
A new species of the “condylarth” Hyopsodus from the middle Eocene of the Erlian Basin, Inner Mongolia, China, and its biostratigraphic implications
BIN BAI, YUAN-QING WANG, XIN-YUE ZHANG, and JIN MENG
Bai, B., Wang, Y.-Q., Zhang, X.-Y., and Meng, J. 2021. A new species of the “condylarth” Hyopsodus from the middle Eocene of the Erlian Basin, Inner Mongolia, China, and its biostratigraphic implications. Acta Palaeontologica Polonica 66 (4): 767–777.
The “condylarth” genus Hyopsodus is diverse and abundantly represented in Eocene mammalian faunas of North America. In contrast, fossil specimens of Hyopsodus are rather sparse in Eurasia. Only four species of Hyopsodus are known from Asia and two from Europe, as compared to the 18 species of Hyopsodus described from North America. Here, we report a new species of Hyopsodus, Hyopsodus arshantensis sp. nov., from the middle part of the Arshanto Formation in the Erlian Basin, Inner Mongolia, China. The holotype and only specimen of the new species, a right mandible with m1–m2, exhibits a unique combination of characters on m1–m2 not present in other species of Hyopsodus, including a moderately lophodont crown, a long trigonid without a paraconid, an obliquely aligned protolophid, an angle between the cristid obliqua and the posthypocristid slightly greater than 90°, a midline position of the hypoconulid, and a relatively large entoconid. The m1–m2 morphology of H. arshantensis is intermediate between specimens of Hyopsodus from the Wasatchian and Uintan North American Land Mammal Ages (NALMA), and is comparable to that of Hyopsodus from the Bridgerian NALMA. Moreover, its relatively large size is near the size range present among the late Bridgerian species of Hyopsodus. Based on those similarities, in combination with a few fossil mammals from overlying layers, the middle part of the Arshanto Formation could be correlated in part to the late Bridgerian, and the upper part of the Arshanto Formation may bracket the time interval equivalent to the Bridgerian/Uintan boundary. That proposed correlation and somewhat different faunas recognized within the Arshanto Formation suggest that it may be necessary to subdivide the Arshantan Asian Land Mammal Age (ALMA) and/or redefine the Arshantan/Irdinmanhan ALMA boundary in future comprehensive studies.
Key words: Mammalia, Hyopsodus, “condylarth”, Eocene, Arshanto Formation, Erlian Basin, China.
Bin Bai [baibin@ivpp.ac.cn], Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing 100044, China; CAS Center for Excellence in Life and Paleoenvironment, Beijing 100044, China.
Yuan-Qing Wang [wangyuanqing@ivpp.ac.cn], Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing 100044, China; CAS Center for Excellence in Life and Paleoenvironment, Beijing 100044, China; College of Earth and Planetary Sciences, University of Chinese Academy of Sciences Beijing 100049, China.
Xin-Yue Zhang [zhangxinyue@ivpp.ac.cn], Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing 100044, China; CAS Center for Excellence in Life and Paleoenvironment, Beijing 100044, China; University of Chinese Academy of Sciences, Beijing 100049, China.
Jin Meng [jmeng@amnh.org], Division of Paleontology, American Museum of Natural History New York 10024, USA; Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing 100044, China; Earth and Environmental Sciences, Graduate Center, City University of New York, New York, 10016, USA.
Received 17 May 2021, accepted 30 September 2021, available online 1 December 2021.
Copyright © 2021 B. Bai et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The “condylarth” Hyopsodus is an archaic ungulate known from the early Eocene to the late Eocene (Archibald 1998). Hyopsodus is included in Hyopsodontidae, a likely polyphyletic group comprising 20 small-sized Paleocene and Eocene genera (Zack et al. 2005b; Rose 2006; Halliday et al. 2017). Hyopsodontids were considered to be ancestral to artiodactyls (Simpson 1945), allied with afrotheres (Godinot et al. 1996; Zack et al. 2005a) or related to South American ungulates (Cifelli 1983; Muizon and Cifelli 2000). Hyopsodus is indeed abundant and diverse in early–middle Eocene North American faunas, and comprises up to 39% of the identified specimens from the Bridger Formation in the US National Museum of Natural History collection (Gazin 1976; West 1979). A brief revision of Hyopsodus recognizes 16 valid species from North America (Archibald 1998), but that work omitted H. lovei from the earliest Uintan North American Land Mammal Age (Flynn 1991). Taking advantage of the abundance and stratigraphic distribution of Hyopsodus, it has been used in evolutionary studies (Gingerich 1974, 1976; Redline 1997) and stable isotope analyses to reconstruct paleoenvironments in combination with other mammalian groups (Secord et al. 2008). More recently, the cranial endocast and inner ear morphology of Hyopsodus have been reconstructed through the use of X-ray computed microtomography (Orliac et al. 2012; Ravel and Orliac 2015). While it is well known from North American deposits, Hyopsodus is much rarer in the Eocene deposits of Asia and Europe. Six species of Hyopsodus and Asiohyopsodus confuciusi (represented by fragmentary maxillae and mandibles) have been reported from the early–middle Eocene of Eurasia (Hooker and Dashzeveg 2003; Tong and Wang 2006), and the validity of some of those Asian species assigned to Hyopsodus is controversial (Kondrashov and Lucas 2004).
Here we report a new species of Hyopsodus and an indeterminate species of Hyopsodus from the early middle Eocene Arshanto Formation in the Erlian Basin, Inner Mongolia, China. These two fragmentary mandibles represent the first record of Hyopsodus from the Arshanto Formation, which is the basis of the Arshanto fauna and the Arshantan Asian Land Mammal Age (Wang et al. 2019b; Speijer et al. 2020). Based on comparisons with other species of Hyopsodus from North America and Eurasia, we further investigate its biostratigraphic implications and propose a probable new correlation between Arshantan and Bridgerian/Uintan NALMA.
Institutional abbreviations.—ACM, Amherst College Museum (Pratt Museum), Amherst, USA; AMNH FM, American Museum of Natural History, Fossil Mammals, New York, USA; IVPP V, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China.
Other abbreviations.—NALMA, North American Land Mammal Ages; ALMA, Asian Land Mammal Age. We follow standard convention in abbreviating tooth families as I, C, P, and M, with upper and lower case letters referring to upper and lower teeth, respectively.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in ZooBank: urn:lsid:zoobank.org:pub:F753C4DF-C373-41DB-BE98-B9B2B7AE3C36
Material and methods
The holotype specimen of the new species (IVPP V 28282) was unearthed from the middle part of the Arshanto Formation at Chaganboerhe in the Erlian Basin, Inner Mongolia, China. The holotype comes from a brownish-red silty mudstone with small calcareous nodules, which is equivalent to the mammalian horizon AS-4 of the Arshanto Formation (Wang et al. 2010). The second specimen identified as Hyopsodus sp. (IVPP V 28283) was collected from the upper level of the lower part of the Arshanto Formation at Nuhetingboerhe. The stratigraphic horizon that produced the second specimen is a brownish-red, muddy siltstone, and is equivalent to the mammalian horizon AS-2 of the Arshanto Formation (Wang et al. 2010). Three species of Hyopsodus previously reported from China are compared with the new material and figured, including H. turpanensis, H. fangxianensis, and H. huashigouensis.
X-ray micro-computerized tomography. The scanning was carried out using the 225 kV micro-CT scanner facility (developed by the Institute of High Energy Physics, Chinese Academy of Sciences) at the Key Laboratory of Vertebrate Evolution and Human Origins, Chinese Academy of Sciences (Wang et al. 2019a). The specimens were scanned with beam energy of 120 kV and a flux of 120 µA. The resolution per pixel for IVPP V 4355 (holotype of H. turpanensis) and IVPP V 12005 (holotype of H. fangxianensis) are 21.956 µm and 18.820 µm, respectively. A 360° rotation with a step size of 0.5° was used. A total of 720 projections were reconstructed in a 2048×2048 matrix of 1536 slices using a two-dimensional reconstruction software developed by the Institute of High Energy Physics, Chinese Academy of Sciences. The three-dimensional reconstructions were created using the software VG Studio 3.2.
