
Late Permian ichthyofauna from the North-Sudetic Basin, SW Poland
DARJA DANKINA, ANDREJ SPIRIDONOV, PAWEŁ RACZYŃSKI, and SIGITAS RADZEVIČIUS
Dankina, D., Spiridonov, A., Raczyński, P., and Radzevičius, S. 2021. Late Permian ichthyofauna from the North-Sudetic Basin, SW Poland. Acta Palaeontologica Polonica 66 (Supplement to 3): 47–57.
The late Permian time was a transformative period before the most severe mass extinction known. Even though fishes constitute a key component of marine ecosystems since the Silurian, their biogeographic patterns during the late Permian are currently insufficiently known. The new ichthyofaunal material described here comes from the southeastern part of the Zechstein Basin, from the calcareous storm sediments alternating with marls, which were deposited in less energetic conditions. Chondrichthyans and osteichthyans are reported here for the first time from the Nowy Kościół quarry in the SW Poland. The assemblage consists of various euselachian dermal denticles, actinopterygian scales and teeth, and isolated hybodontoid tooth putatively assigned as extremely rare ?Gansuselache sp. from the Permian. The diverse actinopterygian tooth shapes show significant ecological differentiation of fishes exploring sclerophagous, durophagous, and herbivory modes of feeding in the given part of the Zechstein Basin suggesting the presence of complex ecosystems even in hyper-saline conditions of an epicontinental sea.
Key words: Euselachii, Actinopterygii, teeth, scales, trophic groups, Permian, Zechstein, Poland.
Darja Dankina [darja.dankina@gmail.com], Andrej Spiridonov [s.andrej@gmail.com], and Sigitas Radzevičius [sigitas.radzevicius@gf.vu.lt], Department of Geology and Mineralogy, Vilnius University, M.K. Čiurlionio st. 21/27, LT03101, Vilnius, Lithuania.
Paweł Raczyński [pawel.raczynski@uwr.edu.pl], Department of Physical Geology, Institute of Geological Sciences, University of Wrocław, Pl. Maksa Borna 9, 50-205 Wrocław, Poland.
Received 14 October 2020, accepted 5 December 2020, available online 2 June 2021.
Copyright © 2021 D. Dankina. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Ever since the Carboniferous, Osteichthyes and Chondrichthyes have been the most successful marine vertebrates (Near et al. 2012; Pindakiewicz et al. 2020). Late Permian fish fossils are widely distributed in the marine and freshwater ecosystems around Pangea (Koot 2013; Romano et al. 2016) including saline, semi-enclosed, lagoon or the playa-like Zechstein Sea in NW Europe. Fish fossils from this basin are known from Germany (Diedrich 2009), England (King 1850), East Greenland (Nielsen 1952), Latvia (Dankina et al. 2020) and Lithuania (Dankina et al. 2017).
Until now only very rare and taxonomically low diversity occurrences of the fish remains were found in the southeastern part of the Zechstein successions. Kaźmierczak (1967) putatively assigned some teeth and scales as Palaeoniscidae in the Zechstein sequences at the Kajetanów quarry in the Holy Cross Mountains, central Poland. Incomplete Platysomus sp. trunks were also found from the copper-bearing Zechstein layers and Palaeoniscus sp. trunks from the Lubin mine in SW Poland, which are stored at the University of Wroclaw.
The current study represents the first record of the ichthyofaunal assemblage from a new paleoichthyological locality—the Nowy Kościół quarry in the SW Poland. The paleoenvironmental and ecological changes in the eastern Zechstein Sea ecosystems are discussed in connection to the ecomorphological features of described teeth assemblages. The described ichthyofaunal assemblage patterns from the SW Poland shed new light on the dispersal patterns and palaeobiogeography of fishes in the Zechstein Sea.
Institutional abbreviations.—VU-ICH-NK, Geological Museum at the Institute of Geosciences of Vilnius University, Vilnius, Lithuania.
Geological setting
The Zechstein Basin in Poland is a result of widespread cyclic carbonate and evaporite sediments (Poszytek and Suchan 2016). The studied sites of the first Zechstein cycle in SW Polish suggested an existence of a narrow zone of upper Permian sediments in the outer part of the North-Sudetic Basin (Gunia and Milewicz 1962; Raczyński 1997; Biernacka et al. 2005; Fig. 1A). The Nowy Kościół area is located in the Leszczyna Syncline in the southeastern, marginal part of the Zechstein Basin (Biernacka et al. 2005; Fig. 1B). Scupin (1933) proposed lithostratigraphic division of the upper Permian association in this part of the Zechstein Basin. According to Scupin’s (1933) division, the limestone-marl association is subdivided into three main units: spotted marl, copper-bearing marl, and lead-bearing marl (Biernacka et al. 2005).
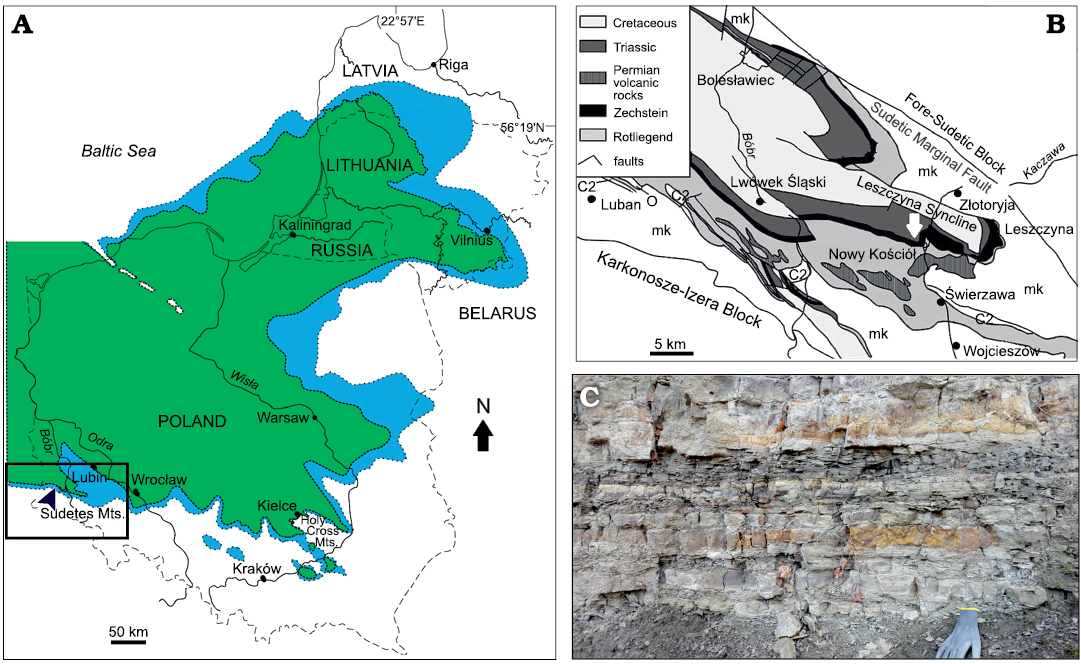
Fig. 1. Location of the late Permian fish-bearing site in SW Poland. A. Map of the Eastern Europe with position of Nowy Kościół (blue, the original distribution of the eastern margin of Zechstein Limestone; green, current distribution of Zechstein sediments in Poland, Russia, Lithuania, and Latvia; Raczyński and Biernacka 2014). B. The geological map of the North-Sudetic Basin showing location of studied site (white arrow) (after Biernacka et al. 2005). C. Photograph of the middle Zechstein limestone sequences of the Nowy Kościół section from 2016. mk, Kaczawa metamorphic rock; C2, Upper Carboniferous.
