
Evidence of two lineages of metriorhynchid crocodylomorphs in the Lower Cretaceous of the Czech Republic
DANIEL MADZIA, SVEN SACHS, MARK T. YOUNG, ALEXANDER LUKENEDER, and PETR SKUPIEN
Madzia, D., Sachs, S., Young, M.T., Lukeneder, A., and Skupien, P. 2021. Evidence of two lineages of metriorhynchid crocodylomorphs in the Lower Cretaceous of the Czech Republic. Acta Palaeontologica Polonica 66 (2): 357–367.
Metriorhynchid crocodylomorphs were an important component in shallow marine ecosystems during the Middle Jurassic to Early Cretaceous in the European archipelago. While metriorhynchids are well known from western European countries, their central and eastern European record is poor and usually limited to isolated or fragmentary specimens which often hinders a precise taxonomic assignment. However, isolated elements such as tooth crowns, have been found to provide informative taxonomic identifications. Here we describe two isolated metriorhynchid tooth crowns from the upper Valanginian (Lower Cretaceous) of the Štramberk area, Czech Republic. Our assessment of the specimens, including multivariate analysis of dental measurements and surface enamel structures, indicates that the crowns belong to two distinct geosaurin taxa (Plesiosuchina? indet. and Torvoneustes? sp.) with different feeding adaptations. The specimens represent the first evidence of Metriorhynchidae from the Czech Republic and some of the youngest metriorhynchid specimens worldwide.
Key words: Crocodylomorpha, Thalattosuchia, Metriorhynchidae, Lower Cretaceous, Valanginian, Czech Republic.
Daniel Madzia [daniel.madzia@gmail.com, ORCID: https://orcid.org/0000-0003-1228-3573], Institute of Paleobiology, Polish Academy of Sciences, Twarda 51/55, 00-818 Warsaw, Poland.
Sven Sachs [Sachs.Pal@gmail.com], Naturkunde-Museum Bielefeld, Abteilung Geowissenschaften, Adenauerplatz 2, 33602 Bielefeld, Germany.
Mark T. Young [mark.young@ed.ac.uk, ORCID: https://orcid.org/0000-0002-7263-6505], School of GeoSciences, Grant Institute, University of Edinburgh, James Hutton Road, Edinburgh, EH9 3FE, UK.
Alexander Lukeneder [alexander.lukeneder@nhm-wien.ac.at], Natural History Museum Vienna, Geological and Palaeontological Department, Burgring 7, 1010 Vienna, Austria.
Petr Skupien [petr.skupien@vsb.cz], Department of Geological Engineering, VŠB – Technical University, 17. listopadu 15, 708 33 Ostrava Poruba, Czech Republic.
Received 13 August 2020, accepted 5 October 2020, available online 20 May 2021.
Copyright © 2021 D. Madzia et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Thalattosuchia was a species-rich and geographically widespread clade of marine and freshwater crocodylomorphs that were particularly abundant between the late Early Jurassic and the Late Jurassic when they regulated the trophic structure of near-shore as well as deep-water settings (e.g., Andrade et al. 2010; Young et al. 2013a; Wilberg 2015; Foffa et al. 2018c, 2019; Johnson et al. 2018; Martin et al. 2018). Thalattosuchian fossils are particularly abundant in western Europe, often preserving complete skeletons (e.g., Eudes-Deslongchamps 1867–1869; Fraas 1902; Andrews 1913). Central and eastern European strata have mostly yielded isolated or highly fragmentary specimens that are rarely able to give a precise taxonomic assignment (e.g., Ochev 1981; Hua et al. 1998; Čerňanský et al. 2019; Madzia et al. 2021). By the end of the Jurassic, thalattosuchian fossils become rarer in Europe, most likely as a consequence of localised sea level regressions. However, both major thalattosuchian lineages, the metriorhynchoids and teleosauroids, crossed the Jurassic–Cretaceous boundary (e.g., Sachs et al. 2020; Young and Sachs 2020). The Cretaceous teleosauroid fossil record is problematic, and currently based on a single unambiguous occurrence from the upper Barremian of Colombia (Cortés et al. 2019; see also Fanti et al. 2016 and discussion in Young and Sachs 2020). In contrast, several metriorhynchoid lineages are known from Cretaceous deposits. Sachs et al. (2020) listed 18 potential Cretaceous occurrences of Metriorhynchoidea, although the age of four of them (Ochev 1981; Herrera et al. 2015; Fernández et al. 2019) remains contentious and may be latest Tithonian (latest Jurassic) or earliest Berriasian (earliest Cretaceous) (see Sachs et al. 2020 for an overview and Discussion herein). Most of the Cretaceous occurrences of metriorhynchoid thalattosuchians originate from the Valanginian (see Sachs et al. 2020). Post-Valanginian metriorhynchoids are exceptionally rare; only two occurrences may be known from the lower Hauterivian (Debelmas 1952; Debelmas and Demains D’Archimaud 1956) and a single record originates from the lowermost Aptian (Chiarenza et al. 2015).
Here we report two new occurrences of Lower Cretaceous metriorhynchoid thalattosuchians, each represented by an isolated tooth crown belonging to two lineages of geosaurin metriorhynchids. Both tooth crowns originate from the upper Valanginian (Lower Cretaceous) of the Kopřivnice Formation in Štramberk, Czech Republic (Fig. 1). Therefore, they are among the youngest representatives of Metriorhynchidae (and Thalattosuchia) worldwide. We describe both crowns, compare their surface enamel structures to other metriorhynchids with well-preserved dentitions, and assess their continuous and discrete characters through a multivariate analysis aimed to explore the morphospace occupation of each of the tooth crowns among thalattosuchian crocodylomorphs.
Fig. 1. A. Geographic location of the studied area. B. Tectonic map of the Outer Western Carpathian area in the eastern part of the Czech Republic showing the location of Štramberk.
Institutional abbreviations.—MOZ, Museo Provincial de Ciencas Naturales “Prof. Dr. Juan A. Olsacher”, Zapala, Argentina; NHMW, Naturhistorisches Museum Wien, Vienna, Austria.
Other abbreviations.—PCoA, principal coordinates analysis.
