
A large hadrosaurid dinosaur from Presa San Antonio, Cerro del Pueblo Formation, Coahuila, Mexico
ROGELIO ANTONIO REYNA-HERNÁNDEZ, HÉCTOR E. RIVERA-SYLVA, LUIS E. SILVA-MARTÍNEZ, and JOSÉ RUBÉN GUZMAN-GUTIÉRREZ
Reyna-Hernández, R.A., Rivera-Sylva, H.E., Silva-Martínez, L.E., and Guzman-Gutiérrez, J.R. 2021. A large hadrosaurid dinosaur from Presa San Antonio, Cerro del Pueblo Formation, Coahuila, Mexico. Acta Palaeontologica Polonica 66 (Supplement to 3): 101–110.
New hadrosaurid postcranial material is reported, collected near Presa San Antonio, Parras de la Fuente municipality, Coahuila, Mexico, in a sedimentary sequence belonging to the upper Campanian of the Cerro del Pueblo Formation, in the Parras Basin. The skeletal remains include partial elements from the pelvic girdle (left ilium, right pubis, ischium, and incomplete sacrum), a distal end of a left femur, almost complete right and left tibiae, right metatarsals II and IV, cervical and caudal vertebrae. Also, partially complete forelimb elements are present, which are still under preparation. The pubis shows characters of the Lambeosaurinae morphotypes, but the lack of cranial elements does not allow us to directly differentiate this specimen from the already described hadrosaurid taxa from the studied area, such as Velafrons coahuilensis, Latirhinus uitstlani, and Kritosaurus navajovius. This specimen, referred as Lambeosaurinae indet., adds to the fossil record of the hadrosaurids in southern Laramidia during the Campanian.
Key words: Dinosauria, Hadrosauridae, Lambeosaurinae, Cretaceous, Campanian, Mexico.
Rogelio Antonio Reyna-Hernández [rogelio.reynahr@gmail.com], Luis E. Silva-Martínez [luis.silva.paleo@gmail.com], Laboratorio de Paleobiología, Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León, Av. Pedro de Alba y Manuel L. Barragán s/n, Ciudad Universitaria, San Nicolás de los Garza, Nuevo León, México. C.P. 66455.
Héctor E. Rivera-Sylva [hrivera@museodeldesierto.org ] (corresponding author), José Rubén Guzman-Gutiérrez [paleovert@yahoo.com.mx], Departamento de Paleontología, Museo del Desierto, Carlos Abedrop Dávila 3745, Saltillo, Coahuila, México. C.P. 25022.
Received 1 October 2020, accepted 15 January 2021, available online 14 June 2021.
Copyright © 2021 R.A. Reyna-Hernández et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Hadrosaurids are a very distinctive and common group of ornithopod dinosaurs in Late Cretaceous ecosystems. Their stratigraphic range begins in the late Santonian and ends together with the other non-avian dinosaurs at the K/Pg extinction event at the end of the Maastrichtian. They also have a wide geographical distribution, as specimens of this clade have been found on all continents except Oceania (Horner et al. 2004; Lund and Gates 2006; Prieto-Márquez 2010a; Longrich et al. 2021). These ornithopods differ from the other groups of dinosaurs due to the adaptations of the nasal region, where the bones have been modified in specialized structures such as cranial crests (e.g., Lull and Wright 1942; Ostrom 1961b). Furthermore, they present a complex dental arrangement made up of different series of rows of teeth and keratinous bills that allowed them a more efficient herbivorous diet (Morris 1970; Nabavizadeh 2014; Nabavizadeh and Weishampel 2016). Most of the diagnostic elements for this family are found in the cranial region (Prieto-Márquez 2010b), from which large number of species has been differentiated. However, the pectoral and pelvic girdles also provide a wide range of taxonomic information that does not always receives the proper importance (Brett-Surman 1989; Campione 2014).
In Mexico, the Hadrosauridae presents the widest record of any group of dinosaurs, with most of the reports coming from the northern region (Ramírez-Velasco et al. 2014; Rivera-Sylva and Carpenter 2014; Ramírez-Velasco and Hernández-Rivera 2015). Despite this, the study of Mexican specimens has been relatively scarce due to the fragmentary condition of most of the specimens, although the number of hadrosaurid studies in Mexico has recently increased (Kirkland et al. 2006; Serrano-Brañas et al. 2006; Ramírez-Velasco et al. 2014; Serrano-Brañas and Espinosa-Chávez 2017; Rivera-Sylva et al. 2019a; Rybakiewicz et al. 2019).
The Cerro del Pueblo Formation is one of the most studied geological formations in Mexico with a very diverse faunal record, composed mainly of dinosaurs and other reptiles (Wolleben 1977; Rodriguez-de la Rosa and Cevallos-Ferriz 1998; Kirkland et al. 2000; Cifuentes-Ruiz et al. 2006; Rivera-Sylva and Espinosa-Chávez 2006; Loewen et al. 2010; Brinkman 2014; Rivera-Sylva et al. 2018, 2019b; Serrano-Brañas et al. 2020), flora (Rodriguez-de la Rosa and Cevallos-Ferriz 1994; Estrada-Ruiz and Cevallos-Ferriz 2007; Estrada-Ruiz et al. 2009; Cevallos-Ferriz and Vázquez-Rueda 2016), and trace fossils (Rodriguez-de la Rosa and Cevallos-Ferriz 1998; Rodríguez-de la Rosa 2007; Rivera-Sylva et al. 2017; Serrano-Brañas et al. 2018a, b, 2019). Three hadrosaurid taxa are known so far from this formation: Kritosaurus navajovius Brown, 1910 (Kirkland et al. 2006; Prieto-Márquez 2014b), Latirhinus uitstlani Prieto-Márquez and Serrano Brañas, 2012 (Prieto-Márquez and Serrano Brañas 2012) and Velafrons coahuilensis Gates, Sampson, Delgado de Jesús, Zanno, Eberth, Hernandez-Rivera, Aguillón-Martínez, and Kirkland, 2007 (Gates et al. 2007). The latter species was described based on cranial elements only, despite the fact that there are a number of preserved postcranial elements.
We describe a new hadrosaurid specimen consisting only of postcranial material from the Cerro del Pueblo Formation collected in Parras de la Fuente municipality, Coahuila, Mexico. The study of these materials would allow for better overview of the faunal diversity within the fossil ecosystems of the Upper Cretaceous in Mexico and provide a more robust anatomical knowledge of the species of the region.