The limestone-marl sequence with the underlying Basal Limestone (micritic limestone) and overlying middle Zechstein (micro-oncolitic limestone) are equivalent to the carbonate rocks of the first evaporitic cyclothem assigned as Zechstein Limestone (Ca1) (Peryt 1978; Raczyński 1997; Biernacka et al. 2005). The duration of sedimentation of the entire first evaporitic cyclothem did not exceed two million years (Menning 1995; Biernacka et al. 2005). The upper Permian limestone-marl association was deposited in the ~20–30 km width and ~100 km length zone along the WNW-ESE stretching lagoon (Biernacka et al. 2005; Fig. 1B).
Material and methods
The studied material was collected in the Nowy Kościół quarry in SW Poland (51°5’19.654’’N, 15°52’43.613’’E) (Fig. 1). Sediments are mostly carbonates consisting of marl and limestone layers. In total eleven samples were collected. Two samples were taken from the Basal Limestone; three samples from copper-bearing marl; three samples from lead-bearing marl; three samples from Middle Zechstein Limestone (Fig. 2). The total weight of the collected samples reached ~128.4 kg. The average mass of each sample was ~14 kg.
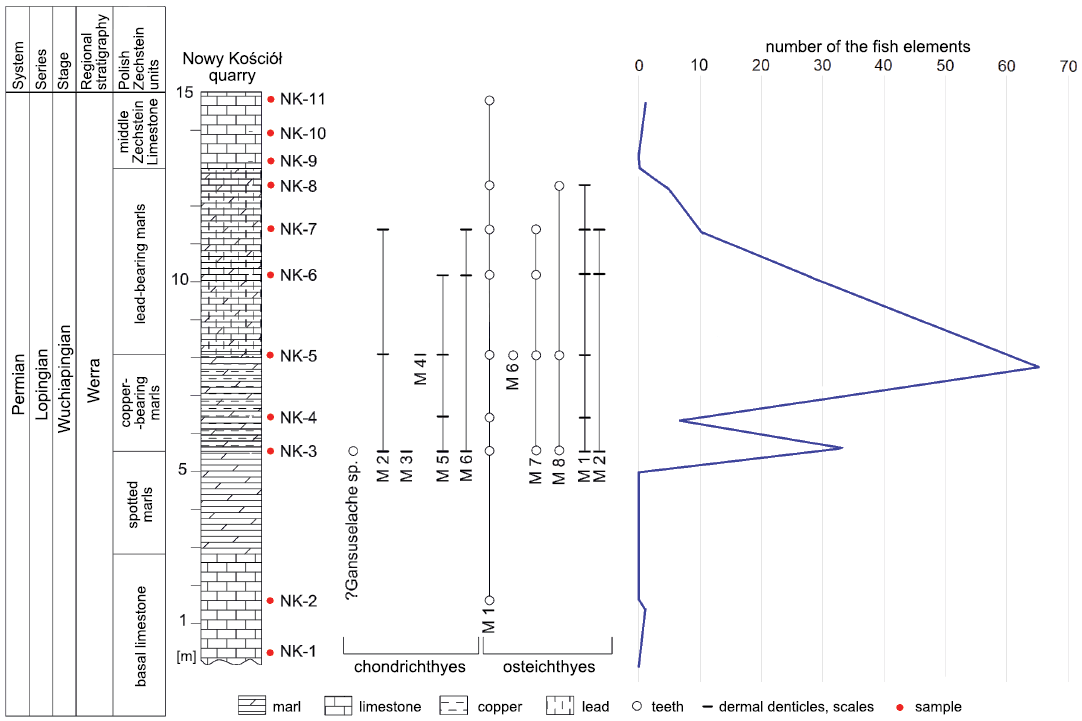
Fig. 2. Stratigraphical profile of the Nowy Kościół quarry with an indication of the late Permian fish assemblage, stratigraphic repartition of the chondrichthyan and osteichthyan taxa based on isolated teeth, dermal denticles and scales; and its vertical distribution. M, morphotype.
The fossil-bearing samples were chemically prepared using standard chemical dissolution technique in buffered formic acid described by Jeppsson et al. (1999). The residues were dried at room temperature and sieved from 0.2 to 0.063 mm sieves in order to more effectively optically spot and pick microremains under the binocular microscope under fixed magnification. Scanning electron microscope (SEM) imaging of the selected fish remains was conducted at the Nature Research Centre (Vilnius, Lithuania).
The collected samples yielded 112 isolated chondrichthyan and osteichthyan fish microremains. The collection is housed in the Geological Museum at the Institute of Geosciences of Vilnius University (VU-ICH-NK).
Systematic palaeontology
Class Chondrichthyes Huxley, 1880
Subclass Elasmobranchii Bonaparte, 1838
Order Hybodontiformes Patterson, 1966
Superfamily Hybodontoidea Owen, 1846
Family Lonchidiidae Herman, 1977
Genus Gansuselache Wang, Zhang, Zhu, and Zhao, 2009
Type species: Gansuselache tungshengi Wang, Zhang, Zhu, and Zhao, 2009; Mazongshan Mountain, Gansu Province, north-western China; Fangshankou Formation, late Permian.
?Gansuselache sp.
Fig. 3A.
Material.—Single isolated tooth (VU-ICH-NK-001) from the upper Permian of the Nowy Kościół Quarry, Leszczyna Syncline, SW Poland.
Description.—The tooth is mesiodistally elongate (slightly less than 3 mm in length), labiolingually compressed, low-crowned, slightly inclined lingually and near-symmetrical. The tooth crown is multicuspid; its central cusp is higher and wider than two pairs of low lateral cusplets. The cusplets are ornamented separately with two or three vertical strong ridges that continue to the root in labial and lingual face of the tooth. The tooth root is massive, higher in labial face than in the lingual one. It has some enlarged and irregular foramina on both lingual and lateral faces, which are filled by sediments. The tooth base is oval and deeply concave.
