
Recurrent volcanic activity recorded in araucarian wood from the Lower Cretaceous Springhill Formation, Patagonia, Argentina: Palaeoenvironmental interpretations
GEORGINA M. DEL FUEYO, MARTÍN A. CARRIZO, DANIEL G. POIRÉ, and MAITEN A. LAFUENTE DIAZ
Del Fueyo, G.M. Carrizo, M.A. Poiré, D.G., and Lafuente Diaz, M.A. 2021. Recurrent volcanic activity recorded in araucarian wood from the Lower Cretaceous Springhill Formation, Patagonia, Argentina: Palaeoenvironmental interpretations. Acta Palaeontologica Polonica 66 (1): 231–253.
This paper describes a petrified trunk collected from a conglomerate bed of the Springhill Formation (Berriasian–Valanginian) in the Estancia El Álamo locality, Santa Cruz Province, Argentina. The fossil trunk is classified within the ubiquitous genus Agathoxylon and the wood anatomy shows a close affinity to that of Araucariaceae. This Patagonian wood has a distinct combination of anatomical characteristics unique among all known species from the Jurassic and Cretaceous of Western Gondwana allowing to diagnose a new fossil taxon Agathoxylon mendezii sp. nov. Sedimentological and megafloristic proxies of the Springhill Formation suggest that Agathoxylon mendezii sp. nov. grew under a warm and wet climate, which indicates a subtropical to temperate palaeoenvironment. However, the large number of frost rings in the earlywood of this araucarian tree suggests that the palaeoenvironment at Estancia El Álamo was subjected to recurrent disturbances, most likely caused by regional continuous volcanic activity originating from volcanoes located far away to the west. This activity would have produced periodic stratospheric veils that promoted rapid decreases in surface air temperature; the wood response to such stressful conditions would have been the formation of numerous (at least five) frost rings. Although recurrent eruptions in Patagonia during the Early Cretaceous are well recorded, this study is the first to register eruptions recorded in a coniferous wood.
Key words:Araucariaceae, Agathoxylon, volcanic eruptions, wood, Berriasian, Valanginian, South America.
Georgina M. Del Fueyo [georgidf@yahoo.com.ar], Martín A. Carrizo [blackdisk@gmail.com], and Maiten A. Lafuente Diaz [maitenlafuentediaz@gmail.com], Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, CONICET, Av. Ángel Gallardo 470, (1405) Buenos Aires, Argentina.
Daniel G. Poiré [poire@cig.museo.unlp.edu.ar], Centro de Investigaciones Geológicas, UNLP-CONICET, calle 1 n. 644, 1900, La Plata, Buenos Aires, Argentina.
Received 11 June 2020, accepted 27 September 2020, available online 24 February 2021.
Copyright © 2021 G.M. Del Fueyo et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The breakup of Gondwana in the Late Triassic to Early Cretaceous times was manifested by intensive rifting processes and appearance of extensional arc systems. Prior to the formation of the Andes in South America, the rifting associated with the opening of the South Atlantic Ocean, and the subduction along the western margin of South America produced a complex series of fore-arc, intra-arc, and retro-arc basins associated by volcanic events that repetitively ravaged this region (Biddle et al. 1986; Kay et al. 2005). As a product of this extensive geological and thermal framework, the Austral-Magallanes Basin developed in South America during the Mesozoic, in the triple junction of the Nazca, South American, and Antarctic plates (Biddle et al. 1986; Varela et al. 2012; Ghiglione et al. 2016; Aramendía et al. 2018, 2019; Cuitiño et al. 2019; Barberón et al. 2019; Gallardo Jara et al. 2019). Subsequently, the Springhill Formation was deposited after the development of the volcano-sedimentary El Quemado Complex, infilling mainly grabens (Galeazzi 1998; Poiré and Franzese 2010; Richiano et al. 2016; Sachse et al. 2016). The Springhill Formation (Thomas 1949) is located in the NNW sector of the Austral-Magallanes Basin, covering the Magallanes Province of Chile, southern Patagonia, and the southern sector of the Continental Platform of Argentina (Nullo et al. 1999; Arbe 2002; Quattrocchio et al. 2006). With widely extending deposits, the Springhill Formation shows strong diachronism from the Tithonian in the southern part of the basin towards the Barremian in the north (Riggi 1957; Riccardi and Rolleri 1980; Kraemer and Riccardi 1997; Ottone and Aguirre Urreta 2000; Aguirre Urreta 2002; Schwarz et al. 2011; Richiano et al. 2013, 2016; Cuitiño et al. 2019; González Estebenet et al. 2019).
The deposits from this formation yield an abundant and diverse megaflora, mostly of excellent preservation, and more recently, additional knowledge on its composition and richness has been acquired (Carrizo 2014; Carrizo et al. 2014; Carrizo and Del Fueyo 2015). A fraction of this megaflora is derived from specimens found in core samples from Chile, Argentina, and the Argentine Continental Platform (Archangelsky 1976; Baldoni 1979, 1980). However, the most complete and numerous fossils are found in outcrops located at the Estancia El Salitral and Río Correntoso localities of the Santa Cruz Province. These consist mainly of compressions and scant impressions of complete to partially complete fronds, leafy branches with different degrees of branching, simple to pinnate leaves, scale leaves, and carbonised fragments of wood that belong to pteridophytes, pteridosperms, Cycadales, Bennettitales, Ginkgoales, and conifers (Baldoni 1977; Baldoni and Taylor 1983; Villar de Seoane 1995, 2001; Carrizo et al. 2014; Carrizo and Del Fueyo 2015). Although some fragments of wood have been found at the Estancia El Salitral and Río Correntoso localities, no diagnostic features were observed due to the high degree of carbonisation; hence, their affinity could not be determined (Carrizo and Del Fueyo 2015).
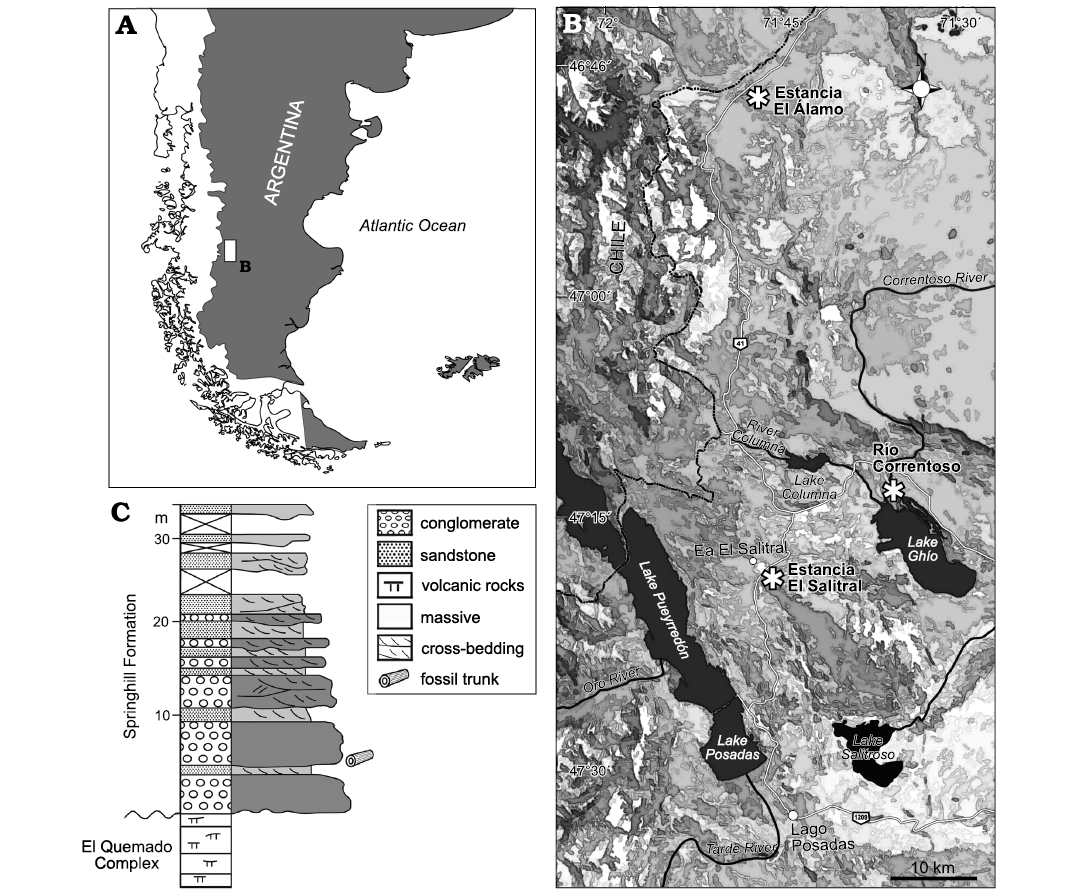
Fig. 1. Map showing location od the study area (A) and the three fossiliferous localities (asterisked) of the Springhill Formation, Santa Cruz Province, Argentina (B). C. Stratigraphic section of the Springhill Formation in the Estancia El Álamo locality.
In this contribution, a petrified conifer trunk found with well-preserved secondary xylem in a new fossiliferous locality of the Springhill Formation, Estancia El Álamo, is comprehensively described. Estancia El Álamo is located 50 km north to the Estancia El Salitral and Río Correntoso localities (Fig. 1). The wood anatomy corresponds to Agathoxylon Hartig, 1848, and the combination of characteristics is notably different from those of other known species of the genus, thereby allowing the creation of the new species, Agathoxylon mendezii sp. nov. The anatomical characteristics of this Patagonian wood show a close affinity to the family Araucariaceae. Palaeoenvironmental evidence during the deposition of the Springhill Formation at the Estancia El Álamo locality suggests that the araucarian trees grew under a warm, possibly subtropical, and wet climate. Nevertheless, the occurrence of numerous frost rings in the early wood of Agathoxylon mendezii sp. nov. indicates that the palaeoenvironment at this locality was subjected to recurrent disturbances. These disturbances are most likely attributable to the recurrent volcanic activity that is well recorded for Patagonia during the Early Cretaceous, promoting severe stressful conditions for plant communities (Archangelsky et al. 1995; Archangelsky 2001a, b; Stothers 2009). Additionally, a hypothetical reconstruction of the palaeoenvironment showing pre-eruption, eruption, and post-eruption successions that occurred at the Estancia El Álamo locality is proposed.
Institutional abbreviations.—MPM-PB, Paleobotany Collection of the Regional Museum Padre Manuel Jesús Molina, Río Gallegos, Santa Cruz Province, Argentina.
Other abbreviations.—EDS, energy dispersive X-ray spectroscopy; LM, light microscopy; SEM, scanning electron microscopy; XRD, X-ray diffraction.
Geological setting
The fossil trunk from the Springhill Formation was found in a massive, whitish, polymictic conglomerate with fine-grained sandy matrix and rounded to sub-angular quartz clasts, chert and minor amount of volcanic and pyroclastic clasts inter-bedded with cross-bedding, quarzitic, pebbly sandstone beds (Figs. 1, 2). Palaeocurrent data from the cross-bedding structures show a primarily northern supply. In the Estancia El Álamo locality outcrop, some volcanic and pyroclastic clasts have been observed, which are unusual in southern localities. Remarkably, the southernmost outcrops of the Springhill Formation exclusively comprise quartzite rocks without pyroclastic content (Arbe 2002; Schwarz et al. 2011; Richiano et al. 2012, 2015, 2016).
The rift stage fill of the Austral-Magallanes Basin involves the El Quemado Complex (Fig. 2A), with its essentially volcanic rocks, accompanied by fine-grained and calcareous sedimentary facies (Fig. 3). In the final part of this rift stage, some grabens were developed with the subsequent deposition of the Springhill Formation (Galeazzi 1998; Poiré and Franzese 2010; Ghiglione et al. 2015; Richiano et al. 2016; Sachse et al. 2016). During the thermal subsidence stage, the Río Mayer Formation and Piedra Clavada Formation were deposited whereas the overlaying Mata Amarilla Formation was developed at the beginning of the foreland stage (Fig. 3).

Fig. 2. Outcrops of the Springhill Formation in the Estancia El Álamo locality. A. Panoramic view of the Springhill Formation outcrop overlying the El Quemado Complex. B. Polymictic conglomerate beds with a sandstone bed intercalation, Springhill Formation. C. Detail of the polymictic conglomerate with siliceous (S) and volcanic (V) clasts. D. Pyroclastic (P) and siliceous (S) clasts in the conglomerate.
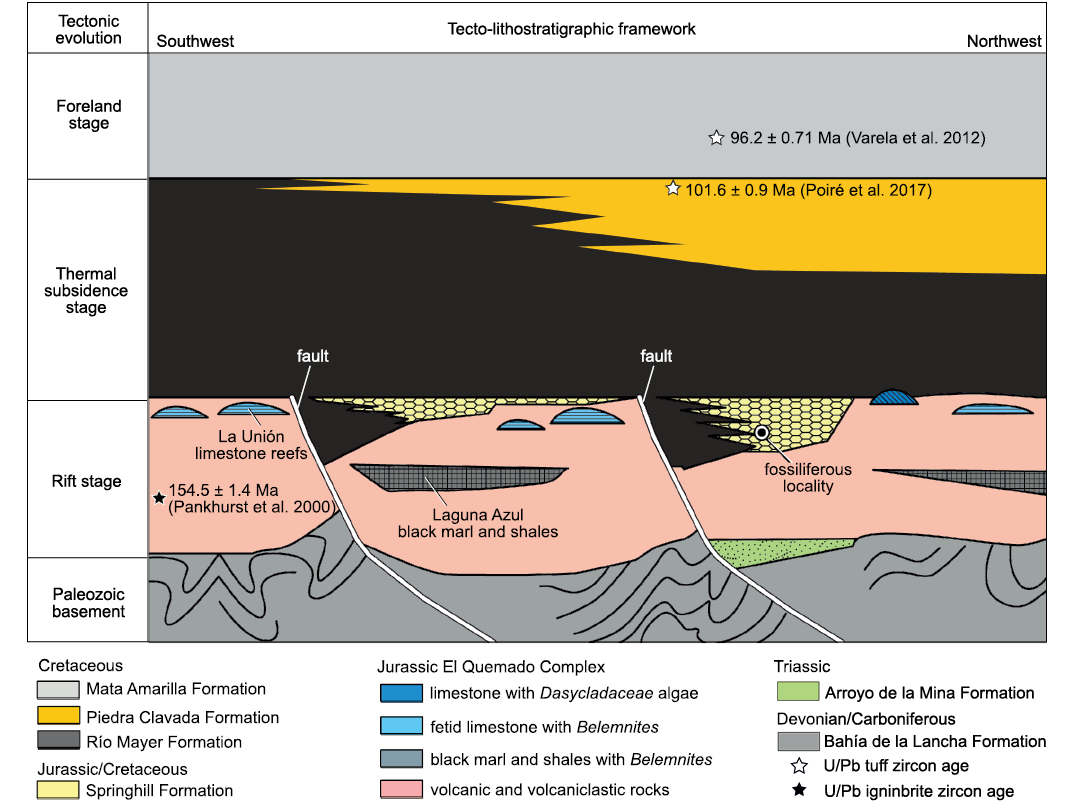
Fig. 3. Trunk location in the tecto-stratigraphic framework of the initial infilling of the Austral-Magallanes Basin, from the rift stage to the beginning of the foreland stage (modified from Poiré et al. 2017). Not to scale.