Systematic palaeontology
Order “Condylarthra” Cope, 1881
Family Hyopsodontidae Trouessart, 1879
Genus Hyopsodus Leidy, 1870
Type species: Hyopsodus paulus Leidy, 1870, Bridger Basin, Wyoming, USA, early–middle Eocene.
Hyopsodus arshantensis sp. nov.
Fig. 1A.
ZooBank LSID: urn:lsid:zoobank.org:act:79A0AB22-A930-4159-8F B8-BBD321777F76
Etymology: In the reference to the Arshanto Formation, where the new species was found.
Holotype: IVPP V 28282, a right mandible with m1–m2.
Type locality: Chaganboerhe, Erlian Basin, Inner Mongolia, China.
Type horizon: Upper level of the middle part of the Arshanto Formation (AS-4), Arshantan ALMA, middle Eocene.
Differential diagnosis.—Differs from other species of Hyopsodus by the combination of following characters of m1–m2: a moderate degree of lophodonty, a relatively longer trigonid without a paraconid, a moderately obliquely aligned protolophid, an angle between the cristid obliqua and posthypocristid slightly greater than 90°, a midline position of the hypoconulid on m2, a relatively larger entoconid, and absence of the cingulids on the buccal and lingual sides. Further differs from contemporaneous Bridgerian species of Hyopsodus in having a generally larger size and in lacking a variably developed metastylid. Further differs from H. marshi by a generally less robust appearance. Differs from Asiohyopsodus confuciusi in having more lophodont molar crowns, a more obliquely aligned protolophid, a more lingually directed cristid obliqua, a more distinct posthypocristid and postentocristid, a larger entoconid on m1–m2, and absence of the paraconid on m1.
Material.—Holotype only.
Description.—m1: The tooth is moderately worn and rectangular in outline with the talonid slightly wider than the trigonid (Table 1). The protoconid and metaconid are conical and obliquely arranged. The protolophid is shallowly notched, joining the posterior walls of the protoconid and metaconid. The paralophid descends from the protoconid anteriorly more than lingually, and then extends lingually to the anterobuccal base of the metaconid. The paraconid is absent, and the trigonid is relatively open and expanded rather than being anteroposteriorly compressed. The hypoconid is as high as the protoconid, extending the cristid obliqua anterolingually to the posterobuccal wall of the metaconid. The posthypocristid, which joins the hypoconid and hypoconulid, is posterolingually extended, and forms an angle of slightly less than 90° with the cristid obliqua at current wear stage. As a result, the hypoconid exhibits a selenodont appearance. The hypoconulid is deeply worn and relatively small, and is positioned closer to the entoconid than to the hypoconid. The entoconid is conical, obliquely arranged relative to the hypoconid, and relatively isolated with a narrow postentocristid joining the hypoconulid. A weak cingulid is present along the anterior border, and the posterior cingulid rises up to form the hypoconulid, with the buccal side of the hypoconulid much more distinct than the lingual one.
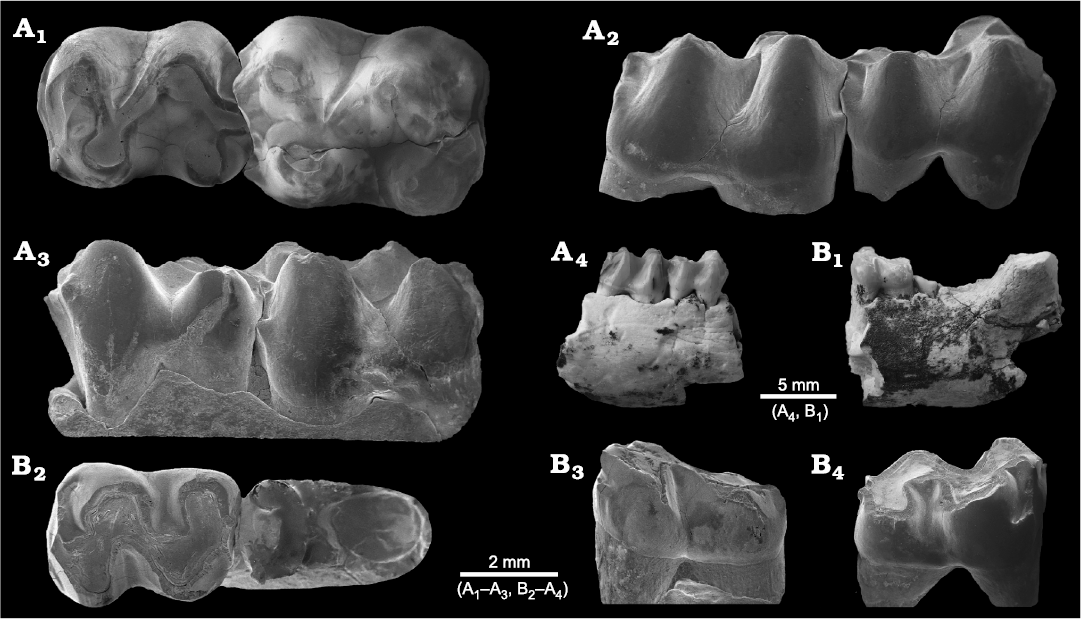
Fig. 1. The middle Eocene “condylarth” mammal Hyopsodus arshantensis sp. nov. (A) and Hyopsodus sp. (B) from the Arshanto Formation of the Erlian Basin, Inner Mongolia, China. A. IVPP V 28282 from Chaganboerhe, m1–m2 in occlusal (A1), buccal (A2), and lingual (A3) views; mandible in buccal view (A4). B. IVPP V 28283 from Huheboerhe, mandible in buccal view (B1); m2 and m3 root in occlusal (B2), buccal (B3), and lingual (B4) views.
Table 1. Measurements (in mm) of lower molars for some species of Hyopsodus from the Eocene of China. Abbreviations: ?, missing data; a, approximate; AW, anterior width; L, length; PW, posterior width.
| |
m1 |
m2 |
m3 |
||||||
|
Species |
L |
AW |
PW |
L |
AW |
PW |
L |
AW |
PW |
|
Hyopsodus arshantensis sp. nov. |
4.05 |
3.10 |
3.08 |
4.60 |
3.60 |
3.38 |
? |
? |
? |
|
Hyopsodus sp. (IVPP V 28283) |
? |
? |
? |
4.10 |
3.20 |
2.90 |
4.05a |
2.12a |
? |
|
Hyopsodus fangxianensis (IVPP V 12005) |
4.10 |
3.20 |
3.45 |
4.41 |
3.80 |
3.50 |
4.00 |
3.10 |
2.25 |
|
Hyopsodus turpanensis (IVPP V 4355) |
3.15 |
2.41 |
2.45 |
3.30 |
3.00 |
2.85 |
4.00 |
2.70 |
2.20 |
|
Hyopsodus huashigouensis (IVPP V 7921) |
? |
? |
? |
4.12 |
3.20 |
2.80a |
? |
? |
? |
m2: The tooth is slightly worn and rectangular in outline with the talonid slightly narrower than the trigonid. The m2 is larger than m1, but is similar to the latter in morphology. The m2 mainly differs from m1 in that: (i) the buccal wall of the hypoconid is lingually slanted, with the angle between the cristid obliqua and posthypocristid becoming narrower during wear, with the current slightly wear stage associated to an angle slightly greater than 90°; (ii) the hypoconulid is positioned near the midline of the talonid; and (iii) the postentocristid is more distinct, joining the entoconid and hypoconulid.
Stratigraphic and geographic range.— Type locality and horizon only.
Hyopsodus sp.
Fig. 1B.
Material.—IVPP V 28283, a left mandible with a heavily worn m2 and roots of m3 from the upper level of the lower part of the Arshanto Formation (AS-2), Huheboerhe, Erlian Basin, Inner Mongolia, China.
