
A new caenogastropod from the upper Rhaetian of Lombardy: Palaeobiogeographical history and implications for the Early Jurassic gastropod recovery
VITTORIO PIERONI, STEFANO MONARI, and JONATHAN A. TODD
Pieroni, V., Monari, S., and Todd, J.A. 2021. A new caenogastropod from the upper Rhaetian of Lombardy: Palaeobiogeographical history and implications for the Early Jurassic gastropod recovery. Acta Palaeontologica Polonica 66 (1): 193–206.
A new gastropod genus and its type species, namely Ederazyga fanchini gen. et sp. nov., are described from the upper Rhaetian deposits of Lombardy (northern Italy) and tentatively placed into the family Zygopleuridae. The first appearance of Ederazyga is recorded in the lower Carnian deposits of Southern Alps and the stratigraphical distribution of the genus ends almost at the Triassic/Jurassic boundary. Its occurrence in Norian beds of the Nayband Basin (central Iran) suggests an eastward extension of the distribution during that time. Ederazyga is probably one of the Alpine gastropod taxa appearing in this area after the formation of the basin. The genus is possibly related to a group of Early Jurassic, medium to large Zygopleura-like species that are well represented in the Sinemurian and Pliensbachian carbonate platform deposits of the Mediterranean region and in the Hettangian to Pliensbachian successions of the European epicontinental shelf. This group shows an apparent species radiation in these areas testifying to the gastropod recovery following the Late Triassic decline in biodiversity. Ederazyga fanchini is shown to be congeneric with Cerithium? lateplicatum Klipstein, 1843, which is the type species of Camponaxis Bandel, 1995. The definition and diagnosis of Camponaxis was based on specimens that are clearly different, at generic and higher taxonomic levels, from the holotype of C.? lateplicatum. They belong to a distinct species here named Camponaxis bandeli sp. nov. Several species have been subsequently ascribed to Camponaxis following its original diagnosis. Therefore, we invoke ICZN Art. 70.3.2 in order to preserve the unambiguous identity of the genus and to ensure its nomenclatural stability. Camponaxis bandeli is fixed as the new type species for Camponaxis and C.? lateplicatum is here assigned to Ederazyga.
Key words: Gastropoda, Zygopleuridae, systematics, palaeobiogeography, faunal recovery, Jurassic, Late Triassic, Italy, Southern Alps.
Vittorio Pieroni [info@marianopieroni.it], Museo Scientifico Naturalistico “A. Stoppani”, Seminario Arcivescovile “Pio XI”, Via Papa Pio XI 32, 21040 Venegono Inferiore (VA), Italy.
Stefano Monari [stefano.monari@unipd.it], Dipartimento di Geoscienze, Università di Padova, Via G. Gradenigo 6, 35131, Padova, Italy.
Jonathan A. Todd [j.todd@nhm.ac.uk], Department of Earth Sciences, The Natural History Museum, Cromwell Road, London SW7 5BD, UK.
Received 7 July 2020, accepted 26 August 2020, available online 4 January 2021.
Copyright © 2021 V. Pieroni et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Late Triassic gastropod record is characterized by a strong and progressive post-Carnian decrease in species richness that reaches its nadir in the Rhaetian (Nützel 2005). This trend most probably reflects a real decrease in overall diversity but detailed studies on the changes in taxonomic diversity and their timing as well as the possible taphonomic control of the record are currently lacking. The interpretation of the Rhaetian record is particularly problematic since the available data are of a quite low overall quality. Apart from the studies on the relatively diverse faunas of the Nayband Basin (central Iran) described by Nützel and Senowbari-Daryan (1999) and Nützel et al. (2003, 2010) and other more sparse contributions (e.g., Bandel 1994; Barker and Munt 1999; Begg and Grant-Mackie 2003; Hasibuan and Grant-Mackie 2007; Ferrari 2015), literature on Rhaetian gastropods is significantly outdated. This is especially the case for the Alpine region. Sources reporting systematic studies, though relatively numerous, date back to the nineteenth century and the more recent ones are at least fifty years old (Escher von der Linth 1853; Winkler 1859, 1861; Stoppani 1857, 1860–1865; Dittmar 1864; Fischer-Ooster 1870; Ammon 1878, 1893; Lepsius 1878; Schäfer 1888; Haas 1909; Kittl and Spengler 1916; Desio 1929; Osswald 1930; Parona 1932; Borghi 1938; Kühn 1942; Chiesa 1949; Zapfe 1949, 1962, 1963, 1965, 1967; Conti 1954). Most of these papers concern gastropod faunas from the thick carbonate platform units of the Northern Calcareous Alps and Southern Alps where the fossils are frequently poorly preserved, difficult to isolate from the hard matrix or preserved as moulds. Their state of preservation obviously limits systematic analysis and the interpretation of the data. On the other hand, the study of these faunas is crucial to understand the reasons for the impoverishment of the Late Triassic gastropod record and the dynamics of the Early Jurassic recovery that helped shape the subsequent evolutionary history of this group.
In this paper a new gastropod genus and its type species are erected on the basis of specimens preserved as moulds obtained from the upper Rhaetian carbonate platform beds of Lombardy (northern Italy). Through taking into account the state of preservation and the poor information on the coeval faunas, a tentative reconstruction of the biogeographical history of the genus is proposed and its possible relationships with Early Jurassic gastropods are investigated. Since the facies, lithology and quality of preservation of most of the Rhaetian Alpine gastropod faunas listed above are comparable to those of the material described here, both the constraints and potential shown by this study are suggestive of what one can expect from a future revision of these faunas.
Institutional abbreviations.—GPIH, Geologisch-Paläontologisches Institut, Universität Hamburg, Hamburg, Germany; MRZ, Museo delle Regole “Rinaldo Zardini”, Cortina d’Ampezzo, Belluno, Italy; MSNVI, Museo Scientifico Naturalistico “A. Stoppani”, Venegono Inferiore, Varese, Italy; NHMUK, The Natural History Museum, London, United Kingdom; RGM, Rijksmuseum van Geologie en Mineralogie, now Naturalis Biodiversity Centre, Leiden, The Netherlands.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in ZooBank: urn:lsid:zoobank.org:pub:30E06C37-5516-476A-B14D-C7CB44A8DFEB.
Geological setting
The material comes from Le Ville (Villa de Grandi and Villa Edera) (DMS 45°51’48”N 8°46’29”E), a locality at about 900 m altitude on the southern side of Monte Tre Croci, a peak of the Monte Campo dei Fiori mountain group (Varese, Lombardy, northern Italy) (Fig. 1A). The Upper Triassic–Lower Jurassic stratigraphy and the palaeogeographic evolution of Lombardy have been the subject of many recent studies (e.g., Jadoul et al. 1994, 2004, 2005; Galli et al. 2005, 2007; Jadoul and Galli 2008; Tackett and Bottjer 2012). The Monte Campo dei Fiori area and Villa Edera section, were specifically treated by Jadoul et al. (2005) to which we refer readers for further information. In the area of Monte Campo dei Fiori, the Rhaetian deposits are represented by platform carbonates of the Zu Limestone Formation. The succession is reduced in thickness and represents the deposition on a structural high bordering the eastern side of the Monte Nudo Basin (western Lombardy). In the Villa Edera section only the upper part of the unit crops out (upper Zu Limestone, upper Rhaetian; Fig. 1B) and consists of grainstone, wackestone and brecciated carbonates representing a cyclic transition from beach to tidal and subaerial environments. The top of the upper Zu Limestone is cut by an unconformity which is easily recognizable across the whole of western Lombardy. This surface is often associated with brecciated limestone and reflects a subaerial exposure event in the Hettangian. The material described here comes from a light nutbrown biocalcarenite bed about three metres below this surface where it is associated with Conchodon infraliasicus Stoppani, 1865, and other bivalves. In the Villa Edera section the Hettangian unconformity is capped by the Saltrio beds, a Sinemurian unit consisting of grey and crinoidal calcarenite deposited in subtidal conditions.
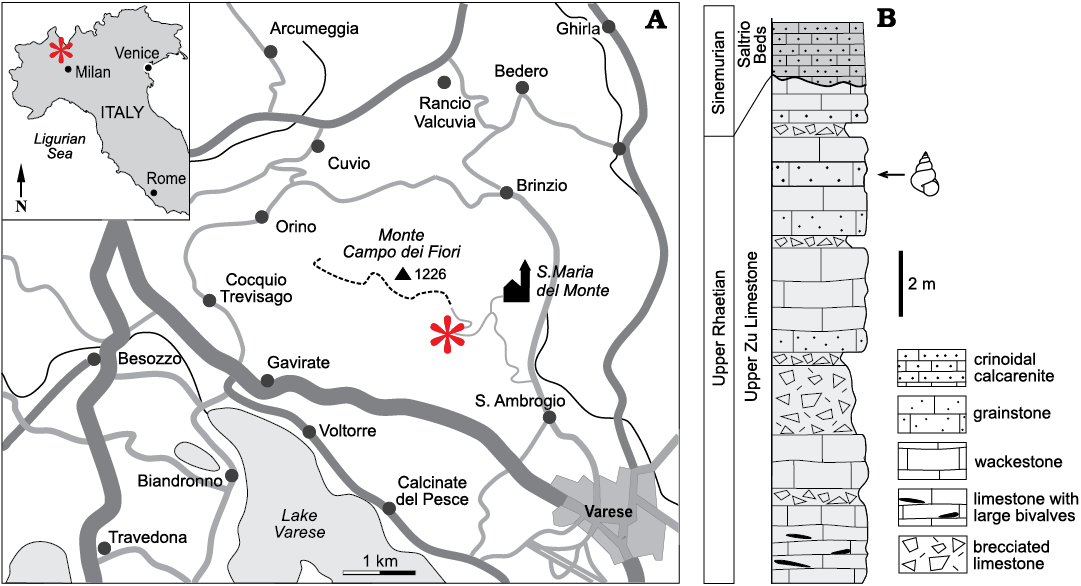
Fig. 1. Location of the outcrop (A) and stratigraphical section (B) modified from Jadoul et al. (2005).
