarctica: Detrital record of rift-, passive-, and active-margin sedimentation. Bulletin of the Geological Society of America 116: 1253–1279. Crossref
Gravestock, D.I., Alexander, E.M., Demidenko, Y.E., Esakova, N.B., Holmer, L.E., Jago, J.B., Lin, T.-R., Melnikova, N., Parkhaev, P.Y., Rozanov, A.Y., Ushatinskaya, G.T., Sang, W.-L., Zhegallo, E.A., and Zhuravlev, A.Y. 2001. The Cambrian biostratigraphy of the Stansbury Basin, South Australia. Transactions of the Palaeontological Institute of the Russian Academy of Sciences 282: 1–341.
Hally, L.A. and Paterson, J.R. 2014. Biodiversity, biofacies and biogeography of middle Cambrian (Series 3) arthropods (Trilobita and Agnostida) on the East Gondwana margin. Gondwana Research 26: 654–674. Crossref
Holmer, L.E, Popov, L., and Wrona, R. 1996. Early Cambrian lingulate brachiopods from glacial erratics of King George Island (South Shetland Islands), In: A. Gaździcki (ed.), Palaeontological Results of the Polish Antarctic Expeditions, Part 2. Palaeontologia Polonica 55: 37–50.
Holmer, L.E., Skovsted, C.B., Brock, G.A., Valentine, J.L., and Paterson, J.R. 2008. The early Cambrian tommotiid Micrina, a sessile bivalved stem group brachiopod. Biology Letters 5: 724–728. Crossref
Holmer, L.E., Skovsted, C.B., Larsson, C., Brock, L.E., and Zhang, Z. 2011. First record of a bivalved larval shell in the early Cambrian tommotiids and its phylogenetic significance. Palaeontology 54: 235–239. Crossref
Hyman, L.H. 1959. The lophophorate coelomates. Phylum Brachiopoda. In: L.H. Hyman (ed.). The Invertebrates: Smaller Coelomate Groups: Chaetognatha, Hemichordata, Pognophora, Phoronida, Ectoprocta, Brachipoda, Sipunculida: the Coelomate Bilateria, Vol. 5, 516–609. McGraw-Hill, New York.
Jeppsson, L., Anehus, R., and Fredholm, D. 1999. The optimal acetate buffered acetic acid technique for extracting phosphatic fossils. Journal of Paleontology 73: 964–972. Crossref
Kruse, P.D. 1990. Cambrian Palaeontology of the Daly Basin. Northern Territory Geological Survey, Reports 7: 58 pp.
Landing, E.D. and Bartowski, K.E. 1996. Oldest shelly fossils from the Taconic allochthon and late Early Cambrian sea-levels in eastern Laurentia. Journal of Paleontology 70: 741–761. Crossref
Landing, E.D., Geyer, G., and Bartowski, K.E. 2002. Latest early Cambrian small shelly fossils, trilobites, and Hatch Hill dysaerobic interval on the Quebec continental slope. Journal of Paleontology 76: 287–305. Crossref
Laurie, J.R. 1986. Phosphatic fauna of the Early Cambrian Todd River Dolomite, Amadeus Basin, central Australia. Alcheringa: An Australasian Journal of Palaeontology 10: 431–454. Crossref
Laurie, J.R. 2006. Early Middle Cambrian trilobites from Pacific Oil & Gas Baldwin 1 well, southern Georgina Basin, Northern Territory. Memoirs of the Association of Australasian Palaeontologists 32: 127–204.
Laurie, J.R. 2012. Biostratigraphy of the Arthur Creek Formation and Thorntonia Limestone, Georgina Basin. In: Central Australian Basins Symposium III, Alice Springs, NT, 16–17 July 2012, 1–6.
Larsson, C.M., Skovsted, C.B., Brock, G.A., Balthasar, U., Topper, T.P., and Holmer, G.A. 2014. Paterimitra pyramidalis from South Australia: Scleritome, shell structure and evolution of a lower Cambrian stem group brachiopod. Palaeontology 57: 417–446. Crossref
Li, G. and Xiao, S. 2004. Tannuolina and Micrina (Tannuolinidae) from the lower Cambrian of East Yunnan, South China and their scleritome reconstruction. Journal of Palaeontology 78: 900–913. Crossref
Liebermann, B.S. 2004. Revised biostratigraphy, systematics, and paleobiology of the trilobites from the middle Cambrian Nelson Limestone, Antarctica. The University of Kansas Paleontological Contributions 14: 1–23.