Description.—The m2 is about 11% smaller than that of Hyopsodus arshantensis sp. nov. (IVPP V 28282) in longitudinal dimensions (Table 1). It differs from H. arshantensis sp. nov. in having a more lingually extended paralophid, and the angle between the cristid obliqua and the posthypocristid is slightly less than 90°. Only the roots of m3 are preserved on the specimen. The m3 was almost as long as m2, but it appears to have been narrower than the latter. The cross section of the anterior root is an anteroposteriorly compressed oval, and that of the posterior one is an elongated ellipsoid.
Discussion
Comparison with North American Hyopsodus.—Hyopsodus was first described by Leidy (1870), and is among the earliest fossil mammal genera to be named from the Eocene of North America (Gazin 1968). The following comprehensive studies of Hyopsodus include those by Osborn (1902), Loomis (1905), and Matthew (1909, 1915). Gazin (1968) thoroughly reviewed the North American record of Hyopsodus and recognized 12 species as valid, including five species from the Wasatchian, five species from the Bridgerian, and two species from the Uintan. However, the taxonomy and validity of some species remain unclear with ongoing debates. Gingerich (1974) recognized 10 species of Hyopsodus from the Wasatchian and studied their changes in size, but Archibald (1998) considered those fossils to represent eight species. West (1979) reported only three species of Hyopsodus from the Bridger Formation, in contrast to the five species from the Bridgerian proposed by Gazin (1968). In terms of Uintan Hyopsodus, Krishtalka (1979) considered H. fastigatus as a junior synonym of H. uintensis, and erected a new species H. sholemi. By contrast, Storer (1984) considered H. fastigatus to be a valid species. Redline (1997) identified two anagenetically evolving “species lineages” of Hyopsodus from the Wasatchian and early Bridgerian: H. paulus and H. powellianus, which are divided into successive, informal segments. For the purposes of our work here, we mainly follow the taxonomy of Hyopsodus species utilized in Archibald’s (1998) brief revision of the genus (Fig. 2). In general, the morphology of the new Chinese fossil material is intermediate between that of the Wasatchian and Uintan species of Hyopsodus, being most comparable with those from the Bridgerian.
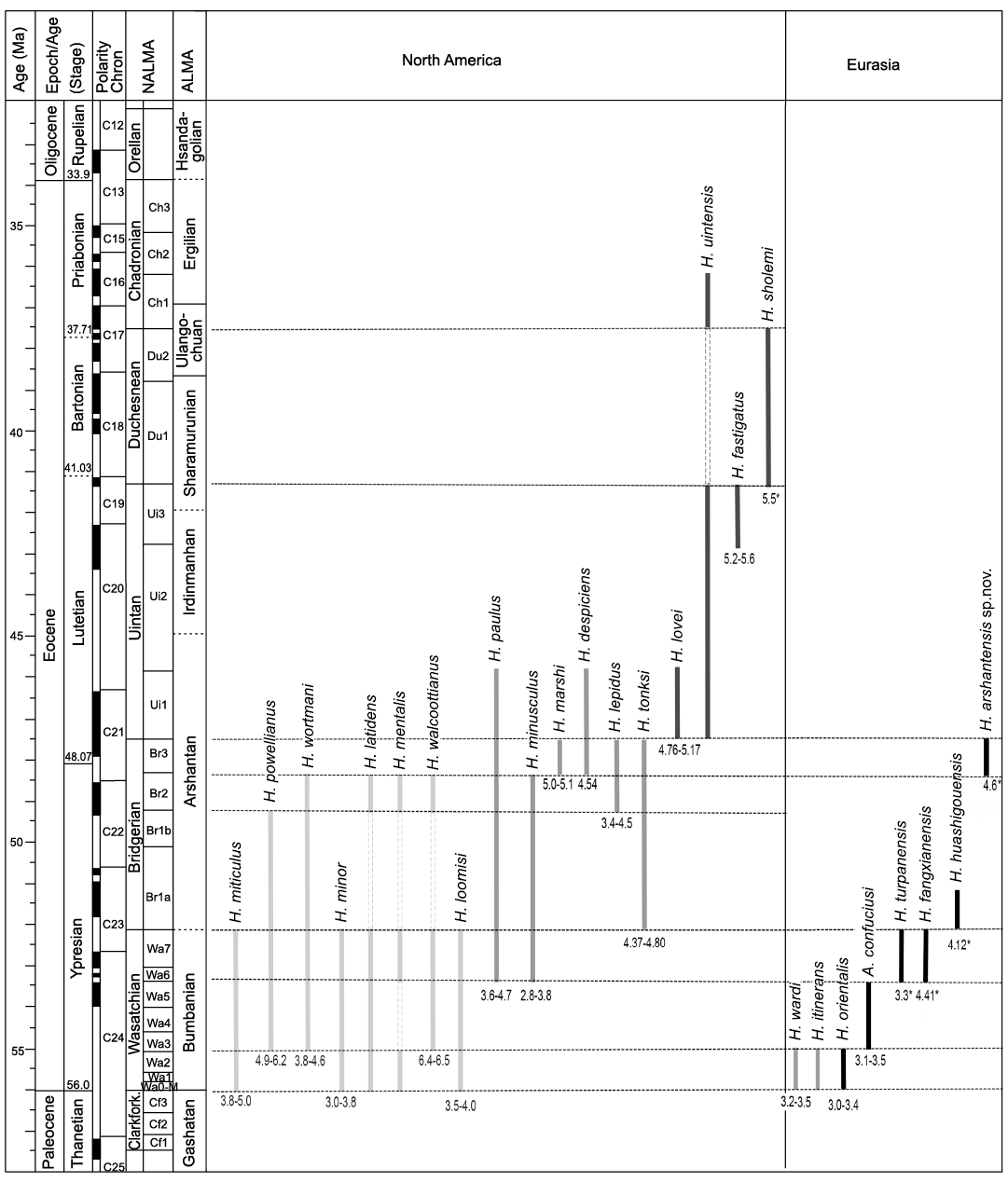
Fig. 2. Temporal distributions of species of Hyopsodus and Asiohyopsodus from North America and Eurasia based on Archibald (1998) and Tong and Wang (2006). The left column shows the correlation between NALMAs and ALMAs modified from Speijer et al. (2020) and Wang et al. (2019b). The numbers below the vertical bars indicate the range or the mean value for lower molar m2 length in millimeters. Numbers with an asterisk mean that the measurement is taken from a single specimen. The traditional Wasatchian, Bridgerian, and Uintan species of Hyopsodus are discriminated by different grey shading. Abbreviations: A., Asiohyopsodus; ALMA, Asian Land Mammal Age; Clarkfork., Clarkforkian; H., Hyopsodus; NALMA, North American Land Mammal Age.
The size of H. arshantensis sp. nov. falls within the ranges of those of H. wortmani (m2 length 3.8–4.6 mm) and H. miticulus (m2 length 3.8–5.0 mm) from the early Wasatchian to the middle Bridgerian, but H. arshantensis sp. nov. is smaller than H. powellianus (= H. browni and H. jacksoni; m2 length 4.9–6.2 mm) and H. walcottianus (m2 length 6.4–6.5 mm) from the middle Wasatchian to the middle Bridgerian (Figs. 2, 3) (Gazin 1968; Redline 1997; Archibald 1998; Tong and Wang 2006). Hyopsodus arshantensis sp. nov. is generally larger than other species of Wasatchian Hyopsodus, including H. loomisi (m2 length 3.5–4.0 mm), H. minor (m2 length 3.0–3.8 mm), H. latidens, and H. mentalis (Gazin 1968; Redline 1997; Archibald 1998). The m1–m2 of H. arshantensis sp. nov. differ from those of Wasatchian Hyopsodus in having more lophodont molar crowns, a relatively longer and more expanded trigonid, a shallower notch of the protolophid, an angle slightly greater than 90° between the cristid obliqua and posthypocristid, and a smaller hypoconulid. Moreover, some Wasatchian species further differ from H. arshantensis sp. nov. in having a paralophid rising up lingually to form a small paraconid (positioned high on the anterior slope of the metaconid on m1 and/or m2), a small entostylid anterior to the entoconid (Gazin 1968), and a distinct cingulid at the base of the ectoflexid.