Molluscs from the Rhaetian succession of Monte Campo dei Fiori are mainly represented by bivalves. The locality was indicated by Stoppani (1857, 1860–1865) as yielding part of the material from which the author erected C. infraliasicus (see Teruzzi 2015 for details). Kutassy (1933) also described some bivalves from outcrops in this area. In contrast, no gastropods were mentioned by Stoppani (1860–1865) from Monte Campo dei Fiori, although the author described numerous Rhaetian species from Lombardy. Chiesa (1949), however, listed about ten species that seemingly come from the same stratigraphical interval yielding the material studied here. In addition, indeterminable molluscs have been frequently recorded in thin section from Rhaetian deposits of this and neighbouring areas (e.g., Wiedenmayer 1963; Jadoul et al. 2005).
Material and methods
The material of the new species Ederazyga fanchini consists of two specimens deposited at the Museo Scientifico Naturalistico “A. Stoppani”, Venegono Inferiore (Varese, Lombardy, Italy) (MSNVI). One of them is represented by an external mould and its internal mould. The systematic study was made possible by the good preservation of the external mould. Its rubber cast represents slightly more than the axial half of the shell and reproduces with good detail both the apical spire and the adult shell. A full definition of the new genus described here required a review of the taxonomic status of Cerithium? lateplicatum Klipstein, 1843, type species of the genus Camponaxis Bandel, 1995, whose type material, stored at the Natural History Museum, London (UK), has been restudied. The systematic arrangement at higher taxonomic rank follows Bouchet et al. (2017). The photographs of E. fanchini have been made with the focus stacking method on the specimens coated with ammonium chloride and elaborated with Helicon Focus Pro 6.7.1 software.
Systematic palaeontology
Class Gastropoda Cuvier, 1795
Subclass Caenogastropoda Cox, 1960
?Superfamily Pseudozygopleuroidea Knight, 1930
?Family Zygopleuridae Wenz, 1938
Remarks.—The uncertain suprageneric attribution of Ederazyga gen. nov. is due to the lack of information on the protoconch. The genus is tentatively placed into the family Zygopleuridae Wenz, 1938, superfamily Pseudozygopleuroidea Knight, 1930, on the basis of the characters of the adult shell and the similarities with other zygopleurid taxa (see below for comparisons). Characters suggesting zygopleurid affinities are the slender shell with strongly convex, pendent whorls and the dominant axial ornament of prominent, opisthocline and sinuous collabral ribs made by distinctly parasigmoidal growth lines.
Genus Ederazyga nov.
Zoobank LSID: urn:lsid:zoobank.org:act:6E6B074A-0672-41C1-8D53- 7177DEA7A187
Etymology: After Villa Edera, type locality of the type species, and the suffix -zyga, referring to the probable zygopleuroidean affinities of the taxon.
Type species: Ederazyga fanchini sp. nov.; see below.
Species included: The following additional species, listed in stratigraphical order, are here ascribed to Ederazyga: Cerithium? lateplicatum Klipstein, 1843 (see below for references), lower Carnian, eastern Dolomites (Southern Alps, northern Italy); Striazyga? sp. in Kaim et al. (2006: 477, fig. 4E, F, H), Carnian, Karavanke Mountains (Slovenia); Katosira? lateplicata (Klipstein, 1843) sensu Nützel et al. (2012: 56, fig. 3G), upper part of middle Norian, Natanz region (central Iran).
Diagnosis.—Shell medium to large sized, fusiform to high-pagodiform. Adult whorls strongly convex, variably pendent in outline, with inflated peripheral region placed at or slightly below middle of whorl surface. Adapical part of whorl surface gently concave. Base subconoidal. Peristome with anterior opening ranging in shape from a short spout to a distinct canal. Collabral ornament composed of prominent opisthocline ribs, slightly to markedly swollen on peripheral band. Spiral ornament composed of sharp threads overlapping collabral ribs and producing a rough shell surface. Base with same ornament pattern as that of spire whorls.
Remarks.—The species belonging to Ederazyga gen. nov. have shells of medium to large adult size, attaining a height of a few centimetres to slightly more than ten centimetres. In addition to the diagnostic characters listed above, the last whorl tends to be more expanded, subventricose, with respect to the spire whorls, and the sharpness of the spiral ornament makes the surface of the shell slightly rough. In the type species, namely Ederazyga fanchini sp. nov., the peristome is reflected outward at the junction of the basal lip with the columellar lip and forms a short, shallow and wide lower spout. The apical spire is poorly preserved. The collabral and spiral ornament are visible from the second preserved whorl and it is not clear if the previous whorl is smooth or provided with a weak collabral ornament. The other described species here assigned to Ederazyga differs from the type species in having a more slender, turriculate-fusiform shell whereas in E. fanchini the shell is subpagodiform. This is the only observable difference and is not sufficient to justify their separation at genus rank.
Cerithium? lateplicatum Klipstein, 1843, shows several distinctive characters of Ederazyga: the size of the shell, the shape of the whorls and the ornament pattern. This species, here re-described (see below), was selected by Bandel (1995) as type species of the genus Camponaxis Bandel, 1995 (family Tofanellidae Bandel, 1995, superfamily Mathildoidea Dall, 1889). However, the definition of this genus, including its formal diagnosis, was based on specimens (Bandel 1995: 27, pl. 13: 9, pl. 14: 1–5) erroneously ascribed by this author to Klipstein’s (1843) species, as also pointed out by Nützel et al. (2012). The differences are obvious especially when one considers the size of the shell (Fig. 2). The type material of C.? lateplicatum is represented by a single fragmentary specimen (NHMUK PI OR 35701) that corresponds to the shell illustrated by Klipstein (1843: 182, pl. 11: 35) (Fig. 2A). Consequently, it is the holotype of the species by monotypy. The specimen, which preserves only the last two whorls, is 28 mm high and the reconstructed height of the shell is about 40 mm. In contrast, the height of the bigger shell illustrated by Bandel (1995) is 2.5 mm (Fig. 2B) and the author reported 4.5 mm as the maximum height observed in his material. According to Bandel (1995), his specimens represent immature shells of Klipstein’s (1843) species. However, some characters described by him, such as the high number of whorls (slightly less than ten), and the full development of the ornament, would indicate that at least the larger specimens represent subadult or adult shells. In addition, the peristome shows a slight anterior spout or sinus whereas in the holotype of C.? lateplicatum the peristome has a distinct anterior canal. Bandel’s (1995) shells belong to a distinct species here named as Camponaxis bandeli sp. nov., pro Camponaxis lateplicata (Klipstein, 1843) in Bandel (1995), non Klipstein (1843). The holotype, here selected, is the shell RGM 219 039 figured by Bandel (1995: pl. 14: 5) and refigured as Fig. 2B herein. Two additional specimens illustrated by Bandel (1995) are selected here as paratypes: one specimen is figured by this author on pl. 13: 9 and pl. 14: 1, 2, 4; the other specimen is figured on pl. 14: 3. As stated by Bandel (1995), the holotype is stored at RGM and comes from the lower Carnian (Trachyceras aonoides Zone) St. Cassian Formation of Dibona, eastern Dolomites (northern Italy) and the paratypes are from the same stratigraphical level but from the locality of Misurina in the same region. The paratypes are apparently registered as RGM 219 040 in Bandel (1995: 27), though noted on the explanation to plate 14 as being housed in the GPIH. The species has been adequately described by Bandel (1995: 27) and its species epithet we dedicate to this author. Several species have been ascribed to the genus Camponaxis (e.g., Bandel 1995; Bandel et al. 2000; Gründel and Nützel 2013, 2015; Nützel and Gründel 2015). They are quite comparable to each other and agree with the original diagnosis of the genus, later slightly emended by Gründel and Nützel (2013). Therefore, in order to preserve the unambiguous identity of the genus and to ensure its nomenclatural stability the taxonomic species involved in the misidentification has been selected as the type species of Camponaxis in accordance with ICZN (1999) Art. 70.3.2. Therefore, Camponaxis bandeli sp. nov. is fixed as the new type species for Camponaxis. As a consequence, we have erected the new genus Ederazyga to include C.? lateplicatum.
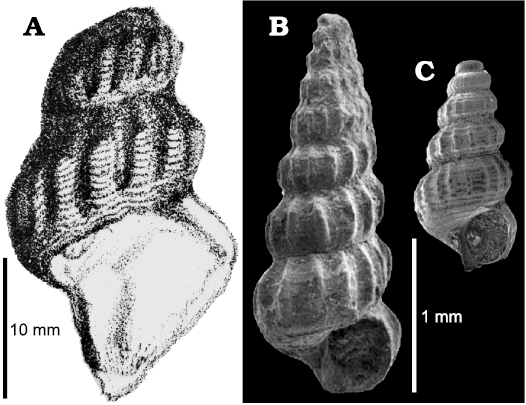
Fig. 2. Comparison between the holotype of zygopleurid? gastropod Cerithium? lateplicatum Klipstein, 1843 and the specimens illustrated by Bandel (1995), here ascribed to a tofanellid Camponaxis bandeli sp. nov.; lower Carnian, eastern Dolomites (Southern Alps, northern Italy). A. Fragmentary type specimen (NHMUK PI OR 35701) of Cerithium? lateplicatum, original illustration from Klipstein (1843: pl. 11: 35). B, C. Camponaxis bandeli sp. nov. B. Holotype, adult shell (RGM 219 039), from Bandel (1995: pl. 14: 5). C. Paratype, juvenile shell (RGM 219 040), from Bandel (1995: pl. 14: 3). Reproduced accordingly to CC-BY 4.0 license.
The genus is close to Stephanocosmia Cossmann, 1895, and Tyrsoecus Kittl, 1892, in the presence of strong collabral ribs that are swollen on the peripheral belt. Tyrsoecus is smaller, more slender and with lower and more numerous whorls. Stephanocosmia lacks the spiral ornament and the collabral ribs attenuate and vanish on the last whorls where they reduce to a peripheral spiral row of nodes. Kaim et al. (2006) and Nützel et al. (2012) tentatively assigned to Striazyga Nützel, 1998, and Katosira Koken, 1892, some species here considered as belonging to Ederazyga (see list above). These genera differ from Ederazyga in having a turriculate shell of much smaller size, with less convex and lower whorls. Moreover, the collabral ribs are much less prominent and not swollen on the peripheral region.