The El Quemado Complex was dated by Pankhurst et al. (2000) as Jurassic (Kimmeridgian; 154.5 ± 1.4 Ma), while the Piedra Clavada Formation was assigned to the latest Early Cretaceous (upper Albian; 101 ± 0.9 Ma; Poiré et al. 2017), and the Mata Amarilla Formation was dated as earliest Late Cretaceous (Cenomanian; 96.2 ± 0.71 Ma; Varela et al. 2012). The deposits of the Springhill Formation are widely distributed and exhibit transgressive characteristics. The resulting heterochrony of this unit makes it difficult to assign it to a single age. Based on the content of palynomorphs, Ottone and Aguirre Urreta (2000) restricted the age of the Springhill Formation for the Estancia El Salitral locality to the late early Hauterivian–early Barremian. Moreover, several authors suggested a Valanginian/Hauterivian and Berriasian?/Valanginian–early Hauterivian age for the subsurface deposits of Springhill Formation in Chile, Tierra del Fuego Province and the Continental Platform (Argentina), based mainly on palynomorphs (Palamarczuk et al. 2000; Archangelsky and Archangelsky 2004; Spalleti et al. 2009). Previous studies of the Springhill Formation in the Estancia El Salitral and Río Correntoso localities suggested an Early Cretaceous age (Archangelsky 1976; Baldoni 1977, 1979; Archangelsky et al. 1981; Cortiñas and Arbe 1981; Baldoni and Taylor 1983). Furthermore, in the Lago San Martín locality, records of the ammonites Jabronella, Neocosmoceras, and Delphinella in the upper part of the Springhill Formation indicate a Berriasian age (Riccardi 1976, 1977, 1988; Riccardi et al. 1992).
The age of the base of the succeeding Río Mayer Formation, in the Estancia El Salitral and Río Correntoso localities, is poorly constrained; on the other hand, in the upper part, a marine molluscan assemblage containing the ammonites Aegocrioceras sp. and Crioceratites sp. suggests an age probably no younger than upper Berriasian for the Springhill Formation (Carrizo et al. 2014). This faunal association is similar to that described by Aguirre Urreta et al. (2007) in Chile and in the Neuquén Basin, which was dated as latest early Hauterivian–early late Hauterivian in age. Taking into consideration the proximity of the Springhill Formation outcrops of the Estancia El Álamo with those of the Estancia El Salitral and Río Correntoso localities, the age of the Springhill Fm. at the Estancia El Álamo locality can be constrained to the Berriasian–Valanginian.
Material and methods
The wood samples were taken from the inner and outer parts of the log. Then, the material was thin-sectioned in standard transverse, tangential, and radial sections and studied under a light microscope (LM). For scanning electron microscopy (SEM) analysis, wood fragments were split into the three views, immersed in a sonic bath to remove loose dust particles, dried at room temperature (20–25ºC), and finally adhered to stubs and coated with gold-palladium. The anatomical wood characteristics are described following the IAWA List of Microscopic Features for Softwood Identification (IAWA Committee 2004). The key for conifer-like wood genera recommended by Philippe and Bamford (2008) was used for systematic assignment of the Patagonian wood. Additionally, the nomenclature recommendations related to Araucaria-like fossil wood made by Rößler et al. (2014) were followed. Comparisons with extant conifer taxa were carried out mainly following Greguss (1955, 1972). Each anatomical wood element was measured considering a set of numbers more than thirty (n >30). Measurements are indicated with their average, minimum, and maximum values in brackets. For the LM observations a Leica DM2500 was used, and micrographs were taken with a Leica DFC 280. SEM observations were made with a Philips XL30 TMP SEM at 15.1 kV at the Argentine Natural Sciences Museum Bernardino Rivadavia.
The mineralogical and chemical composition of the trunk was determined via XRD and SEM/EDS. XRD analyses were carried out on finely ground sample material (mesh 230 ASTM = bulk sample) and very-fine material (<10 µm = clay fraction). The clay fraction sample was analysed through three XRD runs on air-dried, ethylene glycol (EG)-solvated, and heated specimens. Samples were measured using a PANalytical X̕ Pert PRO diffractometer, with a Cu lamp (kα = 1.5403 Å) operated at 40 mA and 40 kV at the Centro de Investigaciones Geológicas (UNLP-CONICET, La Plata). The chemical elemental composition was determined through SEM/EDS. Their spectra were obtained using an FEI Quanta 200 SEM-EDAX Phoenix 40 with an accelerating voltage of 20 kW and a spot size from 3 to 4 mm at the Facultad de Ingeniería, Universidad Nacional de La Plata.
The fossil wood is housed in the Paleobotany Collection of the Regional Museum Padre Manuel Jesús Molina, Río Gallegos, Santa Cruz Province, Argentina, under the acronym MPM-PB.
Mineral and chemical composition of the fossil trunk
The XRD analyses revealed that the trunk is composed exclusively of quartz without any other silica mineral such as cristobalite, tridymite, A-opal, or CT-opal (Fig. 4A, B). Moreover, the clay fraction patterns show only quartz without clay minerals (Fig. 4B). The SEM/EDS spectra (Fig. 4E–I) confirmed the mineralogical results obtained through the XRD analyses, which detect presence of Si and O. Additionally, C has been recognized, while Au and Pd are part of the sample coating.
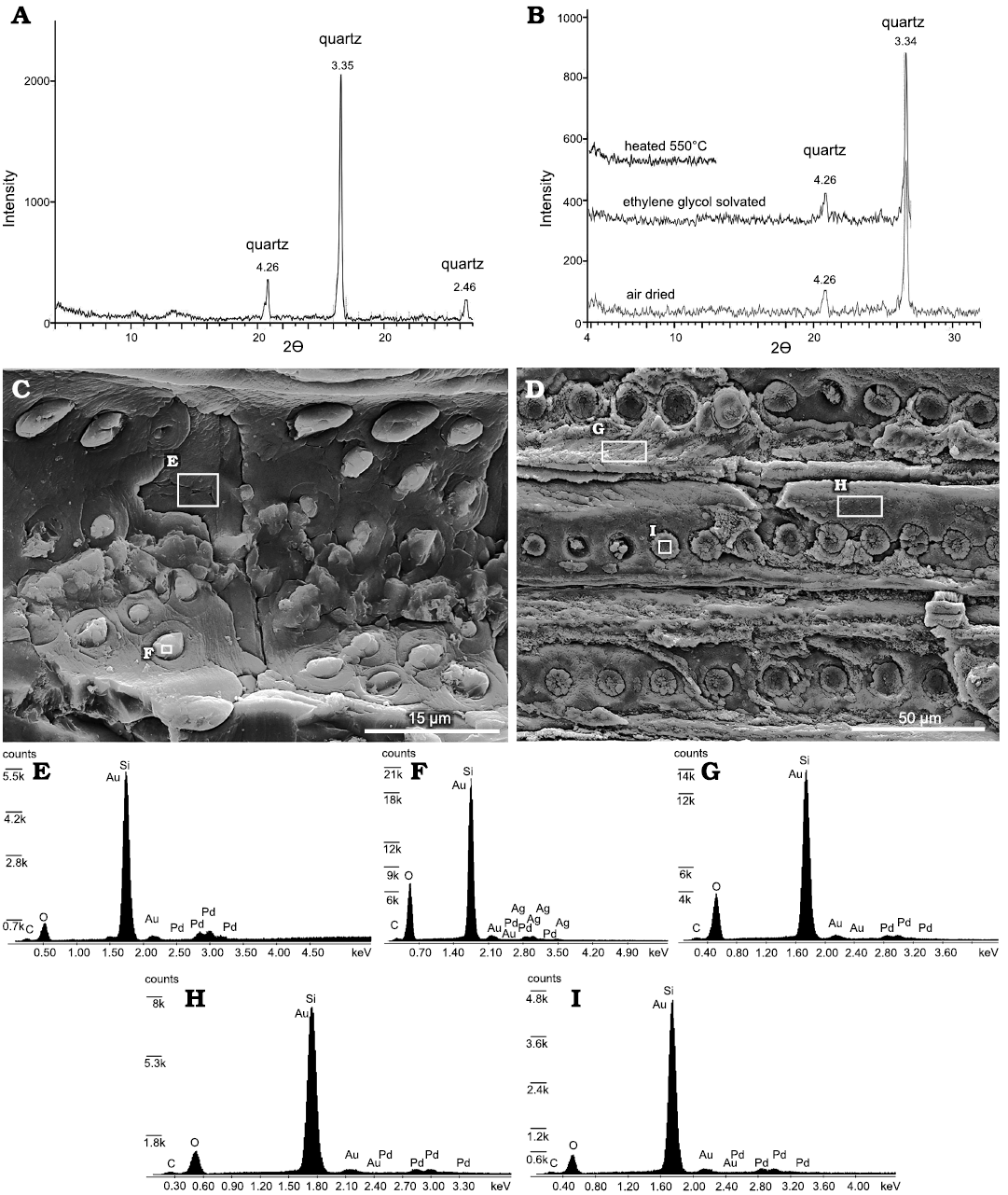
Fig. 4. XRD and SEM/EDS mineralogical and chemical analysis of Agathoxylon mendezii sp. nov. (MPM-PB-15596), Estancia El Álamo, Santa Cruz Province, Argentina, Berriasian–Valanginian. A. XRD pattern of the bulk sample showing quartz composition of the trunk. B. XRD pattern of the clay fraction showing no presence of clay minerals in the trunk. C, D. SEM of cross-field pits and tracheids in radial section. Note that quadrangular and rectangular areas correspond to the spot analysis shown in E–I. E–I. EDS patterns of xylem elements. E. Parenchyma ray cell wall. F. Inner aperture in cross-field pits. G, H. Tracheids cell walls. I. Tracheid pit cavity. All the EDS patterns are showing Si and O components.
Detailed SEM/EDS determinations on parenchyma cells, tracheids, and cross-field pits taken by spot analysis showed that the Si distribution is homogenous (Fig. 4C–I). Moreover, inner apertures in the cross-field pits are infilled with cement composed entirely of Si (Fig. 4C, F). Therefore, the wood consists of a siliceous cellular petrifaction that has retained a good histological structure.
Systematic palaeobotany
Clade Spermatophyta
Clade uncertain
Genus Agathoxylon Hartig, 1848
Type species: Agathoxylon cordaianum Hartig, 1848; Triassic, Coburg, Germany.
Agathoxylon mendezii sp. nov.
Figs. 5–9.
Etymology: Dedicated to Ignacio Méndez, owner of the Estancia El Álamo, who kindly allowed us to visit and collect the fossil wood.
Holotype: MPM-PB-15596 (silicified wood fragment).
Type locality: Estancia El Álamo, Santa Cruz Province, Argentina, GPS S46°49´25”; W71°50´22”.
Type horizon: Fine-grained, whitish, polymictic, conglomerate bed, Springhill Formation, Berriasian–Valanginian.
Diagnosis.—Pycnoxylic homoxylic secondary wood. Growth rings slightly marked. Tracheid pitting of araucarian type, predominantly uniseriate; when biseriate, alternate to sub-opposite; pits always contiguous, circular. Cross-fields of araucarioid type, 5–16 pits in three to four contiguous alternate rows; pits rounded with circular inner apertures. Rays homocellular, predominantly uniseriate, medium. Frequent resin plugs.
Description.—The studied specimen is a decorticated trunk with preserved pycnoxylic and homoxylic secondary wood. The trunk is at least 0.72 m in diameter and 3.60 m long with conspicuous branching of 0.36 m in diameter and 0.27 m of incomplete length (Fig. 5). Transverse sections of the trunk exhibit growth rings slightly marked by 1 (mostly) to 3 layers of latewood tracheids ranging from 11 to 16 mm in width (Fig. 6A, B). The rays have a rectilinear trajectory and appear to be filled by dark contents, probably resins, making it difficult to recognise individual parenchyma cells. Rays are separated by 1–7 rows of tracheids (Fig. 6C). The latewood tracheids are rectangular and flattened with a radial diameter of 14.5 (11.7–18.3) µm and a tangential diameter of 30.5 (23.6–40.5) µm, whereas those of the earlywood are polygonal with rounded to oval lumen with a radial diameter of 40 (20–55.5) µm and a tangential diameter of 38 (23–51) µm (Fig. 6B). The tracheid wall is approximately 5 (2–8.5) µm thick. Many tracheids show dark contents, presumably of the same origin as that of the rays (Fig. 6C). Inter-cellular axial spaces are frequent. Axial parenchyma cells are absent.
In the tangential section, the rays are uniseriate ranging 3–20 cells, most commonly 3–5 cells high and less frequently 6–12 or sporadically 13–20 cells high. Partially biseriate rays are less common; when present, they are 6–7 cells high (Fig. 6D–G). The transverse and tangential walls of the parenchyma cells are smooth. Each of these cells is approximately barrel-shaped and 23 (16–30) µm high and 22 (16–28) µm wide (Fig. 6E). Bordered pits frequently occur on the tangential walls of the tracheids in one single row and are mostly contiguous; they are circular in outline with a diameter of 15 (11–19) µm, and the inner aperture is circular with a diameter of 5 (4–6) µm. Resin plugs are frequently observed (Fig. 6E).
