
The first possible remingtonocetid stem whale from North America
MARK D. UHEN and CARLOS MAURICIO PEREDO
Uhen, M.D. and Peredo, C.M. 2021. The first possible remingtonocetid stem whale from North America. Acta Palaeontologica Polonica 66 (1): 77–83.
Remingtonocetid cetaceans are a group of stem whales known from the Indo-Pakistan and North African Tethys Ocean. An unusual tooth was discovered by Peter J. Harmatuk in 1973 in the middle Eocene Superior Stone Quarry (now the Martin Marietta Quarry) near Castle Hayne, North Carolina, USA. Here we identify this tooth as a premolar of a possible member of the Remingtonocetidae, which would extend the range of this family across the Atlantic to eastern North America. This partial tooth includes most of the crown (missing the mesial end) and the posterior root. The tooth bears a single central cusp and a worn accessory cusp on the posterior end. This tooth most closely resembles premolars of Remingtonocetus and is rather dissimilar to premolars of other archaeocetes known from the middle Eocene of North America, such as the families Protocetidae and Basilosauridae. This new record potentially expands the geographic distribution of the amphibious cetacean family Remingtonocetidae across the Atlantic.
Key words: Mammalia, Archaeocete, Remingtonocetidae, middle Eocene, Castle Hayne Formation, North Carolina, USA.
Mark D. Uhen [muhen@gmu.edu], George Mason University, Department of Atmospheric, Oceanic, and Earth Sciences, MSN 6E2, Fairfax, VA 22030, USA.
Carlos M. Peredo [cmperedo@umich.edu], Univeristiy of Michigan, Department of Earth and Environmental Science, Ann Arbor, MI 48109, USA; Texas A&M University Galveston, Department of Marine Biology Galveston, TX 77553, USA.
Received 30 August 2020, accepted 22 September 2020, available online 27 January 2021.
Copyright © 2021 M.D. Uhen and C.M. Peredo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Remingtonocetidae are a family of stem cetaceans with extremely long snouts and proportionally long mandibles and mandibular symphyses (Kumar and Sahni 1986). This family was originally described and known only from Indo-Pakistan (Kumar and Sahni 1986; Gingerich et al. 1995; Thewissen and Hussain 2000; Bajpai and Thewissen 2000) until remingtonocetid remains were also reported from North Africa (Bebej et al. 2015). Here we report a single tooth from the middle Eocene Castle Hayne Formation of North Carolina that differs markedly in morphology from the protocetids and basilosaurids previously described from the middle Eocene of North America. The Castle Hayne Formation has produced Crenatocetus rayi (McLeod and Barnes 1990, 1996, 2008); Pachycetus wardii (Uhen 1999, 2001; Gol’din and Zvonok 2013; van Vliet et al. 2020); Cynthiacetus maxwelli (Uhen 2005); and various specimens of indeterminate basilosaurids (Uhen 2013). However, the morphology of this new specimen does not match the features found in any of these taxa, nor any closely related taxa. Instead, it more closely resembles a remingtonocetid, despite the family being known only from the Tethys Ocean.
The biogeographic distribution of stem cetaceans has been largely linked to their perceived locomotor capabilities. Pakicetids, ambulocetids, and remingtonocetids have been found only within the Tethys Ocean and have been perceived as more terrestrial, whereas the more aquatic protocetids are now known to have dispersed to the Americas (e.g., Hulbert et al. 1998; Uhen 1998; Geisler et al. 2005; Uhen 2014; Lambert et al. 2019a) and the fully aquatic basilosaurids are cosmopolitan (e.g., Köhler and Fordyce 1997; Uhen and Tichy 2000; Martínez-Cáceres and de Muizon 2011; Uhen et al. 2011; Uhen 2013, 2018). The tooth described here is provisionally identified as the first known remingtonocetid from the North America. This extends the range of remingtonocetids not only out of Indo-Pakistan, but out of the Tethys and across the Atlantic, thus challenging perceived limitations of remingtonocetid locomotion and habitat range.
Institutional abbreviations.—IITR-SB, Indian Institute of Technology, Roorkee, Department of Geosciences, Uttaranchel, India; KPG-M, Université de Montpellier, Institut des Sciences de l’Évolution, Montpellier, France; NHML, Natural History Museum, London, UK; USNM, US National Museum of Natural History, Washington, D.C., USA.
Other abbreviations.—dP/p, deciduous premolar upper/lower; EFA, elliptic Fourier analysis; PCA, principal component analysis.
