

A new pliosaurid from the Oxford Clay Formation of Oxfordshire, UK
HILARY F. KETCHUM and ROGER B.J. BENSON
Ketchum, H.F. and Benson, R.B.J. 2022. A new pliosaurid from the Oxford Clay Formation of Oxfordshire, UK. Acta Palaeontologica Polonica 67 (2): 297–315.
We describe the anatomy of a new longirostrine pliosaurid, Eardasaurus powelli gen. et sp. nov., based on a substantially complete skeleton from the Middle Jurassic Oxford Clay Formation of Oxfordshire, UK. The specimen brings the number of pliosaurid genera known from the Oxford Clay Formation up to a minimum of six, contributing to our understanding of the most taxonomically rich pliosaurid assemblage. The teeth of Eardasaurus powelli gen. et sp. nov. have “carina-like” apicobasal enamel ridges, previously only identified in Liopleurodon ferox among Middle Jurassic pliosaurids, and which may be more widespread among early pliosaurids than has been recognised. Our phylogenetic analysis shows that Eardasaurus powelli gen. et sp. nov. is nested within Middle Jurassic thalassophonean pliosaurids, slightly more derived than Peloneustes philarchus, as sister to a group comprising “Pliosaurus” andrewsi, Simolestes vorax, Liopleurodon ferox, Pliosaurus spp. and brachauchenines.
Key words: Plesiosauria, Pliosauridae, Oxford Clay Formation, Jurassic, Callovian, England.
Hilary F. Ketchum [hilary.ketchum@oum.ox.ac.uk], Oxford University Museum of Natural History, Parks Road, Oxford, OX1 3PW, UK.
Roger B. J. Benson [roger.benson@earth.ox.ac.uk], Department of Earth Sciences, University of Oxford, 3 South Parks Road, Oxford, OX1 3AN, UK.
Received 13 March 2021, accepted 13 December 2021, available online 23 May 2022.
Copyright © 2022 H.F. Ketchum and R.B.J. Benson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Pliosaurid plesiosaurians first appeared in the fossil record in the earliest Jurassic as small-bodied animals around 2 m in length and evolved long-snouted piscivorous forms by the late Early Jurassic (Benson et al. 2011). They underwent evolutionary increases in body size, reductions in neck length, and increases in relative head size to attain characteristic “pliosauromorph” body proportions by the Middle Jurassic (e.g., O’Keefe and Carrano 2005; Soul and Benson 2017), with the Late Jurassic and Cretaceous pliosaurids attaining body lengths of 10–12 m (Romer and Lewis 1959; Tarlo 1960; Benson et al. 2013), remaining as apex predators in marine ecosystems until their extinction in the early Late Cretaceous (e.g., Schumacher 2011; Schumacher et al. 2013).
The Middle Jurassic Oxford Clay Formation (Callovian–early Oxfordian) of England has produced a large number of plesiosaurian fossils for over 100 years (e.g., Andrews 1910, 1913; Tarlo 1960; Brown 1981; Ketchum and Benson 2011a, b). It remains one of the most important and well-studied assemblages of pliosaurids anywhere in the world, both in terms of taxonomic diversity and number of specimens. Five species of pliosaurids from the Oxford Clay Formation are currently considered valid: Simolestes vorax, Liopleurodon ferox, Peloneustes philarchus, “Pliosaurus” andrewsi, and Marmornectes candrewi (see Andrews 1913; Tarlo 1960; Noè 2001; Ketchum and Benson 2011a, b); plus Pachycostasaurus dawni—a “pliosauromorph” taxon of uncertain affinities described on the basis of a highly juvenile individual (Cruickshank et al. 1996).
To date, the Oxford Clay Formation plesiosaurian assemblage shows high taxonomic diversity compared to younger assemblages (Benson and Druckenmiller 2014; Foffa et al. 2018) such as the Sundance Formation (Oxfordian of the USA; O’Keefe and Wahl 2003); the Jagua Formation (Oxfordian of Cuba; e.g., Gasparini 2009); the Kimmeridge Clay Formation (Kimmeridgian–Tithonian of the UK; e.g., Benson et al. 2013); La Caja Formation (Kimmeridgian–lower Berriasian of Mexico; Buchy et al. 2003); the Vaca Muerta Formation (Tithonian–lower Volanginian of Argentina; e.g., Gasparini and O’Gorman 2014; O’Gorman et al. 2018), the Agardhfjellet Formation (Tithonian, Svalbard Archipelago, Norway; e.g., Knutsen et al. 2012) and several Late Jurassic formations of Russia (e.g., Arkhangelsky et al. 2019; Zverkov et al. 2017: 1). However, this may be due to high sampling intensity, and the fossil record of pliosaurid teeth suggests that the group attained their highest disparity of dental morphology and body sizes during the Late Jurassic and earliest Cretaceous (Zverkov et al. 2018).
Many pliosaurid species of the Oxford Clay Formation are known from multiple specimens (Andrews 1913; Tarlo 1960; e.g., 21 specimens of Peloneustes philarchus; Ketchum and Benson 2011b). Most specimens, including the holotypes of Simolestes vorax (NHMUK PV R 3319) and “Pliosaurus” andrewsi (NHMUK PV R 3891) were collected from clay pits in and around Peterborough, Cambridgeshire, by Charles and Alfred Leeds (Andrews 1913). Both of the brothers, but particularly Alfred, became widely known as pioneers of modern methods of collecting and preserving fossil vertebrates in the latter half of the 19th century (Anon 1917). Since then, both the number of working Oxford Clay pits and the number of finds has diminished substantially, due to increasing mechanisation combined with the reduction in demand for common bricks (Bloodworth et al. 2001).
In spite of the diminishing rates of excavation and discovery, ongoing exploration of the extant outcrops of the Oxford Clay Formation demonstrate its continuing potential to yield new species of pliosaurids. For example, a partial skeleton from Bedfordshire discovered in 1998 (BEDFM 1999.201) was reported as a new genus and species, Marmornectes candrewi (Ketchum and Benson 2011a). This is surprising given the high numbers of specimens discovered historically. Here, we report a substantially complete skeleton discovered by H. Philip Powell, a former assistant curator of Oxford University Museum of Natural History, near Yarnton, Oxfordshire in August 1994. The specimen was extracted by H. Philip Powell, Richard M. Wilkins, Juliet Hay, Simon Phillips, John B. Cooke, Nigel T.J. Hollingworth, and Terry R. Hardaker, and represents a new genus and species, Eardasaurus powelli gen. et sp. nov.
Institutional abbreviations.—BEDFM, The Higgins Bedford, Bedford, UK; NHMUK, Natural History Museum, London, UK; OUMNH, Oxford University Museum of Natural History, Oxford, UK.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in ZooBank: urn:lsid:zoobank.org:pub:DD3E3280-B008-465B-883F-726EED54E488.
Geological setting
The Oxford Clay Formation of the Ancholme Group extends in a band across England from Dorset in the south to Yorkshire in the north. The Peterborough Member (formerly the “Lower Oxford Clay”: Sigaloceras enodatum Ammonite Subzone (Sigaloceras calloviense Zone)–Kosmoceras phaeinum Ammonite Subzone (Peltoceras athleta Zone); late early–early late Callovian (e.g., Callomon 1968; Duff 1980; Hudson and Martill 1994; Gradstein et al. 2020) is dominated by brownish-grey, fissile, organic-rich mudstone (Hudson and Martill 1994). It reaches around 28 m thick in the Oxford area, but much of the outcrop is concealed under unconformable Pleistocene sand and gravel deposits (Hollingworth and Wignall 1992; Bloodworth et al. 2001). The Oxford Clay formed within the North West European epieric sea, surrounded by islands and large land-masses (Hudson and Martill 1994).
The new specimen was collected from a temporary exposure of the Peterborough Member at the former ARC Cassington gravel pit near Yarnton, Oxfordshire, following the construction of a deep silt pond (H. Philip Powell, personal communication 2019 to HFK). The bed from which it was collected contained a prominent layer of septarian concretions in addition to abundant Gryphaea and ammonites including Indosphinctes, Cadoceras, Reineckia, and Kosmoceras jason. The pliosaurid skeleton thus derives from the Kosmoceras jason Ammonite Subzone of the Kosmoceras jason Zone (lower middle Callovian; Gradstein et al. 2020) of the Peterborough Member (H. Philip Powell, personal communication 2019 to HFK). OUMNH PAL-J.2247 was partially embedded in six concretions, which enclosed the posterior two-thirds of the cranium, parts of the vertebral series, and the hindlimb.
Material and methods
The present study is based on a single, well-preserved, substantially complete and partially articulated skeleton housed in the Oxford University Museum of Natural History, OUMNH PAL-J.2247 (Fig. 1). It measures approximately 4.7 m long from the tip of the snout to the end of the (incomplete) tail, as preserved, and includes the cranium, lower jaw, four scleral ossicles, two hyoids, more than 40 teeth and tooth fragments (including both disarticulated and in situ teeth), atlas-axis complex, 57 postaxial vertebrae (comprising 18 cervicals, 3 pectorals, 24 dorsals, 4 sacrals, and 8 caudals, along with disarticulated neural arches, neural spines, ribs, and chevrons), and 11 gastralia. The preserved appendicular skeleton includes the distal end of the right humerus plus a partial left forelimb discovered in articulation, which includes the radius, ulna, proximal and distal carpals, and 17 phalanges. There is a partial left hindlimb comprising the femur, proximal and distal tarsals. In total 40 complete phalanges, plus six partial phalanges were found disarticulated and cannot confidently be assigned to any particular limb. Of the limb girdles, only the interclavicle and a clavicle are preserved.
We describe the anatomy of the skull in detail, and a brief description of the postcranial skeleton is presented in the SOM 1 (Supplementary Online Material available at http://app.pan.pl/SOM/app67-Ketchum_Benson_SOM.pdf). We also present 3D surface scans of elements from all major portions of the skeleton, which are available at Morphosource (www.morphosource.org/projects/000371976).
Prior to photography, the teeth were coated with a thin layer of ammonium chloride to highlight the pattern of apicobasal ridges.
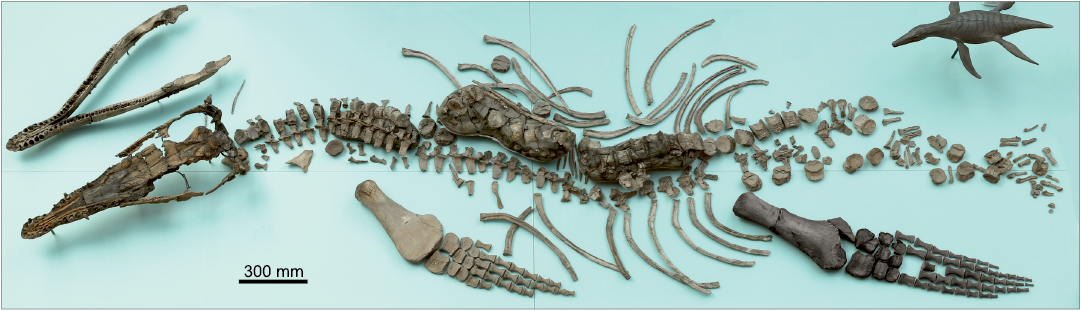
Fig. 1. Holotype of the pliosaurid plesiosaur Eardasaurus powelli gen. et sp. nov. (OUMNH PAL-J.2247) from the Middle Jurassic of Yarnton, Oxfordshire, UK. Photograph of the specimen on display in OUMNH. Skeleton laid out as discovered, with the exception of the left hindlimb (highlighted in black), which was originally disarticulated. An artist’s reconstruction of E. powelli is partially visible in the top right-hand side of the image.
Systematic palaeontology
Sauropterygia Owen, 1860
Plesiosauria de Blainville, 1835
Pliosauridae Seeley, 1874a
Thalassophonea Benson and Druckenmiller, 2014
Genus Eardasaurus nov.
Zoobank LSID: urn:lsid:zoobank.org:act:36CC5781-89C0-4475-8BDE- 79459C263356
Etymology: From Old English eard, home, native soil or dwelling, from which the village name Yarnton was derived; and from Greek saurus, lizard.
Type species: Eardasaurus powelli sp. nov.; see below.
Diagnosis.—As for type and only species.
Eardasaurus powelli sp. nov.
Figs. 1–13.
ZooBank LCID: urn:lsid:zoobank.org:act:E8E70687-2F84-4875-BDE8-820EB5D05A7E
Etymology: After H. Philip Powell, former Assistant Curator at the Oxford University Museum of Natural History.
Holotype: OUMNH PAL-J.2247; well-preserved, partially complete skeleton (Fig. 1).
Type locality: Gravel pit near Yarnton, Oxfordshire, UK. National Grid Reference SP 479 108; latitude (decimal) 51.7938, longitude (decimal) -1.3069.
Type horizon: Kosmoceras jason Zone, Peterborough Member, Oxford Clay Formation, Ancholme Group, middle Callovian, Middle Jurassic.