The radial section shows homocellular rays composed of rectangular cells 67 (35–132) µm long and 20 (15–31) µm wide and quadrangular cells 32 (26–40) µm long and 24 (20–27) µm wide (Fig. 8A). Most of the tangential walls are straight (74 %), and a few are oblique (26 %). In this section, the unpitted nature of the horizontal and tangential walls is evident. The arrangement of the pits on the radial walls of the tracheids is mainly in one row (69 %) and less commonly in two rows (31 %). When uniseriate, the pits are contiguous with neighbouring pits, whereas when biseriate, the pits are contiguous and alternate to sub-opposite (Fig. 8B, C). In most of the cases, the pits are circular in outline with a diameter of 13 (9–17) µm (Fig. 9A, B). In a few cases, uniseriate pits are slightly flattened (Fig. 9A). The inner aperture is circular (59 %) with a diameter of 4.5 (2.5–6.5) µm and elliptical (41 %) with a width of 4.3 (3–6) µm and a length of 5 (4–6.5) µm (Fig. 9B). Occasionally, the transition from sub-opposite/alternate to uniseriate pits occurs on the same tracheid radial wall (Fig. 8B). The length of the tracheids is difficult to measure owing to poor preservation; nevertheless, some individual elements are fusiform in outline. Cross-fields show numerous (5–16) contiguous bordered pits alternately placed in three to four vertical rows (Fig. 9C–E). Individual pits are circular in outline with a diameter of 6 (3.5–9.5) µm, and the inner aperture is circular with a diameter of 2 (1.5–3.5) µm. In very few cases, the innermost pits are polygonal in outline (Fig. 9D). Additionally, some inner apertures appear infilled with siliceous cement, resembling an ellipse in outline (Fig. 9E).

Fig. 5. Araucarian wood Agathoxylon mendezii sp. nov. (MPM-PB-15596), Estancia El Álamo, Santa Cruz Province, Argentina, Berriasian–Valanginian. A, B. Trunk showing diameter and incomplete length. Note branches (arrows). C. Deep intrusion of the trunk into the deposits. D. Detail of the decorticated trunk.

Fig. 6. Araucarian wood Agathoxylon mendezii sp. nov. (MPM-PB-15596), Estancia El Álamo, Santa Cruz Province, Argentina, Berriasian–Valanginian. Wood sections observed under LM. A–C. Transverse view. A. Slightly marked growth ring (arrows). B. Detail of growth ring, arrow shows layers of rectangular-flattened latewood tracheids. C. Detail of earlywood tracheids and rectilinear trajectory of rays (arrow). D–G. Longitudinal tangential view. D. General aspect, arrows indicate partially biseriate rays. E–G. Details of biseriate rays (arrows), arrowhead shows resin plugs. Scale bars: A, 500 µm; B, C, 150 µm; D–G, 100 µm.
Remarks.—Transverse sections of this Patagonian wood exhibit, in at least 5 annual rings, layers of ca. 10 compressed and distorted cells making it difficult to recognise the limit of each growth ring (Fig. 7A, B). This abnormal tissue appears in the earlywood and consists of inner and outer parts (Fig. 7C). The inner part has approximately 5 layers of normal- to irregular-shaped tracheids that grade into 1–2 layers of deformed tracheids, which appear tangentially flattened. All these layers occur in almost regular rows followed by a conspicuous amorphous dark layer of highly collapsed cells (Fig. 7C). Towards the outer part, a few layers of remarkably distorted axial tracheids are observed, but they are difficult to recognise individually. These gradually grade into normal tracheids of regular shape and in regular rows (Fig. 7C). Through the entire abnormal tissue, the normal rectilinear trajectory of the parenchyma ray cells becomes sinuous and distended. In addition, most of the tracheids and parenchyma ray cell lumens appear to be filled by dark contents attributable to resins (Fig. 7C).
Geographic and stratigraphic range.—Type locality and horizon only.

Fig. 7. Araucarian wood Agathoxylon mendezii sp. nov. (MPM-PB-15596), Estancia El Álamo, Santa Cruz Province, Argentina, Berriasian–Valanginian. Transverse sections observed under LM. A. General aspect showing numerous, at least five frost rings (brackets). B, C. Detail of frost rings. B. Two frost rings. Note normal and rectilinear trajectory of rays (arrows) alternate with more sinuous and distended rays (arrowheads). C. Detail of frost ring cell layers, from inside to outside. Note normal tracheids that gradually grade into irregular shaped tracheids (bar) followed by a dark layer of collapsed dead cells (arrow) followed by a layer of distorted axial tracheids difficult to recognise individually. Also note dark contents in lumen cells. Scale bars: A, 3 mm; B, 1.5 mm; C, 150 µm.

Fig. 8. Araucarian wood Agathoxylon mendezii sp. nov. (MPM-PB-15596), Estancia El Álamo, Santa Cruz Province, Argentina, Berriasian–Valanginian. Radial sections observed under SEM. A. General aspect showing ray cells (arrow), cross-fields pits (circle), axial tracheids (arrowhead). B. Detail of tracheids with uniseriate, and contiguous pits, arrow shows transition from biseriate to uniseriate pit rows. C. Detail of tracheids with biseriate, contiguous, and alternate to sub-opposite pits. Scale bars: A, B, 200 µm; C, 100 µm.
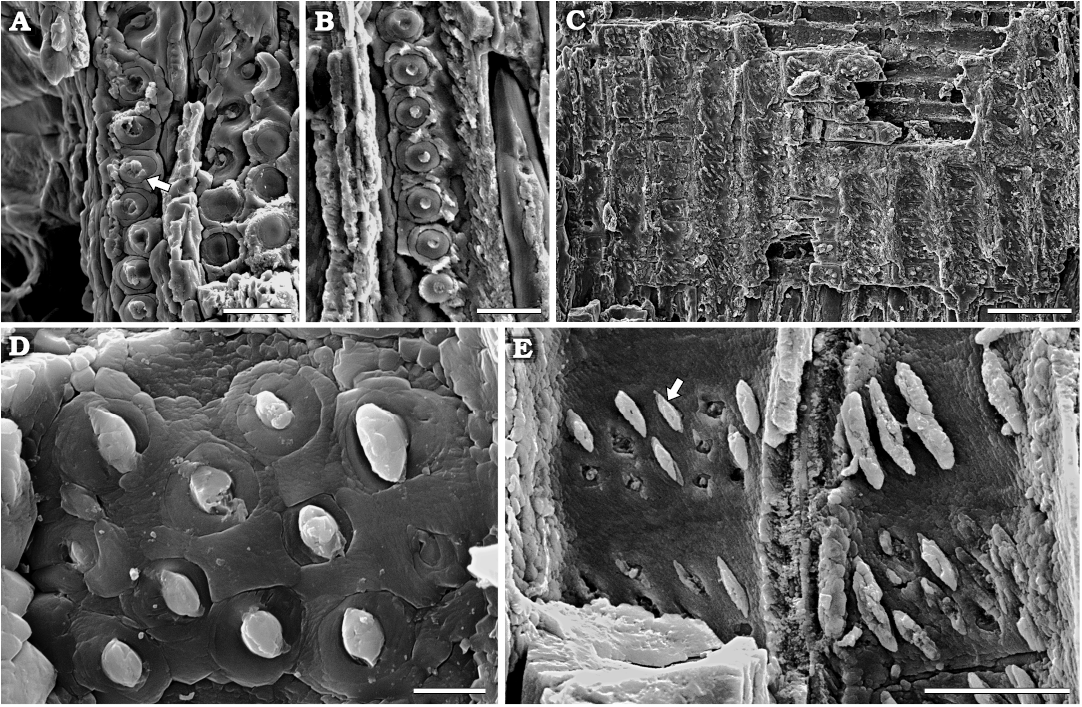
Fig. 9. Araucarian wood Agathoxylon mendezii sp. nov. (MPM-PB-15596), Estancia El Álamo, Santa Cruz Province, Argentina, Berriasian–Valanginian. Details of radial sections under SEM. A. Tracheid with slightly flattened pits (arrow). B. Tracheid showing contiguous pits with circular inner aperture. C–E. Araucarioid cross-field pits. C. General aspect. D. Contiguous alternate bordered pits placed in four vertical rows. Note circular pits in outline and circular inner aperture. E. Detail of inner apertures infilled with Si cement in elliptical form (arrow). Scale bars: A, B, E, 25 µm; C, 100 µm; D, 5 µm.
Discussion
Nomenclatural background and botanical affinity.—Araucaria-like fossil woods have been indistinctly included within three commonly known genera, Araucarioxylon Kraus, 1870, Dadoxylon Endlicher, 1847, and Agathoxylon Hartig, 1848. However, in most cases, nomenclatural rules were not strictly followed. In an attempt to avoid such proliferation of the genera and to adhere to the International Code of Botanical Nomenclature, Philippe (1993, 2011) and Bamford and Philippe (2001) pointed out that Araucarioxylon and Dadoxylon are both illegitimate and superfluous synonyms of Pinites Lindley and Hutton, 1831, whereas Agathoxylon is a legitimate name because it has priority over the two other genera and its validity has been established. More recently, Rößler et al. (2014) reinforced these ideas and strongly advocated the use of Agathoxylon for fossil woods with Araucaria-like anatomy. Therefore, in accordance with these recommendations, Agathoxylon is used in this contribution.
Agathoxylon is a globally distributed genus created on the basis of a brief list of characteristics that include woods with araucarian tracheid pitting, araucarioid cross-field pits, and axial parenchyma (Hartig 1848). However, in an extensive work on Jurassic woods from France, Philippe (1995) noted that the occurrence of axial parenchyma was of little systematic value and thus amended the original definition of Agathoxylon to include fossil woods both with and without axial parenchyma, with mainly uniseriate rays and smooth walls in the parenchyma cells.
Accordingly, the Patagonian fossil wood described here is classified within the genus Agathoxylon based on the distinctive araucarian pitting on the radial walls of the tracheids, araucarioid-type cross-fields, and unpitted ray-cell walls. The combination of several anatomical characteristics observed in the fossil wood is quite distinct from other species of the genus, thus allowing the creation of a new fossil species, Agathoxylon mendezii sp. nov. Notably, the anatomy of the Patagonian wood shows similarity with that of members of Araucariaceae. However, as reported by several authors, Agathoxylon is a ubiquitous genus to which many different and unrelated groups of plants were related such as the Palaeozoic Cordaitales, Pentoxylales or Glossopteridales; the Mesozoic Pteridospermales and extinct Araucariaceae or Cheirolepidiaceae; and, extant Araucariaceae (Philippe et al. 2004; Philippe and Bamford 2008; Taylor et al. 2009; among others).
In Patagonia, reliable records of seed cones, wood, and leaves referring to Araucariaceae indicate that the family was well established by the Middle Jurassic (Stockey 1978; Falaschi et al. 2011; Del Fueyo et al. 2019). During the Cretaceous, the diversity of Araucariaceae reached its peak, as evidenced by records of seed cones, pollen cones, dispersed araucarian pollen grains, leaves, and wood (Del Fueyo et al. 2007). In particular, the finding of dispersed pollen grains, Araucariacites, Cyclusphaera, and Balmeiopsis, and bract-scale complexes assigned to section Eutacta of Araucaria from the Springhill Formation indicate that the plant assemblage of the Early Cretaceous also includes Araucariaceae (Archangelsky and Archangelsky 2004; Carrizo and Del Fueyo 2015). Towards the Cenozoic, most taxa of Araucariaceae became extinct. At present, the only surviving taxon in Patagonia is Araucaria araucana, section Araucaria of Araucaria, restricted to cool temperate forests in the SW of Patagonia (Panti et al. 2011; Wilf et al. 2014).
Recent studies based on molecular phylogeny and fossil records of six extant family-level conifer clades (Araucariaceae, Cupressaceae, Podocarpaceae, Pinaceae, Sciadopityaceae, and Taxaceae) estimated late Palaeozoic to early Mesozoic divergence ages with initial crown splits in families occurring between 190 and 160 mya in the Early to Middle Jurassic (Leslie et al. 2018). Additionally, Escapa and Catalano (2013) confirmed the monophyly of the group through combined phylogenetic analyses of extant and fossil Araucariaceae and proposed that the first diversification of the family may have occurred at least during the Early Jurassic.
Taking into consideration the wood anatomy, fossil records, time-calibrated molecular phylogeny of conifers, and combined phylogenetic analyses on extant and fossil Araucariaceae, it is reasonable they are all assumed to be sufficient supports to suggest that Agathoxylon mendezii sp. nov. is most likely related to Araucariaceae.
Comparison among selected Mesozoic Agathoxylon species from Western Gondwana.—The tree communities inhabiting Western Gondwana during the Jurassic and Cretaceous periods were dominated by conifers of the families Araucariaceae and Podocarpaceae, whereas Cheirolepidiaceae and Cupressaceae were subordinate groups (Bamford and Corbett 1994; Falcon-Lang and Cantrill 2001; Del Fueyo et al. 2007; Quattrocchio et al. 2007; Bodnar et al. 2013; Vera and Césari 2015; Brea et al. 2016; Greppi et al. 2020). Remains of these assemblages are mainly preserved as petrified woods, among which fossils of Araucariaceae logs are the most abundant and frequently occurring. Taking into account that a substantial number of these woods are poorly preserved or incompletely studied, in this work, we compare Agathoxylon mendezii sp. nov. to taxa with anatomically complete descriptions, and thus, those informally published or left in open nomenclature are not considered here. Moreover, because many fossil woods of pre-Jurassic age that were assigned to Araucariaceae show doubtful affinities with this family, comparisons with the Patagonian wood are restricted to those fossils of Jurassic and Cretaceous provenances.