Institutional abbreviations.—AEHM, Amur Natural History Museum of the Far Eastern Institute of Mineral Resources, FEB RAS, Blagoveshchensk, Russia; CPC, Colección Paleobiológica de Coahuila, Museo del Desierto, Saltillo, Mexico; FCBUANL, Colección Paleobiológica, Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León, San Nicolás de los Garza, Mexico; GMH, Geological Museum of Heilongjiang, Harbin, China; IGM, Museo de Paleontología, Instituto de Geología, Universidad Nacional Autónoma de México, Mexico City, Mexico; LACM, Natural History Museum of Los Angeles County, Los Angeles, USA.
Material and methods
The specimen FCBUANL_2711 was collected by a team of the Laboratorio de Paleobiología, FCB UANL, during 1992 near the Ejido Presa San Antonio, located in the Parras de la Fuente municipality, Coahuila state, northeastern Mexico (Fig. 1). The specimen was deposited in the Colección Paleobiológica of the UANL Facultad de Ciencias Biológicas. The preparation process has been slow due to space restrictions. Mechanical techniques have been used for this purpose with tools as electric engravers and brushes. The present material was photographed with a Canon EOS Rebels XT camera and Canon 28 mm lens and Canon PowerShot SX280 HS.
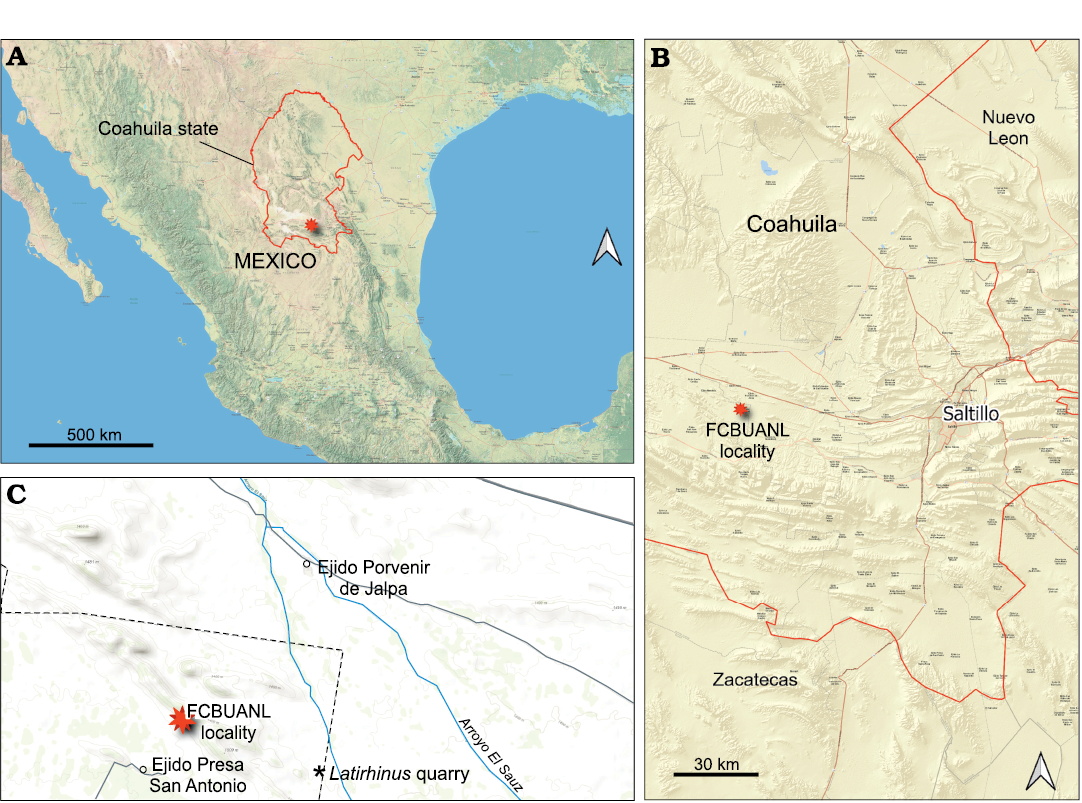
Fig. 1. Map showing location of the Coahuila state (A) and FCBUANL locality (B) where studied fossils were found. Detailed map indicating the principal localities of Cerro del Pueblo Formation (C), FCBUANL_2711 locality represented by red star.
Systematic palaeontology
Dinosauria Owen, 1842
Ornithischia Seeley, 1887
Ornithopoda Marsh, 1881
Hadrosauridae Cope, 1870
Lambeosaurinae Parks, 1923
Lambeosaurinae indet.
Material.—FCBUANL_2711, left ilium, right pubis, right ischium, partial sacrum, distal end of left femur, right and left tibiae, II and IV right metatarsals, one cervical and 21 caudal vertebrae (Fig. 2); from Near Ejido Presa San Antonio, Parras de la Fuente municipality, Coahuila, Mexico. The material was recovered from a thick lodolite layer dominated by the gastropod Tympanotonus nodosa with thin sandstone intercalations, at Cerro del Pueblo Formation, upper Campanian.
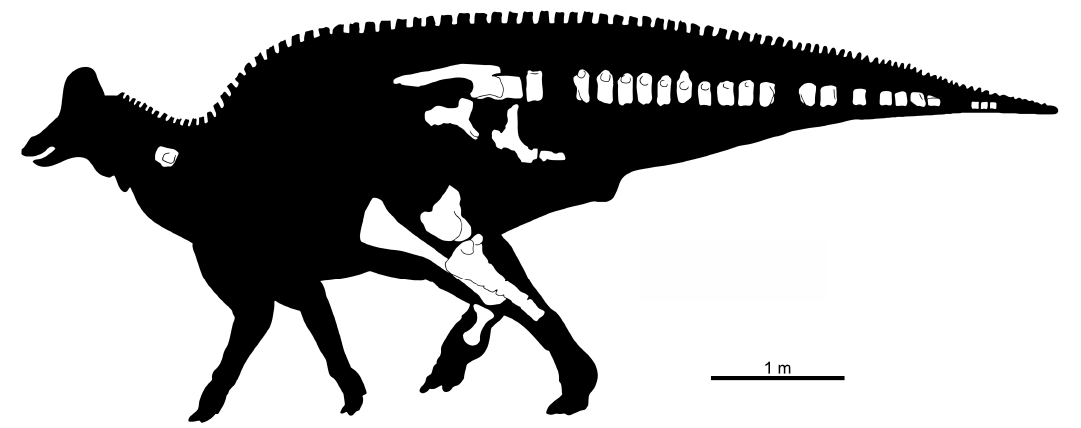
Fig. 2. Skeletal composition of the FCBUANL_2711. The silhouette was obtained from phylopic.org vectorized by T. Michael Keesey from the work of Dmitry Bogdanov.