The Early Jurassic genus Pseudokatosira Nützel and Gründel, 2007, is reminiscent of Ederazyga in the size and general characters of the shell, and in the presence of a distinct anterior canal. It differs in its more slender shell with less convex whorls. As detailed by Nützel and Gründel (2007, 2015), in Pseudokatosira the collabral ribs of the early teleoconch attenuate after several whorls and then become broad, swollen on the periphery, on the last whorls. This ontogenetic change is absent in Ederazyga.
Ederazyga is also reminiscent of Maturifusus Szabó, 1983, type genus of the family Maturifusidae Gründel, 2001. The systematic position of this genus and of the family have been discussed by many authors (Szabó 1983; Bandel 1993, 2006; Schröder 1995; Gründel 1998, 1999, 2001, 2005; Riedel 2000; Guzhov 2004; Kaim 2004; Kaim and Beisel 2005; Nützel 2010; Bandel and Dockery 2012; Bouchet et al. 2017; Webster and Vermeij 2017). Most of them interpreted the Maturifusidae as a stem group of neogastropods or an ancient group of Latrogastropoda Riedel, 2000, probably close to the Buccinidae Rafinesque, 1815, or to the Purpurinidae Zittel, 1895. Ederazyga shares with Maturifusus the general shape of the shell, the peristome provided with an anterior opening and the strong ornament of spiral threads/cords overlapping collabral ribs. In Ederazyga the spire is more slender and the suture is more inclined. More relevant, the growth lines and, consequently, the collabral ribs are opisthocline and more distinctly parasigmoidal. About ten Jurassic species of Maturifusus have been described to date (Szabó 1983; Schröder 1995; Gründel 1998, 1999, 2001, 2005; Guzhov 2004; Kaim 2004) and all of them invariably show prosocline growth lines.
The same difference distinguishes Ederazyga from Pseudotritonium Wenz, 1940, a genus provisionally assigned by Nützel (2010) to the family Maturifusidae on the basis of its adult shell characters. Moreover, in Pseudotritonium the peristome is expanded on the outer lip. According to Nützel (2010), the affinities between Pseudotritonium and Maturifusus, if confirmed by the study of the protoconch, currently unknown in the type species of both genera, would indicate that the family Maturifusidae is a junior synonym of the family Pseudotritonidae Golikov and Starobogatov, 1987 (see also Bouchet et al. 2017).
Brachytrema? despujolsi Dubar, 1948, from the lower Pliensbachian of the High Atlas of Morocco (Dubar 1948: 62, pl. 3: 31–33), is similar to Ederazyga species in the general shape of the shell and in the ornament pattern with axial ribs that are swollen in the peripheral region. However, this species clearly belongs to a different genus in showing a markedly expanded aperture which is provided with a strong varix, and by prosocline collabral ribs and growth lines. In this respect, it seems more closely related to Pseudotritonium.
Obviously, the differential characters mentioned above, although suggesting closer relationships of Ederazyga with the Pseudozygopleuroidea Knight, 1930, are not sufficient to exclude the attribution of this genus to the Maturifusidae. A reliable and more precise taxonomic placement of Ederazyga requires information on its protoconch and on that of the other, comparable taxa.
The large shell, the pendent outline of the whorls and the opisthocline collabral ribs that are more prominent and swollen on the periphery make Ederazyga close to a group of currently unrevised Zygopleura-like species known especially from the Early Jurassic carbonate platforms of western Tethys. Since this group seemingly played an important role in the Early Jurassic gastropod recovery, comparisons with Ederazyga and a discussion of their possible evolutionary relationships are discussed below in the conclusion.
Stratigraphic and geographic range.—The genus ranges from the early Carnian to the late Rhaetian and its distribution extends from the Southern Alps to central Iran.
Ederazyga fanchini sp. nov.
Fig. 3.
Zoobank LSID: urn:lsid:zoobank.org:act:B3326419-D750-4FA6-8C62- 984931816D37
Etymology: Species dedicated to Piero Fanchin, collaborator of MSNVI, who collected the material.
Holotype: MSNVI 042/049, internal and external moulds (Fig. 3A).
Type locality: Villa Edera section in Jadoul et al. (2005), southern side of Monte Tre Croci (Monte Campo dei Fiori group), Lombardy, northern Italy.
Type horizon: Upper Zu Limestone (sensu Jadoul et al. 2005), light nut-brown biocalcarenite, upper Rhaetian.
Material.—Type material and plaster cast replica of an internal mould MSNVI 042/049a (the original specimen has been lost).
Measurements.—Holotype MSNVI 042/049: height of the shell (not complete) 92.7 mm; height of the last whorl 58.1 mm; width of the shell 63.6 mm; apical angle 47°; pleural angle 50°. MSNVI 042/049a: height of the shell (not complete) 81.5 mm; height of the last whorl 67.1 mm; width of the shell 83.6 mm.
Diagnosis.—Shell moderately large, high-pagodiform and composed of about ten whorls. Height of last whorl slightly exceeding half-height of shell. Height of spire whorls about half of their width. Earliest teleoconch whorls regularly convex. Periphery of subsequent whorls slightly below mid-whorl surface. Peristome with wide anterior spout. Collabral ribs rather swollen on peripheral band and forming axially elongated nodes. Each whorl sculptured by 11 collabral ribs. Two orders of spiral threads on spire whorl. Up to four orders of spiral threads on last whorl.
Description.—The shell is large, pagodiform and composed of about ten whorls. The outline of the apical spire is almost conoid or feebly cyrtoconoid whereas the fully adult part of the shell is weakly coeloconoid. The spire whorls have a height slightly less than half of their width and the last whorl extends a little more than half of the shell height. The first two preserved teleoconch whorls appear regularly convex. On the subsequent whorls the periphery rapidly becomes prominent and corresponds to a rounded spiral belt where the axial ornament is rather strong. It lies slightly below mid-whorl and gives to the whorl surface an angular outline. The ramp is slightly concave whereas the surface below the periphery is convex and inclined towards the spire axis. The suture is distinctly impressed, almost grooved. The base is subconoid with a moderately convex surface and most probably anomphalous. The aperture is drop-shaped tending to subpentagonal. The peristome at the junction of the basal lip with the columellar lip shows a short, low and wide anterior spout marked by the outward reflection of the shell surface. The teleoconch is ornamented by collabral ribs and spiral threads. Opisthocline and opisthocyrt collabral riblets and, seemingly, some spiral threads are visible at least on the second preserved whorl. On subsequent whorls the collabral riblets become strong ribs overlapped by cord-like spiral threads. On the fourth preserved whorl the collabral ribs are slightly opisthocline and extend from suture to suture. They are crossed by about six, equally spaced and sized cord-like threads that are slightly stronger when passing on the collabral ribs. The most adapical spiral thread bears a row of little nodes marking the abapical edge of the sutural groove (Fig. 3A7). During the growth, the spiral threads increase in number by intercalation. Six primary spiral threads ornament the ramp of the penultimate whorl. They are intercalated by secondary threads running approximately along the mid-line of the intervals between the primary threads. The same number and pattern of spiral threads cover the remaining part of the whorl surface. Third and fourth orders threads appear sparsely on the last whorl making the spiral pattern less regular. The collabral ribs are rounded, widely spaced and opisthocline. They strengthen during growth and become rather prominent on the peripheral band where they form rounded and axially elongated nodes. On the ramp, the collabral ribs attenuate gradually towards the adapical suture whereas they keep their prominence on the outer face. Each whorl is sculptured by 11 collabral ribs which are not aligned axially with the ribs of the adjacent whorl and appear almost to alternate in position. The base is ornamented by the same spiral pattern as that of the whorl surface whereas the collabral ribs are weakly prosocyrt and seem to attenuate towards the axial region. The whorl surface is crossed by opisthocline and feebly opisthocyrt growth lines, slightly prosocyrt on the subsutural band. On the last whorl, the growth lines occasionally form irregular collabral thickenings that are more marked on the ramp. Although the growth lines are not clearly identifiable on the basal surface, the shape of the collabral ribs and traces of growth thickenings indicate that they are prosocyrt.

Fig. 3. Zygopleurid? gastropod Ederazyga fanchini gen. et sp. nov.; upper Rhaetian, Villa Edera (Lombardy, northern Italy). A. Holotype MSNVI 042/049, inner mould in apertural (A1), basal (A2), and dorsal (A3) views; external mould in general view (A4), rubber cast of the dorsal view (A5), detail of the apical spire (A6), and detail of the penultimate and last whorls (A7). B. Plaster cast replica of MSNVI 042/049a, inner mould in apertural (B1), basal (B2), and dorsal (B3) views.
Remarks.—The holotype is represented by an external mould and internal mould. The external mould preserves only part of the base which seems to be anomphalous. The internal mould lacks the apical spire and shows a deep axial cavity presumably reflecting the presence of a smooth and robust columella. The aperture is incomplete. Its shape and the morphology of the base indicate that the columellar lip is slightly downward elongated in the axial direction. On the other available specimen (MSNVI 042/049a), which is represented by the plaster cast replica of an internal mould, the junction of the columellar lip with the basal lip is outwardly reflected. Here the basal lip, though incomplete, forms an evident curve reflecting the presence of a short and wide anterior spout (Fig. 3B2, B3). The external mould of the holotype shows details of the ornament and the apical spire. These characters are well reproduced on its rubber cast. The apical spire is almost complete but poorly preserved.