Four species of Hyopsodus have been named from the Uintan to the Chadronian NALMA (Fig. 2). Hyopsodus lovei is known from the earliest Uintan (Flynn 1991), H. uintensis from the Uintan and early Chadronian (Osborn 1902; Krishtalka 1979; Archibald 1998), H. fastigatus from the late Uintan (Storer 1984), and H. sholemi from the Duchesnean (Krishtalka 1979). It is necessary to mention that the species H. paulus, H. despiciens, and H. lepidus, which occur mostly in the Bridgerian, are also known from the early Uintan (Archibald 1998; Murphey and Kelly 2017). The size of H. arshantensis sp. nov. is similar to that of H. uintensis (m1 length 4.1 mm; Osborn, 1902), but H. arshantensis sp. nov. is smaller than H. lovei (m2 length 4.76–5.17 mm; Flynn 1991), H. fastigatus (m2 length 5.2–5.6 mm; Storer 1984) and H. sholemi (m2 length 5.5 mm; Krishtalka 1979) (Fig. 3). The m1–m2 of typical Uintan Hyopsodus differ from the new species in the presence of the following derived features: a greater degree of lophodonty with a more crescentic protoconid and hypoconid; a more obliquely oriented trigonid; and a greatly enlarged and more isolated entoconid (Krishtalka 1979; Storer 1984; Flynn 1991). The m1–m2 of typical Uintan Hyopsodus differ further from H. arshantensis sp. nov. in having a hypoconulid placed close to the entoconid, and a metastylid positioned on the posterior slope of the metaconid (Russell and Wickenden 1933; Flynn 1991). The m1–m2 of H. arshantensis sp. nov. share some similarities with that of typical Uintan Hyopsodus in having a reduced hypoconulid and a relatively longer trigonid on m2. The p4–m3 of H. fastigatus and H. sholemi are also characterized by being more hypsodont (Storer 1984).
Gazin (1968) considered five species of Hyopsodus to be valid and present mainly during the Bridgerian, including H. paulus from the late Wasatchian to early Uintan, H. minusculus from the late Wasatchian to middle Bridgerian, H. lepidus from the middle Bridgerian to early Uintan, H. marshi from the late Bridgerian, and H. despiciens from the late Bridgerian to early Uintan (Archibald 1998; Tong and Wang 2006; Murphey and Kelly 2017). However, West (1979) recognized only three species of Hyopsodus from the Bridgerian, and proposed both H. marshi and H. despiciens as junior synonyms of H. paulus. By contrast, H. marshi and H. despiciens still are considered as valid by some authors (Archibald 1998; Tsukui 2016; Murphey and Kelly 2017), and this viewpoint is followed here (Fig. 2). The species of Bridgerian Hyopsodus can be differentiated mainly on the basis of their relative sizes and stratigraphic distributions (Storer 1984), or by their cingulids (Tsukui 2016). Hyopsodus arshantensis sp. nov. is similar in size to H. paulus (m2 length 3.6–4.7 mm; Gazin 1968), H. despiciens (mean of m2 length 4.54 mm; Gazin 1968), and H. tonksi (m2 length 4.37–4.80 mm; Eaton, 1982), smaller than H. marshi (m2 length 5.0–5.1 mm; Gazin 1968), larger than H. minusculus (m2 length 2.8–3.8 mm; Storer 1984), and H. lepidus (m2 length 3.4–4.5 mm, mean 4.24 mm; Storer 1984) (Fig. 3). The m1–m2 of H. arshantensis sp. nov. are generally similar to those of Bridgerian Hyopsodus in lacking a paraconid and entostylid, and in having moderately lophodont molar crowns, a distinct entoconid, and a midline position of the hypoconulid. However, the Erlian Basin material differs from Bridgerian Hyopsodus in having a longer trigonid, and in lacking variably developed metastylid and cingulids on the buccal and lingual sides. Hyopsodus arshantensis sp. nov. further differs from H. paulus by a greater angle between the cristid obliqua and posthypocristid, and from H. despiciens and H. lepidus by a less posteriorly directed posthypocristid and a more distinct postentocristid joining the hypoconulid. The species Hyopsodus tonksi has many unique dental morphological features and its assignment within Hyopsodus is even controversial (Eaton 1982; Flynn 1991). Thus, in terms of size and morphology, H. arshantensis sp. nov. is most comparable to the late Bridgerian species of Hyopsodus (except for H. tonksi) with relatively larger sizes. However, H. marshi is known mainly from upper dentitions, which are characterized by a well-developed protocone on P2–P4, a hypocone as prominent as the protocone on M1–M2, and conical paracone and metacone on the upper molars (Osborn 1902). The morphology of the lower teeth in H. marshi was rarely mentioned and is likely indistinguishable from other Bridgerian species of Hyopsodus, which in turn differ from H. arshantensis sp. nov. as discussed above. Hyopsodus marshi probably further differs from H. arshantensis sp. nov. by a generally more robust appearance (Tsukui 2016). The discovery of the upper dentition of H. arshantensis sp. nov. in the future could provide more differential characters in comparison with Bridgerian Hyopsodus.
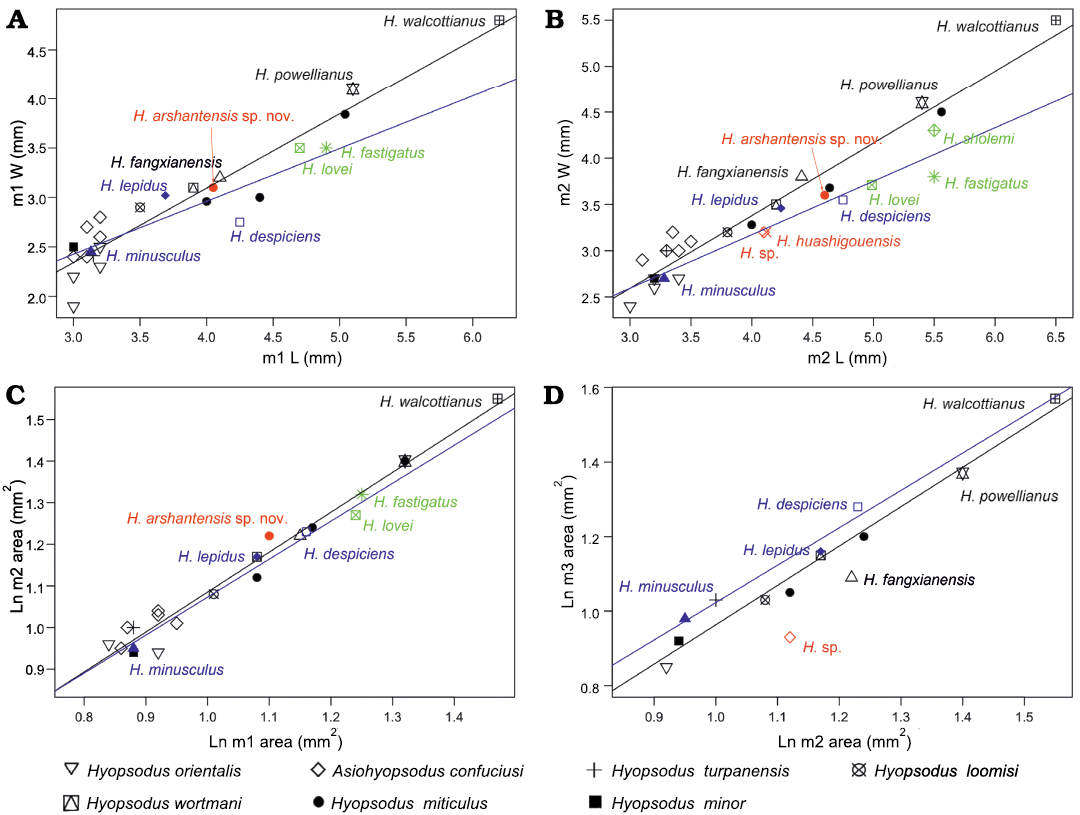
Fig. 3. Scatter plots of lower molar m1–m2 width (W) versus length (L), and size proportions of Hyopsodus and Asiohyopsodus. m1 (A) and m2 (B) width versus length, and regression lines for width as a function of length in species of Hyopsodus and Asiohyopsodus. Size proportions of m2 versus m1 (C) and m3 versus m2 (D), and regression lines for tooth size as a function of its preceding tooth size in species of Hyopsodus and Asiohyopsodus. Black symbols and lines, Wasatchian, blue symbols and lines, Bridgerian, red symbols represents the middle Eocene Hyopsodus from China, and the green symbols represent the Uintan Hyopsodus from North America. Abbreviation: H., Hyopsodus. (Raw data: SOM, the Supplementary Online Material available at http://app.pan.pl/SOM/app66-Bai_etal_SOM.pdf).