Description.—Ilium: Only the left ilium is preserved, missing the posterior region, including the supracetabular crest. The length of the preserved portion is 780 mm (Fig. 3A1). The preacetabular process is thick and shows a deep projection close to the anterior end, extruding posteriorly in the direction of the tip. The lateral surface is flattened and in dorsal view it shows a slight concavity with respect to the lateral plane (Fig. 3A2). This process is 597 mm in length and shows a minimal degree of ventral deflection (164°). The ratio between the length of the preacetabular process and the length of the acetabulum is 3.14. Although the length measurements seem to indicate that this process has a considerable elongation compared to the ilium, it is not possible to confirm this assumption this without knowing the proportions of the postacetabular process.
Pubis: The pubis (Fig. 3B) has a wide and short pubic blade. The dorsal margin starts to project notably dorsally at the beginning of the prepubic process, while the ventral margin does the same in the ventral direction. Due to the absence of the dorsal and ventral margins of the distal portion of the prepubic process, the shape of this region cannot be determined. However, this process is hypothesized to be short, based on its distal margin. The iliac peduncle is massive and wide, with a triangular section and mediodorsally expanded in the distal region. The distal end of the ischiadic peduncle is absent, and the proximal region shows a protuberance at the base. The distal region of the postpubic process is not preserved and the base is slightly ventrally projected. The acetabular margin is concave and an angle of 134° is observed between the portion of the margin that corresponds to the iliac peduncle and the margin of the ischial peduncle, giving the appearance of an abrupt change of direction in the margin.
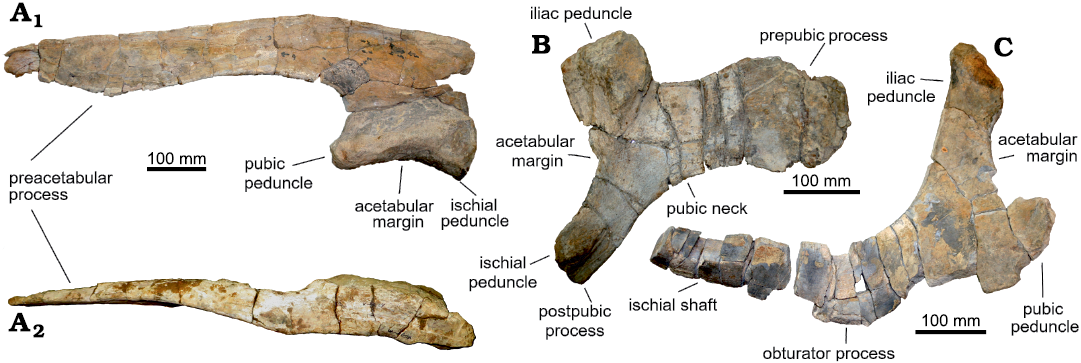
Fig. 3. Lambeosaurinae indet., pelvic gridle elements of FCBUANL_2711, from Ejido Presa San Antonio, Coahuila, Mexico, upper Campanian. A. Left ilium in lateral (A1) and dorsal (A2) views. B. Right pubis in lateral view. C. Right ischium in lateral view.
Ischium: Only the proximal region of the right ischium is present, and it is poorly preserved (Fig. 3C). The iliac peduncle is strongly dorsally projected, giving a longer appearance in comparison with the pubic peduncle, which is eroded. The dorsal and ventral margins of the iliac peduncle are convergent. The dorsal one is nearly straight from the base to the tip of the iliac peduncle, and the vertex between dorsal and articular margin is slightly dorsally projected. Only the base of the obturator process is preserved, and it is strongly eroded. The ischiadic shaft shows a clockwise torsional deformation at the base.
Cervical vertebra: Only one cervical vertebra was collected (Fig. 4A). It presents a markedly opisthocoelus condition with a length close to 110 mm. The centrum is wider than high, and the neural canal is wide. There is no neural arch associated with this vertebra and both transverse processes are short and centrally located.
Sacrum: The sacrum (Fig. 4D) is preserved as a single fragmented block composed of 3 sacral vertebrae. It has an approximate length of 350 mm. The last centrum is as tall as wide. Dorsally, it has a neural arch and associated spines. The ventral side is heavily eroded.
Caudal vertebrae: 21 elements are preserved, eight proximal, five middle and eight distal caudal vertebrae (Fig. 4B, C, E, F). Table 1 lists the measurements of the caudal vertebrae. The proximal caudal vertebrae (Fig. 4C) have amphicoelus centra, rectangular to pentagonal in anterior view, being taller than wide and compressed craniocaudally. The neural canal is relatively narrow. Vascular foramina are present in the lateral sides of the centra. In the first vertebra the transverse processes on the centrum are eroded and missing, whereas the following vertebrae present them on the upper lateral part. Also, the articular surfaces of the chevrons are absent on the first vertebra, but they are present on the following elements. The neural arches and spines are present however they are still under preparation. The middle caudal vertebrae (Fig. 4B) are smaller than the proximal caudal vertebrae and hexagonal in anterior view. The articular surfaces of the chevrons are slightly projected ventrally. The centra are less compressed than the proximal vertebrae, maintaining the amphicoelus centra and the narrow neural canal. The distal caudal vertebrae (Fig. 4E) have the smallest centra with cubic proportions. They lack the transverse processes, and the articular surfaces of the chevrons are completely reduced.

Fig. 4. Vertebrae of Lambeosaurinae indet.(FCBUANL_2711) from Ejido Presa San Antonio, Coahuila, Mexico, upper Campanian. A. Cervical vertebrae, in cranial (A1), lateral (A2), and caudal (A3) views. B. Middle caudal vertebra, in lateral (B1) and cranial (B2) views. C. Proximal caudal vertebra, in cranial (C1) and lateral (C2) views. D. Sacrum in ventral view. E. Distal caudal vertebra in cranial (E2) and lateral (E2) views. F. Caudal vertebrae in lateral view.