Among the anatomical features observed in the wood of Agathoxylon mendezii sp. nov., four features, namely, uniseriate to biseriate (alternate to sub-opposite) radial pitting with contiguous circular pits, uniseriate to partially biseriate rays 3–20 cells high, abundant resin plugs, and up to 16 circular contiguous alternate pits per cross-field, distinguish this taxon from those species recorded in Jurassic and Cretaceous strata of Chile, Argentina, and Antarctica (Table 1). The high number of pits per cross-field differentiates Agathoxylon mendezii sp. nov. from most (14) of the considered species, which show cross-fields with up to 4 or 8 pits. In addition, in 8 of these taxa: Agathoxylon antarcticus (Poole and Cantrill, 2001) Pujana, Santillana, and Marenssi, 2014; Agathoxylon (Araucarioxylon) floresii Torres and Lemoigne, 1989; Agathoxylon (Araucarioxylon) kellerense Lucas and Lacey, 1981; Agathoxylon (Araucarioxylon) ohzuanum (Nishida, Ohsawa, Nishida, and Rancusi, 1992) Gnaedinger and Herbst, 2009; Agathoxylon (Araucarioxylon) parachoshiense Nishida and Nishida, 1987; Agathoxylon protoaraucana (Brea, 1997) Gnaedinger and Herbst, 2009; Agathoxylon pseudoparenchymatosum (Gothan, 1908) Pujana, Santillana, and Marensi, 2014; Agathoxylon santalense (Sah and Jain, 1964) Kloster and Gnaedinger, 2018; the number of ray cells is much lower, never reaching more than 12 cells high (Gnaedinger and Herbst 2009; Lucas and Lacey 1981; Nishida and Nishida 1987; Pujana et al. 2014; Torres and Lemoigne 1989), whereas in the 5 other species: Agathoxylon agathioides (Kräusel and Jain, 1964) Kloster and Gnaedinger, 2018; Agathoxylon (Araucarioxylon) arayaii Torres, Valenzuela, and Gonzalez, 1982; Agathoxylon (Araucarioxylon) pluriresinosum Torres and Biro-Bagoczky, 1986; Agathoxylon (Araucarioxylon) resinosum Torres and Biro-Bagoczky, 1986; Agathoxylon termieri (Attims, 1965) Gnaedinger and Herbst, 2009; the tracheid radial pitting is in 2–3 rows of alternate mostly polygonal pits (Torres and Biro-Bagoczky 1986; Torres et al. 1982), emphasizing the differences from Agathoxylon mendezii sp. nov. Furthermore, the thirteenth species Agathoxylon liguanensis Torres and Philippe, 2002, also differs from this latter taxon because the rays are up to 56 cells high, and the pits on the tracheid radial walls are notably flattened (Torres and Philippe 2002; Table 1).
Table 1. Comparison among selected Mesozoic Agathoxylon species from Western Gondwana; n.a., data about growth ring width, tangential pitting and resin plugs not available; –, absent.
|
Taxon and references |
Age and |
Growth rings |
Radial pitting |
Tangen- |
Rays |
Cross-fields |
Axial paren- -chyma |
Resin plugs |
|
Agathoxylon
mendezii |
Early |
slightly distinct, |
uniseriate to biseriate alternate to opposite; circular |
present |
uniseriate, rarely biseriate, |
5–16 |
– |
present |
|
Agathoxylon agathiodes (Kräusel and Jain, 1964) Kloster and Gnaedinger, 2018 |
Middle Jurassic, Argentina |
indistinct |
uniseriate, biseriate, less frecuently triseriate alternate; hexagonal |
– |
uniseriate, rarely biseriate, |
2–6 |
– |
present |
|
Agathoxylon
antarcticus (Poole and Cantrill, 2001) Pujana,
Santillana, and Marenssi, 2014 |
Middle Jurassic, Argentina; |
distinct, |
uniseriate, less frecuently
biseriate alternate; circular |
– |
uniseriate, |
2–9 |
– |
present |
|
Agathoxylon
(Araucarioxylon)
arayaii Torres, Valenzuela, and Gonzalez, 1982 |
Jurassic and |
slightly distinct |
uniseriate, biseriate alternate, and rarely biseriate opposite and triseriate alternate; hexagonal |
n.a. |
uniseriate, |
2–5 |
– |
present |
|
Agathoxylon (Araucarioxylon) chapmanae Poole and Cantrill, 2001 |
Late Cretaceous, Antarctica |
distinct |
predominantly biseriate, alternate, sometimes uniseriate, rarely triseriate; circular to hexagonal |
n.a. |
1–25 cells high |
2–11 |
– |
present |
|
Agathoxylon (Araucarioxylon) floresii Torres and Lemoigne, 1989 |
Late Cretaceous, Antarctica |
slightly distinct |
uniseriate to biseriate alternate; polygonal |
– |
uniseriate, |
1–4 |
present |
present |
|
Agathoxylon (Araucarioxylon) kellerense Lucas and Lacey, 1981 (Nishida et al. 1992) |
Late Cretaceous, Chile; |
distinct |
uniseriate to triseriate alternate; flattened to hexagonal |
n.a. |
uniseriate, |
1–4 |
n.a. |
n.a. |
|
Agathoxylon
liguanensis Torres and Philippe, 2002 |
Early Jurassic, |
slightly distinct to distinct, 10–13 mm width |
uniseriate to biseriate alternate, some times opposite; usually flattened |
present |
uniseriate, |
1–3 |
– |
– |
|
Agathoxylon (Araucarioxylon) ohzuanum (Nishida, Ohsawa, Nishida, and Rancusi, 1992) Gnaedinger and Herbst, 2009 |
Late |
indistinct |
uniseriate to biseriate alternate; circular |
present |
uniseriate, |
2–4 |
present |
– |
|
Agathoxylon (Araucarioxylon) parachoshiense Nishida and Nishida, 1987 |
Late |
distinct |
uniseriate to biseriate alternate; polygonal |
– |
uniseriate, |
2–4 |
– |
n.a. |
|
Agathoxylon (Araucarioxylon) pichasquensis Torres and Rallo, 1981 (Nishida et al. 1992) |
Late |
slightly distinct |
uniseriate, sometimes biseriate alternate |
n.a. |
uniseriate, |
5–10 |
– |
n.a. |
|
Agathoxylon (Araucarioxylon) pluriresinosum Torres and Biro-Bagoczky, 1986 |
Late |
distinct, |
uniseriate to tetraseriate; hexagonal |
n.a. |
uniseriate, |
2–8 |
present |
present |
|
Agathoxylon protoaraucana (Brea, 1997) Gnaedinger and Herbst, 2009 |
Early Jurassic, |
distinct, 35–75 mm width |
uniseriate, very rarely with
transition to biseriate alternate |
present |
uniseriate to partially biseriate, 2–12 cells high |
1–8 |
n.a. |
n.a. |
|
Agathoxylon
pseudoparenchymatosum (Gothan, 1908) |
Late Cretaceous, Antarctica and Cretaceous, Chile |
distinct, |
uniseriate, rarely biseriate alternate; hexagonal |
– |
uniseriate, |
2–6 |
– |
present |
|
Agathoxylon
(Araucarioxylon) resinosum
Torres and |
Late |
distinct, |
uniseriate to triseriate alternate; hexagonal |
present |
uniseriate, rarely biseriate, |
2–3 |
– |
present |
|
Agathoxylon santacruzense Kloster and Gnaedinger, 2018 |
Middle Jurassic, Argentina |
distinct, |
mostly biseriate; rarely uniseriate or triseriate; alternate, hexagonal |
– |
uniseriate, |
2–14 |
– |
– |
|
Agathoxylon santalense (Sah and Jain, 1964) Kloster and Gnaedinger, 2018 |
Middle Jurassic, Argentina |
distinct, |
rarely uniseriate; mostly biseriate, occasionally triseriate; alternate, hexagonal |
– |
uniseriate, |
2–6 |
– |
present |
|
Agathoxylon termieri (Attims, 1965) Gnaedinger and Herbst, 2009 (Kloster and Gnaedinger 2018) |
Middle Jurassic, Argentina |
distinct, |
uniseriate to biseriate, occasionally triseriate; alternate, circular |
n.a. |
uniseriate, |
1–8 |
n.a. |
n.a. |
The remaining 3 taxa, Agathoxylon (Araucarioxylon) chapmanae Poole and Cantrill, 2001, Agathoxylon santacruzense Kloster and Gnaedinger, 2018, and Agathoxylon (Araucarioxylon) pichasquensis Torres and Rallo, 1981 (Torres and Rallo 1981), share the characteristic of numerous pits per cross-field with Agathoxylon mendezii sp. nov., although they have fewer pits, up to 11, 14, and 10 pits, respectively. Moreover, the tracheid radial pitting in the 2 former taxa is in 2–3 rows of alternate pits that are remarkably polygonal in outline, whereas in Agathoxylon (Araucarioxylon) pichasquensis, the rays are exclusively uniseriate and resin plugs are absent features not observed in Agathoxylon mendezii sp nov. (Table 1). Notably, Late Cretaceous woods from Chile assigned to Agathoxylon (Araucarioxylon) pichasquensis by Nishida et al. (1992) show uniseriate to biseriate tracheid radial pitting with distinct polygonal pits in outline and lower rays at 2–14 cells high, indicating differences from Agathoxylon mendezii sp. nov.
Comparison of Agathoxylon mendezii sp. nov. with extant Araucariaceae.—The wood anatomy boundaries between the three extant genera, Agathis Salisbury, 1807, Araucaria Jussieu, 1789, and Wollemia Jones, Hill, and Allen, 1995, are generally difficult to establish (Greguss 1955, 1972; Heady et al. 2002). However, when Agathoxylon mendezii sp. nov. is quantitatively compared with these taxa, some distinctions are noted.
Agathoxylon mendezii sp. nov. shares the characteristic of numerous pits per cross field with the mono-specific Wollemia nobilis; however, the Australian taxon has fewer pits (3–11); the radial pitting is uniseriate to triseriate with contiguous flattened to polygonal pits, and the rays are lower at 1–11 cells high (Heady et al. 2002).
Most species of Agathis show 3–5, sometimes up to 6, conspicuous polygonal pits on the radial walls of the tracheids, e.g., Agathis australis Salisbury, 1807, Agathis borneenensis Warburg, 1900, Agathis endertii Meijer Drees, 1940, Agathis labillardieri Warburg, 1900, Agathis lanceolata Lindley ex Warburg, 1900, Agathis philippinensis Warburg, 1900, Agathis robusta F.M. Bailey, 1883 (Greguss 1955, 1972), which is notably distinct from the radial pitting observed in Agathoxylon mendezii sp. nov. Furthermore, several Agathis species share uni- to biseriate radial pitting with the Patagonian fossil taxon. However, in the Australasian taxa, the uniseriate pits are always flattened, whereas the biseriate pits are markedly polygonal (e.g., Agathis microstachya J.F. Bailey and C.T. White, 1918). In addition, some taxa show fewer pits per cross-field (3, 5, or up to 8), e.g., Agathis celebica Warburg, 1900, Agathis macrophylla Masters, 1892, Agathis ovata Warburg, 1900; following Greguss (1955, 1972).
The secondary xylem of the majority of the Araucaria species exhibit mainly uniseriate, and less frequently biseriate, radial pitting similar to that observed in Agathoxylon mendezii sp. nov. However, this Patagonian fossil wood differs from the two South American taxa of Araucaria section Araucaria because in both Araucaria araucana K. Kocher, 1873, and Araucaria angustifolia Kuntze, 1898, the rays are exclusively uniseriate and the cross-fields have up to 10 or 12 pits, respectively (Tortorelli 1956). Additionally, in Araucaria angustifolia, radial pitting is also triseriate, emphasizing the differences from the fossil wood.
Agathoxylon mendezii sp. nov. resembles the taxa of Araucaria sections Bunya (Araucaria bidwillii Hooker, 1843) and Intermedia (Araucaria klinkii Lauterbach, 1913, Araucaria hunsteinii K. Schumann, 1889) in that they have ray heights of up to 20, 16, or 23 cells, respectively; however, in these extant species, the pits per cross-field are fewer than those of the fossil taxa, 1–6, 2–4, and 2–6, respectively (Greguss 1955, 1972).
On the other hand, the Patagonian wood shows several differences from species within the Araucaria section Eutacta, which are described as having secondary xylem. Agathoxylon mendezii sp. nov. notably differs from four of them: Araucaria heterophylla Franco, 1952, Araucaria humboldtensis Buchholz, 1949, Araucaria muelleri Brongniart and Grisebach, 1871, Araucaria rulei F. Mueller, 1860; in that they have lower rays up to 10 cells high and the cross-field pits are much less abundant (up to 4 or 8). On the other hand, rays in Araucaria montana Brongniart and Grisebach, 1871, are up to 22 cells high, similar to Agathoxylon mendezii sp. nov.; however, in the New Caledonia taxon, the pits per cross-field are up to 9 and radial pitting is bi- to triseriate (Greguss 1955, 1972). Furthermore, in 3 extant species: Araucaria columnaris Hooker, 1852, Araucaria cunninghami D. Don, 1837, and Araucaria subulata Vieillard, 1862; the pits per cross-field are numerous (up to 12 or 14), similar to the fossil wood studied here, although the ray heights in the 3 former taxa are much lower (up to 4 or 12 cells high) (Greguss 1955, 1972).
