
Comparative cranial osteology of subadult eucentrosauran ceratopsid dinosaurs from the Two Medicine Formation, Montana, indicates sequence of ornamentation development and complex supraorbital ontogenetic change
JOHN P. WILSON and JOHN B. SCANNELLA
Wilson, J.P. and Scannella, J.B. 2021. Comparative cranial osteology of subadult eucentrosauran ceratopsid dinosaurs from the Two Medicine Formation, Montana, indicates sequence of ornamentation development and complex supraorbital ontogenetic change. Acta Palaeontologica Polonica 66 (4): 797–814.
The eucentrosauran centrosaurines Einiosaurus procurvicornis and Achelousaurus horneri are the two most commonly recovered ceratopsids from the Campanian Two Medicine Formation of northwestern Montana, USA. Einiosaurus procurvicornis is known from at least 15 individuals recovered from two monospecific bonebeds, while Achelousaurus horneri is primarily known from one articulated adult cranium as well as two isolated subadult individuals previously referred to the taxon. Previous assessments of ontogeny in these taxa, alongside closely related centrosaurines, focused primarily on crania of mature individuals and disarticulated elements of immature individuals. Here we describe an articulated subadult Einiosaurus procurvicornis skull (MOR 456 8-8-87-1) from the Einiosaurus procurvicornis type locality bonebed and compare its cranial ornamental development with the only identically sized articulated subadult eucentrosauran skull from the Two Medicine Formation, MOR 591. These individuals represent the only known articulated subadult skulls from the hypothesized eucentrosauran lineage in the Two Medicine Formation, thereby enabling comparison of early ontogenetic developmental sequence and timing of all three primary cranial ornaments (nasal, supraorbital, and parietosquamosal frill). Comparison indicates that parietosquamosal frill and supraorbital ornamentation development may have preceded nasal horncore development in these taxa. MOR 456 8-8-87-1 fills a gap between the plesiomorphic morphology of juvenile Einiosaurus procurvicornis supraorbital horncores and the rounded, spheroid mass of bone which characterizes adults. The complete left squamosal of MOR 456 8-8-87-1 is of adult size, in contrast to its shorter face and immature facial ornamentation, which suggests that in Einiosaurus procurvicornis, the face and facial ornamentation development occurred after the parietosquamosal frill had reached adult size.
Key words: Dinosauria, Ceratopsia, Centrosaurine, ontogeny, Cretaceous, Two Medicine Formation, USA.
John P. Wilson [jackwilson1899@gmail.com], Varricchio Lab, Department of Earth Sciences, Montana State University, Bozeman, Montana, USA.
John B. Scannella [john.scannella@montana.edu], Museum of the Rockies and Department of Earth Sciences, Montana State University, Bozeman, Montana, USA.
Received 30 July 2020, accepted 9 October 2020, available online 15 December 2021.
Copyright © 2021 J.P. Wilson and J.B. Scannella. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Ceratopsids are among the most common dinosaur taxa preserved in the Upper Cretaceous terrestrial formations of the Western Interior of North America, and are known from nearly fifty taxa comprising two subfamilies, Chasmosaurinae and Centrosaurinae (Dodson et al. 2004; Ryan and Evans 2005; Horner et al. 2011). Commonly recovered taxa with large sample sizes and well-constrained stratigraphic occurrences, such as the chasmosaurine Triceratops from the Hell Creek Formation, provide insight into ceratopsid evolutionary trends and ontogeny (Horner and Goodwin 2006, 2008; Scannella and Horner 2010; Scannella and Fowler 2014; Scannella et al. 2014). Monodominant bonebeds of some ceratopsids, such as the centrosaurines Centrosaurus apertus, Coronosaurus brinkmani, Styracosaurus albertensis, and Pachyrhinosaurus lakustai, likewise reveal the ontogenetic changes within single, contemporaneous populations and enable a variety of ecological inferences (Ryan et al. 2001; Ryan and Russell 2005; Ryan et al. 2007; Currie et al. 2008).
The two most commonly recovered ceratopsids from the Campanian Two Medicine Formation of northwestern Montana, USA, are the eucentrosauran (Centrosaurini plus Pachyrhinosaurini; Chiba et al. 2017) centrosaurines Einiosaurus procurvicornis Sampson, 1995, and Achelousaurus horneri Sampson, 1995 (Horner et al. 1992; Sampson 1995; Sampson et al. 1997; Chiba et al. 2017). Einiosaurus procurvicornis is known from at least 15 individuals recovered from two monospecific, multi-individual bonebeds (Museum of the Rockies localities Dino Ridge Quarry [TM-023] and Canyon Bonebed [TM-046; type locality]), which are situated 47 and 45 meters, respectively, below the upper contact between the Two Medicine Formation and the overlying Bearpaw Formation (Horner et al. 1992; Rogers 1990; Sampson 1995). Achelousaurus horneri is primarily known from an articulated near-complete adult cranium with additional associated material, as well as from two single, isolated subadult individuals (Horner et al. 1992; Sampson 1995). Adult individuals of Einiosaurus procurvicornis (e.g., holotype MOR 456 8-9-6-1) are characterized by an elongate, heavily procurved nasal horncore, spheroid and convex masses of bone as supraorbital ornamentation, and a parietal bearing a single pair of elongate, straight processes at the P3 locus (Sampson 1995). The single adult specimen of Achelousaurus (holotype MOR 485) is diagnosed by its low nasal boss, high-ridged supraorbital bosses, and a parietal bearing a single pair of elongate, laterally curved P3 processes (Sampson 1995).
Here we describe an articulated subadult Einiosaurus procurvicornis skull, MOR 456 8-8-87-1, from the Einiosaurus procurvicornis type locality TM-046 bonebed, and compare its cranial ornamental development with the only other identically sized (facial length 44.5 cm; Table 1) articulated subadult eucentrosauran skull from the uppermost Two Medicine Formation, MOR 591 (Fig. 1). As the only articulated subadult skulls collected from the Two Medicine Formation from the hypothesized lineage containing Einiosaurus and Achelousaurus (Horner et al. 1992; Wilson et al. 2020), these specimens enable comparison of early ontogenetic developmental sequence and timing of nasal, supraorbital, and parietosquamosal ornamentation of single individuals of these eucentrosauran centrosaurines. Previous assessment of early Einiosaurus ontogenetic change was limited to disarticulated juvenile and subadult nasal and supraorbital specimens which could not unambiguously be assigned to single individuals (Sampson et al. 1997). Additionally, MOR 456 8-8-87-1 presents new information regarding the complexity and sequence of changes in the ontogeny of Einiosaurus supraorbital ornamentation, filling a gap between the plesiomorphic morphology found in juvenile supraorbital horncores and the rounded, spheroid mass of bone which characterizes adults. While MOR 456 8-8-87-1 lacks the majority of the parietal, its completely preserved left squamosal is of adult size, in contrast to its shorter face and immature facial ornamentation, which suggests that Einiosaurus experienced later expansion of the face and development of the facial ornamentation relative to the parietosquamosal frill. These subadult specimens provide details of the early ontogenetic changes which characterize the hypothesized eucentrosauran lineage present within the uppermost Two Medicine Formation, and provide a basis for hypotheses regarding the paleobiological significance of such trends in the evolution of ceratopsid dinosaurs.

Fig. 1. Lateral views of articulated crania of subadult eucentrosaurans from the Campanian Two Medicine Formation, Montana, USA. A. Einiosaurus procurvicornis Sampson, 1995, MOR 456 8-8-87-1. B. Eucentrosauran (Einiosaurus procurvicornis or Achelousaurus horneri Sampson, 1995), MOR 591. Photographs (A1 and B1), osteological line drawings (A2, B2). Face of MOR 591 mirrored, with parietosquamosal frill superimposed. The contact outlines between the postorbital, jugal, and squamosal in A2 are estimated due to lack of externally visible sutures and may appear different from MOR 591 due to some breaks and gaps. P4–P7, parietal processes.
Institutional abbreviations.—MOR, Museum of the Rockies, Bozeman, Montana, USA; TM, Museum of the Rockies Two Medicine Formation localities.
Other abbreviations.—EPS, epiparietosquamosal; P, parietal process; S, squamosal process.
Geological setting, material
and methods
This study primarily examines the cranial ornamentation (nasal and supraorbital horncores, jugals, and parietosquamosal frills) of subadult Einiosaurus procurvicornis MOR 456 8-8-87-1 and subadult eucentrosauran MOR 591 (previously referred to Achelousaurus; see below) within a comparative osteological framework, as they are the only articulated subadult eucentrosauran centrosaurine skulls of near-identical size (Table 1) from the Two Medicine Formation, and therefore the only subadult individuals of each taxon which preserve both the nasal and supraorbital ornamentation from single individuals (Fig. 1). Additional juvenile and subadult Einiosaurus procurvicornis specimens from the Canyon Bonebed (TM-046; MOR 456) are described which expand the cranial ontogenetic series of this taxon, though these are single elements from disarticulated individuals and therefore cannot be unambiguously corresponded to one another. Here, we follow the terminology and recommendations of Hone et al. (2016) in defining our specific ontogenetic assignments, and we consider “juveniles” as individuals which express plesiomorphic conditions with no ontogenetic modification towards the adult condition, and consider “subadults” as being individuals which express some modification of elements and ornamentation towards the adult condition, which may include a combination of both plesiomorphic and modified morphologies and appear ontogenetically transitional between the juvenile and adult conditions. Adults are here identified as individuals which express the diagnostic ornamental conditions of their taxon and bear osteological indicators of skeletal maturity (or cessation of rapid growth, but see Reizner 2010 and Reizner et al. 2020).
