
Ungulates of the middle Miocene Monarch Mill Formation, Churchill County, Nevada, USA
KENT S. SMITH, NICHOLAS J. CZAPLEWSKI, and MARGERY C. COOMBS
Smith, K.S., Czaplewski, N.J., and Coombs, M.C. 2022. Ungulates of the middle Miocene Monarch Mill Formation, Churchill County, Nevada, USA. Acta Palaeontologica Polonica 67 (1): 239–256.
A middle Miocene, early Barstovian land mammal age vertebrate assemblage, the Eastgate local fauna (LF), is known in the basal-most part of the Monarch Mill Formation. This rich assemblage of fossil vertebrates occurs within the Middlegate Basin in Churchill County, Nevada, USA. The Eastgate LF includes the fossil remains of fish, amphibians, reptiles, birds, and over 25 families of mammals. Previous studies on the mammalian remains have revealed several endemic taxa of rodents and carnivores. Herein, we describe the ungulates, which include two taxa from two families of Artiodactyla (Ticholeptus sp., Merycoidodontidae and Barbouromeryx trigonocorneus, Palaeomerycidae) and three taxa from three families of Perissodactyla (Equidae gen. et sp. indet., Moropus merriami, Chalicotheriidae, and Teleoceras sp., Rhinocerotidae). Independent paleobotanical evidence nearly contemporaneous with the vertebrates indicates forest and shrubland paleovegetation, and suggests that the area had been uplifted to 2700–2800 m paleoaltitude. Therefore, this local fauna adds a rare glimpse of a medium-to high-altitude vertebrate community in the intermountain western interior of North America. Temporally-restricted taxa (especially the rodent Tardontia nevadans and chalicothere Moropus merriami) reinforce the early Barstovian age of the Eastgate LF and are consistent with tephrochronological dates and radiometric analyses. The presence of Barbouromeryx trigonocorneus at Eastgate is significant in that its occurrence potentially represents a temporal range extension into the early Barstovian (previously known from latest Arikareean to middle Hemingfordian), and extends the paleobiogeographic range from the Great Plains to the Great Basin. Unequivocally, the ungulates and other mammals of the Eastgate LF support the presence of a temperate forest ecosystem in the Great Basin just subsequent to the Miocene Climatic Optimum.
Key words: Mammalia, Ungulata, paleoecology, palaeoclimate, Barstovian, Miocene, Middlegate Basin, Nevada.
Kent S. Smith [kent.smith@okstate.edu], Office of American Indians in Medicine and Science and Department of Anatomy and Cell Biology, Oklahoma State University Center for Health Sciences, 1111 W 17th St., Tulsa, OK 74107 USA.
Nicholas J. Czaplewski [nczaplewski@ou.edu], Sam Noble Museum, 2401 Chautauqua Ave., Norman, OK 73072 USA.
Margery C. Coombs [mccc@bio.umass.edu], Department of Biology, University of Massachusetts, Amherst, MA 01003 USA.
Received 15 May 2021 accepted 22 November 2021, available online 30 March 2022.
Copyright © 2022 K.S. Smith et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Monarch Mill Formation, that has yielded Miocene vertebrate fossils, is the stratigraphically higher of two sedimentary rock units within a small depositional basin (Middlegate Basin) located in Churchill County, Nevada, USA. (Fig. 1). The second and stratigraphically lower sedimentary rock unit in the basin is the Middlegate Formation, which has produced Miocene plant fossils. Axelrod (1956, 1985) described these two sedimentary rock units and one igneous rock unit (Clan Alpine volcanics) while studying fossil plants of the region. Fossil plant remains from the Middlegate Basin have only been recovered from the upper-most part of the Middlegate Formation. In contrast, Axelrod (1956, 1985) reported on four separate fossil mammal discoveries for the Monarch Mill Formation as follows: (i) About 30 m stratigraphically above and east of Axelrod’s (1956) principal plant locality (UCMP PA 5101). At this site, a tooth, jaw, and cervical vertebra of a rhinoceros were collected and sent to Robert W. Wilson, then at South Dakota School of Mines and Technology, who identified the remains as Aphelops (ii) South of U.S. Highway 50, southeast of the town of Middlegate, Nevada, remains were identified for Axelrod (1956) by Donald E. Savage, University of California Museum of Paleontology (UCMP) as belonging to a gomphotheriid (Proboscidea) and a camel. There is no UCMP locality number for these specimens and they were apparently not preserved in a collection. (iii) Locality UCMP V5200 was discovered farther south and east of the second locality in the basal-most part of the upper member of the Monarch Mill Formation. Axelrod (1956) referred to the faunal assemblage from UCMP V5200 as the Monarch Mill fauna. Richard H. Tedford, American Museum of Natural History, identified the mammal material recovered from this locality as species of machairodont (Felidae), Teleoceras (Rhinocerotidae), cf. Paracamelus and cf. Pliauchenia or Procamelus (Camelidae), and Gomphotheriidae, suggesting a Hemphillian land mammal age (Axelrod 1956). (iv) In 1967, another fossil mammal locality was discovered and reported by Axelrod (1985) in his description of a nearby fossil plant locality that yielded the Eastgate flora. This locality is about 0.8 km southwest of the site from which the Eastgate flora was collected, and yielded the first specimens of the Eastgate LF, considered to be early Barstovian in age. The site was called “Nevaxel” by Savage and Russell (1983: 252). Subsequent prospecting for vertebrate fossils near this locality by UCMP and Oklahoma Museum of Natural History (OMNH) field parties revealed additional localities and fossils in the vicinity (Smith 2002; Smith et al. 2006, 2016). To date, the Eastgate LF has revealed fossil remains of fish, amphibians, reptiles, birds, and mammals (25 families) that have included one new genus and species of rodent (Smith et al. 2006) and two new genera and three new species of carnivorans (Smith et al. 2016).

Fig. 1. Location of the study area within USA (A) and state of Nevada, Churchill County (B), Eastgate study area indicated as black rectangle. Modified from Smith et al. 2016.
Based on fossil mammals, the age of the Monarch Mill Formation spans from the early Barstovian to the Hemphillian North American Land Mammal Ages (NALMAs: see Woodburne 2004), thus potentially representing as much as 11 million years (~16–5 Ma) of geologic history in the Great Basin. The Monarch Mill Formation unconformably overlies the Middlegate Formation, which has revealed two fossil plant localities yielding the Middlegate flora, UCMP PA 5101 (Axelrod 1956) and the Eastgate flora, UCMP PA 533 (Axelrod 1985). These fossil floras occur in the upper member of the Middlegate Formation, but differ markedly in their composition (see below). Currently, no fossil plants have been reported from the Monarch Mill Formation; however, more than 20 vertebrate localities representing two separate vertebrate assemblages have been recovered from this formation: (i) Eastgate LF (early Barstovian) and (ii) Monarch Mill fauna (Hemphillian). The Eastgate LF occurs in the basal-most unit of the Monarch Mill Formation, whereas the Monarch Mill fauna was discovered in the upper-most part of the Monarch Mill Formation. Herein we provide a succinct review of the geological and paleoecological context of the Middlegate Basin, for which detailed study is merited. The main focus of the present study, however, is the ungulates of the Eastgate LF, with emphasis on their systematics.
Institutional abbreviations.—AMNH F:AM, Frick American Mammals, American Museum of Natural History, New York, USA; OMNH, Oklahoma Museum of Natural History, Norman, Oklahoma, USA; SCMNH, Sierra College Museum of Natural History, Rocklin, USA; UCMP, University of California Museum of Paleontology, Berkeley, California, USA; UO, Museum of Natural and Cultural History, University of Oregon, Eugene, Oregon, USA; USNM, National Museum of Natural History, Washington, D.C., USA. OMNH and UCMP numbers preceded by a “V” refer to fossil vertebrate localities; UCMP numbers preceded by a “PA” refer to fossil plant localities.
Other abbreviations.—AP, greatest anteroposterior length of tooth; LF, local fauna; NALMA, North American Land Mammal Age; T, greatest transverse width of tooth.
Geological setting
Geological setting of the Middlegate Basin.—The Middlegate Basin occurs between the southern end of the Clan Alpine Mountains to the west and a spur of the Desatoya Mountains to the east. The former mountains run along a north-south axis, with their south end near the low pass and town known as Middlegate, Nevada. The Eastgate Wash drains the Middlegate Basin and exits through Middlegate Pass. Axelrod (1956) first described and mapped the eastward-dipping exposures in the middle and western parts of the Middlegate Basin where he designated the rocks cropping out from the southeastern side of the Clan Alpine Mountains east toward the town of Eastgate as the Middlegate Formation, with a maximum thickness of 396 m.
Axelrod (1956) examined the geology of the area, in part to determine the stratigraphic context of the Middlegate flora within the Middlegate Formation. The Middlegate Formation consists mainly of sedimentary rocks that rest unconformably on the Clan Alpine volcanics. For purposes of mapping, Axelrod (1956) divided the Middlegate Formation into three informal units as follows: (i) lower member ranges from ~1–91 m thick and consists of sedimentary breccia, conglomerate, and sandstone beds (derived from the underlying volcanics); (ii) the middle member (including the Middlegate flora) reaches a maximum thickness of 122 m and consists of a siliceous shale with alternating soft tuffaceous claystone; and (iii) the upper member includes sandstone and rhyolitic tuff, interbedded with claystone and thin shale beds of lacustrine origin.
The Monarch Mill Formation was named by Axelrod (1956) after a nearby stamp mill in the Middlegate Basin. His original description was based on exposures in the middle and western parts of the Middlegate Basin. The Monarch Mill Formation unconformably overlies the Middlegate Formation and is about 700 m thick in the central part of the Middlegate Basin, thinning at the margins (Stewart et al. 1999). For the Monarch Mill Formation, Axelrod (1956) mapped six sedimentary units from the lower to upper-most parts of the stratigraphic section and described their dominant lithologies as follows: (i) tuffaceous claystone and conglomerate; (ii) sandstone; (iii) rhyolitic tuff and pumice; (iv) diatomite and tuffaceous siltstone; (v) volcanic pebble conglomerate; and (vi) rhyolitic claystone, pumice, sandstone, and siltstone.