Remarks.—The teeth of ?Ganunselache sp. are rare in late Permian material. However, the analysed tooth here is assigned to ?Gansuselache sp. as it matches the following diagnosed characters: multicuspid crown, with central high cusp and lower two pairs of cusplets which are covered by vertical strong ridges. Similar teeth were found in the late Permian material of the Fangshankou Formation in north-western China (Wang et al. 2009: fig.7C).
Cohort Euselachii Hay, 1902
Euselachii indet.
Fig. 3B–R.
Material.—44 isolated dermal denticles were found from the upper Permian of the Nowy Kościół Quarry, Leszczyna Syncline, SW Poland. The dermal denticles are represented here by SEM microphotographs of microremains VU-ICH-NK-002–018.
Description.—The microremains are identified as euselachian-type dermal denticles based on resembling material from the middle Permian of the Apache Mountains in West Texas, USA (Ivanov et al. 2013); Permian of the Kanin Peninsula in Russia (Ivanov and Lebedev 2014); Lower Triassic of Oman (Koot et al. 2015); Carboniferous of Oklahoma, USA (Ivanov et al. 2017); upper Permian of Lithuania (Dankina et al. 2017), and Latvia (Dankina et al. 2020); and are divided into morphotypes based on the morphological differences between their crown, neck, and base.
The morphotype numeration and description in this study is taken from previous studies of late Permian euselachian material from southern Latvia (Dankina et al. 2020; Tables 1, 2).
Table 1. The different morphotypes distribution of late Permian fish remains in Latvia, Lithuania, and Poland.
| |
Morphotype |
Poland |
Lithuania (Karpėnai) |
Latvia (Kūmas) |
|
Euselachii |
M1 |
– |
– |
+ |
|
M2 |
+ |
+ |
+ |
|
|
M3 |
+ |
+ |
+ |
|
|
M4 |
+ |
+ |
+ |
|
|
M5 |
+ |
+ |
+ |
|
|
M6 |
+ |
+ |
+ |
|
|
Actinopterygii teeth |
M1 |
+ |
+ |
+ |
|
M2 |
– |
– |
+ |
|
|
M3 |
– |
+ |
+ |
|
|
M4 |
– |
+ |
+ |
|
|
M5 |
– |
– |
+ |
|
|
M6 |
– |
+ |
+ |
|
|
M7 |
+ |
– |
– |
|
|
M8 |
+ |
– |
– |
|
|
Actinopterygii scales |
M1 |
+ |
+ |
+ |
|
M2 |
+ |
– |
+ |
|
|
M3 |
– |
– |
+ |
|
|
M4 |
– |
+ |
+ |
|
|
M5 |
– |
– |
+ |
|
|
M6 |
– |
– |
+ |
Table 2. The main characteristics of the different morphotypes of Euselachii dermal denticles. M1–6, morphotype 1–6.
|
Morphotype |
Crown ridges |
Crown symmetry |
Horizontal crown |
Visible neck |
Roots foramina |
|
M1 |
no |
yes |
yes |
yes |
yes |
|
M2 |
yes |
yes |
yes |
yes |
no |
|
M3 |
partly |
yes |
partly |
yes |
yes |
|
M4 |
yes |
no |
yes |
partly |
no |
|
M5 |
partly |
partly |
no |
no |
no |
|
M6 |
yes |
partly |
no |
no |
no |
Morphotype 2: Seven dermal denticles from this morphotype were found in the Nowy Kościół quarry (Fig. 3B–D). The denticles have a trident or nearly trident crown with a high, slender, and narrow neck (Fig. 3B), hidden under the crown in apical view (Fig. 3C, D). The crown sits horizontally or slightly obliquely up on the neck. The exterior of the crown is sculptured with numerous gentle convex ridges and furrows originating at the longitudinal crest and reaching 0.1–0.4 mm length. The base has a rhomboid surface and one roundish canal opening in proximal view. The denticle reaches 0.3–0.4 mm height, and 0.4–0.5 mm crowns length and width.
Morphotype 3: Four dermal denticles from this morphotype were found in the Nowy Kościół quarry (Fig. 3E–G). The roundish crown is smooth, thick, without ornamentation (Fig. 3E, F). The crown sits horizontally on the neck and reaches around 0.4–0.8 mm in diameter. The neck is wide and massive. The wide base is slightly curved, multipetaloid in shape, and with concave canal opening in the proximal view (Fig. 3G).
Morphotype 4: Two dermal denticles from this morphotype were found in the Nowy Kościół quarry (Fig. 3H, I). The denticle has a curved outline of the crown; strongly convex, with three continuous ridges joined on the top of the surface (Fig. 3H). Sometimes, the crown is flat, thick, without ornament, has a serrated margin (Fig. 3I1). The crown is placed horizontally on the slender neck. The neck is short and narrow. The base has an indeterminate sinuous shape with deep vertical grooves along base outline. Dermal denticles reach 0.3–0.6 mm in length and 0.5–0.6 mm in width.
Morphotype 5: 21 dermal denticles from this morphotype were found in the Nowy Kościół quarry (Fig. 3J–L). The denticles have an anteriorly-inclined crown ornamented by some short ridges (4–5 ridges) on the anterior side (Fig. 3J, L). This type of denticles have a smooth, drop-like crown margin surface, which sits evidently obliquely up on the wide low neck (Fig. 3K). The crown reaches 0.4–0.5 mm length. The neck and crown widths are almost identical and approximately equal 0.3–0.4 mm (Fig. 3J). The base is flat, sinuous, multipetaloid in shape (Fig. 3L) with concave canal openings in the proximal view.
Morphotype 6: 10 dermal denticles from this morphotype were found in the Nowy Kościół quarry (Fig. 3M–R). The denticles have a complex crown shape, from very narrow, subparallel with straight ridges (Fig. 3O, Q) to wide triangular being striated by curved, branched ridges (Fig. 3P). The neck is poorly developed. The base is low and flat. Its profile has a rhomboid (Fig. 3M2) or polygonal outline (Fig. 3N2), with a slightly convex basal surface (Fig. 3M2) and without any foramina (Fig. 3R). These dermal denticles could reach 0.7–2.0 mm in length and 0.5–0.8 mm in height.

