
The giant pliosaurid that wasn’t—revising the marine reptiles from the Kimmeridgian, Upper Jurassic, of Krzyżanowice, Poland
DANIEL MADZIA, TOMASZ SZCZYGIELSKI, and ANDRZEJ S. WOLNIEWICZ
Madzia, D., Szczygielski, T., and Wolniewicz, A.S. 2021. The giant pliosaurid that wasn’t—revising the marine reptiles from the Kimmeridgian, Upper Jurassic, of Krzyżanowice, Poland. Acta Palaeontologica Polonica 66 (1): 99–129.
Marine reptiles from the Upper Jurassic of Central Europe are rare and often fragmentary, which hinders their precise taxonomic identification and their placement in a palaeobiogeographic context. Recent fieldwork in the Kimmeridgian of Krzyżanowice, Poland, a locality known from turtle remains originally discovered in the 1960s, has reportedly provided additional fossils thought to indicate the presence of a more diverse marine reptile assemblage, including giant pliosaurids, plesiosauroids, and thalattosuchians. Based on its taxonomic composition, the marine tetrapod fauna from Krzyżanowice was argued to represent part of the “Matyja-Wierzbowski Line”—a newly proposed palaeobiogeographic belt comprising faunal components transitional between those of the Boreal and Mediterranean marine provinces. Here, we provide a detailed re-description of the marine reptile material from Krzyżanowice and reassess its taxonomy. The turtle remains are proposed to represent a “plesiochelyid” thalassochelydian (Craspedochelys? sp.) and the plesiosauroid vertebral centrum likely belongs to a cryptoclidid. However, qualitative assessment and quantitative analysis of the jaws originally referred to the colossal pliosaurid Pliosaurus clearly demonstrate a metriorhynchid thalattosuchian affinity. Furthermore, these metriorhynchid jaws were likely found at a different, currently indeterminate, locality. A tooth crown previously identified as belonging to the thalattosuchian Machimosaurus is here considered to represent an indeterminate vertebrate. The revised taxonomy of the marine reptiles from Krzyżanowice, as well as the uncertain provenance of the metriorhynchid specimen reported from the locality, cast doubt on the palaeobiogeographic significance of the assemblage.
Key words: Pliosauridae, Metriorhynchidae, Crocodylomorpha, Thalassochelydia, Kimmeridgian, Jurassic, Poland.
Daniel Madzia [daniel.madzia@gmail.com; ORCID: https://orcid.org/0000-0003-1228-3573], Tomasz Szczygielski [t.szczygielski@twarda.pan.pl; ORCID: https://orcid.org/0000-0001-5108-8493], and Andrzej S. Wolniewicz [wolniewicz@twarda.pan.pl; ORCID: https://orcid.org/0000-0002-6336-8916], Institute of Paleobiology, Polish Academy of Sciences, ul. Twarda 51/55, 00-818 Warszawa, Poland.
Received 21 July 2020, accepted 14 September 2020, available online 12 January 2021.
Copyright © 2021 D. Madzia et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Late Jurassic was an important time interval in the evolutionary history of several clades of Mesozoic marine reptiles. Some groups, such as ophthalmosaurid ichthyosaurs, plesiosaurs (pliosaurids and cryptoclidids, in particular), and thalattosuchian crocodylomorphs had already become globally distributed and underwent significant taxonomic and ecomorphological diversification by the Late Jurassic (e.g., Bardet et al. 2014; Stubbs and Benton 2016; Foffa et al. 2018c; Zverkov et al. 2018; Fernández et al. 2019). Other marine reptile clades, such as thalassochelydian turtles and pleurosaurs (rhynchocephalians) were more restricted in their geographic distribution and represented some of the earliest-diverging, fully marine representatives within Testudinata and Lepidosauria, respectively (Bardet et al. 2014; Anquetin et al. 2017).
In Europe, abundant and well-preserved fossils of Late Jurassic marine reptiles are known from the Kimmeridge Clay Formation (Kimmeridgian) of the United Kingdom (e.g., Owen 1842; Seeley 1869, 1875; Lydekker 1889; Andrews 1921; Tarlo 1960; Brown 1981; Brown et al. 1986; Taylor and Cruickshank 1993; Sassoon et al. 2012; Young et al. 2013b; Benson et al. 2013; Benson and Bowdler 2014; Pérez-García 2015b, c; Püntener et al. 2015; Anquetin and Chapman 2016; Moon and Kirton 2018), the Reuchenette Formation (Kimmeridgian) of Switzerland (e.g., Rütimeyer 1873; Bräm 1965; Meyer 1994; Comment et al. 2015; Püntener et al. 2015, 2017a, b; Sullivan and Joyce 2017; Raselli and Anquetin 2019; Anquetin and Püntener 2020), the “Solnhofen Limestone” (Tithonian) of Southern Germany (e.g., Meyer 1839a; Parsons and Williams 1961; Gaffney 1975b; Bardet and Fernández 2000; Dupret 2004; Young and de Andrade 2009; de Andrade et al. 2010; Young et al. 2012; Anquetin and Joyce 2014; Arratia et al. 2015; Anquetin et al. 2017; Bever and Norell 2017), several Kimmeridgian and Tithonian localities of France (e.g., Thiollière 1850; Meyer 1860; Lortet 1892; Rieppel 1980; Broin 1994; Lapparent de Broin et al. 1996; Bardet et al. 1997; Pérez-García 2015b) and Iberia (e.g., Lapparent de Broin and Murelaga 1999; Pérez-García and Ortega 2011; Pérez-García 2015c), as well as several fossil-bearing horizons spanning the Volgian (Tithonian–lowermost Berriasian) of European Russia (e.g., Zverkov et al. 2015a, b, 2018; Arkhangelsky et al. 2018; Zverkov and Efimov 2019; Zverkov and Prilepskaya 2019) and the Slottsmøya Member of the Aghardfjellet Formation of Spitsbergen (e.g., Druckenmiller et al. 2012; Knutsen et al. 2012a–c; Roberts et al. 2014, 2020; Delsett et al. 2019).
Recent research has provided evidence for extensive faunal interchange between Late Jurassic seas located in Northern, Western and Eastern Europe (e.g., Arkhangelsky et al. 2018; Zverkov and Efimov 2019), with the seaway covering Poland acting as an important dispersal route between them (Tyborowski 2016; Tyborowski and Błażejowski 2019a, b). Therefore, marine reptile fossils from the Upper Jurassic of Poland have the potential for providing data important for testing previously proposed hypotheses on the dispersal and taxonomic diversification of European Late Jurassic marine reptiles. However, Late Jurassic marine reptile fossils occur rarely in Poland and are often fragmentary or very incomplete (Fig. 1 and Table 1). This hinders their correct taxonomic identification, crucial for the proper placement of these fossils into biogeographic and evolutionary contexts.

Fig. 1. Geographic distribution of Jurassic marine reptile fossil occurrences in Poland. See Table 1 for details. Silhouettes obtained from phylopic.org: Geosaurinae (Dmitry Bogdanov, CC BY 3.0), Ichthyosauria, Metriorhynchinae, and Teleosauroidea (Gareth Monger, CC BY 3.0), indeterminate reptile clade (public domain), Plesiosauroidea (Adam Stuart Smith, CC BY-SA 3.0), Pliosauridae (Nobu Tamura, CC BY-SA 3.0), and Testudinata (public domain).
Several localities yielding fragmentary fossils of Late Jurassic marine reptiles were reported from the territory of Poland during the 19th and 20th centuries (Fig. 1 and Table 1). However, the majority of these findings were only briefly described or merely mentioned in Polish or German literature (e.g., Jentzsch 1884; Dames 1888; Gallinek 1895, 1896; Deecke 1907; Hirszberg 1924; Molenda 1997; see Table 1) and have received little scientific attention until very recently. Nevertheless, the last decade saw a marked rise of research interest in Late Jurassic marine reptiles from Poland. Skrzycki (2010) reported a specimen comprising a series of cross-sections through the rostrum of an indeterminate ophthalmosaurid ichthyosaur discovered in Morawica quarry (Oxfordian), which was formally described nine years later (Tyborowski et al. 2019). Three-dimensionally preserved, partial skeletons of the ophthalmosaurid ichthyosaur Undorosaurus kielanae (Tyborowski, 2016) (Delsett et al. 2019; see also Zverkov and Prilepskaya 2019) and the pancryptodiran turtle Owadowia borsukbialynickae Szczygielski, Tyborowski, and Błażejowski, 2018, as well as fragmentary remains of crocodylomorphs (Błażejowski and Tyborowski 2016; Błażejowski et al. 2016; Tyborowski 2016; Tyborowski et al. 2016), were reported from the Tithonian of Owadów-Brzezinki (Kin and Błażejowski 2012; Kin et al. 2012, 2013). A partial skeleton of an ichthyosaur was also reported, but not yet described, from the Kimmeridgian of Szczerców (Grabowska 2013; Maliszewski 2017; Krzeczyńska et al. 2020). Finally, several teeth collected from Zalas (originaly reported by Molenda 1997), Wapiennik, and Częstochowa (initially described by Maryańska 1972) were recently revised as belonging to thalassophonean pliosaurids (Lomax 2015; Tyborowski 2019).
Table 1. Historical material of marine reptiles from the Jurassic of Poland. Abbreviations: A, Aalenian; B, Bathonian; C, Callovian; O, Oxfordian; K, Kimmeridgian; T, Tithonian; * Piotr Sterkowicz, personal communication 2020; ** unpublished; *** this study. References: 1, Hoffmann 2005; 2, Hirszberg 1924; 3, Deecke 1907; 4, Hoffmann and Bickelmann 2008; 5, Dzik 1992; 6, Dzik 1997; 7, Young et al. 2014; 8, Dzik 2003; 9, Dzik 2011; 10, Maryańska 1972; 11, Tyborowski 2019; 12, Jentzsch 1884; 13, Young et al. 2013a; 14, Gallinek 1895; 15, Gallinek 1896; 16, Zatoń 2007; 17, Rehbinder 1913; 18, Hoffmann 2007; 19, Borsuk-Białynicka and Młynarski 1968; 20, Lapparent de Broin and Murelaga 1999; 21, Lapparent de Broin 2001; 22, Joyce et al. 2011; 23, Anquetin et al. 2017; 24, Joyce 2017; 25, Tyborowski and Błażejowski 2019b; 26, Tyborowski and Błażejowski 2019a; 27, Skrzycki 2010; 28, Tyborowski et al. 2019; 29, Tyborowski et al. 2016; 30, Błażejowski et al. 2016; 31, Błażejowski and Tyborowski 2016; 32, Tyborowski 2016; 33, Zverkov and Efimov 2019; 34, Zverkov and Jacobs 2020; 35, Szczygielski et al. 2018; 36, Kin et al. 2012; 37, Błażejowski et al. 2014; 38, Feldmann et al. 2015; 39, Tyborowski 2017; 40, Wierzbowski et al. 2016; 41, Pusch 1837; 42, Krzeczyńska et al. 2020; 43, Maliszewski 2017; 44, Grabowska 2013; 45, Groß 1944; 46, Preussner 1886; 47, Sadebeck 1865; 48, Dames 1888; 49, Krebs 1967; 50, Molenda 1997; 51, Borszcz and Zatoń 2009; 52, Lomax 2015; 53, Ketchum and Benson 2011. A The specimen list is compiled based on the literature, it may thus not be exhaustive. Please note that to retain clarity and minimize taxon splitting, the taxonomic revisions are not considered at the specimen level (which would often be impossible due to complete specimen lists rarely provided by the authors), but rather whole groups of specimens as defined by the original authors (or the first revising authors to mention specimen numbers), unless the data are sufficient to act otherwise. In certain cases the intent of the revising authors is ambiguous and this should be taken into account. B Deecke (1907) mentioned “teeth and dermal plates of Machimosaurus and Steneosaurus” from Czarnogłowy. Since no specimen numbers nor illustrations were provided, it is not possible to verify whether these specimens were later revised by Hoffmann (2005) and Hoffmann and Bickelmann (2008), but this seems likely. C The specimens GG303-43 and GG303-44 are described by Hoffmann (2005) as vertebral centra of Thalattosuchia indet., but under the same numbers figured are two teeth referred to Metriorhynchidae indet. (Hoffmann 2005: tab. 5, figs. 2, 3). D Dzik (1992: fig. 9.20C) figures specimen GG303-30, a partial mandible (captioned as a palate of Machimosaurus sp.), identified later by Hoffmann (2005), Hoffmann and Bickelmann (2008), and Young et al. (2014) as belonging to Steneosaurus sp. Dzik (1997: fig. 10.5) provided the same photograph (this time captioned as palate of Machimosaurus hugii) and additionally mentioned teeth and skeleton fragments of Machimosaurus sp. In the newer editions (Dzik 2003, 2011) the photograph is no longer present, but the teeth and skeleton fragments of Machimosaurus sp. remain mentioned. None of these specimens are figured and no specimen numbers are given by Dzik, but it seems likely that at least part of the specimens interpreted by him as belonging to Machimosaurus sp. are in agreement with those identified as Machimosaurus sp. by Hoffmann (2005) and Hoffmann and Bickelmann (2008). E Tyborowski and Błażejowski (2019b) also mentioned jaws and skull fragments collected from the canary-yellow Nerinea limestone, but the specimen figured is MZ VIII Vr-72 from Załęcze Wielkie, earlier described and figured by Maryańska (1972). F Specimen ZPAL P.16/O-B/1 interpreted initially as a mandible of Pleurosaurus sp. (Kin et al. 2012) or ?Pleurosaurus sp. (Kin and Błażejowski 2012) was later reinterpreted as the actinopterygian fish Furo sp. (Błażejowski et al. 2014, 2015). G Dames (1888) also mentioned a tooth of Megalosaurus sp. from Wrzosowo; it seems likely that this specimen in fact belonged to some indeterminate marine reptile.
|
Locality |
Age |
Material |
Specimen numbersA |
Original taxonomic referral |
Revised taxonomic referral |
|
Bardy (Bartin) |
K |
bone fragment |
not given |
Reptilia indet.1 |
not revised |
|
Bolęcin |
C |
tooth |
not given |
Plesiosauroidea? indet.* |
not revised |
|
Brzustówka |
T |
vertebral centrum |
not given |
Ophtalmosaurus [sic!] sp.2 |
not revised |
|
vertebral centrum |
not given |
Cimoliosaurus
[sic!] |
not revised |
||
|
Czarnogłowy |
K |
hyoplastra, shell fragments |
GG303-25, GG303-26, GG303-27, GG303-28 |
Plesiochelys sp.3 |
Chelonioidea indet.1 |
|
Plesiochelyidae cf. Plesiochelys
|
|||||
|
vertebral centrum |
GG303-29 |
Ophthalmosauridae indet.1 |
cf. Brachypterygius cf. extremus4 |
||
|
teeth |
GG303-32, GG303-33 |
Steneosaurus jugleri1,3,4 |
not revised |
||
|
teeth, vertebrae |
GG303-31 |
Steneosaurus sp.1,4 |
not revised |
||
|
osteoderm |
GG303-41 |
Teleosauridae indet.1 |
Steneosaurus sp.4 |
||
|
vertebral centra |
GG303-43, GG303-44, GG303-45, GG303-46, GG303-47C |
Thalattosuchia indet.1 |
Steneosaurus sp.4 |
||
|
mandible fragment |
GG303-30 |
Machimosaurus sp.5,D |
Machimosaurus hugii6,D |
||
|
Steneosaurus sp.1,4,7 |
|||||
|
teeth |
GG303-43, |
Metriorhynchidae indet.1 |
Dakosaurus maximus4 |
||
|
teeth, mandible fragment, vertebrae |
GG303-34, GG303-35, GG303-36, GG303-40 |
Machimosaurus hugii5,6,8,D |
Machimosaurus sp.1 |
||
|
Machimosaurus hugii4,9,D |
|||||
|
teeth |
GPIT/RE/328, |
Machimosaurus
cf. |
not revised |
||
|
Częstochowa (Mirów) |
O |
skull fragments and teeth |
M.Cz. V 1293 |
Pliosaurus cf. andrewsi10 |
Thalassophonea indet.11 |
|
Faustianka |
?B |
? |
not given |
Plesiosauroidea indet.** |
not revised |
|
Inowrocław |
O |
teeth |
not given |
Dakosaurus maximus12 |
cf. Tyrannoneustes?13 |
|
? |
not given |
Pliosaurus giganteus12 |
not revised |
||
|
O–K |
unspecified remains |
not given |
Reptilia indet.14 |
not revised |
|
|
tooth |
not given |
Dacosaurus
[sic!] |
non Dakosaurus maximus16 |
||
|
cf. Tyrannoneustes?13 |
|||||
|
Jastrząb |
B |
tooth |
not given |
Plesiosauria indet.17 |
not revised |
|
Kłęby (Klemmen) |
O |
teeth |
GG303-37 |
Machimosaurus sp.1,18 |
not revised |
|
Krzyżanowice |
K |
shell fragments |
MZ VIII Vr-71 |
Tretosternon
aff. |
Testudinata indet.20–22 |
|
“Plesiochelyidae” indet.23–26 |
|||||
|
Craspedochelys? sp.*** |
|||||
|
skull fragments |
MZ VIII Vr-72 |
Pliosaurus sp.25,26 |
Metriorhynchidae indet.*** |
||
|
teeth |
not given |
Pliosauridae indet.26 |
not revised |
||
|
vertebral centra |
MZ VIII Vr-73 |
Elasmosauridae indet.25 |
Plesiosauroidea cf.
Elasmosauridae |
||
|
Cryptoclididae? indet.*** |
|||||
|
teethE |
ZPAL V. 69/1 |
Machimosaurus sp.26 |
Vertebrata indet.*** |
||
|
Małogoszcz |
K |
skull fragments |
not given |
Ichthyosauria indet.** |
not revised |
|
Mirów |
O |
tooth |
not given |
Pliosauridae indet.** |
not revised |
|
Młynka |
? |
tooth |
not given |
Pliosauridae indet.* |
not revised |
|
Morawica |
O |
skull fragments |
MNKI/P/162/1, MNKI/P/162/2 |
Ophthalmosaurus? sp.27 |
Ophthalmosauridae indet.28 |
|
Ogrodzieniec |
C or O |
tooth |
GIUS 8–3044 |
Metriorhynchidae indet.16 |
cf. Tyrannoneustes13 |
|
B |
tooth |
not given |
Pliosauridae indet.** |
not revised |
|
|
B |
vertebral centra |
not given |
Reptilia indet.** |
not revised |
|
|
Owadów- |
T |
partial skeleton, teeth, ribs |
WNG UŁ 3579-81 (GMUL 3579-81) |
Ichthyosauria indet.29 |
Ophthalmosauridae cf. Cryopterygius30,31 |
|
Cryopterygius kielanae32 |
|||||
|
Undorosaurus kielanae33,34 |
|||||
|
bones, teeth |
not given |
?Ichthyosauria non |
not revised |
||
|
skull fragment, osteoderms, teeth, partial skeleton |
not given |
Metriorhynchidae |
Thalattosuchia indet.32 |
||
|
mandible fragment, limb bones |
ZPAL V/O-B/1959 |
Cryptodira indet.29–32 |
Owadowia borsukbialynickae35 |
||
|
partial cranium, bone fragments |
not given |
Reptilia indet.30,36–40 |
not revisedF |
||
|
Piekło |
K |
vertebral centrum |
not given |
Mammalia indet.41 |
Cryptocleidus [sic!] kimmeridgensis2 |
|
Sułoszowa |
?O |
skull fragment |
not given |
Ichthyosauria indet.** |
not revised |
|
Szczerców |
K |
?partial skeleton |
not given |
Ichthyosauria indet.42–44 |
not revised |
|
Wapiennik |
O |
tooth |
not given |
Pliosaurus ferox45 |
not revised |
|
? |
tooth |
M.Cz. V 265 |
Thalassophonea indet.11 |
not revised |
|
|
Wolin (Wollin) |
A |
vertebra |
not given |
Ichthyosaurus sp.3,46 |
not revised |
|
tooth |
not given |
Plesiosaurus sp.3 |
not revised |
||
|
Wrzosowo |
K |
teeth |
GG303-38, GG303-39 |
Ichthyosaurus? sp.47 |
Machimosaurus ?sp. nov.48 |
|
Machimosaurus sp.1,3 |
|||||
|
Machimosaurus hugii4,49 |
|||||
|
tooth |
GG303-42 |
Dakosaurus sp.1 |
Dakosaurus maximus4 |
||
|
shell fragments |
not given |
Plesiochelys sp.48 |
Testudines indet.3 |
||
|
Zalas |
O |
teeth |
GM001, GM002, GM003 |
Ichthyosauria indet.50 |
Reptilia indet.51 |
|
Thalassophonea indet.52 |
|||||
|
bone fragment |
GM004 |
Ichthyosauria? indet.50 |
Reptilia indet.52 |
||
|
?O |
vertebral centrum |
not given |
Reptilia indet.50 |
not revised |
|
|
skull fragments |
not given |
Reptilia indet.** |
not revised |
||
|
Załęcze Wielkie |
O |
skull fragments |
MZ VIII Vr-72 |
Peloneustes sp.10 |
Teleosauridae indet.26,53 |
A diverse assemblage of marine reptiles from the upper Kimmeridgian of Krzyżanowice was recently described by Tyborowski and Błażejowski (2019a, b) based on the revision of historical specimens (Borsuk-Białynicka and Młynarski 1968) and the discovery of new fossil material. Although both the historical and newly discovered specimens from Krzyżanowice are very fragmentary, Tyborowski and Błażejowski (2019a, b) proposed that they represented a notable diversity of marine reptiles, including indeterminate “plesiochelyid” turtles, thalattosuchian crocodylomorphs (Machimosaurus sp.) and pliosaurid (Pliosaurus sp.) and elasmosaurid (Elasmosauridae indet.) plesiosaurs. The taxonomic composition of this assemblage was suggested to be similar to that of the Kimmeridge Clay Formation of the United Kingdom and both assemblages were established as part of the newly proposed “Matyja-Wierzbowski Line”—a transitional palaeobiogeographic region separating marine faunas from Northern and Southern Europe (Tyborowski and Błażejowski 2019a, b). However, when reading the accounts of Tyborowski and Błażejowski (2019a, b) we identified the following problems regarding the taxonomic identification, provenance, and historical background provided for the fossils described therein:
(i) The new material (MZ VIII Vr-71) referred to Plesiochelyidae indet., which was described and figured by Tyborowski and Błażejowski (2019a, b) was determined to be the same material previously described and figured by Młynarski and Borsuk-Białynicka (1968). The historical nature of the material was not recognised by Tyborowski and Błażejowski (2019a, b) and no attention was given to the original, controversial designation of MZ VIII Vr-71 as “Tretosternon aff. punctatum” Owen, 1842.