Table 1. Select measurements of MOR 456 8-8-87-1 and MOR 591 compared with the holotypes of Einiosaurus procurvicornis (MOR 456 8-9-6-1) and Achelousaurus horneri (MOR 485) and Einiosaurus procurvicornis supraorbitals from the TM-046 bonebed. Facial length measured from anteriormost point of the nasal to the posterior margin of the orbit. Squamosal length is measured from the anteriormost process to the EPS. Supraorbital ornamentation width measured from lateral margin to medialmost extent of ornamentation. Nasal horn length measured from dorsal margin of nares to dorsalmost preserved extend of horncore. Note, much of the rostrum of MOR 485 is reconstructed. Abbreviations: L, left; R, right; *, incomplete element; #, measured using ImageJ (Rueden et al. 2017).
| |
MOR 456 8-8-87-1 |
MOR 591 |
MOR 456 8-10-87-20 |
MOR 456 8-8-87-19 |
MOR 456 8-9-7-3 |
MOR 456 2020-C-1 |
MOR 456 8-23-87 |
MOR 456 2020-C-2 |
MOR 456 8-9-6-1 (holotype) |
MOR 485 |
|||
|
Basal skull length |
|
|
|
|
|
|
|
|
67 (Sampson 1995) |
~ 78 |
|||
|
Facial length |
42.5 (L) |
46.5 (R) |
44.5 |
|
|
|
|
|
|
54.5 |
~ 46.5*# |
||
|
Squamosal length |
31 (L) |
|
|
|
|
|
|
|
29 (L) |
30.9 (L) |
|||
|
Squamosal length (otic notch to EPS) |
33 |
34.5 |
|
|
|
|
|
|
31.5 |
|
|||
|
Supraorbital ornamentation length |
6.2 (L) |
6.5 (R) |
7.3 (R) |
5.2* |
5.4 |
5.3 |
5.5 |
6.7 |
7.2 |
6.2 (L) |
8.7 (R) |
12# (L) |
14.4# (R) |
|
Supraorbital ornamentation antpost length |
16.6 (L) |
|
15.6 (R) |
6.8 |
7 |
7 |
7.6 |
12.6 |
12.1* |
10.3 (L) |
14.6 (R) |
25.3 (L) |
24.3 (R) |
|
Supraorbital ornamentation width |
5.1 (L) |
5.3 (R) |
6.4 (R) |
4.5 |
5.5 |
6 |
5.6 |
10 |
11.1 |
9.5 (L) |
9.7 (R) |
16.8 (L) |
16.5 (R) |
|
Nasal horn length |
12.5* |
16.7* |
|
|
|
|
|
|
26.6 |
13.6# |
|||
|
Nasal horn ant-post length |
15.2 |
13.8 |
|
|
|
|
|
|
23.8 |
24.6# |
|||
MOR 456 8-8-87-1 is an articulated partial subadult Einiosaurus procurvicornis skull roof preserving paired nasals, lacrimals, palpebrals, postorbitals, frontals, and the complete left jugal, epijugal, quadratojugal, squamosal, and partial left lateral parietal bar. MOR 456 8-8-87-1 was recovered from the Canyon Bonebed (TM-046) in 1987. This site is a monospecific centrosaurine bonebed which produced multiple mature skulls referable to Einiosaurus procurvicornis, including the holotype skull (MOR 456 8-9-6-1) and additional disarticulated, mature cranial ornamentation diagnostic of Einiosaurus procurvicornis (Rogers 1990; Sampson 1995). No diagnostic adult material was recovered from this site which could be referred to any centrosaurine taxon other than Einiosaurus procurvicornis. Canyon Bonebed occurs in Lithofacies 5 (Horner et al. 2001) of the uppermost Two Medicine Formation, 45 meters below the upper contact between the Two Medicine Formation and the overlying Bearpaw Formation, within the Landslide Butte area, immediately adjacent to the USA/Canadian border and 40 km northwest of the town of Cut Bank, Montana (Sampson 1995: fig. 2; Rogers 1990: fig. 1). Additional postorbitals are described which were also recovered from the Canyon Bonebed, including complete left postorbital MOR 456 8-10-87-20, complete left postorbital MOR 456 8-9-7-3, partial right postorbital MOR 456 8-8-87-19, partial right postorbital MOR 456 2020-C-1, complete right postorbital and articulated palpebral and frontal MOR 456 8-23-87, and partial right postorbital MOR 456 2020-C-2.
MOR 591 is an articulated partial eucentrosauran skull with associated postcrania which preserves the rostral, palatal elements, paired premaxillae, partial right and left maxillae, paired nasals, right lacrimal, complete right and partial left palpebrals, complete right and partial left postorbitals, complete right and partial left frontals, complete right jugal and partial left jugal, complete right quadratojugal, proximal right squamosal and near-complete left squamosal, left lateral parietal bar, and elements of both lower jaws. The cranium is broken into three primary segments: the rostrum, including the nasal ornamentation, the supraorbital-jugal complex, and the frill. MOR 591 was discovered in 1989 as an isolated individual (MOR locality TM-077) within Lithofacies 5 (Horner et al. 2001) of the uppermost Two Medicine Formation, within 50 meters of the upper contact between the Two Medicine Formation and the overlying Bearpaw Formation (Jack Horner, personal communication 2020). The specimen was collected from a low, isolated hill of outcrop in an area with minimal exposure of the Two Medicine Formation along Blacktail Creek, in southern Glacier County (Sampson 1995; Jack Horner, personal communication 2020).
MOR 591 was collected approximately 5.7 km southwest of MOR 571 (TM-072), a partial subadult Achelousaurus cranium bearing incipient nasal and supraorbital bosses. The upper contact between the Two Medicine Formation and overlying Bearpaw Formation is visible at the top of the hill from which MOR 571 was collected (Jack Horner, personal communication 2020). MOR 591 was previously referred to Achelousaurus by Sampson (1995) and Sampson et al. (1997) on the basis of low ridges on the medial surface of the supraorbital horncore which were interpreted as incipient boss formation as well as the overall similarity of the parietal to both Einiosaurus and Achelousaurus; this referral has been maintained in subsequent studies (e.g., McDonald 2011). MOR 591 is referable to Eucentrosaura based on the presence of a prominent, pointed nasal horncore (which is not present in Nasutoceratopsini or in more basal centrosaurines such as Diabloceratops and Medusaceratops (Kirkland and DeBlieux 2010; Chiba et al. 2017), as well as diminutive, pointed supraorbital horncores, which is the plesiomorphic condition of Eucentrosaura. The plesiomorphic nasal and supraorbital horncores of MOR 591 align its affinities with the hypothesized eucentrosauran lineage containing Einiosaurus and the stratigraphically successive Achelousaurus. The adult holotype skull of Achelousaurus is known from the uppermost 20 meters of the Two Medicine Formation, while Einiosaurus is known from two bonebeds which occur 47 and 45 meters below the upper contact with the Bearpaw Formation (Sampson 1995). Therefore, with an ambiguous stratigraphic placement within the uppermost 50 meters of the formation, MOR 591 may pertain to either Einiosaurus or Achelousaurus; ontogenetic implications of both taxonomic referrals are discussed.
Results
Surface texture.—Bone surface texture of immature ceratopsids is typically striated, long-grained, and smooth, indicative of rapid growth that is ongoing (Sampson et al. 1997; Brown et al. 2009; Scannella and Horner 2010; Tumarkin-Deratzian 2010). As individuals mature and approach full body size, surface texture becomes mottled at transitional locations, and then rugose, as vessel canals become imprinted into the bone surface (Brown et al. 2009). However, despite its small size and state of cranial ornamentation development, MOR 456 8-8-87-1 expresses only well-preserved rugose bone and does not express any striated surface textures (Figs. 1–5). The anterior nasal processes and the lateral surfaces of the nasal horncore, the lateral surfaces of the supraorbital horncores, and the entire dorsal surface of the skull roof, express the most rugose bone on this skull, imprinted with deep vessel traces. The left jugal likewise lacks any long-grained or striated texture, and is instead dense with a subtle, rugose texture and several shallow vessel indentations. The anterior end of the left squamosal approaching the postorbital and jugal bears a rugose, pebbled texture. The body of the squamosal is likewise pebbled, with small dimples, several wide but shallow vessel canals, and an overall texture similar to the left jugal. The squamosal marginal crenulations express a more rugose texture with deeper canal traces than the majority of the body of the squamosal. In comparison, Einiosaurus procurvicornis holotype MOR 456 8-9-6-1 and a large, undescribed articulated skull from the same bonebed MOR 456 8-12-86-2, likewise bear dense and well-developed bone surfaces punctuated with prominent vessel canal impressions. In contrast to these specimens however, the vessel traces of MOR 456 8-8-87-1 are deeper in some places, such as on the nasal and supraorbital ornamentation, and approximately equivalent on non-ornamental surfaces.
Conversely, MOR 591 expresses surface textures more typical of an immature centrosaurine (Figs. 1–5). The premaxillae are predominantly smooth and long-grained in texture. The nasals bear several large foramina on the postnarial aprons, and the nasal horncore exhibits rugose texture with several deep vessel canals oriented dorsoventrally. The dorsal surface of the posterior nasals, along with the lateral face of the right postorbital, including the horncore, have an overall rugose appearance, due to vessel canals imprinted in the bone, though the fine texture of the bone is visibly woven. The dorsal surfaces of the postorbitals and frontals are considerably more rugose, with deep vessel canals, numerous small foramina, and an overall deeply pebbled texture. The lateral surfaces of the right jugal and quadratojugal are smooth, with some long-grained striations and three deep but short vessel canals. The anterior end of the right squamosal transitions to a more subtle mottled texture, with many small dimples imprinting the surface, though it has an overall smooth appearance. The dorsal surface of the left squamosal is somewhat mottled anteriorly, though with directional striations still visible extending towards the margin of the frill. The squamosal texture resumes an exclusively long-grained texture more posteriorly. Conversely, the ventral surface of the left squamosal is almost entirely mottled, with long-grained texture only occurring within about 3 cm of the frill margin. The dorsal surface of the anteriormost portion of the left lateral parietal bar is striated and long-grained, with the striations oriented towards the lateral margin of the parietal. Moving posteriorly along the dorsal surface of the left lateral bar, large portions of the surface are damaged, but preserved surfaces become somewhat mottled, with numerous small, shallow vessel traces, interspersed with regions of striated, long-grained texture.