The Monarch Mill Formation was traced into the eastern and southeastern parts of the Middlegate Basin by Axelrod (1985), where he reported coarser clastic sediments than those from the middle and western parts. He placed the base of the Monarch Mill Formation at the bottom of a widespread sedimentary breccia that grades basinward into a conglomerate 4–7 m thick, containing cobble- to small boulder-size particles. He noted that the conglomerate scattered throughout the east and southeast region of the Middlegate Basin was composed of coarse, pebbly sandstone that is poorly indurated and poorly bedded; between this conglomerate and slopes covered with rubble and vegetation were mudstone, shale, and tuffs. Axelrod (1985) placed the mammalian locality (Eastgate LF = Nevaxel) in the basal breccia/conglomerate of the Monarch Mill Formation.
Additional studies of the geology in the Middlegate Basin area were conducted by Barrows (1971), who examined the geology of the Desatoya Mountains, and Noble (1972), who described the Middlegate area as representing a boundary for middle Miocene volcanism. Stewart et al. (1999: 12) characterized the sedimentary rocks of the Middlegate and Monarch Mill formations as, “lacustrine, deltaic, fluvial, and landslide deposits,” and noted that, “the Monarch Mill Formation is primarily a fluvial unit, although lacustrine strata are probably also present … Ash-layers in the Middlegate and Monarch Mill formations indicate air-fall from distant sources, mostly the Snake River Plain.”
There are 18 vertebrate-bearing localities yielding fossil mammals occurring within a limited area (1.61 km2) near Eastgate separated by topographic features and including structural deformations that challenge direct stratigraphic correlation. This correlation difficulty was noted by stratigraphers (Darrin Pagnac and Ian Browne, personal communication 2014) and a geochronologist (Alan Deino, personal communication 2014) who joined the lead author (KSS) to collect data on detailed lithologic descriptions, stratigraphic mapping, and sample basalts for radiometric dates of the Monarch Mill and Middlegate formations. These efforts improved understanding of the local stratigraphic relationships among selected Eastgate LF localities within the basal-most part of the Monarch Mill Formation and provided two preliminary radiometric dates at select stratigraphic levels within the upper-most part of the Middlegate Formation and lower-most part of the Monarch Mill Formation (Fig. 2). However, more detailed work needs to be done to provide comprehensive data on the lithologies, stratigraphic mapping, and radiometric dates that will bridge the fossil plant locality in the upper-most part of the Middlegate Formation and the fossil vertebrate localities in the lower and upper parts of the Monarch Mill Formation. Additional data would help answer questions regarding faunal composition, species diversity, and evolutionary changes over time within a local geographic area, lend a better understanding of the local biochronology, and better constrain the NALMA of the Eastgate LF.

Fig. 2. Basic stratigraphic column of the Monarch Mill Formation showing the dominant lithologies (modified from Axelrod 1956). Stratigraphic positions of the Eastgate LF localities is indicated with the “bone” symbol. Adjacent plot of the stratigraphic positions of select localities plotted above the base of the Monarch Mill Formation relative to the “clay change” that seems to delineate the boundary between the Middlegate and Monarch Mill formations (indicated by wavy lines at the base of the column; Darrin Pagnac, personal communication 2014). Basalts for preliminary radiometric dates reported were collected between UCMP V70147 and UCMP V67245.
Age of the Middlegate and Monarch Mill formations.—The absolute age for the Middlegate and Monarch Mill formations has been pursued by several researchers including Swisher (1992: table 5.3.2), who provided 40Ar/39Ar ages of about 15.5 Ma on biotite, 15.95 Ma on plagioclase, and 15.86 Ma on anorthosite from a Middlegate Tuff (sample no. 86CS-MG1). Unfortunately, he did not provide detailed stratigraphic or geographic data associated with these radiometric dates. The Middlegate Formation unconformably underlies the Monarch Mill Formation; thus, these dates provide a maximum possible absolute age for localities yielding the Eastgate LF. Stewart et al. (1999) sampled several units within the Monarch Mill Formation for the chemical composition of volcanic glass shards in the tephra layers of these units. Based on their tephrochronology, an overall age range of 14.7 Ma to 9.8 Ma was indicated for the Monarch Mill Formation. Stewart et al. (1999) suggested that the correlations on which the age range is based are problematic. One sample (1-37-26J), which closely matched a sample taken from tephras of the Virgin Valley Formation located nearby in northwestern Nevada, had an extrapolation age of 15.54–15.41 Ma, which is inconsistent with the high stratigraphic position of this sample within the section. Therefore, Stewart et al. (1999) estimated an age of about 10 Ma for this sample. The preliminary radiometric dates from basalts collected in 2014 from the lower-most part of the Monarch Mill Formation and the upper-most part of the Middlegate Formation ranged from 14.84 to 14.65 Ma. These data were provided for the present study with the caveat that radiometric dates on basalt deposits are difficult to produce with accuracy and precision and additional analyses will be forthcoming (Al Deino, personal communication 2014).
Paleobotany and paleoenvironment of the Middlegate Basin.—Axelrod (1956) described the Middlegate and Eastgate paleofloras based on leaf assemblages from the middle Miocene of the Middlegate Basin. These leaf assemblages were collected from the Middlegate Formation and thus are somewhat older (~16–15.5 Ma as indicated above) than the Eastgate LF of the overlying Monarch Mill Formation. Axelrod (1956, 1985) and Graham (1999) analyzed the Middlegate flora and inferred a chaparral-shrubland that was dominated by sclerophyllous shrubs and trees including Arbutus prexalapensis, Cedrela trainii, Cercocarpus antiquus, C. pacifica, Lithocarpus nevadensis, Quercus hannibali, Q. shrevoides, and Q. simulata. These authors interpreted this plant assemblage as reflecting the vegetation of relatively dry south-facing slopes. The younger Eastgate flora, by contrast, indicates mixed coniferous and hardwood forest, probably from relatively mesic north-facing canyons. It includes Abies concoloroides, Larix cassiana, L. nevadensis, Amelanchier grayi, and Aesculus preglabra. These two paleofloras provided the first example of topographic-climate control of a Miocene forest in a local region in western North America (Axelrod 1985).
The paleoaltitude and other environmental parameters for the Eastgate and Middlegate floras were determined by Wolfe et al. (1997), who suggested that these two paleofloras (16–15.5 Ma) grew at relatively high altitude (predicted as ca. 2700–2800 m above sea level), whereas later paleofloras in the region (ranging from 14–12 Ma) yield estimates near present-day altitudes. At present, the Middlegate flora locality is at 1600 m altitude. Wolfe et al. (1997) interpreted the difference to indicate subsidence of this part of the Great Basin since the early middle Miocene. The paleofloras and the Eastgate LF existed during the Miocene Climatic Optimum, when extensive ice probably occurred in the Antarctic but prior to the buildup of ice sheets in the Northern Hemisphere (Flower and Kennett 1994; Zachos et al. 2008). Although the Middlegate Basin fossil leaf assemblages occur lower in the local stratigraphic section than the Eastgate LF, they provide an approximate paleoenvironmental setting for the ungulates described herein.
The Middlegate Basin includes nearly 1200 m of rocky outcrops that house fossil vertebrates and plants. This basin could provide a wealth of information from high altitude faunas that bracket the Hemingfordian and Barstovian NALMAs boundaries (~18–12.5 Ma; Tedford et al. 2004) and the Miocene Climatic Optimum (~17–14 Ma; Steinthorsdottir et al. 2020; Westerhold et al. 2020). The Eastgate LF, discovered in the basal-most part of the Monarch Mill Formation, is uniquely positioned geographically between other significant middle Miocene faunas known from several rock units (formations) in the Great Basin (Barstow Formation), southern part of the Basin and Range province, and the Columbia Plateau. Interestingly, there are a few taxa of rodents and carnivores unique to the Eastgate LF that are not known from other middle Miocene faunas. The Middlegate Basin could be an ideal place to test empirically the hypothesis that climate change over complex topography stimulated diversification as proposed by Badgley et al. (2014) using paleofaunal databases (MIOMAP and FAUNMAP) that produce a query list of fossil localities that match search criteria and can create interactive maps of geographic occurrences of mammal taxa.
Material and methods
The locality information recorded for the Eastgate LF by earlier workers is modified herein. For example, 11 of the 18 UCMP V numbers representing the Eastgate localities were assigned to isolated occurrences of individual specimens (Donald E. Savage, personal communication 1999). In addition, past workers provided locality numbers (UCMP V numbers) for each specimen collected side-by-side and from the same horizon within a very restricted geographic locality. Therefore, we use a single locality number (UCMP V and OMNH V) to represent all vertebrate fossils collected within a very restricted geographic area of 5 m2 or less and from within the same stratigraphic horizon (see Table 1). In addition, OMNH V numbers were assigned to established UCMP V localities where fossil specimens were collected by OMNH field crews (see Table 1). During a scientific expedition in 1995, four new fossil localities were discovered. These additional localities (OMNH V974–977) were mapped and rock samples were collected for water screenwashing (Cifelli et al. 1996).