(ii) The specimen referred to as Pliosaurus sp. (MZ VIII Vr-72) was found to have a premaxillary count unusually high for Pliosaurus in particular, and Pliosauridae in general (Knutsen 2012; Benson et al. 2013; Madzia et al. 2019). Furthermore, the published photographs indicated the absence of the trihedral and subtrihedral tooth crown morphologies characteristic for Pliosaurus (see, e.g., Knutsen 2012; Benson et al. 2013; Zverkov et al. 2018). Finally, the specimen was also found to have the same catalogue number as a specimen from the Oxfordian of Załęcze Wielkie referred to Peloneustes sp. by Maryańska (1972) and later re-identified as a teleosaurid crocodylomorph by Ketchum and Benson (2011).
(iii) The assignment of an isolated centrum (“ZPAL V/KRZ-32”) to an elasmosaurid by Tyborowski and Błażejowski (2019a) was based on a very limited set of morphological comparisons. If correct, it would make the specimen the first Jurassic elasmosaurid and the oldest representative of the clade discovered to date, older by at least 10 Ma than the oldest known occurrences (see e.g., Sachs et al. 2017; Serratos et al. 2017; Madzia and Cau 2020). Interestingly, Tyborowski and Błażejowski (2019b) also gave an alternative, more generalized identification of this specimen as a plesiosauroid. Due to the editorial error of the venue in which the paper of Tyborowski and Błażejowski (2019a) was published, the specimen, at the moment of the completion of this study, held different catalogue numbers in the HTML (MZ VIII Vr-73) and the paginated PDF (“ZPAL V/KRZ-32”) versions of the paper, while being physically deposited in the MZ collection. However, the collection numbering and transfer were not discussed and agreed upon with the ZPAL collections curator. Later, it was confirmed that the specimen belonged to MZ collections, therefore the number ZPAL V/KRZ-32 is invalid (Jolanta Kobylińska, personal communication 2020). In the text we will use ZPAL numbers in quotation marks to emphasize the original, invalid collection assignment.
(iv) One of the blocks representing MZ VIII Vr-72, originally described as Peloneustes sp. by Maryańska (1972) and referred to Teleosauridae indet. by Ketchum and Benson (2011) from the Oxfordian of Załęcze Wielkie, was figured as a crocodylomorph from the Kimmeridigan of Krzyżanowice (Tyborowski and Błażejowski 2019b: fig. 6).
These controversies raise concerns not only about the correct taxonomic identification of the marine reptile specimens from Krzyżanowice, but also create doubts about the proper acknowledgment of the work of previous generations of palaeontologists on this material. Furthermore, they also raise the possibility that at least some of the specimens reported as discovered from Krzyżanowice were in reality not found at this locality.
The aim of this contribution is threefold: (i) to provide a detailed taxonomic revision of the marine reptile material from Krzyżanowice, for the purpose of using the information in future comparative studies of other marine reptiles from the Upper Jurassic of Europe; (ii) to provide a complete review of the history of findings at Krzyżanowice; and (iii) to revise the significance of the Krzyżanowice marine reptile assemblage for hypotheses related to the dispersal of marine reptiles in the Late Jurassic seas of the European Archipelago.
Institutional abbreviations.—GG, Institut für Geographie und Geologie, Ernst-Moritz-Arndt-Universität Greifswald, Germany; GIUS, Institute of Earth Sciences, Faculty of Natural Sciences, University of Silesia, Katowice, Poland; GMUL (WNG UŁ), Geological Museum, University of Łódź, Poland; GPIT, Palaeontological Collection of the University of Tübingen, Germany; M.Cz., Museum of Częstochowa, Poland; MJSN, Jurassica Museum, Porrentruy, Switzerland; MNKI, National Museum in Kielce, Poland; MZ, Polish Academy of Sciences Museum of the Earth, Warsaw, Poland; NHMUK, Natural History Museum, London, UK; NKMB, Naturkunde-Museum Bamberg, Germany; NMS, Naturmuseum Solothurn, Switzerland; SMNS, Staatliches Museum für Naturkunde, Stuttgart, Germany; ZPAL, Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland.
Other abbreviations.—PCoA, principal coordinates analysis.
Material and methods
Specimens MZ VIII Vr-71 (complete series), MZ VIII Vr-72, MZ VIII Vr-73 (“ZPAL V-KRZ/32”), and ZPAL V. 69/1 (“ZPAL V-KRZ/33”) were studied personally by the authors. Comparisons with Jurassic and Cretaceous turtles were based on personal observation of specimens in the collections of NHMUK, NMS, and SMNS by TS.
3D imaging of MZ VIII Vr-71 was performed using the Shining 3D EinScan Pro 2X 3D scanner fixed on a tripod with EinScan Pro 2X Color Pack (texture scan), Ein-Turntable (alignment based on features), and EXScan Pro 3.2.0.2 software. The meshing was done using the Watertight Model and High Detail pre-sets. The snapshots of MZ VIII Vr-71 were taken after exporting the 3D models into MeshLab 2016.12 (orthographic view and Radiance Scaling [Lambertian] shader enabled). See SOM 1 (Supplementary Online Material at http://app.pan.pl/SOM/app66-Madzia_etal_SOM.pdf).
In order to explore the morphospace occupation of MZ VIII Vr-72 among Jurassic plesiosaurs and thalattosuchians, we performed a principal coordinates analysis (PCoA). We used a modified version of the dataset first published by Foffa et al. (2018c) that was constructed to focus on the dental traits of Middle and Late Jurassic plesiosaur, thalattosuchian, and ichthyosaur taxa. Because the dentition of MZ VIII Vr-72 differed markedly from that of ophthalmosaurid ichthyosaurs (e.g., Fischer et al. 2016), the character list was modified to exclude features present or variable only in ichthyosaurs (discrete characters 16 [D16] and 17 [D17]) and the taxon sampling was reduced to include pliosaurids, plesiosauroids, metriorhynchids, and teleosauroids only. In addition, restricting the dataset only to plesiosaurs and thalattosuchions allowed for a clearer determination of the position of MZ VIII Vr-72 within their tooth morphospace. The taxon scores and extended results of the PCoA are provided in SOM 2 (modified dataset spreadsheet) and SOM 3, respectively. The characters are numbered as in Foffa et al. (2018c: supplementary information). The PCoA was performed in PAST 4.01 (Hammer et al. 2001). As in Foffa et al. (2018c), continuous characters were z-transformed, and the Gower similarity index was used as it is well suited for datasets which include both continuous and discrete variables (Gower 1971).
Note that the original version of the dataset, as published in Foffa et al. (2018c), did not include the raw measurements used for continuous characters C1–C5. Instead, the z-transformed values of these measurements were published. We were provided with the raw measurements by Davide Foffa (National Museums Scotland, Edinburgh, UK) upon request. One of us (DM) has subsequently noticed that some of the z-transformed values of Foffa et al. (2018c) were incorrect and misplaced in the wrong columns (original character C3 should have been in C2) and D. Foffa confirmed these errors (personal communication 2020). It should be noted that re-analyses of the dataset of Foffa et al. (2018c), which take into account corrections to the discovered errors, did not alter the results (D. Foffa, personal communication 2020). Nevertheless, it is encouraged that future studies should use the raw data provided in this paper, which include the original raw measurements collected by D. Foffa for Foffa et al. (2018c) and the recalculated z-transformed values (SOM 2: original dataset spreadsheet).
Systematic palaeontology
Testudinata Klein, 1760
Thalassochelydia Anquetin, Püntener, and Joyce, 2017
“Plesiochelyidae” Baur, 1888
Genus Craspedochelys Rütimeyer, 1873
Type species: Craspedochelys picteti Rütimeyer, 1873, Solothurn, Switzerland, upper Kimmeridgian, Upper Jurassic.
Craspedochelys? sp.
Figs. 2–6.
Material.—MZ VIII Vr-71, shell fragments from the canary-yellow Nerinea limestone, upper Kimmeridgian of Krzyżanowice, Poland (Dąbrowska 1957; Borsuk-Białynicka and Młynarski 1968).
The specimen series collectively labelled MZ VIII Vr-71 was collected in 1962 from three pits located 15–20 m apart and consists of hundreds of shell fragments of varying sizes belonging to three or four individuals of different ontogenetic stages and supposedly representing a single species (Borsuk-Białynicka and Młynarski 1968). The elements from each pit present little to no overlap, therefore their attribution to a single taxon is uncertain. They nonetheless originate from a single stratigraphic layer from the same locality and exhibit congruent characteristics of the shell surface, morphology of the sulci, shell thickness and degree of ossification. Due to the absence of significant differences, these elements are herein considered as representing a single taxonomic entity. Borsuk-Białynicka and Młynarski (1968) stated that the series included a central fragment of carapace, several carapace and plastron fragments likely belonging to another individual, two plastron fragments from a much larger individual, and numerous smaller pieces. The original description of this material (Borsuk-Białynicka and Młynarski 1968) was very brief and no actual specimens were figured. Two of the numerous fragments were recently redescribed and figured (Tyborowski and Błażejowski 2019a: fig. 4B, C, 2019b: fig. 4); however, the descriptions contain numerous errors, insufficient and selective comparisons and do not discuss nor reflect the relevant literature. Therefore, a thorough redescription was needed.
Description.—A central fragment of carapace (Fig. 2): This is the only piece figured as an interpretive drawing in Borsuk-Białynicka and Młynarski (1968: fig. 2), although in a very stylized form, and as photographs in Tyborowski and Błażejowski (2019a: fig. 4C, D; 2019b: fig. 4C). The fragment is pieced together from numerous small fragments and a substantial part is reconstructed with plaster. Currently it is split into two parts, separated along the neurocostal connection.
Borsuk-Białynicka and Młynarski (1968) described this fragment as comprising the third and fourth neural and the costals: third to fifth on the right and third on the left. The layout of the sulci, however, indicates that the costals preserved on the right side of the fragment are either the second to fourth or fourth to sixth (the suture between the middle and posteriormost costal is fused and visible only viscerally), and thus the neurals represent either the second and the third or the fourth and the fifth (Fig. 6A: 1). As mentioned by Tyborowski and Błażejowski (2019a), small fragments of another, preceding costal are preserved at the anterior right edge of the piece. The left preserved costal is attached incorrectly and more likely corresponds to the second preserved costal on the right side (the fourth according to Borsuk-Białynicka and Młynarski 1968, but actually likely either the third or the fifth). This is clearly evidenced by a sulcus on its external surface. Despite the damage, the sulcus interpreted by Borsuk-Białynicka and Młynarski (1968) as an interpleural sulcus is clearly sinuous and approaching the costoneural suture. These aspects make it markedly different from the unambiguous interpleural sulcus preserved on the contralateral side of the fragment, which is straighter and well-separated from the neurals, but congruent with the morphology of intervertebral sulci in the same piece. A supposed intervertebral sulcus was also marked in red close to the anterior end of the first preserved neural. Although plausible, this identification is ambiguous—such a position of the sulcus would require a departure from the typical turtle layout in which the intervertebral sulci cross only the odd-numbered neurals, and it cannot be ruled out that the observed groove might have originated as a result of damage. Despite the intervertebral sulci being sinuous, the areas of the vertebral scutes lack regular and well-defined radial striations, which are typical for many Jurassic turtles (Meyer 1860; Wagner 1861; Maack 1869; Rütimeyer 1873; Bräm 1965; Joyce 2003; Jansen and Klein 2014; Sullivan and Joyce 2017), but particularly for Eurysternum wagleri Meyer, 1839a and Pelobatochelys blakii Seeley, 1875 (see Meyer 1839a; Seeley 1875; Anquetin and Joyce 2014; Pérez-García 2015c; Anquetin et al. 2017). Only several, short and irregularly placed groves are present along the anterior edge of the vertebral scute area (Fig. 2A1). The intervertebral sulcus is predominantly horizontal and the vertebral lacked a sagittal tip, unlike the vertebral scutes in, e.g., Achelonia formosa Meyer, 1860, Eurysternum wagleri, Idiochelys fitzingeri Meyer, 1839b, or Palaeomedusa testa Meyer, 1860 (e.g., Meyer 1860; Maack 1869; Rütimeyer 1873; Joyce 2003; see Anquetin and Joyce 2014; Joyce and Mäuser 2020). The scute areas are completely flat, unlike in Platychelys oberndorferi Wagner, 1853 (e.g., Meyer 1860; Wagner 1861; Bräm 1965; Lapparent de Broin 2001; Sullivan and Joyce 2017). The vertebrals are moderately wide, about five times the width of the neurals. This proportion resembles Thalassemys hugii Rütimeyer, 1873, Plesiochelys etalloni (Pictet and Humbert, 1857) (with the exception of NMS 79 with exceptionally wide vertebrals) and Plesiochelys bigleri Püntener, Anquetin, and Billon-Bruyat, 2017a, as well as Craspedochelys spp., but is smaller than that in Achelonia formosa, Chelonides wittei Maack, 1869, Eurysternum wagleri, Idiochelys fitzingeri, Jurassichelon moseri (Bräm, 1965), Jurassichelon oleronensis Pérez-García, 2015b, Thalassemys bruntrutana Püntener, Anquetin, and Billon-Bruyat, 2015, Thalassemys marina Fraas, 1903, Solnhofia parsonsi Gaffney, 1975b, Solnhofia brachyrhyncha Anquetin and Püntener, 2020, and likely Pelobatochelys blakii, and larger than in Tropidemys langii Rütimeyer, 1873 and Tropidemys seebachi Portis, 1878 (see Meyer 1839b, 1860; Maack 1869; Rütimeyer 1873; Seeley 1875; Portis 1878; Fraas 1903; Andrews 1921; Rieppel 1980; Joyce 2000; Anquetin et al. 2014b, 2017; Karl et al. 2012a; Püntener et al. 2014, 2015, 2017b; Anquetin and Joyce 2014; Pérez-García 2015a, c; Anquetin and Chapman 2016; Raselli and Anquetin 2019; Joyce and Mäuser 2020). MZ VIII Vr-71 differs from Tropidemys seebachi in the lack of infravertebral scutes (Joyce and Mäuser 2020).

Fig. 2. The thalassochelydian turtle Craspedochelys? sp., MZ VIII Vr-71, from the Kimmeridgian of Krzyżanowice. Central part of the carapace (Fig. 6A: 1), in external (A1, A2) and visceral (A3, A4) views. A1, A3, photographs; A2, A4, explanatory drawings.
All the costals are incomplete distally and the rib necks are broken off at the level of the entrance into the costal plates. The neurals are coffin-shaped in outline, longer than wide, and wider anteriorly than posteriorly, unlike in platychelyids, and differ from the elongated but more rectangular and nearly parallel-sided neurals of Neusticemys neuquina (Fernández and de la Fuente, 1988) (see Meyer 1860; Wagner 1861; Bräm 1965; Fernández and de la Fuente 1993; Gasparini et al. 1997; Lapparent de Broin 2001). Their anterolateral edges are much shorter than the posterolateral edges, measuring (as preserved) 12 mm and 34 mm for the first and 13 mm and 39 mm for the second preserved neural (contra 10 mm and 15 mm given by Tyborowski and Błażejowski 2019a), respectively. This, together with the anteroposterior elongation of the neurals and lack of the sagittal keel, distinguishes MZ VIII Vr-71 from Tropidemys spp. (Rütimeyer 1873; Portis 1878; Karl et al. 2012a; Anquetin et al. 2014b, 2017; Püntener et al. 2014; Pérez-García 2015c; Anquetin and Chapman 2016; Joyce and Mäuser 2020). The lack of any keeling also distinguishes it from Pelobatochelys blakii (see Seeley 1875; Pérez-García 2015c; Anquetin et al. 2017). The first preserved neural is missing its anterior right part, has its corners less pronounced than the second preserved neural, and shows a convex rather than concave anterior edge, resulting in an overall rounder outline. However, this morphology seems to be taphonomic. The interneural suture is posteriorly convex. The vertebrae are not preserved, the neural processes are broken just below the neural plates. Unlike Idiochelys fitzingeri, MZ VIII Vr-71 does not show any signs of neural reduction (Meyer 1839b, 1860; Rütimeyer 1873; Lortet 1892; Anquetin et al. 2017). Unlike in Thalassemys spp., there are no striations perpendicular to the sutures (Rütimeyer 1873; Anquetin et al. 2014b, 2017; Pérez-García 2015a; Püntener et al. 2015). The illustration presented for that specimen in the original paper (Borsuk-Białynicka and Młynarski 1968: fig. 2) does not accurately reflect its shape and completeness, showing some elements that are not currently present and, judging from the original description, have likely never been. In the original paper, the illustration is described as “reproducing, according to the principles of symmetry, the lacking parts of plates and damaged edges, as well as […] aligning to each other the disposition of particular parts” (Borsuk-Białynicka and Młynarski 1968: 219). Still, the specimen is recognizable and, therefore, it is puzzling why Tyborowski and Błażejowski (2019a: 745) describe it as a “new specimen”. Curiously, the dimensions of the specimen given in the text and shown in the figures in Tyborowski and Błażejowski (2019a, b) are clearly reduced. For example, the costal preserved on the left side of the fragment measures about 72 × 43 mm, not 51 × 29 mm, and the first preserved costal on the right side of the fragment (the most complete, but still broken distally) measures about 144 × 39 mm (length to width ratio equals 3.7), not 102 × 35 mm.
Two fragments of plastron and carapace with peripherals, likely belonging to a single individual (Figs. 3, 4A–C): One of the pieces (Fig. 3A) was interpreted by Borsuk-Białynicka and Młynarski (1968) as consisting of the first neural, fragments of the first two costals and part of the hyoplastron with a pectoroabdominal sulcus. The identification of the carapace elements seems correct (Fig. 6A: 2), as suggested by their shapes, interconnections, and by what appears to be a very gentle transverse sulcus crossing the supposed first neural and first costal—an appropriate location for the sulcus between the first two vertebral scutes. Posteriorly to the first neural, part of the second neural seems to be present as well. Neither of the elements are preserved completely. The first neural has its anterior part broken but seems to have a rounded rectangular shape in outline. The second neural has only its left part preserved but it appears to be coffin-shaped, with the anterior part wider than the posterior part and very short craniolateral edges. The interneural suture is posteriorly convex. All of these features are common for the first two neurals of thalassochelydians (e.g., Anquetin et al. 2014b). Both neurals lack a keel. The preserved proximal parts of the first and the second left costal are broken into irregular fragments and provide little morphological data. The fragment of the hyoplastron is trapezoidal and bears a single, nearly straight transverse sulcus. If the spatial correlation with the associated carapace fragments is any indication, this would indicate that the sulcus represents the humeropectoral, rather than the pectoroabdominal sulcus. There is, however, no clear morphological indication that would favour one interpretation over the other, because the lateral edge of the element is damaged.