Material and methods
Whale teeth, particularly those of heterodont archaeocetes, can vary widely in form even within an individual. Traditional measures for analyzing shape, such as linear measurements or geometric morphometrics, have proved difficult to apply to whale teeth. Linear measurements often fail to capture the complex forms of multi-cusped cheek teeth. Geometric morphometrics work better, but ultimately still rely on landmarks and therefore lose some of the complexity of morphology (Crampton 1995). Other studies have attempted to quantify shape complexity using orientation patch count (OPC) (Peredo et al. 2018), but this process measures overall complexity of the crown rather than differences in shape, and is not useful for testing the identity of the North Carolina tooth.
In contrast to linear measurements and geometric morphometrics, elliptic Fourier analysis (EFA) creates a complete outline of the shape in question and therefore represents a more accurate representation of the complex morphologies observed (Bonhomme et al. 2014 and references therein). Previous studies have used EFA to classify shape in other taxa such as plants (Neto et al. 2006), bivalves (Ferson et al. 1985), fish otoliths (Tracey et al. 2006), pinniped whiskers (Ginter et al. 2012), and shark teeth (Cullen and Marshall 2019). Here, we use EFA to compare the overall outline morphology of teeth from remingtonocetids, protocetids, and basilosaurids as a means of testing the taxonomic identity of USNM 449550.
Data collection.—We sampled 37 teeth spanning the breadth of potential taxa for the North Carolina tooth. Our sample includes nine remingtonocetid, 14 protocetid, and 13 basilosaurid teeth in addition to USNM 449550. Given that the tooth originally bore 2 roots, USNM 449550 is clearly a cheek tooth. Its morphology is inconsistent with a first premolar or a molar and given that it has no lingual expansion for any sort of protocone or protocone remnant, it likely represents an upper premolar, although it could also represent a lower premolar. Accordingly, we restricted our dataset to only second through fourth premolars, and our dataset does include both upper and lower teeth to make our analysis more conservative.
Photographs of the teeth in our dataset were either taken by the authors or were acquired from the literature (see Table 1 for specific sources). We cropped each photograph in Adobe Photoshop to include only the crown of the tooth to exclude potential biases introduced by varying degrees of preservation of the dental roots. Because EFA is sensitive to orientation, each tooth was rotated such that the occlusal surface was up and the anterior face was to the right (teeth were mirrored if necessary). USNM 449550 preserves only the central cusp and posterior end. Therefore, we conservatively estimated the anterior end based on other archaeocete premolars to create an acceptable reconstruction for this analysis.
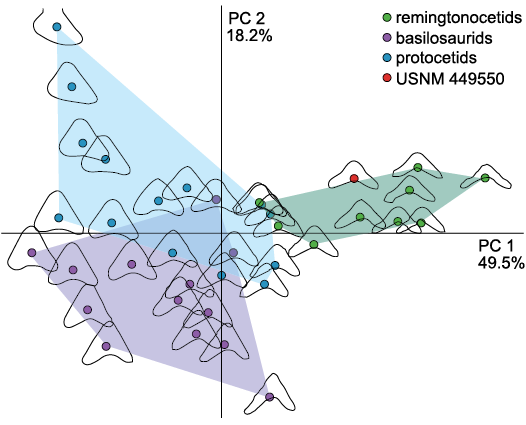
Fig. 1. PCA ordinations of differences in tooth morphology for sampled archaeocetes. This plot includes data from both adult and deciduous premolars. The numbers next to the axis labels indicate the percentage of explained variation in morphology for that axis. The tooth shapes around each point represent the outline generated using the harmonic coefficients produced by elliptic Fourier analysis to achieve 99.9% harmonic power. See SOM: table 1 for a list of included specimens.
EFA and PCA analyses.—We conducted an elliptic Fourier analysis (EFA), and subsequently a principal component analysis (PCA), using the R package Momocs (Bonhomme et al. 2014). Following the methods of Cullen and Marshall (2019), we centered and scaled the outlines of each tooth to remove the effect of tooth size. Then, each tooth was smoothed using a simple moving average with nine iterations to reduce noise from the preparation process. To capture as much morphological accuracy as possible, we chose the number of harmonics to describe 99.9% of the total variation in shape. We then evaluated the variation in tooth shapes using a principal component analysis, which facilitates the visualization of individual teeth within morphological space (Fig. 1). In doing so, we directly tested the taxonomic identity of the North Carolina tooth by plotting it in morphological space alongside the teeth with certain taxonomic identity.
Systematic palaeontology
Cetacea Brisson, 1762
?Remingtonocetidae Kumar and Sahni, 1986
?Remingtonocetidae indet.
Fig. 2.
Material.—USNM 449550, partial premolar (Fig. 2B) from middle Eocene, Martin Marietta Quarry (formerly Superior Stone Quarry), near Castle Hayne, North Carolina, USA (34°21’30” N, 77°52’ W, Paleobiology Database collection 5459).