Cranial sutures.—Though it has recently been demonstrated that cranial suture closure and obliteration is not a reliable indicator of archosaur maturity (Bailleul et al. 2016), and that immature ceratopsids may not express visibly open cranial sutures (Currie et al. 2016), sutures and contacts between bones are sometimes visible in immature dinosaurs. Some of the cranial sutures and contacts in MOR 591 are visible, including the contact between the premaxillae, the junction between premaxillae and anterior processes of the nasals, and the contacts between the right and left posterior processes of the premaxillae and the right and left postnarial aprons. The contacts between the right and left maxillae and the right and left posterior processes of the premaxillae are visible. In dorsal view, the intranasal contact is visible in the anterior processes of the nasals and the posterodorsal portion of the nasals. The anterior edge of the nasal horncore is complete and does not expose any of the intranasal suture, though the posterior edge of the nasal horncore is damaged, and the intranasal suture is visible as a wavy contact in this cross section. In ventral view, the intranasal contact is visible for the entire anteroposterior length of the nasals. The majority of the posterior half of the right maxilla is missing, though the jugal process of the maxilla is preserved in articulation with the anterior margin of the right jugal, with the contact being partially visible. Fewer sutures are visible posterior to the rostrum. No sutures visibly delineate the palpebrals and lacrimals. The contact between the right jugal and postorbital is not immediately visible, though the contact between the jugal and anterior end of the right squamosal is partially visible immediately dorsal to the lateral temporal fenestra. The posterior margin of the right postorbital is somewhat damaged, though it appears that the suture between the postorbital and the anterior portion of the squamosal remained open. The contact between the jugal and quadratojugal is clear. Dorsally, the sutures between the postorbitals and frontals are not visible. The intersection between the posterior margin of the nasals, the prefrontals, and the anterior margin of the frontals is partially visible. The contact between the left squamosal and the left lateral bar of the parietal is visible both dorsally and ventrally along the entire length of the parietosquamosal contact. In the preserved vertebrae of MOR 591, the neural arches remain articulated with their centra, with sutures being at least partially closed but still externally visible.
In contrast to MOR 591, almost none of the cranial sutures of MOR 456 8-8-87-1 are visible. The bonebed from which MOR 456 8-8-87-1 was recovered contains fully articulated, partially articulated, and fully disarticulated crania, and thus articulation and sutural condition does not appear to be taphonomically biased. In dorsal view, the intranasal contact is only visible posterior to the nasal horncore, and is not visible in the anterior nasal processes or within the nasal horncore itself. However, like MOR 591, the intranasal contact is visible for its entire anteroposterior length in ventral view. None of the sutures between the nasals, lacrimals, palpebrals, and postorbitals are visible, though this preorbital region is crushed and deformed on both the left and right sides of the skull. Likewise, the dorsal aspect of the skull roof has been somewhat transversely compressed at the skull midline, though regardless of this no sutures between the nasals, prefrontals, frontals, and postorbitals are visible in the undeformed areas. The contact between the left jugal and the left squamosal is only visible for approximately 3 cm immediately ventral to the lateral temporal fenestra, though the rest of the suture between the jugal and squamosal is not visible. Like MOR 591, the parietosquamosal contact is visible along its entire preserved length.
Nasal horncores.—The nasal horncores of MOR 456 8-8-87-1 and MOR 591 are near-identical and are both simple, erect, and recurved, tapering towards a pointed apex, which is the plesiomorphic condition for Eucentrosaura and “unmodified” compared to diagnostic, mature individuals of Einiosaurus procurvicornis and Achelousaurus horneri (Sampson et al. 1997; and Fig. 2 here). The nasal horncore of MOR 456 8-8-87-1 is damaged and missing its apex, but the anterior and posterior margins are both curved in posteriorly-oriented arcs so as to make the horncore recurved. The anterior margin of the nasal horncore first rises steeply and with less curvature than the posterior margin, then curves posteriorly in an abrupt manner approximately 4 cm above the base of the horncore, whereas the posterior margin of the horncore rises in a more gentle arc. There is no evidence that the missing apex would have expressed any procurvature, as is seen in more mature individuals of Einiosaurus procurvicornis. As noted above, the right and left lateral surfaces of the MOR 456 8-8-87-1 nasals are deeply imprinted with numerous vessel canal traces, contributing to an overall rugose texture of the nasals.

Fig. 2. Right lateral views of nasal horncores of Einiosaurus procurvicornis Sampson, 1995, MOR 456 8-8-87-1 (A) and eucentrosauran (Einiosaurus procurvicornis or Achelousaurus horneri Sampson, 1995), MOR 591 (B, mirrored), from the Campanian Two Medicine Formation, Montana, USA.
The nasal horncore of MOR 591 is near-identical to that of MOR 456 8-8-87-1 and is erect, recurved, and tapers to a pointed apex. The anterior margin of the horncore likewise rises steeply then curves posterodorsally towards the horncore apex, and the posterior margin rises more gradually towards the apex. The horncore of MOR 591 is more complete than that of MOR 456 8-8-87-1, and likewise shows no evidence of procurvature or boss formation. The horncore portion of the nasals of MOR 591 is likewise punctuated on its lateral surfaces with numerous vessel canal impressions and is rugose in texture.
Supraorbital ornamentation.—In contrast to the near-identical nasal ornamentation, the supraorbital ornamentation of MOR 456 8-8-87-1 and MOR 591 is markedly disparate (Figs. 3, 4). MOR 456 8-8-87-1 preserves both postorbitals and associated supraorbital ornamentation. The left supraorbital ornamentation is near-complete, with minor damage to the apex which gives it the artificial appearance of being anteroposteriorly bifurcated in lateral view. In lateral view, the left supraorbital ornamentation is sub-circular and rounded, with a near-flat lateral surface; the dorsolateral margins of the ornamentation on either side of the damaged apex rise gradually and likely would have contributed to a rounded apex lacking a sharp point. In anterior view, however, the majority of the supraorbital ornamentation projects medially in a near-flat manner, and does not slope downwards towards the skull midline (Fig. 4A). This gives the left supraorbital ornamentation the appearance of being an inflated and flattened platform of bone, rather than being a discrete horncore. Only the anteriormost edge of the supraorbital ornamentation, anterior to the damaged apex, slopes downwards towards the skull midline, whereas the majority of the ornamentation projects medially, sub-perpendicular to the flat lateral face of the ornamentation.
The right postorbital and supraorbital ornamentation is essentially identical to the corresponding left supraorbital ornamentation. In lateral view, it is sub-circular and rounded, lacking a pointed apex, and the lateral face of the postorbital is near-flat. Shallow pitting occurs at the dorsolateral margin of the ornamentation, which may contribute to the lack of a pointed apex (Fig. 3A). Like the left supraorbital ornamentation, when viewed anteriorly, the right supraorbital ornamentation projects medially as a flat platform of bone in a perpendicular plane to the lateral face of the postorbital, such that it does not slope downwards as in distinct horncores (Fig. 4A). The anterior margin of the right ornamentation slopes medially less than the left ornamentation. When the entire dorsal skull roof is viewed anteriorly, the surface between the postorbitals is near flat, with the apices of the ornamentation being barely perceptible, and instead appearing as slightly dorsally inflected dorsolateral “corners” of the postorbitals. While the skull of MOR 456 8-8-87-1 is transversely compressed, this compression appears to have only affected the frontoparietal fontanelle by closing it; the solid masses of bone that form the dorsal roof of the postorbitals do not appear to be affected by this compression and are non-deformed (Fig. 4A). The postorbitals have not been rotated or repositioned in such a way that would artificially produce the appearance of the flattened medial surfaces of the right and left supraorbital ornamentation.

Fig. 3. Right lateral views of supraorbital ornamentation of Einiosaurus procurvicornis Sampson, 1995, MOR 456 8-8-87-1 (A) and eucentrosauran (Einiosaurus procurvicornis or Achelousaurus horneri Sampson, 1995), MOR 591 (B) from the Campanian Two Medicine Formation, Montana, USA.

Fig. 4. Anterior views of supraorbital ornamentation of Einiosaurus procurvicornis Sampson, 1995, MOR 456 8-8-87-1 (A) and eucentrosauran (Einiosaurus procurvicornis or Achelousaurus horneri Sampson, 1995), MOR 591 (B), from the Campanian Two Medicine Formation, Montana, USA.
In contrast to the modified supraorbital ornamentation of MOR 456 8-8-87-1, MOR 591 expresses unmodified, plesiomorphic eucentrosauran supraorbital horncores typical of immature individuals (Figs. 3B, 4B). MOR 591 preserves a complete right postorbital and associated supraorbital horncore, while the left postorbital is missing approximately half of the dorsal aspect, including the apex, only preserving the anteriormost portion of the horncore. In lateral view, the right supraorbital horncore is triangular in profile, with a sharper apex than in MOR 456 8-8-87-1. Like MOR 456 8-8-87-1, the lateral face of the right supraorbital horncore is near flat, and is punctuated by several shallow vessel indentations. In anterior view, the right supraorbital horncore rises steeply dorsally, such that the medial surface of the horncore is near-vertical, and is transversely narrow and blade-like (Fig. 4B). The left supraorbital horncore preserves neither the lateral surface nor the apex, but the small preserved portion of the medial surface is likewise dorsoventrally steep. Whereas the medial surface of the MOR 591 supraorbital horncores slope towards the skull midline in a steep, near-vertical manner, the medial surfaces of the MOR 456 8-8-87-1 supraorbital ornamentation project near-horizontally. Sampson (1995) notes that MOR 591 bears shallow ridges on the medial surface of the right supraorbital horncore; this appears to be textural as opposed to incipient diagnostic ridges, and resembles the rugose surface textures which cover most of the dorsal aspect of the MOR 591 cranium.
