
First record of chimaeroid fish Ischyodus from the Upper Jurassic of southwestern Gondwana
RODRIGO A. OTERO, CONSTANZA FIGUEROA BRAVO, PAULA SOTO-HUENCHUMÁN, SARA FERNÁNDEZ-COLLEMANN, ANA M. VALENZUELA-TORO, and CAROLINA S. GUTSTEIN
Otero, R.A., Figueroa Bravo, C., Soto Huenchumán, P., Fernández-Collemann, S., Valenzuela-Toro, A.M., and Gutstein, C.S. 2021. First record of chimaeroid fish Ischyodus from the Upper Jurassic of southwestern Gondwana. Acta Palaeontologica Polonica 66 (3): 623–630.
This study presents two specimens of Chimaeriformes from Upper Jurassic strata of central Chile. The material was recovered from Tithonian levels of the Baños del Flaco Formation and includes two different individuals, one preserving two articulated mandibular plates, and the second, a fragment of an isolated palatine plate. Morphologic traits allow us to refer the material to Ischyodus townsendi and Ischyodus sp., respectively. These are the oldest Chimaeriformes known to date in the Southern Hemisphere and the first Late Jurassic record from Gondwana. The presence of I. townsendi in the Tithonian of central Chile evidences this taxon as part of the proposed faunal interchange between the northern Tethys and the southeastern Pacific during the Late Jurassic.
Key words: Chondrichthyes, Holocephali, Callorhinchidae, cartilaginous fishes, paleobiogeography, Mesozoic, Jurassic, Tithonian.
Rodrigo A. Otero [otero2112@gmail.com], Consultora Paleosuchus Ltda. Huelén 165 Oficina C, Providencia, Santiago, Chile; Red Paleontológica U-Chile. Laboratorio de Ontogenia y Filogenia, Departamento de Biología, Facultad de Ciencias, Universidad de Chile. Las Palmeras 3425, Santiago, Chile; Museo de Historia Natural y Cultural del Desierto de Atacama. Inferior Parque El Loa s/n, Calama, Chile.
Constanza Figueroa Bravo [constanza.figueroa@paleoconsultores.cl] and Sara Fernández Collemann [sara.fernandez@paleoconsultores.cl], Consultora Paleosuchus Ltda. Huelén 165 Oficina C, Providencia, Santiago, Chile.
Paula Soto-Huenchumán [paula.soto@paleoconsultores.cl], Consultora Paleosuchus Ltda. Huelén 165 Oficina C, Providencia, Santiago, Chile; Red Paleontológica U-Chile. Laboratorio de Ontogenia y Filogenia, Departamento de Biología, Facultad de Ciencias, Universidad de Chile. Las Palmeras 3425, Santiago, Chile; Corporación Laguna de Taguatagua, Av. Libertador Bernardo O’Higgins 351, Santiago, Chile.
Ana M. Valenzuela-Toro [avalenzuela.toro@gmail.com], Consultora Paleosuchus Ltda. Huelén 165 Oficina C, Providencia, Santiago, Chile; Department of Ecology and Evolutionary Biology, University of California Santa Cruz, 130 McAllister Way, Santa Cruz, CA 95060, USA; Department of Paleobiology, National Museum of Natural History, Smithsonian Institution, 10th & Constitution NW, Washington, DC 20560 USA.
Carolina S. Gutstein [carolina.sg@paleoconsultores.cl], Consultora Paleosuchus Ltda. Huelén 165 Oficina C, Providencia, Santiago, Chile; Red Paleontológica U-Chile. Laboratorio de Ontogenia y Filogenia, Departamento de Biología, Facultad de Ciencias, Universidad de Chile, Las Palmeras 3425, Santiago, Chile.
Received 13 January 2020, accepted 7 January 2021, available online 2 August 2021.
Copyright © 2021 R.A. Otero et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Chimaeriformes (Chondrichthyes, Holocephali) is a clade of cartilaginous fishes that appeared in the fossil record during the Paleozoic, reaching a high diversity during the Mesozoic and onwards (López-Arbarello et al. 2008). During the Jurassic, the group is well-recorded in the Northern Hemisphere and represented by several genera and species (López-Arbarello 2004; López-Arbarello et al. 2008).
The oldest austral fossil Chimaeriformes are known from the Lower Cretaceous of Australia (represented by the Ischyodus Egerton, 1843 and Pyctoptychion Lees, 1986), becoming abundant during the Upper Cretaceous with records in New Zealand, South America, and Antarctica (e.g., Otero et al. 2013a, b). Then, during the Cenozoic, Chimaeriformes have been recorded globally (Stahl 1999). However, prior to the Cretaceous, there are no records of this clade along the Southern Hemisphere. This contribution presents the oldest Chimaeriformes known to date in the Southern Hemisphere and the first Upper Jurassic record from Gondwana. The material consists of two specimens which preserve enough diagnostic features for a taxonomic identification, being identified as Ischyodus townsendi (Buckland, 1835), while the second specimen is referred to as Ischyodus sp. The coeval occurrence of I. townsendi during the Tithonian in the northern Tethys, and now in southwestern Gondwana, is discussed here.
Institutional abbreviations.—MHNC, Museo de Historia Natural de Concepción, Chile; MJSN, Jurassica Museum (formerly Musée Jurassien des Sciences Naturelles), Porrentruy, Switzerland; NHMUK, Natural History Museum, London, UK.