Fig. 3. Euselachian dermal denticles and hybodontoid tooth from the upper Permian of the Nowy Kościół quarry, Poland. A. ?Gansuselache sp. tooth VU-ICH-NK-001, labial (A1) and lingual (A2) views. B–D. Euselachian-type dermal denticles of morphotype 1. B. VU-ICH-NK-002, crown view. C. VU-ICH-NK-003, crown view. D. VU-ICH-NK-004, lateral view. E–G. Euselachian-type dermal denticles of morphotype 2. E. VU-ICH-NK-005, crown view. F. VU-ICH-NK-006, crown view. G. VU-ICH-NK-007, basal view. H, I. Euselachian-type dermal denticles of morphotype 3. H. VU-ICH-NK-008, apex crown view. I. VU-ICH-NK-008, apex crown (I1) and basal (I2) views. J–L. Euselachian-type dermal denticles of morphotype 4. J. VU-ICH-NK-010, crown view. K. VU-ICH-NK-011, lateral view. L. VU-ICH-NK-012, apex crown. M–R. Euselachian-type dermal denticles of morphotype 5. M. VU-ICH-NK-013, lateral crown (M1) and lateral basal (M2) views. N. VU-ICH-NK-014, apex crown (N1) and basal (N2) views. O. VU-ICH-NK-015, lateral crown view. P. VU-ICH-NK-016, crown view. Q. VU-ICH-NK-017, lateral crown view. R. VU-ICH-NK-018, lateral basal view.
Remarks.—Complex shape of the morphotype 2 denticles is morphologically similar to Carboniferous chondrichthyan scales of Oklahoma, USA (Ivanov et al. 2017). Also, this morphotype is similar to Devonian–early Carboniferous ctenacanthid scales in Belarus (Ivanov and Plax 2018); Carboniferous in Lublin area, SE Poland (Ginter and Skompski 2019); and Devonian of the Holy Cross Mountains (Liszkowski and Racki 1992). The identical dermal denticles of morphotype 3 have been found and described as late Permian euselachian-type dermal denticles from Naujoji Akmenė Formation in Lithuania (Dankina et al. 2017) and Latvia (Dankina et al. 2020). A roundish denticle of morphotype 4 was interpreted as being a ?hybodont/synechodontiform scale from the Lower Triassic in Oman (Koot et al. 2015). Similar denticles but with different neck have been found and described as late Permian euselachian-type denticles from the Naujoji Akmenė Formation in Lithuania (Dankina et al. 2017) and Latvia (Dankina et al. 2020). The morphological similarities of the morphotype 5 dermal denticles (shape of the crown, roots, slender neck) have been found and described as Triassic Hybodontidae dermal denticles in Spitsbergen (Reif 1978) and as late Permian euselachian-type denticles from the Naujoji Akmenė Formation in Lithuania (Dankina et al. 2017) and Latvia (Dankina et al. 2020). The morphotype 6 denticles with similar features (shape of the crown, ornament, base) were identified as hybodontiform scales from the Upper Triassic in Germany (Reif 1978) and Middle Triassic in Spain (Manzanares et al. 2014). Also, the denticles with morphological similarities (size, trident crown, ornament) have been found and described as late Permian euselachian-type dermal denticles from the Naujoji Akmenė Formation in Lithuania (Dankina et al. 2017) and Latvia (Dankina et al. 2020).
Superclass Osteichthyes Huxley, 1880
Class Actinopterygii Cope, 1887
Actinopterygii indet. (teeth)
Fig. 4A–L.
Material.—38 isolated teeth (Fig. 4A–L) from the upper Permian of the Nowy Kościół Quarry, Leszczyna Syncline, SW Poland. The teeth are represented here by SEM microphotographs of microremains VU-ICH-NK-019–030.
Description.—The teeth divided into three morphotypes based on their different shape and enameloid microstructure. The morphotype numeration and description in this study is continued on from previous studies of the late Permian actinopterygian material from southern Latvia (Dankina et al. 2020; Tables 1, 3).
Table 3. The main characteristics of the different morphotypes of Actinopterygii teeth. M1–8, morphotype 1–8.
|
Morphotype |
Surface ornament |
Flat crown top |
Visible roots |
Tooth symmetry |
Micro- |
|
M1 |
no |
no |
partly |
yes |
yes |
|
M2 |
yes |
yes |
partly |
yes |
yes |
|
M3 |
yes |
partly |
no |
partly |
yes |
|
M4 |
no |
partly |
partly |
yes |
yes |
|
M5 |
no |
yes |
no |
yes |
yes |
|
M6 |
yes |
partly |
no |
partly |
yes |
|
M7 |
no |
partly |
no |
partly |
partly |
|
M8 |
no |
no |
partly |
yes |
partly |
Morphotype 1: 26 teeth from this morphotype were found in the Nowy Kościół quarry (Fig. 4A–E). The canine-like teeth are conical, straight (Fig. 4A, B) or curved characterised by the “horn-like” shape (Fig. 4C), thin (Fig. 4A) or wide and convex in the central part (Fig. 4E), with an acrodin cap. Tooth surface is smooth, with no distinct visible ornament, although the microtubercles are well-developed (Fig. 4D2). The microtubercles are proximo-distally elongated, narrow and blend together in oblique rows (Fig. 4D2). Morphotype 1 teeth reach 0.4–1.2 mm in width and 0.6–1.9 mm in length.
Morphotype 7: Five teeth from this morphotype were found in the Nowy Kościół quarry (Fig. 4F–I). The molar teeth are roundish (Fig. 4F), short, slightly depressed in labial-lingual face (Fig. 4G), cylindrical in shape (Fig. 4H), and with convex outline in lateral view (Fig. 4I). The surface of these teeth is smooth with a slightly concave central part (Fig. 4F1, I1). Some teeth have well-developed, vertically elongated, narrow microtubercles (Fig. 4G1). These teeth reach a maximum of 0.2–0.6 mm in width, 0.6–1.2 mm in length and 0.4–1.0 mm in height.
Morphotype 8: Five teeth from this morphotype were found in the Nowy Kościół quarry (Fig. 4J–L). The teeth are straight, wide, and narrow (depressed in labial-lingual face) and with well-developed, vertically elongated, narrow microtubercles (Fig. 4J). Some teeth consist of the three layers: upper layer is dark grey; middle, light grey; lower, medium grey; according to the SEM picture (Fig. 4K, L) while under microscope the tooth’s upper part has the light amber acrodin (almost white); middle part is dark brown; and lower part—mid-amber colour. Rarely the tooth has concave top of the central part (Fig. 4K2, L). The surface is smooth, without any ornament (Fig. 4K, L), sometimes with preserved sediments in the lower part of the tooth (Fig. 4L). These teeth reach 0.7 mm in width and 1.2 mm in height.