Table 1. Equivalency chart of the localities constituting the Eastgate LF that have been assigned UCMP V and OMNH V numbers. No OMNH locality numbers were created for the following UCMP localities: UCMP V74104, UCMP V87139, UCMP V88079, UCMP V88088, UCMP V92020.
|
Locality UCMP V |
Locality OMNH V |
|
UCMP V67245 |
OMNH V966 |
|
UCMP V 70138 UCMP V70139 UCMP V70141 |
OMNH V967 |
|
UCMP V70140 |
OMNH V968 |
|
UCMP V70142 |
OMNH V969 |
|
UCMP V70143 |
OMNH V970 |
|
UCMP V70144 UCMP V70145 UCMP V 70147 |
OMNH V972 |
|
UCMP V74103 |
OMNH V973 |
According to the records at UCMP, two additional localities are on file for the Eastgate LF (UCMP V70146 and V70148). Unfortunately, these sites do not appear on the topographical map (1:24000; Eastgate, NEV.) on file at UCMP and could not be relocated in the field. In addition, the following localities (UCMP V70148, V87139, V88079, V88080, V92020) are included on the topographic map (Eastgate, NEV.) on file at UCMP but the fossil specimens associated with these localities have been lost.
We follow a standard convention in abbreviating tooth type as I (incisor), C (canine), P (premolar), and M (molar), with upper and lower case letters referring to upper and lower teeth, respectively. The letters D or d preceding the tooth type designation indicates upper or lower deciduous teeth, respectively. In addition, the number preceding the specific tooth type represents the specific locus of the tooth (M1, first upper molar). Dental terminology for ruminant artiodactyls follows Bärmann and Rössner (2011), that for rhinocerotids follows Prothero (1998).
Systematic palaeontology
Order Artiodactyla Owen, 1848
Family Merycoidodontidae Thorpe, 1923
Subfamily Ticholeptinae Schultz and Falkenbach, 1941
Genus Ticholeptus Cope, 1878
Type species: Ticholeptus zygomaticus Cope, 1878; Bridgeport Quarries, Nebraska, USA, early Hemingfordian.
Stratigraphic and geographic range.—Early Hemingfordian through late Barstovian, early to middle Miocene (Lander 1998) in NALMA chronostratigraphic scheme. The earliest known occurrence of Ticholeptus is in the Runningwater Formation of Nebraska, which is considered early Hemingfordian (Hunt 1981; Skinner et al. 1977). Although not listed in Lander (1998), this taxon is known from the late Hemingfordian Massacre Lake LF in northwestern Nevada (Morea 1981). Identifiable remains of this taxon are also known from the Skull Ridge Fauna of New Mexico (Morgan 2015), which is considered early Barstovian in age. Ticholeptus is also known from the Cold Spring fauna of Texas (Hesse 1943; Wilson 1960; Patton 1969; Prothero and Sereno 1980; Tedford et al. 1987; Hulbert 1988) and the Bijou Hills LF of South Dakota, which are considered late Barstovian in age (Green and Holman 1977; Green 1985). Geographic range: USA: California (Hidden Treasure Spring LF), Florida (Willacoochee Creek LF), Idaho (Upper Railroad Canyon LF); Montana (Deep River LF, Anceney LF, and Flint Creek beds); Nebraska (Bridgeport Quarries, Channel Prospect, Site 2 Prospect, Pepper Creek, J.L. Ranch, Lower Snake Creek F, Sand Canyon beds, Observation Quarry, and Survey Quarry); Nevada (Eastgate LF); New Mexico (Skull Ridge Fauna); South Dakota (Bijou Hills LF), and Texas (Cold Spring fauna); Canada: Saskatchewan (Topham LF and Wood Mountain LF).
Ticholeptus sp.
Fig. 3.
Material.—UCMP 141873, left dentary fragment with a complete m1 and an incomplete m2; OMNH 77341, left DP3; OMNH 77348, left astragalus; OMNH 77479, right astragalus; from localities UCMP V70142 and OMNH V974, Eastgate, Churchill County, Nevada, USA; Monarch Mill Formation, early Barstovian, middle Miocene.
Measurements.—UCMP 141873 m1, AP = 14.50 mm; T = 10.73 mm: m2, AP = 15.95 mm; T = 11.89 mm.
Description.—Dentary and cheek teeth: The dentary fragment has no foramina or other features present. The molar teeth present include an m1 and m2, which are semi-hypsodont and completely selenodont. The m1 has a well-developed protoconid and hypoconid with an intercrescentic valley closed by a low, labial cingulum. The paraconid is missing. An anterior cingulum is present but is not continuous with the labial cingulum. A posteromedially-directed metastylid occurs midway between the metaconid and entoconid. The hypoconulid is distinct. Internal ribs (vertical ribs along the lingual aspects of the lingual cusps) of the metaconid and entoconid and posterior cingulid are weak. The m2 is longer and wider than the m1, has a paraconid, and the anterior cingulum is continuous with the labial cingulum. The hypoconulid and posterior cingulum of the m2 are missing.
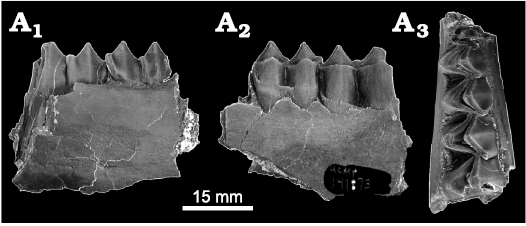
Fig. 3. Merycoidodontid artiodactyl Ticholeptus sp. indet. from locality UCMP V70142, Eastgate Churchill County, Nevada, USA; early Barstovian, middle Miocene. UCMP 141873, partial left dentary fragment with complete m1 and partial m2 in lingual (A1), labial (A2), and occlusal (A3) views.
Remarks.—Lander (1998) listed several characteristics of the m1–m3 for the Merycoidodontidae that are similar to those of the Eastgate specimen (UCMP 141873). These structures include completely selenodont lingual cusps (metaconid and entoconid) with a posteromedially-directed metastylid that occurs midway between the metaconid and entoconid, reduced stylids, and the reduced metaconid and entoconid ribs. The molar teeth of the Eastgate specimen are similar to those of Ticholeptus cf. obliquidens (UO 22798; see Shotwell 1968: 56, fig. 27) from the Red Basin fauna, Oregon in Shotwell (1968). The specimen (UO 22798) described by Shotwell (1968) has lower molars with a labial cingulum closing off the intercrescentic valleys, no protostylid, and other stylids present but not distinct. Morea (1981) identified 11 dentaries of Ticholeptus cf. zygomaticus from the Massacre Lake LF, Washoe County, Nevada; these specimens also are similar to the Eastgate fossil specimen. No measurements were given by Shotwell (1968) for UO 22798 or UO 26798. The greatest length (AP) of the Eastgate m1 occurs within the range of those listed by Morea (1981) for the m1s of T. cf. zygomaticus (m1 AP = 12.70–14.60 mm, T = 8.30–10.10 mm); however, the greatest width (T) is slightly larger for the Eastgate specimen. In addition to size, the specimen from Eastgate is similar in characteristics to those listed above by Shotwell (1968) for T. cf. obliquidens. Teeth are not particularly useful in identifying various taxa of merycoidodontids, and deciduous teeth in the group are little described in the literature. Thin enamel and a low crown height suggest that OMNH 77341 is a deciduous tooth, which is tentatively referred to this genus because no other merycoidodontid is known in the Eastgate LF. Similarly, we tentatively refer the two astragali listed above to Ticholeptus sp. because of their relatively large size, morphology, and differences from the only other artiodactyl astragali in the fauna, which likely pertain to the ruminant palaeomerycid described below. In addition, the specimens extend the known age for Ticholeptus in the Great Basin from the late Hemingfordian (Massacre Lake LF) to the early Barstovian (Eastgate LF).
Family Palaeomerycidae Lydekker, 1883
Subfamily Dromomerycinae Frick, 1937
Tribe Cranioceratini Frick, 1937
Genus Barbouromeryx Frick, 1937
Type species: Barbouromeryx trigonocorneus (Barbour and Schultz, 1934); Antelope Creek, Nebraska, USA; early Hemingfordian.
Barbouromeryx trigonocorneus (Barbour and Schultz, 1934)
Figs. 4–6.
Material.—OMNH 54976, left dentary fragment with p2–m3; OMNH 54977, right dentary with dp3, partial dp4, alveolus with posterior root of m1, and developing-erupting m2; OMNH 77355, right dentary fragment with dp2 or p2 and alveoli for ?dp1; OMNH 77369, partial C1; OMNH 77344, right M1 or M2; OMNH 77368, associated right m1, m3, and fragments of dentary; UCMP 141499, right dentary fragment with m1; UCMP 141518, right dentary fragment with m1 fragment and m2–m3; UCMP 141524, seven isolated cheek teeth and a maxilla fragment with M1-M2; UCMP 141527, median phalanx; UCMP 141650, right dentary fragment with p3–p4; UCMP 141654, right dentary fragment with p3–p4; UCMP 141655, left maxilla fragment with P2-M1; UCMP 141669, left dentary fragment with p3–m3; UCMP 141702, right dentary fragment with p2–m3; UCMP 141863, left maxilla fragment with M2–M3; UCMP 141664, right dp4; UCMP 141667, left DP3; UCMP 141693, left m2 fragment. From localities OMNH V974; UCMP V70138, V70142, and V70140, Eastgate, Churchill County, Nevada; Monarch Mill Formation, early Barstovian, middle Miocene.
Measurements.—See Table 2.
Description.—Upper teeth: All maxillae of Barbouromeryx trigonocorneus at Eastgate are very fragmentary. The upper premolars have three roots, an ectoloph with a low anterior style, external rib, and a posterior style. The upper molars are brachydont and increase in length and width from M1 to M3; however, the M2 is only slightly shorter and narrower than the M3. Other than size, the molars are similar in the following characteristics: four primary cusps (protocone, paracone, metacone, and metaconule); parastyle, mesostyle, metastyle, and entostyle; strong, anteriorly-inflected external rib on the labial surface of the paracone that forms a small extra loph that will shorten with increasing wear; strong, thin anterior cingulum with deep gutter; and posterior cingulum weak (or absent on M3). In all upper molars the postprotocrista curves anteriorly at its labial end and is bifurcated in little-worn teeth, with the anterior branch approaching or contacting the base of the paracone and the posterior branch contacting the premetaconulecrista. The premetaconulecrista may also be bifurcated at its labial end and in contact with the postprotocrista but not in contact with the paracone or metacone. An external rib is absent on the metacones of all upper molars. The metaconule folds are weak.