Material and methods
The specimens described herein are deposited at the collections of the Naturhistorisches Museum Wien, Vienna, Austria, under the catalogue numbers NHMW 2020/0025/ 0001 (Fig. 2) and NHMW 2020/0025/0002 (Fig. 3). They have been noticed by one of us (SS) during collection surveys in NHMW in June 2020. According to the museum data and the label associated with the specimens, the teeth were discovered in the “Tithonian” strata of the “Nesseldorfer Schichten” of “Stramberg”, acquired in 1912 from “R. Wessely”, and identified as “Teleosaurus suprajurensis” (which according to Fraas 1902 is a subjective junior synonym of Dakosaurus maximus). The geological context (including the stratigraphic provenance) and taxonomic affiliation are merely of historical value and are revised below.

Fig. 2. Tooth crown of the metriorhynchid crocodylomorph Plesiosuchina? indet. (NHMW 2020/0025/0001) from Štramberk, Czech Republic; Valanginian (Lower Cretaceous); in mesial (A1), apical (A2), labial (A3), basal (A4), lingual (A5), and distal (A6) views.
The morphospace occupation of NHMW 2020/0025/0001 and NHMW 2020/0025/0002 among thalattosuchian crocodylomorphs was explored through a principal coordinates analysis (PCoA) using the dataset of Foffa et al. (2018c) that focused on the dental traits of Jurassic thalattosuchians, plesiosaurs, and ichthyosaurs. Because the original version of the dataset did not include the raw measurements necessary to replicate the analyses of Foffa et al. (2018c), we obtained the matrix from Madzia et al. (2021) who were provided these data by Davide Foffa (see Madzia et al. 2021 for details). Owing to the fact that NHMW 2020/0025/0001 and NHMW 2020/0025/0002 differ markedly from the teeth of plesiosaurs (e.g., Foffa et al. 2018a, b; Zverkov et al. 2018; Gao et al. 2019) and ichthyosaurs (e.g., Fischer et al. 2016) we reduced the taxon sampling to include only thalattosuchian representatives. The character list was modified to exclude characters D2, D12, D16 and D17 (the second, twelfth, sixteenth and seventeenth discrete character). Character D2 was scored “1” for all thalattosuchian taxa, character D12 is variable only in pliosaurid plesiosaurs and characters D16 and D17 are variable only in ichthyosaurs. The taxon scores and extended results of the PCoA are provided in the SOM 1 and 2 (Supplementary Online Material available at http://app.pan.pl/SOM/app66-Madzia_etal_SOM.pdf), and the character states are numbered as in Foffa et al. (2018c: Supplementary Information). The PCoA was performed in PAST 4.01 (Hammer et al. 2001). As in Foffa et al. (2018c) and Madzia et al. (2021), continuous characters were z-transformed, and the Gower similarity index was used as it is better suited for datasets which combine continuous and discrete variables (Gower 1971).
Geological setting
Historical Mesozoic outcrops are located around Štramberk in the Silesian Unit (nappe) of the Outer Western Carpathians of northeast Czech Republic (Vašíček et al. 2013; Vašíček and Skupien 2019; Fig. 1). The hillside area with the famous invertebrate (e.g., ammonoids, gastropods) localities comprise several quarries on the Kotouč hill, Castle hill, Horní Skalka hill and Dolní Skalka hill (Hohenegger 1861; Zittel 1868, 1873; Mojsisovics 1870; Böhm 1883; Uhlig 1890; Remeš 1897; Leicher 1931; Houša 1975, 1990; Houša and Vašíček 2004).
Tithonian to Hauterivian limestone and marl represent a deposit that formed in a carbonate platform belt along the northern Tethyan margin during the latest Jurassic–Early Cretaceous. Today they can be traced in the area of the Outer Western Carpathians. Block accumulations of the limestone (especially Štramberk Limestone) form part of the continental-rise sediments of the Baška Facies in the Silesian Unit, which were deposited in the flysch trough of the Baška Subunit (for more details, see Picha et al. 2006).
The true Štramberk Limestone occurs in numerous outrcops around Štramberk. Ammonites of the Štramberk Limestone indicate the age from early Tithonian to the earliest Berriasian (Vašíček and Skupien 2016). Concerning the Štramberk area, Houša and Vašíček (2004) showed that during the Early Cretaceous, deposition of the Štramberk Limestone intermittently passed into carbonate sedimentation (the Olivetská hora and Kopřivnice Limestone).
Lower Cretaceous rocks were considered by Houša and Vašíček (2004) to be infill of fissures, clefts, crevices and cavities or, alternatively, overlie the original surfaces of the Štramberk Limestone. Units of Early Cretaceous age form the Čupek Formation (grey micritic limestone; previously lower part of the Olivetská Hora Formation; uppermost lower Berriasian to lowermost Valanginian), Gloriet Formation (grey breccia; previously upper part of the Olivetská Hora Formation; lowermost upper Valanginian), Kopřivnice Formation (i.e., so-called Kopřivnice [Nesselsdorf] Limestones; upper Valanginian) and Plaňava Formation (black-grey claystone and siltstone of late Valanginian to early Hauterivian age; see Houša and Vašíček 2004).
Limestones of the Kopřivnice Formation are partly redeposited from older sediments, including the Čupek and Gloriet formations (Houša and Vašíček 2004), and are typical for ammonite fragments. Remeš (1897) already reported a mixture of latest Jurassic and Early Cretaceous faunal elements in the red limestones (= “rote Kalke von Nesselsdorf” in Hohenegger 1861) of the Kopřivnice Formation. Most macrofaunal remains (especially crinoid and echinoid elements, but also belemnites, brachiopods, aptychi and other materials) originate from the biodetrital beds of the Čupek Formation. However, there is no evidence to suggest that either of the tooth crowns (NHMW 2020/0025/0001 nor NHMW 2020/0025/0002) were redeposited from the Berriasian of the Čupek Formation. Redeposited material is usually preserved in grey (micritic) limestone that differs from the matrix that is preserved in the basal section (observable in basal view) of the tooth crowns, which instead indicates the late Valanginian age (Neocomites peregrinus or Criosarasinella furcillata Ammonite Zone) for both specimens. Even though there are no precise locality data available for either specimen, the Kopřivnice limestone was mined at two Blücher quarries (Horní Blücherův lom and Dolní Blücherův lom), both located in Štramberk. The teeth likely originate from one of the two quarries.
Systematic palaeontology
Crocodylomorpha Hay, 1930 (sensu Nesbitt 2011)
Thalattosuchia Fraas, 1901 (sensu Young and Andrade 2009)
Metriorhynchoidea Fitzinger, 1843 (sensu Young and Andrade 2009)
Metriorhynchidae Fitzinger, 1843 (sensu Young and Andrade 2009)
Geosaurinae Lydekker, 1889 (sensu Young and Andrade 2009)
Geosaurini Lydekker, 1889 (sensu Cau and Fanti 2011)
Plesiosuchina Young, Andrade, Cornée, Steel, and Foffa, 2014a
Plesiosuchina? indet.