Comparison with Asian Hyopsodus and Asiohyopsodus.—Four species of Hyopsodus have been named from Asia, including H. orientalis (Dashzeveg 1977; Kondrashov and Agadjanian 1999; Hooker and Dashzeveg 2003), H. turpanensis (Zhai 1978; Tong 1989), and H. fangxianensis (Huang 1995) from the early Eocene, and H. huashigouensis (Tong 1989) from the middle Eocene (Fig. 2). Tong and Wang (2006) named a new genus and species, Asiohyopsodus confuciusi, from the early Eocene of the Wutu fauna (Shandong Province, China), and suggested that H. orientalis and the North American H. loomisi (McKenna 1960) are likely members of Asiohyopsodus. However, Rose et al. (2012) still considered the species H. loomisi more reasonably placed in Hyopsodus than in Asiohyopsodus and doubted the validity of Asiohyopsodus, because whether the relatively short and wide p3–p4 and reduced m3 in Asiohyopsodus confuciusi merit generic distinction from Hyopsodus needs further comparison with larger samples. Moreover, Kondrashov and Lucas (2004) synonymized H. turpanensis and H. fangxianensis with H. orientalis and H. huashigouensis, respectively. The taxonomy of five species of Hyopsodus and Asiohyopsodus previously reported from Asia (Tong and Wang 2006) is followed here (Fig. 2).
Hyopsodus arshantensis sp. nov. is similar in size to H. fangxianensis, and larger than other previously described Asian species of Hyopsodus or Asiohyopsodus (Fig. 3, Table 1). The m1–m2 of the new material differ from those of Asiohyopsodus confuciusi and H. orientalis in having more lophodont crown morphology, more obliquely aligned protoconid and metaconid, and a more distinct posthypocristid and postentocristid, and in lacking a paraconid on m1. The m1–m2 of Asiohyopsodus confuciusi is further characterized by a midline direction of the cristid obliqua and a smaller entoconid. The m1–m2 of H. orientalis further differ from those of H. arshantensis sp. nov. in having a hypoconulid located close to the entoconid. However, the three species are similar in having a relatively longer trigonid, an obtuse angle between the cristid obliqua and posthypocristid (H. orientalis with an angle slightly less than 90°), and in lacking cingulids on the buccal and lingual sides on m1–m2.
Hyopsodus turpanensis is known from a left mandible with a broken p4 talonid and m1–m3 (Zhai 1978; Tong 1989; Fig. 4A). The m1 of the holotype is heavily worn, and the m2 is moderately worn. The m1–m2 of H. arshantensis sp. nov. differ mainly from those of H. turpanensis in having a longer trigonid and a midline position of the hypoconulid. They are similar in the shared absence of the paraconid and cingulids on the buccal and lingual sides of m1–m2, and in having the obliquely aligned trigonid on m1–m2. Hyopsodus turpanensis also has a single-rooted p2 (Fig. 4A5, A6), an elongated m3, and the hypoconulid and entoconid almost fused into a ridge with a posthypocristid joining the hypoconulid in a relatively low position. The fused hypoconulid and entoconid, which almost form a ridge, are also present on m3 of H. wortmani (IVPP FV 0416, cast of AMNH FM 4716).
Hyopsodus fangxianensis is known from a right mandible with m1–m3 (Huang 1995; Fig. 4B). The m1 is heavily worn, and the m2 is moderately worn. The m1–m2 of H. arshantensis sp. nov. is similar to that of H. fangxianensis in having a relatively long trigonid and a midline position of the hypoconulid. However, H. fangxianensis can be differentiated by its more lingually directed paralophid on m1–m2, distinct entostylid on the anterior slope of the entoconid, relatively wider m1 and m2, and more robust horizontal ramus of the mandible (Fig. 4B4). The entostylid is also present in some Wasatchian Hyopsodus (Gazin 1968), such as the holotypes of H. “simplex” (as observed on IVPP FV 0418, cast of ACM 2290) and H. “jacksoni” (as observed on IVPP FV 0411, cast of ACM 3246).
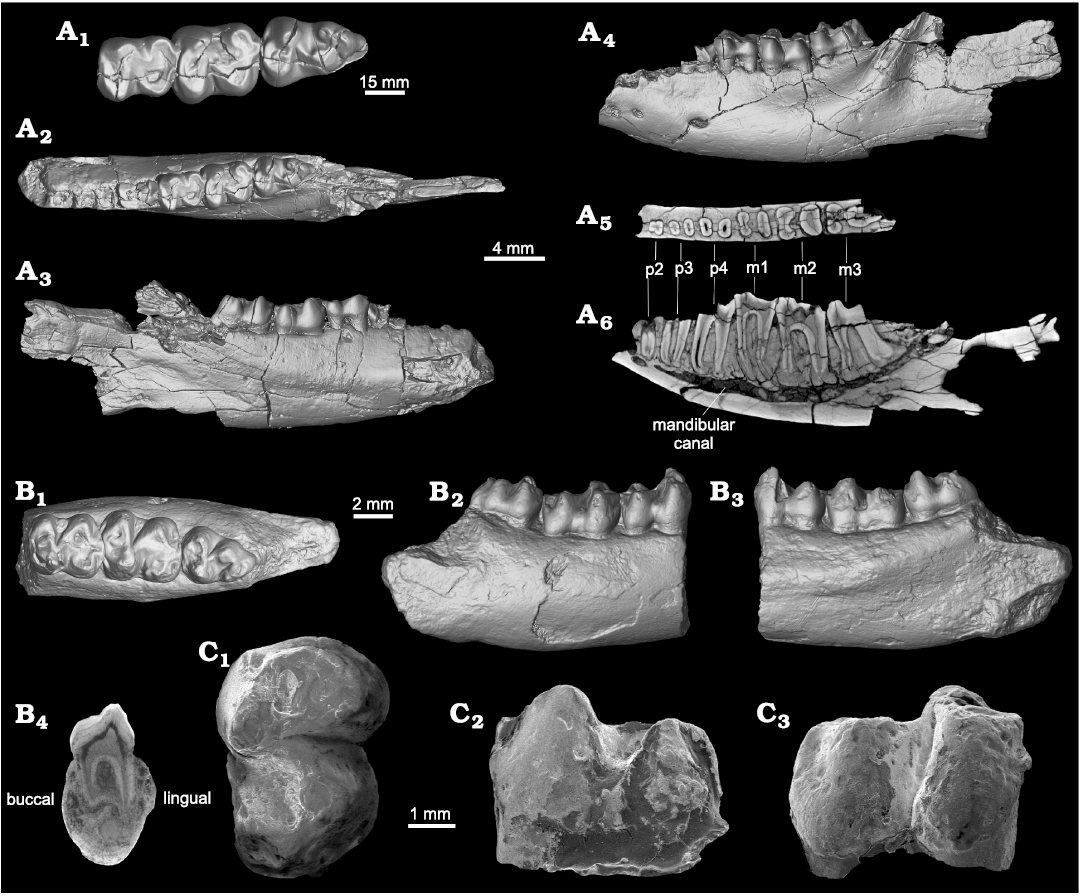
Fig. 4. Mandible fragments and teeth for several species of “condylarth” mammal Hyopsodus previously reported from China utilizing µ-CT scanning (A, B) and SEM images (C). A. Hyopsodus turpanensis Zhai, 1978 (IVPP V 4355) from the early Eocene Shisanjianfang Formation, Turpan Basin, Xinjiang, China. The left mandible with p2–p3 root, p4 talonid and m1–m3 in occlusal (A1, A2), lingual (A3), and buccal (A4) views, horizontal (A5) and longitudinal (A6) sections. B. Hyopsodus fangxianensis Huang, 1985 (IVPP V 12005) from the early Eocene Youping Formation, Fangxian, Hubei, China. The right mandible with m1–m3 in occlusal (B1), buccal (B2), and lingual (B3) views; cross section at the level of m2 trigonid (B4). C. Hyopsodus huashigouensis Tong, 1989 (IVPP V 7921) from the middle Eocene Üqbulak Formation, Junggar Basin, Xinjiang, China. The right m2 in occlusal (C1), lingual (C2), and buccal (C3) views.