Palaeoenvironment of the Estancia El Álamo locality and wood response to volcanic events.—Depositional environment: During deposition of the Springhill Formation in the Estancia El Álamo locality (palaeolatitude of approximately 46°S), Agathoxylon mendezii sp. nov. exhibited a significant diameter of 0.72 m and likely a notable height, clearly suggesting that it was a well-grown tree. Although the log lacks preserved roots and bark, it shows a conspicuous branch, well-preserved xylematic elements, and no signals of compression due to sediment weight. The regional depositional environment of the lower Springhill Formation has been interpreted as continental (Sanchez and Le Roux 2003; Spalletti et al. 2009; Poiré and Franzese 2010; Schwarz et al. 2011; Richiano et al. 2013, 2016), which infilled longitudinally oriented half-grabens with the hanging-wall to the west (Richiano et al. 2016; Horton 2018). Many authors agree upon the existence of a transgressive sequence from distal fluvial facies close to the sea to shoreface deposits (Sanchez and Le Roux 2003; Schwarz et al. 2011; Richiano et al. 2016). However, the sedimentary record in the Estancia El Álamo locality suggests an alluvial fan/fluvial braided palaeoenvironment related to the continental rift deposition. This palaeogeography configuration indicates that the trunk occurs in an alluvial/fluvial channel deposit with a preferred orientation parallel to the palaeocurrents and was probably transported some short distance from its growth site before burial. Hence, the Agathoxylon mendezii sp. nov. tree is considered para-autochthonous, and it most likely grew in the uplands that dominated the Estancia El Álamo locality. In this area of Patagonia, the Springhill Formation strata are poorly exposed, showing no records of other buried logs or fossil flora that could indicate the type of plant assemblage to which Agathoxylon mendezii sp. nov. belongs. Nevertheless, the strata in which this fossil wood is embedded appear to be coeval with southern adjacent deposits (Río Correntoso and Estancia El Salitral localities ca. 50 km south; see Fig. 1) containing the compressed remains of a rich plant community composed of Ginkgophyta, Pteridophyta, Pteridospermophyta, Cycadophyta, and Pinophyta. Within the latter group, the Araucariaceae are represented by bract-scale complexes of Araucaria, referring to the Eutacta section of the genus Araucaria (Carrizo and Del Fueyo 2015). Accordingly, these plant assemblages are assumed to have inhabited the flood plains that graded into the coastal plains followed by an open-marine depositional system (Cortiñas and Arbe 1981; Arbe 2002; Schwarz et al. 2011; Richiano et al. 2013, 2015). This assumption is consistent with the rifting model suggested by Poiré and Franzese (2010), in which the Springhill Formation infilled the grabens as part of a synrift deposition. The specimen of Agathoxylon mendezii sp. nov. may represent a well-developed tree belonging to a fossil forest, which grew in the northern uplands neighbouring the southern Río Correntoso and Estancia El Salitral Cretaceous plant communities inhabiting a fluvial system.
Palaeoclimate during the deposition of the Springhill Formation: The sedimentary evidence at the Estancia El Álamo locality indicates a temperate to warm, humid climate.
The quarzitic composition of the clasts and the abundant kaolinite shown in the clay mineral analysis of the lower Springhill Formation indicates a strong weathering and a high leaching in a temperate-warm, humid climate (Riccardi 1971; Iñiguez Rodríguez and Decastelli 1984; Spalletti et al. 2006; Richiano et al. 2012, 2015). Moreover, the low content of volcanic and pyroclastic clasts of the polymictic conglomerates suggest that the sedimentation rate in the alluvial fan was high preventing the weathering of the labile clasts (Zanchetta et al. 2004; Ventra and Clarke 2018).
Similarly, sedimentological studies by Riccardi (1971) on Springhill Formation deposits located further south in the Bahía de la Lancha locality near Lago San Martín indicate a warm and humid depositional environment, inferred from to the existence of large quantities of quartz, a moderate percentage of kaolinite, and limited feldspar. Moreover, considering the presence of massive greyish mudstones beds showing initial paedogenetic characteristics, thin layers of coal, and large amounts of plant remains, Schwarz et al. (2011) proposed that the lower section of the Springhill Formation in southeastern Argentina most likely developed in humid climate conditions.
Global warm temperature during the Cretaceous was already estimated with the use of 97 data points for both sea surface and continental temperatures and the construction of crossplots (Frakes 1999). Different related palaeotemperature isotopic data have also been reported for the Austral-Magallanes Basin. Gómez Dacal et al. (2019) estimated a seawater palaeotemperature of 25°C on average for the Austral-Magallanes Basin area during the Berriasian–early Valanginian from calcite belemnite-rostra δ18O isotope results. Previously, Price and Gröcke (2002) obtained temperatures of 10°C on average for the Barremian–Albian seawater column interval from belemnite samples in the Malvinas-Falkland Plateau at a similar palaeolatitude. In the Neuquén Basin area approximately 2000 km to the north, isotopic analysis based on oyster shells indicate a warm period during the Tithonian–Valanginian, with seawater temperatures of ~25°C on average (Gómez Dacal et al. 2018). Therefore, the 100–250 m depth seawater column temperature during the deposition of the Springhill Formation ranges from ~10–25°C, which suggests ~20–35°C for the sea surface temperature (Mutterlose et al. 2010; Gómez Dacal et al. 2019). These data indicate temperate to warm temperature for the Estancia El Álamo area, which is consistent with the Early Cretaceous global warm climate already suggested by several authors (Frakes 1999; Zeebe 2001; Föllmi 2012; Hay and Floegel 2012; among others).
On the other hand, palaeofloristic studies along with cuticular analysis of the megaflora recovered in the above-mentioned Río Correntoso and Estancia El Salitral localities have revealed a humid environmental condition, under which this plant community grew during the deposition of the Springhill Formation (Carrizo et al. 2014; Carrizo and Del Fueyo 2015). These palaeoenvironmental deductions are also supported by extensive palynological studies performed on the Springhill Formation from core samples in Argentina and Chile; these studies proposed the development of a plant assemblage during the sedimentation of this unit under a warm and humid environment. Particularly, this plant community was mainly composed of conifers such as Araucariaceae, Cheirolepidiaceae, and Podocarpaceae and abundant ferns (Baldoni and Archangelsky 1983; Archangelsky and Archangelsky 2004; Guler et al. 2015). Additionally, Martínez et al. (2016) found pollen grains of Calamoideae (Palmae) in the Springhill Formation strata, considering which they suggested the existence of a subtropical to tropical palaeoenvironment for this subfamily.
Growth rings in Agathoxylon mendezii sp. nov. are slightly marked, suggesting that its growth environment in the Estancia El Álamo locality was characterised by a warm climate with weakly defined seasons. However, according to several opinions, this palaeoclimatological inference is of little value because the majority of living and fossil araucarian woods display poorly developed rings independent of the local environment or regional climate (Creber and Chaloner 1984; Falcon-Lang 2000; Philippe et al. 2015). Hence, Brison et al. (2001) pointed out that Araucariaceae do not have the ability to develop well-marked growth rings even when the inhabiting sites have strong seasonality. In addition, Agathoxylon never shows thick latewood. Therefore, its growth rings have been interpreted as being controlled genetically rather than by environmental factors (Brison et al. 2001).
Nevertheless, taking into consideration the above discussion and the additional sedimentological evidence, the clay mineral composition, and palaeoflora-based proxies of the Springhill Formation, it could be suggested that Agathoxylon mendezii sp. nov. grew during the Berriasian–Valanginian under a temperate to warm and wet climate, thus strongly suggesting a subtropical environment with abundant water supply. These inferences are correlated with the existence of a well-recorded global warm greenhouse climate during the Early Cretaceous (Kauffman and Johnson 2009), particularly in the Patagonian Province within the Austral domain, as recognised by Vakhrameev (1991), where the Estancia El Álamo locality is situated.
Wood anomaly records: The occurrence of abnormal tissue in numerous tree rings of Agathoxylon mendezii sp. nov. indicates that cambial cells were somehow injured during the period of wood growth and may, therefore, imply that the palaeoenvironment of the Estancia El Álamo locality was subjected to recurrent disturbances. The injured cells within the tree rings in this Patagonian wood include, from inside to outside, deformed axial tracheids that are tangentially flattened, a dark layer of strongly collapsed cells, and abnormal tracheids, whereas parenchyma ray cells are sinuous and distended and lumen cells appear full of resin contents. According to several authors, coniferous annual rings showing this type of injury in their xylem cells are referred to as frost rings (Bailey 1925; Glerum and Farrar 1966; Hantemirov et al. 2000; Gurskaya and Shiyatov 2002).
Deformed or collapsed tracheids and sinuous rays are typical of other wood anomalies. They are often observed when the wood was compressed by the weight of a heavy layer of sediments, promoting mainly the collapse and folding of the thin-walled earlywood cells (Schweingruber et al. 2006). As it was mentioned above, Agathoxylon mendezii sp. nov., shows no evidence of compression by sediments. Fletcher et al. (2015) found that crush zones with characteristic zigzags in a Cretaceous podocarp wood form Australia were due to flooding that increased water availability, leading to a burst of fast growth of the thin-walled earlywood cells. In Agathoxylon mendezii sp. nov., evidence of neither zigzags due to crash zones nor abrupt increases in earlywood cells by flooding were recorded (see Fig. 6A). This fact is also supported by the sedimentological studies. There is no evidence of any flooding during the sedimentation of the Springhill Formation in Estancia El Álamo locality. This is due to the absence of the characteristic fine-grained (pelitic) deposits (see Fig. 1).
As all the cell layers characteristic of typical frost injuries are anatomically well recognised in the wood anomalies of Agathoxylon mendezii sp. nov. (Glerum and Farrar 1966; Gurskaya and Shiyatov 2002; Schweingruber 2007), they can certainly be attributed to frost rings caused most likely by extreme weather conditions. In particular, the most remarkable feature in this araucarian wood is the presence of at least five frost rings.
Following Schweingruber (2007), frost rings are anatomically abnormal and ecophysiologically pathological structures produced when the air temperature drops below sub-zero values during the period of cambium activity and xylem cell growth, promoting incomplete maturation of these tissues and interrupting the normal continuity of the radial walls of the tracheids. The presence of ice crystals produced by frost in the meristematic area is believed to have altered the cell wall thickness, modifying the matrix of the cellulose, hemicellulose, and lignin contents (Arco Molina et al. 2016).
The sedimentological evidence indicates that the warm and humid palaeoenvironment of Estancia El Álamo inhabited by Agathoxylon mendezii sp. nov. was also affected by recurrent volcanic activity produced by the volcanic arc located far away to the west, which originated at the end of the rift stage (Biddle et al. 1986; Poiré and Franzese 2010; Varela et al. 2012, 2013; Horton 2018). Periodic volcanic eruptions producing large amounts of cool ash fall are well recorded to have generated continuous stress on plant communities inhabiting Patagonia during most of the Early Cretaceous (Archangelsky et al. 1995; Passalia 2009; Carrizo et al. 2011).
Several studies (e.g., Rampino and Self 1982; Stothers 2009) have revealed that large explosive eruptions eject large quantities of silicate dust and sulfur compounds into the stratosphere, increasing the optical thickness of the aerosol layer. In addition, after the ejection, the aerosol is rapidly dispersed longitudinally and latitudinally in the stratosphere, causing pronounced regional or global effects on the weather and climate depending on the magnitude of eruption (Self et al. 1981). One remarkable effect is the blocking of sunlight, promoting a rapid decrease in surface air temperature, leading to significant cooling in the vicinity of the volcano (Stothers 2009).
Hantemirov et al. (2004) and Salzer and Hughes (2007) identified the occurrence of light rings (rings with thin-walled latewood cells) and extremely narrow tree rings in coniferous wood (Juniperus sibirica Burgsdorff, 1787, Pinus longaeva D.K. Bailey, 1971, and Pinus aristata Engelmann, 1862, respectively) corresponding to colder weather induced by volcanic eruptions. When air temperature falls below the freezing point due to the eruptions, the so-called frost rings consequently form (LaMarche and Hirschboeck 1984; Schweingruber 1989).
LaMarche and Hirschboeck (1984) studied subalpine pines (Pinus longaeva D.K. Bailey, 1970 and Pinus aristata Engelmann, 1862) in western USA and reported the occurrence of an unusually high number of latewood frost rings, in some cases more than six times, directly related to major volcanogenic atmospheric veils produced by volcanoes such as Krakatoa (Java), St. Helens (USA), Etna (Italy), Santorini (Greece), and White River (Canada). In addition to this coincidence, these authors demonstrated that the multiple occurrences of frost damage were found in the wood of old and well-established trees rather than in the expected tree rings of seedlings, branches, or very young trees, which usually show higher incidences of frost injury (Glerum and Farrar 1966; Gurskaya and Shiyatov 2002).
A similar correlation to LaMarche and Hirschboeck̕s (1984) investigations was also found by Filion et al. (1986) in specimens of Picea mariana Britton, Sterns, and Poggenburg, 1888, growing at the treeline in northern Québec. In this particular case, the high frequency of light rings in the latewood of black spruces was closely associated with major volcanic eruptions like those of Laki, Iceland, and Tambora, Indonesia, which induced a temperature drop during the growing season in that region of Canada (Filion et al. 1986).
Although with some uncertainties, Stahle (1990) also linked frost injury tree rings in deciduous oaks (Quercus stellate Wangenheim, 1787, Quercus alba Linné, 1753) of the Southern Plains (USA) to large-scale explosive volcanic activity that occurred from 1814 to 1819. This activity may have forced the amplification of the El Niño-Southern Oscillation (ENSO) phenomenon, which appears to have been responsible for the unusual climate conditions (abnormally warm winter followed by severe freeze in spring) over that area during that period of time.
Taking into account this evidence and considering that Agathoxylon mendezii sp. nov. grew in a warm, almost subtropical palaeoenvironment (Fig. 10A) influenced by regionally continuous volcanic activity (Fig. 10B), the wood response to such stressful conditions would most likely be the occurrence of earlywood frost rings (Fig. 10C). The record of five frost rings in this Patagonian wood suggests the possibility of the temperature rising to its original values after the ceasing of volcanic injection (Fig. 10D). In addition, the cambium may have started to produce normal cells until the next volcanic activity. These probable scenarios (pre-eruption, eruption, and post-eruption) may have repeatedly occurred at the Estancia El Álamo locality at least four more times, promoting the formation of abnormal tissue alternated with normal tissue in the Agathoxylon mendezii sp. nov. wood (Fig. 7A). The eruption may have also ejected a large amount of cool ash because the Agathoxylon mendezii sp. nov. wood shows no sign of charring prior to permineralisation.