Lower cheek teeth: All dentaries are fragmentary; thus, we provide no information on the coronoid process, condyle, ramus, or foramina. The cheek teeth are brachydont and p1 is absent. One specimen (OMNH 77355) contains two small alveoli that might have held a dp1. The premolars are unreduced relative to the molars and are not molariform. In two specimens (UCMP 141669 and OMNH 54976), the premolar row is long relative to the molar row length and entire cheek tooth row length (Table 2); the specimens fit within the ranges of these dimensions provided for Barbouromeryx trigonocorneus by Prothero et al. (2014: fig. 5). The p3 and p4 each have five transverse lophids. The p4 anterior valley is not closed by an anterolingual cristid of the mesolingual conulid (“metaconid” of previous authors), as there is no anterolingual cristid. In p4 the posterolingual conid and posterior stylid are crest-like and converge posterolingually, but a vertical groove remains between their lingual ends. In one dentary (OMNH 54977) of a juvenile are teeth that we interpret as dp3, partial dp4, alveolus with posterior root of m1, and m2 in the crypt; the deciduous premolars of this specimen differ in morphology from the permanent premolars in other Eastgate B. trigonocorneus dentaries. The dp3 is complete and has five lophs: anterior stylid, anterior conid, mesolingual conid, posterior cristid, and posterior stylid. These lophs are similar to those of the available permanent p3s except for minor differences. The distal (lingual) end of the anterior conid sweeps posteriorly as it descends to the base of the crown, partly closing off the bottom of the anterior valley. The mesolingual conid has its posterolingual cristid angled posterolingually. The posterior cristid is confluent with the posterior stylid, closing off the back valley with no notch or vertical groove on the posterolingual corner of the tooth. The crown of the dp4 is mostly broken away except for an anterior portion, but the alveolus and root fragments indicate that it had three lobes like a normal ruminant dp4. There is a portion of the posterior root remaining in a socket that is confluent with the labially situated socket and its small root that supported the second lobe. The first (anterior) lobe has a large low anterolabial conid with a tiny anterior stylid that juts forward; with increasing wear this stylid would have joined the anterior cingulid. The anterior cristids of the anterolabial and anterolingual conids are confluent. A low anterior ectostylid is present. The m1 is completely broken away and is represented only by its posterior root. The erupting m2 in this specimen is identical to those of the other specimens described below. In its unworn state it bears a short external postmetacristid that will disappear into an external rib of the metaconid with wear.
All lower molars have strong external postprotocristids (the “Palaeomeryx-folds” of previous authors) that form a small extra lophid. The m1 and m2 have prominent anterior and posterior cingulids and ectostylids. The m3 has a strong anterior cingulid, a moderate ectostylid in the anterior external valley, a weak ectostylid in the posterior external valley, and a weak or absent posterior cingulid on the posterior extremity of the tooth. The m3 posterior lobe imitates the shapes of the trigonid and talonid lobes, having a lingual preentoconulidcristid–entoconulid–postentoconulidcristid and labial selene formed by the prehypoconulidcristid–hypoconulid–posthypoconulidcristid, and the back fossa is closed posteriorly.
Table 2. Measurements (in mm) of Barbouromeryx trigonocorneus teeth from the Eastgate fauna. Brackets indicate estimated measurement of a broken specimen. Abbreviations: AP, greatest anteroposterior length of tooth; T, greatest transverse width of tooth; TA, greatest transverse width of anterior half of tooth; TAW, greatest width of talonid part of tooth; TP, greatest transverse width of posterior half of tooth; TRW, greatest width of trigonid part of tooth.
|
Repository number |
Tooth series or locus |
AP |
T |
TA or TRW |
TP or TAW |
|
UCMP 141669 |
p2–m3 |
60.4 |
|
|
|
|
p2–p4 |
24.6 |
|
|
|
|
|
m1–m3 |
36.2 |
|
|
|
|
|
p3 |
9.3 |
5.2 |
|
|
|
|
p4 |
10.6 |
5.7 |
|
|
|
|
m1 |
11.3 |
|
6.2 |
6.8 |
|
|
m2 |
11.4 |
|
7.0 |
7.1 |
|
|
m3 |
15.4 |
|
7.2 |
6.8 |
|
|
OMNH 54976 |
p2–m3 |
64.4 |
|
|
|
|
OMNH 54976 |
p2–p4 |
25.2 |
|
|
|
|
m1–m3 |
39.1 |
|
|
|
|
|
p2 |
6.5 |
3.3 |
|
|
|
|
p3 |
9.2 |
5.0 |
|
|
|
|
p4 |
10.1 |
6.3 |
|
|
|
|
m1 |
11.3 |
|
7.0 |
7.4 |
|
|
m2 |
12.6 |
|
7.7 |
8.0 |
|
|
m3 |
16.4 |
|
[7.6] |
7.2 |
|
|
OMNH 77355 |
dp2 or p2 |
7.3 |
3.0 |
|
|
|
UCMP 141518 |
m1 |
|
|
|
7.4 |
|
m2 |
11.7 |
|
8.1 |
8.1 |
|
|
UCMP 141518 |
m3 |
17.7 |
|
7.9 |
7.5 |
|
OMNH 77368 |
m1 |
[11.1] |
|
6.0 |
6.6 |
|
m3 |
15.8 |
|
[7.6] |
6.9 |
|
|
OMNH 54977 |
dp2 or p2 |
8.1 |
3.9 |
|
|
|
dp3 or p3 |
|
[4.7] |
|
|
|
|
OMNH 77344 |
M1 or M2 |
11.7 |
|
12.4 |
[10.9] |
|
UCMP 141863 |
M2 |
11.8 |
|
11.1 |
10.3 |
|
M3 |
10.0 |
|
10.8 |
8.9 |
|
|
OMNH 77369 |
C1 |
8.5 |
5.0 |
|
|
Remarks.—We follow Janis and Scott (1987) and Prothero and Liter (2008) in placing Dromomerycinae in the family Palaeomerycidae. Janis and Manning (1998) provided characters for the identification of the subfamilies and tribes, including dental characters. Members of this family are largely diagnosed on cranial appendages (Janis and Manning 1998; Prothero and Liter 2007, 2008), none of which were found with the Eastgate fauna.
The ungulate upper canine (OMNH 77369) from Eastgate was unassociated with cheek teeth or other remains. It is tentatively referred to Barbouromeryx trigonocorneus because the type specimen for the species, a cranium, was found with a large saber-like tusk associated near the palate and assumed to belong to the cranium (Barbour and Schultz 1934). Although no measurements of the tusk were provided by Barbour and Schultz (1934), reviewing published photographs aided in determining the size of the C1 alveolus of the cranium of the type specimen published in Prothero and Liter (2008). The C1 anteroposterior length in the type is about 1cm. OMNH 77369 has an anteroposterior length of 8.5 mm (see Table 2). The Eastgate tusk is missing the tip of its crown as well as the base of the root. The crown in cross-section has a “teardrop” shape with the anterior edge rounded and the posterior edge pointed and covered with extremely thin enamel (Fig. 4).

Fig. 4. Palaeomerycid artiodactyl tentatively referred to Barbouromeryx trigonocorneus (Barbour and Schultz, 1934), from locality OMNH V974, Eastgate, Churchill County, Nevada, USA; early Barstovian, middle Miocene. OMNH 77369, partial upper tusk (C1) in labial view with remnant strips of very thin enamel (A1), lingual view (A2), cross-section at natural break in crown (A3).
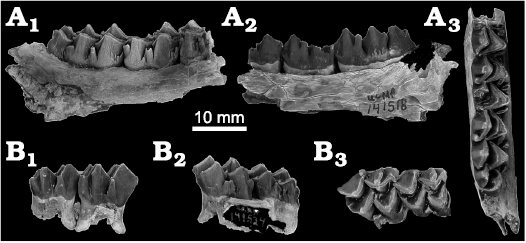
Fig. 5. Palaeomerycid artiodactyl tentatively referred to Barbouromeryx trigonocorneus (Barbour and Schultz, 1934), from locality UCMP V70138, Eastgate Churchill County, Nevada, USA; early Barstovian, middle Miocene. A. UCMP 141518, right dentary fragment with m1 fragment and m2–m3. B. UCMP 141524, maxilla fragment with M1–M2. In labial (A1, B1), lingual (A2, B2), and occlusal (A3, B3) views.
Among the Palaeomerycidae, only members of the tribe Aletomerycinae (Aletomeryx and Sinclairomeryx) and the tribe Cranioceratini (only the Barbouromeryx) possess upper canines (Prothero and Liter 2008). The palaeomerycids Aletomeryx and Sinclairomeryx have upper canines that are reduced, but the Eastgate dentaries bear molars having strong external postprotocristids (“Palaeomeryx-folds”), so the dentaries cannot be referred to Aletomerycinae, in which these folds are weak to absent (Janis and Manning 1998; Prothero and Liter 2008). The Eastgate premolars are unreduced relative to molar size, are not molariform, and are mesodont; dp1 is not retained. These characteristics do not match those of aletomerycines or dromomerycines as diagnosed by Janis and Manning (1998) and Prothero and Liter (2008). Thus, based on the cheek teeth, the Eastgate jaws pertain to a member of the Cranioceratini, either Bouromeryx or Barbouromeryx. As revised by Prothero and Liter (2008), these two genera are essentially identical dentally except that Barbouromeryx has upper canines while Bouromeryx lacks canines (Prothero and Liter 2008). Based on known temporal ranges of these two genera, Bouromeryx extends into the Barstovian while Barbouromeryx is only known prior to the Barstovian. The Eastgate palaeomerycid could possibly represent Bouromeryx unless the isolated canine belongs to the same taxon as the multiple dentaries. If the Eastgate palaeomerycid represents Bouromeryx, the presence of an unassociated but enlarged artiodactyl canine is difficult to explain unless more than one palaeomerycid taxon is present, one of which bears upper canines. Given the currently available sample, we believe it is more parsimonious to assume that a single dromomerycine taxon is present at Eastgate. If the Eastgate canine represents the tusked genus Barbouromeryx, with one recognized species, B. trigonocorneus, ranging from latest Arikareean to middle Hemingfordian (Prothero and Liter 2008), then the temporal range of B. trigonocorneus must be extended into the Barstovian. Alternatively, the Eastgate isolated canine might represent a moschid or gelocid for which no cheek teeth or other skeletal elements have been found (Prothero 2008; Webb 1998, 2008).