Missarzhevsky, V.V. [Missarževskij, V.V.] 1970. New generic name Tommotia nom. nov. [in Russian] Paleontologičeskij žurnal 1970 (2): 100.
Murdock, D.J.E., Bengtson, S., Marone, F., Greenwood, J.M., and Donoghue, P.C.J. 2014. Evaluating scenarios for the evolutionary assembly of the brachiopod body plan. Evolution and Development 16: 13–24. Crossref
Myrow, P.M., Pope, M.C., Goodge, J.W., Fischer, W., and Palmer, A.R. 2002. Depositional history of pre-Devonian strata and timing of Ross orogenic tectonism in the central Transantarctic Mountains, Antarctica. Geological Society of America Bulletin 114: 1070–1088. Crossref
Palmer, A. and Gatehouse, C. 1972. Early and middle Cambrian trilobites from Antarctica. U.S. Geological Survey Professional Paper 456-D: 1–36. Crossref
Palmer, A.R. and Rowell, A.J. 1995. Early Cambrian trilobites from the Shackleton Limestone of the central Transantarctic Mountains. Memoir of the Paleontological Society 48: 1–28. Crossref
Pan, B., Skovsted, C.B., Brock, G.A., Topper, T.P., Holmer, L.E., Li, L., and Li, G. 2019. Early Cambrian organophosphatic brachiopods from the Xinji Formation, Shuiyu section, North China. Palaeoworld [published online, http: doi.org/10.1016/j.palwor.2019.07.001] Crossref
Paterson, J.R., Skovsted, C.B., Brock, G.A., and Jago, J.B. 2007. An early Cambrian faunule from the Koolywurtie Limestone Member (Parara Limestone), Yorke Peninsula, South Australia and its biostratigraphic significance. Memoirs of the Association of Australasian Palaeontologists 34: 131–146.
Paulsen, T.S., Encarnación, J., Grunow, A.M., Layer, P.W., and Watkeys, M. 2007. New age constraints for a short pulse in Ross orogen deformation triggered by East-West Gondwana suturing. Gondwana Research 12: 417–427. Crossref
Peel, J.S. and Skovsted, C.B. 2005. Problematic cap-shaped fossils from the Lower Cambrian of North-East Greenland. Paläontologische Zeitschrift 79: 461–470. Crossref
Rees, M.N., Pratt, B.R., and Rowell, A.J. 1989. Early Cambrian reefs, reef complexes and associated lithofacies of the Shackleton Limesonte, Transantarctic Mountains. Sedimentology 36: 341–361. Crossref
Rode, A.L., Liebermann, B.S., and Rowell, A.J. 2003. A new early Cambrian bradoriid (Arthropoda) from East Antarctica. Journal of Palaeontology 77: 691–697. Crossref
Rowell, A.J., Evans, K.R., and Rees, M.N. 1988. Fauna of the Shackleton Limestone. Antarctic Journal of the United States 23: 13–14.
Rowell, A.J., Rees, M.N., and Evans, K.R. 1992a. Depositional setting of the Lower and Middle Cambrian in the Pensacola Mountains. Antarctic Journal of the United States 25: 40–42.
Rowell, A.J., Rees, M.N., and Evans, K.R. 1992b. Evidence of major Middle Cambrian deformation in the Ross orogen, Antarctica. Geology 20: 31–34. Crossref
Rowell, A.J., van Schmus, W.R., Storey, B.C., Fetter, A.H., and Evans, K.R. 2001. Latest Neoproterozoic to mid-Cambrian age for the main deformation phases of the Transantarctic Mountains: new stratigraphic and isotope constraints from the Pensacola Mountains, Antarctica. Journal of the Geological Society, London 158: 295–308. Crossref
Skovsted, C.B. 2004. Mollusc fauna of the Early Cambrian Bastion Formation of North-East Greenland. Bulletin of the Geological Society of Denmark 51: 11–37. Crossref
Skovsted, C.B., Balthasar, U., Brock, G.A., and Paterson, J.R. 2009a. The tommotiid Camenella reticulosa from the early Cambrian of South Australia: Morphology, scleritome reconstruction, and phylogeny. Acta Palaeontologica Polonica 54: 525–540. Crossref
Skovsted, C.B., Balthasar, U., Vinther, J., and Sperling, E.A. 2020. Small shelly fossils and carbon isotopes from the early Cambrian (Stage 3–4) Mural Formation of western Laurentia. Papers in Palaeontology [published online https://doi.org/10.1002/spp2.1313]. Crossref
Skovsted, C.B., Betts, M.J., Topper, T.P., and Brock, G.A. 2015. The early Cambrian tommotiid genus Dailyatia from South Australia. Memoirs of the Association of Australasian Palaeontologists 48: 1–117.