Fig. 3. The thalassochelydian turtle Craspedochelys? sp., MZ VIII Vr-71, from the Kimmeridgian of Krzyżanowice, parts of the carapace and plastron likely belonging to a single individual. A. Anterior part of the carapace and plastron (Fig. 6A: 2) in dorsal (A1) and ventral (A2) views. B. Posterior part of the carapace and plastron (Fig. 6A: 3) in dorsal (B1) and ventral (B2) views. The fragments likely belong to the same individual as the material shown in Fig. 4A–C.
The second fragment (Fig. 3B), likely belonging to the same individual, has been described as strongly damaged fragments probably pertaining to the seventh costal and neural together with a fragment of hypoplastron with the abdominofemoral sulcus (Borsuk-Białynicka and Młynarski 1968). This interpretation seems to be incorrect—there appears to be no neural (although it cannot be ruled out that it was initially present and became separated over the years) and the external surface apparently bears two oblique sulci, one of which is faint and predominantly aligned anteroposteriorly, and the other sinuous and nearly transverse. This layout refutes the identification of the element as the seventh costal, but supports its recognition as the eight costal, which would agree with the relatively small size of the piece (Fig. 6A: 3). Just like in the previous piece, the costal is incomplete and broken into several fragments. The hypoplastron is subtriangular, has damaged edges, and bears an oblique sulcus, which may be the femoroanal, rather than the abdominofemoral sulcus, as suggested by its straight, rather than curved trajectory and proximity of the (supposedly) eighth costal. Nonetheless, the interpretation of this fragment is ambiguous.
According to Borsuk-Białynicka and Młynarski (1968), the gathered material also includes a fragment of the carapacial rim bearing the sulci between the anteriormost four marginals (Fig. 4A) and two peripherals interpreted as the third right (Fig. 4B) and the fifth or sixth peripheral from an indeterminate side (Fig. 4C), all likely belonging to the same individual as the associated carapace and plastron fragments. The identification of the former fragment (Fig. 4A) seems to be accurate (Fig. 6A: 4); this is supported by the presence of an acute anterior corner of the supposed first pleural and a gently downturned free edge. The element comes from the left side of the body. The areas of the first and the fourth marginal are incomplete anteriorly and posteriorly, respectively. In dorsal view, the first marginal was apparently narrower than the rest and had a straight, rather than protruding anterolateral edge. This differs from the anteriorly bulging first marginal area of Craspedochelys passmorei (Andrews, 1921), Jurassichelon moseri, Jurassichelon oleronensis, and Plesiochelys etalloni (Fig. 6B, C), but resembles most specimens of Craspedochelys jaccardi (Pictet, 1860) (Fig. 6D) (with the possible exception of the holotype, which is slightly divergent in morphology from the specimens from Solothurn, see Pictet 1860; Anquetin et al. 2014b), Craspedochelys picteti (Rütimeyer, 1873) (Fig. 6E), Plesiochelys bigleri (with the exception of MJSN BSY007-257, see Püntener et al. 2017b), and Thalassemys hugii (see Rütimeyer 1873; Portis 1878; Andrews 1921; Bräm 1965; Rieppel 1980; Anquetin et al. 2014a, b, 2017; Pérez-García 2015a; Püntener et al. 2017b; Raselli and Anquetin 2019). The marginals are narrower than in pleurosternids such as Dorsetochelys typocardium (Seeley, 1869), Pleurosternon bullocki (Owen, 1842), Riodevemys inumbragigas Pérez-García, Royo-Torres, and Cobos, 2015, or Selenemys lusitanica Pérez-García and Ortega, 2011 (see Owen 1853; Milner 2004; Pérez-García and Ortega 2011; Pérez-García 2014; Pérez-García et al. 2015). Both the first and the second marginal formed an extensive contact with the first vertebral scute, unlike in Achelonia formosa, Palaeomedusa testa, Parachelys eichstaettensis Meyer, 1864, Plesiochelys spp., and (with the exception of Tropidemys seebachi NKMB Watt09/162) most specimens of Tropidemys spp. (Fig. 6B, C; see also Meyer 1860; Rütimeyer 1873; Bräm 1965; Lapparent de Broin et al. 1996; Joyce 2003; Karl et al. 2012a; Anquetin et al. 2014a, b; Püntener et al. 2014, 2017a, b; Anquetin et al. 2017; Raselli and Anquetin 2019; Joyce and Mäuser 2020). In Craspedochelys spp. the contact between the second marginal and the first vertebral was apparently variable, but it was present at least in Craspedochelys picteti NMS 9149 (former NMS 608; Fig. 6D, E; see also Bräm 1965; Anquetin et al. 2014b). Borsuk-Białynicka and Młynarski (1968) mentioned that the areas of the first three pleurals were preserved in MZ VIII Vr-71, but this was incorrect; in addition to most of the lateral margin of the first pleural, only the craniolateral corner of the first vertebral is preserved. If at the time of the original description there was an additional part preserved posterior to the existing series that bore the sulcus between the first and the second pleural, it has been detached since. The sulci are indistinguishable ventrally in a manner similar to, e.g., Craspedochelys jaccardi (see Bräm 1965; Anquetin et al. 2014b, 2017), either as a result of very limited ventral exposition of the anterior marginals (body wall being unusually close to the edge of the carapace) or as a taphonomic effect. In marginal aspect, the fragment is gently sinuous, reaching the dorsal peak at the level of the second marginal scute, and the third and fourth marginals increase in dorsoventral diameter due do the downturn of their free edges becoming more pronounced. As mentioned by Borsuk-Białynicka and Młynarski (1968), the sutures are not visible in that specimen and there are no costoperipheral fontanelles. The latter may differentiate MZ VIII Vr-71 from, e.g., Achelonia formosa, Chelonides wittei, Eurysternum wagleri, Hydropelta meyeri (Thiollière, 1850), Idiochelys fitzingeri, Jurassichelon spp., Neusticemys neuquina, Pelobatochelys blakii, probably Solnhofia spp., Thalassemys spp., and Tropidemys seebachi (Meyer 1839b, 1860; Maack 1869; Seeley 1869, 1875; Rütimeyer 1873; Portis 1878; Lortet 1892; Fraas 1903; Rieppel 1980; Fernández and de la Fuente 1993; Gasparini et al. 1997; Joyce 2000; Anquetin and Joyce 2014; Anquetin et al. 2014b, 2017; Pérez-García 2015a; Püntener et al. 2015; Anquetin and Püntener 2020; Joyce and Mäuser 2020), although the preserved section comes from the anterior part of the carapace, so the presence of more posterior fontanelles cannot be excluded. The isolated bone described as the third right peripheral (Figs. 4B, 6A: 5) indeed resembles its counterpart from the left side both in the layout of the sulci and in the gradual increase of the downturn of its edge. The identification of the piece described as the fifth or sixth peripheral (Fig. 4C) is, however, dubious: the downturn of the edge and the dorsoventral diameter do not match the fourth peripheral of the preserved series, and the fragment does not show any intermarginal sulcus nor trace of the bridge. Both isolated peripherals are broken off rather than separated along the suture lines. Neither of the fragments shows whether the bridge was ligamentous or osseous.

Fig. 4. The thalassochelydian turtle Craspedochelys? sp., MZ VIII Vr-71, from the Kimmeridgian of Krzyżanowice. A. Anterior left part of the carapacial rim (Fig. 6A: 4) in external (A1), visceral (A2), and peripheral (A3) views. B. Fragment of the anterior right part of the carapacial rim (Fig. 6A: 5) in external (B1), visceral (B2), and peripheral (B3) views. C. Unidentified part of the carapacial rim in external (C1), visceral (C2), and peripheral (C3) views. D. Fragment of the posterior part of the carapace (Fig. 6A: 6). A–C likely belong to the same individual as the material shown in Fig. 3. The numbers indicate marginal scutes.
There is at least one other fragment with well-preserved sutures and a broken ridge on the visceral surface (Figs. 4D, 6A: 6), likely belonging to a different individual and representing a part of the intermediate element present in thalassochelydians (which is thought to at least partially articulate with the vertebral column; Anquetin et al. 2014b), the posterior left part of the preceding neural, and the proximal portion of the last left costal. The incompleteness of that fragment, however, makes this identification tentative. The preserved part of the neural suggests it was coffin-shaped, as the more anterior neurals. Its dorsal surface bears several oblique grooves, but it is not certain whether they were present in life or are of taphonomic origin (and, in the former case, whether they are pathological or not). The intermediate element receives the posterior edge of the costal via a posteriorly convex suture and expands posterolaterally. Neither of the elements is crossed in the preserved part by a sulcus, but the last intervertebral sulcus was located either more anteriorly, in the anterior part of the last neural, or more posteriorly, in the posterior part or behind the intermediate element (as in, e.g., Craspedochelys jaccardi NMS 102a; see Anquetin et al. 2014b: fig. 4M, N). All of the fragments are in agreement with the central part of the carapace (Fig. 2) in suture and sulci morphology, whenever the sutures and/or sulci are preserved.
Six large fragments of costals and indeterminate parts of the carapace (Fig. 5A–F): The costals were interpreted by Borsuk-Białynicka and Młynarski (1968) as the first, the third or fourth, and the seventh or eighth, but no justification for these identifications was provided. For certain, MZ VIII Vr-71 includes two fragments of a second, fourth, or sixth costal (based on the presence of the pleurovertebral and interpleural sulci in one [Fig. 5A] and no sulci in the proximal part of the second [Fig. 5F]), a fragment of a probable third or fifth costal (based on the presence of a sinuous, likely intervertebral sulcus [Fig. 5B]), and at least seven large (over 30 mm) parts, tentatively identified as costals but lacking sulci on the external surfaces (Fig. 5D–E). The latter may represent areas of either vertebral or pleural scutes, but their state of preservation makes it impossible to establish whether the preserved parts are proximal or distal, and therefore their sequential identity is unknown. There is no isolated element readily identifiable as the first costal (contra Borsuk-Białynicka and Młynarski 1968; Tyborowski and Błażejowski 2019a, b). Since Borsuk-Białynicka and Młynarski (1968) did not figure nor describe in detail the fragment mentioned as an isolated first costal, it is impossible to verify whether they singled out the same piece that was described and figured by Tyborowski and Błażejowski (2019a: fig. 4B, 2019b: fig. 4A, B). In any case, although the general outline of the latter element superficially resembles the shape of the first costal in “plesiochelyids” (e.g., Anquetin et al. 2014b), such an identification is incorrect and refuted by the lack of any sulci on its external surface and rib-related structures viscerally, as well as by the presence of a single, natural, non-sutural edge (Fig. 5G; the fragment was broken into several pieces during DM, TS, and ASWs personal examination for this study, but the figures show its appearance before it became damaged). This edge is unlike the borders of the intercostal fontanelles, as it is nearly straight, spans along the entire preserved length of the element, and is not associated with a longitudinally striated free distal part of a rib (compare with, e.g., Meyer 1839b, 1860; Maack 1869; Rütimeyer 1873; Seeley 1875; Portis 1878; Lortet 1892; Fernández and de la Fuente 1988; Joyce 2000; Karl et al. 2012b; Anquetin and Joyce 2014; Anquetin and Püntener 2020; Joyce and Mäuser 2020; Püntener et al. 2020). Furthermore, the edge itself is recessed, i.e., it is deep to the external surface and even slightly deeper than the visceral surface, forming a low lip viscerally. The area between the edge of the element and its main body is set at a conspicuous, obtuse angle to the external surface and the outline of the latter is gently concave along their border between them, yet both surfaces show similar microsculpture suggesting that they were overlain by epidermal scutes. Given all of these characteristics, the interpretation of the element as a hyo- or hypoplastron fragment close to the inguinal or axillary notch is more likely. Just like for the central part of the carapace, the element is significantly larger than indicated by Tyborowski and Błażejowski (2019a, b), measuring about 75 × 40 mm. In all the isolated costals, the morphology of the sulci and sutures, whenever preserved, is congruent with the remaining fragments.

Fig. 5. The thalassochelydian turtle Craspedochelys? sp., MZ VIII Vr-71, from the Kimmeridgian of Krzyżanowice, shell fragments. A. Middle part of a second, fourth, or sixth costal in external view. B. Proximal part of a third or fifth costal in external view. C, D. Parts of unidentified costals in external view. E. Parts of two unidentified costals retaining a sutural contact in external view. F. Proximal part of a second, fourth, or sixth costal in external (F1) and visceral (F2) views. G. Probable part of the plastron in external (G1–G2), visceral (G3–G4), ?posterior (G5), and ?lateral (G6) views. Note that the position of the element within the shell is uncertain, so precise orientation is not possible. G2, G3, G5, and G6 are 3D models based on surface scans in the orthographic view with the Radiance Scaling shader enabled to optimize the lighting and present the raised ?lateral edge (asterisk). H. Part of a plastron of a large individual in external (H1) and visceral (H2) view and natural cross-section (H3). Note that the fragments likely belong to various individuals.
Two thick plastron fragments (Fig. 5H): The pieces are undiagnostic beyond identification as plastral elements. We agree with Borsuk-Białynicka and Młynarski (1968) in their conclusion that these belong to a large individual, as indicated by their thickness, and that their position within the shell is indeterminate.
Small, unidentifiable shell fragments: Hundreds of small, unidentifiable shell fragments are present as part of the MZ VIII Vr-71 series. Some of these bear sulci, but due to their small size and damage it is impossible to reliably identify their position within the shell.
Discussion.—Borsuk-Białynicka and Młynarski (1968) attributed all the gathered material (not part of it, contra Tyborowski and Błażejowski 2019b) to Tretosternon aff. punctatum, a taxon with a complicated taxonomic history (see Lapparent de Broin et al. 1996; Lapparent de Broin and Murelaga 1999; Milner 2004; Joyce 2017), which is now considered a nomen dubium (Milner 2004; Joyce 2017). Borsuk-Białynicka and Młynarski (1968: 219) mistakenly cited the geological age (apparently unaware that the British material was dated to the Early Cretaceous rathen than the Kimmeridgian, see Joyce 2017 and references therein), flatness of the carapace and “its typical sculpture” as “very characteristic of the genus”.
The sculpture, which for over a century was considered diagnostic for Tretosternon spp., was problematic in itself. Based on one of the two syntype specimens, which have never been figured and are now lost (Lydekker 1889; Delair 1958; Lapparent de Broin et al. 1996; Lapparent de Broin and Murelaga 1999; Milner 2004; Joyce 2017), Owen (1842: 165) stated that in his newly described Tretosternon punctatum “the external surface of […] the carapace is closely pitted with minute irregular impressions, smaller than a pin’s head”. However, he also referred the specimens figured earlier by Mantell (1827: pl. 6: 1, 3, 5) and (at least some of them) already described as “Trionyx bakewelli” Mantell, 1833 (currently “Helochelydra” bakewelli, see Joyce 2017) at that time, which clearly show a finely tuberculated surface, to T. punctatum. It is unknown whether this inconsistency was the result of a misinterpretation of Mantell’s (1827, 1833) figures by Owen (1842). This led to a confusion causing helochelydrid-like tubercles rather than pits to be cited as characteristic for the taxon (e.g., Lydekker 1889). The description of the pitted, lost syntype was more recently interpreted as being consistent with the morphology present in Pleurosternon bullocki, suggesting synonymy (Lapparent de Broin et al. 1996; Lapparent de Broin and Murelaga 1999; Milner 2004), but Owen (1842) never noted any particular similarity in the sculpture of the two, despite describing them side by side in the same paper. The surficial characteristics of the specimens figured by Mantell (1827, 1833) are typical for helochelydrid turtles, to which many of the specimens historically attributed to the genus “Tretosternon” are now referred (Lapparent de Broin et al. 1996; Lapparent de Broin and Murelaga 1999; Milner 2004; Joyce 2017). The shells/carapace of these turtles are commonly ornamented in little tubercles and granulations (Lapparent de Broin and Murelaga 1999; Milner 2004; Joyce et al. 2011; Scheyer et al. 2014; Joyce 2017; Herzog 2019), which in some species are prone to breakage, leaving minute pitting (Joyce et al. 2011; Scheyer et al. 2014; Joyce 2017; Herzog 2019). Arguably, these are more similar to the trionychid sculpturing cited by Owen (1842) than is the shell surface of Pleurosternon bullocki (NHMUK 21351, TS personal observation). It may be thus speculated that the “impressions” observed by Owen (1842) were in fact such damaged tubercles, but this cannot be proven until the missing specimen is rediscovered in the future. The tubercles of “Helochelydra” bakewelli are, admittedly, not easily broken off (Joyce et al. 2011; Joyce 2017), but their sculpture is otherwise extremely similar to that in some other helochelydrids in which the tubercles are prone to breakage (Joyce et al. 2011), so such a mistake seems plausible. The Pleurosternon bullocki hypothesis was favoured over the helochelydrid one by Lapparent de Broin et al. (1996) and Lapparent de Broin and Murelaga (1999) due to the presence of striations perpendicular to the sutures in the specimen described by Owen (1842) which, as the authors argued, are known only in pleurosternids (Owen 1853; Lapparent de Broin et al. 1996; Lapparent de Broin and Murelaga 1999; Milner 2004). One of the helochelydrid specimens presented by Joyce et al. (2011: fig. 2B) and several described and pictured by Herzog (2019), however, have tubercles coalescing near the sutures, producing perpendicular ridges/striations, and prone to breakage tubercles on the main body of the bones. These linear structures do span “for the extent of two or three lines” (slightly over four to six millimetres), potentially matching Owen’s (1842: 165) description better than the longer, over 15 mm striations in Pleurosternon bullocki (TS personal observation). In any case, the sculpture of the carapace of MZ VIII Vr-71, which is inconsistent with that of “Tretosternon” was noted multiple times (Lapparent de Broin and Murelaga 1999; Lapparent de Broin 2001; Joyce et al. 2011; Anquetin et al. 2017; Joyce 2017). As a result, modern authors considered the specimen either an indeterminate turtle (Lapparent de Broin and Murelaga 1999; Lapparent de Broin 2001; Joyce et al. 2011) or an indeterminate “plesiochelyid” (Anquetin et al. 2017; Joyce 2017; Tyborowski and Błażejowski 2019a, b). Indeed, the external surface of MZ VIII Vr-71 is mostly smooth, even despite it being covered by numerous larger, irregular pits of varying depths (Figs. 2A1, 3, 4A1, B1, C1, 5A–E, F1, G1). Similar pits occur commonly in European Jurassic aquatic turtles, regardless of their taxonomic affinities, and their distribution and morphology is asymmetric and inconsistent between individuals (e.g., Rütimeyer 1873; Bräm 1965; Lapparent de Broin et al. 1996; Meyer 2011; Anquetin et al. 2014b; Püntener et al. 2014; Anquetin and Püntener 2020; TS personal observation). Thus the pits likely have extrinsic origins (pathologic, taphonomic, a result of weathering or preparatory work) and lack taxonomic value. Very few fragments exhibit finer, more concentrated pitting on the surface (Fig. 5A, D, F1), but its extent is very limited, its origin appears to be taphonomic and the dorsal surface of the fragments is in some cases coarse (Fig. 5F1). This matter will be tackled in a separate paper (Tomasz Szczygielski, Justyna Słowiak, Irena Raselli, and Dawid Surmik, unpublished material).