Diagnosis.—Longirostrine pliosaurid with the following autapomorphies: (i) maxilla with a very high number of alveoli (34 or 35); (ii) jugal with “fluted” ornamentation along the orbital margin; (iii) step-like contact between the jugal and squamosal; (iv) “lacrimal” (neomorphic ossification in the anteroventral orbit margin of pliosaurids) dorsoventrally slender with a posterior rod-like projection. Eardasarus powelli gen. et sp. nov. can also be differentiated from other pliosaurids by the following combination of characters: (i) pattern of enamel ridges on larger (i.e., mesial) teeth similar to those of Peloneustes philarchus (Seeley, 1869) (and different to other Oxford Clay Formation pliosaurids; Tarlo 1960), including relatively few enamel ridges on the convex (labial) tooth surface and most ridges terminate far short of tooth apex, except for a small number of “carina-like” apicobasal enamel ridges on each tooth; (ii) five-six premaxillary teeth; (iii) prominent mediolateral constriction of rostrum at premaxilla–maxilla suture, similar to most Middle Jurassic pliosaurids by only weakly developed in Marmornectes candrewi Ketchum and Benson, 2011a, and “Pliosaurus” andrewsi Tarlo, 1960; (iv) diastema between premaxillary and maxillary dentitions, similar to Liopleurodon ferox Sauvage, 1873, but differing from other Middle Jurassic pliosaurids Marmornectes candrewi, Peloneustes philarchus, and “Pliosaurus” andrewsi; (v) maxilla excluded from medial margin of external naris by an anterior extension of the frontal, differing from other Middle Jurassic pliosaurids; (vi) frontal extends anterior to the external naris, excluding the maxilla from the medial margin, differing from Simolestes vorax Andrews, 1909, and Liopleurodon ferox (among Middle Jurassic pliosaurids) in which the frontal does not extend as far anteriorly; (vii) proportionally elongate posterior interpterygoid vacuities compared to other Middle Jurassic pliosaurids, especially “Pliosaurus” andrewsi; (viii) mediolaterally expanded mandibular symphysis; (ix) nine pairs of dentary teeth adjacent to the mandibular symphysis, a relatively small count compared to other longirostrine Middle Jurassic pliosaurids (e.g., 13–15 symphysial alveoli in Peloneustes philarchus; 12 in Marmornectes candrewi) except for “Pliosaurus” andrewsi (11 symphysial alveoli); (x) splenial encloses the posterior margin of the anterior opening of Meckel’s canal (rather than being limited to the ventral margin as in most other pliosaurids); (xi) coronoid exposed on the lateral surface of the mandible; (xii) strongly convex medial expansion of the surangular in dorsal view; characters (xi) and (xii) are unlike in most pliosaurids except for NHMUK PV R2443 (referred to “Pliosaurus” andrewsi by Tarlo 1960, but considered of uncertain taxonomic status herein); (xiii) proportionally short atlas-axis complex due to anteroposterior shortening of the axial centrum, similar to Liopleurodon ferox and Simolestes vorax but differing from the proportionally long atlas-axis complex of other Middle Jurassic pliosaurids, including Marmornectes candrewi, Peloneustes philarchus, “Pliosaurus” andrewsi, and the holotype of “Peloneustes” evansi (Seeley, 1869) (referred to Peloneustes philarchus by Tarlo [1960]), but considered of uncertain taxonomic status herein); (xiv) cervical centra with midline ventral ridges, shared only with Peloneustes philarchus and the holotype of “Peloneustes” evansi among Middle Jurassic pliosaurids; (xv) cervical neural spines with mediolaterally narrow cross-sections and only weak mediolateral expansion at their dorsal tips, similar to Peloneustes philarchus and Liopleurodon ferox, but differing from other Middle Jurassic pliosaurids including the holotype of “Peloneustes” evansi; (xvi) proportionally short epipodials, similar to those of “Pliosaurus” andrewsi and Marmornectes, but differing from other Middle Jurassic pliosaurids, including NHMUK PV R 2443.
Description.—Cranium: The skull of Eardasaurus powelli gen. et sp. nov. measures 890 mm along the dorsal midline, with a pre-orbital portion 485 mm long, thus comprising 54% of skull length (Figs. 2, 3). This is similar to the proportional snout length of Liopleurodon ferox (55% in NHMUK PV R2680) and longer than in Simolestes vorax (50%; Andrews 1913), but is shorter than the more longirostrine Middle Jurassic forms Peloneustes philarchus and “Pliosaurus” andrewsi (approximately 58–59%; Ketchum and Benson 2011b; NHMUK PV R3891). The posterior two-thirds of the cranium were preserved in a septarian concretion, which was partially removed using a combination of acid and manual preparation (H. Philip Powell and Juliet Hay, personal communication 2020 to HFK). The bone in this region is highly fragmented, with numerous areas of broken bone and coloured filler, making many sutures unclear. By comparison, the snout, which was outside of the concretion, is relatively well preserved, although it has been slightly crushed near the tip (Fig. 4).
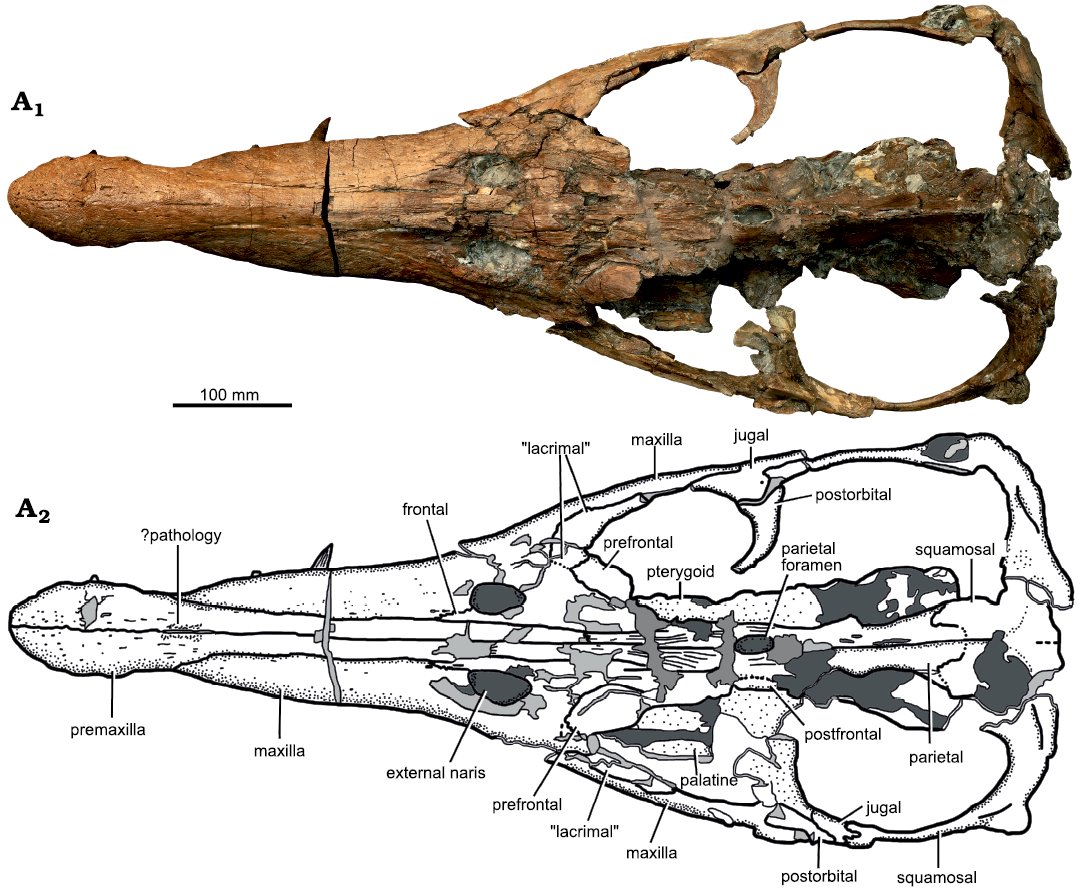
Fig. 2. Holotype of the pliosaurid pleasiosaur Eardasaurus powelli gen. et sp. nov. (OUMNH PAL-J.2247) from the Middle Jurassic of Yarnton, Oxfordshire, UK. Cranium in dorsal view. Photograph (A1), interpretive drawing (A2), light grey, broken bone; mid grey, filler; dark grey, matrix.
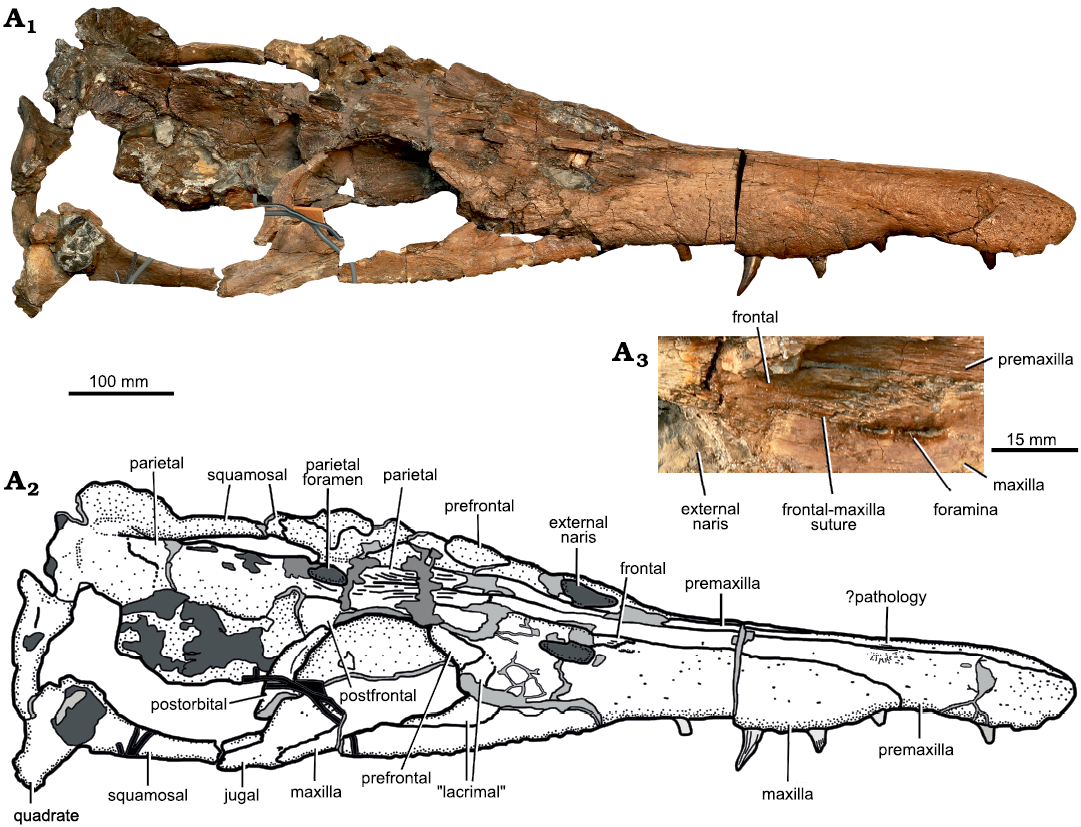
Fig. 3. Holotype of the pliosaurid pleasiosaur Eardasaurus powelli gen. et sp. nov. (OUMNH PAL-J.2247) from the Middle Jurassic of Yarnton, Oxfordshire, UK. Cranium in right lateral view. Photograph (A1), interpretive drawing (A2), light grey, broken bone; mid grey, filler; dark grey, matrix; detail showing region around external naris including frontal–maxilla suture (A3).
Premaxilla: The conjoined premaxillae form the anterior portion of the rostrum (Fig. 4). They contact the maxillae laterally, the parietals posteromedially, and the frontals posterolaterally (in the region of the external naris and more posteriorly) (Figs. 2, 3). Anteriorly, the lateral and dorsal surfaces of the premaxillae bear numerous neurovascular foramina with diameters of 2–4 mm (Fig. 4). The right premaxilla contains five alveoli, whereas the left contains six (Figs. 4, 5). Five or six premaxillary alveoli are also present in all other pliosaurids from the Oxford Clay Formation (Andrews 1913; six are present in Peloneustes philarchus, Marmornectes candrewi, and “Pliosaurus” andrewsi; Ketchum and Benson 2011a, b, and five in Liopleurodon ferox, and possibly Simolestes vorax). Five or six premaxillary alveoli are also present among species of Pliosaurus (Tarlo 1960; Knutsen et al. 2012; Benson et al. 2013; O’Gorman et al. 2018). By contrast, seven premaxillary alveoli are present in species of the early pliosaurid Hauffiosaurus (Benson et al. 2011; Vincent 2011)and the early brachauchenine Luskhan itilensis Fischer, Benson, Zverkov, Soul, Arkhangelsky, Lambert, Stenshin, Uspensky, and Druckenmiller, 2017 (Fischer et al. 2017), whereas four are present in Brachauchenius lucasi Williston, 1903 (Williston 1907), Megacephalosaurus eulerti Schumacher, Carpenter, and Everhart, 2013 (Schumacher et al. 2013), Acostasaurus pavachoquensis Gómez-Pérez and Noè, 2017 (Gómez-Pérez and Noè 2017), Sachicasaurus vitae Páramo-Fonseca, Benavides-Cabra, and Gutiérrez, 2018 (Páramo-Fonseca et al. 2018), and Stenorhynchosaurus munozi Páramo-Fonseca, Gómez-Pérez, Noè, and Etayo-Serna, 2016 (Páramo-Fonseca et al. 2019).