Table 1. Measurements (in mm) of the caudal vertebrae centra of Lambeosaurinae indet. FCBUANL_2711. Element numbers are tentative and do not consider the missing vertebrae.
|
Element |
Height |
Width |
Length |
|
1 |
172 |
194 |
83 |
|
2 |
182 |
179 |
98 |
|
3 |
187 |
166 |
91 |
|
4 |
177 |
176 |
93 |
|
5 |
178 |
157 |
91 |
|
6 |
174 |
142 |
85 |
|
7 |
165 |
152 |
82 |
|
8 |
155 |
144 |
95 |
|
9 |
152 |
120 |
98 |
|
10 |
141 |
135 |
92 |
|
11 |
147 |
120 |
92 |
|
12 |
123 |
129 |
96 |
|
13 |
119 |
114 |
101 |
|
14 |
113 |
84 |
79 |
|
15 |
91 |
83 |
84 |
|
16 |
81 |
85 |
88 |
|
17 |
96 |
90 |
88 |
|
18 |
? |
80 |
91 |
|
19 |
51 |
63 |
55 |
|
20 |
54 |
66 |
52 |
|
21 |
47 |
50 |
58 |
Femur: Only the distal end of the left femur is preserved (Fig. 5A2). The cranial portion is absent, the outer condyle is smaller and more lateromedially compressed than the inner condyle. The latter is more caudally projected, the intercondylar groove is narrow distally, widening proximally (Fig. 5A1).
Tibiae: Both tibiae are present (Fig. 5B, C). However, the left one is missing the distal end and the shaft is transversely fragmented multiple times (Fig. 5B). The right tibia is complete (Fig. 5C), with minor eroded areas and a total length of about 1300 mm. Both tibiae have a wide craniocaudal expansion at their proximal ends and the inner condyle is larger than the lateral condyle, forming a well-defined intercondylar groove. The cnemial crests are eroded on both tibiae. The central shaft of the tibia is sub-cylindrical and very thin compared to the width of the proximal end. In the right tibia the lateral malleolus is smaller than the medial malleolus.
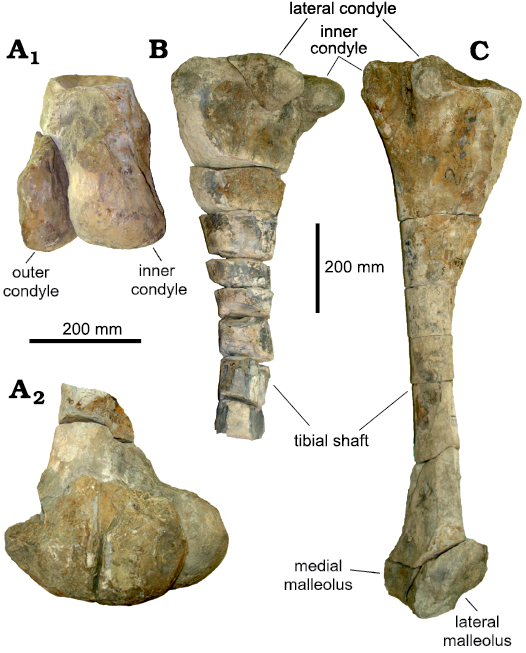
Fig. 5. Hindlimb elements of Lambeosaurinae indet. (FCBUANL_2711) from Ejido Presa San Antonio, Coahuila, Mexico, upper Campanian. A. Distal end of the left femur, in caudal (A1) and lateral (A2) views. B. Left tibia in lateral view. C. Right tibia in lateral view.
Metatarsals: The right metatarsal IV (Fig. 6B) measures about 330 mm in length. Near mid-length, the bone is strongly deflected laterally. The proximal surface is markedly concave. The distal surface is heavily eroded on the sides. The right metatarsal II (Fig. 6A) is much better preserved than the previous one, with a length of approximately 340 mm. The proximal surface is flat, unlike metatarsal IV, and expanded dorsoplantarly. A medial tuberosity is located above the distal end of metatarsal II, where the bone curves medially.

Fig. 6. Metatarsals of Lambeosaurinae indet. (FCBUANL_2711) from Ejido Presa San Antonio, Coahuila, Mexico, upper Campanian. A. Metatarsus II, in dorsal (A1) and lateral (A2) views. B. Metatarsus IV, in dorsal (B1) and lateral (B2) views.
Discussion
Comparisons with Latirhinus and Velafrons.—Latirhinus uitstlani Prieto-Márquez and Serrano Brañas, 2012 is the only species described so far with postcranial material from the Cerro del Pueblo Formation (Prieto-Márquez and Serrano Brañas 2012). The ilium of FCBUANL_2711 (Fig. 7A) differs markedly from L. uitstlani IGM 6583 (Fig. 7B) by presenting a sigmoidal curvature in the ventral margin of the preacetabular process, which narrows anteriorly. The distal end of the iliac peduncle of one of the ischias is also assigned to IGM 6583, but due to its fragmentary condition, a direct comparison is not possible.
The holotype of the hadrosaurid Velafrons coahuilensis (CPC 59) includes the right ilium, left pubis and both ischia, but these elements were never described. The ilium of CPC 59 (Fig. 7C) has a total length of 690 mm and shows a wide and straight preacetabular process in lateral view (Fig. 8A1), whereas in dorsal view this process is strongly craniolaterally curved (Fig. 8A2). The preacetabular notch is wide and the ischiadic peduncle is not strongly projected cranially. In contrast, the preacetabular process of FCBUANL_2711 is concave in dorsal view with respect to the lateral axis but it does not present a deformation as marked as in CPC 59. Furthermore, the preacetabular notch is narrower, highlighting the cranial projection of the pubic peduncle, which is more robust than in CPC 59. The ilium of FCBUANL_2711, despite being incomplete, is longer than the ilium of CPC 59, which has been previously regarded as a juvenile specimen (Gates et al. 2007). Both specimens show a ventral deflection of the preacetabular process greater than 150°, 153° in CPC 59 and 164° in FCBUANL_2711. According to Brett-Surman and Wagner (2007), this character is highly variable, even between the two ilia of the same individual, therefore a difference of 10° of ventral deflection is not sufficient for the differentiation of FCBUANL_2711 from V. coahuilensis. The designation of FCBUANL_2711 as an adult specimen of V. coahuilensis is possible, however Prieto-Márquez (2014a) indicates that in Edmontosaurus annectens Marsh, 1892, there is an ontogenetic change in the ventral deflection of the preacetabular process of the ilium. In juveniles it is slightly deflected (157° to 165°) whereas adults shows a more pronounced deflection (138° to 147°) (Prieto-Marquez 2014a). A more detailed study of this character in other species is necessary to determine whether ontogenetic variation of the deflection of this process is generalized among hadrosaurids, or if it is only present in E. annectens.