Skovsted, C.B., Brock, G.A., Paterson, J.R., Holmer, L.E., and Budd, G.E. 2008. The scleritome of Eccentrotheca from the lower Cambrian of South Australia: Lophophorate affinities and implications for tommotiid phylogeny. Geology 36: 171–174. Crossref
Skovsted, C.B., Brock, G.A., Topper, T.P., Paterson, J.R., and Holmer, L.E. 2011. Scleritome construction, biofacies, biostratigraphy and systematics of the Tommotiid Eccentrotheca helenia sp. nov. from the early Cambrian of South Australia. Palaeontology 54: 253–286. Crossref
Skovsted, C.B., Clausen, S., Álvaro, J.J., and Ponlevé, D. 2014. Tommotiids from the early Cambrian (Series 2, Stage 3) of Morocco and the evolution of the tannuolinid scleritome and setigerous shell structures in stem group brachiopods. Palaeontology 57: 171–192. Crossref
Skovsted, C.B., Holmer, L.E., Larsson, C.M., Högström, A.E.S., Brock, G.A., Topper, T.P., Balthasar, U., Petterson Stolk, S., and Paterson, J.R. 2009b. The scleritome of Paterimitra: an early Cambrian stem group brachiopod from South Australia. Proceedings of the Royal Society B 276: 1651–1656. Crossref
Smith, T.E, Kelman, A.P., Nicoll, R., Edwards, D., Hall, L., Laurie, J.R., and Carr, L.K. 2013. An updated stratigraphic framework for the Georgina Basin, NT and Queensland. The APPEA Journal 53: 487–487. Crossref
Squire, R.J. and Wilson, C.J.L. 2005. Interaction between collisional orogenesis and convergent-margin processes: evolution of the Cambrian proto-Pacific margin of East Gondwana. Journal of the Geological Society 162: 749–761. Crossref
Sundberg, F.A., Geyer, G., Kruse, P.D., McCollum, L.B., Pegel’, T.V., Żylińska, A., and Zhuravlev, A.Y. 2016. International correlation of the Cambrian Series 2–3, Stages 4–5 boundary interval. Australasian Palaeontological Memoirs 49: 83–124.
USGS 2008. Landsat Image Mosaic of Antarctica. United States Geological Survey, available at:
Torsvik, T.H. and Cocks, L.R.M. 2013. New global palaeogeographical reconstructions for the Early Palaeozoic and their generation. In: D.A.T. Harper and T. Servais (eds.), Early Palaeozoic Biogeography and Palaeogeography. Geological Society, London, Memoirs 38: 5–24. Crossref
Wrona, R. 1989. Cambrian limestone erratics in the Tertairy glacio-marine sediments of King George Island, West Antarctica. Polish Polar Research 10: 533–553.
Wrona, R. 2004. Cambrian microfossils from glacial erratics of King George Island, Antarctica. Acta Palaeontologica Polonica 49: 13–56.
Wrona, R. 2009. Early Cambrian bradoriide and phosphatocopide arthropods from King George Island, West Antarctica: biogeographic implications. Polish Polar Research 30: 347–377. Crossref
Yang, B., Steiner, M., and Keupp, H. 2015. Early Cambrian palaeobiogeography of the Zhenba-Fangxian Block (South China): Independent terrane or part of the Yangtze Platform? Gondwana Research 28: 1543–1565. Crossref
Acta Palaeontol. Pol. 66 (1): 207–229, 2021
https://doi.org/10.4202/app.00758.2020