Hyopsodus huashigouensis is known from a fragmentary right mandible with m2 (Tong 1989; Fig. 4C). The tooth is slightly worn with an incomplete entoconid. The m2 of H. arshantensis sp. nov. is similar to that of H. huashigouensis in having a relatively long trigonid, an obtuse angle between the cristid obliqua and posthypocristid, a midline position of the hypoconulid with a small size, and in lacking the cingulids at the buccal and lingual bases. However, the m2 of H. huashigouensis is different from that of H. arshantensis sp. nov. in its smaller size, a slightly more lophodont crown, more distinct postentocristid, and absence of the posterior cingulid.
Comparison with European Hyopsodus.—Two species of Hyopsodus have been reported from the early Eocene of Europe: Hyopsodus wardi and H. itinerans (Godinot 1978; Hooker 1979; Hooker and Dashzeveg 2003). Hooker and Dashzeveg (2003) considered H. wardi and H. itinerans as closely related to the North American H. loomisi and Asian H. orientalis, respectively. However, Tong and Wang (2006) suggested that the European species probably represent a unique group and could be treated as a new genus. The m1–m2 of H. arshantensis sp. nov. are different from those of H. wardi and H. itinerans in having a larger size, a more lophodont crown morphology, a longer trigonid without the paraconid, a more obliquely oriented trigonid, an obtuse angle between the cristid obliqua and posthypocristid, and a midline position of the hypoconulid. Hyopsodus wardi is further characterized by a prominent entostylid on the anterior slope of m1–m2 entoconid, and a distinct cingulid at the base of the ectoflexid on m1–m2 (Hooker 1979).
Biostratigraphic implications.—Compared with North American Bridgerian Hyopsodus species, the relatively large size of H. arshantensis sp. nov. is close to that of species of Hyopsodus from the late Bridgerian (Archibald 1998). Although H. despiciens, H. paulus, and H. lepidus are not restricted in the late Bridgerian as is H. marshi, the specimens of Bridgerian Hyopsodus with relatively larger sizes likely evolved later in the Bridgerian (Fig. 2). Considering the similarities in size and morphology between H. arshantensis sp. nov. and the late Bridgerian species of Hyopsodus, the age of the middle part of the Arshanto Formation, where the holotype of H. arshantensis sp. nov. was collected, likely can be correlated to the late Bridgerian. In addition, the perissodactyl Ephyrachyus woodi from the upper part of the Arshanto Formation (AS-4) is similar to Ephyrachyus implicatus from the Washakie Formation in the Washakie Basin of Wyoming (Bai et al. 2020). Wood (1934) suggested that E. implicatus was probably unearthed from the late Bridgerian deposits, however, the species is actually distributed through Ui1b to Ui3 (Roehler 1973; Gunnell et al. 2009). Thus, the age of the upper part of the Arshanto Formation is most likely correlated to the early Uintan. Furthermore, the age of the Irdin Manhan Formation is intermediate between the late Bridgerian and late Uintan, as indicated by bearing helaletid Desmatotherium mongoliensis and Paracolodon fissus, the former being more derived than late Bridgerian D. intermedius and the latter being more primitive than late Uintan Colodon? kayi, C.? woodi (Bai et al. 2017). The early primate Tarkops from the Indin Manhan Formation was considered being relatively more primitive than Tarka from the Ui1b of the Tepee Trail Formations, East Fork Basin, Wyoming (Gunnell et al. 2009; Ni et al. 2010). Thus, these correlations would mean that the upper part of the Arshanto Formation was deposited in the time interval of the early Uintan, and may bracket the period equivalent to the Bridgerian/Uintan boundary (Fig. 2).
Hyopsodus huashigouensis was recorded from the bone bed A of the Üqbulak Formation, Üqbulak area of the Junggar Basin, Xinjiang, China and the Üqbulak Formation is considered equivalent to the Bridgerian (Tong 1989). Bone beds B and C of the Üqbulak Formation are correlated to the Arshantan and Irdinmanhan, respectively. Tong et al. (1990) later considered the entire Üqbulak Formation (bone beds A to C), as middle Eocene in age (Li 2018). Bone bed B of the Üqbulak Formation has produced specimens of Mesonyx uqbulakensis, also known from the base of the Arshanto Formation (Tong 1989; Jin 2012). That distribution suggests that the stratigraphically lower bone bed A is no younger than the early Arshantan, and that H. huashigouensis is somewhat older than H. arshantensis sp. nov..
The early Eocene Hyopsodus turpanensis from the Shisanjianfang Formation of the Turpan Basin, Xinjiang, China (Zhai 1978) and H. fangxianensis from the Youping Formation of Fangxian, Hubei Province, China (Huang 1995) are more derived than H. orientalis and Asiohyopsodus confuciusi. Those hypothesized relationships support the idea that the Shisanjianfang and Youping formations are equivalent to the Heptodon Interval Zone of the Bumbanian and can be correlated to the Lysitean and Lostcabinian subzones of the Wasatchian (Ting 1998). The Wutu fauna, which produced Asiohyopsodus confuciusi, is correlated to the Homogalax Interval Zone of the Bumbanian and correlated to the Graybullian subzone of the Wasatchian (Ting 1998). Hyopsodus orientalis from the Bumban fauna of Mongolia is correlated to the Orientolophus Interval Zone of the Bumbanian and to the Sandcouleean of the Wasatchian (Ting 1998).
Conclusions
The middle Eocene “condylarth” mammal Hyopsodus arshantensis sp. nov. from the middle part of the Arshanto Formation represents the first record of hyopsodontids from the Erlian Basin, Inner Mongolia, China. Its morphology is somewhat intermediate between Wasatchian and Uintan species of Hyopsodus, and is comparable to species of Hyopsodus from the Bridgerian. The relatively large size of H. arshantensis sp. nov. is close to the species of Hyopsodus from the late Bridgerian, suggesting that the middle part of the Arshanto Formation most likely can be correlated to the late Bridgerian. Thus, the Bridgerian/Uintan boundary could be correlated to a level lying in the upper part of the Arshanto Formation, rather than to the contact of the Arshanto and Irdinmanha formations. It is necessary to mention that the fauna from the upper part of the Arshanto Formation (AS-4 and AS-5) is somewhat different from those of the lower and middle parts of the Arshanto Formation (AS-1–3), and shows the first appearance of some new rodents and perissodactyls (Li and Meng 2015; Li 2016; Bai et al. 2018). The study of other fossil mammal groups (e.g., artiodactyls and deperetellids) from the Arshanto and Irdin Manhan formations are underway, and more evidence could point to the necessity of subdividing the Arshantan and/or redefining the Arshantan/Irdinmanhan boundary in future studies.