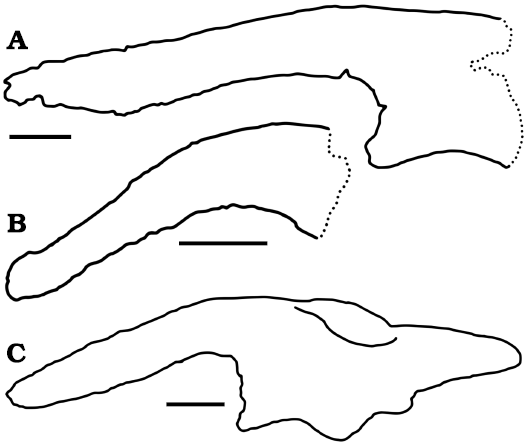
Fig. 7. Comparision of ilia of hadrosaurids from Cerro del Pueblo Formation. A. Lambeosaurinae indet., FCBUANL_2711. B. Latirhinus uitstlani (Prieto-Márquez and Serrano Brañas 2012). C. Velafrons coahuilensis, CPC 59 (Gates et al. 2007). Scale bars 100 mm.

Fig. 8. Ilium of hadrosaurid dinosaur Velafrons coahuilensis Gates, Sampson, Delgado de Jesús, Zanno, Eberth, Hernandez-Rivera, Aguillón-Martínez, and Kirkland, 2007 (CPC 59) from Rincón Colorado, Coahuila, Mexico, upper Campanian; in lateral (A1) and dorsal (A2) views.
The left pubis of CPC 59 (Fig. 9B) shows the common morphology of lamebosaurines, presenting a high iliac peduncle with triangular cross-section and a short prepubic constriction. The prepubic process is incompletely preserved. The margins of the prepubic constriction are divergent, and the dorsal margin arises dorsally without reaching the level of the proximal end of the iliac peduncle. The pubis of FCBUANL_2711 and that of CPC 59 are similar, differing in the more robust iliac peduncle in the former and the slightly narrower prepubic constriction in CPC 59.
CPC 59 preserves both complete ischia (Fig. 9A, C), characterized by the “thumb-like” shape of the dorsal and articular margin of the iliac peduncle. The distal end of the ischiadic shaft ends in the typical lambeosaurine “foot” shape, but it lacks the “heel”, a feature also absent in species of Corythosaurus and Lambeosaurus (Brett-Surmann 1989). The ischium is the least well preserved element of the three pelvic bones of FCBUANL_2711, showing the “thumb like” iliac peduncle as well, although erosion reduces the visibility of this character. Both the pubic peduncle, the obturator process, and the notch are not well preserved, preventing a more extensive comparison with the material assigned to Velafrons. The absence of the distal end of the ischial shaft precludes a more accurate assignment of this material to Lambeosaurinae, as well as its differentiation from V. coahuilensis.
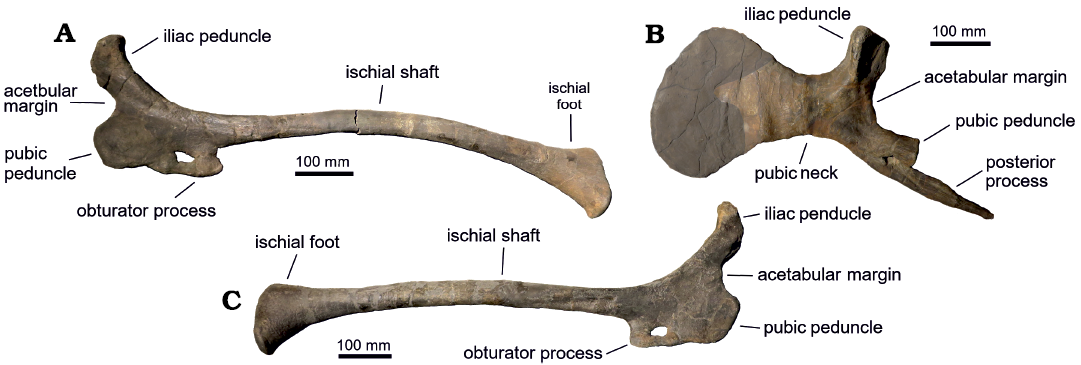
Fig. 9. Pelvic gridle elements of hadrosaurid dinosaur Velafrons coahuilensis Gates, Sampson, Delgado de Jesús, Zanno, Eberth, Hernandez-Rivera, Aguillón-Martínez, and Kirkland, 2007 (CPC 59) from Rincón Colorado, Coahuila, Mexico, upper Campanian. A. Left ischium in lateral view. B. Left pubis in lateral view, shading indicates the reconstructed area. C. Right ischium in lateral view.
Comparisons with other hadrosaurids.—The angle of ventral deflection of the ilium greater than 150°, present in FCBUANL_2711 and Velafrons coahuilensis, is also present in the material tentatively referred to the Asian hadrosaurine Kundurosaurus nagornyi Godefroit, Bolotsky, and Lauters, 2012b, AENM 2/922-7L (Godefroit et al. 2012b), relegated as a junior synonym of Kerberosaurus manakini Bolotsky and Godefroit, 2004, by Xing et al. (2014), in the south American hadrosaurine Bonapartesaurus rionegrensis Cruzado-Caballero and Powell, 2017 (Cruzado-Caballero and Powell 2017), and the Asian lambeosaurine Sahaliyania elunchunorum Godefroit, Shulin, Tingxiang, and Lauters, 2008 (Godefroit et al. 2008). The ilium of FCBUANL_2711 differs from K. manakini by having a wide preacetabular process, a notable characteristic in lambeosaurines (Brett-Surman 1989). The preacetabular process of the ilium also shows a ventral projection close to the cranial end where it narrows distally, as in other lambeosaurine specimens like FMNH P27393 Parasaurolophus cyrtocristatus Ostrom, 1961a (Ostrom 1961a), LACM 20874 Magnapaulia laticaudus Morris, 1981 (Prieto-Márquez et al. 2012) and AEHM 2/846 Olorotitan arharensis Godefroit, Bolotsky, and Alifanov, 2003 (Godefroit et al. 2012a). The ilium shows a similar morphology to the GMH W103 an ilium referred to S. elunchunorum (Godefroit et al. 2008), but the preacetabular notch of FCBUANL_2711 is narrower than in GMH W103. The absence of the supracetabular crest and postacetabular process prevents a more extensive comparison, since in these regions possess important diagnostic characters for the clade (Prieto-Márquez 2010b).