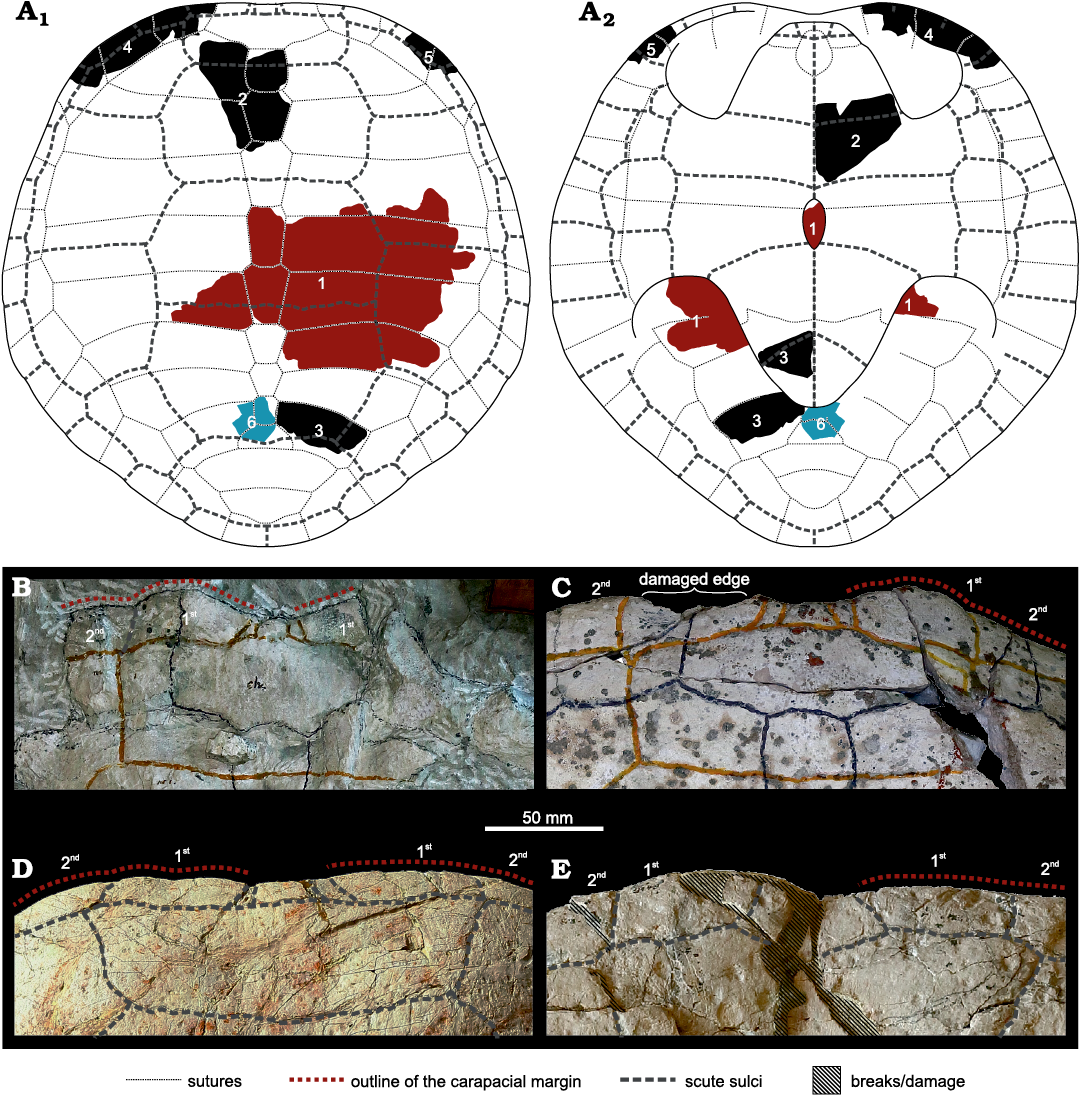
Fig. 6. A. Approximate position of the identifiable fragments of the thalassochelydian turtle Craspedochelys? sp., MZ VIII Vr-71, within the shell, in dorsal (A1) and ventral (A2) views (schematic reconstruction of Craspedochelys spp. based on Anquetin et al. 2014, 2017, and TS personal observations, modified to fit the observed morphologies). Numbers represent the elements illustrated in the text: 1, Fig. 2; 2, Fig. 3A; 3, Fig. 3B; 4, Fig. 4A; 5, Fig. 4B; 6, Fig. 4D. Fragments likely belonging to the same individual are indicated by the same colour. B–E. Anterior edges of the carapace of Plesiochelys etalloni (Pictet and Humbert, 1857), NMS 8514/NMS 118 (B) and NMS 8727/NMS 116 (C), Craspedochelys jaccardi (Pictet, 1860), NMS 101 (D), and Craspedochelys picteti (Rütimeyer, 1873), NMS 9149/NMS 608 (E); all from the Kimmeridgian of Solothurn, Switzerland. The numbers indicate marginal scutes. Note the anteriorly protruding first marginal of Plesiochelys etalloni (B, C) and straight anterior edge in Craspedochelys spp. (D, E).
The smoothness of the shell and the lack of consistent, small-scale ornamentation distinguishes MZ VIII Vr-71 from helochelydrids and Jurassic and Berriasian European pleurosternids, Dorsetochelys typocardium, Pleurosternon bullocki, “Pleurosternon” portlandicum Lydekker, 1889, Riodevemys inumbragigas, and Selenemys lusitanica (see Owen 1842, 1853; Lapparent de Broin and Murelaga 1999; Milner 2004; Joyce et al. 2011; Pérez-García and Ortega 2011; Scheyer et al. 2014; Pérez-García 2014; Pérez-García et al. 2015; Herzog 2019; Joyce and Anquetin 2019). The shell is relatively thick, differentiating it from the “eurysternids” (e.g., Anquetin et al. 2017; Püntener et al. 2020). There is no trace of costoperipheral fontanelles, further refuting attribution to numerous “eurysternid” taxa, Achelonia formosa, Thalassemys spp., Tropidemys seebachi, Jurassichelon spp., Neusticemys neuquina, and Pelobatochelys blakii (see above). The shape of the neurals is more regular than, e.g., in platychelyids (Meyer 1860; Wagner 1861; Bräm 1965; Lapparent de Broin 2001; Sullivan and Joyce 2017). MZ VIII Vr-71 is therefore most similar to “plesiochelyids”, namely Craspedochelys spp. and Plesiochelys spp. The only identifiable characters pointing towards a Craspedochelys spp. affinity are the anteroposterior dimension of the first marginal, which in MZ VIII Vr-71 was smaller compared to the second marginal, unlike the more bulbous and protruding first marginal of Plesiochelys etalloni (Fig. 6B, C), a well-defined contact between the second marginal and the first vertebral (absent in Plesiochelys spp., but present at least in Craspedochelys picteti NMS 9149, Fig. 6B–D) and the proportions of the costals. The shape of the first marginal may be of little taxonomic value, since some variation of that nature is present in thalassochelydians, in general, and “plesiochelyids”, in particular (e.g., Anquetin et al. 2014b; Püntener et al. 2017b). The longest, most complete (although still broken distally) costal (either the second or the fourth), however, attains the length/width ratio of 3.7. This already exceeds the ratios given for the fourth costal by Anquetin et al. (2014b) for Plesiochelys etalloni (3.1–3.6), encompassing also Plesiochelys bigleri (see Püntener et al. 2017a; Raselli and Anquetin 2019), and it seems realistic that, when complete, it could have reached the range within Craspedochelys spp. (4.3–4.8). Unfortunately, the midsection of the plastron and the bridge are not preserved, so there are no data on whether the bridge was ligamentous or osseous and if any plastral fontanelles were present. The material does not allow any comparisons with Owadowia borsukbialynickae, Plesiochelys planiceps (Owen, 1842), Portlandemys mcdowelli Gaffney, 1975a, and Portlandemys gracilis Anquetin, Püntener, and Billon-Bruyat, 2015. Due to its incompleteness, in accordance with tentative inferences of some previous authors (Anquetin et al. 2017; Joyce 2017; Tyborowski and Błażejowski 2019a, b), we identify MZ VIII Vr-71 as an indeterminate “plesiochelyid” resembling Craspedochelys spp.
Archosauria Cope, 1869
Pseudosuchia Zittel, 1887
Thalattosuchia Fraas, 1901
Metriorhynchidae Fitzinger, 1843
Metriorhynchidae indet.
Figs. 7, 8.
Material.—MZ VIII Vr-72, partial jaws and dentition from an unknown locality and age (see Discussion below); reported by Tyborowski and Błażejowski (2019a) as originating from the light yellow marl, upper Kimmeridgian of Krzyżanowice, Poland (Dąbrowska 1957; Borsuk-Białynicka and Młynarski 1968).
Tyborowski and Błażejowski (2019a, b) reported a limestone block allegedly preserving a left premaxilla and right dentary of Pliosaurus (Tyborowski and Błażejowski 2019a), collectively catalogued as MZ VIII Vr-72 (Fig. 7). As already noted, the same catalogue number (MZ VIII Vr-72) has been previously assigned to incomplete jaws originating from the Oxfordian of Załęcze Wielkie that were described and illustrated by Maryańska (1972: pl. 1: 2) and referred to Peloneustes sp. More recently the specimen was proposed to represent a possible teleosaurid thalattosuchian (Ketchum and Benson 2011: appendix S1, p. 11).
Description.—The interpretation of MZ VIII Vr-72 by Tyborowski and Błażejowski (2019a) is ambiguous, both morphologically and taxonomically, and the description contains numerous errors and misinterpretations. The “left premaxilla” (inconsistently referred to as the “right premaxilla” in other parts of their manuscript; e.g., see Tyborowski and Błażejowski 2019a: 745) was described as bearing 10 alveoli, with the first alveolus being supposedly reduced. The presumed posterior margin of the “premaxilla” (the premaxilla-maxilla contact) was considered to form a “zig-zag” suture, and the teeth preserved in the alveoli were described as trihedral. These purportedly observed characters (specifically the reduced first alveolus, “zig-zag” suture at the premaxilla-maxilla contact, and presence of trihedral teeth) then served as the basis to assign the specimen to the globally distributed Late Jurassic pliosaurid taxon Pliosaurus.
In their description of the “right dentary”, in turn, Tyborowski and Błażejowski (2019a: 746) stated that “[a]ll of the dentary teeth are preserved” and that “[t]he crowns of all teeth are robust, conical, gently recurved lingually and display gently longitudinal ridges throughout”, with the “longitudinal ridges” being “regularly and evenly spaced throughout”. Tyborowski and Błażejowski (2019a: 746) further noted that “[t]he preserved dentary teeth share trihedral morphology” which, again, supposedly supports the identification of the specimen as Pliosaurus. However, an imprint of a tooth crown, preserved on the same block as the “premaxilla” and the “dentary” was said to show a subtrihedral morphology because it was “conical, curved and its cross-section has flattened labial and lingual surfaces” (Tyborowski and Błażejowski 2019a: 746); its “distal surface bears prominent, apicobasally-oriented carinae” and its “labial surface […] bears many apicobasal enamel ridges”. Both the “premaxillary” and “dentary” dentitions were further characterized as being heterodont and anisodont though the two terms just appear to be used interchangeably (Tyborowski and Błażejowski 2019a: 745, 746).
Even though the “left premaxilla” is badly preserved, the observed tooth count makes it highly unlikely that it is indeed a premaxilla. The element likely represents either a right maxilla or a left dentary (see below). Furthermore, the dental morphology and the distribution of the outer enamel structural elements clearly show that the specimen cannot be referred to Pliosaurus. In all species currently assigned to that taxon, the premaxillae have typically five or six trihedral or subtrihedral teeth (Knutsen 2012; Benson et al. 2013; Zverkov et al. 2018). Although Tyborowski and Błażejowski (2019a) noted that the teeth of MZ VIII Vr-72 were all trihedral (except for a single tooth, illustrated in their figure 6, that was characterized as subtrihedral), this is incorrect. In trihedral teeth, their cross section is nearly triangular. The labial surface of tooth crowns is almost flat and the flat segment is bordered by prominent carinae (see e.g., Zverkov et al. 2018: fig. 2A). Subtrihedral teeth, in contrast, have their cross sections somewhat transitional between trihedral and subcircular, their labial surfaces mostly lack ridges, and are only slightly flattened. The term “subtrihedral” was introduced by Benson et al. (2013) to describe the teeth of Pliosaurus kevani Benson, Evans, Smith, Sassoon, Moore-Fay, Ketchum, and Forrest, 2013, though it has been used for other taxa as well (for a detailed overview, see the supplementary information in Zverkov et al. 2018: table S1).
Our personal examination of MZ VIII Vr-72 reveals that the cross sections of all the “premaxillary” and “dentary” teeth are actually subcircular, or very indistinctly labiolingually compressed at most. None of the “premaxillary” nor “dentary” crowns show “gently longitudinal ridges” similar to those of plesiosaurs. Two carinae are present, one placed mesially and one distally. In addition, only subtle structural elements, unlike the well-developed apicobasal ridges in pliosaurid teeth, can be observed. The basal half of the crowns is ornamented with short, apicobasally oriented elements resembling ridglets (sensu Zverkov et al. 2018; see Fig. 8). The ornamentation becomes finer apically and forms a vermicular pattern.
The apical and basal structural elements are separated by a short segment of smoother enamel (Fig. 8A1 and A2), although this is not a rule as in some teeth the ridglets and the vermicular pattern meet at mid-section (Fig. 8A3).
Other discussed features are problematic or misinterpreted as well. There is no evidence indicating that the first “premaxillary” alveolus was substantially reduced. The anterior part of the “premaxilla” is incomplete and the first preserved tooth (possibly originating from that region) is markedly displaced (see Fig. 7A1). Nothing suggests that either the “premaxillary” or “dentary” dentition is heterodont (variable with respect to shape) or clearly anisodont (variable with respect to size), though minor differences in size can be observed. Furthermore, the “dentary” certainly does not preserve the complete set of teeth (as stated; see above). It is also apparent that the posterior portion of the “premaxilla” does not indicate the presence of the “zig-zag” suture as observable in Pliosaurus (see e.g., Benson et al. 2013: 3A). The element is irregularly broken, posteriorly, and the broken surface only superficially resembles a “zig-zag” shape.

Fig. 7. Jaw fragments of an indeterminate metriorhynchid (MZ VIII Vr-72) from an unknown locality and age, preserved on the opposite sides of a limestone block: the “premaxilla” (A1) and the “dentary” (A2) of Tyborowski and Błażejowski (2019a, b).
Tyborowski and Błażejowski (2019b) also noted that the “Pliosaurus” remains discovered at Krzyżanowice indicated the presence of a taxon comparable in size to Pliosaurus funkei Knutsen, Druckenmiller, and Hurum, 2012b from the middle Volgian (Tithonian) of Svalbard, Norway, and Pliosaurus kevani from the lower Kimmeridgian of Dorset, United Kingdom (Benson et al. 2013). Despite the fact that both taxa are mostly known from non-overlapping parts of their skeletons, which makes comparisons between them difficult, both Pliosaurus funkei and Pliosaurus kevani had skulls approximately 2 m long and their total body lengths could have reached, and possibly exceeded, 10 m (Knutsen et al. 2012b; Benson et al. 2013). However, Tyborowski and Błażejowski (2019b) did not specify how their estimates for the total body size of MZ VIII Vr-72 were obtained. In their first paper on the assemblage from Krzyżanowice, the authors noted that their study was preliminary (Tyborowski and Błażejowski 2019a: 741) and that they reported “only a small part of the enormous collection of marine reptile bones from the Krzyżanowice site” (Tyborowski and Błażejowski 2019a: 743), but they also noted that the material they described comprises some of the most informative specimens (Tyborowski and Błażejowski 2019a: 743). We assume, therefore, that their size estimates are based on the published “premaxilla” and “dentary”. If so, our reassessment of the material does not indicate the taxon could have approached the size of the aforementioned pliosaurids. The heights of the largest tooth crowns, the only elements that could be used to infer some size-related information, reach up to 40 mm. Even though the crowns of Pliosaurus kevani are up to 50 mm high, the size of pliosaurid teeth would be a weak proxy for their total body length. For example, whereas the tooth crowns of Pliosaurus kevani are apicobasally short comparable to the teeth of some other species referred to Pliosaurus (see Fischer et al. 2017: supplemental information data S3; Zverkov et al. 2018: fig. 6B), the reconstructed skull length of that taxon suggests it represented one of the largest Late Jurassic pliosaurids (Benson et al. 2013).
We propose that, rather than a 10-meter-long representative of Pliosaurus, MZ VIII Vr-72 actually represents a metriorhynchid thalattosuchian (Crocodylomorpha) that probably did not reach half the size (see e.g., Young et al. 2011). The teeth of MZ VIII Vr-72 share a number of characteristics present in dentitions of members of both major metriorhynchid subclades, the metriorhynchines and the geosaurines; though not developed in all of them. These include lingually curved and subcircular to indistinctly labiolingually compressed cross sections near the base of the crowns; indistinctly blunt apices; well-developed and continuous carinae; no constriction at the crown/root junction; and ornamentation that includes a combination of structural elements restricted to particular segments of the tooth crowns. When metriorhynchids with such dental morphology and general characteristics of enamel structural elements are considered, MZ VIII Vr-72 shares the presence of short, apicobasally oriented ridges/ridglets limited to the basal part of their crowns and changing apically into an indistinct to conspicuous vermicular pattern with some taxa, such as the geosaurines Suchodus brachyrhynchus (Eudes-Deslongchamps, 1868b), Torvoneustes spp., and Tyrannoneustes lythrodectikos Young, de Andrade, Brusatte, Sakamoto, and Liston, 2013a, and the metriorhynchines Gracilineustes leedsi Andrews, 1913, and Metriorhynchus superciliosus (Blainville, 1853) (Andrade et al. 2010; Young et al. 2013a, b). All of these taxa, however, appear to lack the smoother mid-section that is present in most teeth of MZ VIII Vr-72, in which the lingual surface is exposed. Also, as in Suchodus spp. and T. lythrodectikos, but unlike in the other metriorhynchids mentioned above, the crown ornamentation in MZ VIII Vr-72 is well-developed on the lingual side but near-absent labially (Fig. 8). The lingual ornamentation on the crowns of G. leedsi and M. superciliosus is better developed than that on the labial side as well but it is also well-noticeable labially.

Fig. 8. Selected tooth crowns of an indeterminate metriorhynchid (MZ VIII Vr-72) from an unknown locality and age, showing observable features of the enamel: “premaxillary” teeth in lingual view (A1, A2), “dentary” teeth (A3, A4), showing the only tooth crown accessible from lingual side (A3) and the smooth labial surface (A4). Note the absence of the smooth enamel band at the mid-section (A3). Abbreviations: dc, distal carina; rgl, ridglets; se, horizontal band of smooth enamel; vp, vermicular pattern. Scale bars 10 mm.
It is worth noting that some teleosauroids, such as the machimosaurins Lemmysuchus obtusidens (Andrews, 1909), Machimosaurus spp., and Yvridiosuchus boutilieri (Eudes-Deslongchamps 1868a), show teeth similar in certain features of their crown ornamentation to those of MZ VIII Vr-72 as well (Young et al. 2014; Johnson et al. 2018, 2020). Specifically, as in MZ VIII Vr-72, the tooth crowns of the machimosaurins show ridged basal halves of the crowns that gradually change into a vermicular pattern at the apex. However, unlike the ornamentation in the crowns of MZ VIII Vr-72, the enamel structural elements in the machimosaurins are of high relief. The tooth crowns of L. obtusidens, Machimosaurus spp., and Y. boutilieri further differ from those of MZ VIII Vr-72 in their morphology; they are conical, without labiolingual compression, and rather robust, and their apices are conspicuously blunt (see Young et al. 2014: figs. 9, 11, 21, 28, 29, 31–35; Johnson et al. 2018: fig. 12; Johnson et al. 2020: figs. 4L, 8D–F).
We were not given permission to examine the microscopic anatomy of the dentition of MZ VIII Vr-72 and so were unable to describe its carinal morphology, which is also used in thalattosuchian taxonomy (e.g., Andrade et al. 2010) and could possibly provide additional data important for determining the taxonomic affinity of the specimen.
Sauropterygia Owen, 1861
Plesiosauria de Blainville, 1835
Plesiosauroidea Welles, 1943
Cryptoclididae Williston, 1925
Cryptoclididae? indet.
Fig. 9.
Material.—MZ VIII Vr-73 (“ZPAL V-KRZ/32”) (see above with respect to the ambiguity regarding the catalogue number of the specimen), a pectoral vertebral centrum reported by Tyborowski and Błażejowski (2019a) as originating from the light yellow marl, upper Kimmeridgian of Krzyżanowice, Poland (Dąbrowska 1957; Borsuk-Białynicka and Młynarski 1968).