Description.— USNM 449550 includes portion of a tooth, including the main central cusp, and the posterior portion of the crown (Fig. 2B). The posterior root is also present, and extends farther posterior than the posterior edge of the crown. The enamel of the crown displays vertical rugosities. There are small cingula on the medial, lateral, and posterior edges. There is also a small secondary cusp on the posterior heel of the tooth. The central cusp displays light apical wear, and the posterior cusp displays heavy apical wear to the point of being almost worn away. No lateral wear is evident on the tooth. The tooth is also very narrow mediolaterally, and long anteroposteriorly. The crown is 9.3 mm wide directly below the central cusp. The anteroposterior distance from the central cusp to the posterior margin, measured parallel to the cingulum is 27.5 mm.
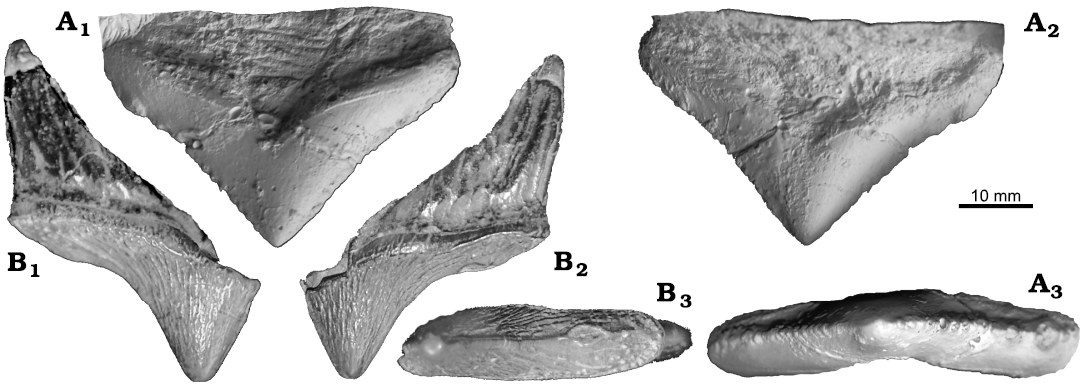
Fig. 2. A. Remingtonocetus sp. (IITR-SB 2630) from Lutetian; Kachchh, India; right P4. B. ?Remingtonocetidae indet. (USNM 449550) from Lutetian–Bartonian; Martin Marietta Quarry (formerly Superior Stone Quarry), near Castle Hayne, North Carolina, USA. Shown as if it were an upper premolar for comparison, but it may also represent a lower premolar as well. Note the extreme narrowness of the tooth. In lateral (A2, B2), medial (A1, B1), and occlusal (A3, B3) views.
Remarks.—Precise information on the horizon where the tooth was collected is lacking. However, Kier (1980) states that the rocks in the quarry are from the Castle Hayne Formation, and that they are Bartonian (late middle Eocene) in age. There is considerable debate in the literature about the age and age range of the Castle Hayne Formation. Some authors consider the Castle Hayne Formation to span from the middle to late Eocene (Harris and Laws 1997). Others restrict the Castle Hayne to the middle Eocene (Coffey and Read 2004) or even some smaller portion of the middle Eocene (Weems et al. 2004). This debate is in part due to different opinions on what is part of the Castle Hayne Formation versus an overlying formation, but is also due to differing opinions on the ages of the rocks themselves. Nevertheless, all parties agree that the rocks in the Superior Stone Quarry are of middle Eocene age, most likely from the Lutetian or early Bartonian.
Results
The only other cetaceans known from the Castle Hayne Formation are protocetid Crenatocetus rayi (McLeod and Barnes 1990, 1996, 2008); and basilosaurids Pachycetus wardii (Uhen 1999, 2001; Gol’din and Zvonok 2013; van Vliet et al. 2020); and Cynthiacetus maxwelli (Uhen 2005); and various specimens of indeterminate basilosaurids (Uhen 2013). Cheek teeth are not known from P. wardii, but alveoli demonstrate that in general the teeth of P. wardii would be much larger and transversely broader than this specimen. The premolars of the Crenatocetus (which is only known from a lower jaw with cheek teeth) are shorter (anteroposteriorly) and broader (mediolaterally) than this specimen. Basilosaurid premolars, both adult and deciduous, bear multiple accessory denticles (Uhen 2004), which USNM 449550 lacks. This specimen is also much too small to represent Cynthiacetus. Measurements of small to moderately sized remingtonocetids, protocetids, and basilosaurids are listed in SOM: table 2 (Supplementary Online Material available at app66-Uhen_Peredo_SOM.pdf).