Figs. 2, 4B, 5C.
Material.—NHMW 2020/0025/0001, tooth crown; from Štramberk, Nový Jičín District, Moravian-Silesian Region, Czech Republic; Neocomites peregrinus or Criosarasinella furcillata Ammonite Zone, middle upper or upper upper Valanginian, Kopřivnice Formation, Silesian Unit, Outer Western Carpathians.
Description.—The morphology of NHMW 2020/0025/0001 (Fig. 2) is very similar to the dental morphology of Plesiosuchus manselii specimens from the Kimmeridgian–Tithonian of the UK (Young et al. 2012a), and a cf. Plesiosuchus sp. tooth from the Tithonian of Slovakia (Čerňanský et al. 2019). It is also similar to the dentition of fragmentary specimens from the Cretaceous referred to Plesiosuchina (Young et al. 2014a; Chiarenza et al. 2015). The tooth is ~22.1 mm high apicobasally and its base is 14.41 mm long mesiodistally and 12.5 mm wide labiolingually. It has a caniniform morphology, is single cusped and labiolingually compressed. The crown is curved lingually, but does not curve throughout, only at the middle and apical sections. The mesial margin of the tooth crown has a pronounced distal (posterior) curvature. While distal tooth curvature is present in many metriorhynchids (e.g., see Young et al. 2012a, 2013a, 2014a, 2015b; Chiarenza et al. 2015), as discussed by Young et al. (2012a, 2015b) the pronounced curvature as seen here is restricted to Plesiosuchina.
The basal section of the crown is wider labiolingually, creating a slightly ovoid cross section. Overall, the tooth crown is robust, i.e., retaining its labiolingual width along most of its apicobasal length. The tooth is broken apically to the crown/root junction, thus only the crown is preserved. When the crown is observed in labial view, it is noticeably wider at the base narrowing apically, creating a sub-triangular profile. The labial surface lacks apicobasal planar surfaces (i.e., “facets”), three labial facets are autapomorphic for the geosaurinan genera Geosaurus and Ieldraan (Young and Andrade 2009; Andrade et al. 2010; Foffa et al. 2018b).
The labial and lingual enamel surface ornamentation is composed of numerous apicobasally aligned ridges, which are fairly well-packed but are of low-relief (very similar to that of Plesiosuchus manselii, Young et al. 2012a). As such, they are best observed with good lighting or optical aids (Fig. 5). The ornamentation differs from the densely packed and high-relief ridges observed in Torvoneustes and members of the “E-clade” (Andrade et al. 2010; Young et al. 2013b; Barrientos-Lara et al. 2016; Abel et al. 2020), and the smooth enamel lacking ridges in Dakosaurus andiniensis (Pol and Gasparini 2009), D. maximus and Geosaurus giganteus (Andrade et al. 2010).
The tooth crown has mesial and distal carinae that are serrated with true denticles. As with other microziphodont metriorhynchids (see Andrade et al. 2010; Young et al. 2013a), the denticles are best seen with the use of optical aids. However, the denticles can be seen with the naked eye in well-lit surroundings, although their morphology cannot be properly discerned. Interestingly, the mesial carina is split (sensu Beatty and Heckert 2009), the bifurcation occurring in the basal third of the preserved crown. The superficial enamel ornamentation does not contact the carinae or denticles (i.e., no false ziphodonty). Overall, the denticles have a largely consistent height (isometric), but the shape and dimensions of each denticle vary substantially (poorly isomorphic). As with other plesiosuchinans (Young et al. 2012a; Chiarenza et al. 2015; Čerňanský et al. 2019), the denticles are rectangular in labial or lingual view, rather than the rounded shape seen in other metriorhynchids (Andrade et al. 2010; Young et al. 2013a). Where the carinae are well-preserved, the denticles are contiguous.
Like Plesiosuchus manselii and indeterminate plesiosuchinans (Chiarenza et al. 2015; Young et al. 2015b), NHMW 2020/0025/0001 has “carinal flanges”. These are concave depressions adjacent to the carinae on the lingual surface of the tooth crown, present at the mid-crown. “Carinal flanges” restricted to that region of the tooth crown appears to be a plesiosuchinan apomorphy (Chiarenza et al. 2015), whereas in Dakosaurus these depressions are present on both the labial and lingual surfaces, and are most prominent at the mid-crown and apex (see Young et al. 2015b).
The apex of NHMW 2020/0025/0001 shows signs of macrowear (Fig. 2A2). The wear facet encompasses most of the apical region, removing the outer layer of enamel and dentine. This pattern of tooth wear is common in geosaurin metriorhynchids (e.g., Andrade et al. 2010; Young et al. 2012a, b, 2013a, b, 2015b) and has been interpreted as being caused by tooth-food abrasion. However, enamel spalling where the wear facet continues basally along the labial surface has been only observed in Dakosaurus and closely related specimens (Young et al. 2012a, b, 2013a).
Geosaurini Lydekker, 1889 (sensu Cau and Fanti 2011)
Torvoneustes? sp.
Figs. 3, 4A, 5A, B.
Material.—NHMW 2020/0025/0002, tooth crown; from Štramberk, Nový Jičín District, Moravian-Silesian Region, Czech Republic; Neocomites peregrinus or Criosarasinella furcillata Ammonite Zone, middle upper or upper upper Valanginian, Kopřivnice Formation, Silesian Unit, Outer Western Carpathians.
Description.—The morphology of NHMW 2020/0025/0002 (Fig. 3) is similar to the dental morphology of species of Torvoneustes from the Kimmeridgian–Tithonian of the UK and Mexico (see Andrade et al. 2010; Young et al. 2013a, b, 2020; Barrientos-Lara et al. 2016). The tooth is 25.94 mm high apicobasally and its base is 19.31 mm long mesiodistally and 15.6 mm wide labiolingually. The tooth has a caniniform morphology, is single cusped and has little labiolingual compression. The crown is curved lingually, but does not curve throughout, only at the middle and apical sections. The mesial margin of the tooth crown has a slight distal curvature, very different from NHMW 2020/0025/0001.