Parietosquamosal frill.—Neither MOR 456 8-8-87-1 nor MOR 591 preserve a complete half of the diagnostic parietal, though MOR 456 8-8-87-1 preserves a complete left squamosal and MOR 591 preserves a near-complete left squamosal (Fig. 1). MOR 456 8-8-87-1 preserves the anteriormost portion of the left lateral parietal bar along the length of the parietosquamosal contact, though it does not preserve any of the anterior crenulation processes which line the lateral margins of the parietal. The left squamosal of MOR 456 8-8-87-1 is complete and is notably the same size as the squamosals of Einiosaurus procurvicornis holotype MOR 456 8-9-6-1, which is an overall larger adult individual (Fig. 8). The MOR 456 8-8-87-1 left squamosal measures 31 cm from the anterior edge of the anteriormost squamosal process to the parietosquamosal contact at the lateral margin, and the adult Einiosaurus holotype MOR 456 8-9-6-1 measures 29 cm. Measured from the dorsalmost extent of the otic notch to the EPS, MOR 456 8-8-87-1 is 33 cm and MOR 456 8-9-6-1 is 31.5 cm. Overall the morphology of the preserved squamosal does not differ from those of other eucentrosaurans.
Four of the five complete squamosals preserved in the stratigraphically lower Einiosaurus bonebed (TM-023) bear four marginal processes plus an EPS structure, with one bearing three processes, whereas the stratigraphically higher Einiosaurus bonebed preserves seven complete squamosals, five of which bear three marginal crenulations and an EPS, while two bear four processes and EPS structures, including MOR 456 8-8-87-1. All of these squamosals are of adult size and bear well-developed surface textures, suggesting that the specimens from each bonebed which deviate from the typical number of marginal crenulations represent individual variation. The apex of process S4 is somewhat damaged, though suggestive of a typical short, pointed process at the anterior corner of the lateral squamosal margin. In contrast to the typical low, rounded, D-shaped profile of eucentrosauran squamosal crenulations, the locus of S3 is rugose and dorsoventrally thickened compared to the squamosal margin between crenulations, but does not project outward from the margin, such that the overall lateral profile of the squamosal gives the appearance of lacking this locus entirely. Processes S2 and S1 are typical, low, rounded crenulations, though the apex of S1 is damaged. The posterolateral margin of the squamosal gently curls dorsally, such that S1 and the EPS at the parietosquamosal contact are both upturned dorsally. As is typical for ceratopsids, an EPS crenulation spans the parietosquamosal contact, with approximately two thirds of the process occurring along the squamosal margin and one third on the parietal. The posteriormost extent of the preserved portion of the parietal appears to bear the locus of P7, though this process is damaged and mostly missing.
The left squamosal of MOR 591 is mostly complete, missing the anterior corner, including S3/S4 (Fig. 1B). Like MOR 456 8-8-87-1, the squamosal of MOR 591 is of adult size. The measurement from the anterior edge of the anteriormost crenulation to the EPS is not possible to assess for MOR 591 because it is missing the anteriormost crenulation, but it can be measured from the dorsalmost extent of the otic notch to the EPS, which is 34.5 cm in MOR 591. An adult sized Achelousaurus horneri squamosal (MOR 485 7-12-87-4) from the holotype Achelousaurus horneri locality measures 31.3 cm from the dorsalmost extent of the otic notch to the EPS. All complete squamosals of Achelousaurus horneri bear three marginal processes plus EPS structures; MOR 591 preserves two squamosal processes plus an EPS. S1 and S2 are typical low, D-shaped processes, though the apex of S1 is damaged. The EPS structure is formed from a small, bulbous, dorsoventrally thickened mass of bone on both the squamosal and left lateral parietal bar on either side of the parietosquamosal contact. Posterior to the EPS on the left lateral parietal bar are the loci of two damaged marginal processes, both of which are imbricated and appear to be low, D-shaped, and unmodified, and therefore likely correspond to P7 and P6 (Wilson et al. 2020). Posterior to the two damaged processes is a low, non-imbricated, D-shaped process with an anteroposteriorly wider base, which would correspond to P5 if the anteriormost two processes are P7 and P6. The posteriormost preserved process, which likely corresponds to P4, is 11.8 cm wide at its base and approximately 11.9 cm long (this homology contrasts that of McDonald 2011 but is consistent with the homology of all known Two Medicine Formation eucentrosaurans, Wilson et al. 2020). The apex of this process is pitted and divided by grooves into multiple bulbous masses, which may correspond to resorption of the spike-like process. The adult holotype of Achelousaurus horneri MOR 485 likewise bears a pitted apex of the P4 process. Conversely, if MOR 591 instead represents a subadult individual of Einiosaurus procurvicornis, this more elongate P4 condition relative to P5–P7 is consistent with the morphology present in the lower Einiosaurus bonebed (Dino Ridge Quarry, TM-023) in which P4 is a short, partially elongate process (Wilson et al. 2020).
Epijugal.—MOR 456 8-8-87-1 preserves the complete left jugal and MOR 591 preserves a complete right and partial left jugal (Fig. 5). The jugals of both individuals are identical in size and do not differ in morphology from each other or from other eucentrosaurans. Notably, however, MOR 456 8-8-87-1 preserves the left epijugal in articulation with the jugal (Fig. 5A). In lateral view, the epijugal is semi-circular, with a rounded posterior edge. The posterior edge of the epijugal projects medially at about a 90° angle to the lateral face of the epijugal, such that the posterior edge is box-like. The lateral surface of the epijugal is roughly parallel to the lateral surface of the jugal and does not face anteriorly, as in some ceratopsids. The rounded ventral margin of the epijugal is contoured identically to the rounded ventral extent of the jugal beneath it. The suture between the epijugal and jugal occurs at an oblique, sub-vertical angle. Interestingly, no other specimens of Einiosaurus thus far reported preserve epijugals. Likewise, MOR 591 does not preserve either epijugal, and none of the additional two individuals of Achelousaurus preserve jugal material. The bone surfaces of the MOR 591 jugals are subtly rugose and undamaged, indicative of unfused epijugal loci, in the locations corresponding to the epijugal attachment site in MOR 456 8-8-87-1. The presence of an articulated epijugal in one subadult individual (MOR 456 8-8-87-1), but the lack of articulated (or fused) epijugals in another subadult individual (MOR 591) and all larger adult individuals may indicate that epijugal fusion did not occur until very late in ontogeny (or perhaps never), or may simply be preservational or taphonomic in nature.

Fig. 5. Lateral views of jugals of Einiosaurus procurvicornis Sampson, 1995, MOR 456 8-8-87-1 (A) and eucentrosauran (Einiosaurus procurvicornis or Achelousaurus horneri Sampson, 1995), MOR 591 (B, mirrored), from the Campanian Two Medicine Formation, Montana, USA. Arrow indicates epijugal.
Ontogenetic series of Einiosaurus procurvicornis supraorbital ornamentation.—The bonebed (TM-046) which produced both MOR 456 8-8-87-1 and Einiosaurus procurvicornis holotype MOR 456 8-9-6-1 preserved an additional six postorbitals and supraorbital complexes (postorbitals in articulation with palpebrals, lacrimals, and frontals, or some combination of those elements) with ornamentation ranging from unmodified horncores, to ornamentation identical to that of MOR 456 8-8-87-1, to masses similar to the ornamentation of holotype MOR 456 8-9-6-1 (Figs. 6, 7). These specimens help to fill the gap in Einiosaurus procurvicornis ontogenetic development between the unmodified plesiomorphic condition and the rounded supraorbital mass of the adult holotype which characterizes the taxon (see Discussion).
The smallest and most gracile postorbital from TM-046, MOR 456 8-10-87-20, is near-complete, preserving all marginal sutures, and missing the majority of the supraorbital horncore (Figs. 6A, 7A). Its bone surfaces medial to the horncore, adjacent to the frontal suture, and posterodorsal to the orbit, are punctuated by numerous vessel canal impressions, though the bone is visibly striated and long-grained. Despite the horncore being damaged, its morphology can be inferred from its preserved base. The base of the horncore is anteroposteriorly elongate, measuring 6.8 cm from its anterior extent at the palpebral suture to its posterior margin, and transversely narrow, being 1.7 cm at its widest point (halfway along its anteroposterior length). The base of the horncore projects dorsolaterally, such that the entire horncore likewise would have had a considerable dorsolateral orientation. While it is not possible to know its dorsoventral height or to what extent the apex would be rounded or pointed, its combined length and width make it narrow and blade-like, consistent with the plesiomorphic condition of eucentrosauran supraorbital horncores which are sub-triangular in lateral profile and bear pointed apices (e.g., as in Spinops, Centrosaurus, Styracosaurus, and Stellasaurus). Therefore, because MOR 456 8-10-87-20 expresses an unmodified plesiomorphic horncore we consider it a juvenile specimen.
The next largest postorbital from TM-046, MOR 456 8-9-7-3, is complete and likewise preserves all sutural margins (Figs. 6C, 7C). The vessel canals punctuating its lateral and dorsal surfaces are more extensively developed than those of the smallest postorbital, MOR 456 8-10-87-20, giving it an overall more rugose appearance, and the bone bears no striations. The supraorbital horncore is complete. In lateral view, the horncore is sub-circular and rounded, without a strongly triangular profile or pointed apex. In anterior view, the medial surface of the anterior ~1/3 of the horncore is minimally sloped towards the skull midline, whereas the posterior ~2/3 of the horncore project horizontally as a flat platform of bone towards the skull midline. In these regards, this specimen is overall similar to MOR 456 8-8-87-1. However, the supraorbital horncore apex of MOR 456 8-9-7-3 is still somewhat more bladed than those of MOR 456 8-8-87-1. Because it expresses some modification from the plesiomorphic condition, MOR 456 8-9-7-3 is considered an early subadult.

Fig. 6. Ontogenetic series of supraorbital ornamentation of Einiosaurus procurvicornis Sampson, 1995 from the Campanian Canyon Bonebed, Two Medicine Formation (TM-046), Montana, USA. A, B. Juvenile, MOR 456 8-10-87-20 (A) and MOR 456 8-8-87-19 (B). C, D. Early subadult, MOR 456 8-9-7-3 (C) and MOR 456 2020-C-1 (D). E, F. Late subadult, MOR 456 8-8-87-1 (E) and MOR 456 8-23-87 (F). G, H. Young adult, MOR 456 2020-C-2 (G) and MOR 456 8-9-6-1 (H). B and D are mirrored. All specimens in anterior view, lateral is to the right in A–D, G, H; E, entire skull width; F, lateral is to the left.