Geological setting
The studied material was recovered in the facilities of Mina Del Fierro (Del Fierro Mine), owned by Minera Río Teno S.A., located in the Cajón del Fierro locality, High Andes, west of Curicó city, Región del Maule, Chile (Fig. 1). Rocks cropping out in this area belong to the Baños del Flaco Formation (Klohn 1960). This sedimentary unit overlies the Río Damas Formation through a concordant contact. The Baños del Flaco Formation is overlain through a discordant contact by a volcanoclastic sequence known as the Brownish-Red Clastic Unit (BRCU; Charrier et al. 1996; Salazar 2012), and through a concordant contact with the Colimapu Formation. The Baños de Flaco Formation was recently divided in two formal units, the Lower and the Upper Member (Salazar 2012). The unit comprises limestone, sandy limestone, and calcareous sandstone mostly grey-colored, with occasional glauconitic sandstone, limestone, and conglomerate. Salazar (2012) estimated thickness of 369 m in the studied locality.
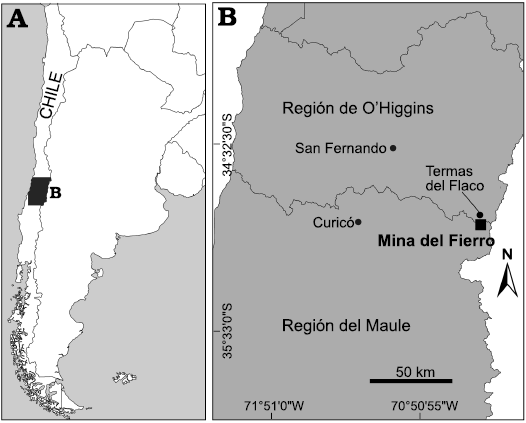
Fig. 1. A. Map indicating location of the studied area in central Chile. B. Enlarged map of Mina Del Fierro area where the material was collected.
The age of the Baños del Flaco has been debated over the years. In this sense, the abundant and diverse fauna of marine invertebrates (including ammonoids, belemnoids, bivalves, gastropods, scaphopods, echinoderms, bryozoans, porifera, foraminifera, algae, cyanobacteriae, and ichnofossils) present in the Baños del Flaco Formation was useful for increasing the precision of its age. Firstly, Klohn (1960) assigned a lower Valanginian–Hauterivian age, based on marine molluscs (mostly ammonoids) present in different outcrops of the unit. After, Covacevich et al. (1976) referred it to the lower Tithonian–Hauterivian? based on the invertebrate assemblage. Later, in more comprehensive studies, Salazar (2012) and Salazar and Stinnesbeck (2015) constrained the age of the unit to the lower middle Tithonian–lowermost Berriasian interval, based on the biostratigraphy of ammonoids.
For the Cajón del Fierro locality, Klohn (1960) described outcrops of Baños del Flaco Formation as fossiliferous marine sediments with N8E/30W strike and dip strike, being represented by a thick succession of limestone and marl with intercalated cross-bedded calcareous sandstone, and conglomerate levels, both concordantly overlying to breccias and conglomerates with andesitic clasts from the Río Damas Formation. Particularly, in the studied locality, Maldonado (2016) described the basal part of the Lower Member of Baños del Flaco Formation, conformed by conglomerate, conglomeratic limestone, sandy limestone, calcareous sandstone, intercalated tuff, calcirudite, and calcareous shale. For this section, the fossil content is abundant (Quinzio and Varela 2015), being present in most levels of the section and comprising bioturbation and marine invertebrates (ammonoids, belemnoids, gastropods, inoceramids, nautiloids, and trigoniids) all of which indicate a Tithonian age. The nature of this assemblage, in addition to the stratigraphic section described by Quinzio and Varela (2015), allows to assess the stratigraphic position of the two vertebrate specimens studied here. These are detailed in Fig. 2. Both specimens were recovered from upper levels of the Lower Member of the Baños del Flaco Formation, thus, having a Tithonian age.
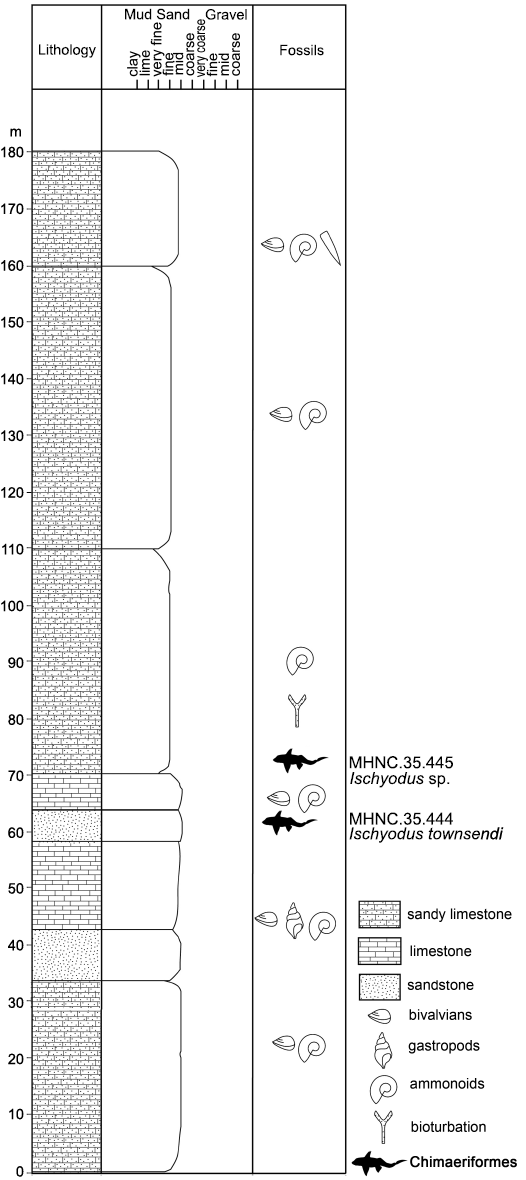
Fig. 2. Stratigraphic section of the levels were the studied material was recovered. Modified from Quinzio and Varela (2015).