The basal section of the crown is wider labiolingually, creating a slightly sub-circular to sub-ovoid cross section. Overall, the tooth crown is robust, i.e., retaining its labiolingual width along most of its apicobasal length. The tooth is broken apical to the crown/root junction, thus only the crown is preserved. When the crown is observed in labial view, it is wider at the base narrowing apically, creating a sub-triangular profile. The labial surface lacks the labial apicobasal facets that are for the autapomorphic of Geosaurus and Ieldraan (Young and Andrade 2009; Andrade et al. 2010; Foffa et al. 2018b).
The labial and lingual enamel surface ornamentation is composed of numerous apicobasally aligned ridges that are very tightly-packed (as seen in other species of Torvoneustes, e.g., see Barrientos-Lara et al. 2016). The ridges themselves are best seen under macrophotography, as it appears as though they have undergone taphonomic distortion. Apically the enamel ridges change in length and orientation. They become much shorter, but also thicker, their shapes are irregular, varying from “bubble”-like shapes to thick ridges to undulating merged ridges creating an anastomosed pattern. In the apical region some of the superficial enamel ornamentation contacts the carinae and denticles (false ziphodonty). In thalattosuchians, this shift in apical enamel ornamentation is characteristic of Torvoneustes (Andrade et al. 2010; Young et al. 2013a, b, 2020; Barrientos-Lara et al. 2016) and machimosaurin teleosauroids (Young et al. 2014b, c, 2015a; Jouve et al. 2016, Johnson et al. 2018, 2020).
The tooth crown has mesial and distal carinae that are serrated with true denticles. The carinae are microziphodont, with the denticles observable with optical aids. The carinae themselves are very prominent, with the keel being very well-developed, an autapomorphic feature of Torvoneustes (Young et al. 2013a, 2020). There are no “carinal flanges” like those seen in Dakosaurus or Plesiosuchina (Young et al. 2015b). In the basal region where the enamel ornamentation does not contact the carinae, the denticles are consistent height (isometric), but the shape and dimensions of the denticles can vary substantially (poorly isomorphic). The interdenticular spaces are variable in dimension as well. Apically where the enamel ornamentation does contact the carinae, the denticles are also isometric but poorly isomorphic, but where the enamel ornamentation fuses with the denticles the morphology becomes more aberrant. Overall, the denticles are contiguous along the carinae, which is one of the characters that differentiates between Torvoneustes and machimosaurin teleosauroids, which have incipient microdenticles that do not form a contiguous series (see Young et al. 2014c, 2015a).
The apex is slightly rounded and has macrowear (Fig. 3A2). The wear facet is largely on the labial surface of the apical region, removing the enamel and exposing the underlying dentine.

Fig. 3. Tooth crown of the metriorhynchid crocodylomorph Torvoneustes? sp. (NHMW 2020/0025/0002) from Štramberk, Czech Republic; Valanginian (Lower Cretaceous); in mesial (A1), apical (A2), labial (A3), basal (A4), lingual (A5), and distal (A6) views.

Fig. 4. Metriorhynchid crocodylomorph tooth crowns from Štramberk, Czech Republic; Valanginian (Lower Cretaceous. A. Torvoneustes? sp., NHMW 2020/0025/0002. B. Plesiosuchina? indet., NHMW 2020/0025/ 0001. SEM micrographs of the carinae (A1, A2, B1, B2) showing the denticle morphologies. General views (A3, B3) indicating the sections where SEM micrographs were obtained. Scale bars 1 mm.

Fig. 5. Metriorhynchid crocodylomorph tooth crown of Torvoneustes? sp., NHMW 2020/0025/0002 (A, B) and Plesiosuchina? indet., NHMW 2020/0025/0001 (C) from Štramberk, Czech Republic; Valanginian (Lower Cretaceous). A. SEM micrograph showing split carina (arrows). B, C. SEM micrograph showing lingual crown surface texture. Scale bars 5 mm.
Results of the principal coordinates analysis
The principal coordinates analysis (PCoA) largely separates metriorhynchids and teleosauroids from one another (Fig. 6). The teleosauroids are exclusively found in the positive section of the second coordinate axis, while metriorhynchids are dispersed across the morphospace. However, metriorhynchines fall within the morphospace occupied by geosaurines. Therefore, their separation from each other might not be expressed through their tooth crown morphology. Both specimens described in this study, NHMW 2020/0025/0001 and NHMW 2020/0025/0002, plot on the negative side of the second axis and are placed far from each other, supporting the interpretation that they belong to distinct taxa. Also, both specimens may be interpreted as plotting within the geosaurine morphospace.
It is essential to note that some dental features have been inferred to develop independently in thalattosuchian lineages sharing a similar ecology (Foffa et al. 2018c). Even though our PCoA apparently “recognizes” the convergent nature of these traits (see e.g., the placement of Torvoneustes and Machimosaurini that share blunt apices and enamel bearing a conspicuous pattern), it cannot be ruled out that the morphospace occupation of NHMW 2020/0025/0001 and NHMW 2020/0025/0002 is at least partly affected by convergence.