Fig. 7. Ontogenetic series of supraorbital ornamentation of Einiosaurus procurvicornis Sampson, 1995 from the Campanian Canyon Bonebed (TM-046), Montana, USA. A, B. Juvenile, MOR 456 8-10-87-20 (A) and MOR 456 8-8-87-19 (B). C, D. Early subadult, MOR 456 8-9-7-3 (C) and MOR 456 2020-C-1 (D). E, F. Late subadult, MOR 456 8-8-87-1 (E) and MOR 456 8-23-87 (F). G, H. Young adult, MOR 456 2020-C-2 (G) and MOR 456 8-9-6-1 (H). B, D–G are mirrored. All specimens in lateral view, anterior is to the left in all images.
MOR 456 8-8-87-19 is a partial postorbital with a complete supraorbital horncore which appears to be approximately the same size and robusticity as MOR 456 8-9-7-3, with similarly rugose surface textures, though because it is damaged their sizes cannot be compared directly (Figs. 6B, 7B). In lateral view, the horncore is likewise short and rounded with a sub-circular profile, and in anterior view the horncore rises more vertically, with a more sharply sloping medial surface which does not project horizontally towards the skull midline. Overall, its ornamental morphology is more similar to the inferred horncore of MOR 456 8-10-87-20, and it is therefore considered a juvenile.
MOR 456 2020-C-1 is an additional partial postorbital with a complete supraorbital horncore which appears to match MOR 456 8-9-7-3 in size and robusticity (Figs. 6D, 7D). Like MOR 456 8-9-7-3, its supraorbital horncore is low, rounded, and sub-circular in lateral view, and in anterior view the medial surface of the horncore minimally slopes towards the skull midline before projecting horizontally, making it an early subadult.
MOR 456 8-23-87 is a supraorbital complex, preserving the complete right postorbital and associated supraorbital ornamentation, palpebral, and partial frontal in articulation, which is near identical to MOR 456 8-8-87-1 in size, degree of rugosity, and morphology (Figs. 6F, 7F). The bone surface texture of MOR 456 8-23-87 is highly rugose, with numerous narrow ridges and deep grooves radiating transversely across the dorsal and posterodorsal aspect of the postorbital, including the supraorbital ornamentation. The medial aspect of the postorbital and the articulated frontal have been partially crushed downward in the regions corresponding to the thinner bone of the skull roof above the sinuses. Like MOR 456 8-8-87-1 and the majority of the other postorbitals described here, the supraorbital ornamentation in lateral view is low, somewhat rounded, and lacks a triangular profile or distinctly pointed apex. The deep grooves which penetrate the dorsal surface of the ornamentation terminate at the lateral edge of the ornamentation and disrupt the rounded profile of its dorsolateral edge. Due to the taphonomic crushing, the ornamentation has the appearance of sloping downwards towards the skull midline, though even when this crushing is not accounted for this sloping is shallow. If the medial aspect of the postorbital was not crushed, it likely would have projected horizontally as a flat platform of bone, as in MOR 456 8-8-87-1. Visible in dorsolateral and anterior view, the dorsal surface of the ornamentation is subtly convex and appears inflated, with convex posterior and medial margins which slope downwards. In this regard, this postorbital is near-identical to and replicates the morphology of MOR 456 8-8-87-1, expressing a subtle dorsal inflation of the more medial aspect of the ornamentation which contributes to an incipient, Einiosaurus procurvicornis holotype-like morphology, and we consider it a late subadult like MOR 456 8-8-87-1 (Table 2).
Table 2. Ontogenetic development and ornamental expression of Einiosaurus procurvicornis and Achelousaurus horneri growth stages. Ornamental expression is based on articulated skulls of subadults Einiosaurus procurvicornis MOR 456 8-8-87-1 and eucentrosauran MOR 591, and on articulated skulls of adults Einiosaurus procurvicornis MOR 456 8-9-6-1 and Achelousaurus horneri MOR 485, as well as disarticulated postorbitals from the TM-046 Einiosaurus bonebed.
|
Growth stage |
Ornamental condition |
|
Juvenile |
plesiomorphic morphology, unmodified erect nasal and supraorbital horncores |
|
Subadult |
early subadult: may express a combination of plesiomorphic and modified features; plesiomorphic and unmodified to slightly medially inflated supraorbital ornamentation; plesiomorphic, unmodified erect nasal horncore; “adult-sized” frill |
|
late subadult: may express a combination of plesiomorphic and modified features; medially inflated and transversely flattened supraorbital ornamentation; plesiomorphic, unmodified erect nasal horn core; “adult-sized” frill |
|
|
Adult |
expresses diagnostic condition of the taxon; procurved nasal horncore or nasal boss; spheroid, rounded supraorbital mass or supraorbital boss |
MOR 456 2020-C-2 is a partial right postorbital which preserves the majority of the supraorbital ornamentation (Figs. 6G, 7G). The surface texture of this specimen is well-developed and is not striated or long grained, but does not express the same degree of rugosity as MOR 456 8-8-87-1 and MOR 456 8-23-87, with rugose texture instead primarily occurring at the anterior extent of the preserved postorbital and supraorbital ornamentation. The supraorbital ornamentation of MOR 456 2020-C-2 is a low, dorsally-inflated spheroid mass of bone with convex edges. In lateral view, it lacks the acute subcircular profile of the subadult supraorbital ornamentation, and is more anteroposteriorly broad. It is most closely similar to the morphology of adult Einiosaurus procurvicornis holotype MOR 456 8-9-6-1, in which the supraorbital ornamentation is a low, convex, spheroid mass of bone dorsal to the orbit. Additionally, the supraorbital mass of holotype MOR 456 8-9-6-1 is near-identical in anteroposterior length (10.3 cm maximum) and transverse width (9.4 cm maximum) to MOR 456 2020-C-2 (9.8 cm maximum anteroposterior length; 9.7 cm transverse width), though the marginal edges of holotype MOR 456 8-9-6-1 are taller and it is overall more dorsally inflated than MOR 456 2020-C-2. We consider MOR 456 2020-C-2 an adult, like adult holotype MOR 456 8-9-6-1.
Discussion
Complex supraorbital ontogeny in Einiosaurus.—There are multiple supraorbital ornamentation morphologies preserved within the TM-046 bonebed, which may reflect ontogenetic change, individual variation, or both. Due to some specimens being incomplete, it is difficult to establish the absolute size of specimens relative to one another, though size alone is not a reliable indicator of maturity (e.g., Griffin and Nesbitt 2016). However, plesiomorphic morphologies are only expressed in small and more gracile specimens, while morphologies approaching the diagnostic morphology of the holotype occur in larger and more robust specimens. This is consistent with the hypothesis that the differences expressed in these specimens primarily represent ontogenetic change. Certain specimens likewise appear intermediate in development between the small, plesiomorphic specimens and the larger holotype bearing the diagnostic mature ornamentation. Because these additional postorbitals express varying degrees of more plesiomorphic ornamentation relative to the adult holotype, it is reasonable that they represent less mature individuals experiencing ontogenetic change. Because ontogenetic change cannot yet be ruled out as the cause for the differing morphologies, the hypothesis that the multiple morphologies of supraorbital ornamentation represent individual variation cannot be favored. If the multiple morphologies preserved in the TM-046 bonebed represent random individual variation without ontogenetic change, that would imply wide polymorphism of the supraorbital ornamentation, with the morphology of the holotype being one of multiple morphologies and therefore not diagnostic. The hypothesis that these specimens represent ontogenetic change could be falsified by discovery of large specimens (i.e., at the full size of the adult holotype) bearing distinct, erect horncores, or small specimens (i.e., at the size of MOR 456 8-10-87-20, the smallest Einiosaurus postorbital) bearing rounded supraorbital masses similar to that of the larger holotype, though evidence of this falsifying nature currently does not exist.
The ornamental morphologies seen in MOR 456 8-8-87-1 (erect, short nasal horncore; transversely flattened platform of bone as supraorbital ornamentation), which bears rugose surface texture, are seen in multiple other specimens from the TM-046 Einiosaurus bonebed (e.g., MOR 456 8-23-87-4 short, erect nasal horncore; MOR 456 8-23-87 flattened supraorbital platform). However, these additional specimens of similar or identical morphology do not express the same degree of bone surface rugosity as in MOR 456 8-8-87-1, perhaps indicating a degree of variability in textural expression in MOR 456 8-8-87-1 or an unknown physiological cause which may warrant more detailed examination. Brown et al. (2020) similarly note relatively advanced surface texture in a subadult specimen of Styracosaurus albertensis. We do not consider the texture alone indicative of MOR 456 8-8-87-1 being a mature individual of a different taxon. Further, Sampson (1995) and Sampson et al. (1997) did not recognize the presence of multiple centrosaurine taxa within the two bonebeds of Einiosaurus procurvicornis, and noted that specimens such as these express ornamental morphologies characteristic of immature individuals. Given that MOR 456 8-8-87-1 lacks the diagnostic parietal ornamentation, and considering that the hypothesis of an ontogenetic relationship between the specimens described here remains unfalsified, it is not taxonomically conservative to consider MOR 456 8-8-87-1 a different taxon from Einiosaurus procurvicornis. The supraorbital ornamentation of MOR 456 8-8-87-1 is additionally not consistent with that of any basal eucentrosaurans, but is consistent with being an intermediate step in the inflation of the spherical mass of bone which characterizes the larger Einiosaurus procurvicornis holotype specimen MOR 456 8-9-6-1. As stated above, in the absence of larger adults in the same bonebed bearing erect nasal horncores and erect, pointed supraorbital horncores, or small immature individuals expressing small procurved nasal horncores or small, spherical supraorbital masses of bone, the presence of multiple centrosaurine taxa within the TM-046 bonebed is not supported. In the absence of any diagnostic adult material of a second centrosaurine taxon from the same locality, it is most taxonomically conservative to consider the small, partially plesiomorphic skull of MOR 456 8-8-87-1 and all other similar such disarticulated specimens from the same bonebed as representing juvenile and subadult Einiosaurus procurvicornis.