USNM 449550 could possibly represent a premolar of a protocetid, rather than a remingtonocetid. Upper premolars of protocetids have lingual expansions of the posterior portion of the tooth, often bearing a protocone in more apomorphic forms, but this structure is lacking in USNM 449550, as is the case in remingtonocetids (Thewissen and Bajpai 2001). USNM 449550 might also represent a protocetid lower premolar, but it is very buccolingually narrow when compared to the lower premolars of known protocetids. Only remingtonocetid premolars and deciduous protocetid lower premolars are buccolingually as narrow as USNM 449550. See SOM: table 2.
USNM 449550 could also represent a deciduous premolar of a protocetid. One characteristic that is suggestive of it being a deciduous tooth is the obtuse angle of the root to the cingulum. Adult archaeocete teeth usually have roots that angle in towards the center of the tooth while deciduous teeth have roots that angle away from the center (Uhen and Gingerich 2001). The root of USNM 449550 is angled slightly away from the center, unlike either adult or deciduous teeth previously described. However, the root is also closed, as in adult teeth, and unlike deciduous teeth. The only known protocetid deciduous teeth are those of Pappocetus lugardi (Andrews 1920; McLeod and Barnes 2008), Indocetus ramani (Bajpai and Thewissen 2014), and Togocetus traversei (Gingerich and Cappetta 2014), as listed in Table 1.
Both known specimens of Pappocetus lugardi (NHML M11414, M11086) dentaries are in the process of erupting their permanent molars (Andrews 1920), which indicates that teeth in the premolar positions are still deciduous teeth (Uhen 2004). The first lower deciduous premolar (dp1) is missing in both specimens, but was clearly a single rooted tooth based on the preserved alveolus in NHML M 11414. The second deciduous lower premolar is present in NHML M 11414, has 2 roots, but the crown is broken. From what is preserved it is likely to have had a single, central cusp, but that cannot be confirmed. Both dp3 and dp4 clearly bear at least two if not more accessory denticles, like those of basilosaurid deciduous and adult teeth, and unlike the permanent premolars of protocetids.
IITR-SB 2986 and IITR-SB 2000-33, Indocetus ramani, both preserve portions of dp3 and complete dp4. Both are similar to the morphology of Pappocetus in the presence of accessory cusps (Bajpai and Thewissen 2014).
Table 1. All known protocetid specimens that include deciduous premolars, with descriptions of those specimens.
|
Specimen |
Taxon |
Teeth |
Notes |
|
NHML M11414 |
Pappocetus lugardi |
dp2–dp4 |
both dp3 and dp4 have some
accessory denticles, but dp2 appears to lack them; |
|
USNM 542448 |
Protocetidae indet. |
dp3? |
missing 2 roots, no accessory denticles; possibly a dp2 based on lack of denticles |
|
IITR-SB 2986 |
Indocetus ramani |
dp3–dp4 |
dp3 is broken, dp4 has accessory denticles |
|
IITR-SB 2000-33 |
Indocetus ramani |
dp3–dp4 |
dp3 is complete, but not described, dp4 has accessory denticles; not figured |
|
KPG-M 10 |
Togocetus traversei |
dP4 |
2 roots, 2 cusps |
|
KPG-M 133 |
Togocetus traversei |
dP3 |
2 roots, 2 cusps |
|
KPG-M 23 |
Togocetus traversei |
dp4 |
3 cusps, 1 main, 1 anterior, 1 posterior |
|
KPG-M 16 |
Togocetus traversei |
dp2 |
2 roots, no extra cusps, but anteroposteriorly short |
|
KPG-M 15 |
Togocetus traversei |
dp3 |
2 roots, 1 main cusp missing posterior portion of crown and posterior root |
Both upper and lower deciduous premolars are known from several isolated teeth of Togocetus traversei (see Table 1). Crowns of dP3 and dP4 have two roots, and two cusps each. They differ in that dP3 lacks any kind of a protocone, but dP4 has a lingual swelling over the posterior root indicating a protocone remnant (Gingerich and Cappetta 2014). Isolated lower deciduous premolars from dp2 to dp4 have also been described. All have two roots. dp2 clearly bears a single cusp and lacks accessory denticles. dp3 lacks anterior accessory denticles, but is broken posteriorly so the presence or absence of accessory denticles cannot be determined. dp4 bears three cusps, a large central cusp and one smaller anterior accessory denticle and one larger posterior accessory denticle, with the possibility of another smaller one absent due to breakage (Gingerich and Cappetta 2014).
USNM 449550 bears a tiny, worn denticle on the posterior carina of the crown. This morphology makes it a poor match for any of the adult protocetid premolars and for the premolars of basilosaurids, which bear numerous accessory denticles. We therefore consider it unlikely that USNM 449550 belongs to either a protocetid or a basilosaurid adult tooth. It could be a protocetid deciduous premolar, but it is double rooted, ruling out identity as a dp1, and given the lack of accessory denticles, it cannot be either an upper deciduous premolar, nor a dp3 or dp4. If USNM 449550 is from a protocetid, then it must be a dp2.