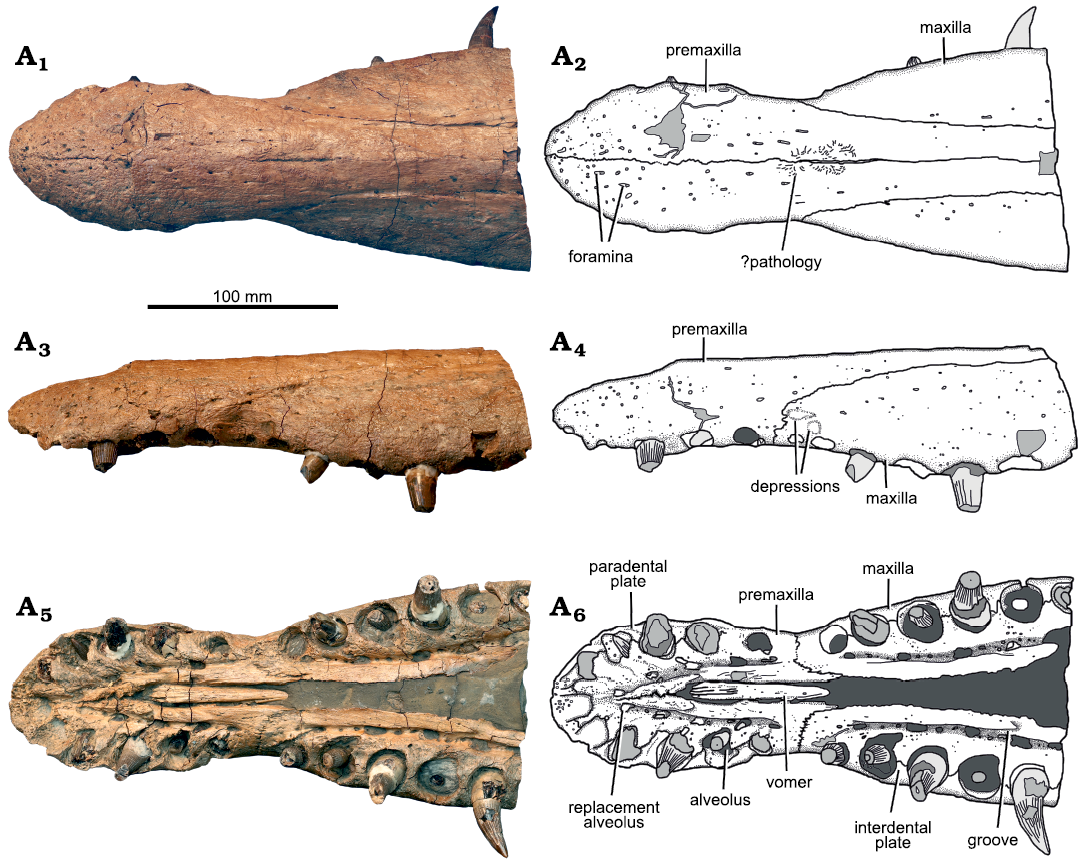
Fig. 4. Holotype of the pliosaurid plesiosaur Eardasaurus powelli gen. et sp. nov. (OUMNH PAL-J.2247) from the Middle Jurassic of Yarnton, Oxfordshire, UK. Rostrum in dorsal (A1, A2), left lateral (A3, A4), and ventral (A5, A6) views. Photographs (A1, A3, A5), interpretive drawings (A2, A4, A6), light grey, broken bone; mid grey, filler; dark grey, matrix.
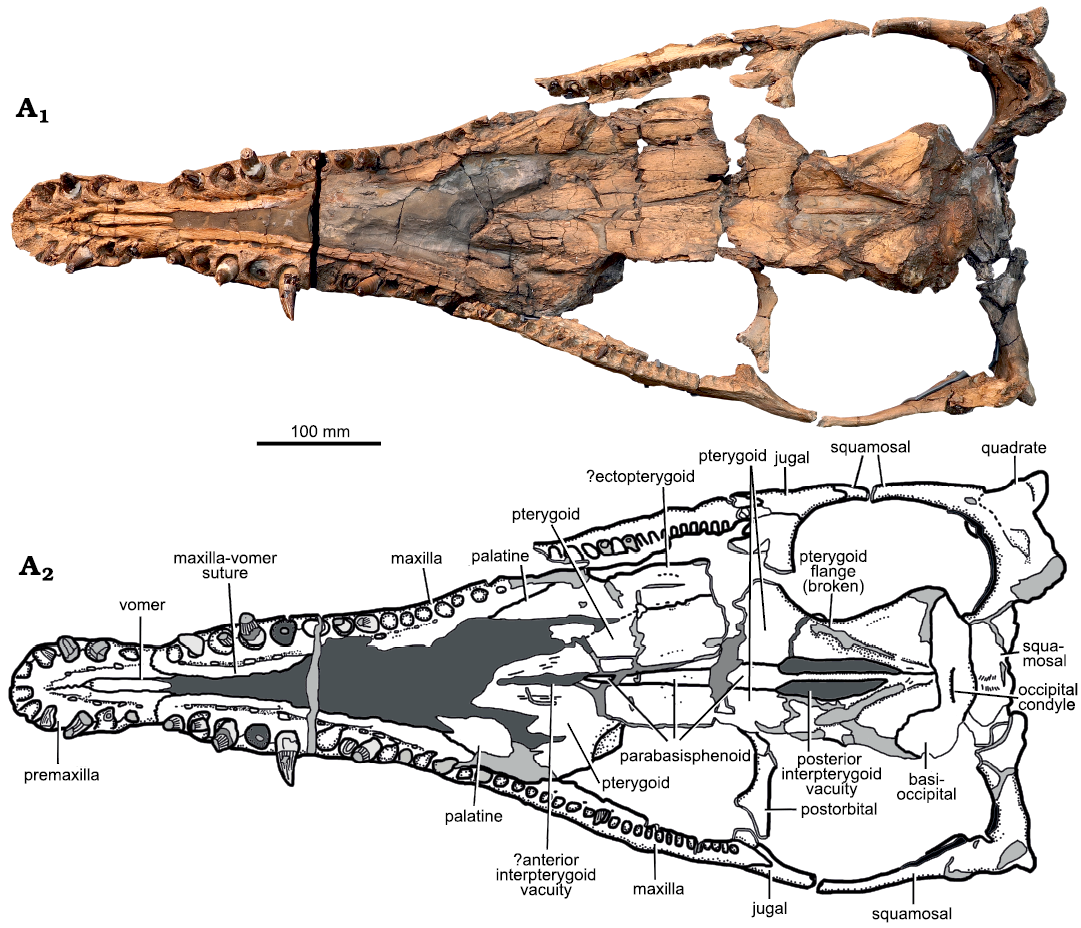
Fig. 5. Holotype of the pliosaurid plesiosaur Eardasaurus powelli gen. et sp. nov. (OUMNH PAL-J.2247) from the Middle Jurassic of Yarnton, Oxfordshire, UK. Cranium in ventral view. Photograph (A1), interpretive drawing (A2), light grey, broken bone; mid grey, filler; dark grey, matrix.
The premaxillary teeth of Eardasaurus powelli gen. et sp. nov. are anisodont, and the penultimate premaxillary alveolus is the largest (Fig. 4). The mesialmost and distalmost alveoli are small compared to adjacent alveoli. However, the mesialmost alveolus is not as substantially reduced as in most species of Pliosaurus (Benson et al. 2013; Benson and Druckenmiller 2014; O’Gorman et al. 2018), in which is it reduced to less than half the diameter of the largest alveolus. The interdental spaces are relatively narrow, as in most other pliosaurids, with maximum lengths of less than half of one alveolar diameter. This differs from the condition in Marmornectes candrewi (Ketchum and Benson 2011b) and Luskhan itilensis (Fischer et al. 2017) in which the alveoli are more widely spaced. The rugose, triangular paradental plates are bounded medially by a groove for the replacement pits (Fig. 4), positioned at the level of the interalveolar walls.
The premaxilla contacts the maxilla at an interdigitating suture that is present within a diastema between the distalmost premaxillary alveolus and the mesialmost maxillary alveolus (Fig. 4). This region is also marked by a prominent mediolateral constriction of the snout, which is clearly evident in dorsal view and which is widespread among Jurassic pliosaurids (e.g., Andrews 1913; O’Keefe 2001), but is weak or absent in is weak or absent in species of Hauffiosaurus, Marmornectes candrewi, and “Pliosaurus” andrewsi (Ketchum and Benson 2011a; Benson et al. 2011; Vincent 2011; NHMUK PV R3891, Andrews 1913). The length of the diastema exceeds (on the right) or is similar to (on the left) that of the largest maxillary alveolus, and the presence of this diastema differs from the condition in most other Middle Jurassic pliosaurids except for Liopleurodon ferox (Andrews 1913; i.e., the diastema is absent in Marmornectes candrewi, Peloneustes philarchus, and “Pliosaurus” andrewsi; Ketchum and Benson 2011a, b; NHMUK PV R3891; the condition is not determined in Simolestes vorax). The premaxilla/maxilla suture extends dorsally, then posterodorsally along the lateral surface of the snout, forming the medial margin of the posteromedian process of the premaxilla. The median contact between the premaxillae on the ventral surface of the skull extends from the anterior tip of the snout posteriorly, up to the level of the third/fourth premaxillary alveolus. In the posterior half of this contact, the ventral surface of the premaxilla bears a posterolaterally oriented, rugose ridge. Together, these ridges on the right and left premaxillae form a V-shaped notch that accommodates the anterior end of the vomer.
The median contact between the premaxillae on the dorsal surface of the skull is approximately straight, with some limited sinuosity in the region anterior to the rostral constriction (Figs. 2–4). Just posterior to this, a mediolaterally narrow, slit-like median opening 38 mm long interrupts the suture. This opening is surrounded by an irregular, raised region marked by fine-scale rugosity and so appears to be pathological (Fig. 4). A similar opening is absent in other pliosaurids, but is present in some rhomaleosaurids, in which it is generally larger and more well-defined (e.g., Smith and Dyke 2008; Smith and Benson 2014). The posteromedian process of the premaxilla extends posteriorly to approximately the level of the anterior margin of the orbit, where it contacts the parietal in an interdigitating suture that is somewhat obscured by fragmentation of the bone surface. This morphology is similar to that seen in many other thalassophoneans, including Peloneustes philarchus (Ketchum and Benson 2011b) and some species of Pliosaurus (Benson et al. 2013). It is unlike the situation in Acostasaurus pavachoquensis, Pliosaurus westburyensis Benson, Evans, Smith, Sassoon, Moore-Fay, Ketchum, and Forrest, 2013, and Simolestes vorax in which the premaxilla extends further posteriorly to around orbital midlength (Andrews 1913; Taylor and Cruickshank 1993; Gómez-Pérez and Noè 2017), and unlike the condition in some brachauchenines such as Luskhan itilensis and Megacephalosaurus eulerti in which the premaxilla terminates further anterior to the orbit (Schumacher et al. 2013; Fischer et al. 2017).
Maxilla: The maxillae form the lateral portions of the snout, including the central and posterior parts of the tooth row (Figs. 2–5). The right maxilla bears 34 or 35 alveoli, and the less complete left maxilla likely bore a similar number (only 30 are preserved but the posterior portion is missing). This is a high number compared to that in other pliosaurids, including early-diverging taxa such as Hauffiosaurus zanoni O’Keefe, 2001 (29; Vincent 2011) and Peloneustes philarchus (30–31; Ketchum and Benson 2011b), and especially in more derived forms such as “Pliosaurus” andrewsi (22; NHMUK PV R3891) Liopleurodon ferox (20; Andrews 1913), species of Pliosaurus (20–27; Knutsen 2012; Benson et al. 2013) and brachauchenines (e.g., 18–19 in Megacephalosaurus eulerti; Schumacher et al. 2013); although the Early Cretaceous brachaucheninine Stenorhynchosaurus munozi has 28–30 (Páramo-Fonseca et al. 2016, 2019). The maxillary dentition of Eardasaurus powelli gen. et sp. nov. is highly anisodont, as in most pliosaurids. The fifth alveolus is the largest, and the diameters of the alveoli decrease posteriorly from there. The tooth row terminates ventral to the posterior part of the postorbital bar. As in the premaxilla and dentary, the maxillary alveoli are bound medially by subtriangular paradental plates.
The maxilla is excluded from the anteroventral and ventral margins of the orbit by the presence of a “lacrimal” (a neomorphic ossification described below and not homologous to the lacrimal of other tetrapods; Figs. 2, 3, 6). The “lacrimal”-maxilla suture is clear in the antorbital region. However, sutures between the maxilla and most other elements in this region are not evident due to damage. Nevertheless, it is clear that the maxilla forms the lateral and anterior margins of the external naris only, and does not extend further posteriorly along the medial margin of the external naris (Figs. 2, 3). Instead, the maxilla terminates anteromedial to the external naris, forming a deeply interdigitating, anteromedially-oriented suture with the frontal (Fig. 3A3). This differs from other Middle Jurassic pliosaurids that preserve this region, in which the maxilla extends further posteriorly to form part of the medial margin of the external naris (Liopleurodon ferox, NHMUK PV R2680; Peloneustes philarchus, Ketchum and Benson 2011b; Simolestes vorax, Noè 2001). The portion of the maxilla anterior to the external naris bears two oval foramina approximately 6–7 mm in maximum anteroposterior diameter (Fig. 3A3). On the right side, these foramina participate in a deep, anteroposteriorly-oriented fissure in the maxilla anterior to the external naris (Fig. 3A3). On the left, both foramina contact the suture between the maxilla and frontal posteriorly.
Interorbital skull roof: Due to fragmentation of the interorbital portion of the skull roof, sutures in this region (between the frontal, prefrontal, postfrontal) are unclear. Nevertheless, the anterodorsal margin of the orbit is embayed by a convex lateral extension of the skull roof (Figs. 2, 3). This extension is widespread among thalassophonean pliosaurids (e.g., Ketchum and Benson 2011b; Gómez-Pérez and Noè 2017).
Jugal: The jugal forms the posteroventral margin of the orbit and is quite large in OUMNH PAL-J.2247 compared to other plesiosaurians (Figs. 2, 3, 6). It extends posteriorly approximately one-third of the length of the temporal fenestra, where it contacts the anterior ramus of the squamosal. It extends dorsally as a convex eminence that contacts the postorbital, therefore forming the ventral part of the postorbital bar. It also extends anteriorly, forming the posteroventral margin of the orbit. At least three large foramina are present, distributed over the lateral surface of the jugal.
The jugal of OUMNH PAL-J.2247 is fluted by a series of grooves that extend posteriorly from the posteroventral margin of the orbit (Fig. 6). A similar fluted texture is present on several bones forming the orbital rim, including the postfrontal, postorbital prefrontal and palpebral, of other thalassophoneans (Ketchum and Benson 2010: fig. A3), and on many of the bones, including the frontal and postfrontal which form the orbital rim in cryptoclidids such as Muraenosaurus leedsii Seeley, 1874b (Andrews 1910: pl. 3; Maisch 1998). However, the presence of this fluted ornamentation on the jugal in Eardasaurus powelli gen. et sp. nov. is unique among pliosaurids reported so far, and is regarded here as an autapomorphy.