Tyborowski and Błażejowski (2019a, b) reported an isolated plesiosaur pectoral centrum (MZ VIII Vr-73 or “ZPAL V-KRZ/32”), though their taxonomic interpretation of the element differed slightly between these two studies. While originally considered to represent an indeterminate elasmosaurid (Tyborowski and Błażejowski 2019a: 747), in their second paper the authors referred the specimen to a plesiosauroid and noted that it resembled the vertebrae of elasmosaurids and cryptoclidids (Tyborowski and Błażejowski 2019b: 1000).
Description.—The centrum clearly pertains to the pectoral series at the cervical-dorsal transition; the rib facet is transected by the neurocentral suture and is formed by the parapophysis on the vertebral centrum and diapophysis on the neural arch (see e.g., Sachs et al. 2013 for discussion on plesiosaur pectorals). The centrum itself is slightly kidney-shaped in anterior/posterior view, wider than high, the rib facets are circular, and the subcentral foramina are widely separated. The dorsal aspect of MZ VIII Vr-73 (“ZPAL V-KRZ/32”) further indicates the presence of a fairly wide neural canal. Such an overall morphology corresponds well with the pectoral morphology of cryptoclidid plesiosauroids, such as Cryptoclidus eurymerus Phillips, 1871 (Brown 1981: fig. 9), Colymbosaurus megadeirus (Seeley, 1869) (Benson and Bowdler 2014: fig. 6), and Ophthalmothule cryostea Roberts, Druckenmiller, Cordonnier, Delsett, and Hurum, 2020 (Roberts et al. 2020: fig. 15).
Considering the morphology of the centrum and its stratigraphic provenance, the taxonomic placement among Cryptoclididae appears to be most likely. However, owing to the fact that no diagnostic features were observed that would allow us to unambiguously assign the centrum to that clade, we refer the specimen to Cryptoclididae? indet.

Fig. 9. A plesiosauroid (Cryptoclididae? indet.) pectoral centrum, MZ VIII Vr-73 (“ZPAL V-KRZ/32”), from the Kimmeridgian of Krzyżanowice, in anterior (A1), posterior (A2), dorsal (A3), ventral (A4), left lateral (A5), and right lateral (A6) views.
Vertebrata indet.
Fig. 10.
Material.—ZPAL V. 69/1 (“ZPAL V-KRZ/33”), an isolated tooth crown from the canary-yellow Nerinea limestone, upper Kimmeridgian of Krzyżanowice, Poland (Dąbrowska 1957; Borsuk-Białynicka and Młynarski 1968).
Tyborowski and Błażejowski noted that crocodylomorph material (“small and fragile teeth and tooth crowns”, Tyborowski and Błażejowski 2019a: 747) were discovered at Krzyżanowice and provided a brief description of a specimen calatogued under the number “ZPAL V-KRZ/33” (correct catalogue number of this specimen is ZPAL V. 69/1, Jolanta Kobylińska, personal communication 2020).
Description.—Although ZPAL V. 69/1 (“ZPAL V-KRZ/33”) was not figured by Tyborowski and Błażejowski (2019a), the crown was reported to show a “robust conical morphology”, “circular cross-section”, and round apex, and its enamel surface reportedly presented “numerous wrinkles” and “several pronounced vertical striae”. These features led Tyborowski and Błażejowski (2019a) to refer the specimen to Machimosaurus sp.
However, the tooth crown measures only ~5 mm in apicobasal height, whereas tooth crowns in specimens unambiguously referable to Machimosaurus are much larger and usually measure >30 mm (see e.g., Young et al. 2014). In addition, the crown has not been fully prepared yet and, as such, some of its characters, including the shape of the cross-section, cannot be assessed with certainty. Nevertheless, we fail to recognise feaures indicating unambiguous machimosaurin affinities. The apical region of the crown lacks the conspicuous anastomosed pattern that is observable in Machimosaurus spp. (Young et al. 2014) and the apicobasal ridges (“pronounced vertical striae” of Tyborowski and Błażejowski 2019a) are very wide, separated by a somewhat vermicular surface (“numerous wrinkles” of Tyborowski and Błażejowski 2019a), and appear to be of a much higher relief than in machimosaurins (Johnson et al. 2018, 2020).
The crown resembles teeth of sharks and coelacanths in some respects (Maciej Pindakiewicz, personal communication 2020), but it is not possible to provide precise comparisons by examination of an unprepared, possibly damaged crown. Pending its full preparation or visualisation via micro-computed tomography, it is difficult to confidently assign ZPAL V. 69/1 (“ZPAL V-KRZ/33”) to a specific taxonomic group, so it is here provisionally regarded as an indeterminate vertebrate.

Fig. 10. Isolated tooth crown ZPAL V. 69/1 (“ZPAL V-KRZ/33”) from the Kimmeridgian of Krzyżanowice, pertaining to an indeterminate vertebrate, incorrectly identified as Machimosaurus sp. by Tyborowski and Błażejowski (2019a).
Principal coordinates analysis of Late Jurassic plesiosaur and thalattosuchian teeth
Our principal coordinates analysis (PCoA), aimed to explore the morphospace occupation of MZ VIII Vr-72 with respect to other Late Jurassic marine reptiles (notably metriorhynchids and pliosaurids), separates all major groups (pliosaurids, plesiosauroids, metriorhynchids, and teleosauroids), with only a small subset of the morphospace occupied by teleosauroids overlapping with that of geosaurine metriorhynchids (Fig. 11). Thalattosuchians are clearly separated from plesiosaurs by the first principal coordinates axis. Most thalattosuchian specimens (all teleosauroids, all metriorhynchines, and the vast majority of geosaurines) are grouped on the negative side of the axis, whereas the plesiosauroids and pliosaurids form distinct clusters on its positive side. MZ VIII Vr-72 plots on the negative side of the first axis and is placed within the geosaurine morphospace. This result supports the referral of MZ VIII Vr-72 to Metriorhynchidae and possibly within Geosaurinae (and Geosaurini), though this needs to be treated with caution. Metriorhynchine morphospace overlaps with that of geosaurines but metriorhynchines are not as well represented as Geosaurinae in the original dataset of Foffa et al. (2018c) which may explain why they occupy only a small area of morphospace near the line separating the positive and negative sections of the first coordinate. It is essential to note that the evolution of craniodental shapes of some secondarily aquatic tetrapods is strongly affected by convergence (e.g., Fischer et al. 2017), which may sometimes hinder taxonomic assignments based on characters related with feeding ecology and lead to ambiguous interpretations. Convergent development of some dental features has been observed in different thalattosuchian lineages that shared a similar ecology. For instance, the tooth crowns pertaining to taxa that were inferred to have a durophagous diet, such as Torvoneustes spp. and Machimosaurini (Foffa et al. 2018c), show a blunt apex and enamel bearing a conspicuous pattern. Even though our PCoA largely separates these particular lineages (thus “recognizing” the convergent nature of the discussed traits), it cannot be ruled out that the morphospace occupation of MZ VIII Vr-72 is at least partly affected by convergence. In addition, MZ VIII Vr-72 could be scored only for 60% of the characters (12 out of 20) and additional preparation/CT scanning may provide new morphological data crucial for further testing the morphospace occupation of MZ VIII Vr-72 in the future.
Regardless, the metriorhynchid affinities of MZ VIII Vr-72 are evident and the assignment of the specimen to Pliosauridae is strongly rejected.
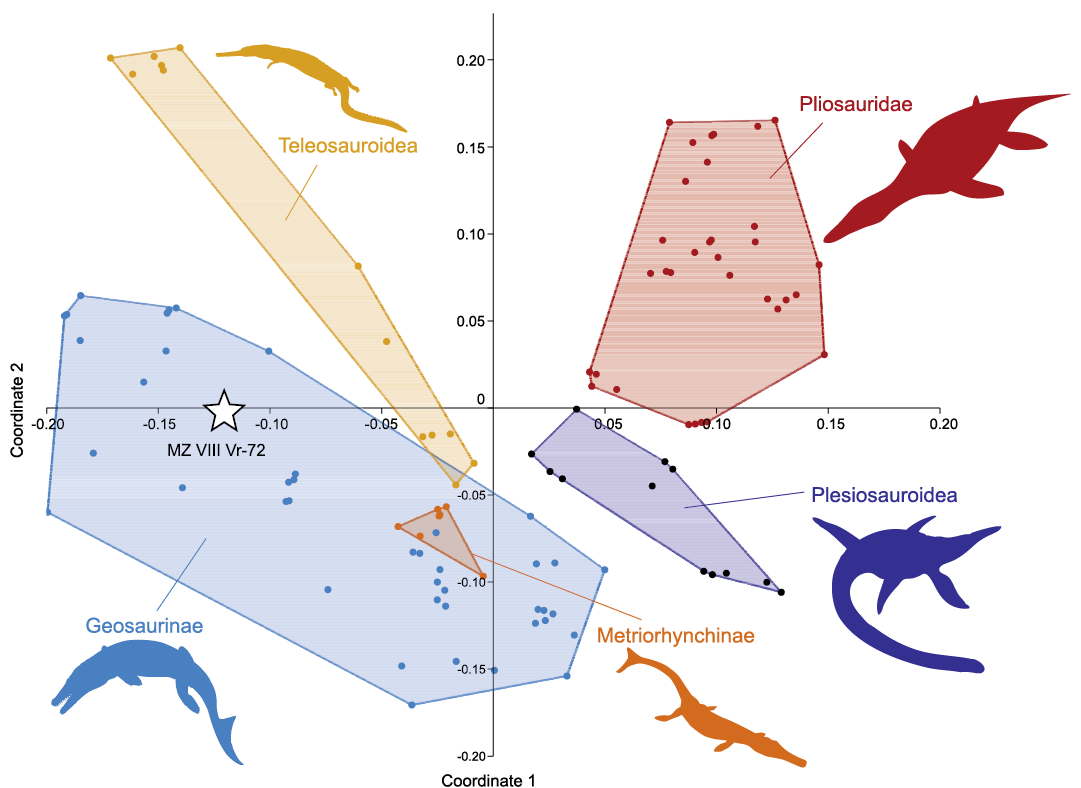
Fig. 11. Morphospace occupation of MZ VIII Vr-72 (white star) among Jurassic marine reptiles (thalattosuchians and plesiosaurs), resulting from principal coordinates analysis of the dataset of Foffa et al. (2018c), segregated along principal coordinates 1 and 2. See SOM 3 for extended results of the principal coordinates analysis. Silhouettes obtained from phylopic.org: Geosaurinae (Dmitry Bogdanov, CC BY 3.0), Metriorhynchinae and Teleosauroidea (Gareth Monger, CC BY 3.0), Plesiosauroidea (Adam Stuart Smith, CC BY-SA 3.0), and Pliosauridae (Nobu Tamura, CC BY-SA 3.0).
Discussion
The uncertain provenance of the specimens from the “Krzyżanowice assemblage”.—Tyborowski and Błażejowski (2019a, b) reported that the material they described and figured was collected recently, in some cases directly contrasting their descriptions against the historically collected specimens. This is particularly clear in the description of the state of preservation observed in the turtle material (Tyborowski and Błażejowski 2019b: 998): the description includes references to the figure showing the historical material (MZ VIII Vr-71), yet it ends with a statement that “[t]he remains of turtles collected during the 60s of the 20th century by the researchers from the PAS [Polish Academy of Sciences] also show a division into the three described preservation categories”. Though the current HTML version of Tyborowski and Błażejowski (2019a) states that “[t]hese specimens were discovered in 1962 by Teresa Maryańska, Gwidon Jakubowski (Museum of Earth, PAS), Zofia Kielan-Jaworowska and Magdalena Borsuk-Białynicka (Institute of Paleobiology, PAS)”, this sentence was not present in the in press version of that article and (at the moment of publication of this paper) has still not been added to the paginated PDF version. Furthermore, although it was clarified that the material originated from the 60s, the reference to its initial description by Borsuk-Białynicka and Młynarski (1968) is still missing.
The history of the metriorhynchid MZ VIII Vr-72 (“Pliosaurus sp.” of Tyborowski and Błażejowski 2019a, b) is enigmatic as well. As already noted, the specimen shares its catalogue number with an indeterminate teleosaurid (“Peloneustes sp.” of Maryańska 1972) from the Oxfordian of Załęcze Wielkie, located over 200 km west from Krzyżanowice. Just like MZ VIII Vr-71, MZ VIII Vr-72 comprises historical material (Daniel Tyborowski, personal communication 2020). According to Tyborowski and Błażejowski (2019a: 743, 2019b: 996–997), the same team that collected the turtle specimens in the 60s also unearthed skull fragments belonging to a pliosaurid, including the specimen that was later assigned the catalogue number MZ VIII Vr-72 (D. Tyborowski, personal communication 2020). However, this raises a few questions:
(i) According to Gwidon Jakubowski (personal communication 2020) who organized the excavations in Krzyżanowice in 1962, during the first week of the fieldwork, when he was present in the outcrop, the team did not find any vertebrate specimens. The second stage of the fieldwork was performed by Magdalena Borsuk-Białynicka and Teresa Maryańska, and during that time Jakubowski was absent. When he returned at the end of the excavations, the collected material had already been packed, so he had no possibility to see the specimens. Jakubowski recalls, nonetheless, that Maryańska spoke about “a jaw with teeth” of a marine reptile that she was excited to work on in the future. In fact, the memory of Jakubowski was the only reason why the specimen was identified by Tyborowski and Błażejowski (2019a, b) as originating from Krzyżanowice (D. Tyborowski, personal communication 2020) because, according to the curator of the palaeozoological collection of MZ (Barbara Studencka, personal communication 2020), it has no associated documentation. Although Jakubowski is convinced that the specimen MZ VIII Vr-72 of Tyborowski and Błażejowski (2019a, b) was found by Maryańska during these excavations, he admits that it might have been in fact collected from some other, neighbouring locality. Interestingly, Magdalena Borsuk-Białynicka (personal communication 2020) explicitly denies that the specimen was found by her and she does not recall Teresa Maryańska (deceased 2019) finding “anything noteworthy”, either.
(ii) No pliosaurid remains from the Kimmeridgian of Krzyżanowice are mentioned either in Borsuk-Białynicka and Młynarski (1968), a paper that was published six years after the excavations, or in Maryańska (1972), a paper specifically tackling the pliosaurid material from Poland authored by an employee of the Museum of the Earth in Warsaw who had supposedly participated in the excavations at this site (Tyborowski and Błażejowski 2019a, b). Also, no such specimens have ever been mentioned in the reviews of the Museum of the Earth’s collections published by Jakubowski et al. (1980) and Jakubowski et al. (1984). In 1984, the collection from Krzyżanowice comprised only MZ VIII Vr-71 (“Remains of the turtle from Krzyżanowice […]; 1 species, 1 specimen (carapace plates)”) and the only other Jurassic specimen in the collection was MZ VIII Vr-72: the material described by Maryańska (1972) from the Oxfordian of Załęcze Wielkie (Jakubowski et al. 1984: 378). As mentioned above, the metriorhynchid jaws described herein (MZ VIII Vr-72) were never recorded in the inventory book of MZ (B. Studencka, personal communication 2020).
(iii) Assigning specimens from two distant localities (Krzyżanowice and Załęcze Wielkie) and two different geological ages (Kimmeridgian and Oxfordian) under a single catalogue number, instead of assigning the Krzyżanowice specimen a new number or even adding it to the assemblage from the very same locality and age (MZ VIII Vr-71, which was already present in the collection), is counterintuitive. This confusion, however, may be caused by an error of Tyborowski and Błażejowski who assumed that the specimen was a part of MZ VIII Vr-72 while it was likely never assigned with any inventory or catalogue number. Tyborowski and Błażejowski never contacted the curator of the palaeozoology collection of MZ to confirm a formal assignment in the inventory (B. Studencka, personal communication 2020). Adding up to the confusion, there is yet another specimen with the catalogue number MZ VIII Vr-72—a dentary fragment of a lepidosauromorph from the Olenekian of Czatkowice that was figured by Borsuk-Białynicka et al. (1999: fig. 5A). The use of that number by Borsuk-Białynicka et al. (1999) is likely a result of Maryańska not entering the record of her original specimen (Maryańska 1972) to the inventory book of MZ and subsequently forgetting that the number MZ VIII Vr-72 had already been preoccupied (B. Studencka, personal communication 2020).
(iv) One of the blocks originally described as MZ VIII Vr-72 from the Oxfordian of Załęcze Wielkie and illustrated by Maryańska (1972: pl. 1: 2) was also depicted by Tyborowski and Błażejowski (2019b: fig. 6) in the context of crocodylomorphs from Krzyżanowice (Tyborowski and Błażejowski 2019b: 1000).
(v) Both the “pliosaurid” specimen of Tyborowski and Błażejowski (2019a, b) and the lightly-coloured matrix surrounding the material differ markedly in colour and mode of preservation from MZ VIII Vr-71 and the typically-developed turmeric-olive-yellow Nerinea-yielding turtle limestone from Krzyżanowice (Dąbrowska 1957). This could be easily explained by their origin from a different horizon within the profile, but according to Tyborowski and Błażejowski (2019a, b) all of the specimens excavated at Krzyżanowice originate from the same level of the Nerinea limestone. However, according to Błażej Błażejowski (personal communication 2020), MZ VIII Vr-72 might have actually originated from the light-coloured limestone located above the turtle limestone. Yet, following Dąbrowska (1957), Borsuk-Białynicka and Młynarski (1968), and Tyborowski and Błażejowski (2019a), the layers overlying the canary-yellow limestone (cream-coloured marl) are devoid of macrofauna.
Considering that (i) no mentions of the specimen referred to Pliosaurus by Tyborowski and Błażejowski (2019a, b) could be traced in the MZ documentation or published historical accounts (Borsuk-Białynicka and Młynarski 1968; Maryańska 1972; Jakubowski et al. 1980, 1984); (ii) the horizon of origin of the specimen within the geological profile is uncertain; (iii) the specimen was published under the same catalogue number as an Oxfordian crocodylomorph specimen from Załęcze Wielkie; and (iv) the materials from Krzyżanowice have been confused with those from that locality (Tyborowski and Błażejowski 2019b), we propose that the “pliosaurid” specimen of Tyborowski and Błażejowski (2019a, b), reinterpreted here as an indeterminate metriorhynchid, might have originally belonged to the series of blocks described by Maryańska (1972). If correct, the following two possibilities arise:
(i) It was among the fragments from the Oxfordian of Załęcze Wielkie, but for some reason (e.g., insufficient preparation) its different morphology went unnoticed. If so, its number, MZ VIII Vr-72, would be correct from the beginning.
(ii) It was among the fragments from the Oxfordian of Częstochowa (Mirów), also described by Maryańska (1972), but it was brought by Maryańska as an uncatalogued specimen or borrowed from the collection of the Museum of Częstochowa, e.g., for additional studies, preparation, or exhibition, and later mistaken as a part of the only other specimen of a Jurassic marine reptile in the collection, namely MZ VIII Vr-72. The morphology of the specimen of Tyborowski and Błażejowski (2019a, b) is reminiscent of M.Cz. V 1293 figured by Maryańska (1972: pl. 2). Unfortunately, the most complete fragment of the latter has been lost during the 90s (Ewa Kaczmarzyk, Museum of Częstochowa, personal communication 2020), so its examination is not possible and the comparison of the mode of preservation is difficult because the photographs in Maryańska’s (1972) paper are black and white. Currently, only a small fragment of the jaw from that specimen is present in the collection of M.Cz. (Tyborowski 2019). Maryańska (1972: 202) described the material from Mirów as “[s]ome fragments of jaws and detached teeth”. As such, the collection might have originally comprised more specimens than today, possibly including those that are currently in the possession of the Museum of the Earth.
Another possible explanation is that the specimen was delivered to Maryańska by an unknown donor later, possibly for description, and comes from an unknown locality and geological time interval.