We compared USNM 449550 to the best remingtonocetid dentition known, that of Remingtonocetus harudiensis, IITR-SB 2630 (Thewissen and Bajpai 2001) (cast = USNM 533999, Fig. 2A1, A3). The tooth is most similar to the P4 of R. harudiensis. The crown of this tooth is 8.6 mm wide directly below the central cusp, and the length from the central cusp to the posterior margin is 29.8 mm, making it slightly narrower than USNM 449550 when compared to the length.
Finally, our shape analysis of the tooth compared to the cheek teeth of other archaeocetes places the shape of the tooth in a morphospace with other remingtonocetid premolars, separate from those of both protocetids and basilosaurids. This further confirms the identity of this tooth as a possible remingtonocetid cheek tooth, extending their range across the Atlantic Ocean to the east coast of North America.
Discussion
The probable identification of a specimen from North Carolina as a remingtonocetid, along with recent discovery of protocetid cetaceans in Peru (Lambert et al. 2019b) greatly alters previous notions of biogeography and locomotor abilities of early cetaceans. Previous distributions of families of archaic whales were such that Pakicetidae (Fig. 3A), Ambulocetidae (Fig. 3A), and Remingtonocetidae (Fig. 3B) were restricted to Indo-Pakistan; Protocetidae (Fig. 3C) were known from Indo-Pakistan, north and west Africa, and eastern North America; while Basilosauridae (Fig. 3D) were globally distributed. Thus, Pakicetidae, Ambulocetidae, and Remingtonocetidae were restricted to the Tethys Ocean, while Protocetidae expanded into the Atlantic and Basilosauridae were globally distributed.
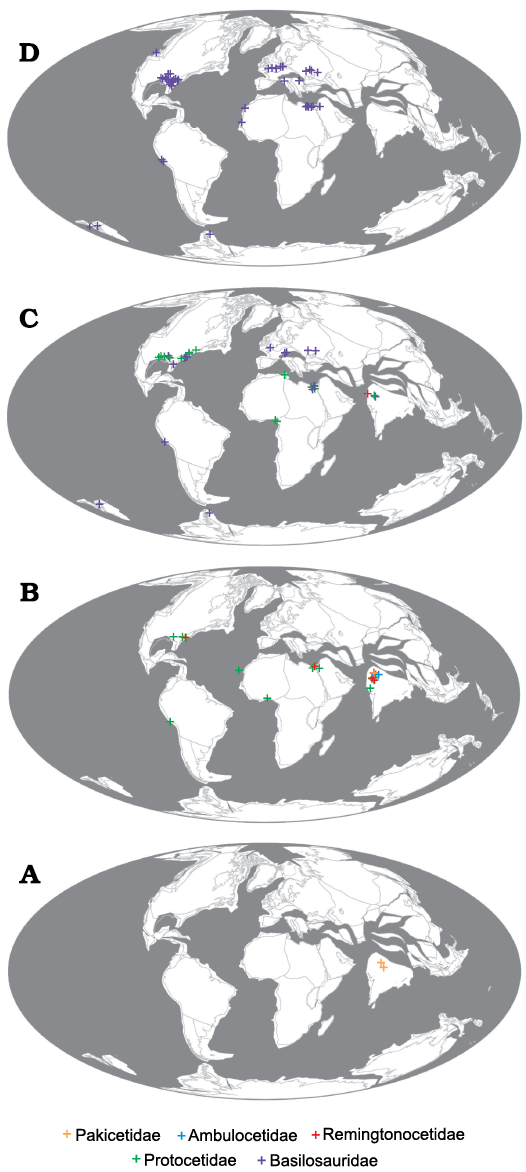
Fig. 3. Paleogeographic reconstructions and distributions of Eocene cetaceans. A. Ypresian, reconstructed at 52 Ma. B. Lutetian, reconstructed at 45 Ma. C. Bartonian, reconstructed at 40 Ma. D. Priabonian reconstructed at 36 Ma. Data are derived from occurrences in the Paleobiology Database and includes every published occurrence of archaeocete cetaceans (Uhen 2020).
This pattern of distribution fit well with hypotheses regarding locomotor capabilities. Pakicetids were quite terrestrial (Madar 2007), while ambulocetids and remingtonocetids are thought to be convergent on crocodile morphology and ecology (Thewissen et al. 1996; Bajpai and Thewissen 2000). Neither family appeared to have ranged far from the Indo-Pakistan region of the Tethys. Protocetids, which were more semi-aquatic like pinnipeds (Gingerich 2003), ranged into and across the Atlantic to North America following prevailing currents at the time (Uhen 1999), while basilosaurids, which were fully aquatic, could disperse across all of the oceans to be found on virtually all continents (Uhen 1998).