Therefore, we hypothesize that the stratigraphically higher bonebed of Einiosaurus procurvicornis (TM-046) preserves a comprehensive ontogenetic series of supraorbital ornamentation, including the adult holotype specimen (MOR 456 8-9-6-1), articulated individuals such as the subadult skull described here (MOR 456 8-8-87-1), and the disarticulated postorbitals (Figs. 6, 7). Likewise, the possible subadult Achelousaurus horneri (MOR 591), together with the isolated subadult MOR 571 which bears incipient nasal and supraorbital bosses and long-grained striated texture on portions of the parietal, and the adult holotype (MOR 485) compose a basic ontogenetic series.
Based on the Einiosaurus procurvicornis ontogenetic series, we can infer that Einiosaurus procurvicornis experienced relatively complex ontogenetic development of the supraorbital ornamentation among ceratopsids (Figs. 6, 7). Hypothesized juvenile and early subadult individuals (e.g., MOR 456 8-10-87-20, MOR 456 8-8-87-19) of Einiosaurus procurvicornis possess plesiomorphic, unmodified supraorbital horncores with steep lateral and medial surfaces, which begin to flatten medially in early subadults, which transition to transversely flattened platforms of bone in late subadults such as MOR 456 8-8-87-1 and MOR 456 8-23-87. This flat platform of bone appears to inflate dorsally into the diagnostic convex, spherical masses expressed by the holotype MOR 456 8-9-6-1 and MOR 456 2020-C-2. MOR 456 8-8-87-1 fills an ontogenetic gap in supraorbital ornamentation development between plesiomorphic, erect horncores and the spherical mass of bone expressed by the holotype MOR 456 8-9-6-1. The specific nature of this remodeling can therefore be characterized as dorsal inflation of the medial aspect of the postorbital to form a transversely flat platform of bone. This inflation continued until the supraorbital ornamentation became a thick, convex, spheroid mass of bone (MOR 456 8-9-6-1).
Among other centrosaurines, with the exception of Pachyrhinosaurus (Fiorillo and Tykoski 2013) and Achelousaurus (though the current ontogenetic series for Achelousaurus is less comprehensive), Einiosaurus expresses relatively more complex ontogenetic development of the supraorbital ornamentation, and currently the earliest occurrence of this supraorbital ontogenetic change within Eucentrosaura (Wilson et al. 2020). Einiosaurus is the earliest occurring eucentrosauran centrosaurine to extend ontogenetic development of its supraorbital ornamentation beyond a simple, erect, pyramidal supraorbital horncore, which is the basal condition of the clade. Whereas Coronosaurus does express supraorbital horncores with laterally deflected apices, its horncores fundamentally remain horncores through growth (Ryan and Russell 2005). Conversely, the ontogenetic remodeling of Einiosaurus supraorbital horncores produces rounded, spherical masses of bone in place of erect horncores with discernable apices, fundamentally differing from other, more basal eucentrosaurans. As the first eucentrosauran to extend the development of the supraorbital ornamentation beyond a plesiomorphic horncore, Einiosaurus may represent a shift in the particular characteristics of the socio-sexual selection acting upon this group (Wilson et al. 2020).
Based on the growth series described here, Einiosaurus procurvicornis likewise appears to have had one of the most complex documented ontogenetic trajectories of supraorbital ornamentation of any ceratopsid. While the number of changes seen in this growth series may seem surprising, there is plentiful evidence that ceratopsids were capable of extensive ontogenetic remodeling of their cranial ornamentation. The eucentrosauran Pachyrhinosaurus, hypothesized as the direct descendant of Einiosaurus and Achelousaurus (Horner et al. 1992; Wilson et al. 2020) likely represents the most complex ontogenetic development of the supraorbital ornamentation of known centrosaurines. In Pachyrhinosaurus lakustai, the smallest and presumed youngest specimens bear supraorbital horncores which are rounded and semi-circular in lateral view with a shallow slope medially, similar to the ontogenetic series described here for Einiosaurus, inflating medially and becoming large and complex bosses (Currie et al. 2008). The most common and well-documented ceratopsid, Triceratops, expresses reorientation of its postorbital horncores through ontogeny, from recurved horncores in juveniles to anteriorly curved in mature individuals (Horner and Goodwin 2006). Other chasmosaurines, such as Chasmosaurus and Regaliceratops, underwent ontogenetic resorption of their postorbital horncores (Brown and Henderson 2015; Campbell et al. 2016). Among the basal eucentrosaurans closely related to Einiosaurus, such as Centrosaurus and Styracosaurus, there appears to be less dramatic development of the supraorbital ornamentation through ontogeny, with the majority of change consisting of increase in size and sometimes resorption of the horncore apices (Ryan et al. 2001; Frederickson and Tumarkin-Deratzian 2014; Brown et al. 2020). Given that mature individuals of Centrosaurus and Styracosaurus typically express simple, erect horncores, similar to those of immature Einiosaurus and Achelousaurus (nasal horncore procurvature is known in some adults of Centrosaurus; Frederickson and Tumarkin-Deratzian 2014; Brown et al. 2020, though to a lesser degree than in Einiosaurus), we would perhaps not expect to see the same degree of change through ontogeny in those more basal taxa.
Sequence of ornamentation ontogenetic change.—Although the sample size of articulated subadult eucentrosauran skulls from the Two Medicine Formation is currently limited to two, we can tentatively infer the order of ontogenetic development between the nasal and supraorbital ornamentation. In both MOR 456 8-8-87-1 and MOR 591, the nasal ornamentation is an erect, recurved, and unmodified horncore, consistent with the plesiomorphic condition of Eucentrosaura. However, whereas the supraorbital horncores of MOR 591 are likewise erect, triangular in profile, unmodified, and plesiomorphic, the supraorbital horncores of MOR 456 8-8-87-1 appear to be ontogenetically modified from the plesiomorphic condition expressed in juvenile postorbitals from the same bonebed, e.g., MOR 456 8-10-87-20 and are instead transversely flat and dorsally inflated. This condition is likewise expressed to a lesser degree in MOR 456 2020-C-1, and to an approximately equivalent degree in the near-identical MOR 456 8-23-87, indicating that this morphology is a genuine step in the ontogenetic development of Einiosaurus procurvicornis supraorbital ornamentation. While the timing of ornamentation development is likely variable individually and through stratigraphic time (characteristic of heterochronic trends; see Brown et al. 2020), the available specimens currently indicate that onset of supraorbital ornamentation modification preceded nasal horncore modification in the uppermost Two Medicine Formation eucentrosaurans, whether within Einiosaurus alone or between Einiosaurus and Achelousaurus, depending on the identity of MOR 591. This tentative hypothesis can be tested by the recovery of additional subadult individuals of these taxa. Likewise, the hypothesis that the timing of the developmental changes noted here was affected by evolutionary change through the time separating the TM-046 Einiosaurus procurvicornis bonebed from the stratigraphically higher Achelousaurus horneri can be further tested with additional articulated, immature specimens.
Facial versus parietosquamosal frill development.—The face of subadult Einiosaurus MOR 456 8-8-87-1, measured from the anteriormost preserved extent of the anterior nasal processes to the posterior margin of the orbit, is 42.5 cm on the left side of the face, though there is minor damage to the anterior margin of the anterior nasal processes and some taphonomic folding anterior to the orbit. Conversely, the right side of the face measures 46.5 cm with the same compression pulling the posterior margin of the orbit more posterior than its in vivo position (Table 1). Therefore, MOR 456 8-8-87-1 has an estimated facial length of 44.5 cm. In comparison, the facial length of MOR 591 is identical to that estimated for MOR 456 8-8-87-1, at 44.5 cm. The faces of both MOR 456 8-8-87-1 and MOR 591 are therefore approximately 81.6% as long as the 54.5 cm face of Einiosaurus procurvicornis holotype MOR 456 8-9-6-1. Interestingly, however, the complete left squamosal of MOR 456 8-8-87-1 and the partial left squamosal of MOR 591 are of approximately equivalent size to that of the adult MOR 456 8-9-6-1. The squamosal of MOR 456 8-8-87-1 measures 31 cm from the anterior edge of the anteriormost squamosal process to the parietosquamosal contact at the lateral margin, while MOR 456 8-9-6-1 measures 29 cm. The squamosal of MOR 485, the holotype of Achelousaurus horneri, measures 31 cm (Table 1). This measurement is not possible to assess for MOR 591 because it is missing the anterior corner of the lateral squamosal margin, though it is possible to measure from the dorsalmost extent of the otic notch to the EPS in MOR 456 8-8-87-1 (33 cm), MOR 591 (34.5 cm), and MOR 456 8-9-6-1 (31.5 cm). This measurement is consistent with the anteriormost marginal process to EPS measurement, with both being slightly longer in MOR 456 8-8-87-1 than in MOR 456 8-9-6-1. Likewise, the squamosal of MOR 591 is slightly longer than MOR 456 8-9-6-1 for the single comparable measurement between these two individuals (from the dorsalmost extent of the otic notch to the EPS). Digital overlay of the squamosals indicates that MOR 456 8-9-6-1 may express slightly more anteroposterior elongation of the squamosal, while MOR 456 8-8-87-1 is somewhat more dorsally extended, in line with the plane of the lateral frill margins (Fig. 8). Unfortunately, none of the additional parietal material from the TM-046 bonebed can be unambiguously assigned to MOR 456 8-8-87-1. While the frills of MOR 456 8-8-87-1 and MOR 456 8-9-6-1 are approximately the same size, the face of MOR 456 8-8-87-1 would require additional growth to reach adult size, suggesting relatively later facial ornamentation development and expansion of the facial region compared to the parietosquamosal frill reaching adult size. Future discoveries of articulated or associated fossils of individuals representing earlier growth stages may reveal if frill growth was similarly allometric or isometric. Other centrosaurines, such as Centrosaurus apertus, appear to attain adult sized or near-adult sized frills relatively early in ontogeny (Frederickson and Tumarkin-Deratzian 2014). Conversely, the order of some developmental events, such as squamosal elongation versus postorbital horn elongation, may be the opposite in other ceratopsid (albeit chasmosaurine) taxa, such as Chasmosaurus in which elongation of squamosals appears to occur after postorbital horn elongation (Mallon et al. 2015). In Arrhinoceratops squamosal elongation occurs while postorbital horncores are recurved, the immature condition (Mallon et al. 2015). The particular relationship (order and timing) between facial versus parietosquamosal frill development is likely dependent upon the particular selection scheme affecting the frill versus facial ornamentation in individual taxa and/or lineages, and it may be expected that uneven selection pressure upon certain ornamental structures likely affects the timing and sequence of ornamentation development (Wilson et al. 2020).