The contact between the jugal and squamosal of Eardasaurus powelli gen. et sp. nov. has a step-like morphology in which a long anterior extension of the squamosal dorsally overlies a posterior process of the jugal for about 35 mm (Fig. 6). This is unlike the condition seen in other pliosaurids, which mostly have a squamosal-jugal suture that is interdigitating and oriented approximately dorsoventrally, and may therefore be an autapomorphy. The morphology in Eardasaurus powelli gen. et sp. nov. is most similar to the morphology in species of Pliosaurus, in which a dorsoventrally slender posterior process of the jugal underlies the squamosal ventrally (Benson et al. 2013). However, it differs from this in that the posterior process of the jugal is thicker, constituting approximately half the dorsoventral height of the subtemporal bar in Eardasaurus powelli gen. et sp. nov.
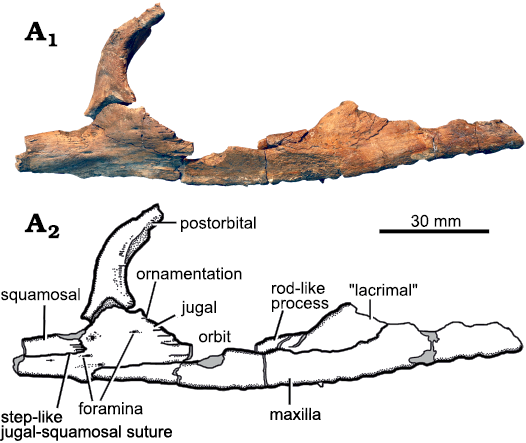
Fig. 6. Holotype of the pliosaurid plesiosaur Eardasaurus powelli gen. et sp. nov. (OUMNH PAL-J.2247) from the Middle Jurassic of Yarnton, Oxfordshire, UK. Detail of right ventral orbital margin in lateral view. Photograph (A1), interpretive drawing (A2), light grey, broken bone.
“Lacrimal”: The presence of a distinct, neomorphic ossification, typically called the “lacrimal” (see Williston 1907; Andrews 1913; Benson et al. 2011; Ketchum and Benson 2011b; although not homologous to the lacrimal of other amniotes) in pliosaurids is debated, with some authors arguing that this is just a long anterior process of the jugal (e.g., Carpenter 1996; O’Keefe 2001; Schumacher et al. 2013). The specimen of Eardasaurus powelli gen. et sp. nov. does not help to resolve this debate because of breakage ventral to the orbit (Figs. 2, 3). Nevertheless, because we have observed a “lacrimal”-jugal suture in other thalassophoneans (Ketchum and Benson 2010, 2011a), we describe the morphology of OUMNH PAL-J.2247 as though it also had a distinct “lacrimal” ossification that is separate from the jugal. The suborbital portion of the “lacrimal” in OUMNH PAL-J.2247 is dorsoventrally thin, approximately 8 mm in dorsoventral thickness on both sides compared to 25 mm for the dorsoventral thickness of the underlying maxilla around orbital midlength (Fig. 6). This narrow morphology of the “lacrimal” differs from other pliosaurids (e.g., Williston 1907; O’Keefe 2001; Ketchum and Benson 2011b), and is considered here to be an autapomophy of Eardasaurus powelli gen. et sp. nov.
Postorbital: The postorbital forms most of the postorbital bar from the skull roof dorsally, extending from its dorsal/medialmost point just lateral to the parietal foramen, down to its ventral contact with the jugal ventrally and squamosal posteroventrally (Figs. 2, 3). Contact with the squamosal is evident from the morphology of the facet for the postorbital on the jugal. However, the posteroventral process of the (better-preserved) right postorbital, which would originally have contacted the squamosal, is broken. The ventral part of the postorbital is anteroposteriorly broad, and it tapers dorsally. It has a subtriangular cross-section, with a flat lateral surface and prominent medial crest (Fig. 6). The lateral surface of the postorbital is rugose adjacent to the orbit. The posteroventral process of the postorbital extends posteriorly over the dorsal surface of the jugal to approximately one-quarter the length of the subtemporal bar.
Parietal: The parietals enclose a large, oval parietal foramen along the midline of the cranium, adjacent to the posterior margin of the orbit (Figs. 2, 3). Anterior to the parietal foramen, the parietals are ornamented with longitudinal ridges and grooves, and contact the premaxillae in an interdigitating suture adjacent to the anterior orbit margin, as in many Middle Jurassic pliosaurids (e.g., Andrews 1913; Ketchum and Benson 2011b). Posterior to the parietal foramen the parietal rises dorsally and forms a narrow sagittal crest. The sagittal crest may be broken along its dorsal margin, so its true original height cannot be confidently determined. More posteriorly, it contacts the squamosal forming a roof over the braincase.
Squamosal: The squamosal is a triradiate bone that forms the posterior and lateral margins of the temporal fenestra (Figs. 2, 3). The conjoined squamosals contact the parietal along the midline in a broad semi-circular suture. Posteriorly, the squamosals expand and rise in height towards the vertex at the posteriormost point of the skull. In Middle Jurassic pliosaurids such as Peloneustes philarchus, the squamosals are usually expanded at the vertex to form a mediolaterally narrow, but prominent “bulb” (e.g., Ketchum and Benson 2011b). In contrast to this, the squamosal morphology of Eardasaurus powelli gen. et sp. nov. appears to be more similar to that of Late Jurassic pliosaurids such as Pliosaurus kevani Benson, Evans, Smith, Sassoon, Moore-Fay, Ketchum, and Forrest, 2013, in which the conjoined squamosals form a mediolaterally broad convexity that projects posteriorly (Benson et al. 2013). However, there is some damage in this area, and it is possible that a squamosal bulb was originally present.
Quadrate: A partial quadrate is preserved on the left side of the skull, bounded by the ventral ramus of the squamosal. However, it is crushed and the morphology is therefore difficult to determine (Fig. 3).
Vomer: The vomer forms a single midline element that emerges anteriorly adjacent to the fourth premaxillary alveolus, as in other Oxford Clay pliosaurids (Andrews 1913). It forms a narrow and ventrally convex bar along the midline, which is ornamented with anteroposteriorly oriented grooves (Figs. 4, 5). Posterior to the premaxilla–maxilla suture, the vomer is completely obscured by matrix, although the maxilla–vomer suture remains visible along the medial margin of the matrix. Posteriorly, the matrix obscures the contact between the vomer and palatine, and the location of the internal nares.
Palatine: The palatines are obscured by matrix for the majority of their length (Fig. 5). Posteriorly, they are exposed at around skull midlength, contacting the maxilla laterally and pterygoid medially. The palatines are dorsoventrally thin and plate-like bones, and are highly fractured and fragmented. Due to the poor preservation in this region, many of the sutures between the palatine, pterygoid and ectopterygoid could not be recognised.
Ectopterygoid: A small fragment of the left ectopterygoid may be preserved lateral to the palatine, although the suture cannot be identified with confidence (Fig. 5). The equivalent area on the right-hand-side of the palate is not preserved.
Pterygoid: The pterygoids are large, complex bones that form much of the palate (Fig. 5). The anterior processes of the pterygoids extend anteriorly along the midline, contacting the ectopterygoids and palatines laterally. Posterior to this, the pterygoids underlap the braincase ventrally, and give rise to posterolateral processes that contact the quadrates. An area of matrix along the midline contact between the anterior processes indicates the possible presence of a slit-like interpterygoid vacuity. However, it is also possible that this results from breakage and partial displacement. Posterior to this, the pterygoids are separated along the midline by the cultriform process of the parasphenoid.
Anteroposteriorly elongate posterior interpterygoid vacuities are present ventral to the braincase, with a length:width ratio of approximately 2.75. This is similar to or slightly greater than the relative length of the posterior interpteryoid vacuities of most other Middle Jurassic pliosaurids (ratio 2.4–2.5 in Peloneustes philarchus, Liopleurodon ferox, and Simolestes vorax; e.g., Andrews 1913; Ketchum and Benson 2011b; NHMUK PV R2680; HFK and RBJB, personal observation), and considerably greater than that of “Pliosaurus” andrewsi (ratio 1.75; NHMUK PV R3891). Lateral to these, the pterygoids form ventrolaterally directed flanges that extend anterolaterally from a point of midline contact posterior to the posterior interpterygoid vacuities. These flanges are present in all thalassophonean pliosaurids (e.g., Williston 1907; Andrews 1913; Schumacher et al. 2013; Ketchum and Benson 2011b), although they are often crushed dorsolaterally, and partially broken, as in Eardasaurus powelli gen. et sp. nov. The flanges of Eardasaurus powelli gen. et sp. nov. contact posteriorly along the midline, as in many early thalassophoneans, but unlike in some brachauchenines (e.g., Williston 1907; Schumacher et al. 2013; Gómez-Pérez and Noè 2017).
Braincase: The braincase is preserved, crushed between the skull roof and pterygoids. This limits the detail with which elements can be observed and we restrict our description to the visible portions, comprising the parabasisphenoid and basioccipital.
Parabasisphenoid: The parasphenoid and basisphenoid are conjoined, and the line of fusion between them cannot be discerned (Fig. 5). The parabasisphenoid bisects the posterior interpterygoid vacuities as a narrow convex bar. Anteriorly, it widens and flattens, extending along the midline of the palate between the pterygoids as the cultriform process.
Basioccipital: The basioccipital forms the posterior part of the basicranium and bears the occipital condyle (Fig. 5). It is highly abraded and the shape of the occipital condyle is difficult to discern, but appears to be low, hemispherical, and separated from the body of the basioccipital by a shallow groove ventrally.
Mandible: The mandible is almost complete and three-dimensionally preserved, missing only the margins of the four anterior-most dentary alveoli (Figs. 7–9). The mandible is 932 mm long, measured from the anterior tip to the posterior margin of the retroarticular process (873 mm long from the anterior tip to the posterior margin of the articular condyles), and 356 mm wide between the articular condyles. The symphysis is 200 mm long anteroposteriorly, with a maximum width of 127 mm adjacent to the fifth dentary alveolus (Fig. 8). Thus, the symphysis is quite strongly expanded mediolaterally, similar to many Middle–Late Jurassic pliosaurids, which exhibit at least moderate expansion of the symphysis (e.g., Andrews 1913), although this expansion is weaker in Marmornectes candrewi (Ketchum and Benson 2011a) and “Pliosaurus” andrewsi (NHMUK PV R3891).
Dentary: The dentary forms the major part of the mandible, including the symphysis, and terminates just posterior to the tooth row, at the level of the coronoid eminence (Fig. 7). Each dentary bears 38 alveoli, of which nine are adjacent to the symphysis (Figs. 7, 8). Thus the symphysis is relatively shorter than those of other longirostrine pliosaurids from the Oxford Clay Formation, such as Peloneustes philarchus (13–15 symphysial alveoli; Ketchum and Benson 2011b), Marmonectes candrewi (12 symphysial alveoli; Ketchum and Benson 2011a), and “Pliosaurus” andrewsi (11; Tarlo 1960), but longer than in Simolestes vorax (five symphysial alveoli) and Liopleurodon ferox (6.5 symphysial alveoli; Andrews 1913). Dentary alveoli 3–7 of Eardasaurus powelli gen. et sp. nov. are the largest in the mandible, and the fifth alveolus is marginally the largest of these, being located in the widest part of the symphysis (Fig. 8). The eighth alveolus is much smaller, being approximately 60% of the diameter of the seventh. Alveolus size decreases gradually from here posteriorly. The posteriormost alveolus of the left dentary is infilled with rugose bone, similar to that described in Peloneustes philarchus (Ketchum and Benson 2011b), and likely pathological. As in the maxilla and premaxilla, the medial walls of the dentary aveoli are bounded by rugose, subtriangular paradental plates (Fig. 8). Medial to the paradental plates is a longitudinal groove containing the replacement alveoli.
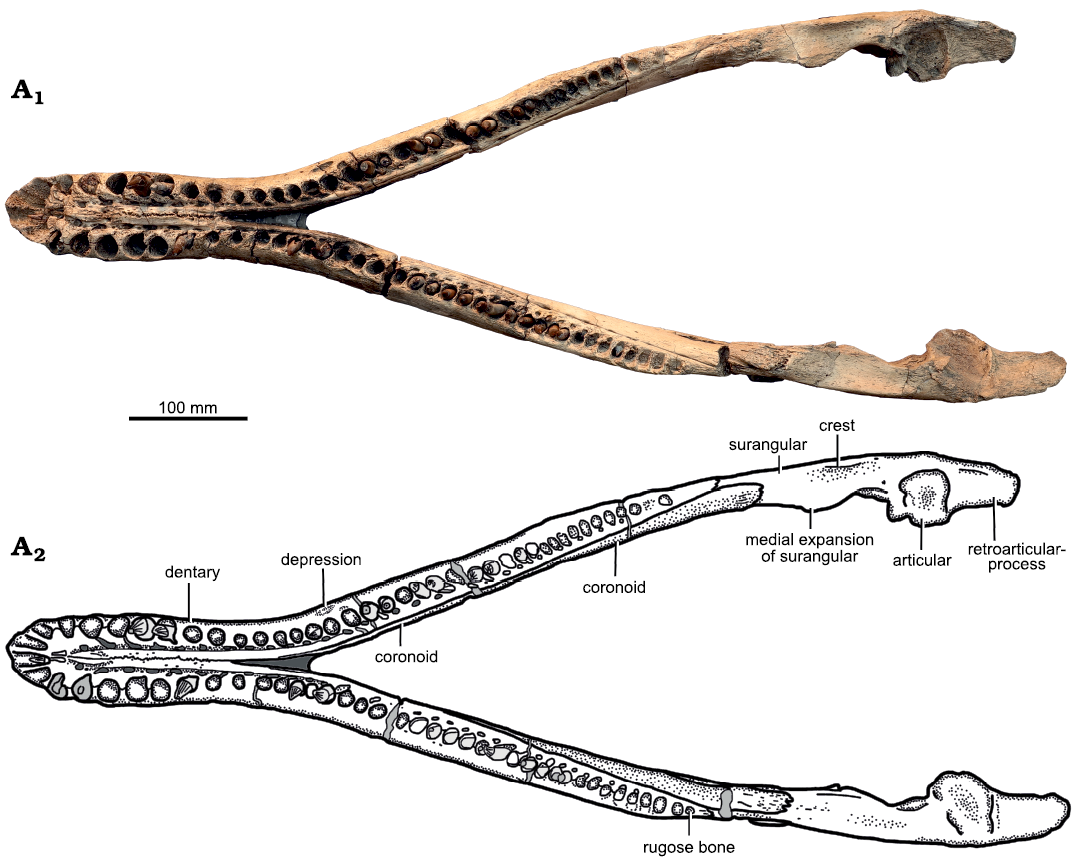
Fig. 7. Holotype of the pliosaurid plesiosaur Eardasaurus powelli gen. et sp. nov. (OUMNH PAL-J.2247) from the Middle Jurassic of Yarnton, Oxfordshire, UK. Mandible in dorsal view. Photograph (A1), interpretive drawing (A2), light grey, broken bone; mid grey, filler; dark grey, matrix.