Ederazyga lateplicata (Klipstein, 1843) and the specimen assigned by Nützel et al. (2012) to the same species differ from Ederazyga fanchini sp. nov. in their smaller size. The shells are more slender and fusiform and the whorls are higher. Moreover, the holotype of E. lateplicata shows sharper collabral ribs, with a less swollen periphery, and a distinct anterior canal on the peristome. Striazyga? sp. described by Kaim et al. (2006), here considered as belonging to Ederazyga, is more slender and acute. Moreover, the whorls are more regularly convex, the collabral ribs are slightly more numerous and the spiral ornament is less sharp.
Stratigraphic and geographic range.—Type locality and horizon only.
Ederazyga lateplicata (Klipstein, 1843) comb. nov.
Figs. 2A, 4.
1843 Cerithium (?) lateplicatum n. sp.; Klipstein 1843: 182, pl. 11: 35.
?1868 Loxonema lateplicata Klipstein, 1845 (sic!); Laube 1868: 62–63, non pl. 24: 14.
1894 Katosira (?) lateplicata Klipstein, 1843 (sic!); Kittl 1894: 164–165, pl. 4: 27–28.
?1978 Katosira seelandica (Kittl, 1894) var. alta n.f.; Zardini 1978: 43–44, pl. 28: 8a, b.
non1995 Camponaxis lateplicata (Klipstein, 1843); Bandel 1995: 27, pl. 13: 9, pl. 14: 1–5.
non2012 Katosira? lateplicata (Klipstein, 1843); Nützel et al. 2012: 56, fig. 3G.
Material.—A single specimen (NHMUK PI OR 35701) is present in the Klipstein’s collection of the NHMUK. It corresponds to the single shell figured by Klipstein (1843: pl. 11: 35) and therefore it is deemed to be the holotype of the species. Saint Cassian Formation, lower Carnian, Kreutzkofel (eastern Dolomites, northern Italy).
Measurements.—Height of the shell (not complete) 28.0 mm; apparent width of the shell 17.8 mm; height of last whorl 21.1 mm; apparent pleural angle 31°.
Description.—The shell is slender, seemingly fusiform-turriculate. The whorls are strongly convex, with a roundedly prominent periphery at their mid-line. The periphery tends to become subangulate on the last whorl. The surface of the whorl is slightly concave below the suture. The width of the penultimate whorl is two times its height and the last whorl is seemingly half the reconstructed height of the shell. The base is anomphalous, conoidal and downward elongated to form a lower neck. The surface of the base is evenly convex and passes smoothly to the whorl side. The peristome is not preserved but the cross section of the last whorl indicates a seemingly ovoid and axially elongated aperture. The columella is stout and straight. It extends downward in axial direction where edges a distinct lower canal. The collabral ornament consists of strong and gently opisthocline ribs. They are moderately swollen on the peripheral region and become thinner towards both the sutures. The penultimate whorl bears about ten ribs. Dense, sharp, subequally spaced and sized spiral threads overlap the collabral ribs. An alternation of finer and coarser spiral threads is visible in a wide area above and below the periphery. The finer threads are about half the width of the coarser ones and possibly occur also in the other parts of the shell. On the ribs, the spiral threads become slightly thicker and seem to form low, spirally elongated nodes. About fifteen spiral threads ornament the penultimate whorl.
Remarks.—The specimen preserves only the last two whorls and it is slightly deformed by compaction. Thought incomplete, the peristome shows the presence of a distinct lower canal. Since the surface of the shell is partly covered by a hard matrix, the details of the ornament, especially the finer spiral threads, are visible only in patches. Most probably the material on which Zardini (1978) (Fig. 4D) instituted Katosira seelandica var. alta Zardini, 1978, belongs to Ederazyga lateplicata (Klipstein, 1843). It differs only in the slightly higher whorls and less numerous collabral ribs. As reported by Kittl (1894), the illustration of the shell ascribed by Laube (1868) to Loxonema lateplicata is a composite from fragmentary specimens belonging to different species. According to him, the most complete specimen belongs to Coronaria subcompressa Kittl, 1894, whereas other two specimens most probably belong to Katosira? lateplicata. Nützel et al. (2012) also maintained that Laube (1868) illustrated a shell different from Klipstein’s (1843) holotype. The specimen from the middle Norian beds of Nayband Formation (central Iran) assigned by Nützel et al. (2012) to K.? lateplicata represents a different and probably new species. Although clearly congeneric, this species differs from E. lateplicata in its more angular and pendent whorls, and in the collabral ribs that are more swollen and less numerous.
Stratigraphic and geographic range.—Lower Carnian, eastern Dolomites, northern Italy.

Fig. 4. Zygopleurid? gastropod Ederazyga lateplicata (Klipstein, 1843); lower Carnian, eastern Dolomites (Southern Alps, northern Italy). A. Holotype NHMUK PI OR 35701, incomplete shell, in apertural (A1), dorsal (A2), and subdorsal (A3) views, detail of the ornament (A4). B. Original labels of the holotype (the label at the top shows an incorrect inventory number). C. Illustration of a specimen classified by Kittl (1894: pl. 4: 28) as Katosira? lateplicata. D. Original illustration of the specimen MRZ3711, ascribed by Zardini (1978: pl. 28: 8a, b) to Katosira seelandica var. alta, incomplete shell, in apertural (D1) and dorsal (D2) views, reproduced with permission Tipografia Ghedina Snc.
Discussion
Palaeobiogeography.—Occurrences of Ederazyga species are sparse and far apart (Fig. 5). Their rather patchy palaeogeographical distribution could reflect a primary rarity of the taxon in Late Triassic gastropod communities. However, the bias of preservation probably played a role especially in the Norian–Rhaetian record of the carbonate platform units. The genus ranges from Carnian to Rhaetian and its palaeogeographical distribution extends from the Southern Alps to central Iran. In the Southern Alps Ederazyga first occurs in the lower Carnian beds of the St. Cassian Formation with Ederazyga lateplicata. The genus is also present in the middle–upper part of the Carnian (post-Cordevolian in a tripartite scheme of the Carnian stage) of the Karavanke Mountains (Slovenia) (Striazyga? sp. in Kaim et al. 2006). The data presented here indicate that in the Southern Alps Ederazyga persisted up to the late Rhaetian.
The occurrence of Ederazyga in the Norian beds of the Nayband Basin (central Iran) (Katosira? lateplicata sensu Nützel et al. 2012) suggests an eastward extension of its distribution during that time. The Nayband Basin originated during the Eo-Cimmerian phase of collision of the Iran plate with the Turan plate (Eurasia) (Fürsich et al. 2005; Barrier and Vrielinck 2008; Brunet et al. 2009; Wilmsen et al. 2009). According to Wilmsen et al. (2009), its formation was the consequence of the onset of subduction of the Neotethys at the southern margin of the Iran plate in middle Norian times. The benthic macrofaunas of the Nayband Basin show affinities with those of both the Alpine region and south-eastern Asia (Fürsich et al. 2005 and references therein). Ederazyga could be interpreted as one of the Alpine gastropod taxa appearing in this area after the formation of the basin. The close relationships highlighted by Nützel and Senowbari-Daryan (1999), at least at the genus rank, between some of the Norian–Rhaetian gastropods of the Nayband Basin and the older, mainly Carnian fauna of the Southern Alps seems to support this suggestion.
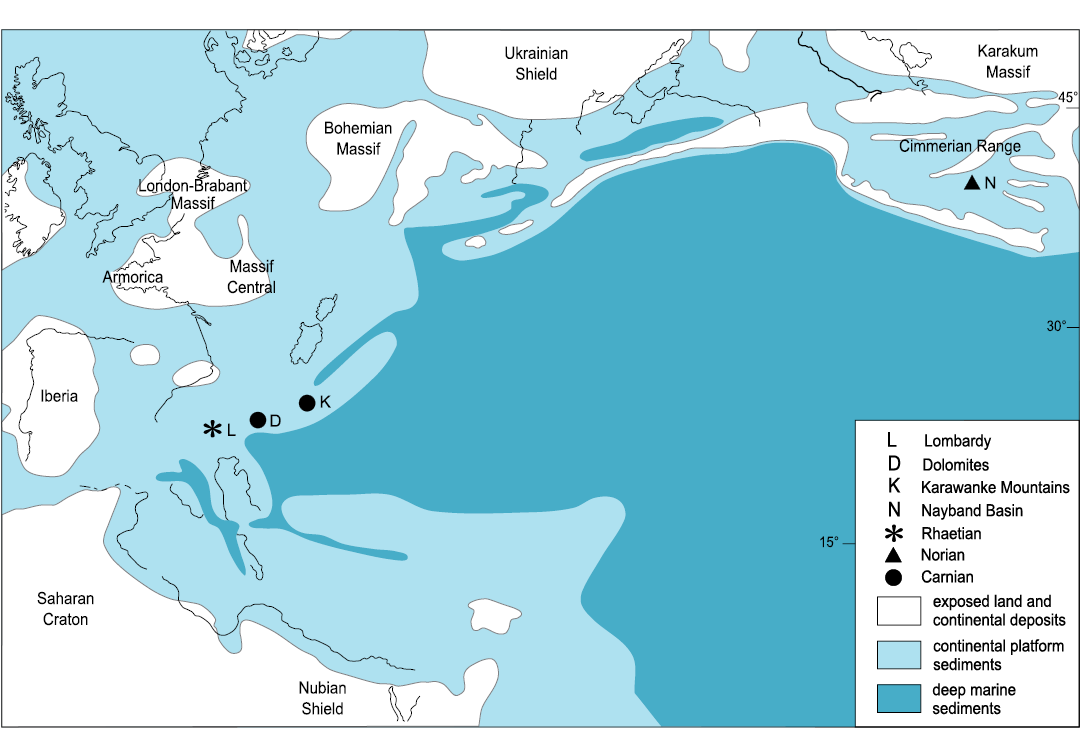
Fig. 5. Palaeogeographical distribution of Ederazyga during the Late Triassic. Map modified from the late Norian maps of Dercourt et al. (2000) and Barrier and Vrielinck (2008).