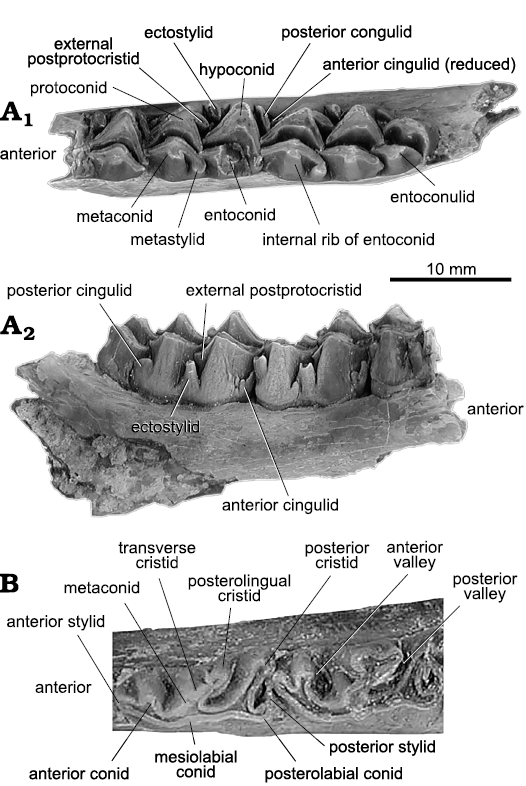
Fig. 6. Dental terminology for palaeomerycid artiodactyl Barbouromeryx trigonocorneus (Barbour and Schultz, 1934), from Eastgate, Churchill County, Nevada, USA; early Barstovian, middle Miocene. A. UCMP 141518 from UCMP V70138; right dentary fragment with m1 fragment and m2–m3 in occlusal (A1) and labial (A2) views. B. UCMP 141669 from UCMP V70140; left dentary fragment with p3–m3 in occlusal view (only p3–p4 figured).
Stratigraphic and geographic range.—Latest Arikareean to middle Hemingfordian (Janis and Manning 1998; Prothero and Liter 2008). The Eastgate LF record extends the range to early Barstovian (Smith 2002). Geographic range: USA: Nebraska (Anderson Ranch Formation, Runningwater Formation, Box Butte Formation), South Dakota (Batesland Formation), Colorado (Martin Canyon Formation), Texas (Oakville Formation), and Wyoming (Split Rock Formation) (Prothero and Liter 2008); and Nevada (Eastgate LF).
Order Perissodactyla Owen, 1848
Family Equidae Gray, 1821
Subfamily Equinae Gray, 1821
Stratigraphic and geographic range.—Early Arikareean to Recent (MacFadden 1998); Eurasia, Africa, and North America (in the Miocene).
Equinae gen. et sp. indet.
Fig. 7.
Material.—UCMP 141537, middle phalanx; UCMP 141698, astragalus; UCMP 141837-01, astragalus; UCMP 141837-02, isolated tooth fragments; UCMP 141899-01, proximal part of humerus; UCMP 141899-02, magnum; UCMP 141899-03 metatarsals II or IV; UCMP 141921, distal part of tibia. From localities UCMP V70138, V70142, V70143, V70145, V70148, Eastgate, Churchill County, Nevada, USA; Monarch Mill Formation, early Barstovian, middle Miocene.

Fig. 7. Articulating distal tibia and astragalus and isolated magnum elements of Equinae gen. et sp. indet. from localities UCMP V70145 (A, B) and UCMP V70142 (C), Eastgate Churchill County, Nevada, USA; early Barstovian, middle Miocene. A. UCMP 141921, distal tibia in anterior view. B. UCMP 141837-01, astragalus in anterior view. C. UCMP 141899-02, magnum in dorsal view.
Remarks.—The elements listed above were identified by Donald E. Savage in the 1950s as belonging to “Merychippus” sp. Traditionally, Merychippus included a paraphyletic/polyphyletic assemblage of subhypsodont to hypsodont horses (Hulbert and MacFadden 1991; MacFadden 1998) that are now placed in several different genera. The fossil horse material from the Eastgate LF does not provide sufficient information to allow taxonomic identification below the subfamily level.
Family Chalicotheriidae Gill, 1872
Subfamily Schizotheriinae Holland and Peterson, 1914
Genus Moropus Marsh, 1877
Type species: Moropus oregonensis (Leidy, 1873), by Coombs et al. (2001), Sutton Mountain, Oregon, USA; upper John Day Formation, late Arikareean.
Moropus merriami Holland and Peterson, 1914
Fig. 8.
Material.—UCMP 80398, olecranon process of ulna, distal metacarpal II, partial lunate; UCMP 141896, duplex = fused proximal and middle phalanges of digit II manus; UCMP 141898, duplex of digit II manus; UCMP 141917, proximal end of proximal phalanx; UCMP 141946, proximal end and shaft of humerus; UCMP 141947-01, metatarsal III; UCMP 141947-02, metatarsal IV; UCMP 141947-03, metatarsal II; UCMP 141948-01, metatarsal III; UCMP 141948-02, metatarsal IV; UCMP 141948--03, metatarsal II; UCMP 141954-01, ungual phalanx; UCMP 141954-02, proximal phalanx; UCMP 141960, left metacarpal IV; UCMP 141978-01, complete tibia; UCMP 141978-02, partial radius; UCMP 141978-03, ulna shaft; UCMP 141978-04, distal fused radius and ulna; UCMP 141839, distal end of right scapula. Additional skeletal elements of M. merriami occur in the UCMP collections from Eastgate and will be presented in a separate study by Margery Coombs. Chalicothere material has been collected from at least seven UCMP Eastgate vertebrate localities: V70141, V70142, V70143, V70144, V70146, V74103, V67245. Most of the fossil materials are from locality V74103, informally called the Chalicothere Quarry or Burmester Chalicothere Quarry in UCMP field records. Although given separate catalog numbers, some of the elements collected from V74103 articulate and probably belonged to one individual.
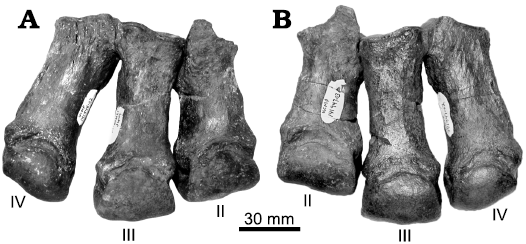
Fig. 8. Articulating right and left hind feet elements of chalicotheriid perissodactyl Moropus merriami Holland and Peterson, 1914, from locality UCMP V74103, Eastgate, Churchill County, Nevada, USA; early Barstovian middle Miocene. A. UCMP 141948-02, -01, -03, right metatarsals IV–II. B. UCMP 141947-03, -01, -02, left metatarsals II–IV).
Remarks.—Holland and Peterson (1914) named Moropus merriami for UCMP material collected from Virgin Valley and the High Rock Canyon area in northwestern Nevada and figured by Merriam (1911) as “Moropus(?) sp.”. They named syntypes that included an astragalus, a calcaneum, fused proximal and middle phalanges (a duplex) of digit II of the manus, two ungual phalanges, a P4, and an m2. Because the syntypes come from several localities, they cannot possibly represent a single individual. Nonetheless, Coombs (2004), while presenting a revised species diagnosis, refrained from designating a lectotype on the grounds that the complete series gives a better sense of the species than any single lectotype would do. Moropus merriami was tentatively referred to either the genus Macrotherium or Chalicotherium by Matthew (1929) and others; however, it is clear on the basis of both dental and postcranial anatomy that the species belongs to the Schizotheriinae, not Chalicotheriinae. Coombs (1978, 1998, 2004) has maintained the original referral to Moropus.
The virtual lack of craniodental material is not a significant barrier in identifying the Eastgate chalicothere material. Skull remnants and teeth are rare in general among Nevada chalicotheres; the type series of M. merriami includes only a P4 and m2. On the other hand, postcranial elements, particularly podials, metapodials, and some phalanges, are very useful in species identification. For example, Coombs (2004) used the proportions and morphology of metatarsals as important aids in the identification of chalicotheres from the Lower Snake Creek LF of the Great Plains as belonging to M. merriami. The well-preserved, robust metatarsals of the Eastgate chalicothere similarly provide important evidence to support assignment to M. merriami (see Table 3). A fuller comparison awaits a more detailed description of all Nevada material belonging to this species (MCC unpublshed data).