Fig. 10. Hypothetical scenario in the Estancia El Álamo locality (Santa Cruz Province, Argentina, Berriasian–Valanginian) following volcanic disturbances. A. Pre-eruption stage. Seedlings, juvenile, and mature trees of Agathoxylon mendezii sp. nov. growing in a warm almost subtropical palaeoenvironment. Note volcanoes in the distance. Photo shows wood with slightly growth ring. B. Initial eruption stage. Volcanoes begin to eject silicate dust and sulfur compounds into the stratosphere. C. Climax eruption stage. Aerosol layer thickness is markedly increased producing the decrease of the surface air temperature below subzero values. Photo shows wood damaged by frost. D. Post-eruption stage. Volcanoes activity begins to cease and temperature begins to rise to original values.
Traumatic frost rings have previously been reported from araucarian wood in the Triassic of Patagonia from Agathoxylon (Araucarioxylon) protoaraucana (Brea, 1997) Gnaedinger and Herbst, 2009, and the development of the frost rings was attributed to un-predictable frosts events during the growing season (Brea 1997). This study reports the first finding of numerous frost rings in Agathoxylon mendezii sp. nov. caused by regional recurrent volcanic eruptions in a Lower Cretaceous coniferous wood from Patagonia, as well as in other Mesozoic coniferous woods from Western Gondwana.
In the extant Araucaria araucana K. Kocher, 1873, wood, endemic to temperate sub-Antarctic Patagonian forests, the record of frost rings for the last 600 years allowed Hadad et al. (2012, 2019) to build reliable frost ring chronologies, indicating the capability of this araucarian taxon to document past extreme climatic events in that area. Accordingly, frost damage in Agathoxylon mendezii sp. nov. will most likely help with research of extreme palaeoclimates, particularly during the Early Cretaceous in Patagonia. However, more specimens of this taxon are required to accurately re-construct such extreme climatic events during that period of time.
Conclusions
Agathoxylon mendezii sp. nov. has araucarian affinities because of its alternate contiguous tracheid pitting and araucarioid cross-field pits that distinguish Araucariaceae woods from all other conifers. The combination of anatomical characteristics in this Patagonian wood (i.e., uniseriate to biseriate and alternate to sub-opposite radial pitting with contiguous circular pits, uniseriate to partially biseriate rays up to 20 cells high, abundant resin plugs, and up to 16 circular contiguous alternate pits per cross-field) is unique among others species of Agathoxylon recorded in Jurassic and Cretaceous strata from Western Gondwana, allowing the creation of a new fossil species, Agathoxylon mendezii sp. nov. In addition, the Patagonian taxon appears to differ from the wood of extant Agathis and Wollemia, showing a close affinity with Araucaria. However, the combination of features in Agathoxylon mendezii sp. nov. cannot be accommodated within any of the four sections of Araucaria, namely Araucaria, Bunya, Eutacta, or Intermedia. Among these sections, only Eutacta is represented in the Springhill Formation strata by bract-scale complexes. With the exception of Intermedia, the other three sections of Araucaria were well established in the Jurassic and Cretaceous deposits of Patagonia and documented by vegetative and reproductive remains. Agathoxylon mendezii sp. nov. is the first araucarian wood to be recorded in the Springhill Formation, increasing the number of taxa within the Araucariaceae family and revealing that this family was more diverse in Patagonia during the Mesozoic than it is today. In Patagonia, Araucariaceae currently consists of solely Araucaria araucana of the Araucaria section.
At the time of the deposition of the Springhill Formation, the trunk of Agathoxylon mendezii sp. nov. reached 0.72 m in diameter indicating that it was a well-developed tree. The Patagonian wood exhibits slightly marked growth rings, suggesting that it inhabited a seasonless palaeoenvironment. Although growth rings in araucarian woods are believed to be independent of environmental influence, this is not the case for Agathoxylon mendezii sp. nov. The ring patterns in this taxon are correlated with sedimentological as well as palaeofloristic evidence that reveal the existence of a warm and wet climate during the deposition of the Springhill Formation in the Estancia El Álamo locality. This reflects a subtropical environment with abundant available water in an alluvial-fan palaeoenvironment.
The presence of numerous frost rings in the wood of Agathoxylon mendezii sp. nov. suggests that the growing conditions were most stressed during the Berriasian–Valanginian at the Estancia El Álamo locality. In agreement with sedimentological evidence, the palaeoenvironment at this locality is interpreted to have been subjected to recurrent volcanic activity produced by a volcanic arc located far away to the west. The repeated volcanic eruptions may have promoted abrupt decreases in the air temperature below the freezing point, consequently producing frost injuries to the cambium that lead to the formation of multiple (at least five) frost rings in Agathoxylon mendezii sp. nov.
Usually, frequent frost rings related to the cooling effect of volcanic eruptions are reported in extant conifers of the genus Pinus. However, this is the first time that numerous frost rings caused by regional recurrent volcanic eruptions are recorded in a Lower Cretaceous coniferous wood from Patagonia and not in other Mesozoic coniferous woods from Western Gondwana.
Frost rings in extant araucarian wood are used to build reliable frost ring chronologies to document past extreme climatic events in areas from where the taxon originates. Thus, the occurrences of frost damage in the Agathoxylon wood described herein would probably be a useful tool for researching extreme palaeoclimates, particularly during the Early Cretaceous in Patagonia.
Acknowledgements
We are grateful to Ignacio Méndez, owner of the Estancia El Álamo, for letting us to explore and collect the fossil in his property. We would like to thank Fidel A. Roig (Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales, CCT CONICET-Mendoza, Departamento de Dendrocronología e Historia Ambiental, Mendoza, Argentina) for valuable discussions about traumatic tissues in woods and Silvia Gnaedinger (Centro de Ecología Aplicada del Litoral, Área de Paleontología, Facultad de Ciencias Exactas y Naturales y Agrimensura, Universidad Nacional del Nordeste, CONICET, Corrientes, Argentina) for constructive comments on the manuscript and for providing bibliography during the coronavirus pandemic. We thank Orlando Cárdenas (Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, CONICET, Buenos Aires, Argentina) for his guidance and help during the fieldtrip and for technical assistance in SEM samples processing. The authors would like to thank the reviewers Marion Bamford (Bernard Price Institute for Palaeontological Research, University of the Witwatersrand, Johannesburg, South Africa) and Marc Philippe (Laboratoire d’écologie de hydrosystèmes naturels et anthropisés, Claude Bernard University Lyon 1, France) for their useful comments that have greatly improved the manuscript. This article was supported in part by grants ANPCyT PICT 2012/528, CONICET PIP 112-201201-00212, and CONICET PIP 1016.
References
Aguirre Urreta, M.B. 2002. Invertebrados del Cretácico Inferior. In: M.J. Haller (ed.), Geología y Recursos Naturales de Santa Cruz. Proceedings of the 15th Congreso Geológico Argentino. El Calafate, Argentina 2002, 439–459. Talleres Trama, Buenos Aires.
Aguirre Urreta, B., Suárez, M., De la Cruz, R., and Ramos, V.A. 2007. Ammonoids Crioceratitinae, Hauterivian) from the Austral Basin. Ameghiniana 44: 387–397.
Aramendía, I., Cuitiño, J.I., Ghiglione, M.C., and Bouza, P.J. 2019. Tectonostratigraphic significance of the Neogene sedimentary record of the northwestern Austral-Magallanes Basin, Argentinean Patagonia. Latin American Journal of Sedimentology and Basin Analysis 26: 99–126.
Aramendía, I., Ramos, M.E., Geuna, S., Cuitiño, J.I., and Ghiglione, M.C. 2018. A multidisciplinary study of the Lower Cretaceous marine to continental transition in the northern Austral-Magallanes basin and its geodynamic significance. Journal of South American Earth Sciences 86: 54–69. Crossref
Arbe, H.A. 2002. Análisis estratigráfico del Cretácico de la Cuenca Austral. In: M.J. Haller (ed.), Geología y Recursos Naturales de Santa Cruz. Proceedings of the 15th Congreso Geológico Argentino. El Calafate, Argentina 2002, 407–420. Talleres Trama, Buenos Aires.
Archangelsky, S. 1976. Vegetales fósiles de la Formación Springhill, Cretácico, en el subsuelo de la Cuenca Magallánica, Chile. Ameghiniana 13: 141–158.
Archangelsky, S. 2001a. Evidences of an Early Cretaceous floristic change in Patagonia, Argentina. VII International Symposium on Mesozoic Terrestial Ecosystems. Argentine Paleontological Association Special Publication 7: 15–19.
Archangelsky, S. 2001b. The Ticó Flora (Patagonia) and the Apitan Extinction Event. Acta Palaeobotanica 41: 115–122.
Archangelsky, S. and Archanglelsky, A. 2004. Palinología estadística en el Cretácico de la Cuenca Austral, Plataforma Continental Argentina. II. Seis perforaciones del área Gallegos. 3. Discusión y conclusiones. Revista del Museo Argentino de Ciencias Naturales (Nueva Serie) 6: 245–255. Crossref
Archangelsky, A., Andreis, R.R., Archangelsky, S., and Artabe, A. 1995. Cuticular characters adapted to volcanic stress in a new Cretaceous cycad leaf from Patagonia, Argentina. Considerations on the stratigraphy and depositional history of the Baqueró Formation. Review of Palaeobotany and Palynology 89: 213–233. Crossref
Archangelsky, S., Baldoni, A., Gamerro, J.C., Palamarczuk, S., and Seiler, J. 1981. Palinología estratigráfica del Cretácico de Argentina Austral. Diagramas de grupos polínicos del suroeste de Chubut y noroeste de Santa Cruz. In: VIII Congreso Geológico Argentino, Actas, 719–742. Buenos Aires.
Arco Molina, J.G., Hadad, M.A., Patón Domínguez D., and Roig F.A. 2016. Tree age and bark thickness as traits linked to frost ring probability on Araucaria araucana trees in northern Patagonia. Dendrochronologia 37: 116–125. Crossref
Bailey, I.W. 1925. The “Spruce Budworm” Biocoenose. 1: Frost rings as indicators of the chronology of specific biological events. Botanical Gazette 80: 93–101. Crossref
Baldoni, A.M. 1977. Ptilophyllum ghiense n. sp., una nueva bennettital de Paso Roballos, provincia de Santa Cruz. Ameghiniana 14: 53–58.
Baldoni, A.M. 1979. Nuevos elementos paleofloristicos de la tafoflora de la Formación Spring Hill, limite Jurásico–Cretácico subsuelo de Argentina y Chile Austral. Ameghiniana 16: 103–119.
Baldoni, A.M. 1980. Nota sobre una nueva especie de Elatocladus (?) (Coniferae) de la Formación Springhill, límite Jurásico–Cretácico, Provincia de Santa Cruz. Ameghiniana 17: 373–378.
Baldoni, A.M. and Archangelsky, S. 1983. Palinología de la Formación Springhill (Cretácico Inferior), subsuelo de Argentina y Chile Austral. Revista Española de Micropaleontología 15: 47–101.
Baldoni, A.M. and Taylor, T.N. 1983. Plant remains from a new Cretaceous site in Santa Cruz, Argentina. Review of Palaeobotany and Palynology 39: 301–311. Crossref
Bamford, M.K. and Corbett, I.B., 1994. Fossil wood of Cretaceous age from the Namaqualand Continental Shelf, South Africa. Palaeontographica Africana 31: 83–95.
Bamford, M. and Philippe, M. 2001. Jurassic–Early Cretaceous Gondwanan homoxylous woods: a nomenclatural revision of the genera with taxonomic notes. Review of Palaeobotany and Palynology 113: 287–297. Crossref
Barberón, V., Ronda, G., Aramendía, I., Suárez, R.J., Ramos, M.E., Naipauer, M., Sue, C., and Ghiglione M.C. 2019. Tectonic evolution of the northern Austral-Magallanes basin in the Southern Patagonian Andes from provenance analysis. Journal of South American Earth Sciences 95: 102234. Crossref
Biddle, K., Uliana, M., Mitchum, R., Fitzgerald, M., and Wright, R. 1986. The stratigraphic and structural evolution of central and eastern Magallanes Basin, Southern America. In: P. Allen and P. Homewood (eds.), Foreland Basins. International Association of Sedimentology Special Publication 8: 41–61. Crossref
Bodnar, J., Escapa, I., Cúneo, N.R., and Gnaedinger, S. 2013. First record of conifer wood from the Cañadón Asfalto Formation (early–Middle Jurassic), Chubut Province, Argentina. Ameghiniana 50: 227–239. Crossref
Brea, M. 1997. Una nueva especie fósil del género Araucarioxylon Kraus 1870, emend. Maheshwari 1972 del Triásico de Agua de la Zorra, Uspallata, Mendoza, Argentina. Ameghiniana 34: 485–496.
Brea, M., Bellosi, E.S., Umazano, A.M., and Krause, J.M. 2016. Aptian–Albian Cupressaceae (sensu stricto) woods from Cañadón Asfalto Basin, Patagonia Argentina. Cretaceous Research 58: 17–28. Crossref
Brison, A., Philippe, M., and Thévenard, F. 2001. Are Mesozoic wood growth rings climate-induced? Paleobiology 27: 531–538. Crossref
Carrizo, M.A. 2014. Morfología, anatomía y sistemática de la Paleoflora Cretácica de la Formación Springhill, Patagonia Austral, Argentina. 269 pp. Ph.D. Thesis, Buenos Aires University, Buenos Aires.
Carrizo, M.A. and Del Fueyo, G.M. 2015. The Early Cretaceous Megaflora of the Springhill Formation, Patagonia. Paleofloristic and Paleonvironmental Inferences. Cretaceous Research 56: 93–109. Crossref
Carrizo, M.A., Del Fueyo, G.M., and Archangelsky, S. 2011. Morfología y anatomía de un helecho creciendo bajo condiciones de estrés en el Aptiano de Santa Cruz, Argentina. Ameghinina 48: 605–617. Crossref
Carrizo, M.A, Del Fueyo, G.M., and Medina, F. 2014. Foliar cuticle of Ruflorinia orlandoi nov. sp (Pteridospermophyta) from the Lower Cretaceous of Patagonia. Geobios 47: 87–99. Crossref
Cortiñas, J.C. and Arbe, H.A. 1981. Un nuevo afloramiento fosilífero de la Formación Springhill, en el Noroeste de la provincia de Santa Cruz. Revista de la Asociación Geológica Argentina 36: 212–214.