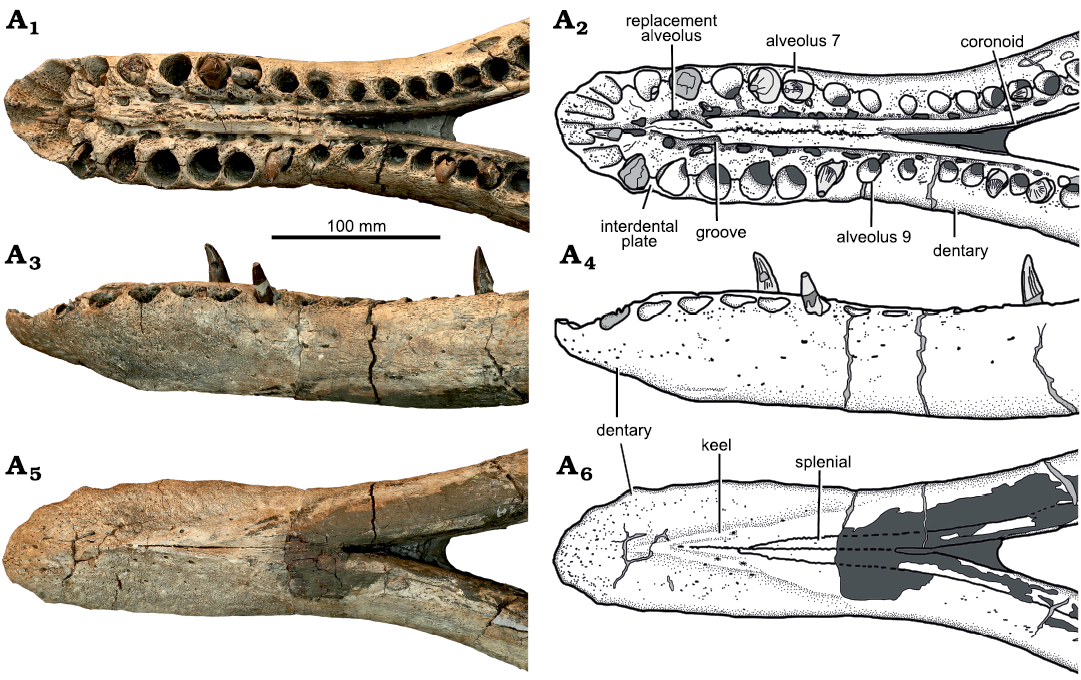
Fig. 8. Holotype of the pliosaurid plesiosaur Eardasaurus powelli gen. et sp. nov. (OUMNH PAL-J.2247) from the Middle Jurassic of Yarnton, Oxfordshire, UK. Mandibular symphysis in dorsal (A1, A2), left lateral (A3, A4), and ventral (A5, A6) views. Photographs (A1, A3, A5), interpretive drawings (A2, A4, A6), light grey, broken bone; mid grey, filler; dark grey, matrix.
The dentaries are tightly joined anteriorly along the midline to form the majority of the mandibular symphysis, other than a small posteroventral contribution of the splenial (Fig. 8A6). The ventral exposure of the interdentary suture is tightly interdigitating at its anteriormost end and becomes straight at approximately midlength up to its posterior end, where it protrudes ventrally as a low median keel. Numerous grooves and foramina are present on the ventral and lateral surfaces of the dentary, and are smaller and more abundant anteriorly (Figs. 7–9).
The dorsal exposure of the interdentary suture interdigitates along its entire length (Fig. 8). The suture is located on a robust longitudinal ridge, delimited laterally on either side by the groove for replacement alveoli. This morphology is similar to that in most other pliosaurids, but is unlike the condition in Peloneustes philarchus in which the symphysis is raised up on an additional, narrower median ridge (Ketchum and Benson 2011b). The median symphysial ridge in Eardasaurus powelli gen. et sp. nov. is embayed by an oblique, posteromedially oriented groove on either side at the level of the fifth alveolus on the left, and between the fourth and fifth alveoli on the right (Fig. 8). These grooves are widespread among plesiosaurians with long symphyses, including both pliosaurids (e.g., Noè et al. 2004; Albright et al 2007a; Gasparini and O’Gorman 2014; Gómez-Pérez and Noè 2017; Holland 2018) and polycotylids (e.g., Albright et al. 2007b; Schumacher and Martin 2015), and are at least slightly asymmetrical (as in Eardasaurus powelli gen. et sp. nov.) in several other pliosaurids, including Peloneustes philarchus (Ketchum and Benson 2011b), Marmornectes candrewi (Ketchum and Benson 2011a), Pliosaurus patagonicus O’Gorman, 2014 (Gasparini and O’Gorman 2014) and Kronosaurus queenslandicus Longman, 1924 (Holland 2018).
Posterior to the symphysis, the lateral surface of the dentary forms the lateral and alveolar surfaces of the mandible, and is overlapped medially by the surangular and angular (Fig. 9). A longitudinal row of small, evenly spaced foramina is present on the lateral surface of the dentary at two-thirds of its total height. This row of foramina continues posteriorly to approximately half the length of the dentary.
The dentary bifurcates posteriorly into a dorsoventrally deep posterodorsal process and a narrower posteroventral process. The posterior margins of both processes are deeply crenulated. A large, anteroposteriorly-elongate fenestra, is present around midheight in the lateral surface of the mandible, just anterior to the coronoid eminence (Figs. 9, 10). We refer to this as the intramandibular fenestra. The anterior and dorsal margins of the intramandibular fenestra are bounded by the split between the posterodorsal and posteroventral processes of the dentary. The intramandibular fenestra penetrates to the medial surface of the mandible by extending posteromedially through the suture between the angular and surangular. A prominent intramandibular fenestra is also present in “Pliosaurus” andrewsi (NHMUK PV R3891; HFK and RBJB, personal observation). Unfortunately its presence or absence in other pliosaurids, including Peloneustes philarchus, is difficult to determine, because crushing and breakage is common in this region (Ketchum and Benson 2011b).
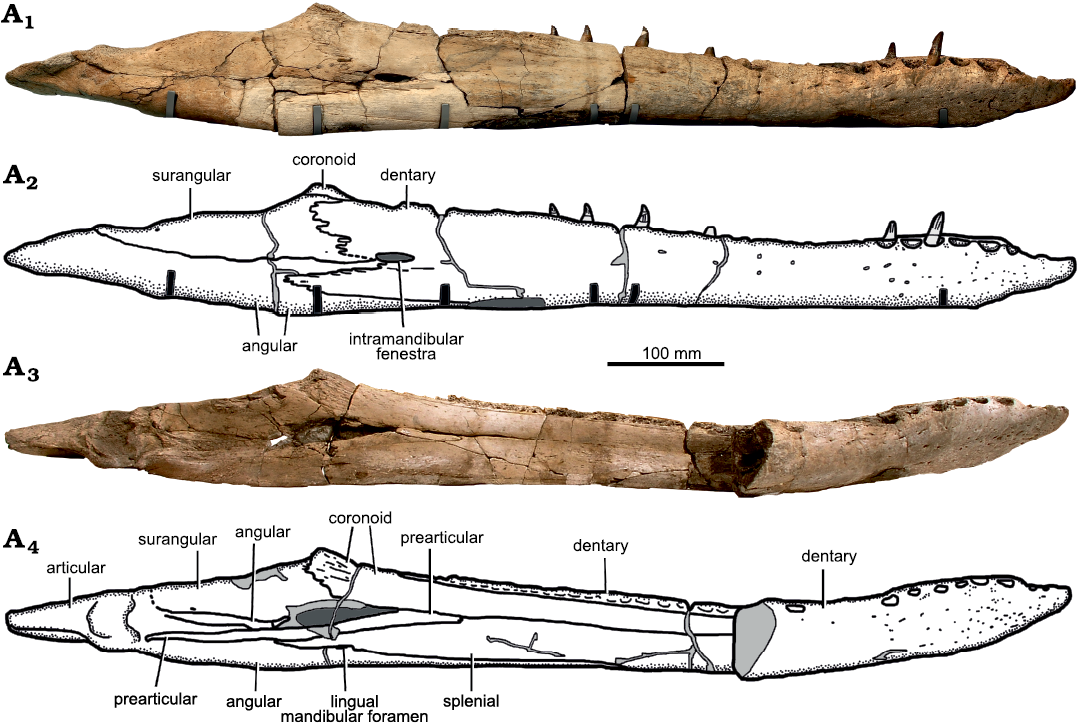
Fig. 9. Holotype of the pliosaurid plesiosaur Eardasaurus powelli gen. et sp. nov. (OUMNH PAL-J.2247) from the Middle Jurassic of Yarnton, Oxfordshire, UK. Mandible in right lateral (A1, A2) and medial (A3, A4) views. Photographs (A1, A3), interpretive drawings (A2, A4), light grey, broken bone; mid grey, filler; dark grey, matrix.
Splenial: The splenial is an anteroposteriorly elongate bone that forms the ventral half of medial surface of the mandible (Fig. 9), covering the medial surface of the dentary anteriorly, and of the angular posteriorly. It extends from the mandibular symphysis anteriorly, to the posterior opening of Meckel’s canal posteriorly, where it contacts the prearticular dorsally. The paired splenials contribute to the posterior half of the ventral surface of the mandibular symphysis (Fig. 8) as in other pliosaurids (e.g., Andrews 1913; Ketchum and Benson 2011a, b; Schumacher et al. 2013; Fischer et al. 2017; Páramo-Fonseca et al. 2019). Their ventral surface is flat, and does not contribute to the median keel of the dentary symphysis. Posterior to the symphysis on the medial surface, the splenial encloses the ventral and posterodorsal margins of the anteromedial opening of Meckel’s canal (Fig. 11), as in Pliosaurus kevani (Benson et al. 2013: figs. 16, 17), but unlike Peloneustes philarchus, Acostasaurus pavachoquensis, Pliosaurus almanzaensis O’Gorman, Gasparini, and Spalletti, 2018, and possibly Sachicasaurus vitae, in which the splenial is limited to the ventral margin (Ketchum and Benson 2011b: text-fig. 12; Gómez-Pérez and Noè 2017: text-fig. 11; O’Gorman et al. 2018: 248; Páramo-Fonseca et al. 2018: fig. 3). Posteriorly, a small, anteroposteriorly elongate foramen penetrates the splenial–angular contact (the lingual mandibular foramen, Fig. 9A4), as in other pliosaurids (e.g., Ketchum and Benson 2011b; Benson et al. 2013).
Coronoid: The coronoid is an anteroposteriorly long, plate-like bone that covers the dorsal half of the medial surface of the mandible and forms the prominent medial portion of the coronoid eminence posteriorly (Figs. 7, 9, 10). Anteriorly, it forms the anterodorsal margin of the opening into Meckel’s canal (Fig. 11). Its anteriormost extent cannot be determined due to the remnants of the septarian concretion obscuring the median surface of the mandible immediately posterior to the symphysis. However, it is not exposed on the posterodorsal surface of the symphysis, unlike in Peloneustes philarchus (Ketchum and Benson 2011b) and Kronosaurus queenslandicus (Holland 2018).
The coronoid portion of the coronoid eminence is tall and rugose, extending dorsal to both the surangular and dentary so it is distinctly visible in lateral view (Fig. 10). This also occurs in NHMUK PV R2443 (Nikolay Zverkov, personal communication to RBJB), a specimen that was referred to “Pliosaurus” andrewsi by Tarlo 1960). However, it is unlike the situation in most other pliosaurids, including Liopleurodon ferox and Simolestes vorax (Andrews 1913), Peloneustes philarchus (e.g., Ketchum and Benson 2011b), Pliosaurus kevani (Benson et al. 2013), Stenorhynchosaurus munozi (Páramo-Fonseca et al. 2016), Acostasaurus pavachoquensis (Gómez-Pérez and Noè 2017) and Kronosaurus queenslandicus (Holland 2018), in which the coronoid is low and therefore concealed by the dentary and surangular in lateral view. The coronoid is low and partially visible in Megacephalosaurus eulerti (Schumacher et al. 2013: fig. 9C, D).
Angular: The angular is a large element, which first appears on the ventral surface of the mandible anteriorly as a narrow longitudinal splint of bone between the dentary and splenial at approximately mandibular midlength. Posteriorly, the external exposure of the angular expands mediolaterally and then dorsoventrally so that it forms the posterior half of the mandible from approximately the level of the coronoid eminence and more posteriorly, ventrally underlapping the articular, including both the glenoid and the retroarticular process (Figs. 7, 9). Ventral to the coronoid eminence, the dorsomedial surface of the angular forms a shallow trough that extends anteriorly to the intramandibular fenestra and posteriorly to the glenoid, where it is overlapped by a rugose anterior extension of the articular.