Jakubowski’s recollection of Maryańska speaking about a jaw and teeth of a marine reptile during the fieldwork season of 1962 may also be explained. A series of Late Cretaceous mosasaurid specimens, including a partial jaw and teeth had been found by Andrzej Sulimski in 1960–1962 in Maruszów and a single tooth was collected around the same time by Teresa Maryańska in Nasiłów (Sulimski 1968; see also Machalski et al. 2003). The specimens of Sulimski are currently housed in ZPAL, but the tooth found by Maryańska is part of the MZ collection (MZ VIII/Vr-66). Although Sulimski (1968) did not provide the discovery date for the former, merely mentioning Maryańska as the donor, the specimen is mentioned as found by “the employees of MZ, 1962” (Jakubowski et al. 1984). It seems, therefore, plausible that Maryańska had already found MZ VIII/Vr-66 when she participated in the original excavations at Krzyżanowice, knew about the discoveries of Sulimski, and discussed them with her co-workers in Krzyżanowice in 1962, possibly confusing Jakubowski who re-joined them later. Due to Maryańska’s demise, this explanation must, unfortunately, remain speculative. In any case, the geographic and temporal origin of the metriorhynchid specimen MZ VIII Vr-72 is uncertain and thus the specimen should not be considered in a palaeobiogeographical context without special caution.
Palaeobiogeographic and palaeoecological implications. —Following their taxonomic assessment of the material, Tyborowski and Błażejowski (2019a: 749–750) noted that the marine reptile assemblage formed a “palaeobiogeographically unique ecosystem” that was characterized by the co-occurrence of Mediterranean/Sub-Mediterranean and Boreal/Sub-Boreal faunal components, and proposed to recognize it as an example of a newly identified “palaeobiogeographic belt” that they termed the “Matyja-Wierzbowski Line”. According to Tyborowski and Błażejowski (2019a: 749–750), the “Matyja-Wierzbowski Line” can be understood as “the hypothetical faunal/biogeographical boundary separating the ecozones/palaeobiogeographical realms of the warm Tethys Ocean and colder seas at the north, a transitional zone between Mediterranean/Sub-Mediterranean Province and Boreal/Sub-Boreal Province” and is established specifically “by the co-occurrence of plesiochelyid turtles (Mediterranean fauna) and large pliosaurid reptiles (Boreal fauna)”.
Unfortunately, this concept is problematic for a number of reasons. Even though the authors specified the extent of the “Matyja-Wierzbowski Line” as extending “through Dorset and Yorkshire regions of England, between Fennoscandia and the Bohemian Massif and through the Teisseyre-Tornquist Zone” (Tyborowski and Błażejowski 2019a: 750), they did not provide any specific details on the faunal composition at localities positioned along the “line” and their evolution during the Late Jurassic. Only general information was provided to support the establishment of the new “palaeobiogeographic belt” and to show the differences between the faunas of the southern and northern provinces. Six European “assemblages” were briefly mentioned; two representing the Boreal fauna, two representing the Mediterranean fauna, and two supposedly lying on the “Matyja-Wierzbowski Line”. The Boreal fauna is characterized as comprising “large pliosaurids”, “plesiosaurids”, and ophthalmosaurid ichthyosaurs; the Mediterranean fauna, in turn, includes “plesiochelyid” turtles, crocodylomorphs, and ophthalmosaurid ichthyosaurs; and the “Matyja-Wierzbowski Line” includes all of the above-mentioned faunal components. Such categories, however, are too vague to be of any value, especially when ignoring their temporal settings (the Boreal assemblages discussed by Tyborowski and Błażejowski 2019a are Volgian [Tithonian–earliest Berriasian], and the Mediterranean and “Matyja-Wierzbowski Line” faunas fall within partly overlapping intervals of Kimmeridgian and Tithonian) and the complex evolutionary history of marine reptile clades during the Middle and Late Jurassic. For example, placing thalattosuchians collectively in a “crocodylomorph bin” significantly oversimplifies their Middle and Late Jurassic dispersal patterns and diverse ecological adaptations (see Young et al. 2013a, b, 2014; Foffa et al. 2015, 2018a–c, 2019; Stubbs and Benton 2016; Johnson et al. 2018, 2020). With respect to plesiosaurs, in turn, pliosaurids clearly occurred south of the “line” in the Kimmeridgian (Bardet et al. 1993). Tyborowski and Błażejowski (2019a: 749) partly specified what they meant when they spoke of “plesiosaurids” by listing Spitrasaurus wensaasi Knutsen, Druckenmiller, and Hurum, 2012b, Spitrasaurus larseni Knutsen, Druckenmiller, and Hurum, 2012b, and Djupedalia engeri Knutsen, Druckenmiller, and Hurum, 2012c as representatives of the group. However, these taxa actually belong to cryptoclidids (all from the Volgian strata of the Agardhfjellet Formation of central Spitsbergen) and are not particularly closely related to Plesiosaurus dolichodeirus Conybeare, 1824, that is one of the basalmost plesiosauroids (e.g., Madzia and Cau 2020; Roberts et al. 2020).
The establishment of the “Matyja-Wierzbowski Line” has been proposed based on the supposed co-occurrence of “plesiochelyid” turtles with a large pliosaurid in the upper Kimmeridgian of Krzyżanowice. Our study demonstrates, however, that the taxonomic interpretation of the assemblage by Tyborowski and Błażejowski (2019a, b) was largely erroneous and that the “pliosaurid’ specimen MZ VIII Vr-72 actually represents a metriorhynchid thalattosuchian (possibly a geosaurine).
Detailed assessment of the biogeographic history of marine reptiles during the Late Jurassic is beyond the scope of the present paper. However, the establishment of narrowly-defined palaeobiogeographic units, such as the proposed “Matyja-Wierzbowski Line”, should be preceded with large-scale specimen-based analyses of occurrence data gathered after careful research of the literature and after considerations of potential biases resulting from uneven temporal and environmental sampling.
Similarly, in light of the questioned taxonomical identification and ambiguous origin of MZ VIII Vr-72, as well as no documentation of fish taxa included in the trophic web reconstructed by Tyborowski and Błażejowski (2019a), their palaeoecological elaborations are dubious. Particularly puzzling is their interpretation of Craspedochelys? sp. (MZ VIII Vr-71) as a durophagous form. Skull material of Craspedochelys spp. is unknown. Among the “plesiochelyid” thalassochelydians, cranial or mandibular material is preserved in Plesiochelys bigleri (skulls [posterior parts], see Püntener et al. 2017a, b; Raselli and Anquetin 2019), Plesiochelys etalloni (skulls and mandibles, see Cuvier 1824; Rütimeyer 1873; Bräm 1965; Gaffney 1975a, 1976; Joyce 2000; Sterli et al. 2010; Carabajal et al. 2013; Anquetin et al. 2015; Anquetin and Chapman 2016), Plesiochelys planiceps (skull and mandible, see Owen 1842, 1884; Gaffney 1975a, 1976), Portlandemys gracilis (skull and mandible, see Anquetin et al. 2015; Püntener et al. 2017a), Portlandemys mcdowelli (skulls and mandible, see Parsons and Williams 1961; Gaffney 1975a, 1976), Tropidemys langii (mandible, see Püntener et al. 2017b), and Tropidemys seebachi (skull, see Joyce 2015; Joyce and Mäuser 2020). The material of Plesiochelys bigleri is too fragmentary to provide relevant data and the rest shows varied expansion of the primary palate, elongation of the snout, and broadening of the triturating surfaces, with Portlandemys mcdowelli exhibiting the strongest development in these aspects (Parsons and Williams 1961; Gaffney 1975a, 1976), but neither is particularly adapted for crushing hard prey. Most other Late Jurassic forms, such as Achelonia formosa (see Lortet 1892), Idiochelys fitzingeri (see Rütimeyer 1873; Lortet 1892), Jurassichelon moseri (see Püntener et al. 2017b), Jurassichelon oleronensis (see Rieppel 1980), Palaeomedusa testa (see Meyer 1860; Joyce 2003), also lack clear specializations for durophagy (although in most cases insufficient preservation or preparation of their cranial material makes it impossible to refute any incipient adaptations). The only three currently known Late Jurassic turtles for which durophagous ecology was explicitly proposed are Solnhofia spp. (see Parsons and Williams 1961; Gaffney 1975b; Broin 1994; Joyce 2000; Püntener et al. 2017b; Anquetin and Püntener 2020), Owadowia borsukbialynickae (see Szczygielski et al. 2018), and, possibly, Eurysternum wagleri (see Meyer 1839a; Anquetin and Joyce 2014). Relevant morphology is best studied in Solnhofia spp., which exhibits a suite of durophagous adaptations, including development of the secondary palate and expansion of the triturating surfaces of the skull and mandible co-occurring with an elongation of the snout (Parsons and Williams 1961; Gaffney 1975b; Broin 1994; Joyce 2000; Anquetin and Püntener 2020). Owadowia borsukbialynickae is a very fragmentary taxon, but its mandible presents a well-developed, posteriorly elongated, triturating surface, the structure of which corresponds better with some Cretaceous or younger turtle taxa than with Solnhofia spp. (Szczygielski et al. 2018). The skull and mandible morphology of Eurysternum wagleri is virtually unknown, save for several published images, all lacking useful details (Meyer 1839a; Anquetin and Joyce 2014), but its durophagous habits can be inferred based on the gut content of one of the specimens containing echinozoan remains (Joyce 2015). The youngest possible thalassochelydian cranial and mandibular material from the Lower Cretaceous of Dorset shows only a moderate expansion of the triturating surface (Anquetin and André 2020). Nonetheless, although the expansion of the triturating surface is correlated with a durophagous diet (Claude et al. 2004; Ferreira et al. 2015), this correspondence is not always strict and some species with only a moderate expansion (such as Owadowia borsukbialynickae and Eurysternum wagleri) may fall within the variability spectrum of non-durophagous forms (Walter Joyce, personal communication 2020). In any case, in light of the lack of any cranial remains referable to turtles from Krzyżanowice, in particular, and Craspedochelys spp., in general, the relative rarity of durophagy in Late Jurassic turtles, and no evidence of such adaptations in the supposedly closely related species (traditionally grouped into “Plesiochelyidae”), the interpretation of MZ VIII Vr-71 as a durophagous form (Tyborowski and Błażejowski 2019a: fig. 3) is unjustified.
Conclusions
Acknowledgements
DM, TS, and ASW conceived and designed the study and collected data from MZ and ZPAL specimens; DM described the metriorhynchid and plesiosaur material, compiled data and performed the principal coordinates analysis and interpreted its results; TS described the turtle material, 3D-scanned MZ VIII Vr-71, and compiled Table 1; DM and TS prepared figures; DM, TS and ASW wrote parts of the manuscript and approved the final draft. All authors consider their contributions equal and are listed in alphabetical order. We would like to express our gratitude to Błażej Błażejowski (ZPAL) for help with facilitating access to the specimens at MZ that originally appeared to remain inaccessible for our study. We thank Sandra Chapman (NHMUK), Rainer Schoch (SMNS), Barbara Studencka (MZ), Silvan Thüring (NMS), and Daniel Tyborowski (MZ) for access to their institutional collections. We also thank Magdalena Borsuk-Białynicka (ZPAL), Davide Foffa (National Museums Scotland, Edinburgh, UK), Niko Hoffmann (Germany), Gwidon Jakubowski (MZ), Katarzyna Jezierska, Ewa Kaczmarzyk (both M.Cz.), Hilary Ketchum (Oxford University Museum of Natural History, UK), Wojciech Knaga (Museum of Wieluń, Poland), Cezary Krawczyński (MZ), Stefan Meng (Institut für Geographie und Geologie, Ernst-Moritz-Arndt-Universität Greifswald, Germany), Remigiusz Molenda, Piotr Olejniczak (Faculty of Geology, Geophysics and Environmental Protection, Geological Museum, Kraków, Poland), Maciej Pindakiewicz (ZPAL), Irena Raselli (MJSN), Aubrey J. Roberts (University of Oslo, Norway), Sven Sachs (Naturkunde-Museum Bielefeld, Germany), Piotr Skrzycki, Piotr Sterkowicz (Poland), Jarosław Stolarski (ZPAL), Przemysław Sztajner (University of Szczecin, Poland), Marlena Świło (Polish Geological Institute-National Research Institute, Warsaw, Poland), Adam Zaremba (ZPAL) and Nikolay Zverkov (Borissiak Paleontological Institute of the Russian Academy of Sciences, Moscow, Russia) for their assistance and helpful discussions at various stages of the preparation of this study. Finally, we thank the Editor, Daniel E. Barta (Oklahoma State University Center for Health Sciences, Tahlequah, USA) for facilitating the processing of this manuscript, and the reviewers, Valentin Fischer (University of Liège, Belgium) and Walter G. Joyce (University of Fribourg, Switzerland), for valuable comments that improved the manuscript. Collection of comparative data for Jurassic and Cretaceous turtles was funded by grant no. 2016/23/N/NZ8/01823 from the National Science Centre (NCN), Kraków, Poland, awarded to TS.
References
Andrade, M.B. de, Young, M.T., Desojo, J.B. and Brusatte, S.L. 2010. The evolution of extreme hypercarnivory in Metriorhynchidae (Mesoeucrocodylia: Thalattosuchia) based on evidence from microscopic denticle morphology. Journal of Vertebrate Paleontology 30: 1451–1465. Crossref
Andrews, C.W. 1909. On some new steneosaurs from the Oxford Clay of Peterborough. Annals and Magazine of Natural History 3 (15): 299–308. Crossref
Andrews, C.W. 1913. A Descriptive Catalogue of the Marine Reptiles of the Oxford Clay. Based on the Leeds Collection in the British Museum (Natural History), London. Part II. 206 pp. Order of the Trustees, London.
Andrews, C.W. 1921. On a new chelonian from the Kimmeridge Clay of Swindon. Annals and Magazine of Natural History 7: 145–153. Crossref
Anquetin, J. and André, C. 2020. The last surviving Thalassochelydia—a new turtle cranium from the Early Cretaceous of the Purbeck Group (Dorset, UK). PaleorXiv [published online, https://doi.org/10.31233/osf.io/7pa5c]. Crossref
Anquetin, J. and Chapman, S.D. 2016. First report of Plesiochelys etalloni and Tropidemys langii from the Late Jurassic of the UK and the palaeobiogeography of plesiochelyid turtles. Royal Society Open Science 3 (1): 150470. Crossref
Anquetin, J. and Joyce, W.G. 2014. A reassessment of the Late Jurassic turtle Eurysternum wagleri (Eucryptodira, Eurysternidae). Journal of Vertebrate Paleontology 34: 1317–1328. Crossref
Anquetin, J. and Püntener, C. 2020. A new species of
the large-headed coastal marine turtle Solnhofia
(Testudinata, Thalassochelydia) from the Late Jurassic of NW
Switzerland. PeerJ 8: e9931.
Crossref
Anquetin, J., Deschamps, S., and Claude, J. 2014a. The rediscovery and redescription of the holotype of the Late Jurassic turtle Plesiochelys etalloni. PeerJ 2: e258. Crossref
Anquetin, J., Püntener, C., and Billon-Bruyat, J.-P. 2014b. A taxonomic review of the Late Jurassic eucryptodiran turtles from the Jura Mountains (Switzerland and France). PeerJ 2: e369. Crossref
Anquetin, J., Püntener, C., and Billon-Bruyat, J.-P. 2015. Portlandemys gracilis n. sp., a new coastal marine turtle from the Late Jurassic of Porrentruy (Switzerland) and a reconsideration of plesiochelyid cranial anatomy. PloS ONE 10 (6): e0129193. Crossref
Anquetin, J., Püntener, C., and Joyce, W.G. 2017. A review of the fossil record of turtles of the clade Thalassochelydia. Bulletin of the Peabody Museum of Natural History 58 (2): 317–369. Crossref
Arkhangelsky, M.S., Zverkov, N.G., Spasskaya, O.S., and Evgrafov, A.V. 2018. On the first reliable record of the ichthyosaur Ophthalmosaurus icenicus Seeley in the Oxfordian–Kimmeridgian beds of European Russia. Paleontological Journal 52 (1): 49–57. Crossref
Arratia, G., Schultze, H.-P., Tischlinger, H., and Viohl, G. (eds.) 2015. Solnhofen. Ein Fenster in die Jurazeit. 620 pp. Verlag Dr. Friedrich Pfeil, Munich.
Bardet, N. and Fernández, M. 2000. A new ichthyosaur from the Upper Jurassic lithographic limestones of Bavaria. Journal of Paleontology 74 (3): 503–511. Crossref
Bardet, N., Duffaud, S., Martin, M., Mazin, J.-M., Pereda-Suberbiola, X., and Vidier, J-P. 1997. Découverte de l’ichthyosaure Ophthalmosaurus dans le Tithonien (Jurassique supérieur) du Boulonnais, Nord de la France. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 205: 339–354. Crossref
Bardet, N., Falconnet, J., Fischer, V., Houssaye, A., Jouve, S., Pereda Suberbiola, X., Pérez García, A., Rage, J.-C., and Vincent, P. 2014. Mesozoic marine reptile palaeobiogeography in response to drifting plates. Gondwana Research 26 (3–4): 869–887. Crossref
Bardet, N., Mazin, J.M., and Martin, M. 1993. Une mandibule de Pliosaurus brachyspondylus (Reptilia, Sauropterygia) dans le Kimméridgien du Boulonnais (France). Neues Jahrbuch für Geologie und Paläntologie Monatshefte 11: 417–424. Crossref
Baur, G. 1888. Osteologische Notizen über Reptilien (Fortsetzung III). Zoologischer Anzeiger 11: 417–424.
Benson, R.B.J. and Bowdler, T. 2014. Anatomy of Colymbosaurus megadeirus (Reptilia, Plesiosauria) from the Kimmeridge Clay Formation of the U.K., and high diversity among Late Jurassic plesiosauroids. Journal of Vertebrate Paleontology 34: 1053–1071. Crossref
Benson, R.B.J., Evans, M., Smith, A.S., Sassoon, J., Moore-Fay, S., Ketchum, H.F., and Forrest, R. 2013. A giant pliosaurid skull from the Late Jurassic of England. PLoS ONE 8: e65989. Crossref
Bever, G.S., and Norell, M.A. 2017. A new rhynchocephalian (Reptilia: Lepidosauria) from the Late Jurassic of Solnhofen (Germany) and the origin of the marine Pleurosauridae. Royal Society Open Science 4 (11): 170570. Crossref
Blainville, H.D. de 1835. Description de quelques espèces de reptiles de la Californie, précédée de l’analyse d’une système générale d’Erpetologie et d’Amphibiologie. Nouvelles Archives du Muséum d’Histoire Naturelle 4: 233–296.
Blainville, H.D. de 1853. Letter by H.D. de Blainville. Bulletin de la Société Linnéenne de Normandie, Série 2 IX: 109–120.
Błażejowski, B. and Tyborowski, D. 2016. Remarkably well-preserved Upper Jurassic fossils of marine and terrestrial biota from the Owadów-Brzezinki Quarry, central Poland. In: A. Kaim, K. Hryniewicz, M.A. Bitner, R.G. Jenkins, N.H. Landman, and J.K. Cochran (eds.), 1st International Workshop on Ancient Hydrocarbon Seep and Cognate Communities, 13–17 June 2016, 35–42. Instytut Paleobiologii PAN, Warszawa.
Błażejowski, B., Gieszcz, P. and Tyborowski, D. 2016. New finds of well-preserved Tithonian (Late Jurassic) fossils from the Owadów-Brzezinki Quarry, Central Poland: A review and perspectives. Volumina Jurassica 14: 123–132.
Błażejowski, B., Lambers, P., Gieszcz, P., Tyborowski, D., and Binkowski, M. 2015. Late Jurassic jaw bones of halecomorph fish (Actinopterygii: Halecomorphi) studied with X-ray microcomputed tomography. Palaeontologia Electronica 18.3.53A: 1–10. Crossref
Błażejowski, B., Matyja, B.A., Wierzbowski, A., and Wierzbowski, H. 2014. A new exceptionally preserved Late Jurassic ecosystem (Owadów-Brzezinki, Central Poland) and its geological background. In: A. Feldman-Olszewska and A. Wierzbowski (eds.), Jurajskie utwory synkliny tomaszowskiej. Jurassica XI. Przewodnik wycieczek terenowych, abstrakty i artykuły, 21–26, Państwowy Instytut Geologiczny-PIB, Warszawa.
Borsuk-Białynicka, M. and Młynarski, M. 1968. The first finding of the Mesozoic marine turtle Tretosternon aff. punctatum Owen, 1848 in Poland. Prace Muzeum Ziemi 12: 217–222.
Borsuk-Białynicka, M., Cook, E., Evans, S.E., and Maryańska, T. 1999. A microvertebrate assemblage from the Early Triassic of Poland. Acta Palaeontologica Polonica 44: 167–188.
Borszcz, T. and Zatoń, M. 2009. Exploring the Jurassic at Zalas Quarry, southern Poland. Deposits Magazine 20: 4–7.