Acknowledgements
We thank Qian Li, Xun Jin, Fang-Yuan Mao, Hai-Bing Wang, Wei Zhou, Shi-Jie Li, Qi Li, Yong-Xing Wang, Yong-Fu Wang, Xiao-Yang Wang, Ran-Cheng Xu (all IVPP), K. Christopher Beard (University of Kansas, Lawrence, USA), and Daniel L. Gebo (Northern Illinois University, DeKalb, USA) for assistance in the field; Xun Jin and Wei Gao (both IVPP) for photography; Ye-Mao Hou (IVPP) for µ-CT scanning; Jack Z. Tseng (University of California, Berkeley, USA) and Ryan McKeller (Royal Saskatchewan Museum, Regina, Saskatchewan, Canada) for valuable references; and Thomas Stidham (IVPP) for comments and improvement of English writing. The comments and suggestions from K. Christopher Beard, Thierry Smith (Royal Belgian Institute of Natural Sciences, Brussels, Belgium), and the editor Oliver Lambert (Royal Belgian Institute of Natural Sciences, Brussels, Belgium) greatly improved the final manuscript. Funding was provided by grants from the Strategic Priority Research Program of Chinese Academy of Sciences (XDB26000000), the National Natural Science Foundation of China (41672014), the Special Research Program of Basic Science and Technology of the Ministry of Science and Technology (2015FY310100), Youth Innovation Promotion Association CAS (2017101), Special Fund for Fossil Excavation and Preparation of Chinese Academy of Sciences, and Frick Funds from the Division of Paleontology, American Museum of Natural History.
References
Archibald, J.D. 1998. Archaic ungulates (“Condylarthra”). In: C.M. Janis, K.M. Scott, and L.L. Jacobs (eds.), Evolution of Tertiary Mammals of North America. Volume 1: Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals, 292–331. Cambridge University Press, Cambridge. Crossref
Bai, B., Meng, J., Zhang, C., Gong, Y.-X., and Wang, Y.-Q. 2020. The origin of Rhinocerotoidea and phylogeny of Ceratomorpha (Mammalia, Perissodactyla). Communications Biology 3: 1–16. Crossref
Bai, B., Wang, Y.Q., Li, Q., Wang, H.B., Mao, F.Y., Gong, Y.X., and Meng, J. 2018. Biostratigraphy and diversity of Paleogene perissodactyls from the Erlian Basin of Inner Mongolia, China. American Museum Novitates 3914: 1–60. Crossref
Bai, B., Wang, Y.Q., Mao, F.Y., and Meng, J. 2017. New material of Eocene Helaletidae (Perissodactyla, Tapiroidea) from the Irdin Manha Formation of the Erlian Basin, Inner Mongolia, China and comments on related localities of the Huheboerhe Area. American Museum Novitates 3878: 1–44. Crossref
Cifelli, R. 1983. The origin and affinities of the South American Condylarthra and early Tertiary Litopterna (Mammalia). American Museum Novitates 2772: 1–49.
Dashzeveg, D. 1977. On the first occurrence of Hyopsodus Leidy, 1870 (Mammalia, Condylarthra) in Mongolian People’s Republic. Transactions of the Joint Soviet-Mongolian Paleontological Expedition 4: 7–13.
Eaton, J. 1982. Paleontology and correlation of Eocene volcanic rocks in the Carter Mountain area, Park County, southeastern Absaroka Range, Wyoming. Contributions to Geology, University of Wyoming 21: 153–194.
Flynn, J.J. 1991. Hyopsodus (Mammalia) from the Tepee Trail Formation (Eocene), northwestern Wyoming. American Museum Novitates 3007: 1–19.
Gazin, C.L. 1968. A study of the Eocene condylarthran mammal Hyopsodus. Smithsonian Miscellaneous Collections 153 (4): 1–90.
Gazin, C.L. 1976. Mammalian faunal zones of the Bridger Middle Eocene. Smithsonian Contributions to Paleobiology 26: 1–25. Crossref
Gingerich, P.D. 1974. Stratigraphic record of early Eocene Hyopsodus and the geometry of mammalian phylogeny. Nature 248: 107–109. Crossref
Gingerich, P.D. 1976. Paleontology and phylogeny; patterns of evolution at the species level in early Tertiary mammals. American Journal of Science 276: 1–28. Crossref
Godinot, M. 1978. Diagnoses de trois nouvelles espèces de mammifères du Sparnacien de Provence. Compte Rendu Sommaire des Séances de la Société Géologique de France 6: 286–288.
Godinot, M., Smith, T., and Smith, R. 1996. Mode de vie et affinités de Paschatherium (Condylarthra, Hyopsodontidae) d’après ses os du tarse. In: M. Godinot and P.D. Gingerich (eds.), Paléobiologie et Évolution des Mammifères Paléogènes: Volume Jubilaire en Hommage à Donald E. Russell. Palaeovertebrata 25: 225–242.
Gunnell, G., Murphy, P., Stucky, R., Townsend, K., Robinson, P., Zonneveld, J., and Bartels, W. 2009. Biostratigraphy and biochronology of the latest Wasatchian, Bridgerian, and Uintan North American Land Mammal “Ages”. In: L.B. Albright and M.O. Woodburne (eds.), Papers on Geology, Vertebrate Paleontology, and Biostratigraphy in Honor of Michael O. Woodburne. Museum of Northern Arizona Bulletin 65: 279–330.
Halliday, T.J., Upchurch, P., and Goswami, A. 2017. Resolving the relationships of Paleocene placental mammals. Biological Reviews 92: 521–550. Crossref
Hooker, J.J. 1979. Two new condylarths (Mammalia) from the early Eocene of southern England. Bulletin of the British Museum (Natural History), Geology Series 32: 43–56.
Hooker, J.J. and Dashzeveg, D. 2003. Evidence for direct mammalian faunal interchange between Europe and Asia near the Paleocene–Eocene boundary. In: S.L. Wing, P.D. Gingerich, B. Schmitz, and E. Thomas (eds.), Causes and Consequences of Globally Warm Climates in the Early Paleogene, Geological Society of America Special Paper 369, 479–500. Geological Society of America, Boulder.
Huang, X.S. 1995. A new Hyopsodus from the early Eocene of Fangxian, Hubei. Vertebrata PalAsiatica 33: 39–46.
Jin, X. 2012. New mesonychid (Mammalia) material from the lower Paleogene of the Erlian Basin, Nei Mongol, China. Vertebrata Palasiatica 50: 245–257.
Kondrashov, P.E. and Agadjanian, A.K. 1999. New material on the genus Hyopsodus (Mammalia, Condylarthra) from the Eocene of Mongolia: Morphological variability and taxonomic position. Paleontological Journal 33 (6): 64–72.
Kondrashov, P.E. and Lucas, S.G. 2004. Revised distribution of condylarths (Mammalia, Eutheria) in Asia. In: S.G. Lucas, K.E. Zeigler, and P.E. Kondrashov (eds.), Paleogene Mammals. New Mexico Museum of Natural History and Science Bulletin 26: 209–214.
Krishtalka, L. 1979. Paleontology and geology of the Badwater Creek area, central Wyoming. Revision of late Eocene Hyopsodus. Annals of Carnegie Museum 48: 377–389.
Leidy, J. 1870. Remarks on a collection offossils from the western Territories. Proceedings of the Academy of Natural Sciences of Philadelphia 22: 109–110.
Li, Q. 2016. Eocene fossil rodent assemblages from the Erlian Basin (Inner Mongolia, China): Biochronological implications. Palaeoworld 25: 95–103. Crossref
Li, Q. and Meng, J. 2015. New ctenodactyloid rodents from the Erlian Basin, Nei Mongol, China, and the phylogenetic relationships of Eocene Asian ctenodactyloids. American Museum Novitates 3828: 1–60. Crossref
Li, S. 2018. A new species of Brontotheriidae from the Middle Eocene of Junggar Basin, Xinjiang, China. Vertebrata PalAsiatica 56: 25–44.