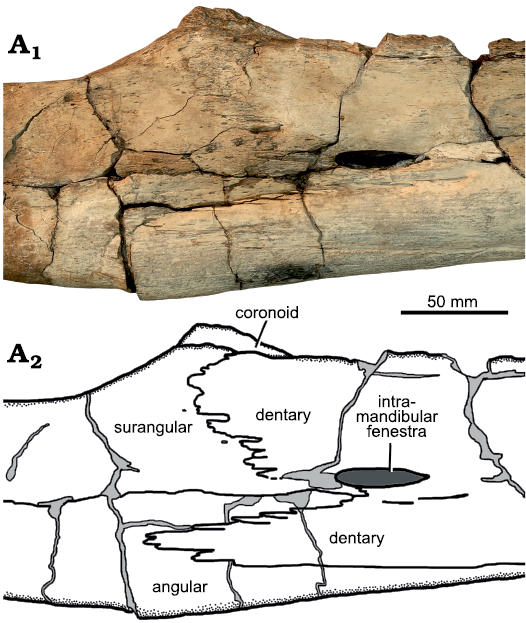
Fig. 10. Holotype of the pliosaurid plesiosaur Eardasaurus powelli gen. et sp. nov. (OUMNH PAL-J.2247) from the Middle Jurassic of Yarnton, Oxfordshire, UK. Detail of right coronoid eminence in lateral view. Photograph (A1), interpretive drawing (A2), light grey, broken bone; dark grey, matrix.
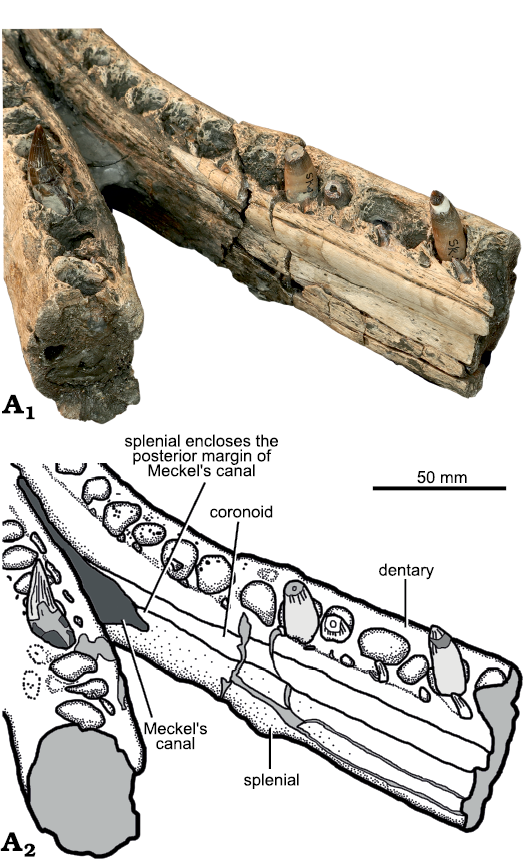
Fig. 11. Holotype of the pliosaurid plesiosaur Eardasaurus powelli gen. et sp. nov. (OUMNH PAL-J.2247) from the Middle Jurassic of Yarnton, Oxfordshire, UK. Detail of posterior end of mandibular symphysis. Photograph (A1), interpretive drawing (A2), light grey, broken bone; dark grey, matrix.
Surangular: The surangular forms the dorsal half of the mandible between the glenoid and the coronoid eminence, and encloses the dorsal margin of the intermandibular fenestra internally (Figs. 7, 9). The lateral surface of the surangular is overlapped by the deeply-crenulated posterodorsal process of the dentary anteriorly, ventral to the coronoid eminence (Fig. 10). The dorsal surface of the surangular bears a shallow anteroposterorly elongate fossa, as in most other pliosaurids (e.g., Ketchum and Benson 2011a; Fig. 7). However, in Eardasaurus powelli gen. et sp. nov. the medial margin of the surangular adjacent to the fossa is strongly expanded compared to most other pliosaurids including Marmornectes candrewi, Peloneustes philarchus (Ketchum and Benson 2011a, b), Simolestes vorax, Liopleurodon ferox (Noè 2001), Kronosaurus queenslandicus (Holland 2018), Acostasaurus pavachoquensis (Gómez Pérez and Noè 2017), and Brachauchenius lucasi (Albright et al. 2007a). Nevertheless, a similar morphology is present in NHMUK PV R2443 (Tarlo 1960) and there may be scope for within-species variation in the morphology of this site of muscle attachment.
The extent of any contribution of the surangular to the ventral surface of the glenoid cannot be determined because its suture with the articular in this region is closed. However, it certainly contributes to the anterolateral margin of the glenoid as a dorsoventrally oriented, crescentic process. Anterior to this, on the medial surface of the jaw, the surangular encloses a small foramen, as in Peloneustes philarchus (Ketchum and Benson 2011b; Fig. 9).
Prearticular: Both the left and right prearticulars are well preserved, which is unusual among pliosaurids, where it is frequently only partly preserved (e.g., Ketchum and Benson 2011b). The prearticular is an elongate, medioventrally thin, splint-like bone that articulates on to the dorsomedial surface of the angular (Fig. 9C, D). It extends anteriorly from its origin, anteroventral to the glenoid, tapering anteriorly to a point just anterior to the coronoid process. Together with the angular it forms the ventral margin of the intramandibular fenestra.
Articular: The articular forms the articular cotyles and the dorsal portion of the retroarticular process, both of which are rotated so their “dorsal” surfaces face dorsomedially (Fig. 9). As in Marmornectes candrewi, but unlike Peloneustes philarchus, the articular also forms the ventrolateral surface of the retroarticular process (Ketchum and Benson 2011a), forming a suture with the surangular at the widest point of the mandibular glenoid. As in other pliosaurids, a rugose, triangular anterior process of the articular extends from the glenoid along a shallow trough in the angular, just dorsal to the prearticular.
Sclerotic ring: Four disarticulated scleral ossicles are associated with the skull. They are irregular in shape, with crenulated margins (Fig. 12A, B). As in Peloneustes philarchus (Ketchum and Benson 2011b), the ossicles are very thin adjacent to the internal margin (forming the corneal aperture) and external margin of the sclerotic ring, and slightly more robust in between. The lateral (outwardly facing) surface of the sclerotic ring is gently convex.
Hyoids: Two elongate elements were found close by to the skull, and are almost certainly hyoid bones (Fig. 12C). As in the hyoid elements of Peloneustes philarchus (NHMUK PV R 8574; Ketchum and Benson 2011b), Pliosaurus almanzaensis (O’Gorman et al. 2018), and Sachicasaurus vitae (Páramo-Fonseca et al. 2018) the shafts are narrow, elongate, and gently curved. However, in Eardasaurus powelli gen. et sp. nov., one end of the shaft tapers to a rounded point, unlike in Peloneustes philarchus and possibly Pliosaurus almanzaensis, in which both ends of the shaft are gently expanded (Ketchum and Benson 2011b: text-fig. 11; O’Gorman et al. 2018: fig. 3).
Dentition: The mesial teeth of Eardasaurus powelli gen. et sp. nov. are large, conical and slightly recurved, with approximately circular cross-sections (Fig. 13). Several teeth bear apical wear facets. The enamel is ornamented with fine apicobasal enamel ridges, which are present on all surfaces of the tooth, but more densely packed on the recurved lingual (concave) surface (Fig. 13A2). Enamel ridges are finer and more widely spaced, mostly restricted to the basal region, on the labial (convex) surface (Fig. 13A4; note that reduction of enamel ornamentation on the lingual surface of the tooth may be a common pattern amongst aquatic predators; McCurry et al. 2019). On all surfaces of the crown, the majority of ridges originate from near the base of the enamel, but a small number originate part way up the tooth. In the largest teeth with intact crowns, between two and five of the enamel ridges are particularly prominent, and extend to the tip of the tooth from either side of the convex (mesial) surface, with the appearance of carinae (Fig. 13A8).

Fig. 12. Holotype of the pliosaurid plesiosaur Eardasaurus powelli gen. et sp. nov. (OUMNH PAL-J.2247) from the Middle Jurassic of Yarnton, Oxfordshire, UK. A, B. Scleral ossicles. C. Hyoid element, orientation unknown.
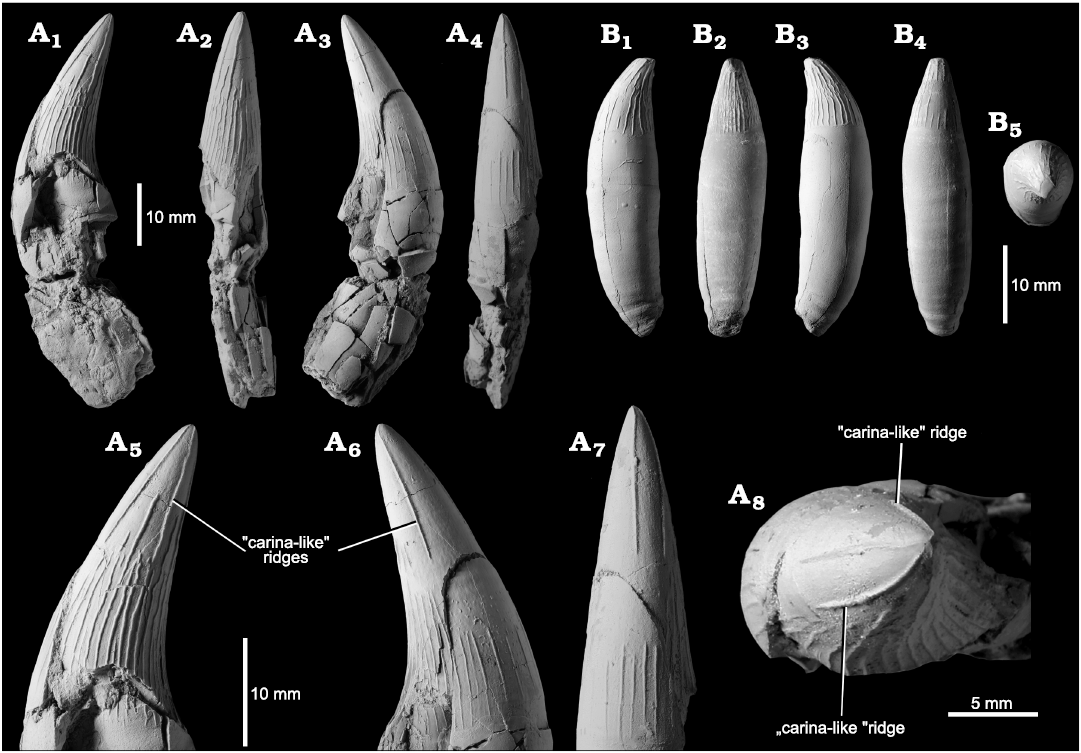
Fig. 13. Holotype of the pliosaurid plesiosaur Eardasaurus powelli gen. et sp. nov. (OUMNH PAL-J.2247) from the Middle Jurassic of Yarnton, Oxfordshire, UK. A. Mesial tooth in anterior or posterior (A1), lingual (A2), anterior or posterior (A3), and labial (A4) views; close-up of crown of tooth showing detail of “carinae” in anterior or posterior (A5–A6), labial (A7), and tip (A8) views. B. Distal (“ratchet” type) tooth in anterior or posterior (B1), lingual (B2), anterior or posterior (B3), labial (B4), and tip (B5) views.
Tooth morphology varies among pliosaurid species and has a long history of use in defining morphotypes (e.g., Madzia and Machalski 2017; Zverkov et al. 2018) and species diagnoses (e.g., Tarlo 1960). The overall tooth morphology and pattern of enamel ridges in the larger (i.e., more mesial) teeth of Eardasaurus powelli gen. et sp. nov. is somewhat similar in overall morphology to those of some other Middle Jurassic pliosaurids, especially those of Peloneustes philarchus, which show some variation but in general are similar to the description given above (Ketchum and Benson 2011b), including the presence of a small number of especially prominent ridges that extend to the tooth apex (e.g., NHMUK PV R1253). The teeth of Eardasaurus powelli gen. et sp. nov. differ from those of Marmornectes candrewi, which shows a set of ridges on either the mesial or distal surface that originate a short distance from the base of the crown (Ketchum and Benson 2011a), and not at the crown base as seen in most other pliosaurids, including Eardasaurus powelli gen. et sp. nov. They differ from those of Liopleurodon ferox and Simolestes vorax, which show a greater proportion of enamel ridges extending to the tooth apex, on all surfaces (e.g., Noè 2001). The teeth of Eardasaurus powelli gen. et sp. nov. differ substantially from those of “Pliosaurus” andrewsi, which bear few enamel ridges, most of which are on the lingual (concave) tooth surface and bear very large wear faces compared to other pliosaurids (NHMUK PV R3891; Andrews 1913; Tarlo 1960). The teeth of Eardasaurus powelli gen. et sp. nov. differ from those of the holotype tooth of “Pliosaurus” grossouvrei (Sauvage, 1873) from the Middle Jurassic (Callovian) of France, in which enamel ridges are almost entirely limited to the concave (lingual) surface of the tooth, and the “carina-like” ridge extends all the way to the tooth base (Sauvage 1873; Foffa et al. 2018).
Smaller, more posterior teeth of Eardasaurus lack the carina-like ridges described above, but nevertheless have well-defined enamel ridges on most surface of the crown, with fewer present on the convex (mesial) surface (Fig. 13B). The apicobasal ridges of these smaller teeth are sinuous, with wavy undulations. Some ridges branch near the base, and some are interrupted by a short stretch of smooth enamel.
Stratigraphic and geographic range.—Type locality and horizon only.
Phylogenetic analysis
We scored Eardasaurus powelli gen. et sp. nov. into the phylogenetic data matrix of Fischer et al. (2018), resulting in a dataset of 108 taxa and 270 characters. We used Bayesian phylogenetic inference in MrBayes 3.2 (Ronquist et al. 2012). The analysis was based on the relaxed (variable rates) Mkv model, run for 60 000 000 generations, with a burn-in faction of 25%. The batch file that executes this analysis, including the data matrix in Nexus format and full specifications of our analysis, are included in the SOM 2.