Table 3. Measurements (in mm) and proportions of metatarsals referable to Moropus merriami. UCMP 141947 (left) and UCMP 141948 (right) from Eastgate probably belong to one individual. Compare proportions with Coombs (1978: table 6, 1979: table 8, 2004: table 1). * corrected from Coombs (2004).
| |
Specimen |
Maximum length |
Maximum distal width |
Minimum shaft width |
Maximum length: maximum distal width |
Maximum length: minimum shaft width |
|
Metatarsal II |
UCMP 141947-03 |
106.0 |
50.8 |
36.2 |
2.1 |
2.9 |
|
UCMP 141948-03 |
103.6 |
49.5 |
32.8 |
2.1 |
3.2 |
|
|
UCMP 11858 |
112.3 |
55.3 |
38.4 |
2.0 |
2.9 |
|
|
SCNHM VMO 545 |
116 (approx.) |
52.1 |
40.0 |
2.2 |
2.9 |
|
|
SCNHM VMO 680 |
96.2 |
43.0 |
35.5 |
2.2 |
2.7 |
|
|
Metatarsal III |
UCMP 141947-01 |
111.1 |
46.5 |
30.7 |
2.4 |
3.6 |
|
UCMP 141948-01 |
109.7 |
46.3 |
31.1 |
2.4 |
3.5 |
|
|
UCMP 35582 |
97.6 |
41.1 |
28.2 |
2.4 |
3.5 |
|
|
UCMP 31417 |
128.7 |
53.8 |
40.7 |
2.4 |
3.2* |
|
|
AMNH F:AM 54176 |
128.3 |
51.1 |
35.3 |
2.5 |
3.6 |
|
|
Metatarsal IV |
UCMP 141947-02 |
116.4 |
45.0 |
36.0 |
2.6 |
3.2 |
|
UCMP 141948-02 |
115.7 |
47.5 |
36.1 |
2.4 |
3.2 |
|
|
SCNHM VMO 547 |
130.2 |
46.3 |
40.8 |
2.8 |
3.5 |
|
|
AMNH F:AM 54172 |
135.5 |
46.6 |
37.4 |
2.9 |
3.6 |
Although chalicothere fossils are numerous at a few unusual sites (most famously Moropus elatus Marsh, 1877, at the Agate Spring Quarries and Morava Ranch Quarry in Nebraska; see Coombs and Coombs 1997), it is more typical to find them sporadically and as isolated elements. Classifying these elements is often an exercise in comparing teeth from one place with footbones from another, with less than conclusive results. Fortunately, the substantial number of chalicothere elements from Nevada allow more complete and definitive comparisons. In particular, the articulating hind foot elements of M. merriami from Eastgate (UCMP locality V74103) help to clarify the proportions of bones belonging to a single animal. Metatarsals III and IV are of subequal length in UCMP 141947 and UCMP 141948 (Fig. 6; Table 2), with metatarsal IV slightly longer. Similar metatarsal IV/metatarsal III proportions occur in M. elatus (Coombs 1978: table 5), while in the North American dome-skulled chalicothere Tylocephalonyx skinneri Coombs, 1979, metatarsal III is clearly the longest metatarsal (Coombs 1979: table 7).
Stratigraphic and geographic range.—Late Hemingfordian to early Barstovian (Coombs 1998, 2004) = early middle Miocene. The earliest known occurrence of M. merriami is in the Massacre Lake local fauna. This fauna is also important for the presence of Zygolophodon, one of the earliest proboscidean occurrences in North America (Morea, 1981; Tedford et al. 2004; Prothero et al. 2008). Other clearly identifiable remains of M. merriami are early Barstovian in age, The known geographic range includes Nevada and northwest Nebraska (USA). North of Eastgate (Churchill County, Nevada), M. merriami occurs at UCMP localities in Washoe and Humboldt counties in the Massacre Lake fauna, Virgin Valley fauna, Fly Canyon fauna, and Smoky Creek fauna, as well as in the Yellow Hills fauna, which was excavated in the High Rock Caldera by field parties from Sierra College (Bromm and Hilton 2011; Hilton and Hausback 2015).
Family Rhinocerotidae Gray, 1821
Subfamily Aceratheriinae Dollo, 1885
Genus Teleoceras Hatcher, 1894
Type species: Teleoceras major Hatcher, 1894; Loup Fork Beds, Nebraska, USA; early–late Barstovian.
Stratigraphic and geographic range.—Early Hemingfordian through late Blancan, middle Miocene to early Pleistocene (Prothero 1998; Tedford et al. 2004) in NALMA chrostratigraphic scheme. One of the earliest known occurrences of Teleoceras is in Colorado, which is considered early Hemingfordian in age (Martin Canyon Quarry A), whereas one of the latest occurrences is considered late Blancan in age (Broadwater LF in Nebraska). USA: California (Mt. Eden LF); Nevada (Panaca LF, Eastgate LF); Arizona (Keams Canyon LF, Jeddito LF); New Mexico (Nambe Fauna, Skull Ridge Fauna, Pojoaque Fauna, Round Mountain Fauna, and San Juan Fauna); Colorado (Wray localities); Nebraska (Burge and Penny Creek faunas, Sand Canyon, Broadwater LF, Snake Creek Formation); Kansas (Sawrock LF); Texas (Lapara Creek fauna, Coffee Creek LF, Clarendon LF); Oklahoma (Optima LF); Florida (Palmetto Fauna); South Dakota (Ash Hollow Formation).
Teleoceras sp.
Material.—OMNH 54980, tooth fragment; OMNH 54987, distal metapodial III; UCMP 83779, right P3 or P4 fragment; UCMP 120127, palate with left and right P1–M3, symphysis of dentary with left and right i2s; UCMP 141947-03, three phalanges; UCMP 141948-03, three phalanges; UCMP 141841, dentary fragment with fragmented cheek tooth. From localities OMNH V 977, UCMP V67245, V70141, V70142, V74103, Eastgate, Churchill County, Nevada, USA; Monarch Mill Formation, early Barstovian, middle Miocene.
Remarks.—At Eastgate, this rhinoceros is represented by a palate with a complete set of upper cheek teeth. Postcranial specimens are present as well. The upper molars have a diagnostic π-shaped pattern formed by the union of several lophs: protoloph, metaloph, and ectoloph (Prothero 1998). These specimens were identified as belonging to Teleoceras sp. based on the presence of the i2 on the symphysis of the dentary (UCMP 120127) and other characters listed by Prothero (1998). The upper cheek teeth are heavily worn and the postcranial material does not afford a specific identification.
Discussion
Fossil vertebrates have been collected from the Monarch Mill Formation in the Middlegate Basin in Churchill County, Nevada, for nearly 70 years by researchers at the UCMP, OMNH, and SDSM. Although most of the fossils obtained by UCMP were recovered by surface collecting, UCMP V74103 was quarried by Neil Burmester and others while excavating chalicothere bones. Field crews from the OMNH quarried selected localities to collect rock matrix for screenwashing to recover small vertebrates. To date, more than 20 vertebrate fossil localities have been discovered from the Monarch Mill Formation. Three localities in the upper part of the Monarch Mill Formation have yielded mammal fossils of Hemphillian age. One of the three localities (UCMP V5200) discovered by Axelrod (1985) has associated documentation on file at UCMP. Unfortunately, the data and vertebrate fossils from the other two Hemphillian age localities reported by Axelrod (1956, 1985) have been lost. A large carnivoran phalanx was found in the same area by OMNH crew in 2016 (Jeff Hargrave, unpublished data). The remaining 18 localities, which were found in the basal part of the Monarch Mill Formation, have yielded the Eastgate LF (Smith 2002; Smith et al. 2006, 2016), of probable early Barstovian age.
No biostratigraphy is yet possible for the Monarch Mill Formation that could contribute to improve the utility of the evolutionary stages of its fossil mammals for NALMAs in the manner supported by Woodburne (2006). This is because of the restricted stratigraphic interval of localities yielding the Eastgate LF near the bottom of the formation (but not placed within measured stratigraphic sections) and the single unnamed Hemphillian assemblage at the top, with no other occurrences yet known in between. Nor is there a geomagnetic polarity stratigraphy available for the Monarch Mill Formation. However, there are several radiometric dates for geochronologic correlation and published temporal spans of taxa from other localities elsewhere for biochronologic correlation by which we interpret the age of the Eastgate LF.
Biochronology.—Woodburne and Swisher (1995) placed the Hemingfordian–Barstovian boundary at about 15.9 Ma. Tedford et al. (1987, 2004) divided the Barstovian into early and late intervals, informally designated as Ba1 and Ba2. Chronologically, the early Barstovian (Ba1) extends from the Hemingfordian–Barstovian boundary (ca. 16.0 Ma) to about 14.7 Ma, and the late Barstovian (Ba2) ends at the Barstovian–Clarendonian boundary at about 12.5 Ma (Woodburne 2004).
The vertebrate fossils of the Monarch Mill Formation in the Middlegate Basin have previously been interpreted as representing a period spanning from early Barstovian (earlier fauna, from the lower-most part of the rock unit; Smith 2002; Smith et al. 2006, 2016) to Hemphillian (later fauna; Axelrod 1956, 1985). No fauna of intermediate age (e.g., Clarendonian) has been found. In this paper we focus on ungulates, also incorporating other identified mammal taxa, to bracket the age of the earlier (Eastgate) local fauna.
Some of the taxa from the Eastgate LF have proved to be unique to the fauna (the eomyid rodent Apeomyoides savagei and the ailurid and mustelid carnivorans Actiocyon parverratis, Brevimalictis chikasha, and Negodiaetictis rugatrulleum; Smith et al. 2006, 2016), thus, they are not helpful in correlation. Temporally-restricted mammal taxa whose temporal ranges are known from other North American faunas help to establish the age of the Eastgate LF (Fig. 9). Especially useful are the rodent Tardontia nevadans and the chalicothere Moropus merriami. Tardontia nevadans is otherwise known only from the nearby Stewart Springs and Tedford Pocket faunas (both in the Esmeralda Formation of southwestern Nevada; Hopkins 2019) of early Barstovian age (Ba1; Janis et al. 2008) and thus suggests that the Eastgate LF is early Barstovian (Smith 2002; Hopkins 2019). Moropus merriami is restricted to the late Hemingfordian and early Barstovian (Coombs 1998, 2004). Therefore, the presence of M. merriami and T. nevadans at the Eastgate LF appears to support an early Barstovian age for this fauna.