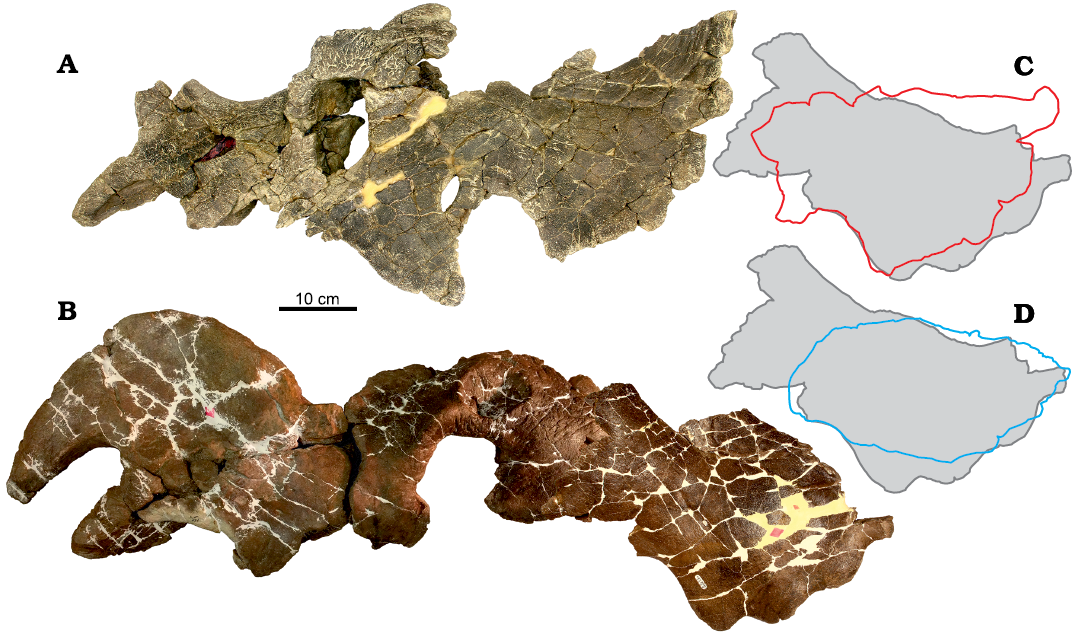
Fig. 8. Size comparison of squamosal versus face in Einiosaurus procurvicornis Sampson, 1995, subadult MOR 456 8-8-87-1 (A) and MOR 456 8-9-6-1, holotype (B, mirrored); squamosal superimposed in B. C. Outline of MOR 456 8-8-87-1 (red) superimposed over outline of MOR 456 8-9-6-1 (grey). D. Outline of MOR 591 (blue) superimposed over outline of MOR 456 8-9-6-1 (grey). Outlines aligned by otic notch.
Among ceratopsids, the parietosquamosal frill is generally considered the most diagnostic portion of the skull, with an array of disparate morphologies expressed between unique taxa (Dodson et al. 2004). Einiosaurus and Achelousaurus are closely related to other eucentrosaurans expressing relatively conservative facial ornamentation (e.g., Spinops, Coronosaurus, Centrosaurus, and Styracosaurus; Ryan and Russell 2005; Ryan et al. 2007; Farke et al. 2011; Chiba et al. 2017). Evolutionary facial change between these taxa is primarily limited to simplification of the supraorbital horncores and elongation of the nasal horncore through time. Conversely, the parietal ornamentation of these taxa differs considerably, from the node clusters which characterize Coronosaurus, to the large, hook-like P1 and P2 processes of Centrosaurus and Spinops, to the hypertrophied P3, P4, and P5 processes of Styracosaurus (Ryan and Russell 2005; Ryan et al. 2007; Farke et al. 2011; Brown et al. 2020). Given that the parietosquamosal frill is generally considered an ornamental structure which functioned in a signaling capacity (e.g., Padian and Horner 2011; Knell and Sampson 2011), and that the ornamental disparity between taxa within Eucentrosaura appears to be greatest within the parietal, it can be hypothesized that a parietal-dominated selection pattern characterized the basal portion of Eucentrosaura (Wilson et al. 2020). In later eucentrosaurans (e.g., Einiosaurus, Achelousaurus, Pachyrhinosaurus) increased complexity of the facial ornamentation relative to the parietosquamosal frill suggests facial-dominated signaling selection (Wilson et al. 2020).
If parietal-dominated selection did characterize Eucentrosaura basally, the signaling capacity of the frill would be maximized by initiating its growth and development earliest among the cranial ornamentation. It is hypothesized that subadult individuals would express more extensive development of the frill towards the adult condition relative to the facial ornamentation. Unfortunately, the articulated skull of subadult MOR 456 8-8-87-1 does not preserve the diagnostic posterior bar and ornamentation of the parietal, but the adult-sized preserved squamosal of this immature individual is consistent with this hypothesis, with a fully-developed and adult-sized squamosal contrasting its plesiomorphic morphology of the nasal horncore and partially-modified supraorbital ornamentation. Likewise, MOR 591 bears an adult-sized squamosal while still expressing unmodified, plesiomorphic nasal and supraorbital horncore morphologies. Einiosaurus and Achelousaurus immediately succeed in time the more basal eucentrosaurans with marked parietal disparity, and their ontogenetic development may maintain some developmental trends seen earlier in Eucentrosaura. If the subtle differences in development between MOR 456 8-8-87-1 and MOR 591 represent genuine trends in the sequence of ornamentation development in Einiosaurus (and possibly Achelousaurus), then parietosquamosal frill growth likely preceded both nasal and supraorbital modification. Likewise, because MOR 591 appears to be relatively less mature than MOR 456 8-8-87-1 based on surface texture and ornamentation development but is equivalent in size, this may indicate that Achelousaurus reached adult size more rapidly if MOR 591 represents Achelousaurus, though more specimens of immature individuals are needed to test this. Supraorbital horncore development appears to have occurred after the parietosquamosal frill reached adult size, but prior to nasal horncore modification. Articulated subadult individuals such as MOR 591 and MOR 456 8-8-87-1 are critical to elucidating trends in the timing and differential development of cranial ornamentation and informing hypotheses regarding the paleobiological significance of such trends and their role in the evolution of ornamented dinosaurs.
Conclusions
This study describes an articulated subadult cranium of the eucentrosauran centrosaurine dinosaur Einiosaurus procurvicornis from the Late Cretaceous Two Medicine Formation of Montana, and compares its ontogenetic development with a similarly sized, articulated subadult eucentrosauran cranium from the same formation. As the only known articulated subadult crania of Two Medicine Formation eucentrosaurans, these specimens allow tentative inference of the order of ontogenetic development between the nasal, supraorbital, and parietosquamosal frill ornamentation; the parietosquamosal frill appears to reach adult size first, and the supraorbital ornamentation appears to begin ontogenetic development before the nasal horncore is modified. The supraorbital ornamentation of the newly described Einiosaurus procurvicornis cranium appears to fill a previous gap in the ontogenetic development from the plesiomorphic short horncore morphology of immature individuals to the rounded, spheroid ornamentation of adult individuals. The ontogenetic change in supraorbital ornamentation seen in Einiosaurus procurvicornis appears to be among the most complex supraorbital ontogenetic change within Ceratopsidae.
Acknowledgments
The bonebed which produced the subadult Einiosaurus procurvicornis skull described here, as well as the holotype specimen, was discovered by Carrie Ancell (Museum of the Rockies, Bozeman, USA) and Jack Horner (Chapman University, Orange, USA), both of whom likewise discovered MOR 591. Carrie Ancell masterfully prepared these specimens. We are indebted to them, as well as all those who assisted in the excavation of these sites, especially Bob Harmon and Patrick Leiggi (both formerly Museum of the Rockies, Bozeman, USA), and the late Bob Makela. The fieldwork was funded by a grant from the National Science Foundation to Jack Horner. We thank the Blackfeet Nation and Gloria Sundquist who graciously permitted the Museum of the Rockies land access and allowed the collection of specimens. We thank Denver Fowler (Badlands Dinosaur Museum, Dickinson, USA), Jacob Gardner, Chris Organ, Holley Flora, Dave Varricchio, Willie Freimuth, and Giulio Panasci (all Montana State University, Bozeman, USA), Jack Horner (Chapman University, Orange, USA), David Evans (Royal Ontario Museum; University of Toronto, Toronto, Canada), Michael Ryan (Carleton University, Ottawa, Canada), and Julie Reizner (Northern Kentucky University, Highland Heights, USA) for helpful discussion. Thanks to MOR and the Department of Earth Sciences at Montana State University, Bozeman, USA, for support. JPW thanks Jack Horner for permission to study these specimens and Amy Atwater (MOR) for facilitating access to MOR collections. Thanks to Andy Farke (Raymond M. Alf Museum, Claremont, USA) and Joseph Frederickson (Weis Earth Science Museum, Menasha, USA) for very helpful and constructive reviews which improved this manuscript.