Creber, G.T. and Chaloner, W.G. 1984. Influence of environmental factors on the wood structure of living and fossil trees. Botanical Review 50: 357–448 Crossref
Cuitiño, J.I., Varela, A.N., Ghiglione, M.C., Richiano, S., and Poiré, D.G. 2019. The Austral-Magallanes Basin (Southern Patagonia): a synthesis of its stratigraphy and evolution. Latin American Journal of Sedimentology and Basin Analysis 26: 155–166.
Del Fueyo, G.M., Gnaedinger, S.C., Lafuente Diaz, M.A., and Carrizo, M.A. 2019. Permineralized conifer-like leaves from the Jurassic of Patagonia, Argentina. Annals of the Brazilian Academy of Sciences 91 (2): 1–19. Crossref
Del Fueyo, G.M., Villar de Seoane, L., Archangelsky, A., Guler, V., Llorens, M., Archangelsky, S., Gamerro, J.C., Musacchio, E.A., Passalia, M.G., and Barreda, V.D. 2007. Biodiversidad de las Paleofloras de Patagonia Austral durante el Cretácico Inferior. Ameghiniana 50° Aniversario. Argentine Paleontological Association Special Publication 11: 101–122.
Escapa, I.H. and Catalano, S.A. 2013. Phylogenetic analysis of Araucariaceae: Integrating molecules, morphology, and fossils. International Journal of Plant Sciences 174: 1153–1170. Crossref
Falaschi, P., Grosfeld, J., Zamuner, A.B., Foix, N., and Rivera, S.M. 2011. Growth architecture and silhouette of Jurassic conifers from La Matilde Formation, Patagonia, Argentina. Palaeogeography Palaeoclimatology Palaeoecology 302: 122–141. Crossref
Falcon-Lang, H.J. 2000. The relationship between leaf longevity and growth ring markedness in modern conifer woods and its implications for palaeoclimatic studies. Palaeogeography, Palaeoclimatology, Palaeoecology 160: 317–328. Crossref
Falcon-Lang, H.J. and Cantrill, D.J. 2001. Gymnosperm woods from the Cretaceous (mid-Aptian) Cerro Negro Formation, Byres Peninsula, Livingston Island, Antarctica: the arborescent vegetation of a volcanic arc. Cretaceous Research 22: 277–293. Crossref
Filion, L., Payette, S., Gauthier, L., and Boutin, Y. 1986. Light rings in subarctic conifers as a dendrochronological tool. Quaternary Research 26: 272–279. Crossref
Fletcher, T.L., Moss, P.T., and Salisbury, S.W. 2015. Wood growth indices as climate indicators from the Upper Cretaceous (Cenomanian–Turonian) portion of the Winton Formation, Australia. Palaeogeography, Palaeoclimatology, Palaeoecology 417: 35–43. Crossref
Föllmi, K.B. 2012. Early Cretaceous life, climate and anoxia. Cretaceous Research 35: 230–257. Crossref
Frakes, L.A. 1999. Estimating the global thermal state from Cretaceous sea surface and continental temperature data. In: E. Barrera and C.C. Johnson (eds.), Evolution of the Cretaceous Ocean-Climate System. Geological Society of America, Special Paper 332: 49–57. Crossref
Galeazzi, J.S. 1998. Structural and stratigraphic evolution of the western Malvinas Basin, Argentina. American Association of Petroleum Geologists Bulletin 82: 594–636.
Gallardo Jara, R.E., Ghiglione, M.C., and Rojas Galliani, L. 2019. Tectonic evolution of the southern Austral-Magallanes Basin in Tierra del Fuego. Latin American Journal of Sedimentology and Basin Analysis 26: 127–154.
Ghiglione, M.C., Naipauer, M., Sue, C., Barberón, V., Valencia, V., Aguirre Urreta, B., and Ramos, A. 2015. U-Pb zircon ages from the northern Austral basin and their correlation with the Early Cretaceous exhumation and volcanism of Patagonia. Cretaceous Research 55: 116–128. Crossref
Ghiglione, M.C., Ramos, V.A., Cuitiño, J.I., and Barberón, V. 2016. Growth of the Southern Patagonian Andes (46–53°S) and their relation to subduction processes. In: A. Folguera, M. Naipauer, L. Sagripanti, M. Ghiglione, D. Orts, and L. Giambiagi (eds.), Growth of the Southern Andes, 201–218. Springer Earth System Sciences, Springer International Publishing, Zurich.
Glerum, C. and Farrar, J.L. 1966. Frost ring formation in the stems of some coniferous species. Canadian Journal of Botany 44: 879–886. Crossref
Gnaedinger, S. and Herbst, R. 2009. Primer registro de maderas gimnospérmicas de la Formación Roca Blanca (Jurásico Inferior), provincia de Santa Cruz, Argentina. Ameghiniana 46: 59–71.
Gnaedinger, S., García Massini, J.L., Bechis, F., and Zavattieri A.M. 2015. Coniferous woods and wood-decaying fungi from the El Freno Formation (Lower Jurassic), Neuquén Basin, Mendoza province, Argentina. Ameghiniana 52: 447–467. Crossref
Gómez Dacal, A.R., Gómez Peral, L.E., Spalletti, L.A., Sial, A.N., and Poiré, D.G. 2019. Evidence of warm seas in high latitudes of southern South America during the early Cretaceous. Cretaceous Research 95: 8–20.
Gómez Dacal, A.R., Gómez Peral, L.E., Spalletti, L.A., Sial, A.N., Siccardi, A., and Poiré, D.G. 2018. First record of the Valanginian positive carbon isotope anomaly in the Mendoza shelf, Neuquén Basin, Argentina: palaeoclimatic implications. Andean Geology 45: 111–129. Crossref
González Estebenet, M.S., Paolillo, M.A., and Guler, M.V. 2019. Marine Cretaceous organic-walled dinoflagellate cysts from the Austral-Magallanes Basin. Latin American Journal of Sedimentology and Basin Analysis 26: 75–98.
Greguss, P. 1955. Identification of Living Gymnosperms on the Basis of Xylotomy. 263 pp. Akadémiai Kiadó, Budapest.
Greguss, P. 1972. Xylotomy of Living Conifers. 329 pp. Akadémiai Kiadó, Budapest.
Greppi, C.D., Pujana, R.R., and Scasso, R.A. 2020. Fossil woods from the Lower Cretaceous Tres Lagunas Formation of central Patagonia (Chubut Province, Argentina). Cretaceous Research 108: 104322. Crossref
Guler, V., Berbach, L., Archangelsky, A., and Archangelsky, S. 2015. Quistes de dinoflagelados y polen asociado del Cretácico Inferior (Formación Springhill) de la Cuenca Austral, Plataforma Continental Argentina. Revista Brasileira de Paleontologia 18: 307–324. Crossref
Gurskaya, M.A. and Shiyatov, S.G. 2002. Formation of two xylem frost injuries in one annual ring in Siberian Spruce under conditions of Western Siberian Forest-Tundra. Russian Journal of Ecology 33: 73–79. Crossref
Hadad, M.A., Amoroso, M.M., and Roig, F.A. 2012. Frost ring distribution in Araucaria araucana trees from the xeric forests of Patagonia, Argentina. Bosque 33: 309–312. Crossref
Hadad, M.A., Arco Molina, J., Roig Juñent, F.A., Amoroso, M.M., Müller, G., Araneo, D., and Tardif, J.C. 2019. Frost record in tree rings linked to atmospheric circulation in northern Patagonia. Palaeogeography, Palaeoclimatology, Palaeoecology 524: 201–211. Crossref
Hantemirov, S.M., Gorlanova, L.A., and Shiyatov, S.G. 2000. Pathological Tree-Ring Structures in Siberian Juniper (Juniperus sibirica Burgsd.) and their use for reconstructing of extreme climatic events. Russian Journal of Ecology 31: 167–173. Crossref
Hantemirov, R.M., Gorlanova, L.A, and Shiyatov, S.G. 2004. Extreme temperature events in summer in northwest Siberia since AD 742 inferred from tree rings. Palaeogeography, Palaeoclimatology, Palaeoecology 209: 155–164. Crossref
Hartig, A. 1848. Beiträge zur Geschichte der Pflanzen und zur Kenntnis der norddeutschen Braunkohlen-Flora. Botanische Zeitung 6; 185–190.
Hay, W.W. and Floegel, S. 2012. New thoughts about the Cretaceous climate and oceans. Earth-Science Reviews 115: 262–272. Crossref
Heady, R.D., Banks, J.G., and Evans, P.D. 2002. Wood anatomy of Wollemi pine (Wollemia nobilis, Araucariaceae). IAWA Journal 23: 339–357. Crossref
Horton, B.K. 2018. Sedimentary record of Andean mountain building. Earth-Science Review 178: 279–309. Crossref
IAWA Committee 2004. International Association of Wood Anatomists list of microscopic features for softwood identification. IAWA Journal 25: 1–70. Crossref
Iñiguez Rodríguez, A.M. and Decastelli, O. 1984. Mineralogía y diagénesis de arcillas de las formaciones cretácicas-terciarias de la Cuenca Austral. In: Actas del IX Congreso Geológico Argentino, Vol. 3, 402–414. San Carlos de Bariloche.
Kauffman, E.G. and Johnson, C.C. 2009. Cretaceous warm climates. In: V. Gornitz (ed.), Encyclopedia of Paleoclimatology and Ancient Environments, 213–206. Springer, Dordrecht. Crossref
Kay, S.M., Mpodozis, C., and Ramos, V.A. 2005. Andes. In: R.C Selley, L.R.M. Cocks, and I.R. Pilmer (eds.), Encyclopedia of Geology, 118–131. Elsevier, Amsterdam. Crossref
Kloster, A.C. and Gnaedinger, S.C. 2018. Coniferous wood of Agathoxylon from the La Matilde Formation, (Middle Jurassic), Santa Cruz, Argentina. Journal of Palaeontology 92: 546–567. Crossref
Kraemer, P.E. and Riccardi, A.C. 1997. Estratigrafía de la región comprendida entre los lagos Argentino y Viedma (49°40’–50°10’ LS), Provincia de Santa Cruz. Revista de la Asociación Geológica Argentina 52: 333–360.
LaMarche, V.C. and Hirschboeck, K.K. 1984. Frost rings in trees as records of major volcanic eruptions. Nature 307: 121–126. Crossref
Leslie, A., Beaulieu, J., Holman, G., Campbell, C.S., Mei, W., Raubeson, L.R., and Mathews, S. 2018. An overview of extant conifer evolution from the perspective of the fossil record. American Journal of Botany 105 (9): 1–14. Crossref
Lucas, R.C. and Lacey, W.S. 1981. A permineralized wood flora of probable early Tertiary from King George Island, South Shetland Islands. British Antarctic Survey Bulletin 53: 147–151.
Martínez, L.C.A., Archangelsky, S., Prámparo, M.B., and Archangelsky, A. 2016. Early Cretaceous palm pollen tetrads from Patagonia, Argentina. Cretaceous Research 59: 129–139. Crossref
Mutterlose, J., Malkoc, M., Schouten, S., Sinninghe Damsté, J.S., and Forster, A. 2010. TEX86 and stable 18O paleothermometry of Early Cretaceous sediments: Implications for belemnite ecology and paleotemperature proxy application. Earth and Planetary Science Letters 298: 286–298. Crossref
Nishida, M. and Nishida, H. 1987. Petrified woods from the Upper Cretaceous of Quiriquina Island, near Concepción, Chile. In: M. Nishida (ed.), Contribution to the Botany in the Andes 2: 5–11. Academic Scientific Book Inc., Tokyo.
Nishida, M., Ohsawa, T., Nishida, H., and Rancusi, M.H. 1992. Permineralized coniferous woods from the XI Region of Chile, Central Patagonia. Research Institute of Evolutionary Biology, Scientifical Report 7: 47–59.
Nullo, F.E., Panza, J.L., and Blasco, G. 1999. Jurásico y Cretácico de la Cuenca Austral. In: R. Caminos (ed.), Geología Argentina. Anales del Servicio Geológico Minero Argentino 29: 528–535.
Ottone, E.G. and Aguirre Urreta, M.B. 2000. Palinomorfos cretácicos de la Formación Springhill en Estancia El Salitral, Patagonia Austral, Argentina. Ameghiniana 37: 379–382.
Palamarczuk, S., Gamerro, J.C., and Barreda, V. 2000. Estudio palinológico en el pozo Chiton MFJ8 x-1, plataforma continental argentina, Cuenca Austral. 11º Simposio Argentino de Paleobotánica y Palinología, Tucumán, Argentina. Suplemento Resúmenes Ameghiniana 37: 59R.
Pankhurst, R.J., Riley, T.R., Fanning, C.M., and Kelley, S.P. 2000. Episodic silicic volcanism in Patagonia and the Antarctic Peninsula: chronology of magmatism associated with the break-up of Gondwana. Journal of Petrology 41: 605–625. Crossref
Panti, C., Pujana, R.R., Zamaloa, M.C., and Romero, E.J. 2011. Araucariaceae macrofossil record from South America and Antarctica. Alcheringa 36:1–22. Crossref
Passalia, M.G. 2009. Cretaceous pCO2 estimation from stomatal frequency analysis of gymnosperm leaves of Patagonia, Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 273: 17–24. Crossref
Philippe, M. 1993. Nomenclature générique des trachéidoxyles fossiles mésozoïques à champs araucarioïdes. Taxon 42: 74–80. Crossref
Philippe, M. 1995. Bois fossiles du Jurassique de Franche-Comté (nord-est de la France): systématique et biogéographie. Palaeontographica B 236: 45–103.