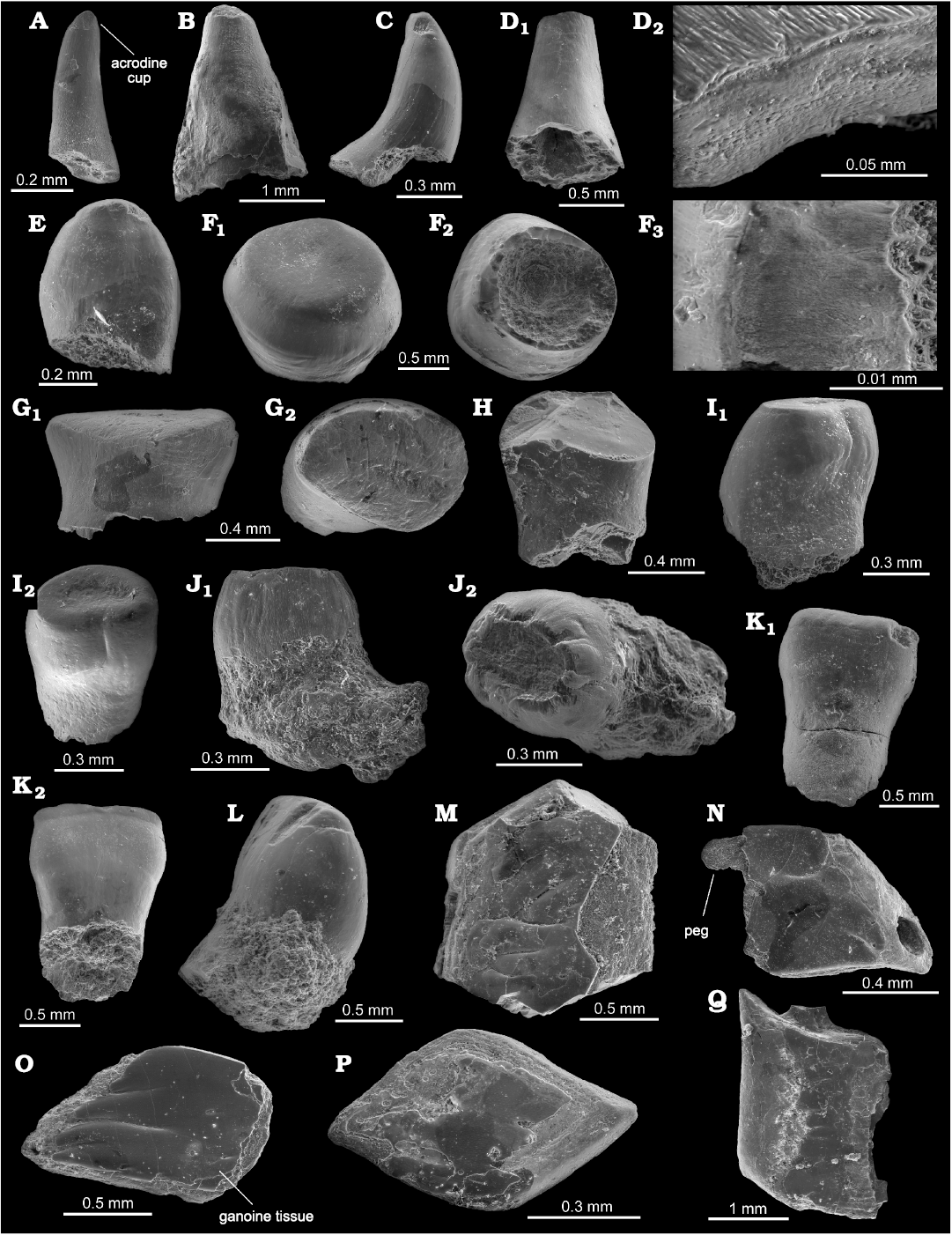
Fig. 4. Actinopterygian remains from the upper Permian of the Nowy Kościół quarry, Poland. A–E. Actinopterygian teeth of morphotype 1. A. VU-ICH-NK-019, lateral view. B. VU-ICH-NK-02-20, lateral view. C. VU-ICH-NK-021, lateral view. D. VU-ICH-NK-022, lateral view (D1), microtubercles structure (D2). E. VU-ICH-NK-023, lateral view. F–I. Actinopterygian teeth of morphotype 3. F. VU-ICH-NK-024, externo-lateral (F1) and basal (F2) views, detailed tissue pattern (F3). G. VU-ICH-NK-025, lateral (G1) and external (G2) views. H. VU-ICH-NK-026, lateral view. I. VU-ICH-NK-027, lateral (I1) and externo-lateral (I2) views. J–L. Actinopterygian teeth of morphotype 4. J. VU-ICH-NK-028, lingo-lateral (J1) and labio-lateral (J2) views. K. VU-ICH-NK-029, lingual (K1) and labial (K2) views. L. VU-ICH-NK-030, lateral view. M–P. Actinopterygian scales of morphotype 1, VU-ICH-NK-031–034, all external view. Q. Actinopterygian scale of morphotype 2, VU-ICH-NK-035, external view.
Remarks.—Similar Actinopterygii teeth of the morphotype 1 are known from Permian fish assemblages in Argentina (Cione et al. 2010), Latvia (Dankina et al. 2020) and Lithuania (Dankina et al. 2017). Moreover, the Gyrolepis albertii conical teeth with transparent acrodin cup are known from the Upper Triassic of the Westbury Formation in southwest Britain (Landon et al. 2017: fig.6A).
Actinopterygii indet. (scales)
Fig. 4M–Q.
Material.—29 scales (Fig. 4M–Q) from the upper Permian of the Nowy Kościół Quarry, Leszczyna Syncline, SW Poland. The scales are represented here by SEM microphotographs of microremains VU-ICH-NK-031–035.
Description.—The scales divided into two morphotypes relatively based on their different shape and ganoine layer characteristics. The morphotype numeration and description in this study is taken from previous studies of late Permian euselachian material from southern Latvia (Dankina et al. 2020; Tables 1, 4).
Table 4. The main characteristics of the different morphotypes of Actinopterygii scales. M1–6, morphotype 1–6.
|
Morphotype |
Ganoine top |
Scale symmetry |
Surface ridges |
Surface grooves |
Micro-tubercles |
Canal openings |
|
M1 |
yes |
yes |
no |
partly |
yes |
yes |
|
M2 |
yes |
partly |
no |
yes |
no |
no |
|
M3 |
yes |
no |
no |
partly |
yes |
yes |
|
M4 |
yes |
no |
yes |
no |
yes |
no |
|
M5 |
yes |
no |
no |
no |
yes |
yes |
|
M6 |
yes |
partly |
no |
partly |
yes |
yes |
Morphotype 1: 26 scales from this morphotype were found in the Nowy Kościół quarry (Fig. 4M–P). The scales are rhombic-shaped, thick, massive and gently convex in their central part. Scales have numerous small, roundish-shaped microtubercles in the outer ganoine-covered field part (Fig. 4M–P). The surface is covered by smooth ganoine tissue (Fig. 4P), with some fine, slightly diagonally oriented, short ridges, which are separated by narrow grooves (Fig. 4O). In rare cases, scales have a well-preserved peg articulation and significantly wide anterior entry of the lateral line canal (Fig. 4N). Scales reach 0.6–1.0 mm in length and 0.4–0.6 mm in width.