However, recent discoveries reveal this narrative is inaccurate. Bebej et al. (2015) reported a remingtonocetid from North Africa, demonstrating that they left Indo-Pakistan into the broader Tethys. Lambert et al. (2019a, b) reported a protocetid from Peru, indicating that protocetids did not just cross the Atlantic but also dispersed into the Pacific. Now, the potential discovery of a remingtonocetid from North America extends a third family of archaeocetes across the Atlantic and suggests that the aquatic locomotory abilities of remingtonocetids may have been better developed than previously thought.
A lack of previously discovered remingtonocetids from North America is unsurprising given that the record of protocetids is also sparse. Only 14 published fossil collections include protocetids from the Americas, with 13 from North America (Uhen 2014), one from South America (Lambert et al. 2019a), and several with very limited material. Indeed, only four of these North American collections are of Lutetian age, and all other known remingtonocetids from the Afro-Eurasia are Lutetian (Uhen 2020). These protocetid occurrences only represent five species in five genera, the first of which were only named in 1998 (Hulbert et al. 1998; Uhen 1998) although fragmentary protocetids had been noted from North America by Kellogg (1936). Thus, lack of previous discoveries of remingtonocetids in North America is likely a problem of poor sampling. New discovery of protocetids from Peru (Lambert et al. 2019a) indicates that protocetids ranged much farther than previously thought, and perhaps, one should not be surprised by a North American remingtonocetid.
Conclusions
The first phase of cetacean evolution is, for the most part, the story of adaptation to the aquatic environment (Pyenson 2017). Changes in feeding, sensory systems, and locomotor systems are apparent in all of the lineages of middle Eocene cetaceans (Uhen 2007). Remingtonocetids and protocetids had been previously known to co-occur in Indo-Pakistan, but the presence of protocetids in North America without remingtonocetids, suggested that the swimming abilities of remingtonocetids were perhaps somewhat more restricted than those of protocetids. The discovery of a potential remingtonocetid in North America calls that interpretation into question. Also, the discovery of protocetids in Peru suggests that protocetids were also strong swimmers, able to cross the Atlantic between Africa and the Americas. Continued discovery of middle Eocene cetaceans in North America, and the southern hemisphere will further test the hypothesized swimming abilities of these early cetaceans.
Acknowledgements
We would like to thank Hans Thewissen (Northeast Ohio Medical University, Rootstown, USA) for access to and permission to use the casts of Remingtonocetus harudiensis. We would also like to thank David J. Bohaska (National Museum of Natural History, Smithsonian Institution, Washington, DC, USA) for assistance with the specimens. We also thank Joshua A. Cullen (University of Florida, Gainesville, USA) for access to the R code used for this analysis. Finally, we thank an anonymous reviewer and Ryan Bebej (Calvin University, Grand Rapids, USA) for helpful comments on earlier versions of this work.
References
Andrews, C.W. 1920. A description of new species of zeuglodont and of leathery turtle from the Eocene of southern Nigeria. Proceedings of the Zoological Society of London 1919: 309–319. Crossref
Bajpai, S. and Thewissen, J.G.M. 2000. A new, diminutive Eocene whale from Kachchh (Gujarat, India) and its implications for locomotor evolution of cetaceans. Current Science 79: 1478–1482.
Bajpai, S. and Thewissen, J.G.M. 2014. Protocetid cetaceans (Mammalia) from the Eocene of India. Palaeontologia Electronica 17 (3-32A): 1–19. Crossref
Bebej, R.M., Zalmout, I., El-Aziz, A.A.A., Antar, M.S.M., and Gingerich, P.D. 2015. First remingtonocetid archaeocete (Mammalia, Cetacea) from the middle Eocene of Egypt with implications for biogeography and locomotion in early cetacean evolution. Journal of Paleontology 89: 882–893. Crossref
Bonhomme, V., Picq, S., Claude, J., and Gaucherel, C. 2014. Momocs: Outline Analysis Using R. Journal of Statistical Software 56 (13): 1–24. Crossref
Coffey, B.P. and Read, J.F. 2004. Integrated sequence stratigraphy of Paleogene outcrop and subsurface strata of the North Carolina Coastal Plain, Southeastern U.S.A. Southeastern Geology 42: 253–278.