Bräm, H. 1965. Die Schildkröten aus dem oberen Jura (Malm) der Gegend von Solothurn. Schweizerische Paläontologische Abhandlungen 83: 1–190.
Broin, F. de 1994. Données préliminaires sur les chéloniens du Tithonien inférieur des calcaires lithographiques de Cajuers (Var, France). Geobios 16: 167–175. Crossref
Brown, D.S. 1981. The English Upper Jurassic Plesiosauroidea (Reptilia) and a review of the phylogeny and classification of the Plesiosauria. Bulletin of the British Museum (Natural History), Geology 35: 253–347.
Brown, D.S., Milner, A.C., and Taylor, M.A. 1986. New material of the plesiosaur Kimmerosaurus langhami from the Kimmeridge Clay of Dorset. Bulletin of the British Museum (Natural History), Geology 40: 225–234.
Carabajal, A.P., Sterli, J., Müller, J., and Hilger, A. 2013. Neuroanatomy of the marine Jurassic turtle Plesiochelys etalloni (Testudinata, Plesiochelyidae). PLoS ONE 8 (7): e69264. Crossref
Claude, J., Pritchard, P.C.H., Tong, H., Paradis, E., and Auffray, J.-C. 2004. Ecological correlates and evolutionary divergence in the skull of turtles: A geometric morphometric assessment. Systematic Biology 53 (6): 933–948. Crossref
Comment, G., Lefort, A., Koppka, J., and Hantzpergue, P. 2015. Le Kimméridgien d’Ajoie (Jura, Suisse): lithostratigraphie et biostratigraphie de la Formation de Reuchenette. Revue de Paléobiologie, Genève 34 (2): 161–194.
Conybeare, W.D. 1824. On the discovery of an almost perfect skeleton of the Plesiosaurus. Transactions of the Geological Society of London, Series 2 1: 381–389. Crossref
Cope, E.D. 1869. Synopsis of the extinct Batrachia, Reptilia and Aves of North America. Part I. Transactions of the American Philosophical Society, New Series 14: 1–104. Crossref
Cuvier, G. 1824. Recherches sur les ossemens fossiles, où l’on rétablit les caractères de plusieurs animaux dont les révolutions du globe ont détruit les espèces, T. 5, Pt. 2. 547 pp. G. Dufour et ed. d’Ocagne, Paris.
Dąbrowska, Z. 1957. Profil warstw pogranicza jury i kredy w Krzyżanowicach pod Iłżą. Biuletyn Instytutu Geologicznego 105: 205–210.
Dames, W. 1888. Verhandlungen der Gesellschaft. 1. Protokoll der November-Sitzung. Zeitschrift der Deutschen geologischen Gesellschaft 40: 777–778.
Deecke, W. 1907. Geologie von Pommern. 302 pp. Gebrüder Borntraeger, Berlin.
Delair, J.B. 1958. The Mesozoic reptiles of Dorset. Part one. Proceedings of the Dorset Natural History & Archaeological Society 79: 47–72.
Delsett, L.L., Roberts, A.J., Druckenmiller, P.S., and Hurum, J.H. 2019. Osteology and phylogeny of Late Jurassic ichthyosaurs from the Slottsmoya Member Lagerstätte (Spitsbergen, Svalbard). Acta Palaeontologica Polonica 64: 717–743. Crossref
Druckenmiller, P.S., Hurum, J.H., Knutsen, E.M., and Nakrem, H.A. 2012. Two new ophthalmosaurids (Reptilia: Ichthyosauria) from the Agardhfjellet Formation (Upper Jurassic: Volgian/Tithonian), Svalbard, Norway. Norwegian Journal of Geology 92: 311–339.
Dupret, V. 2004. The pleurosaurs: anatomy and phylogeny. Revue de Paléobiologie, Volume spécial 9: 61–80.
Dzik, J. 1992. Dzieje życia na Ziemi. Wprowadzenie do paleobiologii. 1st Edition. 447 pp. Państwowe Wydawnictwo Naukowe, Warszawa.
Dzik, J. 1997. Dzieje życia na Ziemi. Wprowadzenie do paleobiologii. 2nd Edition. 515 pp. Wydawnictwo Naukowe PWN, Warszawa.
Dzik, J. 2003. Dzieje życia na Ziemi. Wprowadzenie do paleobiologii. 3rd Edition. 523 pp. Wydawnictwo Naukowe PWN, Warszawa.
Dzik, J. 2011. Dzieje życia na Ziemi. Wprowadzenie do paleobiologii. 4th Edition 577 pp. Wydawnictwo Naukowe PWN, Warszawa.
Eudes-Deslongchamps, J.A. 1868a. Notes sur le squelette et la restauration du Teleosaurus cadomensis. Bulletin de la Société Linnéenne de Normandie, Série 2 3: 124–221.
Eudes-Deslongchamps, J.A. 1868b. Note sur un groupe de vertèbres et d’écailles rapportées au Teleosaurus hastifer et provenant des assises kimméridgiennes du Cap de la Hève. Bulletin de la Société Linnéenne de Normandie, Série 2 1: 146–155.
Feldmann, R.M., Schweitzer, C.E., and Błażejowski, B. 2015. A new species of lobster (Glypheoidea: Mecochiridae) from the Late Jurassic (late Tithonian) Lagerstätte from central Poland. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 275: 107–114. Crossref
Fernández, M.S. and de la Fuente, M.S. 1993. Las tortugas casiquelidias de las calizas litograficas Titonianas del area Los Catutos, Neuquen, Argentina. Ameghiniana 30: 283–295.
Fernández, M.S. and de la Fuente, M.S. 1988. Nueva tortuga (Cryptodira: Thalassemydidae) de la Formacion Vaca Muerta (Jurasico, Tithoniano) de la provincia del Neuquen, Argentina. Ameghiniana 25: 129–138.
Fernández, M.S., Herrera, Y., Vennari, V.V., Campos, L., de la Fuente, M., Talevi, M., and Aguirre-Urreta, B. 2019. Marine reptiles from the Jurassic/Cretaceous transition at the High Andes, Mendoza, Argentina. Journal of South American Earth Sciences 92: 658–673. Crossref
Ferreira, G.S., Rincón, A.D., Solórzano, A., and Langer, M.C. 2015. The last marine pelomedusoids (Testudines: Pleurodira): a new species of Bairdemys and the paleoecology of Stereogenyina. PeerJ 3: e1063. Crossref
Fischer, V., Bardet, N., Benson, R.B.J, Arkhangelsky, M.S., and Friedman, M. 2016. Extinction of fish-shaped marine reptiles associated with reduced evolutionary rates and global environmental volatility. Nature Communications 7 (1): 1–11. Crossref
Fischer, V., Benson, R.B.J., Zverkov, N.G., Soul, L.C., Arkhangelsky, M.S., Lambert, O., Stenshin, I.M., Uspensky, G.N., and Druckenmiller, P.S. 2017. Plasticity and convergence in the evolution of short-necked plesiosaurs. Current Biology 27: 1667–1676. Crossref
Fitzinger, L.J.F.J. 1843. Systema Reptilium. 106 pp. Braumüller et Seidel, Wien.
Foffa, D., Johnson, M.M., Young, M.T., Steel, L. and Brusatte, S.L. 2019. Revision of the Late Jurassic deep-water teleosauroid crocodylomorph Teleosaurus megarhinus Hulke, 1871 and evidence of pelagic adaptations in Teleosauroidea. PeerJ 7: e6646. Crossref
Foffa, D., Young, M.T., and Brusatte, S.L. 2015. Evidence of macrophagous teleosaurid crocodylomorphs in the Corallian Group (Oxfordian, Late Jurassic) of the UK. PeerJ 3: e1497. Crossref
Foffa, D., Young, M.T., and Brusatte, S.L. 2018a. Filling the Corallian gap: New information on Late Jurassic marine reptile faunas from England. Acta Palaeontologica Polonica 63: 287–313. Crossref
Foffa, D., Young, M.T., Brusatte, S.L., Graham, M.R., and Steel, L. 2018b. A new metriorhynchid crocodylomorph from the Oxford Clay Formation (Middle Jurassic) of England, with implications for the origin and diversification of Geosaurini. Journal of Systematic Palaeontology 16: 1123–1143. Crossref
Foffa, D., Young, M.T., Stubbs, T.L., Dexter, K.G., and Brusatte, S.L. 2018c. The long-term ecology and evolution of marine reptiles in a Jurassic seaway. Nature Ecology & Evolution 2: 1548–1555. Crossref
Fraas, E. 1901. Die Meerkrokodile (Thalattosuchia n. g.) eine neue sauriergruppe der Juraformation. Jahreshefte des Vereins fur vaterlandische Naturkunde, Wurttemberg 57: 409–418.
Fraas, E. 1903. Thalassemys marina E. Fraas aus dem oberen weissen Jura von Schnaithei nebst Bemerkungen über die Stammesgeschichte der Schildkröten. Jahreshefte des Vereins für vaterlandische Naturkunde in Württemberg 59: 72–104.
Gaffney, E.S. 1975a. A taxonomic revision of the Jurassic turtles Portlandemys and Plesiochelys. American Museum Novitates 2574: 1–19.
Gaffney, E.S. 1975b. Solnhofia parsonsi, a new cryptodiran turtle from the Late Jurassic of Europe. American Museum Novitates 2576: 1–25.
Gaffney, E.S. 1976. Cranial morphology of the European Jurassic turtles Portlandemys and Plesiochelys. Bulletin of the American Museum of Natural History 157: 489–543.
Gallinek, E. 1895. Ueber den weissen Jura von Inowraclaw. Jahres-Bericht der Schlesischen Gesellschaft für Vaterländische Cultur 72: 23.
Gallinek, E. 1896. Der obere Jura bei Inowrazlaw in Posen. Verhandlungen der Kaiserlichen RussischenMineralogischen Gesellschaft zu St. Petersburg, Zweite Serie 33: 353–427.
Gasparini, Z., Spalletti, L., and de la Fuente, M.S. 1997. Tithonian marine reptiles of the western Neuquén Basin, Argentina. Facies and palaeoenvironments. Geobios 30: 701–712. Crossref
Gower, J.C. 1971. A general coefficient of similarity and some of its properties. Biometrics 27: 857–871. Crossref
Grabowska, A. 2013. Rybojaszczur do Gigantów. EKspress 36 (7): 6.
Groß, K. 1944. Über den Bau der Zähne von Pliosaurus ferox (Sauvage). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen B 1944: 227–230.
Hammer, Ø., Harper, D.A.T., and Ryan, P.D. 2001. PAST: PAleontological STatistics software package for education and data analysis. Palaeontologia Electronica 4 (1): 1–9.
Herzog, L.L. 2019. Morphological characteristics and variability of helochelydrid turtles in North America: Evidence for a new species.North Carolina State University, 88pp.
Hirszberg, F. 1924. Sur quelques vertèbres des ichtyosauriens et des plesiosauriens du Kimmeridgien et du Portlandien des environs de Tomaszów Rawski (sur la Pilica) [in Polish, with French summary]. Prace Polskiego Instytutu Geologicznego 1 (2–5): 199–232.
Hoffmann, N. 2005. Die Vertebraten des Pommerschen Malms von Czarnogłowy, Wrzosowo, Kłęby und Bardy (Oxford/Kimmeridge, Nordwestpolen). 72 pp. Institut für Geographie und Geologie der Ernst-Moritz-Arndt-Universität Greifswald, Greifswald.
Hoffmann, N. 2007. Upper Jurassic marl quarry of Kłęby (Klemmen). In: H.G. Röhling, C. Breitkreuz, T. Duda, W. Stackebrandt, A. Witkowski, and O. Uhlmann (eds.), Geo-Pomerania Szczecin 2007. Geology Cross-Bordering the Western and Eastern European Platform, 47. Państwowy Instytut Geologiczny, Warsaw.
Hoffmann, N. and Bickelmann, C. 2008. Upper Jurassic marine reptiles from Northwest Poland. Palherp 2008.
Jakubowski, G., Jakubowski, K., Kohlman-Adamska, A., Kosmowska-Ceranowicz, B., and Popiel-Barczyk, E. 1980. The Museum of the Earth in Warsaw, its collections, aims, and publications. Journal of the Society for the Bibliography of Natural History 9: 435–447. Crossref
Jakubowski, G., Mączyńska, S., and Popiel-Barczyk, E. 1984. Muzeum Ziemi Polskiej Akademii Nauk. Zbiory paleozoologiczne. Przegląd Zoologiczny 28: 370–387.
Jansen, M. and Klein, N. 2014. A juvenile turtle (Testudines, Eucryptodira) from the Upper Jurassic of Langenberg Quarry, Oker, Northern Germany. Palaeontology 57: 743–756. Crossref
Jentzsch, A. 1884. Über den Jura der Gegend von Inowrazlaw. Schriften der Physikalisch-Ökonomischen Gesellschaft zu Königsberg 24: 41–45.
Johnson, M.M., Young, M.T., and Brusatte, S.L. 2020. Re-description of two contemporaneous mesorostrine teleosauroids (Crocodylomorpha: Thalattosuchia) from the Bathonian of England and insights into the early evolution of Machimosaurini. Zoological Journal of the Linnean Society 189 (2), 449–482. Crossref
Johnson, M.M., Young, M.T., Steel, L., Foffa, D., Smith, A.S., Hua, S., Havlik, P.E., Howlett, E.A., and Dyke, G. 2018. Re-description of “Steneosaurus” obustidens Andrews, 1909, an unusual macrophagous teleosaurid crocodylomorph from the Middle Jurassic of England. Zoological Journal of the Linnean Society 182: 385–418. Crossref
Joyce, W.G. 2000. The first complete skeleton of Solnhofia parsonsi (Cryptodira, Eurysternidae) from the Upper Jurassic of Germany and its taxonomic implications. Journal of Paleontology 74: 684–700. Crossref
Joyce, W.G. 2003. A new Late Jurassic turtle specimen and the taxonomy of Palaeomedusa testa and Eurysternum wagleri. PaleoBios 23 (3): 1–8.
Joyce, W.G. 2015. Schildkröten (Testudines). In: G. Arratia, H.-P. Schultze, H. Tischlinger, and G.Viohl, (eds.), Solnhofen. Ein Fenster in Die Urzeit, 415–421. Verlag Dr. Friedrich Pfeil, Munich.
Joyce, W.G. 2017. A review of the fossil record of basal Mesozoic turtles. Bulletin of the Peabody Museum of Natural History 58 (1): 65–113. Crossref
Joyce, W.G. and Anquetin, J. 2019. A review of the fossil record of nonbaenid turtles of the clade Paracryptodira. Bulletin of the Peabody Museum of Natural History 60: 129–155. Crossref
Joyce, W.G. and Mäuser, M. 2020. New material of named fossil turtles from the Late Jurassic (late Kimmeridgian) of Wattendorf, Germany. PLoS ONE 15 (6): e0233483. Crossref
Joyce, W.G., Chapman, S.D., Moody, R.T.J., and Walker, C.A. 2011. The skull of the solemydid turtle Helochelydra nopcsai from the early Cretaceous of the Isle of Wight (UK) and a review of Solemydidae. Special Papers in Palaeontology 86: 75–97.
Karl, H.-V., Gröning, E., and Brauckmann, C. 2012a. Revision of Tropidemys seebachi Portis, 1878 (Testudines: Eucryptodira) from the Kimmeridgian (Late Jurassic) of Hannover (northwestern Germany). Studia Palaeocheloniologica 4: 11–24.
Karl, H.-V., Tichy, G., and Valdiserri, D. 2012b. Sontiochelys cretacea Stache, 1905 and new description of the families Eurysternidae Dollo, 1886 and Thalassemydidae Rütimeyer, 1873 (Testudines: Cryptodira). Studia Geologica Salmanticensia 48: 45–76.
Ketchum, H.F. and Benson, R.B.J. 2011. The cranial anatomy and taxonomy of Peloneustes philarchus (Sauropterygia, Pliosauridae) from the Peterborough Member (Callovian, Middle Jurassic) of the United Kingdom. Palaeontology 54: 639–665. Crossref
Kin, A. and Błażejowski, B. 2012. Polskie Solnhofen [Polish Solnhofen]. Przegląd Geologiczny 60: 375–379.
Kin, A., Błażejowski, B., and Binkowski, M. 2012. The “Polish Solnhofen”: A long awaited alternative? Geology Today 28: 91–94. Crossref
Kin, A., Gruszczyrski, M., Martill, D.M., Marshall, J., and Błażejowski, B. 2013. Palaeoenvironment and taphonomy of a Late Jurassic (Late Tithonian) Lagerstätte from central Poland. Lethaia 46: 71–81. Crossref
Klein, J.T. 1760. Classification und kurze Geschichte der vierfüßigen Thiere. 431 pp. Jonas Schmidt, Lübeck.
Knutsen, E.M. 2012. A taxonomic revision of the genus Pliosaurus (Owen, 1841a) Owen, 1841b. Norwegian Journal of Geology 92: 259–276.
Knutsen, E.M., Druckenmiller, P.S., and Hurum, J.H. 2012a. A new plesiosauroid (Reptilia: Sauropterygia) from the Agardhfjellet Formation (Middle Volgian) of central Spitsbergen, Norway. Norwegian Journal of Geology 92: 213–234.
Knutsen, E.M., Druckenmiller, P.S., and Hurum, J.H. 2012b. A new species of Pliosaurus (Sauropterygia: Plesiosauria) from the Middle Volgian of central Spitsbergen, Norway. Norwegian Journal of Geology 92: 235–258.
Knutsen, E.M., Druckenmiller, P.S., and Hurum, J.H. 2012c. Two new species of long-necked plesiosaurians (Reptilia: Sauropterygia) from the Upper Jurassic (Middle Volgian) Agardhfjellet Formation of central Spitsbergen. Norwegian Journal of Geology 92: 187–212.
Krebs, B. 1967. Der Jura-Krokodilier Machimosaurus H.v. Meyer. Paläontologische Zeitschrift 41: 46–59. Crossref
Krzeczyńska, M., Wierzbowski, A., Woźniak, P., Świło, M., and Chećko, A. 2020. Działania Muzeum Geologicznego Państwowego Instytutu Geologicznego-Państwowego Instytutu Badawczego prowadzone w celu wykorzystania edukacyjnego i ochrony starych kamieniołomów. Przegląd Geologiczny 68: 187–211.
Lapparent de Broin, F. de 2001. The European turtle fauna from the Triassic to the Present. Dumerilia 4: 155–217.
Lapparent de Broin, F. de and Murelaga, X. 1999. Turtles from the Upper Cretaceous of Laño (Iberian Peninsula). Estudios del Museo de Ciencias Naturales de Alava 14 (S1): 135–211.
Lapparent de Broin, F. de, Lange-Badré, B., and Dutrieux, M. 1996. Nouvelles decouvertes de tortues dans le Jurassique supérieur du Lot (France) et examen du taxon Plesiochelyidae. Revue de Paleobiologie 15: 533–570.
Lomax, D.R. 2015. The first plesiosaurian (Sauropterygia, Pliosauridae) remains described from the Jurassic of Poland. Palaeontologia Electronica 18 (2): 29A. Crossref
Lortet, L. 1892. Les reptiles fossiles du Bassin du Rhone. Archives du Muséum d’Historie Naturelle de Lyon 5: 3–28.
Lydekker, R. 1889. Catalogue of fossil Reptilia and Amphibia. Part III. Containing the order Chelonia. 239 pp. Order of the Trustees, London.
Maack, G.A. 1869. Die bis jetzt bekannten fossilen Schildkröten und die im oberen Jura bei Kelheim (Bayern) und Hannover neu aufgefundenen ältesten Arten derselben. Palaeontographica 18: 193–336.
Machalski, M., Jagt, J.W.M., Dortangs, R.W., Mulder, E.W.A., and Radwański, A. 2003. Campanian and Maastrichtian mosasaurid reptiles from central Poland. Acta Palaeontologica Polonica 48: 397–408.
Madzia, D. and Cau, A. 2020. Estimating the evolutionary rates in mosasauroids and plesiosaurs: discussion of niche occupation in Late Cretaceous seas. PeerJ 8: e8941. Crossref
Madzia, D., Sachs, S., and Lindgren, J. 2019. Morphological and phylogenetic aspects of the dentition of Megacephalosaurus eulerti, a pliosaurid from the Turonian of Kansas, USA, with remarks on the cranial anatomy of the taxon. Geological Magazine 156: 1201–1216. Crossref
Maliszewski, G. 2017. Skarby ukryte w kopalni. Tygodnik 7 Dni 1327: 12.