Material and methods
The material is included in a single sandstone block with two articulated mandibular plates; no other skeletal portions were found nearby. This specimen was discovered in 2019 by personnel of Mina Del Fierro as part of a monitoring policy for mitigation of the mining operation, following the Chilean heritage law. Therefore, as the fossil-bearing geologic unit (Baños del Flaco Formation) is known to host frequent marine invertebrates, the area was the subject of a periodic paleontological survey, as requested by the local Chilean heritage authority (Consejo de Monumentos Nacionales) which was carried out under a paleontological excavation permit of the Chilean authority for one of the authors (CFB; Ord. CMN N° 2275/17).
The site of the finding was firstly inspected by two of the authors (SFC and PSH) during the paleontological survey of the mining activities, recognizing the stratigraphic provenance of the material. The block was originally removed from its natural occurrence by mine blasting, together with several large blocks of calcareous rocks.
Among the rescued material, a single sandstone block was recovered, originally showing a partial broken dental plate and part of a second, likely complete and articulated plate. The articulated plates were later prepared by one of the authors (RAO) during December 2019 and January 2020 in facilities of the Red Paleontológica U-Chile, Laboratorio de Ontogenia y Filogenia, Departamento de Biología, Universidad de Chile (Santiago, Chile). The extraction of the sediment was carried out with a 9100 air scribe, while the fragile specimen was consolidated with Paraloid B-76 using different concentrations, and cyanoacrylate for bonding. The second isolated plate was not prepared, and it is shown in its natural occurrence, which is enough for its anatomical interpretation. After, both specimens were included in the formal repository of the Museo de Historia Natural de Concepción, together with the rest of the fossil invertebrate collection from the studied locality.
The studied material was compared with Late Jurassic species within the Ischyodus, based on literature review. The anatomical terminology used here follows Popov and Machalski (2014), adding an anatomical topology (plm, prominence of the labial margin) discussed in the diagnosis of the Ischyodus by Stahl (1999).
Systematic palaeontology
Chimaeroidei Patterson, 1965
Callorhinchidae Garman, 1901
Callorhinchinae Stahl, 1999
Genus Ischyodus Egerton, 1843
Type species: Chimaera townsendi Buckland, 1835; Tithonian of England.
Ischyodus townsendi (Buckland, 1835)
Fig. 3A.
Material.—MHNC.35.444, two articulated mandibular plates from Mina Del Fierro, Andes Cordillera, Región del Maule, Chile; Baños del Flaco Formation, Tithonian (Upper Jurassic).
Description.—Both plates are visible in basal view (Fig. 3A). The mandibular plates remain articulated in nearly anatomical position. The right mandibular plate is the most damaged by diagenesis or transport, with the basal surface lost and a few tritors broken, being exposed in different sections. This preservation allows us to observe the tritor distribution. Its posterior occlusal surface is preserved as a thin bone layer attached to the matrix, which is why this was not removed. The preserved surface indicates a sudden mechanical break-up, judging by the fresh bone exposed. This is consistent with damage derived from rock blasting, discarding erosive agents. On the broken surface (basal view) it can be recognized a larger tritor pad placed medially, and therefore, being identified as the median tritor which covers two-thirds of the plate width. A cylindrical tritor, identified as the posterior outer tritor (pot), flanks labially to the median tritor (mt). Over the labial margin of each plate there is a prominence. The broken right mandibular plate shows the anterior outer tritor (aot) near to this prominence. The left mandibular shows two marked ridges (r) over the basal surface, which reflect the internal position of the symphyseal tritor (syt) of laminated pleromin and the median tritor, respectively. On its basal margin, the symphyseal tritor is broken, leaving a cavity. The median tritor is visible, it is also partially broken, but prominent. Flanking the latter, there is a posterior exposure of the posterior outer tritor. The beak (bk) is preserved in both plates, being prominent and slightly recurved laterally. Its anteriormost tip is worn. Measurements of both specimens are provided on Table 1.
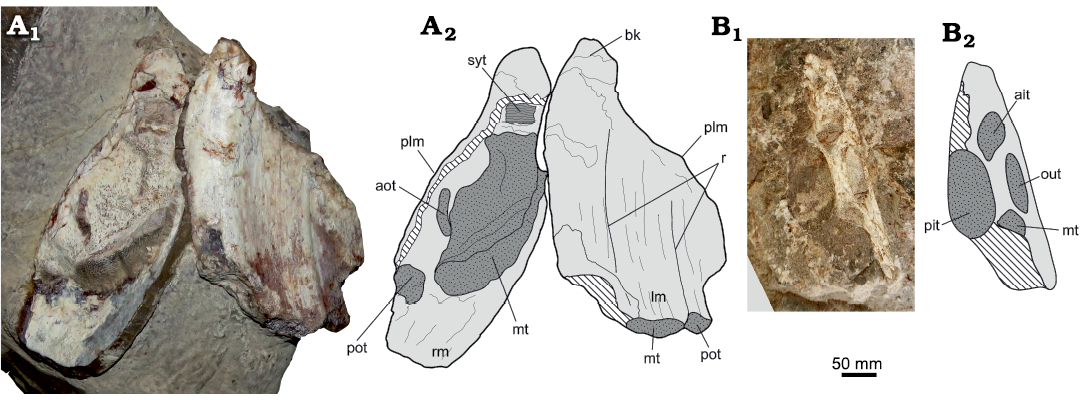
Fig. 3. Chimaeroid fishes from Mina Del Fierro, Andes Cordillera, Región del Maule, Chile; Baños del Flaco Formation, Tithonian (Upper Jurassic). A. Ischyodus townsendi (Buckland, 1835) (MHNC.35.444); two articulated mandibular plates in basal view. B. Ischyodus sp. (MHNC.35.445); ventral view of the bony contour of a right palatine attached to the matrix, preserving the occlusal surface of the tritors. Abbreviations: ait, anterior inner tritor; aot, anterior outer tritor; bk, beak; lm, left mandibular; mt, median tritor; out, outer tritor; pit, posterior inner tritor; plm, prominence of the labial margin; pot, posterior outer tritor; r, ridges; rm, right mandibular; syt, symphyseal tritor. Photographs (A1, B1), schematic illustrations (A2, B2).