Philippe, M. 2011. How many species of Araucarioxylon? Comptes Rendus Palevol 10: 201–208. Crossref
Philippe, M. and Bamford, M.K. 2008. A key to morphogenera used for Mesozoic conifer-like woods. Review of Palaeobotany and Palynology 148: 184–207. Crossref
Philippe, M., Bamford, M., McLoughlin, S., Alves, L.S.R., Falcon-Lang, H.J., Gnaedinger, S., Ottone, E.G., Pole, M., Rajanikanth, A., Shoemaker, R.E., Torres, T., and Zamuner, A. 2004. Biogeographic analysis of Jurassic–Early Cretaceous wood assemblages from Gondwana. Review of Palaeobotany and Palynology 129: 141–173. Crossref
Philippe, M., Pacyna, G., Wawrzyniak, Z., Barbacka, M. Boka, K., Filipiak, P. Marynowski, L., Thévenard, F., and Uhl, D. 2015. News from an old wood Agathoxylon keuperianum (Unger) nov. comb. in the Keuper of Poland and France. Review of Palaeobotany and Palynology 221: 3–91. Crossref
Poiré, D.G. and Franzese, J.R. 2010. Mesozoic clastic sequences from a Jurassic rift to Cretaceous foreland basin, Austral Basin, Patagonia, Argentina. In: C. del Papa and R. Astini (eds.), 18th International Sedimentological Congress, Mendoza, Argentina 2010 Field Excursion Guidebook, FE-C13, 1–53. Editorial IAS, Buenos Aires.
Poiré, D.G., Iglesias, A., Varela, A.N., Richiano, S., Ibañez Mejías, M., and Strömberg, C.A.E. 2017. Edades U-Pb en zircones de tobas de la Fm. Piedra Clavada, Pcia. de Santa Cruz, Argentina: Un marcador albiano tardio para la evolución tectónica y biológica de la Cuenca Austral. In: Actas del XX Congreso Geológico Argentino, 95–98. San Miguel de Tucumán.
Poole, I. and Cantrill, D.J. 2001. Fossil woods from Williams Point beds, Livingston Island, Antarctica: a late Cretaceous southern high latitude flora. Palaeontology 44: 1081–1112. Crossref
Price, G.D. and Gröcke, D.R. 2002. Strontium-isotope stratigraphy and oxygen- and carbon-isotope variation during the Middle Jurassic–Early Cretaceous of the Falkland Plateau, South Atlantic. Palaeogeography, Palaeoclimatology, Palaeoecology 183: 209–222. Crossref
Pujana, R., Santillana, S., and Marenssi, S. 2014. Conifer fossil woods from the La Meseta Formation (Eocene of Western Antarctica): Evidence of Podocarpaceae-dominated forests. Review of Palaeobotany and Palynology 200: 122–137. Crossref
Quattrocchio, M.E., Martínez, M.A., and Volkheimer, W. 2007. Las floras jurásicas de la Argentina. Ameghiniana 50° Aniversario of the Argentine Paleontological Association, Special Publication 11: 87–100.
Quattrocchio, M.E., Martínez, M.A., Pavisich, A.C., and Volkheimer, W. 2006. Early Cretaceous palynostratigraphy, palynofacies and palaeoenvironments of well sections in northeastern Tierra del Fuego, Argentina. Cretaceous Research 27: 584–602. Crossref
Rampino, M.R. and Self, S. 1982. Historic eruptions of Tambora (1815), Krakatau (1883), and Agung (1963), their stratospheric aerosols, and climatic impact. Quaternay Research 18: 127–143. Crossref
Riccardi, A.C. 1971. Estratigrafía en el oriente de la Bahía de la Lancha, Lago San Martín, Santa Cruz, Argentina. Revista del Museo de La Plata. Geología (Nueva Serie) 7: 245–318.
Riccardi, A.C. 1976. Paleontología y edad de la Formación Springhill. In: Actas del Primer Congreso Geológico Chileno, Santiago, Chile, C41–C56. Universidad de Chile, Santiago.
Riccardi, A.C. 1977. Berriasian invertebrate fauna from the Springhill Formation of Southern Patagonia. Neues Jahrbuch für Geologie und Paläontologie 155: 216–252.
Riccardi, A.C. 1988. The Cretaceous System of southern South America. Memoir of the Geological Society of America 168: 1–161. Crossref
Riccardi, A.C. and Rolleri, E.O. 1980. Cordillera Patagónica Austral. In: J.C.M. Turner (ed.), Segundo Simposio de Geología Regional Argentina, Córdoba 1980, 1173–1306. Academia Nacional de Ciencias de Córdoba, Córdoba.
Riccardi, A.C., Gulisano, C.A., Mojica, J., Palacios, O., Schubert, C., and Thomson, M.R.A. 1992. Western South America and Antarctica. In: G.E.G. Westermann (ed.), The Jurassic of the Circum-Pacific, 122–161. Cambridge University Press, Cambridge. Crossref
Richiano, S., Poiré, D.G., and Varela, A.N. 2013. Icnología del Cretácico inferior en la Formación Río Mayer, SW Gondwana, Patagonia, Argentina. Ameghiniana 50: 273–286. Crossref
Richiano, S., Varela, A.N., and Poiré, D.G. 2016. Heterogeneous distribution of trace fossils across initial transgressive deposits in rift basins: an example from the Springhill Formation, Argentina. Lethaia 49: 524–539. Crossref
Richiano, S., Varela, A.N., Cereceda, A., and Poiré, D.G. 2012. Evolución paleoambiental de la Formación Río Mayer, Cretácico inferior, Cuenca Austral, provincia de Santa Cruz, Argentina. Latin American Journal of Sedimentology and Basin Analysis 19: 3–26.
Richiano, S., Varela, A.N., Gómez Peral, L.E., Cereceda, A., and Poiré, D.G. 2015. Composition of the Lower Cretaceous source rock from the Austral Basin (Río Mayer Formation, Patagonia, Argentina): Regional implication for unconventional reservoirs in the Southern Andes. Marine and Petroleum Geology 66: 764–790. Crossref
Riggi, J.C. 1957. Resumen geológico de la zona de los lagos Pueyrredón y Posadas, provincia de Santa Cruz. Revista de la Asociación Geológica Argentina 12: 65–97.
Rößler, R., Philippe, M., Van Konijnenburg-van Cittert, J.H.A., McLoughlin, S., Sakala, J., and Zijlstra, G. 2014. Which name(s) should be used for Araucaria-like fossil wood?—Results of a poll. Taxon 63: 177–184. Crossref
Sachse, V.F., Strozyk, F., Anka, Z., Rodriguez, J.F., and di Primio, R. 2016. The tectono-stratigraphic evolution of the Austral basin and adjacent areas against the back-ground of Andean tectonics, southern Argentina, South America. Basin Research 28: 462–482. Crossref
Salzer, M.W. and Hughes, K. 2007 Bristlecone pine tree rings and volcanic eruptions over the last 5000 yr. Quaternary Research 67: 57–68. Crossref
Sanchez, J.A. and Le Roux, J.P. 2003. Mesozoic sedimentation on an isolated platform at the eastern entrance to the Strait of Magellan, Tierra del Fuego (Chile). Andean Geology 30: 143–157. Crossref
Schwarz, E., Veiga, G.D., Spalletti, L.A., and Massaferro, J.L. 2011. The transgressive infill of an inherited-valley system: The Springhill Formation (lower Cretaceous) in southern Austral Basin, Argentina. Marine and Petroleum Geology 28: 1218–1241. Crossref
Schweingruber, F.H. 1989. Tree Rings. Basics and Applications of Dendrochronology. 276 pp. Kluwer Academic Publishers, Dordrecht.
Schweingruber, F.H. 2007. Wood Structure and Environment. 279 pp. Springer, Berlin.
Schweingruber, F.H., Börner, A., and Schulze, E.D. 2006. Atlas of Woody Plant Stems. Evolution, Structure, and Environmental Modifications. 229 pp. Springer, Berlin.
Self, S.J, Rampino, M.R., and Barbera, J.J. 1981. The possible effects of large 19th and 20th century volcanic eruptions on zonal and hemispheric surface temperatures. Journal of Volcanology and Geothermal Research 1: 41–60. Crossref
Spalletti, L.A., Matheos, S.D., Sánchez, E., and Oyarzábal, F. 2006. Análisis diagenético de la Formación Springhill (Santa Cruz, Argentina). Boletín de Informaciones Petroleras (Nueva Serie) 4: 60–73.
Spalletti, L.A., Schwarz, E., Veiga, G.D., Matheos, S.D., Haring, C., and Covellone, G. 2009. Análisis paleoambiental y estratigráfico secuencial de alta resolución de la Formación Springhill al este de Tierra del Fuego (Cuenca Austral, Argentina). In: Actas del XII Congreso Geolólogico Chileno, S10_037. Universidad de Chile, Santiago.
Stahle, D.W. 1990. Tree-Ring Record of False Spring in the South Central USA. 272 pp. Ph.D. Thesis, Arizona State University, Tempe.
Stockey, R.A. 1978. Reproductive biology of Cerro Cuadrado fossil conifers: ontogeny and reproductive strategies in Araucaria mirabilis (Spegazzini) Windhausen. Palaeontographica B 166: 1–15.
Stothers, R.B. 2009. Volcanic eruptions and climate change. In: V. Gornitz (ed.), Encyclopedia of Paleoclimatology and Ancient Environments, 976–979. Springer, Dordrecht. Crossref
Taylor, T.N., Taylor, E.L., and Krings, M. 2009. Paleobotany, the Biology and Evolution of Fossil Plants. 1253 pp. Academic Press, London.
Thomas, C.R. 1949. Geology and petroleum exploration in Magallanes Province, Chile. American Association of Petroleum Geologists Bulletin 33: 1553–1578. Crossref
Torres, T. and Biro-Bagoczky, L. 1986. Xilotomía de coníferas fósiles de la Isla Quiriquina, Chile. Comunicaciones 37: 65–80.
Torres, T. and Lemoigne, Y. 1989. Hallazgos de maderas fósiles de Angiospermas y Gimnospermas del Cretácico Superior en punta Williams, isla Livingston, islas Shetland del Sur, Antártida. Serie Científica del Instituto Antártico Chileno 39: 9–29.
Torres, T. and Philippe, M. 2002. Nuevas especies de Agathoxylon y Baieroxylon del Lías de La Ligua (Chile) con una evaluación del registro paleoxilológico en el Jurásico de Sudamérica. Revista Geológica de Chile 29: 151–165. Crossref
Torres, T. and Rallo, M. 1981. Anatomía de troncos fósiles del Cretácico superior de Pichasca, en el Norte de Chile. In: Actas del II Congreso Latino-Americano de Paleontología, 385–398. Porto Alegre.
Torres, T., Calderón, M., and Fadel Cury, L. 2009. Maderas fósiles de Gimnospermas en la Formación Yahgan, estratos de Tekenika, Isla Hoste, Chile. In: Actas del XII Congreso Geológico Chileno, S12_020. Universidad de Chile, Santiago.
Torres, T., Galleguillos, H., and Philippe, M. 2000. Maderas fósiles en el Monte Flora, Bahía Esperanza, Península Antartica. In: Actas del IX Congreso Geológico Chileno, 386–390. Puerto Varas.
Torres, T., Valenzuela, E., and Gonzalez, I. 1982. Paleoxilología de Península Byers, Isla Livingston, Antartica. In: Actas del III Congreso Geológico Chileno, Vol. 2, 321–342. Concepción.
Tortorelli, L.A. 1956. Maderas y bosques argentinos. 910 pp. Editorial Acme, Buenos Aires.
Vakhrameev, V.A. 1991. Jurassic and Cretaceous Floras and Climates of the Earth. 381 pp. Cambridge University Press, Cambridge.
Varela, A.N., Gómez Peral, L.E., Richiano, S., and Poiré, D.G. 2013. Distinguishing similar volcanic source areas from an integrated provenance analysis: Implications for foreland andean basins. Journal of Sedimentary Research 83: 258–276. Crossref
Varela, A.N., Poiré, D.G., Martin, T., Gerdes, A., Goin, F.J., Gelfo, J.N., and Hoffmann, S. 2012. U-Pb zircon constraints on the age of the Cretaceous Mata Amarilla Formation, Southern Patagonia, Argentina: its relationship with the evolution of the Austral Basin. Andean Geology 39: 359–379. Crossref
Ventra, D. and Clarke, L.E. 2018. Geology and geomorphology of alluvial and fluvial fans: current progress and research perspectives. In: D. Ventra and L.E. Clarke (eds.), Geology and Geomorphology of Alluvial and Fluvial Fans: Terrestrial and Planetary Perspectives. Geological Society, London, Special Publication 440: 1–21. Crossref
Vera, E.I. and Césari, S.N. 2015. New species of conifer wood from the Baqueró Group (early Cretaceous) of Patagonia. Ameghiniana 52: 468–471. Crossref
Vera, E.I., Perez, V.S., Llorens, M., and Passalia, M.G. 2020. Agathoxylon Hartig in the Lower Cretaceous Arroyo del Pajarito Member (Los Adobes Formation), Chubut Province, Argentina. Journal of South American Earth Sciences 100: 102562. Crossref
Villar de Seoane, L. 1995. Estudio cuticular de nuevas Bennettitales eocretácicas de Santa Cruz, Argentina. In: Actas del 6to Congreso Argentino de Paleontología y Bioestratigrafía, 247–254. Trelew.
Villar de Seoane, L. 2001. Cuticular study of Bennettitales from the Springhill Formation, Lower Cretaceous of Patagonia, Argentina. Cretaceous Research 22: 461–479. Crossref
Wilf, P., Escapa, I.H., Cuneo, N.R., Kooyman, R.M., Johnson, K.R., and Iglesias, A. 2014. First South American Agathis (Araucariaceae), Eocene of Patagonia. American Journal of Botany 101: 156–179. Crossref
Zamuner, A.B. and Falaschi, P. 2005. Agathoxylon matildense n. sp., leño araucariaceo del Bosque Petrificado del cerro Madre e Hija, Formación La Matilde (Jurásico medio), provincia de Santa Cruz, Argentina. Ameghiniana 42: 339–346.
Zanchetta, G., Sulpizio R., and Di Vito, M.A. 2004. The role of volcanic activity and climate in alluvial fan growth at volcanic areas: an example from southern Campania (Italy). Sedimentary Geology 168: 249–280. Crossref
Zeebe, R.E. 2001. Seawater pH and isotopic paleotemperatures of Cretaceous oceans. Palaeogeography, Palaeoclimatology, Palaeoecology 170: 49–57. Crossref
Acta Palaeontol. Pol. 66 (1): 231–253, 2021
https://doi.org/10.4202/app.00783.2020