Crampton, J.S. 1995. Elliptic Fourier shape analysis of fossil bivalves: Some practical considerations. Lethaia 28: 179–186. Crossref
Cullen, J.A. and Marshall, C.D. 2019. Do sharks exhibit heterodonty by tooth position and over ontogeny? A comparison using elliptic Fourier analysis. Journal of Morphology 280: 687–700. Crossref
Ferson, S., Rohlf, F.J., and Koehn, R.K. 1985. Measuring shape variation of two-dimensional outlines. Systematic Biology 34: 59–68. Crossref
Geisler, J.H., Sanders, A.E., and Luo, Z.-X. 2005. A new protocetid whale (Cetacea: Archaeoceti) from the Late Middle Eocene of South Carolina. American Museum Novitates 3480: 1–65. Crossref
Gingerich, P.D. 2003. Land-to-sea transition in early whales: evolution of Eocene Archaeoceti (Cetacea) in relation to skeletal proportions and locomotion of living semiaquatic mammals. Paleobiology 29 (3): 429–454. Crossref
Gingerich, P.D. and Cappetta, H. 2014. A new archaeocete and other marine mammals (Cetacea and Sirenia) from Lower Middle Eocene phosphate deposits of Togo. Journal of Paleontology 88: 109–129. Crossref
Gingerich, P.D., Arif, M., and Clyde, W.C. 1995. New archaeocetes (Mammalia, Cetacea) from the Middle Eocene Domanda Formation of the Sulaiman Range, Punjab (Pakistan). Contributions from the Museum of Paleontology, The University of Michigan 29 (11): 291–330.
Ginter, C.C., DeWitt, T.J., Fish, F.E., and Marshall, C.D. 2012. Fused traditional and geometric morphometrics demonstrate pinniped whisker diversity. PLoS One 7 (4): e34481. Crossref
Gol’din, P. and Zvonok, E. 2013. Basilotritus uheni, a new cetacean (Cetacea, Basilosauridae) from the Late Middle Eocene of Eastern Europe. Journal of Paleontology 87: 254–268. Crossref
Harris, W.B. and Laws, R.A. 1997. Paleogene stratigraphy and sea-level history of the North Carolina Coastal Plain: Global coastal onlap and tectonics. Sedimentary Geology 108: 91–120. Crossref
Hulbert, R.C., Jr., Petkewich, R.M., Bishop, G.A., Bukry, D., and Aleshire, D.P. 1998. A new middle Eocene protocetid whale (Mammalia: Cetacea: Archaeoceti) and associated biota from Georgia. Journal of Paleontology 72: 907–927. Crossref
Kellogg, R. 1936. A Review of the Archaeoceti. Carnegie Institution of Washington Special Publication 482: 1–366.
Kier, P.M. 1980. The echinoids of the Middle Eocene Warley Hill Formation, Santee Limestone, and Castle Hayne Limestone of North and South Carolina. Smithsonian Contributions to Paleobiology 39: 1–102. Crossref
Köhler, R. and Fordyce, R.E. 1997. An archaeocete whale (Cetacea: Archaeoceti) from the Eocene Waihao Greensand, New Zealand. Journal of Vertebrate Paleontology 17: 574–583. Crossref
Kumar, K. and Sahni, A. 1986. Remingtonocetus harudiensis, new combination, a middle Eocene archaeocete (Mammalia, Cetacea) from western Kutch, India. Journal of Vertebrate Paleontology 6: 326–349. Crossref
Lambert, O., Bianucci, G., Salas-Gismondi, R., Di Celma, C., Steurbaut, E., Urbina, M., and de Muizon, C. 2019a. An amphibious whale from the Middle Eocene of Peru reveals early South Pacific dispersal of quadrupedal cetaceans. Current Biology 29: 1–8. Crossref
Lambert, O., Bianucci, G., Salas-Gismondi, R., Di Celma, C., Steurbaut, E., Urbina, M., and de Muizon, C. 2019b. A new protocetid from the middle Eocene of Peru provides insights on the colonization of the New World by African four-legged whales. In: T. Smith and A. Folie (eds.), International Symposium PalEurAfrica. Program and Abstracts, 24. Royal Belgian Institute of natural Sciences, Brussels.
Madar, S.I. 2007. The postcranial skeleton of early Eocene pakicetid cetaceans. Journal of Paleontology 81 (1): 176–200. Crossref
Martínez-Cáceres, M. and de Muizon, C. 2011. A new basilosaurid (Cetacea, Pelagiceti) from the late Eocene to early Oligocene Otuma Formation of Peru. Comptes Rendus Palevol 10: 517–526. Crossref
McLeod, S.A. and Barnes, L.G. 1990. Archaeocete cetaceans from the Atlantic Coastal Plain of the United States, including a new protocetid. Journal of Vertebrate Paleontology 9 (3): 35A.
McLeod, S.A. and Barnes, L.G. 1996. The systematic position of Pappocetus lugardi and a new taxon from North America (Archaeoceti: Protocetidae). The Paleontological Society Special Publication 8: 270. Crossref
McLeod, S.A. and Barnes, L.G. 2008. A new genus and species of Eocene protocetid archaeocete whale (Mammalia, Cetacea) from the Atlantic Coastal Plain. Contributions in Science, Natural History Museum of Los Angeles County 41: 73–98.