Results.—The phylogenetic analysis recovered a tree that is broadly consistent with other recent analyses regarding global relationships among plesiosaurians (e.g., Benson and Druckenmiller 2014; Serratos et al. 2017; Fischer et al. 2017, 2018). Eardasaurus powelli gen. et sp. nov. is recovered as an early-diverging thalassophonean pliosaurid, more derived than the non-thalassophonean Marmornectes candrewi, but outside of the derived group comprising “Pliosaurus” andrewsi, Simolestes vorax, Liopleurodon ferox, Pliosaurus spp., and Brachaucheninae (Fig. 14). Eardasaurus powelli gen. et sp. nov. was recovered as the immediate sister taxon to that derived group, as the most derived taxon in a pectinate grade comprising Anguanax zignoi Cau and Fanti, 2015, Peloneustes philarchus, and Eardasaurus powelli gen. et sp. nov.
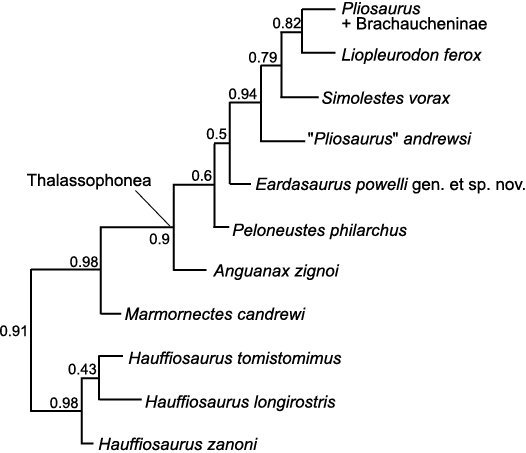
Fig. 14. Phylogenetic topology for Pliosauridae, focussing on Early–Middle Jurassic representatives, based on Bayesian Mkv analysis. Numbers indicate posterior support for nodes demonstrating strong support for many aspects, but weaker support for relationships among Peloneustes philarchus, Anguanax zignoi, and Eardasaurus powelli gen. et sp. nov., which are tentatively recovered as grade leading to more derived pliosaurids.
The following characters indicate that Eardasaurus powelli gen. et sp. nov. is more derived than Hauffiosaurus, Marmornectes candrewi, and Peloneustes philarchus within Pliosauridae:
– In Eardasaurus powelli gen. et sp. nov., the alveolar margin of the upper jaw in lateral view is undulating, forming a “scalloped” margin (character 13, state 1), as in Simolestes vorax, Liopleurodon ferox, and Pliosaurus spp. More basal members of Pliosauridae such as Marmornectes candrewi and Peloneustes philarchus have a weakly convex margin (state 13.0).
– The mandibular symphysis of Eardasaurus powelli gen. et sp. nov. is intermediate in length (character 112, state 1), as in Simolestes vorax, Liopleurodon ferox, “Pliosaurus” andrewsi, and Brachaucheninae. This is unlike more basal members of Pliosauridae, which are relatively longirostrine, such as Peloneustes philarchus, Marmornectes candrewi, and species of Hauffiosaurus (state 112.0).
The cranial characters that indicate that Eardasaurus powelli gen. et sp. nov. is excluded from the group comprising “Pliosaurus” andrewsi, Simolestes vorax, Liopleurodon ferox, Pliosaurus spp., and Brachacheninae, and especially from the group comprising Pliosaurus spp. + Brachacheninae include:
– Eardasaurus powelli gen. et sp. nov. has only a weakly interdigitating premaxilla–maxilla suture (character 24, state 0), as in Peloneustes philarchus, Marmornectes candrewi, and Liopleurodon ferox, but unlike Simolestes vorax, species of Pliosaurus, and the majority of brachauchenines, which have a “zig-zag” shaped suture (state 24.1), that is especially pronounced in Pliosaurus (Benson et al. 2013).
– The posteromedial portion of the maxilla of Eardasaurus powelli gen. et sp. nov. is not subdivided, but forms a simple sheet of bone (character 30, state 0), as in other Middle Jurassic pliosaurids from the Oxford Clay (Peloneustes philarchus, Simolestes vorax, Liopleurodon ferox) but unlike younger Upper Jurassic and Cretaceous pliosaurids including Pliosaurus spp., Acostasaurus pavachoquensis, and Brachauchenius lucasi, in which the maxilla is subdivided by anteroposteriorly oriented fissures (state 30.1).
–The parietal vault of Eardasaurus powelli gen. et sp. nov. in dorsal view is mediolaterally narrow, with the lateral surfaces weakly convex or slightly sinuous (character 49, state 0), as in Peloneustes philarchus, Simolestes vorax, and Liopleurodon ferox. This is unlike Pliosaurus spp. and brachachenines, in which the parietal vault is strongly expanded (state 49.2).
– As in all Middle Jurassic pliosaurids, the ventral surface of the parasphenoid in Eardasaurus powelli gen. et sp. nov. is visible through a V-shaped notch in the posterior pterygoid contact anterior to the posterior interpterygoid vacuities (character 86, state 1). In Cretaceous brachauchenines, the parasphenoid is covered by the pterygoids anterior to the posterior interpterygoid vacuities (state 86.0).
– In Eardasaurus powelli gen. et sp. nov., the midpoint of the posterior interpterygoid vacuities is posterior to the anterior margin of the subtemporal fossa (character 104, state 0), like the more basal species of Hauffiosaurus and Peloneuestes. This is unlike more derived pliosaurids such as Simolestes vorax, Liopleurodon ferox, and Pliosaurus spp., in which the midpoint is approximately level with the anterior margin of the fossa, or more anterior (state 104.1).
– As in Middle–Late Jurassic pliosaurids, the dentary of Eardasaurus powelli gen. et sp. nov. forms a raised ventral platform or keel adjacent to the mandibular symphysis (character 114, state 1). This is unlike the Cretaceous brachauchenines, which do not have a ventral elaboration (state 114.0).
– In Eardasaurus powelli gen. et sp. nov., the retroarticular process is longer than the mandibular glenoid (character 116, state 1) as in the longirostrine pliosaurids ’species of Hauffiosaurus, Peloneustes philarchus, and Marmornectes candrewi and “Pliosaurus” andrewsi. This is unlike Liopleurodon ferox, Pliosaurus spp., and the brachauchenines (other than Acostasaurus pavachoquensis), in which the retroarticular process is shorter or subequal to the anteroposterior length of the glenoid (state 116.0).
Discussion
The teeth of Eardasaurus powelli gen. et sp. nov. bear several prominent apicobasal enamel ridges on the mesial and distal surfaces of the crown, which extend to the tip of the tooth, thus forming distinct cutting edges similar to carinae. These structures have potential ecological importance, indicating enhanced cutting function of the teeth and have often been taken as evidence for a macropredatory diet (Massare 1987). Carinae, or “carina-like” enamel ridges are present in many other groups of marine reptiles, including in trihedral-toothed pliosaurids such as species of Pliosaurus and some brachauchenines (Fischer et al. 2015; Zverkov 2015). Potentially “carina-like” ridges are also present in an isolated pliosaurid tooth from the Cenomanian of Poland (Madzia and Machalski 2017: text-fig. 4). They are also present in the conical-toothed pliosaurid Liopleurodon ferox, in which the teeth have “carinate tips” formed by a few of the coarse apicobasal enamel ridges reaching the apex of the crown (Massare 1987). Nevertheless, so far, carina-like ridges have rarely been reported in plesiosaurians. The carina-like ridges of Liopleurodon ferox are ornamented with fine, transverse ribs, which form serrated cutting edge (Massare 1987), as also occurs in a more prominent form in the trihedral-toothed Cretaceous pliosaurid Makhaira rossica Fischer, Arkhangelsky, Stenshin, Uspensky, Zverkov, and Benson, 2015 (Fischer et al. 2015: fig. 3f–h).
The carina-like apicobasal ridges of Eardasaurus gen. nov. are relatively much finer than those of Liopleurodon and Makhaira and bear only a very weakly-defined serration-like texture. However, they result in a distinctive overall dental morphology with a clear distinction between the carina-like ridges and the finer (“normal”) enamel ridges because these finer enamel ridges extend only to approximately one-third or mid-height of the crown on most surfaces (Fig. 13A). During the present work, we reviewed the distribution of these structures in other Middle Jurassic pliosaurid teeth from the UK. We note that they are absent in some taxa (Simolestes vorax, Noè 2001; Marmornectes candrewi, Ketchum and Benson 2011a); but may be present and previously unreported in others (e.g., Peloneustes philarchus, NHMUK PV R1253; “Pliosaurus” andrewsi, NHMUK PV R3891, although the tips of the crowns are frequently worn in “P.” andrewsi). These observations suggest that carina-like dental morphologies may be more widespread in pliosaurids, and perhaps in plesiosaurians, than had previously been recognised.
Conclusions
Eardasaurus powelli gen. et sp. nov. is a new pliosaurid from the Oxford Clay Formation of Oxfordshire, UK, identified by several autapomorphies and a unique combination of character states. Eardasaurus gen. nov. brings the total number of pliosaurid genera known from the Oxford Clay Formation up to a minimum of six (or seven, including Pachycostasaurus dawni), contributing to our understanding of the most taxonomically rich assemblage of pliosaurids known. In our view, it is likely that this high pliosaurid diversity results from intensive sampling over more than a century and may be representative for marine communities of the late Middle Jurassic, which remain more poorly sampled elsewhere. The teeth of Eardasaurus powelli gen. et sp. nov. have distinct “carina-like” apicobasal enamel ridges, similar to those of the much larger pliosaurid Liopleurodon ferox. We find that carina-like ridges are more widespread among pliosaurids than has previously been recognised and that their distribution among plesiosaurians requires further investigation.
Acknowledgements
We are grateful to William J. Kennedy (OUMNH) for his assistance applying ammonium chloride and photographing the teeth in Fig. 12C. We thank Eliza Howlett, Juliet Hay, Pete Johnson, and Adam Fisk (all OUMNH); Sandra Chapman, Lil Stevens, and Mike Day (all NHMUK); and Matthew Riley (Sedgwick Museum of Earth Sciences, University of Cambridge, UK) for access to specimens, and H. Philip Powell (OUMNH) for discussion. JulietHay and H. Philip Powell carried out preparation work on the skull and postcranial skeleton following the excavation in 1994, and Stephanie Wright made 3D surface scans of part of the postcranial skeleton. We thank the DCMS/Wolfson Museums and Galleries Improvement Fund, which enabled the preparation of the skull to be completed by Scott Moore-Fay (Wavecut Platforms Ltd., Sundridge, UK) in 2018. We thank Nikolay Zverkov ((Russian Academy of Sciences, Moscow, Russia) and one anonymous reviewer for their comments, which greatly improved the manuscript.
References
Albright, L.B., III, Gillette, D.D., and Titus, A.L. 2007a. Plesiosaurs from the Upper Cretaceous (Cenomanian–Turonian) Tropic Shale of southern Utah, part 1: new records of the pliosaur Brachauchenius lucasi. Journal of Vertebrate Paleontology 27: 31–40. Crossref
Albright, L.B., III, Gillette, D.D., and Titus, A.L. 2007b. Plesiosaurs from the Upper Cretaceous (Cenomanian–Turonian) Tropic Shale of southern Utah, part 2: Polycotylidae. Journal of Vertebrate Paleontology 27: 41–58. Crossref
Andrews, C.W. 1909. On some new Plesiosauria from the Oxford Clay of Peterborough. Annals and Magazine of Natural History 4 (23): 418–429. Crossref
Andrews, C.W. 1910. A Descriptive Catalogue of the Marine Reptiles of the Oxford Clay. Part 1. xvii + 202 pp. British Museum, London.
Andrews, C.W. 1913. A Descriptive Catalogue of the Marine Reptiles of the Oxford Clay. Part 2. xxiv + 206 pp. British Museum, London.
Anon. 1917. Obituary. Alfred Nicholson Leeds, F.G.S. Geological Magazine 4: 478–480. Crossref
Arkhangelsky, M.S., Zverkov, N.G., Rogov, M.A., Stenshin, I.M., and Baykina, E.M. 2019. Colymbosaurines from the Upper Jurassic of European Russia and their implications for palaeobiogeography of marine reptiles. Palaeobiodiversity and Palaeoenvironments 100: 197–218. Crossref
Benson, R.B.J. and Druckenmiller, P.S. 2014. Faunal turnover of marine tetrapods during the Jurassic–Cretaceous transition. Biological Reviews 89: 1–23. Crossref
Benson, R.B.J., Evans, M., Smith, A.S., Sassoon, J., Moore-Fay, S., Ketchum, H.F., and Forrest, R. 2013. A giant pliosaurid skull from the Late Jurassic of England. PLoS ONE 8(5): e65989. Crossref
Benson, R.B.J., Ketchum, H.F., Noè, L.F., and Gómez-Pérez, M. 2011. New information on Hauffiosaurus (Reptilia, Plesiosauria) based on a new species from the Alum Shale Member (Lower Toarcian: Lower Jurassic) of Yorkshire, UK. Palaeontology 54: 547–571. Crossref
Bloodworth, A.J., Cowley, J.F., Highley, D.E., and Bowler, G.K. 2001. Brick Clay: Issues for planning. 44 + 81 pp. HSMO for the British Geological Survey, London.
Brown, D.S. 1981. The English Upper Jurassic Plesiosauroidea (Reptilia) and a review of the phylogeny and classification of the Plesiosauria. Bulletin of the British Museum (Natural History), Geology 35: 253–344.
Buchy, M.C., Frey, E., Stinnesbeck, W., and López-Olivia, J. 2003. First occurrence of a gigantic pliosaurid plesiosaur in the late Jurassic (Kimmeridgian) of Mexico. Bulletin de la Société Géologique de France 174: 271–278. Crossref
Callomon, J.H. 1968. The Kellaways Beds and the Oxford Clay. In: P.C. Sylvester-Bradley and T.D. For (eds.), The Geology of the East Midlands, 264–290. Leicester University Press, Leicester.