Table 1. Measurements (in mm) of the two studied specimens. Mesiodistal length definition follows Popov and Machalski (2014).
|
Taxon |
Specimen number |
Plate |
Mesiodistal length |
|
Ischyodus townsendi |
MHNC.35.444 |
left mandibular |
995 |
|
Ischyodus sp. |
MHNC.35.444 |
right mandibular |
102 |
|
MHNC.35.445 |
right palatine |
721 |
Remarks.—MHNC.35.444 can be referred to the genus Ischyodus based on the possession of mandibular plates with a large central median tritor flanked by a short anterior outer tritor, an elongate posterior outer tritor, and the existence of a symphyseal tritor of laminated pleromin at the mesial tip of the tooth plate (Stahl 1999; Popov and Machalski 2014). The occurrence of two small tritors exposed at the prominence on the wavy labial margin of the mandibular plate has been considered as another diagnostic feature of the genus (Stahl 1999: 131); however, this feature is not present in the holotype of Ischyodus townsendi, which only has a single tritor (anterior outer) in this position. Nevertheless, the absence of a second tritor is related to the wearing suffered in larger/old mandibular plates, being instead a consequence of the ontogeny rather than being a diagnostic value (Ward and Grande 1991; Popov and Machalski 2014).
Stratigraphic and geographic.—Baños del Flaco Formation, Tithonian (Upper Jurassic). Mina Del Fierro, Andes Cordillera, Región del Maule, Chile.
Ischyodus sp.
Fig. 3B.
Material.—MHNC.35.445, an isolated, fragmentary right palatine plate from Mina Del Fierro, Andes Cordillera, Región del Maule, Chile; Baños del Flaco Formation, Tithonian (Upper Jurassic).
Description.—MHNC.35.445 is an isolated and poorly preserved palatine plate attached to the matrix. The basal surface is lost, allowing us to see the internal part of the tritors. The labial and symphyseal contour are preserved as a cast in the sandstone matrix, allowing us to observe the triangular shape of the plate. The posterior inner tritor is suboval and it is the larger tritor in the plate. The labial margin shows three aligned tritors, identified as fragments of the median tritor, the outer tritor, and the anterior inner tritor.
Remarks.—MHNC.35.445 possesses a large, oval posterior inner tritor, a median tritor likely smaller, an outer tritor axially elongated, and an anterior inner tritor slightly rounded. These features are present in different species of the Ischyodus (Stahl 1999: fig. 138A, C; Popov and Machalski 2014: fig. 4D1). The incompleteness of this mandibular plate precludes a more specific identification.
Discussion
Late Jurassic records of Ischyodus and comparison with the studied specimen.—Twenty-two nominal species within Ischyodus have been described from the Jurassic of Europe, with eight species known in the Tithonian (Table 2). However, more recent reviews (Stahl 1999; Duffin 2004; Lauer et al. 2019) have reduced this initial great diversity. Tithonian species from Germany initially included Ischyodus schuebleri Quendstedt, 1858, Ischyodus suevicus Philippi, 1897, Ischyodus quendstedti Wagner, 1857, Ischyodus rostratus Meyer, 1859, and Ischyodus acutus Meyer, 1859. From these, Ischyodus rostratus and I. suevicus were synonymized to I. schuebleri by Riess (1887) and by Heimberg (1949), respectively. The most informative Tithonian species from Germany is represented by I. quendstedti, which is based in a complete holotype and several referred specimens, including well preserved mandibular plates (Leuzinger et al. 2017).
Table 2. List of Jurassic species within the Ischyoudus Egerton, 1843. Modified from Stahl (1999).
|
Species |
Age |
Geographic occurrences |
References |
|
Ischyodus aalensis |
Bajocian |
Württemberg, Germany |
|
|
Ischyodus personati |
Bajocian |
Württemberg, Germany |
|
|
Ischyodus colei |
Bathonian |
England |
Agassiz 1837, tome III |
|
Ischyodus curvidens |
Bathonian |
England |
|
|
Ischyodus ferrugineus |
Bathonian |
Württemberg, Germany |
|
|
Ischyodus emarginatus |
Bathonian |
England |
|
|
Ischyodus obruchevi |
“Middle Jurassic” |
Russia |
|
|
Ischyodus egertoni |
Callovian–Kimmeridgian |
south England |
|
|
Ischyodus dufrenoyi |
Kimmeridgian |
Boulogne, France |
|
|
Ischyodus baeumonti |
Kimmeridgian |
Boulogne, France |
|
|
Ischyodus beaugrandi |
Kimmeridgian |
Boulogne, France |
|
|
Ischyodus bouchardi |
Kimmeridgian |
Boulogne, France |
|
|
Ischyodus suprajurensis |
Kimmeridgian |
Boulogne, France |
|
|
Ischyodus sauvaugei |
Kimmeridgian |
Boulogne, France |
|
|
Ischyodus schuebleri |
Tithonian |
Württemberg, Germany |
|
|
Ischyodus suevicus |
Tithonian |
Württemberg, Germany |
|
|
Ischyodus quendstedti |
Tithonian |
Bavaria, Germany |
|
|
Ischyodus rostratus |
Tithonian |
Hannover, Germany |
|
|
Ischyodus acutus |
Tithonian |
Hannover, Germany |
|
|
Ischyodus dutertrei |
Tithonian |
Boulogne, France |
|
|
Ischyodus avitus |
Tithonian |
Solnhofen, Germany |
|
|
Ischyodus townsendi |
Tithonian |
south England |
Duffin (in Stahl 1999) considered I. rostratus, I. suevicus, and I. schuebleri to be synonyms of I. quendstedti. Besides the latter, the remaining species I. acutus was based in a unique known vomerine (Meyer 1859), making any direct comparison to the Chilean specimen impossible. Finally, Ischyodus avitus (Meyer, 1862) from the Tithonian of Solnhofen, Germany, is known by several fairly complete specimens, but their dentition was not sufficiently exposed for a good description (Stahl 1999). This species was later referred to Ischyodus avitus (Riess, 1887). More recently, Duffin (2004) and Lauer et al. (2019) transferred the species to the genus Elasmodectes Newton, 1878. Therefore, the currently valid known Ischyodus species from the Tithonian of Germany can be reduced to only two species, Ischyodus quendstedti and I. acutus, the latter known by a single vomerine, rising reasonable doubts about its eventual synonymy with I. quendstedti.