Neto, J.C., Meyer, G.E., Jones, D.D., and Samal, A.K. 2006. Plant species identification using elliptic Fourier leaf shape analysis. Computers and Electronics in Agriculture 50: 121–134. Crossref
Peredo, C.M., Peredo, J.S., and Pyenson, N.D. 2018. Convergence on dental simplification in the evolution of whales. Paleobiology 44: 1–10. Crossref
Pyenson, N.D. 2017. The ecological rise of whales chronicled by the fossil record. Current Biology 27: R558–R564. Crossref
Thewissen, J.G.M. and Bajpai, S. 2001. Dental morphology of Remingtonocetidae (Cetacea, Mammalia). Journal of Paleontology 75: 463–465. Crossref
Thewissen, J.G.M. and Hussain, S.T. 2000. Attockicetus praecursor, a new remingtonocetid cetacean from marine Eocene sediments of Pakistan. Journal of Mammalian Evolution 7: 133–146. Crossref
Thewissen, J.G.M., Madar, S.I., and Hussain, S.T. 1996. Ambulocetus natans, an Eocene cetacean (Mammalia) from Pakistan. Courier Forschungsinstitut Senckenberg 191: 1–86.
Tracey, S.R., Lyle, J.M., and Duhamel, G. 2006. Application of elliptical Fourier analysis of otolith form as a tool for stock identification. Fisheries Research 77: 138–147. Crossref
Uhen, M.D. 1998. New protocetid (Mammalia, Cetacea) from the late Middle Eocene Cook Mountain Formation of Louisiana. Journal of Vertebrate Paleontology 18: 664–668. Crossref
Uhen, M.D. 1999. New species of protocetid archaeocete whale, Eocetus wardii (Mammalia, Cetacea), from the middle Eocene of North Carolina. Journal of Paleontology 73: 512–528. Crossref
Uhen, M.D. 2001. New material of Eocetus wardii (Mammalia, Cetacea), from the middle Eocene of North Carolina. Southeastern Geology 40: 135–148.
Uhen, M.D. 2004. Form, function, and anatomy of Dorudon atrox (Mammalia, Cetacea): An archaeocete from the middle to late Eocene of Egypt. The University of Michigan Museum of Paleontology Papers on Paleontology 34: 1–222.
Uhen, M.D. 2005. A new genus and species of archaeocete whale from Mississippi. Southeastern Geology 43: 157–172.
Uhen, M.D. 2007. The terrestrial to aquatic transition in Cetacea. In: J.S. Anderson and H.-D. Sues (eds.), Major Transitions in Vertebrate Evolution, 392–408. Indiana University Press, Bloomington.
Uhen, M.D. 2013. A review of North American Basilosauridae. Alabama Museum of Natural History Bulletin 31 (2): 1–45.
Uhen, M.D. 2014. New specimens of Protocetidae (Mammalia, Cetacea) from New Jersey and South Carolina. Journal of Vertebrate Paleontology 34: 211–219. Crossref
Uhen, M.D. 2018. Basilosaurids and kekenodontids. In: B. Würsig, J.G.M. Thewissen, and K.M. Kovacs (eds.), Encyclopedia of Marine Mammals, 78–80. Academic Press, London. Crossref
Uhen, M.D. 2020. Cetacea. Paleobiology Database Data Archive 9. doi: 10.13021/as3j-2d21.
Uhen, M.D. and Gingerich, P.D. 2001. New genus of dorudontine archaeocete (Cetacea) from the middle-to-late Eocene of South Carolina. Marine Mammal Science 17: 1–34. Crossref
Uhen, M.D. and Tichy, G. 2000. A new basilosaurid archaeocete from Austria. Journal of Vertebrate Paleontology 20 (3, Supplement): 74A– 75A.
Uhen, M.D., Pyenson, N.D., DeVries, T.J., Urbina, M., and Renne, P.R. 2011. New Middle Eocene whales from the Pisco Basin of Peru. Journal of Paleontology 85 (5): 955–969. Crossref
van Vliet, H.J., Bosselaers, M., Vahldiek, B.-W., Paymans, T., and Verheijen, I. 2020. Eocene cetaceans from the Helmstedt region, Germany, with some remarks on Platyosphys, Basilotritus and Pachycetus. Cainozoic Research 20: 121–148.
Weems, R.E., Self-Trail, J.M., and Edwards, L.E. 2004. Supergroup stratigraphy of the Atlantic and Gulf Coastal Plains (Middle? Jurassic through Holocene, eastern North America). Southeastern Geology 42: 191–216.
Acta Palaeontol. Pol. 66 (1): 77–83, 2021
https://doi.org/10.4202/app.00799.2020