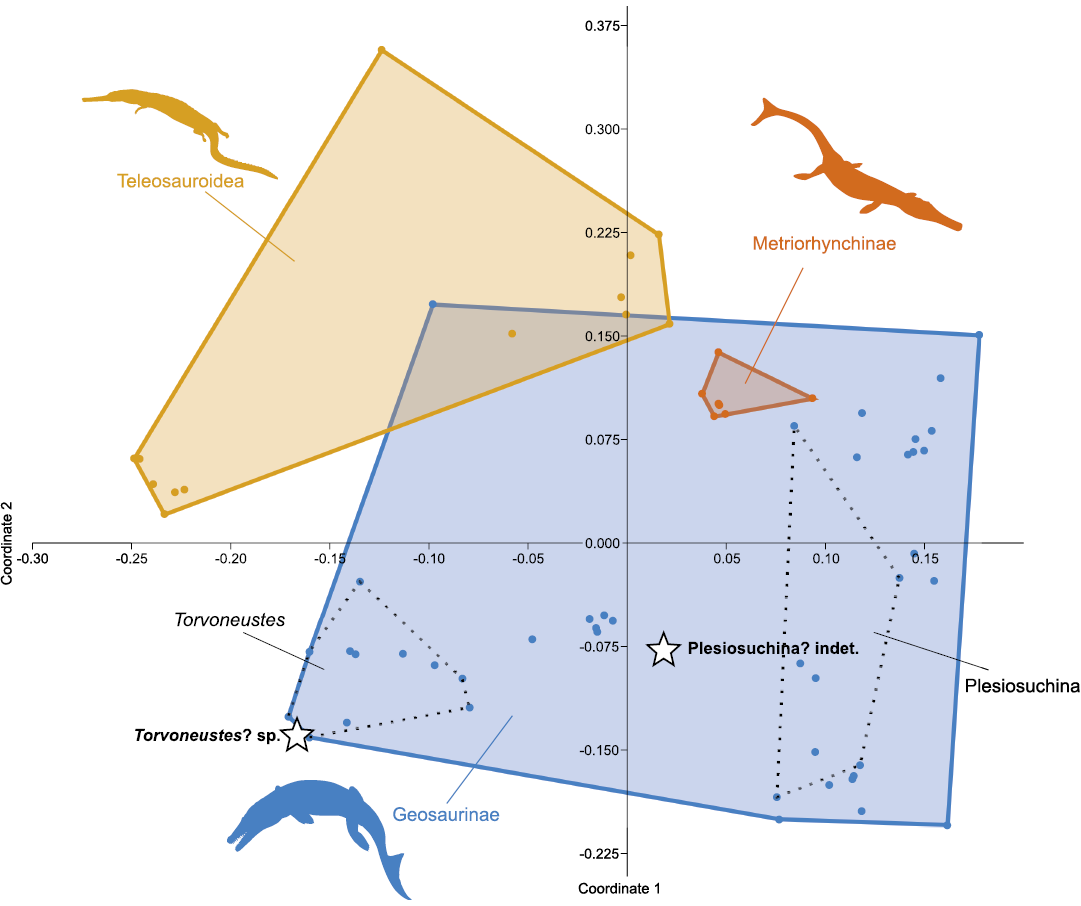
Fig. 6. Morphospace occupation of Plesiosuchina? indet. (NHMW 2020/0025/0001) and Torvoneustes? sp. (NHMW 2020/0025/0002) among the thalattosuchian crocodylomorphs using principal coordinates 1 and 2. See SOM 1 and 2 for extended results of the principal coordinates analysis. Silhouettes obtained from phylopic.org: Geosaurinae (Dmitry Bogdanov, CC BY 3.0), Metriorhynchinae and Teleosauroidea (Gareth Monger, CC BY 3.0).
The comparisons between NHMW 2020/0025/0001 and other known metriorhynchids indicate that the crown shows affinities to plesiosuchinan geosaurines. The results of PCoA are slightly ambiguous with that respect. NHMW 2020/0025/0001 does not fall within the morphospace of Plesiosuchus manselii (and Plesiosuchus-like material) and Suchodus durobrivensis, the taxa that form Plesiosuchina (e.g., Foffa et al. 2018b; Cau 2019; Young et al. 2020), as expressed through the convex hulls although it falls within the plesiosuchinan 95% confidence ellipse. Considering these results, combined with the comparisons, we regard our designation of NHMW 2020/0025/0001 as Plesiosuchina? indet. to be acceptable. Note, the tooth crown described here differs from the Jurassic members of Plesiosuchina in having observable enamel ridges (much like the Plesiosuchina specimen from the Valanginian of France; Young et al. 2014a).
The affinities of NHMW 2020/0025/0002 to other geosaurines is more clear, as the specimen is placed in close proximity to the morphospace occupied by the species of Torvoneustes (closer to T. carpenteri than to T. coryphaeus).
Discussion
Implications for the diversity of Cretaceous metriorhynchids.—Cretaceous metriorhynchids have so far been known from 18 records, four of which have uncertain stratigraphic placement and may be latest Tithonian (latest Jurassic) or earliest Berriasian (earliest Cretaceous) in age (see Sachs et al. 2020: table 1). Regardless, these records show that at least five lineages crossed the Jurassic–Cretaceous boundary, two within Rhacheosaurini, Plesiosuchina, Geosaurina, and “Dakosaurina”. Some other specimens appear to be undiagnostic beyond Metriorhynchidae indet., which may also hinder estimations of the actual diversity patterns within the Early Cretaceous metriorhynchid crocodylomorphs. Note that Sachs et al. (2020) listed Dakosaurus andiniensis to be among the taxa with the uncertain latest Tithonian or earliest Berriasian age, while in fact one specimen referred to the taxon (MOZ 6140P) originates from the lower Berriasian strata (Pol and Gasparini 2009); hence the presence of “Dakosaurina” among the lineages crossing the Jurassic–Cretaceous boundary (above).
Nevertheless, following the interpretation of NHMW 2020/0025/0002 as Torvoneustes? sp., we propose that the “Subclade T” (sensu Foffa et al. 2018b) has survived to the Cretaceous as well and reached at least the middle late Valanginian, prolonging the existence of the lineage by ~10 Ma and increasing the taxic diversity of Cretaceous metriorhynchids.
Feeding ecology of the Štramberk metriorhynchids.—The morphologies of NHMW 2020/0025/0001 and NHMW 2020/0025/0002 indicate that the tooth-bearing taxa belonged to different feeding guilds. Species of Plesiosuchus and taxa sharing similar dental morphologies (such as NHMW 2020/0025/0001), show macrophagous adaptations (Foffa et al. 2018c) and may have been specialists feeding on other large-bodied vertebrates, such as other marine reptiles (Young et al. 2012a), while the stouter teeth with blunt apices and conspicuous ornamentation of specimens belonging or similar to Torvoneustes spp. (such as NHMW 2020/0025/0002) are suggestive of the durophagous diet (Foffa et al. 2018c).
Conclusions
We report two isolated crocodylomorph tooth crowns (NHMW 2020/0025/0001 and NHMW 2020/0025/0002) from the upper Valanginian (Lower Cretaceous) of the Kopřivnice Formation in the Štramberk area of the Czech Republic. The specimens represent the first evidence of Metriorhynchidae from the Czech Republic and some of the youngest metriorhynchid records worldwide.
Our assessment of the material shows that the teeth represent two distinct taxa of geosaurin metriorhynchids, designated here as Plesiosuchina? indet. (for NHMW 2020/0025/ 0001) and Torvoneustes? sp. (NHMW 2020/0025/0002). The crown morphologies indicate that the tooth-bearing taxa had different feeding adaptations; NHMW 2020/0025/0001 likely represented a specialist feeding on other large-bodied vertebrates, while taxa with teeth similar to NHMW 2020/0025/0002 have previously been inferred to have a durophagous diet (Foffa et al. 2018c).
Finally, the presence of a taxon related to Torvoneustes spp. in the late Valanginian of the European archipelago prolongs the existence of the Torvoneustes lineage (“Subclade T” of Foffa et al. 2018b) by ~10 Ma and increases the taxic diversity of the late-surviving metriorhynchids.