Relationships with the Early Jurassic gastropods.—The stratigraphical distribution of Ederazyga does not cross the Triassic/Jurassic boundary. In fact, the genus is absent from the well documented earliest Jurassic gastropod faunas of Europe (see Monari et al. 2011 for references). Its last occurrence, i.e., E. fanchini, is only a few meters below the stratigraphical discontinuity that, according to Jadoul et al. (2005) marks the Triassic/Jurassic boundary in western Lombardy.
Table 1. List, references, localities, and stratigraphical position of the Early Jurassic Zygopleura-like species possibly related to Ederazyga.
|
Species |
Localities |
Stratigraphical levels |
References |
|
Zygopleura
vinosimonensis |
Vendée (western France) |
Hettangian |
Fischer and Weber
1997: 16; Cossmann 1902: 189, pl. 4:
2, 4, described as Zygopleura subnodosa;
|
|
Melania
theodori |
Lorraine (eastern France) |
upper Hettangian |
Terquem 1855: 257, pl. 14: 6, 6a |
|
southern Luxembourg |
upper Hettangian |
Meier and Meiers 1988: 40, pl. 9: 25a, b, described as Zygopleura etalensis; SM, personal observation 2019 |
|
|
Zygopleura subnodosa (d’Orbigny, 1850) |
Calvados (northern France) |
upper Pliensbachian |
Eudes-Deslongchamps 1843: 219, pl. 12: 1, described as Melania nodosa; d’Orbigny 1850: 226; d’Orbigny 1851: 37, pl. 237bis: 6; Fischer and Weber 1997: 15, pl. 1: 8 |
|
Chemnitzia tatia Gemmellaro, 1878 |
north-western Sicily (southern Italy) |
Sinemurian |
Gemmellaro 1878: 135, pl. 6: 1–3; De Gregorio 1886: 6 |
|
eastern Sicily (southern Italy) |
Sinemurian |
Di Stefano 1887: 431, pl. 2: 6a, b |
|
|
Tuscany (central Italy) |
Sinemurian |
Fucini 1895: 315 |
|
|
eastern High Atlas (Morocco) |
?Sinemurian |
?Bourrouilh 1966:
45, pl. 1: 6a, b, described as |
|
|
Chemnitzia polyplecta Gemmellaro, 1878 |
north-western Sicily (southern Italy) |
Sinemurian |
Gemmellaro 1878: 137, pl. 6: 7, 8 |
|
eastern Sicily (southern Italy) |
Sinemurian |
Di Stefano 1887: 432, pl. 2: 11, 12 |
|
|
Tuscany (central Italy) |
Sinemurian |
Fucini 1895: 316, pl. 12: 5, 5a |
|
|
Trento Platform (northern Italy) |
Pliensbachian |
Dal Piaz 1909: 9, pl. without number: 8, 9 |
|
|
Chemnitzia moorei Gemmellaro, 1878 |
north-western Sicily (southern Italy) |
Sinemurian |
Gemmellaro 1878: 136, pl. 6: 4, 5 |
|
Tuscany (central Italy) |
Sinemurian |
Fucini 1895: 317 |
|
|
Chemnitzia veturia Gemmellaro, 1878 |
north-western Sicily (southern Italy) |
Sinemurian |
Gemmellaro 1878: 139, pl. 6: 6 |
|
Tuscany (central Italy) |
Sinemurian |
Fucini 1895: 319, pl. 12: 12, 12a |
|
|
Chemnitzia
catacyclus |
eastern Sicily (southern Italy) |
Sinemurian |
Di Stefano 1887: 432, pl. 2: 7, 8 |
|
Chemnitzia appenninica Gemmellaro, 1878 |
north-western Sicily (southern Italy) |
Sinemurian |
Gemmellaro 1878: 138, pl. 6: 10, pl. 9: 1, 2 |
|
High Atlas (Morocco) |
lower Pliensbachian |
Dubar 1948: 98, pl . 9: 1a, b |
|
|
Chemnitzia paradisi Böhm, 1884 |
Trento Platform (northern Italy) |
Pliensbachian |
Böhm, 1884: 782, pl. 26: 5 |
|
High Atlas (Morocco) |
lower Pliensbachian and lower Toarcian |
Dubar 1948: 99, pl. 7: 11, 12 |
A number of Early Jurassic species, namely Chemnitzia tatia Gemmellaro, 1878, Chemnitzia polyplecta Gemmellaro, 1878, Chemnitzia moorei Gemmellaro, 1878, Chemnitzia appenninica Gemmellaro, 1878, Chemnitzia veturia Gemmellaro, 1878, Chemnitzia catacyclus Di Stefano, 1887, and Chemnitzia paradisi Boehm, 1884 (Table 1, Fig. 6A–K; see also Gatto and Monari 2010), share with Ederazyga the shape and the medium to large size of the shell, the outline of the whorls and the ornament of strong, opisthocline collabral ribs that are swollen or subnodose in the peripheral region. The only appreciable difference is their lack of a prominent spiral ornament. These species are reported only in very old literature sources or have been no longer quoted after their initial description, and revision probably will lead to the reduction of their number. In any case, they occur frequently in the Sinemurian–Pliensbachian carbonate platform deposits of the Mediterranean region, such as those of the Trapanese Platform (north-western Sicily, southern Italy), the Longi-Taormina unit of the Calabrian Arc (eastern Sicily, southern Italy), the Tuscany Zone (central Italy), the Trento Platform (northern Italy) and the High Atlas (Morocco) (Fig. 7).
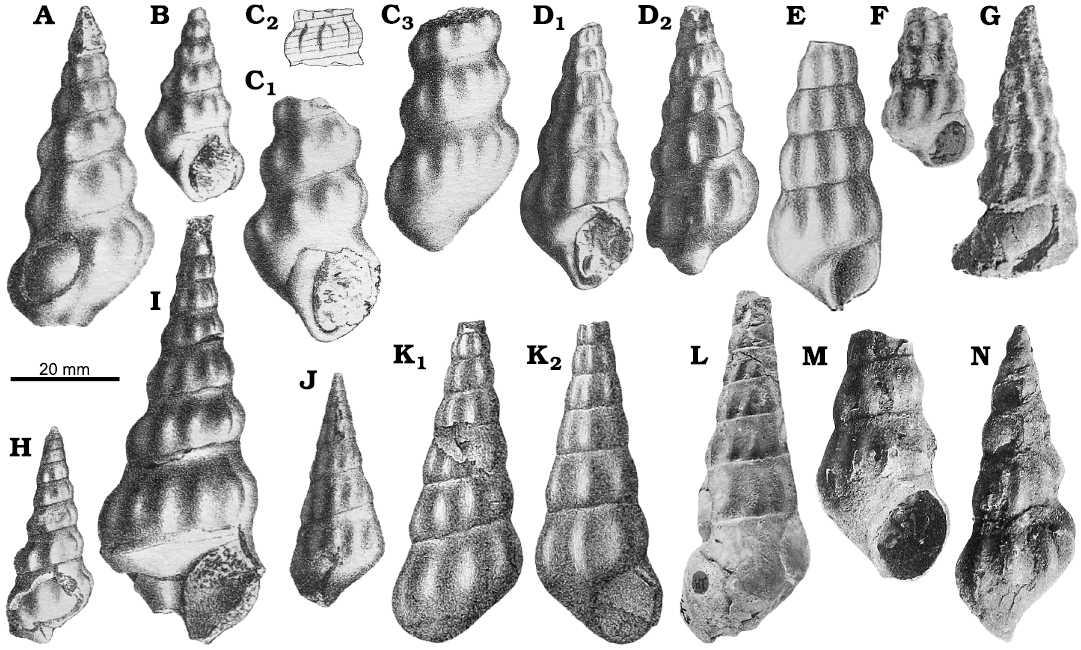
Fig. 6. Original illustrations of the Early Jurassic Zygopleura-like species probably closely related to Ederazyga. A, B. Chemnitzia moorei Gemmellaro, 1878 (pl. 6: 4, 5), Sinemurian, Rocca Busambra (north-western Sicily, southern Italy). C. Chemnitzia tatia Gemmellaro, 1878 (pl. 6: 1–3), Sinemurian, Rocca Busambra (north-western Sicily, southern Italy), in apertural view (C1), detail showing the spiral ornament (C2), and dorsal view (C3). D. Chemnitzia polyplecta Gemmellaro, 1878 (pl. 6: 7, 8), Sinemurian, Rocca Busambra (north-western Sicily, southern Italy), in apertural (D1) and dorsal (D2) views. E. Chemnitzia catacyclus Di Stefano, 1887 (pl. 2: 7b), Sinemurian, Taormina (eastern Sicily, southern Italy). F, G. Specimens figured by Dubar (1948: pl. 7: 11a, 12) as Zygopleura paradisi (Böhm, 1884), lower Pliensbachian (F) and lower Toarcian (G), Djebel Bou-Dahar, (High Atlas, Morocco). H, I. Chemnitzia appenninica Gemmellaro, 1878 (pl. 9: 1, 2), Sinemurian, Rocca Busambra (north-western Sicily, southern Italy). J. Chemnitzia veturia Gemmellaro, 1878 (pl. 6: 6), Sinemurian, Rocca Busambra (north-western Sicily, southern Italy). K. Specimen figured by Fucini (1895: pl. 12: 5, 5a) as Zygopleura polyplecta (Gemmellaro, 1878), Sinemurian, Monte Pisano (Tuscany, central Italy). L. Zygopleura subnodosa (d’Orbigny, 1850), holotype figured by Fischer and Weber (1997: pl. 1: 8), upper Pliensbachian, Calvados (northern France), in dorsal (K1) and apertural (K2) views. M, N. Zygopleura vinosimonensis Fischer and Weber, 1997, syntypes figured by Cossmann (1902: pl. 4: 2, 4) as Zygopleura subnodosa, Hettangian, Vendée (western France).