Carpenter, K. 1996. A review of short-necked plesiosaurs from the Cretaceous of the Western Interior, North America. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 201: 259–287. Crossref
Cau, A. and Fanti, F. 2015. High evolutionary rates and the origin of the Rosso Ammonitico Veronese Formation (Middle–Upper Jurassic of Italy) reptiles. Historical Biology 28: 952–962. Crossref
Cruickshank, A.R.I., Martill, D.M., and Noè, L.F. 1996. A pliosaur (Reptilia, Sauropterygia) exhibiting pachyostosis from the Middle Jurassic of England. Journal of the Geological Society of London 153: 873–879. Crossref
de Blainville, H.D. 1835. Description de quelques espèces de reptiles de la Californie, précédé de l’analyse d’un système general d’Erpétologie et d’Amphibiologie. Nouvelles Annales du Museum National d’Histoire Naturelle, Paris 4: 233–296.
Duff, K.L. 1980. Callovian correlation chart. In: J.C.W. Cope, K.L. Duff, C.F. Parsons, H.S. Torrens, W.A. Wimbledon, and J.K. Wright (eds.), A Correlation of Jurassic Rocks in the British Isles, Part Two: Middle and Upper Jurassic, 45–60. Geological Society of London Special Report, London.
Fischer, V., Arkhangelsky, M.S., Stenshin, I.M., Uspensky, G.N., Zverkov, N.G., and Benson, R.B.J. 2015. Peculiar macrophageous adaptations in a new Cretaceous pliosaurid. Royal Society Open Science 2: 150552. Crossref
Fischer, V., Benson, R.B.J., Druckenmiller, P.S., Ketchum, H.F., and Bardet, N. 2018. The evolutionary history of polycotylid plesiosaurians. Royal Society Open Science 5: 172177. Crossref
Fischer, V., Benson, R.B.J., Zverkov, N.G., Soul, L.C., Arkhangelsky, M.S., Lambert, O., Stenshin, I.M., Uspensky, G.N., and Druckenmiller, P.S. 2017. Plasticity and convergence in the evolution of short-necked plesiosaurs. Current Biology 27: 1667–1676.e3. Crossref
Foffa, D., Young, M., and Brusatte, S. 2018. Filling the Corallian gap: new information on Late Jurassic marine reptile faunas from England. Acta Palaeontologica Polonica 63: 287–313. Crossref
Gasparini, Z. 2009. A new Oxfordian pliosaurid (Plesiosauria, Pliosauridae) in the Caribbean Seaway. Palaeontology 52: 661–669. Crossref
Gasparini, Z. and O’Gorman, J.P. 2014. A new species of Pliosaurus (Sauropterygia, Plesiosauria) from the Upper Jurassic of Northwestern Patagonia, Argentina. Ameghiniana 51: 269–283. Crossref
Gómez-Pérez, M. and Noè, L.F. 2017. Cranial anatomy of a new pliosaurid Acostasaurus pavachoquensis from the Lower Cretaceous of Colombia, South America. Palaeontographica A 310: 5–52. Crossref
Gradstein, F.M., Ogg, J.G., Schmitz M.D and Ogg G.M. 2020. Geologic Time Scale 2020. 1357 pp. Elsevier, Amsterdam. Crossref
Holland, T. 2018. The mandible of Kronosaurus queenslandicus Longman, 1924 (Pliosauridae, Brachaucheniinae), from the Lower Cretaceous of northwest Queensland, Australia. Journal of Vertebrate Paleontology 38: e1511569. Crossref
Hollingworth, N.T.J. and Wignall, P.B. 1992. The Callovian–Oxfordian boundary in Oxfordshire and Wiltshire based on two new temporary sections. Proceedings of the Geologists’ Association 103: 15–30. Crossref
Hudson, J.D. and Martill, D. 1994. The Peterborough Member (Callovian, Middle Jurassic) of the Oxford Clay Formation at Peterborough, UK. Journal of the Geological Society of London 151: 133–124. Crossref
Ketchum, H.F. and Benson, R.B.J. 2010. Global interrelationships of Plesiosauria (Reptilia, Sauropterygia) and the pivotal role of taxon sampling in determining the outcome of phylogenetic analyses. Biological Reviews 85: 361–392. Crossref
Ketchum, H.F. and Benson, R.B.J. 2011a. A new pliosaurid (Sauropterygia, Plesiosauria) from the Oxford Clay Formation (Middle Jurassic, Callovian) of England: evidence for a gracile, longirostrine grade of Early–Middle Jurassic pliosaurids. Special Papers in Palaeontology 86: 109–129.
Ketchum, H.F. and Benson, R.B.J. 2011b. The cranial anatomy and taxonomy of Peloneustes philarchus (Sauropterygia, Pliosauridae) from the Peterborough Member (Callovian, Middle Jurassic) of the United Kingdom. Palaeontology 54: 639–665. Crossref
Knutsen, E.M. 2012. A taxonomic revision of the genus Pliosaurus (Owen, 1841a) Owen, 1841b. Norwegian Journal of Geology 92: 259–276.
Knutsen, E.M., Druckenmiller, P.S., and Hurum, J.H. 2012. A new species of Pliosaurus (Sauropterygia: Plesiosauria) from the middle Volgian of central Spitsbergen, Norway. Norwegian Journal of Geology 92: 235–258.
Longman, H.A. 1924. A new gigantic marine reptile from the Queensland Cretaceous. Memoirs of the Queensland Museum 8: 26–28.
Madzia, D. and Machalski, M. 2017. Isolated pliosaurid teeth from the Albian–Cenomanian (Cretaceous) of Annopol, Poland. Acta Geologica Polonica 67: 393–403. Crossref
Maisch, M.W. 1998. Notes on the cranial osteology of Muraenosaurus Seeley, 1874 (Sauropterygia, Jurassic), with special reference to the neurocranium and its implications for sauropterygian phylogeny. Neues Jahrbuch für Geologie und Paläontologie. Abhandlungen 207: 207–253. Crossref
Massare, J.A. 1987. Tooth morphology and prey preference of Mesozoic marine reptiles. Journal of Vertebrate Paleontology 7: 121–137. Crossref
McCurry, M.R., Evans, A.R., Fitzgerald, E.M.G., McHenry, C.R., Bevitt, J., and Pyenson, N.D. 2019. The repeated evolution of dental apicobasal ridges in aquatic-feeding mammals and reptiles. Biological Journal of the Linnaean Society 127: 245–259. Crossref
Noè, L.F. 2001. A Taxonomic and Functional Study of the Callovian (Middle Jurassic) Pliosauroidea (Reptilia; Sauropterygia). Volume 1: xxxii + 347 pp, Volume 2: xix + 182 pp. Ph.D. Thesis, School of Environmental and Applied Sciences, University of Derby, Derby.
Noè, L.F., Smith, D.T.J., and Walton, D.K. 2004. A new species of Kimmeridgian pliosaur (Reptilia; Sauropterygia) and its bearing on the nomenclature of Liopleurodon macromerus. Proceedings of the Geologists’ Association 115: 13–24. Crossref
O’Gorman, J.P., Gasparini, Z., and Spalletti, L.A. 2018. A new Pliosaurus species (Sauropterygia. Plesiosauria) from the Upper Jurassic of Patagonia: new insights on the Tithonian morphological disparity of mandibular symphyseal morphology. Journal of Paleontology 92: 240–253. Crossref
O’Keefe, F.R. 2001. A cladistic analysis and taxonomic revision of the Plesiosauria (Reptilia: Sauropterygia). Acta Zoologica Fennica 213: 1–63.
O’Keefe, F.R. and Carrano, M.T. 2005. Correlated trends in the evolution of the plesiosaur locomotor system. Paleobiology 31: 656–675. Crossref
O’Keefe, F.R. and Wahl, W., jr 2003. Current taxonomic status of the plesiosaur Pantosaurus striatus from the Upper Jurassic Sundance Formation, Wyoming. Paludicola 4: 37–47.
Owen, R. 1860. On the orders of fossil and recent Reptilia, and their distribution in time. Report of the British Association for the Advancement of Science for 1859 29: 153–166.
Páramo-Fonseca, M.E., Benavides-Cabra, C.D., and Gutiérrez, I.E. 2018. A new large pliosaurid from the Barremian (Lower Cretaceous) of Sáchica, Boyacá, Colombia. Earth Sciences Research Journal 22: 223–238. Crossref
Páramo-Fonseca, M.E., Benavides-Cabra, C., and Gutiérrez, I.E. 2019. A new specimen of Stenorhynchosaurus munozi Páramo-Fonseca et al., 2016 (Plesiosauria, Pliosauridae) from the Barremian of Colombia: new morphological features and ontogenetic implications. Journal of Vertebrate Paleontology 39: e1663426. Crossref
Páramo-Fonseca, M.E, Gómez-Pérez, M., Noè, L.F., and Etayo-Serna, F. 2016. Stenorhynchosaurus munozi, gen. et sp. nov. a new pliosaurid from the Upper Barremian (Lower Cretaceous) of Villa de Leiva, Colombia, South America. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 40: 84–103. Crossref
Romer, A.S. and Lewis, A.D. 1959. A mounted skeleton of the giant plesiosaur Kronosaurus. Breviora 112: 1–15.
Ronquist, F., Telsenko, M., Van der Mark, P., Ayres, D.L., Darling, A., Höhna, S., Largey, B., Liu, L., Suchard, M.A., and Huelsenbeck, J.P. 2012. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biology 61: 539–542. Crossref
Sauvage, H.E. 1873. Notes sur les reptiles fossiles. Bulletin de la Société Géologique de France, Series 3 1: 366–386.
Schumacher, B.A. 2011. A ‘woollgari-zone mosasaur’ (Squamata; Mosasauridae) from the Carlile Shale (Lower Middle Turonian) of central Kansas and the stratigraphic overlap of early mosasaurs and pliosaurid plesiosaurs. Transactions of the Kansas Academy of Science 114: 1–14. Crossref
Schumacher, B.A., Carpenter, K., and Everhart, MJ. 2013. A new Cretaceous pliosaurid (Reptilia, Plesiosauria) from the Carlile Shale (middle Turonian) of Russell County, Kansas. Journal of Vertebrate Paleontology 33: 613–628. Crossref
Schumacher, B.A. and Martin, J. 2015. Polycotylus latippinis Cope (Plesiosauria, Polycotylidae), a nearly complete skeleton from the Niobrara Formation (early Campanian) of southwestern South Dakota. Journal of Vertebrate Paleontology 36: e1031341. Crossref
Seeley, H.G. 1869. Index to the fossil remains of Aves, Ornithosauria, and Reptilia, from the Secondary System of Strata arranged in the Woodwardian Museum of the University of Cambridge. xxiii + 143 pp. Deighton, Bell and Co., London.
Seeley, H.G. 1874a. Note on some of the generic modifications of the plesiosaurian pectoral arch. Quarterly Journal of the Geological Society of London 30: 436–449. Crossref
Seeley, H.G. 1874b. On Muraenosaurus leedsii, a plesiosaurian from the Oxford Clay. Part 1. Quarterly Journal of the Geological Society of London 30: 197–208. Crossref
Serratos, D.J., Druckenmiller, P., and Benson, R.B.J. 2017. A new elasmosaurid (Sauropterygia, Plesiosauria) from the Bearpaw Shale (Late Cretaceous, Maastrichtian) of Montana demonstrates multiple evolutionary reductions of neck length within Elasmosauridae. Journal of Vertebrate Paleontology 37: e1278608. Crossref
Smith, A.S. and Benson, R.B.J. 2014. Osteology of Rhomaleosaurus thorntoni (Sauropterygia: Rhomaleosauridae) from the Lower Jurassic (Toarcian) of Northamptonshire, England. Monographs of the Palaeontographical Society 168: 1–40. Crossref
Smith, A.S. and Dyke, G.J. 2008. The skull of the giant predatory pliosaur Rhomaleosaurus cramptoni: implications for plesiosaur phylogenetics. Naturwissenschaften 95: 975–980. Crossref
Soul, L.C. and Benson, R.B.J. 2017. Developmental mechanisms of macroevolutionary change in the tetrapod axis: a case study of Sauropterygia. Evolution 71: 1164–1177. Crossref
Tarlo, L.B. 1960. A review of the Upper Jurassic pliosaurs. Bulletin of the British Museum (Natural History), Geology Series 4: 147–189.
Taylor, M.A. and Cruickshank, A.R.I. 1993. Cranial anatomy and functional morphology of Pliosaurus brachyspondylus (Reptilia: Plesiosauria) from the Upper Jurassic of Westbury, Wiltshire. Philosophical Transactions of the Royal Society of London B 341: 399–418. Crossref
Vincent, P. 2011. A re-examination of Hauffiosaurus zanoni, a pliosauroid from the Toarcian (Early Jurassic) of Germany. Journal of Vertebrate Paleontology 31: 340–351. Crossref
Williston, S.W. 1903. North American plesiosaurs. Part I. Field Columbian Museum Publication 73, Geological Series 2: 1–79. Crossref
Williston, S.W. 1907. The skull of Brachauchenius, with observations on the relationships of the plesiosaurs. Proceedings of the National Museum 32: 477–493. Crossref
Zverkov, N.G. 2015. On a typically Late Jurassic pliosaur from the Lower Cretaceous of Crimea. In: E.Y. Baraboshkin and D.E. Bykov (eds.), The International Scientific Conference on the Jurassic/Cretaceous Boundary, September 7–13, Samara (Russia), Proceedings Volume, 89–94. Kassandra, Tolyatti.
Zverkov, N.G., Fischer, V., Madzia, D., and Benson, R.B.J. 2018. Increased pliosaurid dental disparity across the Jurassic–Cretaceous transition. Palaeontology 61: 825–846. Crossref
Zverkov, N.G., Shmakov, A.S., and Arkhangelsky, M.S. 2017. Jurassic marine reptiles of Moscow and surroundings. In: M.A. Rogov and V.A. Zakharov, (eds.), Jurassic Deposits of the Southern Part of the Moscow Syneclyse and their Fauna, 230–263. GEOS, Moscow.
Acta Palaeontol. Pol. 67 (2): 297–315, 2022
https://doi.org/10.4202/app.00887.2021