Mantell, G. 1827. Illustrations of the Geology of Sussex Containg a General View of the Geological Relations of the South-Eastern Part of England; With Figures and Descriptions of the Fossils of Tilgate Forest. 92 pp. Lupton Relfe, London.
Mantell, G. 1833. The Geology of the South-East of England. 415 pp. Longman, Rees, Orme, Brown, Green, and Longman, London. Crossref
Maryańska, T. 1972. Aberrant pliosaurs from the Oxfordian of Poland. Prace Muzeum Ziemi 20: 201–205.
Meyer, C.A. 1994. Depositional environment and paleoecology of the Solothurn Turtle Limestone (Kimmeridgian, Northern Switzerland). Geobios 16: 227–236. Crossref
Meyer, C.A. 2011. Amazing graze—grazing traces of sea urchins on turtles—an example from the Late Jurassic of Switzerland. Annalen des Naturhistorischen Museums in Wien A 113: 555–565.
Meyer, H. von 1839a. Eurysternum wagleri, Münster. Eine Schildkröte aus dem Kalkschiefer von Solnhofen. Beiträge zur Petrefacten-Kunde 1: 75–82.
Meyer, H. von 1839b. Idiochelys fitzingeri, eine Schildkröte aus dem Kalkschiefer von Kelheim. Beiträge zur Petrefacten-Kunde 1: 59–74.
Meyer, H. von 1860. Zur Fauna der Vorwelt. Reptilien aus dem lithographischen Schiefer des Jura in Deutschland und Frankreich. 144 pp. Heinrich Keller, Frankfurt am Main.
Meyer, H. von 1864. Parachelys eichstättensis aus dem lithographischen Schiefer von Eichstätt. Palaeontographica 11: 289–295.
Milner, A.R. 2004. The turtles of the Purbeck Limestone Group of Dorset, southern England. Palaeontology 47: 1441–1467. Crossref
Molenda, R. 1997. Zalas, Bear Mountain—History of Mining, Geology, Ecology. 32 pp. Studio Graficzne Szelerewicz i S-ka, Cracow.
Moon, B.C. and Kirton, A.M. 2018. Ichthyosaurs of the British Middle and Upper Jurassic. Part 2. Brachypterygius, Nannopterygius, Macropterygius and taxa invalida. Monographs of the Palaeontographical Society 172 (650): 85–177. Crossref
Owen, R. 1842. Report on British fossil reptiles. Part II. Report of the British Association for the Advancement of Science 11: 60–204.
Owen, R. 1853. Monograph of the Fossil Reptilia of the Wealden and Purbeck Formations. Part I. Chelonia (Pleurosteron &c.) [Purbeck.]. 12 pp. The Palaeontographical Society, London. Crossref
Owen, R. 1861. Palaeontology, or a Systematic Summary of Extinct Animals and Their Geological Relations. 463 pp. Adam and Charles Black, Edinburgh. Crossref
Owen, R. 1884. A History of British Fossil Reptiles. Vol. II. 719 pp. Cassell & Company Limited, London.
Parsons, T.S. and Williams, E.E. 1961. Two Jurassic turtle skulls: A morphological study. Bulletin of the Museum of Comparative Zoology 125: 41–107.
Pérez-García, A. 2014. Revision of the poorly known Dorsetochelys typocardium, a relatively abundant pleurosternid turtle (Paracryptodira) in the Early Cretaceous of Europe. Cretaceous Research 49: 152–162. Crossref
Pérez-García, A. 2015a. Largest shell of a basal eucryptodiran turtle reveals Late Jurassic Thalassemys hugii in the British record. Comptes Rendus Palevol 14: 555–561. Crossref
Pérez-García, A. 2015b. New data on the poorly-known Late Jurassic European turtles Thalassemys and Enaliochelys and description of a new basal eucryptodiran taxon. Journal of Iberian Geology 41: 21–30. Crossref
Pérez-García, A. 2015c. Revision of the British record of Tropidemys (Testudines, Plesiochelyidae) and recognition of its presence in the Late Jurassic of Portugal. Journal of Iberian Geology 41: 11–20. Crossref
Pérez-García, A. and Ortega, F. 2011. Selenemys lusitanica, gen. et sp. nov., a new pleurosternid turtle (Testudines: Paracryptodira) from the Upper Jurassic of Portugal. Journal of Vertebrate Paleontology 31: 60–69. Crossref
Pérez-García, A., Royo-Torres, R., and Cobos, A. 2015. A new European Late Jurassic pleurosternid (Testudines, Paracryptodira) and a new hypothesis of paracryptodiran phylogeny. Journal of Systematic Palaeontology 13: 351–369. Crossref
Phillips, J. 1871. Geology of Oxford and the Valley of the Thames. 523 pp. Clarendon Press, Oxford. Crossref
Pictet, F.-J. 1860. Deuxième partie. Description des fossiles. In: F.-J. Pictet and A. Jaccard (eds.), Description de Quelques Débris de Reptiles et de Poissons Fossiles Trouvés Dans l’étage Jurassique Supérieur (Virgulien) Du Jura Neuchâtelois, 15–88. J. Kessmann, Geneve.
Pictet, F.-J. and Humbert, A. 1857. Description d’une émyde nouvelle (Emys etalloni) du terrain Jurassique supérieur des environs de St. Claude. In: F.-J. Pictet (ed.), Matériaux Pour La Paléontologie Suisse, Première Série, 1–10. J. Kessmann, Geneve.
Portis, A. 1878. Über fossile Schildkröten aus dem Kimmeridgien von Hannover. Palaeontographica 25: 125–140.
Preussner 1886. Verhandlungen der Gesselschaft. 1. Protokoll der November-Sitzung. Zeitschrift der Deutschen Geologischen Gesellschaft 38: 916.
Püntener, C., Anquetin, J., and Billon-Bruyat, J.-P. 2015. Thalassemys bruntrutana n. sp., a new coastal marine turtle from the Late Jurassic of Porrentruy (Switzerland), and the paleobiogeography of the Thalassemydidae. PeerJ 3: e1282. Crossref
Püntener, C., Anquetin, J., and Billon-Bruyat, J.-P. 2017a. The comparative osteology of Plesiochelys bigleri n. sp., a new coastal marine turtle from the Late Jurassic of Porrentruy (Switzerland). PeerJ 5: e3482. Crossref
Püntener, C., Anquetin, J., and Billon-Bruyat, J.-P. 2017b. Vertébrés mésozoïques. Tortues. 357 pp. Office de la Culture, Porrentruy.
Püntener, C., Anquetin, J., and Billon-Bruyat, J.-P. 2020. New material of “Eurysternidae” (Thalassochelydia, Pan-Cryptodira) from the Kimmeridgian of the Swiss Jura Mountains. Palaeovertebrata 43 (1): e2. Crossref
Püntener, C., Billon-Bruyat, J.-P., Bocat, L., Berger, J., and Joyce, W.G. 2014. Taxonomy and phylogeny of the turtle Tropidemys langii Rütimeyer, 1873, based on new specimens from the Kimmeridgian of the Swiss Jura Mountains. Journal of Vertebrate Paleontology 34: 353–374. Crossref
Pusch, G.G. 1837. Polens Paläontologie oder Abbildung und Beschreibung der vorzüglichsten und der noch unbeschriebenen Petrefakten aus den Gebirgsformationen in Polen, Volhynien und den Karpathen nebst einigen allgemeinen Beiträgen zur Petrefaktenkunde und einem Versuch zu. 218 pp. E. Schweizerbart, Stuttgart.
Raselli, I. and Anquetin, J. 2019. Novel insights into the morphology of Plesiochelys bigleri from the early Kimmeridgian of Northwestern Switzerland. PLoS ONE 14 (5): e0214629. Crossref
Rehbinder, B. 1913. Die mitteljurassischen eisenerzführenden Tone längs dem südwestlichen Rande des Krakau-Wieluner Zuges in Polen. Zeitschrift der Deutschen Geologischen Gesellschaft 65: 181–349.
Rieppel, O. 1980. The skull of the Upper Jurassic cryptodire turtle Thalassemys, with a reconsideration of the chelonian braincase. Palaeontographica A 171: 105–140.
Roberts, A.J., Druckenmiller, P.S., Cordonnier, B., Delsett, L.L., and Hurum, J.H. 2020. A new plesiosaurian from the Jurassic–Cretaceous transitional interval of the Slottsmøya Member (Volgian), with insights into the cranial anatomy of cryptoclidids using computed tomography. PeerJ 8: e8652. Crossref
Roberts, A.J., Druckenmiller, P.S., Sætre, G.P., and Hurum, J.H. 2014. A new Upper Jurassic ophthalmosaurid ichthyosaur from the Slottsmøya Member, Agardhfjellet Formation of central Spitsbergen. PLoS ONE 9 (8): e103152. Crossref
Rütimeyer, L. 1873. Die fossilen Schildkröten von Solothurn und der übrigen Juraformation. Neue Denkschrift der allgemeinen schweizerischen naturforschenden Gesellschaft 25: 1–185.
Sachs, S., Hornung, J.J., and Kear, B.P. 2017. A new basal elasmosaurid (Sauropterygia: Plesiosauria) from the Lower Cretaceous of Germany. Journal of Vertebrate Paleontology 37: e1301945. Crossref
Sachs, S., Kear, B.P., and Everhart, M.J. 2013. Revised vertebral count in the “longest-necked vertebrate” Elasmosaurus platyurus Cope, 1868, and clarification of the cervical-dorsal transition in plesiosauria. PLoS ONE 8: e70877. Crossref
Sadebeck, A. 1865. Die oberen Jurabildungen im Pommern. Zeitschrift der Deutschen Geologischen Gesellschaft 17: 651–701.
Sassoon, J., Noe, L.F., and Benton, M.J. 2012. Cranial anatomy, taxonomic implications and palaeopathology of an Upper Jurassic pliosaur (Reptilia: Sauropterygia) from Westbury, Wiltshire, UK. Palaeontology 55: 743–773. Crossref
Scheyer, T.M., Pérez-García, A., and Murelaga, X. 2014. Shell bone histology of solemydid turtles (stem Testudines): palaeoecological implications. Organisms Diversity and Evolution 15: 199–212. Crossref
Seeley, H.G. 1869. Index to the Fossil Remains of Aves, Ornithosauria, and Reptilia, From the Secondary System of Strata Arranged in the Woodwardian Museum of the University of Cambridge. 143 pp. Deighton, Bell and Co., Cambridge.
Seeley, H.G. 1869. Index to the Fossil Remains of Aves, Ornithosauria,and Reptilia in the Woodwardian Museum of the University of Cambridge. 143 pp. Deighton, Bell and Co., Cambridge.
Seeley, H.G. 1875. Note on Pelobatochelys blakii and other vertebrate fossils exhibited by the Rev. J.F. Blake in illustration of his paper on the Kimmeridge Clay. Quarterly Journal of the Geological Society of London 31: 234–237. Crossref
Serratos, D.J., Druckenmiller, P.S., and Benson, R.B.J. 2017. A new elasmosaurid (Sauropterygia, Plesiosauria) from the Bearpaw Shale (Late Cretaceous, Maastrichtian) of Montana demonstrates multiple evolutionary reductions of neck length within Elasmosauridae. Journal of Vertebrate Paleontology 37: e1278608. Crossref
Skrzycki, P. 2010. Large ichthyosaur from the Oxfordian of Morawica. In: Z. Dubicka, P. Gorzelak, and Ł. Rakowicz (eds.), Konferencja Naukowa „Młodzi w Paleontologii”, 259–265. Institute of Paleobiology, Polish Academy of Sciences, Warsaw.
Sterli, J., Müller, J., Anquetin, J., and Hilger, A. 2010. The parabasisphenoid complex in Mesozoic turtles and the evolution of the testudinate basicranium. Canadian Journal of Earth Sciences 47: 1337–1346. Crossref
Stubbs, T.S. and Benton, M.J. 2016. Ecomorphological diversifications of Mesozoic marine reptiles: the roles of ecological opportunity and extinction. Paleobiology 42: 547–573. Crossref
Sulimski, A. 1968. Remains of Upper Cretaceous Mosasauridae (Reptilia) of Central Poland. Acta Palaeontologica Polonica 13: 243–250.
Sullivan, P.M. and Joyce, W.G. 2017. The shell and pelvic anatomy of the Late Jurassic turtle Platychelys oberndorferi based on material from Solothurn, Switzerland. Swiss Journal of Palaeontology 136: 323–343. Crossref
Szczygielski, T., Tyborowski, D., and Błażejowski, B. 2018. A new pancryptodiran turtle from the Late Jurassic of Poland and palaeobiology of early marine turtles. Geological Journal 53: 1215–1226. Crossref
Tarlo, B. 1960. A review of the Upper Jurassic pliosaurs. Bulletin of the British Museum (Natural History), Geology 4: 145–189.
Taylor, M.A. and Cruickshank, A.R.I. 1993. Cranial anatomy and functional morphology of Pliosaurus brachyspondylus (Reptilia: Plesiosauria) from the Upper Jurassic of Westbury, Wiltshire. Philosophical Transactions of the Royal Society B: Biological Sciences 341: 399–418. Crossref
Thiollière, V. 1850. Continuation des indications sommaires sur les espèces fossiles d’animaux et végétaux recueillies dans le gisement de Cirin. Annales des Sciences Physiques et Naturelles d’Agriculture et d’Industrie de Lyon 32: 128–129.
Tyborowski, D. 2016. A new ophthalmosaurid ichthyosaur from the Late Jurassic of Owadów-Brzezinki Quarry, Poland. Acta Palaeontologica Polonica 61: 791–803. Crossref
Tyborowski, D. 2017. Large predatory actinopterygian fishes from the Late Jurassic of Poland studied with X-ray microtomography. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 283: 161–172. Crossref
Tyborowski, D. 2019. Jurassic pliosaur remains in the collections of the Museum of Częstochowa [in Polish, with English abstract]. Rocznik Muzeum Częstochowskiego 18: 73–76.
Tyborowski, D. and Błażejowski, B. 2019a. New marine reptile fossils from the Late Jurassic of Poland with implications for vertebrate faunas palaeobiogeography. Proceedings of the Geologists’ Association 130: 741–751. Crossref
Tyborowski, D. and Błażejowski, B. 2019b. A marine vertebrate assemblage from the Upper Jurassic of Krzyżanowice near Iłża—with turtles, crocodylomorphs and large pliosaurs (NE margin of the Holy Cross Mountains, Poland) [in Polish, with English abstract]. Przegląd Geologiczny 67: 996–1001.
Tyborowski, D., Błażejowski, B., and Krystek, M. 2016. Reptile remains from the Upper Jurassic limestones of the Owadów-Brzezinki Quarry (central Poland) [in Polish, with English abstract]. Przegląd Geologiczny 64: 564–569.
Tyborowski, D., Skrzycki, P., and Dec, M. 2019. Internal structure of ichthyosaur rostrum from the Upper Jurassic of Poland with comments on ecomorphological adaptations of ophthalmosaurid skull. Historical Biology 20: 201–206.
Wagner, A. 1853. Beschreibung einer fossilen Schildkröte und etlicher anderer Reptilien-Ueberreste aus den lithographischen Schiefern und dem Grünsandstein. AAbhandlungen der Mathematisch-Physikalischen Klasse der Königlich Bayerischen Akademie der Wissenschaften 7: 241–264.
Wagner, A. 1861. Schildkröten und Saurier aus dem lithographischen Schiefer. Abhandlungen der Mathematisch-Physikalischen Klasse der Königlich Bayerischen Akademie der Wissenschaften 9: 67–124.
Welles, S.P. 1943. Elasmosaurid plesiosaurs, with description of new material from California and Colorado. University of California Memoirs 13: 125–254.
Williston, S.W. 1925. Osteology of the Reptiles. 300 pp. Harvard University Press, Cambridge, Massachusetts.
Wierzbowski, H., Dubicka, Z., Rychliński, T., Durska, E., Olempska-Roniewicz, E., and Błażejowski, B. 2016. Depositional environment of Sławno limestones (uppermost Jurassic, central Poland): evidence from microfacies analysis, microfossils and geochemical proxies. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 282: 81–108. Crossref
Young, M.T. and Andrade, M.B. de 2009. What is Geosaurus? Redescription of Geosaurus giganteus (Thalattosuchia: Metriorhynchidae) from the Upper Jurassic of Bayern, Germany. Zoological Journal of the Linnean Society 157: 551–585. Crossref
Young, M.T., Andrade, M.B. de, Brusatte, S.L., Sakamoto, M., and Liston, J. 2013a. The oldest known metriorhynchid super-predator: a new genus and species from the Middle Jurassic of England, with implications for serration and mandibular evolution in predacious clades. Journal of Systematic Palaeontology 4: 475–513. Crossref
Young, M.T., Andrade, M.B. de, Etches, S., and Beatty, B.L. 2013b. A new metriorhynchid crocodylomorph from the Lower Kimmeridge Clay Formation (Late Jurassic) of England, with implications for the evolution of dermatocranium ornamentation in Geosaurini. Zoological Journal of the Linnean Society 169: 820–848. Crossref
Young, M.T., Bell, M.A., Andrade, M.B. de, and Brusatte, S.L. 2011. Body size estimation and evolution in metriorhynchid crocodylomorphs: implications for species diversification and niche partitioning. Zoological Journal of the Linnean Society 163: 1199–1216. Crossref
Young, M.T., Brusatte, S.L., Andrade, M.B. de, Desojo, J.B., Beatty, B.L., Steel, L., Fernández, M.S., Sakamoto, M., Ruiz-Omeñaca, J.I., and Schoch, R. R. 2012. The cranial osteology and feeding ecology of the metriorhynchid crocodylomorph genera Dakosaurus and Plesiosuchus from the Late Jurassic of Europe. PLoS One 7 (9): e44985. Crossref
Young, M.T., Hua, S., Steel, L., Foffa, D., Brusatte, S.L., Thüring, S., Mateus, O., Ruiz-Omeñaca, J.I., Lepage, Y., Havlik, P.E., and Andrade, M.B. de 2014. Revision of the Late Jurassic teleosaurid genus Machimosaurus (Crocodylomorpha, Thalattosuchia). Royal Society Open Science 1: 1–42. Crossref
Zatoń, M. 2007. A tooth of a marine crocodylomorph from the Jurassic of the Cracow-Częstochowa Upland [in Polish, with English abstract]. Przegląd Geologiczny 55 (11): 961–964.
Zittel, K.A. von 1887. Handbuch der Paläontologie. Abteilung 1. Paläozoologie Band III. Vertebrata (Pisces, Amphibia, Reptilia, Aves). 900 pp. Oldenbourg, Munich.
Zverkov, N.G. and Efimov, V.M. 2019. Revision of Undorosaurus, a mysterious Late Jurassic ichthyosaur of the Boreal Realm. Journal of Systematic Palaeontology 17: 1183–1213. Crossref
Zverkov, N.G. and Jacobs, M.L. 2020. Revision of Nannopterygius (Ichthyosauria: Ophthalmosauridae): reappraisal of the “inaccessible” holotype resolves a taxonomic tangle and reveals an obscure ophthalmosaurid lineage with a wide distribution. Zoological Journal of the Linnean Society: 1–48. Crossref
Zverkov, N.G. and Prilepskaya, N.E. 2019. A prevalence of Arthropterygius (Ichthyosauria: Ophthalmosauridae) in the Late Jurassic—earliest Cretaceous of the Boreal Realm. PeerJ 7: e6799. Crossref
Zverkov, N.G., Arkhangelsky, M.S., and Stenshin, I.M. 2015a. A review of Russian Upper Jurassic ichthyosaurs with an intermedium/humeral contact. Reassessing Grendelius McGowan, 1976. Trudy Zoologičeskogo Instituta RAN 319 (4): 558–588.
Zverkov, N.G., Arkhangelsky, M.S., Pardo, P.J., and Beznosov, P.A. 2015b. On the Upper Jurassic ichthyosaur remains from the Russian North. Trudy Zoologičeskogo Instituta RAN 319 (1): 81–97.
Zverkov, N.G., Fischer, V., Madzia, D., and Benson, R.B.J. 2018. Increased pliosaurid dental disparity across the Jurassic–Cretaceous transition. Palaeontology 61: 825–846. Crossref
Acta Palaeontol. Pol. 66 (1): 99–129, 2021
https://doi.org/10.4202/app.00795.2020