Morphotype 2: Three scales from this morphotype were found in the Nowy Kościół quarry (Fig. 4Q). Elongated rhombic-shaped scale with almost right-angled (~90°) all four corners. The base is thick and convex in the central part. The scale has preserved a fragment of the peg articulation. The ganoine-covered field is smooth without any ornament. The scale reaches 2.3 mm length and 1.4 mm width.
Remarks.—The scales collection of morphotype 1 with similar morphological characteristics (rhombic shape and ganoine pattern) was found in late Permian actinopterygian scales from Lithuania (Dankina et al. 2017) and Latvia (Dankina et al. 2020). Also, rhomboidal scales are known from the middle Permian of the Apache Mountains in West Texas, USA (Ivanov et al. 2013); Permian in England (King 1850); and Permian in Argentina (Cione et al. 2010). The morphotype 2 scale seems to be similar to the Middle Triassic Gyrolepis sp. rhomboidal scales from the North-Sudetic Basin in Poland (Chrząstek 2013).
Discussion
The hybodontoids are one of the best-known groups of fossil sharks and their teeth, spines and scales are common fossils in several Mesozoic rock formations (Wang et al. 2009). The new material (isolated chondrichthyan tooth) described herein is attributed to the family Lonchidiidae. This tooth has the most morphological similarities (elongate shape, low-crowned, higher central cusp and two pairs of low lateral cusplets, ornament, deep root, with irregular, numerous foramina) to the teeth of Gansuselache from the upper Permian of north-western China (Wang et al. 2009). Although Lissodus teeth are similarly shaped (Ginter et al. 2010), the main differences between them and the described tooth are blunt cusps and an absence of the coarse vertical ridge ornamentation, which descends from the tip of the cusp downwards. Additionally, no obvious labial peg was observed, which is typical for Lissodus. On the other hand, the geographic distribution of the genus Gansuselache Wang et al. (2009) first described from the late Permian deposits in north-western China, is poorly known. However, the Lonchidiidae remains are more widely distributed and known from the Permian of the Kanin Peninsula, Arkhangelsk Region in Russia (Ivanov and Lebedev 2014); lower Permian of Germany (Hampe 1996); and middle Permian of central Japan (Yamagishi and Fujimoto 2011).
Herein, actinopterygian teeth are divided into three different morphotypes based on the various shapes. The tooth form is mainly determined by the different types of food ray-finned fishes consume. Every trophic group is characterised by a certain mode of feeding, suited type of diet, and specific morphofunctional adaptations of jaws and teeth (Esin 1997). According to Esin (1997) who described major trophic groups of fish, “morphotype 1” with a small canine-like shape suggests “specialised sclerophagous” and “small predator” diets. The small predatory teeth are conical with acute terminations (Fig. 4A, B). Sometimes this morphotype of teeth are high, conic, slightly turned back into the mouth (Fig. 4C) and high-conic with blunt top shapes (Fig. 4E). Smaller teeth (presumably of juvenile individuals) of this type are covered with numerous acute microtubercles (Fig. 4D). Fishes with similar teeth shape mostly fed on crustaceans with hard shells and also on aquatic insects (Esin 1997). The earliest aquatic insects are known from the early Permian (Sinitshenkova 2003). This type of feeding could be compared to that of modern chubs or perch, which diets include arthropods, small fishes and vegetation (Esin 1997). The Permian genera, such as Acropholis, Acrolepsis, Palaeoniscum, Elonichthys, Varialepis are characterised by this tooth ecomophotype (Esin 1997). Acropholis remains are known from the Permian of Greenland (Aldinger 1937) and Russia (Nurgaliev et al. 2015); Acrolepsis was found in Permian sequences of Germany (Diedrich 2009), the United Kingdom (King 1850) and Russia (Nurgaliev et al. 2015); Permian Palaeoniscum was found in United Kingdom (King 1850); Germany (Diedrich 2009), Russia (Nurgaliev et al. 2015), Turkey (Hoşgör and Štamberg 2014); Permian Elonichthys are known from Greenland (Aldinger 1937) and Russia (Nurgaliev et al. 2015); while Permian Varialepis was found in Russia (Tverdokhlebov et al. 2005; Nurgaliev et al. 2015), and USA (Ivanov et al. 2013).
Morphotype 7 teeth suggest durophagous-type mode of feeding. These teeth are rod-shaped or slightly depressed cylinders with rounded and almost rounded flat or concave apex (Fig. 4F–I). These teeth most likely were used for crushing and grinding of hard external skeletons, such as shells (Purnell and Darras 2015) of molluscs or brachiopods as well as possibly soft-bodied invertebrates (polychaetes) (Esin 1997). Productid brachiopods, and many species of bivalves were common in the Zechstein strata and are known from the Holy Cross Mountains, Poland (Kaźmierczak 1967), England (Ramsay 1878), and Lithuania (Suveizdis 1975). Also, polychaetes are known from the upper Permian of Poland (Szaniawski 1968). This type of teeth can be compared to the modern breams in their mode of life (Esin 1997). Morphotype 8 teeth are interpreted here to represent a “grazing” trophic mode (Fig. 4J–L). These teeth come in a variety of different shapes, others are characterised by the saw-edge or even are fused into the “beak”, like in modern parrotfishes. The wide and thin teeth are adapted for feeding on water vegetation, apparently used for cutting thin threadlike algae (Esin 1997). This morphotype of teeth is characterised by the herbivorous diet, which is very similar to Recent freshwater teleosts, such as characiforms and cichlids (Pindakiewicz et al. 2020).
The morphotype diversity of fish teeth from the upper Permian in the SW Poland indicates that ray-finned fishes already started wide exploration of different feeding modes before the onset of the Mesozoic.
Conclusions
The micropaleontological study of the late Permian fish assemblages from the distal storm deposits of the Nowy Kościół quarry revealed assemblages that are composed of non-abundant but relatively diverse isolated microremains of different fish taxa, including euselachian dermal denticles, diverse actinopterygians scales and teeth, and ?Gansuselache sp. tooth. The finding of the later taxon significantly extends the known geographic distribution of this hybodontiform genus (or closely related forms), which was previously known only from China.