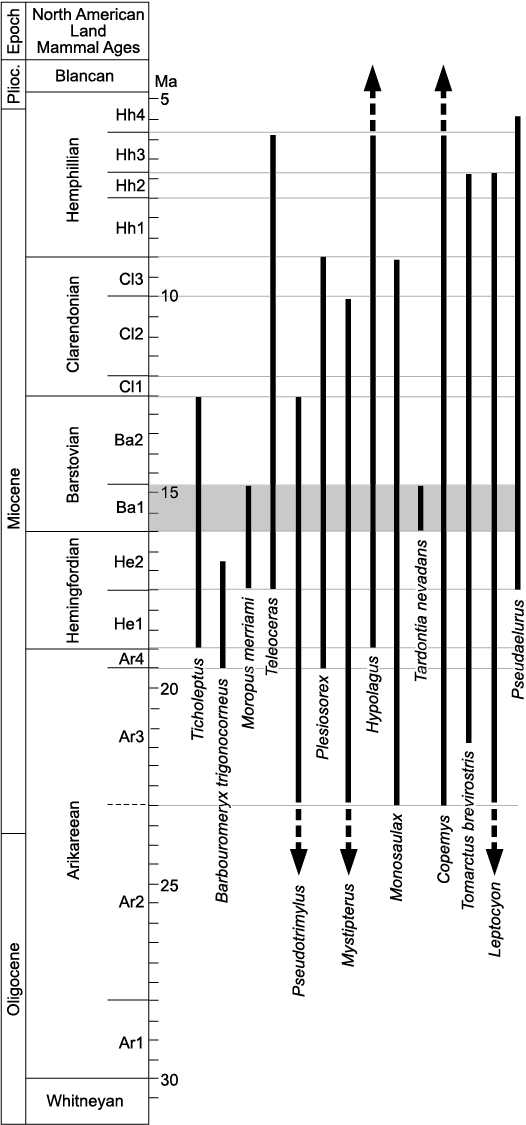
Fig. 9. Biochronologic ranges of land mammals of the Eastgate local fauna (horizontal grey zone). North American Land Mammal ages and subages and absolute time scale in Ma adapted from Tedford et al. (2004: fig. 6.2). Dashed lines and arrows indicate that the temporal range for the taxon continues beyond the Miocene. Ranges for taxa taken from Munthe (1998); Lander (1998); Martin (1998); Dawson (2008); Flynn and Jacobs (2008a, b); Gunnell et al. (2008); Lindsay (2008); Prothero and Liter (2008).
Tedford et al. (2004) and Woodburne (2004) defined the onset of the Barstovian on the first North American occurrences of Plithocyon (a hemicyonine ursid) and on the North American “Proboscidean Datum”, the first occurrence of Zygolophodon. Unfortunately, neither proboscideans nor Plithocyon are known from the Middlegate or Monarch Mill formations. Except for the palaeomerycid Barbouromeryx trigonocorneus, most other Eastgate LF mammals are long-ranging but do not conflict with an early Barstovian age. B. trigonocorneus is known to range from latest Arikareean to middle Hemingfordian (Prothero and Liter 2008) and thus would suggest an earlier age for the Eastgate LF. For reasons noted above, even though diagnostic horn cores are unavailable from Eastgate, we provisionally maintain this identification for the Eastgate palaeomerycid and argue that its occurrence there potentially represents its highest stratigraphic datum, and a temporal range extension for the taxon into the early Barstovian.
Morgan (2015) provided a comprehensive review of Barstovian faunas from the southern Basin and Range province in the Española Basin of New Mexico. These included one early Barstovian (Ba1) fauna (Skull Ridge Fauna) and 10 late Barstovian (Ba2) faunas (Chama-El Rito; Ojo Caliente; Pojoaque, Santa Cruz sites and Bluffs sites; Jacona Microfauna Quarry; Cerro Conejo, Rincon Quarry, and Red Cliff Prospect; and Cerro Conejo, Alamo Ranch and Benavidez Ranch). Teleoceras is the only ungulate from the Skull Ridge Fauna (Ba1) that is known from the Eastgate LF. All known Barstovian sites in New Mexico and many elsewhere have Equinae, but until the species in the Eastgate LF can be identified, these specimens are not helpful for biochronologic correlation. All New Mexico middle Miocene faunas lack Moropus merriami found at the Eastgate LF. Chalicothere fossils are rare, so the absence of M. merriami in the Skull Ridge Fauna is not surprising. Alternatively, the absence of M. merriami from the Skull Ridge Fauna could be related to paleoenvironment. Coombs and Reynolds (2015) described material of Moropus sp. in late Hemingfordian and early Barstovian faunas of the Cajon Valley Formation of southern California; in contrast no chalicothere has ever been found in the well-studied early Barstovian faunas of the nearby Barstow Formation. Coombs and Reynolds (2015) attributed the Cajon Valley occurrences to moister and more vegetated environmental conditions. Pagnac and Reynolds (2010) described an analogous distribution of the equine horse Archaeohippus mourningi, which is common in the Cajon Valley Formation but rare in the Barstow Formation.
Radiometric dates.—Absolute dates have been reported from several middle Miocene rock units (formations) within the Great Basin and Great Plains of North America. Swisher (1992) dated the rocks of the Virgin Valley Formation (including the Virgin Valley fauna) and the Massacre Lake basalt (underlying the Massacre Lake fauna) at about 15.5 Ma and 16.3 Ma, respectively. He named the latter unit “Massacre Lake basalt” and reported a date of 16.5 Ma. Swisher’s (1992) date of 15.96 Ma from the upper-most part of the Middlegate Formation provided a maximum possible age for the Eastgate LF at the base of the overlying Monarch Mill Formation.
Stewart et al. (1999) used tephrochronology to support an indirect age range of 9.8 to 14.7 Ma for eight tephras within the Monarch Mill Formation. Their results revealed a temporal range in the lower part of the formation consistent with a Barstovian age based on mammalian biochronology (Tedford et al. 2004) for the Eastgate LF. However, Stewart et al. (1999) noted several uncertainties in their stratigraphic correlations. Because the Monarch Mill Formation unconformably overlies the Middlegate Formation, there is a temporal gap of unknown duration between the two rock units. Stewart et al. (1999: fig. 3) provided tephra dates of 15.41–15.18 Ma in the Middlegate Formation just below the unconformity and 14.72–14.43 Ma in the lower Monarch Mill Formation above the unconformity. In 2014, we collected basalt samples to obtain radiometric dates specifically to further calibrate the Eastgate LF localities. Tentative ages on basalts we collected in the upper-most part of the Middlegate Formation and the lower-most part of the Monarch Mill Formation range from 14.85–14.65 Ma, respectively.
Paleobiogeography.—The ungulates of the Eastgate LF include two taxa of Artiodactyla (Merycoidodontidae, Ticholeptus sp., and Palaeomerycidae, Barbouromeryx trigonocorneus) and three taxa of Perissodactyla (Equinae, Moropus merriami, and Teleoceras sp.). Three of the five ungulate taxa (Ticholeptus, Moropus, and Equinae) are known from other middle Miocene faunas (Massacre Lake, Virgin Valley, Fly Canyon, Smoky Creek, and Yellow Hills) from the Columbia Plateau, whereas one of the taxa (Ticholeptus) of the Eastgate LF taxa is known from an early Barstovian fauna (Skull Ridge Fauna) in the Española Basin of New Mexico, and all taxa (Ticholeptus, Barbouromeryx, Equinae, Moropus, Teleoceras) of the Eastgate LF are known from Great Plains faunas.
Ticholeptus was reported by Morea (1981) as the most abundant taxon in the Massacre Lake LF in the Columbia Plateau physiographic province. In the Great Plains province, Ticholeptus has been recovered from the Wood Mountain fauna in the southernmost part of Saskatchewan (Storer 1975, 1978, 1993), in a late Barstovian fauna from the lower part of the Valentine Formation of Nebraska (Tedford et al. 2004), and from the Eubanks fauna described by Galbreath (1953) from the lower part of the Pawnee Creek Formation (early Barstovian) in northeastern Colorado. Finally, in the Gulf of Mexico Coastal Plain, another early Barstovian fauna, Willacoochee Creek LF, of Florida included Ticholeptus (Bryant 1991, 1992; Tedford et al. 2004). The paleobiogeographic occurrence of this taxon thus ranges from the southeastern to the northwestern regions of North America.
Barbouromeryx trigonocorneus was previously known from a limited geographic occurrence in a few faunas reported from Nebraska that ranged in age from the late Arikareean to middle Hemingfordian. The earliest representative of B. trigonocorneus is from the latest Arikareean fauna of the Upper Harrison beds (= Anderson Ranch Formation; see Hunt 2002), Nebraska. Other Great Plains occurrences are from the Hemingfordian in Nebraska, South Dakota, Colorado, and Wyoming (Prothero and Liter 2008). The presence of B. trigonocorneus in the Eastgate LF extends the paleogeographic range for this taxon from the Great Plains to the Great Basin in west-central Nevada.
In the middle Miocene, Equinae are known throughout North America. Without knowing the taxon or taxa to which the Eastgate horse belongs, we cannot comment on the paleobiogeography of the horse(s) at Eastgate.
Although early Barstovian chalicothere fossils are rare, fragmentary material is available from several sites. The schizotheriine chalicothere Moropus merriami is known from the nearby early Barstovian Virgin Valley fauna of the Columbia Plateau. It is also known from the Massacre Lake fauna (Morea 1981; Coombs 1998) from northwestern Nevada. This late Hemingfordian fauna shares similar faunal elements with faunas of the Great Plains of the same age (Tedford et al. 2004). In the case of chalicotheres, overlap is with Great Plains early Barstovian (Coombs 2004), not late Hemingfordian, when a different taxon, Tylocephalonyx skinneri is found in the Sheep Creek Formation (Coombs 1979).
The rhinocerotid genus Teleoceras is widespread in North America, with several species identified. In late Hemingfordian and Barstovian faunas the genus occurs in the Great Basin (Massacre Lake LF, Virgin Valley beds, and Esmeralda Formation, Nevada), Columbia Plateau (Oregon), Pacific Coast (California), Great Plains (Nebraska and Colorado), southern Rocky Mountain System (New Mexico), and Gulf of Mexico Coastal Plain (Texas) (Prothero 2005; Morgan 2015).