The pubis of FCBUANL_2711 resembles the V morphotype proposed by Brett-Surmann (1989) which corresponds to the Parasaurolophini clade, while differs from the morphotypes of the Hadrosaurinae by having a short and wide prepubic constriction. Despite the incomplete condition, the prepubic process is similar to the morphology of the Hypacrosaurus stebingeri Horner and Currie, 1994, pubis (see Guenther 2009: fig. 10c). The material referred to S. elunchunorum includes a left pubis (GMH W179) that differs from FCBUANL_2711 in having an almost straight ventral margin of the prepubic process, resulting in a more elongated appearance of the bone, similar to the V morphotype of Brett-Surmann (1989). The pubis of GMH W179 bears a robust iliac peduncle, although to a lesser extent than in FCBUANL_2711. The incomplete condition of the prepubic process complicates a better systematic resolution, but the recognized features suggest lambeosaurine affinities.
Conclusions
The hadrosaurid specimen described in this paper shows lambeosaurine affinities, therefore distinct from Kritosaurus navajovius and Latirhinus uitstlani, both present in the Cerro del Pueblo Formation. The differentiation from to Velafrons coahuilensis is not clear; because FCBUANL_2711 is a large individual, it is possible that it is a fully developed adult individual of this species, but an osteohistological analysis would be necessary to confirm this. The lack of cranial elements in FCBUANL_2711 prevents the assignment of the specimen to V. coahuilensis or to a different species. Therefore, this specimen is referred only as Lambeosaurinae indet. Paleontological studies of this clade, along with other groups of dinosaurs, have increased in recent years in Mexico, but several other materials need to be studied from other geological formations of Campanian–Maastrichtian Mexico. This will allow a better understanding of the terrestrial environments during these ages in southern Laramidia.
Acknowledgements
We thank Claudia Hernández Pérez, Rogelio Reyna de Labra, and José Martínez (Monterrey, Mexico) for their collaboration and help with the photographs and assistance in the preparation of the figures. Thanks to the reviewer Filippo Bertozzo (Queen’s University Belfast, UK) and the anonymous reviewer for their detailed and helpful comments on the manuscript. We would like to thank Erick C. Oñate-González and Hidalgo Rodríguez-Vela (both Universidad Autónoma De Nuevo León, Mexico) for their reviews and the helpful comments. Financial support for the field work was provided by Programa de Apoyo a la Investigación Científica y Tecnológica (PAICYT) Proyecto N° CN1181-11 and Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León.
References
Bolotsky, Y.L. and Godefroit, P. 2004. A new hadrosaurine dinosaur from the Late Cretaceous of Far Eastern Russia. Journal of Vertebrate Paleontology 24: 351–365. Crossref
Brett-Surman, M.K. 1989. A Revision of the Hadrosauridae (Reptilia: Ornithischia) and Their Evolution During the Campanian and Maastrichtian. 272 pp. Unpublished Ph.D. Thesis, George Washington University, Washington D.C.
Brett-Surman, M.K. and Wagner, J.R. 2007. Discussion of character analysis of the appendicular anatomy in Campanian and Maastrichtian North American hadrosaurids—variation and ontogeny. In: K. Carpenter (ed.), Horns and Beaks, 135–170. Indiana University Press, Bloomington. Crossref
Brinkman, D.B. 2014. Turtles of the Mesozoic of Mexico. In: H.E. Rivera-Sylva, K. Carpenter, and E. Frey (eds.), Dinosaurs and Other Reptiles from the Mesozoic of Mexico, 30–43. Indiana University Press, Bloomington.
Brown, B. 1910. The Cretaceous Ojo Alamo beds of New Mexico with description of the new dinosaur genus Kritosaurus. Bulletin of the American Museum of Natural History 28: 267–274.
Campione, N.E. 2014. Postcranial anatomy of Edmontosaurus regalis (Hadrosauridae) from the Horseshoe Canyon Formation, Alberta, Canada. In: D.A. Eberth, D.C. Evans, and P.E. Ralrick (eds.), Hadrosaurs, 208–244. Indiana University Press, Bloomington.
Cevallos-Ferriz, S.R.S. and Vázquez-Rueda, M.A. 2016. Permineralized infructescence from the Cerro del Pueblo Formation, Upper Cretaceous, Coahuila, Mexico. Botany 94: 773–785. Crossref
Cifuentes-Ruiz, P., Vršanský, P., Vega, F.J., Cevallos-Ferriz, S.R.S., González-Soriano, E., and Delgado de Jesús, C.R. 2006. Campanian terrestrial arthropods from the Cerro del Pueblo Formation, Difunta Group in northeastern Mexico. Geologica Carpathica 57: 347–354.
Cope, E.D. 1870. Synopsis of the extinct Batrachia and Reptilia of North America. Transactions of the American Philosophical Society 14: 1–252. Crossref
Cruzado-Caballero, P. and Powell, J. 2017. Bonapartesaurus rionegrensis, a new hadrosaurine dinosaur from South America: implications for phylogenetic and biogeographic relations with North America. Journal of Vertebrate Paleontology 37: e1289381. Crossref
Estrada-Ruiz, E. and Cevallos-Ferriz, S.R.S. 2007. Infructescences from the Cerro del Pueblo Formation (Late Campanian), Coahuila, and El Cien Formation (Oligocene–Miocene), Baja California Sur, Mexico. International Journal of Plant Sciences 168: 507–519. Crossref
Estrada-Ruiz, E., Calvillo-Canadell, L., and Cevallos-Ferriz, S.R.S. 2009. Upper Cretaceous aquatic plants from Northern Mexico. Aquatic Botany 90: 282–288. Crossref
Gates, T.A., Sampson, S.D., Delgado de Jesús, C.R., Zanno, L.E., Eberth, D., Hernandez-Rivera, R., Aguillón-Martínez, M.C., and Kirkland, J.I. 2007. Velafrons coahuilensis, a new lambeosaurine hadrosaurid (Dinosauria: Ornithopoda) from the Late Campanian Cerro del Pueblo Formation, Coahuila, Mexico. Journal of Vertebrate Paleontology 27: 917–930. Crossref
Godefroit, P., Bolotsky, Y.. and Alifanov, V. 2003. A remarkable hollow-crested hadrosaur from Russia: An Asian origin for lambeosaurines. Comptes Rendus Palevol 2: 143–151. Crossref
Godefroit, P., Bolotsky, Y.L., and Bolotsky, I.Y. 2012a. Osteology and relationships of Olorotitan arharensis, a hollow-crested hadrosaurid dinosaur from the latest Cretaceous of far eastern Russia. Acta Palaeontologica Polonica 57: 527–560. Crossref
Godefroit, P., Bolotsky, Y.L., and Lauters, P. 2012b. A new saurolophine dinosaur from the Latest Cretaceous of far eastern Russia. PLoS ONE 7 (5): e36849. Crossref
Godefroit, P., Shulin, H., Tingxiang, Y., and Lauters, P. 2008. New hadrosaurid dinosaurs from the uppermost Cretaceous of northeastern China. Acta Palaeontologica Polonica 53: 47–74. Crossref
Guenther, M.F. 2009. Influence of sequence heterochrony on hadrosaurid dinosaur postcranial development. Anatomical Record 292: 1427–1441. Crossref
Horner, J.R., Weishampel, D.B., and Foster, C.A. 2004. Hadrosauridae. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria, 438–463. University of California Press, Berkeley. Crossref
Kirkland, J.I., Hernández Rivera, R., Aguillón Martínez, M.C., Delgado de Jesús, C.R., Gómez Nuñez, R., and Vallejo, I. 2000. The Late Cretaceous Difunta Group of the Parras Basin, Coahuila, Mexico, and its vertebrate fauna. In: Society of Vertebrate Paleontology Annual Meeting 2000 Field Trip Book, 133–172. UNAM, Mexico City.