The stratigraphic distribution of fish remains and in particular their abundance in Nowy Kościół site shows high congruence with the Werra marine transgression. This transgression created the favourable conditions for flourishing of fish fauna in the Zechstein Basin. The highest abundance of fish microremains and their diversity was found at the boundary between copper-bearing and lead-bearing marl sequences in Nowy Kościół quarry. The association of transgression with higher abundance of ichtyofauna could be explained by two-fold effect. Transgression should have positively affected abundance by increasing runoff of nutrients from surrounding terrain in a warmer climate, and diluting hyper-saline waters. On other hand warmer climates, which accompany transgressions, should have promoted effectiveness of biomass transfer to the higher trophic levels occupied by fishes by physiologically reinvigorating predation efficiency (Britten and Sibert 2020).
The diverse morphology of actinopterygian denticulation demonstrates the prevalence of three trophic groups in the eastern Zechstein Sea margin. Those groups are inferred based on the three ecomorphotypes, which dominate the assemblages: (i) morphotype 1, teeth of specialised sclerophagous and small predators; (ii) morphotype 7, the crushing and grinding teeth of durophagous-type; (iii) morphotype 8, peg-like teeth adapted for herbivory or general grazing. The actinopterygian teeth material indicates that small generalized predators dominated among the recorded ichthyofauna. The diversity of different types of teeth indicates an onset of active ecomorphological specialization among the fish taxa, which further strengthens the case that the roots of Mesozoic Marine Revolution were set in Palaeozoic.
Acknowledgements
The authors would like to thank Michał Ginter (University of Warsaw, Poland) and anonymous reviewer for significantly improving the manuscript. Also, we would like to express our thanks to Vaida Kirkliauskaitė (Vilnius University, Lithuania), who helped safely transport samples. Many thanks to Laurynas Šiliauskas (Nature Research Centre, Lithuania) for the SEM micrographs. This research was carried out by the Open Access to the research infrastructure of the Nature Research Centre (Vilnius) under the Lithuanian open access network initiative. The field work in Poland was financed by Sepkoski Grant 2018 of Paleontological Society (PaISIRP).
References
Aldinger, H. 1937. Permische ganoidfische aus Ostgrönland. Meddelelser om Grønland 102 (3): 1–392.
Biernacka, J., Borysiuk, K., and Raczyński, P. 2005. Zechstein (Ca1) limestone-marl alternations from the North-Sudetic Basin, Poland: depositional or diagenetic rhythms? Geological Quarterly 49: 1–14.
Bonaparte, C.L.J.L. 1838. Selachorum tabula analytica. Nuovi Annali delle Scienze Naturali 1: 195–214.
Britten, G.L. and Sibert, E.C. 2020. Enhanced fish production during a period of extreme global warmth. Nature communications 11: 1–6. Crossref
Chrząstek, A. 2013. Trace fossils from the Lower Muschelkalk of Raciborowice Górne (North Sudetic Synclinorium, SW Poland) and their palaeoenvironmental interpretation. Acta Geologica Polonica 63: 315–353. Crossref
Cione, A.L., Gouiric-Cavalli, S., Mennucci, J.A., Cabrera, D.A. and Freije, R.H. 2010. First vertebrate body remains from the Permian of Argentina (Elasmobranchii and Actinopterygii). Proceedings of the Geologists’ Association 121: 301–312. Crossref
Cope, E.D. (1887). Zittel’s manual of paleontology. American Naturalist 17: 1014–1019.
Dankina, D., Chahud, A., Radzevičius, S., and Spiridonov, A. 2017. The first microfossil record of ichthyofauna from the Naujoji Akmenė Formation (Lopingian), Karpėnai Quarry, North Lithuania. Geological Quarterly 61: 602–610. Crossref
Dankina, D., Spiridonov, A., Stinkulis, Ģ., Manzanares, E., and Radzevičius, S. 2020. A late Permian ichthyofauna from the Zechstein Basin, Lithuanian-Latvian Region. Journal of Iberian Geology 46: 1–21. Crossref
Diedrich, C.G. 2009. A coelacanthid-rich site at Hasbergen (NW Germany): taphonomy and palaeoenvironment of a first systematic excavation in the Kupferschiefer (upper Permian, Lopingian). Palaeobiodiversity and Palaeoenvironments 89: 67–94. Crossref
Esin, D.N. 1997. Peculiarities of trophic orientation changes in palaeoniscoid assemblages from the upper Permian of the European part of Russia. Modern Geology 21: 185–196.
Ginter, M. and Skompski, S. 2019. The squamation of “Ctenacanthus” costellatus (Chondrichthyes: Ctenacanthiformes) from the Carboniferous of Lublin area, south-eastern Poland. Acta Geologica Polonica 69: 571–582.
Ginter, M., Hampe, O., Duffin, C.J., and Schultze, H.-P. 2010. Handbook of Paleoichthyology. Volume 3D. Chondrichthyes. Paleozoic Elasmobranchii: Teeth. 168 pp. Verlag Dr. Friedrich Pfeil, München.
Gunia, T. and Milewicz, J. 1962. Development of the Zechstein in the North-Sudetic Basin. Biuletyn Państwowego Instytutu Geologicznego 173: 115–126.
Hampe, O. 1996. Dermale Skelettreste von Lissodus (Chondrichthyes: Hybodontoidea) aus dem Unterperm des Saar-Nahe-Beckens. Paläontologische Zeitschrift 70: 225–243. Crossref
Hay, O.P. 1902. Bibliography and catalogue of the fossil vertebrata of North America. Bulletin of the United States Geological Survey 179: 1–868. Crossref
Herman, J. 1977. Les Sélaciens des terrains néocrétacés & paléocènes de Belgique & des contrées limitrophes. Eléments d’une biostratigraphie intercontinentale. Mémoires pour servir à l’explication des Cartes géologiques et minières de la Belgique 15: 1–450.
Hoşgör, I. and Štamberg, S. 2014. A first record of late middle Permian actinopterygian fish from Anatolia, Turkey. Acta Geologica Polonica 64: 147–159. Crossref
Huxley, T.H. 1880. On the application of the laws of evolution to the arrangement of the Vertebrata and more particularly of the Mammalia. Proceedings of the Zoological Society of London 43: 649–662.
Ivanov, A.O. and Lebedev, O.A. 2014. Permian chondrichthyans of the Kanin Peninsula, Russia. Paleontological Journal 48: 1030–1043. Crossref
Ivanov, A.O. and Plax, D.P. 2018. Chondrichthyans from the Devonian–Early Carboniferous of Belarus. Estonian Journal of Earth Sciences 67: 43–58. Crossref
Ivanov, A.O., Nestell, G.P., and Nestell, M.K. 2013. Fish assemblage from the Capitanian (middle Permian) of the Apache Mountains, West Texas, USA. The Carboniferous–Permian Transition: Bulletin 60: 152–160.