Additional Tithonian records of Ischyodus are known from France and England, represented by Ischyodus dutertrei Egerton, 1843, and Ischyodus townsendi (Buckland, 1835), respectively. Agassiz (1837, tome III) mentioned I. dutertrei “as very similar to I. townsendi, but being one third smaller. The exterior [= labial margin] of the lower mandible [= mandibular plate] is not strongly grooved as in Ischyodus townsendi; [...] the most evident difference lays in the obtuse angle formed by the tooth crown [= beak] with the upper board of the mandible [= posterior part of the labial margin]. In other words, the tritorial surfaces are used in a more obliquous position in its mandibles [= mandiblar plates]”.
The description of Agassiz (1837, tome III) indicated that the occlusal curvature of the plate was more pronounced in I. townsendi than in I. dutertrei. This feature, plus the larger size of the former, could be better explained by the different ontogenetic stages of each specimen. Otherwise, the topology of the tritors is very similar in both species (see Sauvage 1896: pl. 21: 1, 2).
Comparison of MHNC.35.444 with the I. townsendi (holotype, BMNH P474; currently under acronym and numeration NHMUK 010039966 PV), I. quendstedti (referred, MJSN SCR010-1000), and I. dutertrei (holotype, specimen number 3402, Musée Boulogne-sur-le-mer, France) revealed a very similar topology of the tritorials in all cases (Fig. 4). Different plate outlines are explainable due to their respective ontogenetic stages. Ward and Grande (1991: fig. 4) brilliantly illustrated how wearing by occlusion and growth define the outline of the mandibular plates of the Ischyodus. The strictly ontogenetic changes reported by these authors are reflected in the enlargement of each tritor pad and the separation between tritors coupled to the plate growth. Wearing can explain the reduction or even the loss of the small tritors located close to the labial margin and the reduction of the beak sharpness.
With all these considerations, MHNC.35.444 shares with I. townsendi, I. dutertrei, and I. quendstedti the presence of a large median tritor, flanked labially by the posterior outer tritor. It differs from I. dutertrei, which lacks the anterior outer tritor (although, this could be a consequence of the wearing). Based on the referred specimen (MJSN SCR010-1000; Leuzinger et al. 2017), the anterior outer tritor of I. quendstedti is placed anterolateraly to its median tritor. In I. townsendi (holotype) and in the Chilean specimen, the anterior outer tritor is placed laterally to their respective median tritor. This condition cannot be assessed in I. dutertrei (holotype) because the anterior outer tritor is not present. Also, the posterior outer tritor of I. quendstedti (referred specimen) is well-exposed in occlusal view. Older individuals of the genus show this tritor embedded within the plate, which is the case of the Ischyodus spp. mandibular plates compared on Fig. 4. On the lingual margin, the Chilean specimen, I. dutertrei (holotype) and I. townsendi (holotype) have partial exposition of the posterior outer tritor, seen in cross-section, and closely placed lateral to the median tritor.
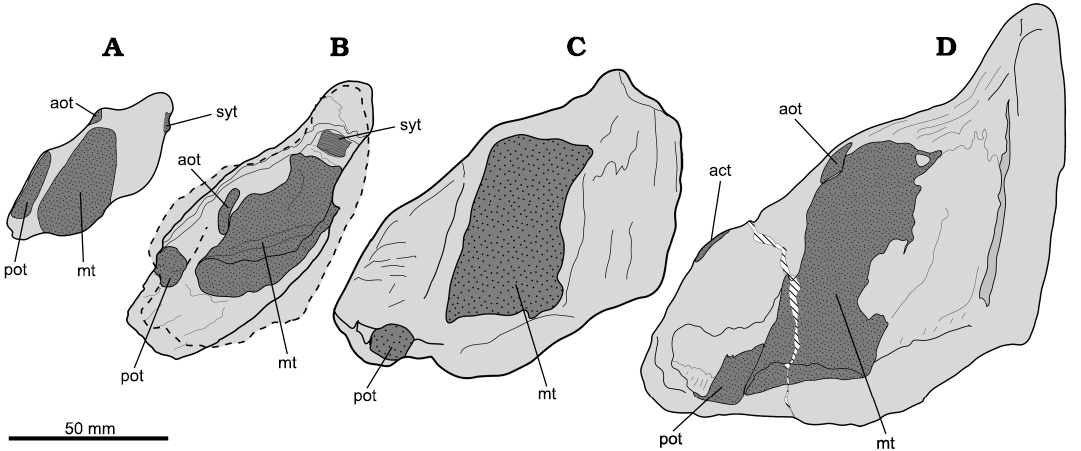
Fig. 4. Comparison of mandibular plates of Tithonian species within Ischyodus Egerton, 1843. All in occlusal view. A. Ischyodus quendstedti, MJSN SCR010-1000), Tithonian of Switzerland; right mandibular, mirrored for comparison (modified from Leuzinger et al. 2017). B. Ischyodus townsendi (MHNC.35.444), Tithonian of central Chile; outline of the left mandibular (only visible in basal view) is overlapped in dashed lines for a better understanding of the complete plate outline. C. Ischyodus dutertrei (holotype, specimen number 3402, Musée Boulogne-sur-le-mer, France), Kimmeridgian–Tithonian of France (modified from Sauvage 1896). D. Ischyodus townsendi (holotype, BMNH P474; currently NHMUK 010039966 PV), Tithonian of England (modified from Natural History Museum, London, Data Portal, data.nhm.ac.uk, https://doi.org/10.5519/0002965). Abbreviations: act, accessory tritor; aot, anterior outer tritor; mt, median tritor; pot, posterior outer tritor; syt, symphyseal tritor.