Kirkland, J.I., Hernández-Rivera, R., Gates, T., Paul, G.S., Nesbitt, S., Serrano-Brañas, C.I., and Garcia-de la Garza, J.P. 2006. Large hadrosaurine dinosaurs from the Latest Campanian of Coahuila, Mexico. New Mexico Museum of Natural History and Science Bulletin 35: 299–315.
Loewen, M.A., Sampson, S.D., Lund, E.K., Farke, A.A., Aguillón-Martínez, M.C., de León, C.A., Rodríguez-de la Rosa, R.A., Getty, M.A., and Eberth, D.A. 2010. Horned dinosaurs (Ornithischia: Ceratopsidae) from the Upper Cretaceous (Campanian) Cerro del Pueblo Formation, Coahuila, Mexico. In: M.J. Ryan, B.J. Chinnery-Allgeier, D.A. Eberth, and P.E. Ralrick (eds.), New Perspectives on Horned Dinosaurs, 99–116. Indiana University Press, Bloomington.
Longrich, N.R., Suberbiola, X.P., Pyron, R.A. and Jalil, N.-E. 2021. The first duckbill dinosaur (Hadrosauridae: Lambeosaurinae) from Africa and the role of oceanic dispersal in dinosaur biogeography. Cretaceous Research 120: 104678. Crossref
Lull, R.S. and Wright, N.E. 1942. Hadrosaurian dinosaurs of North America. Geological Society of America Special Papers 40: 1–272. Crossref
Lund, E.K. and Gates, T.A. 2006. A historical and biogeographical examination of hadrosaurian dinosaurs. New Mexico Museum of Natural History and Science Bulletin 35: 263–276.
Marsh, O.C. 1881. Principal characters of American Jurassic dinosaurs. Part IV. American Journal of Science 21: 167–170. Crossref
Marsh, O.C. 1892. Notice of new reptiles from the Laramie Formation. American Journal of Science 43: 449–453. Crossref
Morris, W.J. 1970. Hadrosaurian dinosaur bills-morphology and function. Contributions in Science 193: 1–14.
Morris, W.J. 1981. A new species of hadrosaurian dinosaur from the Upper Cretaceous of Baja California—?Lambeosaurus laticaudus. Journal of Paleontology 55: 453–462.
Nabavizadeh, A. 2014. Hadrosauroid jaw mechanics and the functional significance of the predentary bone. In: D.A. Eberth, D.C. Evans, and P.E. Ralrick (eds.), Hadrosaurs, 467–483. Indiana University Press, Bloomington.
Nabavizadeh, A. and Weishampel, D.B. 2016. The predentary bone and its significance in the evolution of feeding mechanisms in ornithischian dinosaurs. The Anatomical Record 299: 1358–1388. Crossref
Ostrom, J.H. 1961a. A new species of hadrosaurian dinosaur from the Cretaceous of New Mexico. Journal of Paleontology 35: 575–577.
Ostrom, J.H. 1961b. Cranial morphology of the hadrosaurian dinosaurs of North America. Bulletin of the American Museum of Natural History 122: 33–186.
Owen, R. 1842. Report on British fossil reptiles. Part II. Reports of the British Association for the Advancement of Science 11: 60–204.
Parks, W.A. 1923. Corythosaurus intermedius, a new species of trachodont dinosaur. University of Toronto Studies, Geological Series 15: 5–57.
Prieto-Márquez, A. 2010a. Global historical biogeography of hadrosaurid dinosaurs. Zoological Journal of the Linnean Society 159: 503–525. Crossref
Prieto-Márquez, A. 2010b. Global phylogeny of hadrosauridae (Dinosauria: Ornithopoda) using parsimony and bayesian methods. Zoological Journal of the Linnean Society 159: 435–502. Crossref
Prieto-Márquez, A. 2014a. A juvenile Edmontosaurus from the late Maastrichtian (Cretaceous) of North America: Implications for ontogeny and phylogenetic inference in saurolophine dinosaurs. Cretaceous Research 50: 282–303. Crossref
Prieto-Márquez, A. 2014b. Skeletal morphology of Kritosaurus navajovius (Dinosauria: Hadrosauridae) from the Late Cretaceous of the North American south-west, with an evaluation of the phylogenetic systematics and biogeography of Kritosaurini. Journal of Systematic Palaeontology 12: 133–175. Crossref
Prieto-Márquez, A. and Serrano Brañas, C.I. 2012. Latirhinus uitstlani, a “broad-nosed” saurolophine hadrosaurid (Dinosauria, Ornithopoda) from the late Campanian (Cretaceous) of northern Mexico. Historical Biology 24: 607–619. Crossref
Prieto-Márquez, A., Chiappe, L.M., and Joshi, S.H. 2012. The lambeosaurine dinosaur Magnapaulia laticaudus from the Late Cretaceous of Baja California, northwestern Mexico. PLoS ONE 7 (6): e38207. Crossref
Ramírez-Velasco, A.A. and Hernández-Rivera, R. 2015. Diversity of Late Cretaceous dinosaurs from Mexico. Boletín Geológico y Minero 126: 63–108.
Ramírez-Velasco, A.A., Hernández-Rivera, R., and Servin-Pichardo, R. 2014. The hadrosaurian record from Mexico. In: D.A. Eberth, D.C. Evans, and P.E. Ralrick (eds.), Hadrosaurs, 340–360. Indiana University Press, Bloomington.