Acknowledgements
We thank Ursula Göhlich (NHMW) for providing access to the specimens under her care and Yanina Herrera (División Paleontología Vertebrados, Museo de La Plata, Facultad de Ciencias Naturales y Museo, UNLP, CONICET, La Plata, Argentina) for discussion on South American metriorhynchids. We further thank Davide Foffa (National Museums Scotland, Edinburgh, UK) for discussion. The reviews by Yanina Herrera and Lorna Steel (Natural History Museum, London, UK) improved the manuscript. This work was supported by a Leverhulme Trust Research Project (grant number RPG-2017-167) (financially supporting MTY). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
Abel, P., Sachs, S. and Young, M.T. 2020. Metriorhynchid crocodylomorphs from the lower Kimmeridgian of Southern Germany: evidence for a new large-bodied geosaurin lineage in Europe. Alcheringa 44: 312–326. Crossref
Andrade, M.B., Young, M.T., Desojo, J.B., and Brusatte, S.L. 2010. The evolution of extreme hypercarnivory in Metriorhynchidae (Mesoeucrocodylia: Thalattosuchia) based on evidence from microscopic denticle morphology. Journal of Vertebrate Paleontology 30: 1451–1465. Crossref
Andrews, C.W. 1913. A Descriptive Catalogue of the Marine Reptiles of the Oxford Clay, Part Two. xxiv + 206 pp., 13 pl. British Museum (Natural History), London.
Barrientos-Lara, J.I., Herrera, Y., Fernández, M.S., and Alvarado-Ortega, J. 2016. Occurrence of Torvoneustes (Crocodylomorpha, Metriorhynchidae) in marine Jurassic deposits of Oaxaca, Mexico. Revista Brasileira de Paleontologia 19: 415–424. Crossref
Beatty, B.L. and Heckert, A.B. 2009. A large archosauriform tooth with multiple supernumerary carinae from the Upper Triassic of New Mexico (USA), with comments on carina development and anomalies in the Archosauria. Historical Biology 21: 57–65. Crossref
Böhm, G.D. 1883. Die Bivalven der Stramberger Schichten. Palaeontographica. Supplementbände II 4: 403–680.
Cau, A. 2019. A revision of the diagnosis and affinities of the metriorhynchoids (Crocodylomorpha, Thalattosuchia) from the Rosso Ammonitico Veronese Formation (Jurassic of Italy) using specimen-level analyses. PeerJ 7: e7364. Crossref
Cau, A. and Fanti, F. 2011. The oldest known metriorhynchid crocodilian from the Middle Jurassic of North-eastern Italy: Neptunidraco ammoniticus gen. et sp. nov. Gondwana Research 19: 550–565. Crossref
Cortés, D., Larsson, H.C.E., Maxwell, E.E., Parra Ruge, M.L., Patarroyo, P., and Wilson, J.A. 2019. An Early Cretaceous teleosauroid (Crocodylomorpha: Thalattosuchia) from Colombia. Ameghiniana 56: 365–379. Crossref
Chiarenza, A., Foffa, D., Young, M.T., Insacco, G., Cau, A., Carnevale, G., and Catanzariti, R. 2015. The youngest record of metriorhynchid crocodylomorphs, with implications for the extinction of Thalattosuchia. Cretaceous Research 56: 608–616. Crossref
Čerňanský, A., Schlögl, J., Mlynský, T., and Józsa, S. 2019. First evidence of the Jurassic thalattosuchian (both teleosaurid and metriorhynchid) crocodylomorphs from Slovakia (Western Carpathians). Historical Biology 31: 1008–1015. Crossref
Debelmas, J. 1952. Un crocodilien marin dans l’Hauterivien des environs de Comps (Var) Dacosaurus maximus Plieninger var. gracilis Quenstedt. Travaux Laboratoire de Géologie de l’université de Grenoble 29: 101–116.
Debelmas, J. and Demains D’Archimaud, J. 1956. Sur un métatarsien de Dacosaure trouvé à Robion, près Castellane (Basses-Alpes). Travaux Laboratoire de Géologie de l’université de Grenoble 33: 101–104.
Eudes-Deslongchamps, E. 1867–1869. Prodrome des Téléosauriens du Calvados. Notes Paléontologiques Caen, 95–162 (year 1867); 163–196 (year 1868); 197–354, 374–379, 385–392 (year 1869); pl. 10–24. Le Blanc-Hardel, Savy.
Fanti, F., Miyashita, T., Cantelli, L., Mnasri, F., Dridi, J., Contessi, M., and Cau, A. 2016. The largest thalattosuchian (Crocodylomorpha) supports teleosaurid survival across the Jurassic–Cretaceous boundary. Cretaceous Research 61: 263–274. Crossref
Fernández, M.S., Herrera, Y., Vennari, V.V., Campos, L., de la Fuente, M., Talevi, M., and Aguirre-Urreta, B. 2019. Marine reptiles from the Jurassic/Cretaceous transition at the High Andes, Mendoza, Argentina. Journal of South American Earth Sciences 92: 658–673.
Fischer, V., Bardet, N., Benson, R.B.J, Arkhangelsky, M.S., and Friedman, M. 2016. Extinction of fish-shaped marine reptiles associated with reduced evolutionary rates and global environmental volatility. Nature Communications 7: 10825. Crossref
Fitzinger, L.J.F.J. 1843. Systema Reptilium. Fasciculus primus. Amblyglossae. 106 pp. Braumüller et Seidel Press, Vienna.
Foffa, D., Johnson, M.M., Young, M.T., Steel, L., and Brusatte, S.L. 2019. Revision of the Late Jurassic deep-water teleosauroid crocodylomorph Teleosaurus megarhinus Hulke, 1871 and evidence of pelagic adaptations in Teleosauroidea. PeerJ 7: e6646. Crossref
Foffa, D., Young, M.T., and Brusatte, S.L. 2018a. Filling the Corallian gap: New information on Late Jurassic marine reptile faunas from England. Acta Palaeontologica Polonica 63: 287–313. Crossref
Foffa, D., Young, M.T., Brusatte, S.L., Graham, M.R. and Steel, L. 2018b. A new metriorhynchid crocodylomorph from the Oxford Clay Formation (Middle Jurassic) of England, with implications for the origin and diversification of Geosaurini. Journal of Systematic Palaeontology 16: 1123–1143. Crossref
Foffa, D., Young, M.T., Stubbs, T.L., Dexter, K.G., and Brusatte, S.L. 2018c. The long-term ecology and evolution of marine reptiles in a Jurassic seaway. Nature Ecology & Evolution 2: 1548–1555. Crossref
Fraas, E. 1901. Die Meerkrokodile (Thalattosuchia n. g.) eine neue Sauriergruppe der Juraformation. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg 57: 409–418.