Under these considerations, I. dutertrei and I. townsendi could likely belong to the same species, as first pointed out by Agassiz (1833–1843) himself. However, a review of the valid species of the genus elsewhere is out of the scope of this study. Nevertheless, the completeness of the I. townsendi holotype compared with other Tithonian species, allows us to establish more detailed comparisons. The presence in I. townsendi of an anterior outer tritor laterally placed relative to the median tritor (contrary to the anterior outer tritor placed anterior to the median tritor in I. quendstedti) in effect occurs in the Chilean specimen. Following Grande and Ward (1991), the ontogenetic stage can cause lateral separation of the tritors, but a rostral migration seems more unlikely. Based on these traits, we refer MHNC.35.444 to the species I. townsendi.
Records of Austral Mesozoic Chimaeriformes.—The studied material represents the first Late Jurassic Chimaeriformes recorded in South America. Previous to this research, the oldest austral Callorhinchidae were represented by Aptian–Albian records from Queensland, Australia, including Ischyodus cf. thurmanni Pictet and Campiche, 1858 (Popov 2011; Popov and Machalski 2014); Ptyktoptychion tayyo Lees, 1986, and Pyctoptychion wadeae Bartholomai, 2008. In addition, Albian–Cenomanian records from western Australia include material referred to Callorhinchus cf. borealis and Ischyodus sp. (Popov 2011; Popov and Machalski 2014). During the Late Cretaceous, the genus Ischyodus was recorded in the Campanian–Maastrichtian of New Zealand (Pictet and Campiche 1858), and in the upper Maastrichtian of Seymour Island, Antarctica (Stahl and Chatterjee 2002). It is also documented during the Paleogene of Antarctica and southern South America (Ward and Grande 1991; Kriwet and Gaździcki 2003; Otero et al. 2013a; Otero and Soto-Acuña 2015). The material from central Chile studied here represents the oldest Chimaeriformes known to date in the Southern Hemisphere and the oldest known occurrence of the group in Gondwana.
Paleobiogeography.—The presence of the Ischyodus in the Tithonian of central Chile adds to a growing body of evidence regarding the interchange of marine vertebrates between the northern Tethys and the southeastern Pacific. Gasparini et al. (2007) synthesized the Middle and Late Jurassic record of ichthyosaurs, thalattosuchians, plesiosaurians, and Testudines from the Neuquén Basin (Argentina and Chile), suggesting that the common occurrence of several genera in the northern Tethys and the southeastern Pacific are likely explained as the result of an interchange through the Caribbean Seaway (sensu Iturralde-Vinent 2003). In addition, Arratia (1994, 2015) reviewed the osteichthyan fossil record from Chile, where phylogenetic analyses endorse a narrow relationship between the osteichthyan faunas from Europe, Cuba, and northern Chile, pointing out the Caribbean Seaway as the main path for such interchange. In this sense, the material here studied represents the first paleobiogeographically valuable evidence, now including chondrichthyans to the proposed connection. Previously, there was a single known record of an indeterminate neoselachian from the Tithonian of central Chile, likely representing a palaeospinacid or a hybodontid (Suárez and Otero 2011). Its still unsolved generic status precludes a palaeobiogeographic approach.
Conclusions
We present two new specimens of the Ischyodus (Chimaeriformes) from Tithonian levels of the Baños del Flaco Formation of central Chile. Available elements include one partial specimen comprising two articulated mandibular plates of a single individual, preserving enough diagnostic traits for an alpha-taxonomic identification. The specimen is here referred to Ischyodus townsendi (Buckland, 1835), a species previously restricted to the Tithonian of England. The common presence of this taxon in the northern Tethys and now in the southeastern Pacific reinforces the Caribbean Seaway as a fully functional path of marine faunal interchange, at least during the Late Jurassic. Evidence of this interhemispheric marine faunal interchange was already noted during the Middle and Late Jurassic on several groups of marine vertebrates (i.e., thalattosuchians, ichthyosaurians, plesiosaurians and osteichthtyes), although, previous to this research, no evidence of such interchange was verified among chondrichthyans. The studied material represents the first record of I. townsendi in the Southern Hemisphere, the first find of a Chimaeriformes in the Upper Jurassic of Gondwana, and the oldest known callorhinchid from South America.
Author’s contributions
PSH, survey of the collecting site, detailed stratigraphy, stratigraphic column, geologic setting, manuscript writing; SFC, survey of the collecting site, manuscript writing and review; CFB, survey of the collecting site, data processing, manuscript writing and review. AVT, data processing, manuscript writing and review; CSG, survey of the collecting site, complementary stratigraphy, data processing, manuscript writing; RAO, preparation of the material, conceptualization of the research, methods, anatomical identification and taxonomic determinations, elaboration of pictures and schemes, manuscript writing and review.