Rivera-Sylva, H.E. and Carpenter, K. 2014. The ornithischian dinosaurs of Mexico. In: H.E. Rivera-Sylva, K. Carpenter, and E. Frey (eds.), Dinosaurs and Other Reptiles from the Mesozoic of Mexico, 156–180. Indiana University Press, Bloomington.
Rivera-Sylva, H.E. and Espinosa-Chávez, B. 2006. Ankylosaurid (Dinosauria: Thyreophora) osteoderms from the Upper Cretaceous Cerro del Pueblo Formation of Coahuila, Mexico. Carnets de Géologie (Notebooks on Geology) 2006: CG2006_L02. Crossref
Rivera-Sylva, H.E., Barrón-Ortíz, C.I., Vivas González, R., Nava Rodríguez, R.L., Guzmán-Gutiérrez, J.R., Cabral Valdez, F., and de León Dávila, C. 2019a. Preliminary assessment of hadrosaur dental microwear from the Cerro del Pueblo Formation (Upper Cretaceous: Campanian) of Coahuila, northeastern Mexico. Paleontología Mexicana 8 (1): 17–28.
Rivera-Sylva, H.E., Frey, E., Schulp, A.S., Meyer, C., Thüring, B., Stinnesbeck, W., and Vanhecke, V. 2017. Late Campanian theropod trackways from Porvenir de Jalpa, Coahuila, Mexico. Palaeovertebrata 41 (2): e1. Crossref
Rivera-Sylva, H.E., Frey, E., Stinnesbeck, W., Amezcua, N., and Flores Huerta, D. 2018. First occurrence of Parksosauridae in Mexico, from the Cerro del Pueblo Formation (Late Cretaceous; late Campanian) at Las águilas, Coahuila. Boletín de la Sociedad Geológica Mexicana 70: 779–784. Crossref
Rivera-Sylva, H.E., Frey, E., Stinnesbeck, W., Amezcua Torres, N., and Flores Huerta, D. 2019b. Terrestrial vertebrate paleocommunities from the Cerro del Pueblo Formation (Late Cretaceous; late Campanian) at Las Aguilas, Coahuila, Mexico. Palaeovertebrata 42 (2): e1. Crossref
Rodríguez de la Rosa, R.A. 2007. Hadrosaurian footprints from the Late Cretaceous Cerro del Pueblo Formation of Coahuila, Mexico. In: E. Díaz-Martínez and I. Rábano (eds.), 4th European Meeting on the Palaeontology and Stratigraphy of Latin America. Cuadernos Del Museo Geominero No. 8, 339–343. Instituto Geológico y Minero de España, Madrid.
Rodriguez-de la Rosa, R.A. and Cevallos-Ferriz, S.R.S. 1994. Upper Cretaceous zingiberalean fruits with in situ seeds from southeastern Coahuila, Mexico. International Journal of Plant Sciences 55: 786–805. Crossref
Rodriguez-de la Rosa, R.A. and Cevallos-Ferriz, S.R.S. 1998. Vertebrates of the El Pelillal locality (Campanian, Cerro Del Pueblo Formation), southeastern Coahuila, Mexico. Journal of Vertebrate Paleontology 18: 751–764. Crossref
Rybakiewicz, S., Rivera-Sylva, H.E., Stinnesbeck, W., Frey, E., Guzmán-Gutiérrez, J.R., Vivas González, R., Nava Rodríguez, R.L., and Padilla-Gutiérrez, J.M. 2019. Hadrosaurs from Cañada Ancha (Cerro del Pueblo Formation; upper Campanian–?lower Maastrichtian), Coahuila, northeastern Mexico. Cretaceous Research 104: 104199. Crossref
Seeley, H.G. 1887. Researches on the structure, organization, and classification of the fossil reptilia. I. On Protorosaurus speneri (von Meyer). Philosophical Transactions of the Royal Society of London B 178: 187–213. Crossref
Serrano-Brañas, C.I. and Espinosa-Chávez, B. 2017. Taphonomic history of a “duck-bill” dinosaur (Dinosauria: Ornithopoda) from the Cerro del Pueblo Formation (Upper Cretaceous, Campanian) Coahuila, Mexico: Preservational and paleoecological implications. Cretaceous Research 74: 165–174. Crossref
Serrano-Brañas, C.I., Espinosa-Chávez, B., and Maccracken, S.A. 2018a. Gastrochaenolites Leymerie in dinosaur bones from the Upper Cretaceous of Coahuila, north-central Mexico: Taphonomic implications for isolated bone fragments. Cretaceous Research 92: 18–25. Crossref
Serrano-Brañas, C.I., Espinosa-Chávez, B., and Maccracken, S.A. 2018b. Insect damage in dinosaur bones from the Cerro del Pueblo Formation (Late Cretaceous, Campanian) Coahuila, Mexico. Journal of South American Earth Sciences 86: 353–365. Crossref
Serrano-Brañas, C.I., Espinosa-Chávez, B., and Maccracken, S.A. 2019. Teredolites trace fossils in log-grounds from the Cerro del Pueblo Formation (Upper Cretaceous) of the state of Coahuila, Mexico. Journal of South American Earth Sciences 95: 102316. Crossref
Serrano-Brañas, C.I., Espinosa-Chávez, B., Maccracken, S.A., Gutiérrez-Blando, C., de León-Dávila, C., and Flores Ventura, J. 2020. Paraxenisaurus normalensis, a large deinocheirid ornithomimosaur from the Cerro del Pueblo Formation (Upper Cretaceous), Coahuila, Mexico. Journal of South American Earth Sciences 101: 102610. Crossref
Serrano-Brañas, C.I., Hernández-Rivera, R., Torres-Rodríguez, E., and Espinosa Chávez, B. 2006. A natural hadrosaurid endocast from the Cerro del Pueblo Formation (Upper Cretaceous) of Coahuila, Mexico. New Mexico Museum of Natural History and Science Bulletin 35: 317–322.
Wolleben, J.A. 1977. Paleontology of the Difunta Group (Upper Cretaceous–Tertiary) in Northern Mexico. Journal of Paleontology 51: 373–398.
Xing, H., Zhao, X., Wang, K., Li, D., Chen, S., Mallon, J.C., Zhang, Y., and Xu, X. 2014. Comparative osteology and phylogenetic relationship of Edmontosaurus and Shantungosaurus (Dinosauria: Hadrosauridae) from the Upper Cretaceous of North America and East Asia. Acta Geologica Sinica (English Edition) 88: 1623–1652. Crossref
Acta Palaeontol. Pol. 66 (Supplement to 3): 101–110, 2021
https://doi.org/10.4202/app.00831.2020