Ivanov, A.O., Seuss, B., and Nützel, A. 2017. The fish assemblage from the Pennsylvanian Buckhorn Asphalt Quarry Lagerstätte (Oklahoma, USA). Paläontologische Zeitschrift 91: 565–576. Crossref
Jeppsson, L., Anehus, R., and Fredholm, D. 1999. The optimal acetate buffered acetic acid technique for extracting phosphatic fossils. Journal of Paleontology 73: 964–972. Crossref
Kaźmierczak, J.O. 1967. Morphology and palaeoecology of the productid Horridonia horrida (Sowerby) from Zechstein of Poland. Acta Palaeontologica Polonica 12: 239–264.
King, W. 1850. The Permian fossils of England. Monograph of the Palaeontographical Society of London 37: 1–253.
Koot, M.B. 2013. Effects of the Late Permian Mass Extinction on Chondrichthyan Palaeobiodiversity and Distribution Patterns. 859 pp. Ph.D. dissertation, University of Plymouth, Plymouth.
Koot, M.B., Gilles, C., Orchard, M.J., Richoz, S., Hart, M.B., and Twitchett, R.J. 2015. New hybodontiform and neoselachian sharks from the Lower Triassic of Oman. Journal of Systematic Palaeontology 13: 891–917. Crossref
Landon, E.N., Duffin, C.J., Hildebrandt, C., Davies, T.G., Simms, M.J., and Benton, M.J. 2017. The first discovery of crinoids and cephalopod hooklets in the British Triassic. Proceedings of the Geologists’ Association 128: 360–373. Crossref
Liszkowski, J. and Racki, G. 1992. Ichthyoliths and deepening events in the Devonian carbonate platform of the Holy Cross Mountains. Acta Palaeontologica Polonica 37: 407–426.
Manzanares, E., Plá, C., Martínez-Pérez, C., Rasskin, D., and Botella, H. 2014. The enameloid microstructure of euselachian (Chondrichthyes) scales. Paleontological Journal 48: 1060–1066. Crossref
Menning, M. 1995. A numerical time scale for the Permian and Triassic Periods. In: P.A. Scholle, T. Peryt, D.S. Ulmer-Scholle (eds.), The Permian of Northern Pangea. Vol. 1: Paleogeography, Paleoclimates, Stratigraphy, 77–97. Springer, Heidelberg. Crossref
Near, T.J., Eytan, R.I., Dornburg, A., Kuhn, K.L., Moore, J.A., Davis, M.P., Wainwright, P.C., Friedman, M., and Smith, W.L. 2012. Resolution of ray-finned fish phylogeny and timing of diversification. Proceedings of the National Academy of Sciences 109: 13698–13703. Crossref
Nielsen, E. 1952. On new or little known Edestidae from the Permian and Triassic of East Greenland. Palaeozoologica Groenlandica 6: 5–55.
Nurgaliev, D.K., Silantiev, V.V., and Nikolaeva, S.V. (eds.) 2015. Type and Reference Sections of the Middle and Upper Permian of the Volga and Kama River Regions. A Field Guidebook of XVIII International Congress on Carboniferous and Permian. 208 pp. Kazan University Press, Kazan.
Owen, R. 1846. Lectures on comparative anatomy and Physiology of Vertebrate Animals. Part 1. Fishes. 51 pp. Royal College of Physicians of London, London. Crossref
Patterson, C. 1966. British Wealden sharks. Bulletin of the British Museum (Natural History) 11: 283–350.
Peryt, T.M. 1978. Sedimentology and paleoecology of the Zechstein Limestone (upper Permian) in the Fore-Sudetic area (western Poland). Sedimentary Geology 20: 217–243. Crossref
Pindakiewicz, M., Tałanda, M., Sulej, T., Niedźwiedzki, G., Sennikov, A.G., Bakaev, A S., Bulanov, V.V., Golubev, V.K., and Minikh, A. 2020. Feeding convergence among ray-finned fishes: teeth of the herbivorous actinopterygians from the latest Permian of East European Platform, Russia. Acta Palaeontologica Polonica 65: 71–79. Crossref
Poszytek, A. and Suchan, J. 2016. A tight-gas reservoir in the basinal facies of the upper Permian Ca1 in the southwestern Zechstein Basin, Poland. Facies 62: 1–13. Crossref
Purnell, M.A. and Darras, L.P. 2015. 3D tooth microwear texture analysis in fishes as a test of dietary hypotheses of durophagy. Surface Topography: Metrology and Properties 4: 014006. Crossref
Raczyński, P. 1997. Depositional conditions and paleoenvironments of the Zechstein deposits in the North Sudetic Basin, SW Poland. Przegląd Geologiczny 45: 693–699.
Ramsay, W. 1878. A synopsis of the genus Pomatorhinus. Ibis 20: 129–145. Crossref
Reif, W.E. 1978. Types of morphogenesis of the dermal skeleton in fossil sharks. Paläontologische Zeitschrift 52: 110–128. Crossref
Romano, C., Koot, M. B., Kogan, I., Brayard, A., Minikh, A.V., Brinkmann, W., Bucher, H., and Kriwet, J. 2016. Permian–Triassic Osteichthyes (bony fishes): diversity dynamics and body size evolution. Biological Reviews 9: 106–147. Crossref
Scupin, H. 1933. Der Buntsandstein der Nordsudeten. Zeitschrift der Deutschen Geologischen Gesellschaft 85: 161–189.
Sinitshenkova, N.D. 2003. Main ecological events in aquatic insects history. Acta Zoologica Cracoviensia 46: 381–392.
Suveizdis, P.I. 1975. Permskaâ sistema Pribaltiki (fauna i stratigrafiâ). 305 pp. Mintis, Vilnius.
Szaniawski, H. 1968. Three new polychaete jaw apparatuses from the upper Permian of Poland. Acta Palaeontologica Polonica 13: 252–281.
Tverdokhlebov, V.P., Tverdokhlebova, G.I., Minikh, A.V., Surkov, M.V., and Benton, M.J. 2005. Upper Permian vertebrates and their sedimentological context in the South Urals, Russia. Earth-Science Reviews 69: 27–77. Crossref
Wang, N.Z., Zhang, X., Zhu, M., and Zhao, W.J. 2009. A new articulated hybodontoid from late Permian of northwestern China. Acta Zoologica 90: 159–170. Crossref
Yamagishi, H. and Fujimoto, T. 2011. Chondrichthyan remains from the Akasaka Limestone Formation (Middle Permian) of Gifu Prefecture, Central Japan. Bulletin of the Kanagawa Prefectural Museum of Natural Science 40: 1–6.
Acta Palaeontol. Pol. 66 (Supplement to 3): 47–57, 2021
https://doi.org/10.4202/app.00839.2020