References
Bailleul, A.M., Scannella, J.B., Horner, J.R., and Evans, D.C. 2016. Fusion patterns in the skulls of modern archosaurs reveal that sutures are ambiguous maturity indicators for the Dinosauria. PLOS ONE 11 (2): e0147687. Crossref
Brown, C.M. and Henderson, D.M. 2015. A new horned dinosaur reveals convergent evolution in cranial ornamentation in Ceratopsidae. Current Biology 25: 1641–1648. Crossref
Brown, C.M., Holmes, R., and Currie, P. 2020. A subadult individual of Styracosaurus albertensis (Ornithischia: Ceratopsidae) with comments on ontogeny and intraspecific variation in Styracosaurus and Centrosaurus. Vertebrate Anatomy Morphology Palaeontology 8: 67–95. Crossref
Brown, C.M., Russell, A.P., and Ryan, M.J. 2009. Pattern and transition of surficial bone texture of the centrosaurine frill and their ontogenetic and taxonomic implications. Journal of Vertebrate Paleontology 29: 132–141. Crossref
Campbell, J.A., Ryan, M.J., Holmes, R.B., and Schröder-Adams, C.J. 2016. A re-evaluation of the chasmosaurine ceratopsid genus Chasmosaurus (Dinosauria: Ornithischia) from the Upper Cretaceous (Campanian) Dinosaur Park Formation of Western Canada. PLoS ONE 11 (1): e0145805. Crossref
Chiba, K., Ryan, M.J., Fanti, F., Loewen, M.A., and Evans, D.C. 2017. New material and systematic re-evaluation of Medusaceratops lokii (Dinosauria, Ceratopsidae) from the Judith River Formation (Campanian, Montana). Journal of Paleontology 92: 272–288. Crossref
Currie, P.J., Holmes, R.B., Ryan, M.J., and Coy, C. 2016. A juvenile chasmosaurine ceratopsid (Dinosauria, Ornithischia) from the Dinosaur Park Formation, Alberta, Canada. Journal of Vertebrate Paleontology 36 (2): e1048348. Crossref
Currie, P.J., Langston Jr. W., and Tanke, D.H. 2008. A new species of Pachyrhinosaurus (Dinosauria, Ceratopsidae) from the Upper Cretaceous of Alberta, Canada. In: P.J. Currie and W. Langston Jr. (eds.), A New Horned Dinosaur from an Upper Cretaceous Bone Bed in Alberta, 1–108. NRC Research Press, Ottawa. Crossref
Dodson, P., Forster, C.A., and Sampson, S.D. 2004. Ceratopsidae. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria, 494–513. University of California Press, Berkeley. Crossref
Farke, A.A., Ryan, M.J., Barrett, P.M., Tanke, D.H., Braman, D.R., Loewen, M.A., and Graham, M.R. 2011. A new centrosaurine from the Late Cretaceous of Alberta, Canada, and the evolution of parietal ornamentation in horned dinosaurs. Acta Palaeontologica Polonica 56: 691–702. Crossref
Fiorillo, A.R. and Tykoski, R.S. 2013. An immature Pachyrhinosaurus perotorum (Dinosauria: Ceratopsidae) nasal reveals unexpected complexity of craniofacial ontogeny and integument in Pachyrhinosaurus. PLOS ONE 8 (6): e65802. Crossref
Frederickson, J.A. and Tumarkin-Deratzian, A.R. 2014. Craniofacial ontogeny in Centrosaurus apertus. PeerJ 2: e252. Crossref
Griffin, C.T. and Nesbitt, S.J. 2016. Anomalously high variation in postnatal development is ancestral for dinosaurs but lost in birds. Proceedings of the National Academy of Sciences 113: 14757–14762. Crossref
Hone, D.W.E., Farke, A.A., and Wedel, M.J. 2016. Ontogeny and the fossil record: what, if anything, is an adult dinosaur? Biology Letters 12 (2): 20150947. Crossref
Horner, J.R. and Goodwin, M.B. 2006. Major cranial changes during Triceratops ontogeny. Proceedings of the Royal Society B: Biological Sciences 273 (1602): 2757–2761. Crossref
Horner, J.R. and Goodwin, M.B. 2008. Ontogeny of cranial epi-ossifications in Triceratops. Journal of Vertebrate Paleontology 28 (1): 134–144. Crossref
Horner, J.R., Goodwin, M.B., and Myhrvold, N. 2011. Dinosaur census reveals abundant Tyrannosaurus and rare ontogenetic stages in the Upper Cretaceous Hell Creek Formation (Maastrichtian), Montana, USA. PloS one 6 (2): e16574. Crossref
Horner, J.R., Schmitt, J.G., Jackson, F., and Hanna, R. 2001. Bones and rocks of the Upper Cretaceous Two Medicine-Judith River clastic wedge complex, Montana. In: C.L. Hill (ed.), Society of Vertebrate Paleontology 61st Annual Meeting: Mesozoic and Cenozoic Paleontology in the Western Plains and Rocky Mountains. Field Trip Guidebook. Museum of the Rockies Occasional Paper 3: 3–14.
Horner, J.R., Varricchio, D.J., and Goodwin, M.B. 1992. Marine transgressions and the evolution of Cretaceous dinosaurs. Nature 358: 59–61. Crossref
Kirkland, J.I. and DeBlieux, D.D. 2010. New basal centrosaurine ceratopsian skulls from the Wahweap Formation (Middle Campanian), Grand Staircase-Escalante National Monument, southern Utah. In: M.J. Ryan, B.J. Chinnery-Allgeier, and D.A. Eberth (eds.), New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium, 117–140. Indiana University Press, Bloomington.
Knell, R.J. and Sampson, S. 2011. Bizarre structures in dinosaurs: species recognition or sexual selection? A response to Padian and Horner. Journal of Zoology 283: 18–22. Crossref
Mallon, J.C., Ryan, M.J., and Campbell, J.A. 2015. Skull ontogeny in Arrhinoceratops brachyops (Ornithischia: Ceratopsidae) and other horned dinosaurs. Zoological Journal of the Linnean Society 175: 910–929. Crossref
McDonald, A.T. 2011. A subadult specimen of Rubeosaurus ovatus (Dinosauria: Ceratopsidae), with observations on other ceratopsids from the Two Medicine Formation. PLoS ONE 6 (8): e22710. Crossref
Padian, K. and Horner, J.R. 2011. The evolution of “bizarre structures” in dinosaurs: biomechanics, sexual selection, social selection or species recognition? Journal of Zoology 283: 3–17. Crossref
Reizner, J.A. 2010. An Ontogenetic Series and Population Histology of the Ceratopsid Dinosaur Einiosaurus procurvicornis. 97 pp. Ph.D. Dissertation, Montana State University-Bozeman, College of Letters & Science, Bozeman.
Reizner, J.A., Woodward, H., and Freedman Fowler, E.A. 2020. Histologic growth dynamic study and population biology of the ceratopsid dinosaur Einiosaurus procurvicornis from a death assemblage of the Upper Cretaceous Two Medicine Formation of Northwestern Montana. In: Journal of Vertebrate Paleontology, SVP Program and Abstracts Book, 279–280. Society of Vertebrate Paleontology.
Rogers, R.R. 1990. Taphonomy of three dinosaur bone beds in the Upper Cretaceous Two Medicine Formation of Northwestern Montana: evidence for drought-related mortality. Palaios 5: 394–413. Crossref
Rueden, C.T., Schindelin, J., Hiner, M.C., DeZonia, B.E., Walter, A.E., Arena, E.T., and Eliceiri, K.W. 2017. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinformatics 18: 529. Crossref
Ryan, M.J. and Evans, D.C. 2005. Ornithischian dinosaurs. In: P.J. Currie and E.B. Koppelhus (eds.), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed, 312–348. Indiana University Press, Bloomington.
Ryan, M.J. and Russell, A.P. 2005. A new centrosaurine ceratopsid from the Oldman Formation of Alberta and its implications for centrosaurine taxonomy and systematics. Canadian Journal of Earth Sciences 42: 1369–1387. Crossref
Ryan, M.J., Holmes, R., and Russell, A.P. 2007. A revision of the Late Campanian centrosaurine ceratopsid genus Styracosaurus from the Western Interior of North America. Journal of Vertebrate Paleontology 27: 944–962. Crossref
Ryan, M.J., Russell, A.P., Eberth, D.A., and Currie, P.J. 2001. The taphonomy of a Centrosaurus (Ornithischia: Certopsidae) Bone Bed from the Dinosaur Park Formation (Upper Campanian), Alberta, Canada, with comments on cranial ontogeny. Palaios 16: 482–506. Crossref
Sampson, S.D. 1995. Two new horned dinosaurs from the Upper Cretaceous Two Medicine Formation of Montana; with a phylogenetic analysis of the Centrosaurinae (Ornithischia: Ceratopsidae). Journal of Vertebrate Paleontology 15: 743–760. Crossref
Sampson, S.D., Ryan, M.J., and Tanke, D.H. 1997. Craniofacial ontogeny in centrosaurine dinosaurs (Ornithischia: Ceratopsidae): taxonomic and behavioral implications. Zoological Journal of the Linnean Society 121: 293–337. Crossref
Scannella, J.B. and Fowler, D.W. 2014. A stratigraphic survey of Triceratops localities in the Hell Creek Formation, northeastern Montana (2006–2010). Geological Society of America Special Papers 503: 313–332. Crossref
Scannella, J.B. and Horner, J.R. 2010. Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae): synonymy through ontogeny. Journal of Vertebrate Paleontology 30: 1157–1168. Crossref
Scannella, J.B., Fowler, D.W., Goodwin, M.B., and Horner, J.R. 2014. Evolutionary trends in Triceratops from the Hell Creek Formation, Montana. Proceedings of the National Academy of Sciences 111: 10245–10250. Crossref
Tumarkin-Deratzian, A.R. 2010. Histologic evaluation of ontogenetic bone surface texture changes in the frill of Centrosaurus apertus. In: M.J. Ryan, B.J. Chinnery-Allgeier, and D.A. Eberth (eds.), New Perspectives on Horned Dinosaurs. The Royal Tyrrell Museum Ceratopsian Symposium, 251–263. Indiana University Press, Bloomington.
Wilson, J.P., Ryan, M.J., and Evans, D.C. 2020. A new, transitional centrosaurine ceratopsid from the Upper Cretaceous Two Medicine Formation of Montana and the evolution of the “Styracosaurus-line” dinosaurs. Royal Society Open Science 7 (4): 1–25. Crossref
Acta Palaeontol. Pol. 66 (4): 797–814, 2021
https://doi.org/10.4202/app.00797.2020