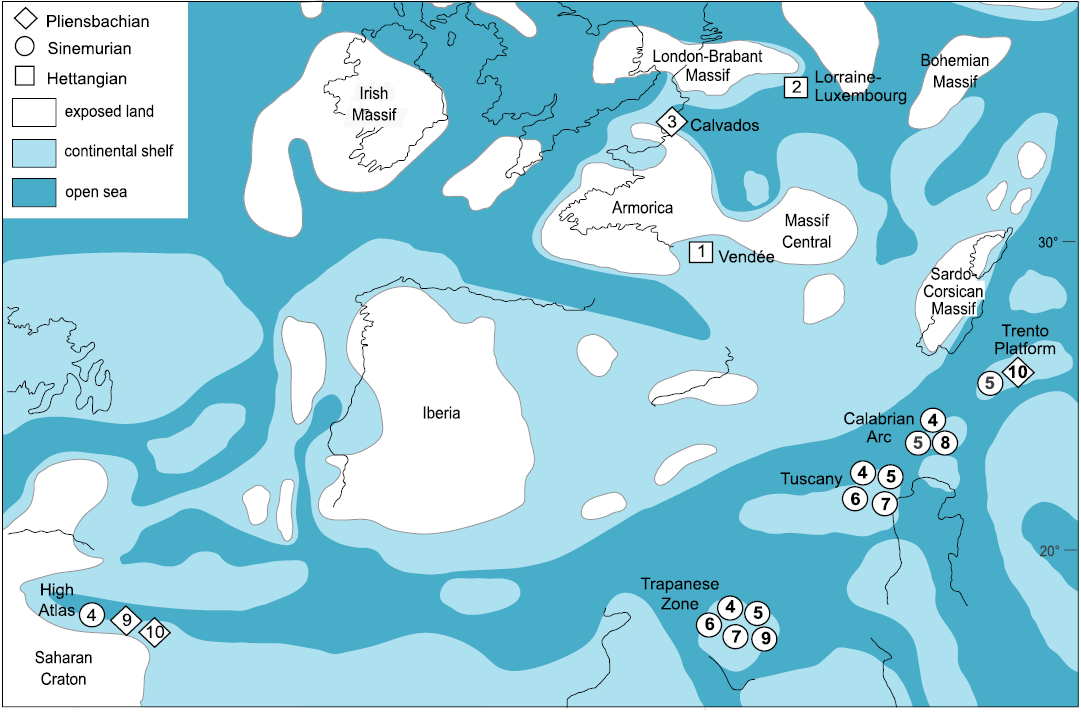
Fig. 7. Early Jurassic palaeogeographical distribution of the Zygopleura-like species listed in Table 1. Map simplified from the late Sinemurian map of Dercourt et al. (2000). Abbreviations: 1, Zygopleura vinosimonensis; 2, Melania theodori; 3, Zygopleura subnodosa; 4,Chemnitzia tatia; 5, Chemnitzia polyplecta; 6, Chemnitzia moorei; 7, Chemnitzia veturia; 8, Chemnitzia catacyclus; 9,Chemnitzia appenninica; 10, Chemnitzia paradisi.
Species showing the same characters are known also from the Hettangian–Pliensbachian epicontinental deposits of the Paris Basin and Vendée (eastern France), namely Zygopleura subnodosa (d’Orbigny, 1850), Zygopleura vinosimonensis Fischer and Weber, 1997, and Melania theodori Terquem, 1855 (Table 1, Figs. 6L–N, 7). According to Cossmann (1907), Z. subnodosa, together with Zygopleura? benoisti Cossmann, 1907, a species from the Bathonian of the Paris Basin (Cossmann 1907; Fischer 1964), differs from the typical Triassic representatives of Zygopleura Koken, 1892, in having a much bigger shell ornamented by collabral ribs that are swollen or subnodose on the peripheral region. In his opinion, they probably belong to a distinct Jurassic genus. This suggestion is plausible if one considers the clear uniformity of the adult characters of these species and of those listed above from the Mediterranean region. However, a confirmation needs the study of their protoconchs, that are currently unknown. Since also Ederazyga lacks information on the protoconch, its relationships with this group of species cannot be fully established and remain a matter for future researches. At the present, based on the available teleoconch characters, one cannot exclude that Ederazyga represents a Triassic member of a group that during the Early Jurassic radiated both in the western Tethyan carbonate platforms and in the European epicontinental shelf, testifying to the gastropod recovery after the Late Triassic decline in biodiversity.
Conclusions
The Late Triassic gastropod record shows a strong and progressive post-Carnian loss of species richness that most probably reflects a real decrease of overall diversity. For this reason, the Rhaetian record is in general rather poor. As far as the Alpine region is concerned, its interpretation is particularly problematic because, in addition to the scarcity of data, the literature sources (see references cited above) are significantly outdated and, in most cases, the gastropod faunas are poorly preserved. However, the present day knowledge of the systematics of the class Gastropoda is incomparably better than in the past, mainly thanks to the numerous contributions made in the last few decades (see Bouchet and Rocroi 2005 and Bouchet et al. 2017 for references). In this context, the work presented here can be regarded as a case study demonstrating that a systematic analysis can allow us to gain useful morphological information even from the study of incomplete and poorly preserved material. On the other hand, the study shows the main constraints imposed by the preservation of the material. They are mostly due to the lack of the protoconch, making uncertain the suprageneric attribution of Ederazyga and, therefore, its systematic relationships with other comparable taxa. However, one should underline that, since the protoconch is frequently not preserved, these uncertainties are common also to many other gastropod faunas represented by better preserved and richer material.
From a preliminary estimate, about 70 species were so far identified from the Rhaetian deposits of the Alpine region, and numerous other species were quoted in open nomenclature. In the light of this study, it seems highly likely that their revision would add important data permitting us to better understand the reasons for the impoverishment of the Late Triassic gastropod record in the Alpine region and to better assess the faunal turnover related to the subsequent Early Jurassic recovery.
Acknowledgements
We thank Piero Fanchin (Cassano Magnago, Varese, Italy), collaborator of the MSNVI for allowing access to the material studied here and for the geographical and stratigraphical information on its finding. We are also indebted to Flavio Jadoul (University of Milan, Italy) for additional data on the stratigraphy of the type locality, Roberto Gatto (University of Padua, Italy) for his critical reading of an earlier version of the paper. Alexander Nützel (Bayerische Staatssammlung für Paläontologie und Geologie and Ludwig Maximilian University, Münich, Germany) kindly provided further information on Cerithium? lateplicatum. Mariel Ferrari (Instituto Patagónico de Geología y Paleontología, Puerto Madryn, Argentina) and Alexander Nützel are acknowledged for their constructive reviews. The photographic work has been executed by Stefano Castelli and the line drawings by Nicola Michelon (both University of Padua, Italy). The paper was supported by the fund GATT-SID17-01-2017 of the University of Padua.
References
Ammon, L. von 1878. Die Gastropoden des Hauptdolomites und Plattenkalkes der Alpen. Abhandlungen des Zoologisch Mineralogischen Vereins Regensburg 11: 1–72.
Ammon, L. von 1893. Die Gastropoden des Hochfellnkalkes und über Gastropoden-Reste aus Ablagerungen von Adnet, vom Monte Nota und den Raibler Schichten. Geognostische Jahreshefte 5: 161–221.
Bandel, K. 1993. Caenogastropoda during Mesozoic times. Scripta Geologica, Special Issue 2: 7–56.
Bandel, K. 1994. Comparison of Upper Triassic and Lower Jurassic gastropods from the Peruvian Andes (Pucará Group) and the Alps (Cassian Formation). Palaeontographica A 233: 127–160.
Bandel, Κ. 1995. Mathildoidea (Gastropoda, Heterostropha) from the Late Triassic St Cassian Formation. Scripta Geologica 111: 1–83.
Bandel, K. 2006. Families of the Cerithioidea and related superfamilies (Palaeo-Caenogastropoda; Mollusca) from the Triassic to the Recent characterized by protoconch morphology—including the description of new taxa. Freiberger Forschungshefte C 511: 59–138.
Bandel, K. and Dockery, D.T. III 2012. Protoconch characters of Late Cretaceous Latrogastropoda (Neogastropoda and Neomesogastropoda) as an aid in the reconstruction of the phylogeny of the Neogastropoda. Freiberger Forschungshefte C 542: 93–128.
Bandel, K., Gründel, J., and Maxwell, P.A. 2000. Gastropods from the upper Early Jurassic/early Middle Jurassic of Kaiwara Valley, North Canterbury, New Zealand. Freiberger Forschungshefte C 490: 67–132.
Barker, M.J. and Munt, M.C. 1999. Gastropods. In: A. Swift and D.M. Martill (eds.), Fossils of the Rhaetian Penarth Group. Palaeontological Association Field Guides to Fossils 9: 75–82.
Barrier, E. and Vrielinck, B. (eds.) 2008. Palaeotectonic Maps of the Middle East (MEBE Program). Tectono-sedimentary-palinspastic Maps from Late Norian to Pliocene. 14 maps. CCGM/CGMW, Paris.
Begg, J.G. and Grant-Mackie, J.A. 2003. New Zealand and New Caledonian Triassic Pleurotomariidae (Gastropoda, Mollusca). Journal of the Royal Society of New Zealand 33: 223–268. Crossref
Böhm, G. 1884. Beiträge zur Kenntniss der Grauen Kalke in Venetien. Zeitschrift der Deutschen Geologischen Gesellschaft 36: 737–782.
Borghi, P. 1938. Su alcune nuove località fossilifere del Trias superiore Lombardo. Atti della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale in Milano 77: 21–35.
Bouchet, P. and Rocroi, J.-P. (eds.) 2005. Classification and nomenclator of gastropod families. Malacologia 47: 1–397.
Bouchet, P., Rocroi, J.-P, Hausdorf, B., Kaim, A., Kano, Y., Nützel, A., Parkhaev, P., Schrödl, M., and Strong, E.E. 2017. Revised classification, nomenclator and typification of gastropod and monoplacophoran families. Malacologia 61: 1–526. Crossref
Bourrouilh, R. 1966. Gastéropodes du Lias inférieur et moyen du domaine atlasique marocain. Notes et Mémoires du Service Géologique du Maroc 196: 25–73.