Fraas, E. 1902. Die Meer-Crocodilier (Thalattosuchia) des oberen Jura unter specieller Berücksichtigung von Dacosaurus und Geosaurus. Palaeontographica 49: 1–72.
Gao, T., Li, D.-Q., Li, L.-F., and Yang, J.-T. 2019. The first record of freshwater plesiosaurian from the Middle Jurassic of Gansu, NW China, with its implications to the local palaeobiogeography. Journal of Palaeogeography 8: 27. Crossref
Gower, J.C. 1971. A general coefficient of similarity andsome of its properties. Biometrics 27: 857–871. Crossref
Hammer, Ø., Harper, D.A.T., and Ryan, P.D. 2001. PAST: PAleontological STatistics software package for education and data analysis. Palaeontologia Electronica 4: 1–9.
Hay, O.P. 1930. Second Bibliography and Catalogue of the Fossil Vertebrata of North America. 1074 pp. Carnegie Institute Washington, Washington DC.
Herrera, Y., Gasparini, Z., and Fernández, M.S. 2015. Purranisaurus potens Rusconi, an enigmatic metriorhynchid from the Late Jurassic–Early Cretaceous of the Neuquén Basin. Journal of Vertebrate Paleontology 35: e904790. Crossref
Hohenegger, L. 1861. Die geognostischen Verhältnisse der Nordkarpathen in Schlesien und der angrenzenden Teilen von Mähren und Galizien, als Erläuterung zur geognostischen Karte der Nordkarpathen. 50 pp. Justus Perthes, Gotha.
Houša, V. 1975. Geology and paleontology of the Stramberg limestone (Upper Tithonian) and the associated Lower Cretaceous Beds. Mémoires du Bureau Recherches Géologiques et Miniers 86: 342–349.
Houša V. 1990. Stratigraphy and calpionellid zonation of the Štramberk Limestone and associated Lower Cretaceous beds. In: G. Pallini, F. Cecca, S. Cresta, and M. Santanorio (eds.), Atti del Secondo Convegno Internattzuionale “Fossili, Evoluzione, Ambiente” in Pergola 1987, 365–370. Comitato Centenario Raffaele Piccinini, Pergola.
Houša, V. and Vašíček, Z. 2004. Ammonoidea of the Lower Cretaceous Deposits (Late Berriasian, Valanginian, Early Hauterivian) from Štramberk, Czech Republic. GeoLines 18: 7–57.
Hua, S., Vignaud, P., and Efimov, V. 1998. First record of Metriorhynchidae (Crocodylomorpha, Mesosuchia) in the Upper Jurassic of Russia. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1998 (8): 475–484. Crossref
Johnson, M.M., Young, M.T., and Brusatte, S.L. 2020. Re-description of two contemporaneous mesorostrine teleosauroids (Crocodylomorpha, Thalattosuchia) from the Bathonian of England, and insights into the early evolution of Machimosaurini. Zoological Journal of the Linnean Society 189: 449–482. Crossref
Johnson, M.M., Young, M.T., Steel, L., Foffa, D., Smith, A.S., Hua, S., Havlik, P., Howlett, E.A., and Dyke, G. 2018. Re-description of “Steneosaurus” obtusidens Andrews, 1909, an unusual macrophagous teleosaurid crocodylomorph from the Middle Jurassic of England. Zoological Journal of the Linnean Society 182: 385–418. Crossref
Jouve, S., Mennecart, B., Douteau, J., and Jalil, N.E. 2016. The oldest durophagous teleosaurid (Crocodylomorpha, Thalattosuchia) from the Lower Bathonian of Central High Atlas, Morocco. Palaeontology 59: 863–876. Crossref
Leicher, J. 1931. Zur Tektonik der Stramberger Tithonklippen. Lotos Prag: 59–93.
Lydekker, R. 1889. On the remains and affinities of five genera of Mesozoic reptiles. Quarterly Journal of the Geological Society 45: 41–59. Crossref
Madzia, D., Szczygielski, T., and Wolniewicz, A.S. 2021. The giant pliosaurid that wasn’t—revising the marine reptiles from the Kimmeridgian (Upper Jurassic) of Krzyżanowice, Poland. Acta Palaeontologica Polonica 66: 99–129. Crossref
Martin, J.E., Suteethorn, S., Lauprasert, K., Tong, H., Buffetaut, E., Liard, R., Salaviale, C., Deesri, U., Suteethorn, V., and Claude, J. 2018. A new freshwater teleosaurid from the Jurassic of northeastern Thailand. Journal of Vertebrate Paleontology 38: e1549059. Crossref
Mojsisovics, E. von 1870. Durchschnitt von Stramberg nach Nesselsdorf. Verhandlungen der Kaiserlich-Königlichen geologischen Reichsanstalt 8: 136–139.
Nesbitt, S.J. 2011. The early evolution of archosaurs: relationships and the origin of major clades. Bulletin of the American Museum of Natural History 352: 1–292. Crossref
Ochev, V.G. [Očev, V.G.] 1981. Marine crocodiles in the Mesozoic of Povolzh’e [in Russian]. Priroda 1981: 103.
Picha, F.J., Stráník, Z., and Krejčí, O. 2006. Geology and hydrocarbon resources of the Outer Western Carpathians and their foreland: geology and hydrocarbon resources. American Association of Petroleum Geologists, Memoir 84: 49–175. Crossref
Pol, D. and Gasparini, Z. 2009. Skull anatomy of Dakosaurus andiniensis (Thalattosuchia: Crocodylomorpha) and the phylogenetic position of Thalattosuchia. Journal of Systematic Palaeontology 7: 163–197. Crossref
Remeš, M. 1897. Ueber den rothen Kalkstein von Nesselsdorf. Bericht vom 31. August. Verhandlungen der Kaiserlich-Königlichen Reichsanstalt 11: 221–229.
Sachs, S., Young, M.T., and Hornung, J.J. 2020. The enigma of Enaliosuchus, and a reassessment of the Lower Cretaceous fossil record of Metriorhynchidae. Cretaceous Research 114: 104479. Crossref
Uhlig, V. 1890. Ergebnisse geologischer Aufnahmen in den westgalizischen Karpathen. II Theil. Der pieninische Klippenzug. Jahrbuch der Kaiserlich-königlichen Reichsanstalt 40: 559–824.