Loomis, F.B. 1905. Hyopsodidae of the Wasatch and Wind River basins. American Journal of Science (4) 19: 416–424. Crossref
Matthew, W.D. 1909. The Carnivora and Insectivora of the Bridger Basin, middle Eocene. Memoirs of the American Museum of Natural History 9 (6): 291–567.
Matthew, W.D. 1915. Part II. Order Condylarthra. Families Hyopsodontidae. In: W.D. Matthew and W. Granger (eds.), A Revision of the Lower Eocene Wasatch and Wind River Faunas. Bulletin of the American Museum of Natural History 34: 311–328.
McKenna, M.C. 1960. Fossil Mammalia from the early Wasatchian Four Mile fauna, Eocene of northwest Colorado. University of California Publications in Geological Sciences 37: 1–130.
Muizon, C.d. and Cifelli, R.L. 2000. The “condylarths” (archaic Ungulata, Mammalia) from the early Palaeocene of Tiupampa (Bolivia): implications on the origin of the South American ungulates. Geodiversitas 22: 47–150.
Murphey, P.C. and Kelly, T.S. 2017. Mammals from the earliest Uintan (middle Eocene) Turtle Bluff Member, Bridger Formation, southwestern Wyoming, USA. Part 2: Apatotheria, Lipotyphla, Carnivoramorpha, Condylartha, Dinocerata, Perissodactyla and Artiodactyla. Palaeontologia Electronica 20.1.29A: 1–51. Crossref
Ni, X.J., Meng, J., Beard, K.C., Gebo, D.L., Wang, Y.Q., and Li, C.K. 2010. A new tarkadectine primate from the Eocene of Inner Mongolia, China: phylogenetic and biogeographic implications. Proceedings of the Royal Society B-Biological Sciences 277: 247–256. Crossref
Orliac, M.J., Argot, C., and Gilissen, E. 2012. Digital Cranial Endocast of Hyopsodus (Mammalia, “Condylarthra’’): A Case of Paleogene Terrestrial Echolocation? PLoS One 7 (2): 1–10. Crossref
Osborn, H.F. 1902. American Eocene primates, and supposed rodent family Mixodectidae. Bulletin of the American Museum of Natural History 16 (17): 169–214.
Ravel, A. and Orliac, M.J. 2015. The inner ear morphology of the “condylarthran” Hyopsodus lepidus. Historical Biology 27: 957–969. Crossref
Redline, A.D. 1997. Revision of the Wind River faunas, early Eocene of central Wyoming. Systematics and phylogenetic pattern of early Eocene Hyopsodus (Mammalia: Condylarthra). Annals of Carnegie Museum 66: 1–81.
Roehler, H.W. 1973. Stratigraphy of the Washakie Formation in the Washakie Basin, Wyoming. U.S. Geological Survey Bulletin 1369: 1–40.
Rose, K.D. 2006. The Beginning of the Age of Mammals. 428 pp. The John Hopkins University Press, Baltimore.
Rose, K.D., Chew, A.E., Dunn, R.H., Kraus, M.J., Fricke, H.C., and Zack, S.P. 2012. Earliest Eocene mammalian fauna from the Paleocene–Eocene Thermal Maximum at Sand Creek Divide, Southern Bighorn Basin, Wyoming. Museum of Paleontology Papers on Paleontology 36: i–ix, 1–122.
Russell, L.S. and Wickenden, R.T.D. 1933. An upper Eocene vertebrate fauna from Saskatchewan. Transactions of the Royal Society of Canada 27 (4): 53–66.
Secord, R., Wing, S.L., and Chew, A. 2008. Stable isotopes in early Eocene mammals as indicators of forest canopy structure and resource partitioning. Paleobiology 34: 282–300. Crossref
Simpson, G.G. 1945. The principles of classification and a classification of mammals. Bulletin of American Museum of Natural History 85: 1–350.
Speijer, R.P., Pälike, H., Hollis, C.J., Hooker, J.J., and Ogg, J.G. 2020. The Paleogene period. In: F.M. Gradstein, J.G. Ogg, M.D. Schmitz, and G.M. Ogg (eds.), Geologic Time Scale 2020, 1087–1140. Elsevier, Oxford. Crossref
Storer, J.E. 1984. Mammals of the Swift Current Creek Local Fauna. Natural History Contributions, Saskatchewan Culture and Recreation Museum of Natural History 7: 1–158.
Ting, S.Y. 1998. Paleocene and early Eocene Land Mammal Ages of Asia. In: C. Beard and M.R. Dawson (eds.), Dawn of the Age of Mammals in Asia. Bulletin of the Carnegie Museum of Natural History 34: 124–147.
Tong, Y.S. 1989. Some Eocene mammals from the Üqbulak area of the Junggar basin, Xinjiang. Vertebrata PalAsiatica 27 (3): 182–196.
Tong, Y.S., Qi, T., Ye, J., Meng, J., and Yan, D.F. 1990. Tertiary stratigraphy of the north of Junggar Basin, Xinjiang. Vertebrata Palasiatica 28 (1): 59–70.
Tong, Y.S. and Wang, J.W. 2006. Fossil mammals from the Early Eocene Wutu Formation of Shandong Province. Palaeontologia Sinica, New Series C 28: 1–195.
Tsukui, K. 2016. Chronology and Faunal Evolution of the Middle Eocene Bridgerian North American Land Mammal “Age”: Achieving High Precision Geochronology. 247 pp. Unpublished PhD Thesis, Columbia University, New York.
Wang, Y.F., Wei, C.F., Que, J.M., Zhang, W.D., Sun, C.L., Shu, Y.F., Hou, Y.M., Zhang, J.C., Shi, R.J., and Wei, L. 2019a. Development and applications of paleontological computed tomography. Vertebrata PalAsiatica 57 (1): 84–92.
Wang, Y.Q., Li, Q., Bai, B., Jin, X., Mao, F.Y., and Meng, J. 2019b. Paleogene integrative stratigraphy and timescale of China. Science China Earth Sciences 62: 287–309. Crossref
Wang, Y.Q., Meng, J., Beard, C.K., Li, Q., Ni, X.J., Gebo, D.L., Bai, B., Jin, X., and Li, P. 2010. Early Paleogene stratigraphic sequences, mammalian evolution and its response to environmental changes in Erlian Basin, Inner Mongolia, China. Science China Earth Sciences 53: 1918–1926. Crossref
West, R.M. 1979. Paleontology and geology of the Bridger formation, southern Green River Basin, southwestern Wyoming. Part 3. Notes on Hyopsodus. Milwaukee Public Museum, Contributions in Biology and Geology 25: 1–52.
Wood, H.E. 1934. Revision of the Hyrachyidae. Bulletin of the American Museum of Natural History 67 (5): 181–295.
Zack, S.P., Penkrot, T.A., Bloch, J.I., and Rose, K.D. 2005a. Affinities of “hyopsodontids” to elephant shrews and a Holarctic origin of Afrotheria. Nature 434: 497–501. Crossref
Zack, S.P., Penkrot, T.A., Krause, D.W., and Maas, M.C. 2005b. A new apheliscine “condylarth” mammal from the late Paleocene of Montana and Alberta and the phylogeny of “hyopsodontids”. Acta Palaeontologica Polonica 50: 809–830.
Zhai, R.J. 1978. More evidence favouring on early Eocene connection between Asia and Neoarctic. In: Reports of Paleontological Expedition to Sinkiang (III)—Permian and Triassic vertebrate fossils of Dzungaria Basin and Tertiary stratigraphy and mammalian fossils of Turfan Basin. Memoirs of the Institute of Vertebrate Paleontology and Paleoanthropology, Academia Sinica 13: 107–115.
Acta Palaeontol. Pol. 66 (4): 767–777, 2021
https://doi.org/10.4202/app.00908.2021