Brunet, M.-F., Wilmsen, M., and Granath, J.W. (eds.) 2009. South Caspian to Central Iran basins. Geological Society, London, Special Publications 312: 1–352. Crossref
Chiesa, C. 1949. Contributo alla conoscenza del Retico della Lombardia occidentale. Rivista italiana di Paleontologia e di Stratigrafia 60: 18–35.
Conti, G. 1954. Stratigrafia e paleontologia della Val Solda (Lago di Lugano). Memorie descrittive della Carta Geologica d’Italia 30: 1–248.
Cossmann, M. 1902. Paléontologie. In: C. Chartron and M. Cossmann (eds.), Note sur l’Infralias de la Vendée et spécialement sur un gisement situé dans la commune du Simon-la-Vineuse. Bulletin de la Société Géologique de France, 4ème série 2: 166–202.
Cossmann, M. 1907. Troisième note sur le Bathonien de Saint-Gaultier (Indre). Bulletin de la Société géologique de France, 4ème série 7: 225–253.
Cossmann, M. 1913. Contribution à la paléontologie française des terrains jurassiques. III. Cerithiacea et Loxonematacea. Mémoires de la Société Géologique de France 46: 1–263.
Dal Piaz, G. 1909. Nuovo giacimento fossilifero del Lias inferiore dei Sette Comuni (Vicentino). Mémoires de la Société Paléontologique Suisse 35 (for 1908): 1–10.
De Gregorio, A. 1886. Essai paléontologique à propos de certains fossiles secondaires de la contrée Casale-Ciciù provenant probablement de l’étage Alpinien De Greg. Annales de Géologie et de Paléontologie 4: 1–7.
Dercourt, J., Gaetani, M., Vrielynck, B., Barrier, E., Biju-Duval, B., Brunet, M.-F., Cadet, J.-P., Crasquin, S., and Sandulescu, M. (eds.) 2000. Atlas Peri-Tethys Palaeogeographical Maps. i–xx + 269 pp., 24 maps. CCGM⁄CGMW, Paris.
Desio, A. 1929. Studi geologici sulla regione dell’Albenza (Prealpi Bergamasche). Memorie della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano 10: 1–156.
Di Stefano, G. 1887. Sul Lias inferiore di Taormina e dei suoi dintorni. Giornale di Scienze Naturali ed Economiche di Palermo 18: 46–184.
Dittmar, A. von 1864. Die contorta-zone (Zone der Avicula contorta Portl.) ihre verbreitung und ihre organischen Einschlüsse. 217 pp. Straub, München.
d’Orbigny, A. 1850. Prodrome de Paléontologie stratigraphique universelle des animaux mollusques et rayonnés, faisant suite au cours élémentaire de paléontologie et de géologie stratigraphiques. Vol. 1. lx + 394 pp. Masson, Paris. Crossref
d’Orbigny, A. 1851. Livraison 66. In: A. d’Orbigny A. (1851–1860). Paléontologie française. Description zoologique et géologique de tous les animaux mollusques et rayonnés fossiles de France, comprenant leur application a la reconnaissance des couches. Terrains jurassiques. Tome 2, contenant les gastéropodes, 33–48. Masson, Paris.
Dresnay, R. du 1966. Observations stratigraphiques au sujet des gisements de gastéropodes liasiques du Causse moyen-atlasique et du Haut Atlas oriental, étudiés pas R. Bourrouilh. Notes et Mémoires du Service Géologique du Maroc 196: 1–23.
Dubar, G. 1948. La faune domeriénne du Djebel Bou-Dahar (prés de Beni-Tajjite). Notes et Mémoires du Service Géologique du Maroc 68: 1–248.
Escher von der Linth, A. 1853. Geologische Bemerkungen über das nördliche Vorarlberg und einige angrenzenden Gegenden. Neue Denkschriften der Allgemeinen Schweizerischen Gesellschaft für die Gesammten Naturwissenschaften 13: 1–135. Crossref
Eudes-Deslongchamps, J.C.A. 1843. Mémoire sur les Mélanies fossiles des terrains secondaires du Calvados. Mémoires de la Société linnéenne de Normandie 7: 215–230.
Ferrari, S.M. 2015. Systematic revision of Late Triassic marine gastropods from Central Perú: considerations on the Late Triassic/Early Jurassic faunal turnover. Andean Geology 42: 71–96.
Fischer, J.-C. 1964. Contribution à l’étude de la faune bathonienne dans la vallée de la Creuse (Indre). Annales de Paléontologie, Invertébrés 50: 21–101.
Fischer, J.-C. and Weber, C. 1997. Révision Critique de la Paléontologie Française d’Alcide d’Orbigny (incluant la réédition de l’original). Volume II, Gastropodes Jurassiques. 300 pp. Muséum National d’Histoire Naturelle and Masson, Paris.
Fischer-Ooster, C. von 1870. Ueber die Rhätische Stufe in der Umgegend von Thun. Mittheilungen der Naturforschenden Gesellschaft in Bern 684 (for 1869): 32–99.
Fucini, A. 1895. Fauna dei calcari bianchi ceroidi con Phylloceras cylindricum Sow. sp. del Monte Pisano. Atti della Società Toscana di Scienze Naturali, Memorie 14: 125–351.
Fürsich, F.T., Hautmann, M., Senowbari-Daryan, B., and Seyed-Emami, K. 2005. The Upper Triassic Nayband and Darkuh formations of east-central Iran: stratigraphy, facies patterns and biota of extensional basins on an accreted terrane. Beringeria 35: 53–133.
Galli, M.T., Jadoul, F., Bernasconi, S.M, and Weissert, H. 2005. Anomalies in global carbon cycling and extinction at the Triassic/Jurassic boundary: evidence from a marine C-isotope record. Palaeogeography, Palaeoclimatology, Palaeoecology 216: 203–214. Crossref
Galli, M.T., Jadoul, E., Bernasconi, S.M., Cirilli, S., and Weissert, H. 2007. Stratigraphy and palaeoenvironmental analysis of the Triassic–Jurassic transition in the Western Southern Alps (Northern Italy). Palaeogeography, Palaeoclimatology, Palaeoecology 244: 52–70. Crossref
Gatto, R. and Monari, S. 2010. Pliensbachian gastropods from Venetian Southern Alps (Italy) and their palaeobiogeographical significance. Palaeontology 53: 771–802. Crossref
Gemmellaro, G.G. 1878. Sui fossili del calcare cristallino delle montagne del Casale e di Bellampo nella provincia di Palermo. Giornale di Scienze Naturali ed Economiche di Palermo 13: 116–212.
Gründel, J. 1998. Archaeo- und Caenogastropoda aus dem Dogger Deutschlands und Nordpolens. Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie) 260: 1–39. Crossref
Gründel, J. 1999. Gastropoden aus dem höheren Lias von Grimmen, Vorpommern (Deutschland). Archiv für Geschiebekunde 2: 629–672.
Gründel, J. 2001. Neritimorpha und weitere Caenogastropoda (Gastropoda) aus dem Dogger Norddeutschlands und des nordwestlichen Polens. Berliner geowissenschaftliche Abhandlungen E36: 45–99.
Gründel, J. 2005. Gastropoden aus dem oberen Callovium (Lamberti-Zone) der Tongrube Dubki bei Saratov, Russische Plattform. Zitteliana A 45: 65–85.
Gründel, J. and Nützel, A. 2013. Evolution and classification of Mesozoic mathildoid gastropods. Acta Palaeontologica Polonica 58: 803–826. Crossref
Gründel, J. and Nützel, A. 2015. Gastropods from the upper Pliensbachian (Amaltheenton-Formation) from NE Bavaria (vicinity of Stauf/Dörlbach/Altdorf) (Franconia, Southern Germany). Zitteliana A 55: 45–76.
Guzhov, A.V. 2004. Jurassic gastropods of European Russia (Orders Cerithiiformes, Bucciniformes, and Epitoniiformes). Paleontological Journal 38 (Supplement 5): S457–S562.
Haas, O. 1909. Bericht über neue Aufsammlungen in den Zlambachmergeln der Fischerwiese bei Alt-Aussee. Beiträge zur Paläontologie und Geologie Österreich-Ungarns und des Orients 22: 144–167.
Hasibuan, F. and Grant-Mackie, J.A. 2007. Triassic and Jurassic gastropods from the Misool Archipelago, Eastern Indonesia. Jurnal Sumber Daya Geologi 17: 257–272.
ICZN 1999. International Code of Zoological Nomenclature. Fourth edition adopted by the international union of biological sciences. 306 pp. International Trust for Zoological Nomenclature, London.
Jadoul, F. and Galli, M.T 2008. The Hettangian shallow water carbonates after the Triassic/Jurassic biocalcification crisis: the Albenza Formation in the western Southern Alps. Rivista ltaliana di Paleontologia e Stratigrafia 114: 453–470.
Jadoul, F., Calabrese, L., Galli, M.T., and Gnaccolini, M. 2005. Stratigraphy of Rhaetian to lower Sinemurian carbonate platforms in western Lombardy (Southern Alps, Italy): paleogeographic implications. Rivista Italiana di Paleontologia e Stratigrafia 111: 285–303.
Jadoul, F., Galli, M.T., Berra, F., Cirilli, S., Ronchi, P., and Paganoni, A. 2004. Field guide book excursion P68. The Late Triassic–Early Jurassic of the Lombardy Basin: Stratigraphy, palaeogeography, and Palaeontology. In: L. Guerrieri, I. Rischia, and L. Serva (eds.), 32nd International Geological Congress, Florence (Italy) 20–28 August, 2004. Memorie Descrittive della Carta geologica d’Italia 63 (6): 1–36.
Jadoul, F., Masetti, D., Cirilli, S., Berra, F., Claps, M., and Frisia, S. 1994. Excursion B1. Norian–Rhaetian stratigraphy and paleogeographic evolution of the Lombardy Basin (Bergamasc Alps). In: G. Carannante and R. Tonielli (eds.), Post Meeting Field Trip Guidebook, 15th IAS Regional Meeting, April 1994, Ischia, Italy, 1–38. De Fede, Napoli.