Vašíček, Z. and Skupien, P. 2016. Tithonian–early Berriasian perisphinctoid ammonites from the Štramberk Limestone at Kotouč Quarry near Štramberk, Outer Western Carpathians (Czech Republic). Cretaceous Research 64: 12–29. Crossref
Vašíček, Z. and Skupien, P. 2019. New Tithonian and lower Berriasian aptychi of Štramberk Limestone from the Kotouč Quarry (Outer Western Carpathians, Czech Republic). Paläontologische Zeitschrift 93: 37–48. Crossref
Vašíček, Z., Skupien, P., and Jirásek, J. 2013. The northernmost occurrence of the lower Berriasian ammonite Pseudosubplanites grandis (Štramberk Limestone, Outer Western Carpathians, Czech Republic). Geologica Carpathica 64: 461–466. Crossref
Wilberg, E.W. 2015. A new metriorhynchoid (Crocodylomorpha, Thalattosuchia) from the Middle Jurassic of Oregon and the evolutionary timing of marine adaptations in thalattosuchian crocodylomorphs. Journal of Vertebrate Paleontology 35: e902846. Crossref
Young, M.T. and Andrade, M.B. 2009. What is Geosaurus? Redescription of Geosaurus giganteus (Thalattosuchia, Metriorhynchidae) from the Upper Jurassic of Bayern, Germany. Zoological Journal of the Linnean Society 157: 551–585. Crossref
Young, M.T. and Sachs, S. 2020. Evidence of thalattosuchian crocodylomorphs in the Portland Stone Formation (Late Jurassic) of England, and a discussion on Cretaceous teleosauroids. Historical Biology [published online, https://doi.org/10.1080/08912963.2019.1709453]. Crossref
Young, M.T., Andrade, M.B., Brusatte, S.L., Sakamoto, M., and Liston J. 2013a. The oldest known metriorhynchid super-predator: a new genus and species from the Middle Jurassic of England, with implications for serration and mandibular evolution in predacious clades. Journal of Systematic Palaeontology 11: 475–513. Crossref
Young, M.T., Andrade, M.B., Cornée, J.-J., Steel, L., and Foffa, D. 2014a. Re-description of a putative Early Cretaceous “teleosaurid” from France, with implications for the survival of metriorhynchids and teleosaurids across the Jurassic–Cretaceous Boundary/Nouvelle description d’un “téléosauridé” hypothétique de France et implications sur la survie des métriorhynchidés et des téléosauridés au passage Jurassique-Crétacé. Annales de Paléontologie 100: 165–174. Crossref
Young, M.T., Andrade, M.B., Etches, S., and Beatty, B.L. 2013b. A new metriorhynchid crocodylomorph from the lower Kimmeridge Clay Formation (Late Jurassic) of England, with implications for the evolution of dermatocranium ornamentation in Geosaurini. Zoological Journal of the Linnean Society 169: 820–848. Crossref
Young, M.T., Beatty, B.L., Brusatte, S.L., and Steel, L. 2015a. First evidence of denticulated dentition in teleosaurid crocodylomorphs. Acta Palaeontologica Polonica 60: 661–671.
Young, M.T., Brusatte S.L., Andrade, M.B., Desojo, J.B., Beatty, B.L., Steel, L., Fernández, M.S., Sakamoto, M., Ruiz-Omeñaca, J.I. and Schoch, R.R. 2012a. The cranial osteology and feeding ecology of the metriorhynchid crocodylomorph genera Dakosaurus and Plesiosuchus from the Late Jurassic of Europe. PLOS ONE 7: e44985. Crossref
Young, M.T., Brusatte, S.L., Beatty, B.L., Andrade, M.B., Desojo, J.B. 2012b. Tooth-on-tooth interlocking occlusion suggests macrophagy in the Mesozoic marine crocodylomorph Dakosaurus. The Anatomical Record 295: 1147–1158. Crossref
Young, M.T., Foffa, D., Steel, L., and Etches, S. 2020. Macroevolutionary trends in the genus Torvoneustes (Crocodylomorpha: Metriorhynchidae) and discovery of a giant specimen from the Late Jurassic of Kimmeridge, UK. Zoological Journal of the Linnean Society 189: 483–493. Crossref
Young, M.T., Hua, S., Steel, L., Foffa, D., Brusatte, S.L., Thüring, S., Mateus, O., Ruiz-Omeñaca, J.I., Havlik, P., Lepage, Y., and Andrade, M.B. 2014b. Revision of the Late Jurassic teleosaurid genus Machimosaurus (Crocodylomorpha, Thalattosuchia). Royal Society Open Science 1: 140222. Crossref
Young, M.T., Sachs, S., Abel, P., Foffa, D., Herrera, Y., and Kitson, J.J.N. 2020. Convergent evolution and possible constraint in the posterodorsal retraction of the external nares in pelagic crocodylomorphs. Zoological Journal of the Linnean Society 189: 494–520. Crossref
Young, M.T., Steel, L., Brusatte, S.L., Foffa, D., and Lepage, Y. 2014c. Tooth serration morphologies in the genus Machimosaurus (Crocodylomorpha, Thalattosuchia) from the Late Jurassic of Europe. Royal Society Open Science 1: 140269. Crossref
Young, M.T., Steel, L., Rigby, M.P., Howlett, E.A., and Humphrey, S. 2015b. Largest known specimen of the genus Dakosaurus (Metriorhynchidae: Geosaurini) from the Kimmeridge Clay Formation (Late Jurassic) of England, and an overview of Dakosaurus specimens discovered from this formation (including reworked specimens from the Woburn Sands Formation). Historical Biology 27: 947–953. Crossref
Zittel, K. 1868. Die Cephalopoden der Stramberger Schichten. Paläontologische Mitteilungen aus dem Museums der Königlich Bayerischen Staatssammlung 2: 33–118.
Zittel, K. 1873. Die Gastropoden der Stramberger Schichten. Paläontologische Mitteilungen aus dem Museums der Königlich Bayerischen Staatssammlung 2: 311–491.
Zverkov, N.G., Fischer, V., Madzia, D., and Benson, R.B.J. 2018. Increased pliosaurid dental disparity across the Jurassic–Cretaceous transition. Palaeontology 61: 825–846. Crossref
Acta Palaeontol. Pol. 66 (2): 357–367, 2021
https://doi.org/10.4202/app.00801.2020