Acknowledgements
The authors thank Sergio Bogan (Universidad Maimónides, Buenos Aires, Argentina), to Jaime Villafaña (University of Vienna, Austria) and to Editor Daniel Barta (Oklahoma State University, Tahlequah, USA) for their valuable reviews and comments which improved our manuscript. Fieldwork resulting in this research was supported by Minera Río Teno S.A. Authorization for paleontologic management in Chile was granted by Consejo de Monumentos Nacionales (National Monuments Council, Chile) on Ord. CMN N° 2275/17. Access to laboratory and research for RAO, CSG, and PSH was supported by the project “New data sources on the fossil record and evolution of vertebrates”, Anillos de Ciencia y Tecnología-ACT-172099, ANID (formerly Conicyt)-Chile. AVT was funded by ANID PCHA/Becas Chile, Doctoral Fellowship; Grant No. 2016-72170286 and a Peter Buck Predoctoral Fellowship, National Museum of Natural History, Smithsonian Institution. All fieldwork and research was permanently supported by Consultora Paleosuchus Ltda.
References
Agassiz, L. 1833–1843. Recherches sur les Poissons fossiles. Tome III. 448 pp. Petitperriere, Neuchâtel.
Arratia, G. 1994 Phylogenetic and paleobiogeographic relationships of the varasichthyid group (Teleostei) from the Late Jurassic of central and South America. Revista Geológica de Chile 21: 119–161.
Arratia, G. 2015. Los peces osteíctios fósiles de Chile y su importancia en los contextos paleobiogeográfico y evolutivo. In: D. Rubilar-Rogers, R.A. Otero, A. Vargas, and M. Sallaberry (eds.), Vertebrados Fósiles de Chile. Publicación Ocasional del Museo Nacional de Historia Natural (Chile) 63: 35–83.
Averianov, A.O. 1992. New Jurassic chimeroids of Russia. Paleontological Journal 3: 57–62.
Bartholomai, A. 2008. Lower Cretaceous chimaeroids (Chondrichthyes: Holocephali) from the Great Artesian Basin, Australia. Memoirs of the Queensland Museum 52: 49‒56.
Buckland, W. 1835. A notice on the fossil beaks of four extinct species of fishes, referrible to the genus Chimaera, what occurs in the oolitic and Cretaceous formations of England. Proceedings of the Geological Society of London 2: 205‒206.
Charrier, R., Wyss, A., Flynn, J., Swisher, C., Norell, M., Zapatta, F., McKenna, M., and Novaceck, M. 1996. New evidence for late Mesozoic–early Cenozoic evolution of the Chilean Andes in the upper Tinguiririca valley (35º S), Central Chile. Journal of South American Earth Sciences 9: 1–30. Crossref
Covacevich, V., Varela, J., and Vergara, M. 1976. Estratigrafıa y Sedimentación de la Formación Baños del Flaco al sur del Rıo Tinguiririca, Cordillera de los Andes. Provincia de Curicó, Chile. Actas I Congreso Geologico Chileno, 191‒211. Santiago.
Duffin, C.J. 2004. Plattenkalk Late Jurassic chimaeroids. In: F.J. Poyato-Ariza (ed.), Fourth International Meeting on Mesozoic Fishes—Systematics, homology, and nomenclature, 83‒86. Miraflores de la Sierra, Madrid.
Egerton, P.M.G. 1843. On some new species of fossil chimaeroid fishes, with remarks on their general affinities. Proceedings of the Geological Society of London 4: 153–157.
Garman, S. 1901. Genera and families of the chimaeroids. Proceedings of the New England Zoological Club 2: 75–77.
Gasparini, Z., Fernández, M., De La Fuente, M., and Salgado, L. 2007. Reptiles marinos jurásicos y cretácicos de la Patagonia Argentina: su aporte al conocimiento de la herpetofauna mesozoica. Asociación Paleontológica Argentina. Publicación Especial Ameghiniana 50º aniversario 11: 125‒136.
Hamy, M.E.T. 1866. Note sur une nouvelle espece d’Ischyodus de l’argile kimmeridgienne de Chatillon, pres de Boulogne-sur-Mer. Bulletin della Societé Geológiqué de France 2 (Series XXIII): 654‒657.
Heimberg, G. 1949. Neue Fischfunde aus dem Weissen Jura von Wurttemberg. Palaeontographica 97: 75‒98.
Iturralde-Vinent, M. 2003. The conflicting paleontologic versus stratigraphic record of the formation of the Caribbean Seaway. In: C. Bartolini, R.T. Buffler, and J. Blickwede (eds.), The Circum-Gulf of Mexico and the Caribbean: Hydrocarbon Habitats, Basin Formation, and Plate Tectonics. American Association Petroleum Geology, Tulsa OK, Memoire 79: 75–88. Crossref
Klohn, G. 1960. Geología de Santiago, O’Higgins, Colchagua y Curicó. Instituto de Investigaciones Geologicas Chile. Boletin 8: 1‒95.
Kriwet, J. and Gaździcki, A. 2003. New Eocene Antarctic chimaeroid fish (Holocephali, Chimaeriformes). Polish Polar Research 24: 29‒51.
Lauer, B., Lauer, R., Bernard, E., Duffin, C., Popov, E., and Ward, D. 2019. Observations on the Mesozoic chimaeroid, Elasmodectes Newton, 1878. In: A. Farke, A. MacKenzie, and J. Miller-Camp (eds.), 79th Annual Meeting of the Society of Vertebrate Paleontology, Meeting Program, 138. Society of Vertebrate Paleontology, Brisbane.
Lees, T.A. 1986. A new chimaeroid Ptyktoptychion tayyo gen. et sp. nov. (Pisces: Holocephali) from the marine Cretaceous of Queensland. Alcheringa 10: 187‒193. Crossref
López-Arbarello, A. 2004. The record of Mesozoic fishes from Gondwana (excluding India and Madagascar). In: G. Arratia and A. Tintori (eds.), Mesozoic Fishes 3—Systematics, Paleoenvironments and Biodiversity. 597‒624. Verlag Dr. Friedrich Pfeil, München.
Lopez-Arbarello, A., Rauhut, O.W.M., and Moser, K. 2008. Jurassic fishes of Gondwana. Revista de la Asociacion Geologica Argentina 63: 586‒612.