Kaim, A. 2004 . The evolution of conch ontogeny in Mesozoic open sea gastropods. Palaeontologia Polonica 62: 1–183.
Kaim, A. and Beisel, A.L. 2005. Mesozoic gastropods from Siberia and Timan (Russia). Part 2: Neogastropoda and Heterobranchia. Polish Polar Research 26: 41–64.
Kaim, A., Jurkovšek, B., and Kolar-Jurkovšek, T. 2006. New associations of Carnian gastropods from Mežica region in the Karavanke Mountains of Slovenia. Facies 52: 469–482. Crossref
Kittl, E. 1894. Die Gastropoden der Schichten von St. Cassian der südalpinen Trias. III. Theil. Annalen des Kaiserlich-königlichen Naturhistorischen Hofmuseums 9: 143–277.
Kittl, E. and Spengler, E. 1916. Halorellenkalke vom Vorderen Gosausee. Annalen des Naturhistorischen Museums in Wien 30: 51–54.
Klipstein A. 1843. Petrefactologischer Theil. Beschreibung neuer Versteinerungen von St. Cassian und einiger anderer Localitäten, Erster Abschnitt. Mollusken. In: A. Klipstein A. 1843–1845. Beiträge zur Geologischen Kenntniss der Östlichen Alpen, 101–207. Heyer, Giessen.
Kühn, O. 1942. Zur Kenntnis des Rhät von Voralberg. Mitteilungen der Geologischen Gesellschaft in Wien 33 (for 1940): 111–157.
Kutassy, A. 1933. Su alcuni Megalodus del Monte Campo dei Fiori (Varese). Atti della Società Italiana di Scienze Naturali 72: 232–241.
Laube, G.C. 1868. Die Fauna der Schichten von St. Cassian. III. Abtheilung. Gastropoden. I. Hälfte. Denkschriften der Kaiserlichen Akademie der Wissenschaften. Mathematisch-naturwissenschaftliche Klasse 28: 29–94.
Lepsius, R. 1878. Das westliche Südtirol. Geologisch Dargestellt. 375 pp. Hertz, Berlin.
Meier, H. and Meiers, K. 1988. Die Gastropodenfauna der “Angulata-zone” des Steinbruchs “Reckingerwald” bei Brouch. Travaux Scientifiques du Musée National d’Histoire Naturelle de Luxembourg 13: 1–87.
Monari, S., Valentini, M., and Conti, M.A. 2011. Earliest Jurassic patellogastropod, vetigastropod, and neritimorph gastropods from Luxembourg with considerations on the Triassic–Jurassic faunal turnover. Acta Palaeontologica Polonica 56: 349–384. Crossref
Nützel, A. 2005. Recovery of gastropods in the Early Triassic. Comptes Rendus Palevol 4: 501–515. Crossref
Nützel, A. 2010. A review of the Triassic gastropod genus Kittliconcha Bonarelli, 1927. Implications for the phylogeny of Caenogastropoda. Zitteliana A 50: 9–24.
Nützel, A. and Gründel, J. 2007. Two new gastropod genera from the Early Jurassic (Pliensbachian) of Franconia (South Germany). Zitteliana A 47: 59–67.
Nützel, A. and Gründel, J. 2015. Early Jurassic (Pliensbachian) gastropods from Franconia, Southern Germany. Palaeontographica A 305: 1–87. Crossref
Nützel, A. and Senowbari-Daryan, B. 1999. Gastropods from the Late Triassic (Norian–Rhaetian) Nayband Formation of central Iran. Beringeria 23: 93–132.
Nützel, A., Aghababalou, B., and Senowbari-Daryan, B. 2012. Gastropods from the Norian (Late Triassic) Nayband Formation near Natanz (Iran). Bulletin of Geosciences 87: 53–65. Crossref
Nützel, A., Hamedani, E., and Senowbari-Daryan, B. 2003. Some Late Triassic gastropods from the Nayband Formation in Central Iran. Facies 48: 127–134. Crossref
Nützel, A., Mannani, M., Senowbari-Daryan, B., and Yazdi, M. 2010. Gastropods from the Late Triassic Nayband Formation (Iran), their relationships to other Tethyan faunas and remarks on the Triassic gastropod body size problem. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 256: 213–228. Crossref
Osswald, K. 1930. Über einige Rhätfossilien aus dem Risserkogelgebiet (südlich Tegernsee). Jahrbuch der Preussischen Geologischen Landesanstalt zu Berlin 50 (1929): 733–750.
Parona, C.F. 1932. Di un particolare carattere paleontologico del Retico di Riva di Solto (Lago d’Iseo). Atti della Reale Accademia delle Scienze di Torino, classe di scienze fisiche, matematiche e naturali 67: 403–408.
Riedel, F. 2000. Ursprung und Evolution der ‘höheren’ Caenogastropoda: eine paläobiologische Konzeption. Berliner Geowissenschaftliche Abhandlungen E 32: 1–240.
Schäfer, R. 1888. Über die geologischen Verhältnisse des Karwendels in der Gegend von Hinterriss und um den Scharfreiter. Inaugural-Dissertation zur Erlangung der Doctorwürde der philosophischen Facultät der K. Ludwigs-Maximilians-Universität zu München. 37 pp. Kutzner, München.
Schröder, M. 1995. Frühontogenetische Schalen jurassischer und unterkretazischer Gastropoden aus Norddeutschland und Polen. Paläontographica A 238: 1–95.
Stoppani, A. 1857. Studii geologici e paleontologici sulla Lombardia. 458 pp. Turati, Milano.
Stoppani, A. 1860–1865. Paléontologie Lombarde, ou, Description des fossiles de Lombardie, publiée à l’aide de plusieurs savants. 3.e Série, Géologie et paléontologie des couches à “Avicula contorta” en Lombardie, comprenant des aperçus sur l’étage infraliasien en Lombardie et en Europe en général, et deux monographies des fossiles appartenant à la zone supérieure et à la zone inférieure des couches à “Avicula contorta” en Lombardie, une note supplémentaire et deux appendices sur l’infralias du versant N.-O. des Alpes et sur les faunes aux limites supérieures et inférieures des couches à “A. contorta”. 267 pp. Bernadoni, Milan.
Szabó, J. 1983. Lower and Middle Jurassic gastropods from the Bakony Montains (Hungary). Part 5. Supplement to Archaeogastropoda; Caenogastropoda. Annales Historico-Naturales Musei Nationalis Hungarici 75: 27–46.
Tackett, L.S. and Bottjer, D.J. 2012. Faunal succession of Norian (Late Triassic) level-bottom benthos in the Lombardian Basin: implications for the timing, rate, and nature of the Early Mesozoic Marine Revolution. Palaios 27: 585–593. Crossref
Terquem, O. 1855. Paléontologie de l’étage inférieur de la formation liasique de la province du Luxembourg, Grand-Duché (Hollande), et de l’Hettange, du département de la Moselle. Mémoires de la Société Géologique de France, 2ème série 5: 219–343.
Teruzzi, G. 2015. The Stoppani collection of large bivalves (Bivalvia, Megalodontida) from the Upper Triassic of Lombardy, Italy. Natural History Sciences 2: 15–24. Crossref
Webster, N.B. and Vermeij, G.J. 2017. The varix: evolution, distribution, and phylogenetic clumping of a repeated gastropod innovation. Zoological Journal of the Linnean Society 180: 732–754. Crossref
Wiedenmayer, F. 1963. Obere Trias bis mittlerer Lias zwischen Saltrio und Tremona (Lombardische Alpen). Die Wechselbeziehungen zwischen Stratigraphie, Sedimentologie und syngenetischer Tektonik. Eclogae Geologicae Helvetiae 56: 529–640.
Wilmsen, M., Fürsich, F.T., Seyed-Emami, K., Reza Majidifard, M., and Taheri, J. 2009. The Cimmerian Orogeny in northern Iran: tectono-stratigraphic evidence from the foreland. Terra Nova 21: 211–218. Crossref
Winkler, G. 1859. Die schishten der Avicula contorta inner- und ausserhalb der Alpen. 51 pp. Joh. Palm, München.
Winkler, G. 1861. Der Oberkeuper in den Bayrischen Alpen. Zeitschrift der Deutschen Geologischen Gesellschaft 13: 459–521.
Zapfe, H. 1949. Eine rhätische Fauna aus dem Gebiet des Eibenberges bei Ebensee in Oberösterreich. Jahrbuch des Oberösterreichischen Musealvereines 94: 235–257.
Zapfe, H. 1962. Beiträge zur Paläontologie der nordalpinen Riffe. Ein Massenvorkommen von Gastropoden im Dachsteinkalk des Tennengebirges, Salzburg. Annalen des Naturhistorischen Museums Wien 65: 57–69.
Zapfe, H. 1963. Beiträge zur Paläontologie der nordalpinen Riffe. Zur Kenntnis der Fauna des oberrhätischen Riffkalkes von Adnet, Salzburg. (Exklus. Riffbildner). Annalen des Naturhistorischen Museums Wien 66: 207–259.
Zapfe, H. 1965. Beiträge zur Paläontologie der nordalpinen Riffe. Die Fauna der “erratischen Blöcke” auf der Falmbergalm bei Gosau, Oberösterreich (Brachiopoda, Scaphopoda, Gastropoda, Cephalopoda). Annalen des Naturhistorischen Museums Wien 68: 279–308.
Zapfe, H. 1967. Beiträge zur Paläontologie der nordalpinen Riffe. Die Fauna der Zlambach-Mergel der Fischerwiese bei Aussee, Steiermark (exkl. Coelenterata und Mikrofossilien). Annalen des Naturhistorischen Museums in Wien 71: 413–480.
Zardini, R. 1978. Fossili cassiani (Trias medio-superiore). Atlante dei gasteropodi della formazione di S. Cassiano raccolti nella regione dolomitica attorno a Cortina d’Ampezzo. 55 pp. Edizioni Ghedina, Cortina d’Ampezzo.
Acta Palaeontol. Pol. 66 (1): 193–206, 2021
https://doi.org/10.4202/app.00792.2020