Leuzinger, L., Cuny, G., Popov, E., and Billon-Bruyat, J.P. 2017. A new chondrichthyan fauna from the Late Jurassic of the Swiss Jura (Kimmeridgian) dominated by hybodonts, chimaeroids and guitarfishes. Papers in Palaeontology 3: 471‒511. Crossref
Meyer, H. 1859. Fossile chimaeriden aus dem Portland von Hannover. Palaeontographica 7: 14‒18.
Meyer, H. 1862. Chimaera (Ganodus) avita aus dem Lithographischen Schiefer von Eichstadt. Palaeontographica 10: 87‒95.
Newton, E.T. 1878. The chimaeroid fishes of the British Cretaceous rocks. Memoirs of the Geological Survey of the United Kingdom 4: 1‒62.
Otero, R.A. and Soto-Acuña, S. 2015. New chondrichthyans from Bartonian–Priabonian levels of Río de Las Minas and Sierra Dorotea, Magallanes Basin, Chilean Patagonia. Andean Geology 42: 268‒283. Crossref
Otero, R.A., Oyarzún, J.L., Soto-Acuña, S., Yury-Yáñez, R., Gutierrez, N., Le Roux, J., Torres, T., and Hervé, F. 2013a. Neoselachians and Chimaeriformes (Chondrichthyes) from the Upper Cretaceous–Paleogene of Sierra Baguales, southernmost Chile. Chronostratigraphic, paleobiogeographic and paleoenvironmental implications. Journal of South American Earth Sciences 48: 13‒30. Crossref
Otero, R.A., Rubilar-Rogers, D., Yury-Yañez, R.E., Vargas, A.O., Gutstein, C.S., Mourgues, F.A., and Robert, E. 2013b. A new species of chimaeriform (Chondrichthyes, Holocephali) from the uppermost Cretaceous of the López de Bertodano Formation, Isla Marambio (Seymour Island), Antarctica. Antarctic Science 25: 99‒106. Crossref
Patterson, C. 1965. The phylogeny of the chimaeroids. Philosophical Transactions of the Royal Society of London B249: 101‒219. Crossref
Philippi, E. 1897. Ueber Ischyodus suevicus nov. sp. Ein Beitrag zur Kenntnissder fossilen Holocephalen. Palaeontographica 44: 1‒10.
Pictet, F.J. and Campiche, G. 1858. Description des fossiles du terrain Crétacé des environs de Sainte-Croix. Matériaux pour la Paléontologie Suisse: 1–380.
Popov, E.V. 2011. New data on chimaeroid fishes (Holocephali; Chimaeroidei) from the Cretaceous and Neogene of Australia. In: O. Lebedev and A. Ivanov (eds.), II International Obruchev Symposium “Paleozoic Early Vertebrates”, Russia, Abstract Volume, 40–41. Alexander Ivanov Publisher, Saint Petersburg.
Popov, E.V. and Machalski, M. 2014. Late Albian chimaeroid fishes (Holocephali, Chimaeroidei) from Annopol, Poland. Cretaceous Research 47: 1‒18. Crossref
Quenstedt, F.A. 1858. Der Jura. 842 pp. Laupp, Tübingen.
Quinzio, L.A. and Varela, N. 2015. Caracterización Paleontológica Mina del Fierro. Proyecto Mina del Fierro de Minera Río Teno S.A. Región del Maule. Report for the Consejo de Monumentos Nacionales (National Heritage Council, Chile), ingreso CMN Nº5287-25/08/2015. 19 pp. Santiago.
Riess, J. 1887. Ueber einige fossile Chimeriden-Reste im Münchenerpalaeontologischen Museum. Palaeontographica 34: 1‒27.
Salazar, C. 2012. The Jurassic–Cretaceous Boundary (Tithonian–Hauterivian) in the Andean Basin, Central Chile: Ammonite fauna, Bio- and Sequence Stratigraphy and Palaeobiogeography. 388 pp. Doctoral Thesis, Universität Heidelberg.
Salazar, C. and Stinnesbeck, W. 2015. Redefinition, stratigraphy and facies of the Lo Valdés Formation (Upper Jurassic-Lower Cretaceous) in central Chile. Boletín del Museo Nacional de Historia Natural, Chile 64: 41–68.
Sauvage, H.E. 1867. Poissons fossiles des formations secondaires du Boulonnais. Mémoires de la Société Académique de l’Arrondissement de Boulogne-sur-Mer 2: 53–152.
Sauvage, H.E. 1896. Les Ischyodus des terrains Jurassiques superieurs du Boulonnais. Bulletin de la Societé Geológiqué du France 24: 456‒465.
Stahl, B.J. 1999. Handbook of Paleoichthyology, Vol. 4: Chondrichthyes III. Holocephali. 164 pp. Verlag Dr. Friedrich Pfeil, München.
Stahl, B.J. and Chatterjee, S. 2002. A Late Cretaceous callorhinchid (Chondrichthyes, Holocephali) from Seymour Island, Antarctica. Journal of Vertebrate Paleontology 22: 848‒850. Crossref
Suárez, M.E. and Otero, R.A. 2011. First record of a neoselachian shark in the Jurassic of Chile. Resúmenes del IV Congreso Latinoamericano de Paleontología de Vertebrados, San Juan Argentina. Ameghiniana 48 (Suplemento): R213.
Wagner, J.A. 1857. Charakteristik neuer Arten von Knorpelfischen aus den lithographischen Schiefern der Umgegend von Solnhofen. Gelehrte Anzeigen der königlich bayerischen Akademie der Wissenschaften 44 (35–36): 288–293.
Ward, D.J. and Grande, L. 1991. Chimaeroid fish remains from Seymour Island, Antarctic Peninsula. Antarctic Science 3: 323‒330. Crossref
Acta Palaeontol. Pol. 66 (3): 623–630, 2021
https://doi.org/10.4202/app.00859.2020