Overall, these Eastgate LF ungulates show paleobiogeographic affinities with western intermontane basins (Columbia Plateau and other Great Basin faunas) and the Great Plains.
Paleoecology.—Based on its early Barstovian NALMA and roughly 14.85–14.65 Ma radiometric age, the Eastgate LF existed during or just after the middle Miocene Climatic Transition (MMCT) to cooler temperatures following the Miocene Climatic Optimum (MCO) (Steinthorsdottir et al. 2020). Smith et al. (2016) briefly summarized paleobotanical and paleoenvironmental evidence from the upper Middlegate Formation collected by Axelrod (1956), Graham (1999), and Wolfe et al. (1997). This evidence basically records a moderately high-paleoaltitude flora undergoing change from a sclerophyllous shrub- and tree-dominated chaparral-shrubland (in the Middlegate paleoflora) to a mixed coniferous and hardwood forest (in the Eastgate paleoflora). Both paleofloras are slightly older than the Eastgate LF. Within that background, the Eastgate LF ungulates provide mostly autecological information.
In regard to Ticholeptus, Webb (1977) suggested that the Ticholeptinae were the only group of merycoidodontids to survive into the middle and late Miocene, when the predominant habitats in North America were undergoing a transition from woodland to savanna. Lander (1998) suggested that this subfamily may have been successful in a more open habitat due to their dentition, which was more hypsodont than that of other members of this group. Conversely, mesowear studies of Ticholeptus (T. zygomaticus) suggest that it was a fruit or leaf browser and that Barbouromeryx trigonocorneus was a leaf browser (Semprebon et al. 2019) Mesowear evidence for these artiodactyls thus supports a woodland or forest habitat for the Eastgate LF.
Of the three perissodactyl taxa in the Eastgate LF, the chalicothere Moropus can be used to infer a forested habitat. Chalicothere fossils have generally been considered to be indicators of treed, well-watered environments. Their body proportions, claws, and dental morphology are all suggestive of browsing on vegetation well above ground level (Osborn 1919; Zapfe 1979; Coombs 1983). This interpretation does not necessarily imply closed forests, however. For example, fossils of Moropus elatus have been found most often in stream and waterhole sediments consistent with riparian environments in semiarid savannas with seasonal rainfall (Hunt 1978, 1990; Coombs and Coombs 1997). Stereomicrowear analysis of molar enamel of M. elatus suggests committed browsing on leaves and twigs with some bark consumption (Semprebon et al. 2011). It also shows some large pitting and gouging, probably associated with dust and grit present in relatively open country habitats. Unfortunately, the dearth of dental remains of Moropus merriami from Nevada allowed only one specimen from High Rock Canyon (UCMP V1107) to be included in the chalicothere stereomicrowear study. Stereomicrowear of this tooth, along with that of five specimens of M. merriami from the Lower Snake Creek fauna in the Olcott Formation of Nebraska (Semprebon et al. 2011: table 1), suggested leaf and twig browsing as in M. elatus, but with less large pitting and gouging, as might be expected in relatively closed, more forested environments. Therefore, M. merriami might have inhabited relatively closed, more forested environments than M. elatus. Thus, it is not surprising to find M. merriami, along with forest indicators such as petauristine and tamiine squirrels and diverse insectivores (Smith 2002), in the Eastgate LF.
Equinae had its greatest diversity during the middle Miocene, which included horses that were mixed feeders (browser/grazer) to grazers and inhabited a wide range of habitats (MacFadden 1998; Semprebon et al. 2019). Unfortunately, the fossil horse remains at Eastgate cannot be identified below the subfamily level and thus provide little insight into the paleoecology of the Middlegate Basin.
The presence of Teleoceras at Eastgate complements the faunal diversity and may provide insights into the local paleoecology. For example, Prothero (1998) indicated that the greater abundance of rhinoceroses in Great Plains faunas was an attribute of these animals’ preference for savannas over the more wooded habitats of the intermountain states. Most stable isotope studies and actual grass fossils found in the throats and ribcages of Teleoceras major at Ashfall Fossil Bed State Park, Nebraska (Voorhies and Thomasson 1979) confirm that these rhinoceroses were grazers or possibly mixed feeders (Prothero 2005), while in Florida Feranec and MacFadden (2006) suggested that Teleoceras proterum potentially inhabited open habitat or riparian areas.
Conclusions
Collectively, the mammals so far described from the Eastgate LF continue to support an early Barstovian age for the fauna. Preliminary radiometric dates between 14.85 Ma and 14.65 Ma are not tied directly to measured stratigraphic sections, but in association with the rocks in which the Eastgate LF localities occur, these dates are consistent with a middle Miocene age, and with an early Barstovian age. Paleoecological information from both fossil plants and mammals support an interpretation of a moderately high-elevation woodland habitat for the Eastgate LF. While terrestrial ecosystems such as grasslands were expanding globally during the Neogene, the Eastgate LF provides a unique glimpse into a relatively high-elevation, temperate forest ecosystem in the Great Basin during the middle Miocene.
The taxonomic composition of ungulates at the Eastgate LF is relatively depauperate compared with the carnivorans and small mammals (e.g., rodents and insectivores), but nevertheless contributes to our understanding of the fauna. The artiodactyls include Ticholeptus sp. and Barbouromeryx trigonocorneus. These taxa have a wide paleogeographic distribution across the USA and the former taxon is also known from Saskatchewan (Canada). Interestingly, Ticholeptus sp. is the only ungulate taxon from Eastgate that shares an affinity with the early Barstovian Skull Ridge fauna of New Mexico, and B. trigonocorneus has not previously been reported from the Great Basin. The occurrence of a species of Ticholeptus at Eastgate extends its occurrence from the latest Arikareean to early Barstovian and is the only group of ticholeptines to survive into the middle and late Miocene. At present, the artiodactyls do not provide additional insights into the paleoecology or paleo-altitude for the Eastgate LF as supported by the perissodacytls (and other mammal taxa) and fossil plants.
The perissodactyls include the equid subfamily Equinae, the chalicotheriid Moropus merriami, and the rhinocerotid Teleoceras sp. Currently, the fossil horse material provides little insight into the paleoecological interpretations or age for the Eastgate LF. Teleoceras has a widespread paleogeographic distributions in the USA, and is known from fossil vertebrate faunas ranging from the late Hemingfordian to late Barstovian in age and likely occurred in forested habitats. The chalicotheriid M. merriami supports previous paleoecological interpretations of a forested environment and an early Barstovian age for the Eastgate LF.
Acknowledgements
KSS has been mentored by many professors but few who possess a rare combination of talents, devotion, enthusiasm, humor, and professionalism which collectively had an impact on him. Over the decades, in various ways and degrees, KSS has tried to mold himself after those individuals. Therefore, KSS (and NJC and MCC) are honored to contribute to this expression of gratitude to one such unique professor, Richard L. Cifelli. For help with fieldwork, the authors thank the late Donald E. Savage and J. Howard Hutchison (both UCMP) and other UCMP field crews; Dale Harber (retired from US Forest Service, Price, USA); Ian Browne (Oklahoma State University Center for Health Sciences College of Osteopathic Medicine OSU-COM, Tulsa, USA); Darrin Pagnac (South Dakota School of Mines and Technology, Rapid City South Dakota, USA); Alan Deino (Berkeley Geochronology Center, Berkeley, USA); David Schmidt (National Museum of Natural History, Washington, D.C., USA); Jeff Hargrave (Native Explorers Foundation, Edmond, USA); Patrick Ballard, Nathan Claver, Linden Cowley, Steven Dyer, Jessica Fisher, Kelly Fitzpatrick, Erin Kratz, Kayci Lewis, and Ryan Sullivan (OSU-COM); and participants in the 2014 Native Explorers Program. We thank Janz Hunter (OSU-COM) for editing raw photographs for this manuscript. For curatorial assistance, we thank Jennifer Larsen and Kyle Davies (both OMNH). For the loan of and curatorial assistants with UCMP specimens, we thank Donald E. Savage, Patricia A. Holroyd, J. Howard Hutchison, Neil Burmester, and Jere H. Lipps (all UCMP). A special thanks is extended to our spouses for their unyielding support of our work. For their logistical support in fieldwork, we thank Patty Smith, Lotsee Patterson, and Cheryl and Jessica Czaplewski (Norman, USA). Richard Hilton (Rocklin, USA) generously facilitated examination of material at Sierra College collected by his field group. The University of Massachusetts Natural History Collections has helped fund the research travel necessary to further this study. Collecting permits from the Bureau of Land Management were provided by Thom Burke and Bryan Hockett of the United States Department of the Interior, Bureau of Land Management at the State office in Reno, Nevada, and Rachel Crews, Kristin Bowen, and Patricia Mecham of the Sierra Front Field Office (formerly the Carson City District Office) in Carson City, Nevada, Antiquities Permit for the State of Nevada were graciously provided by Gene Hattori (Curator of Anthropology), Margaret Brown (Curation Assistant), and James Barmore (Director) of the Nevada State Museum in Carson City, Nevada. We sincerely thank Gary Morgan (New Mexico Museum of Natural History and Science, Albuquerque, USA) and Darrin Pagnac (South Dakota School of Mines and Technology, Rapid City, USA) and editors Brian Davis (University of Louisville, USA) and Mathew Wedel (Western University of Health Sciences, Pomona, USA) of Acta Palaeontologica Polonica for their thoughtful and thorough reviews of an earlier draft that improved the final version of the paper. Funding for this project was provided by the Chickasaw Nation; Whitten-Newman Foundation; Oklahoma State University Center for Health Sciences; University of Oklahoma Department of Zoology; Oklahoma State Regents for Higher Education Doctoral Study Grant; Southern Regional Education Board Doctoral Scholars Program Dissertation Year Fellowship; the American Indian Graduate Center, Albuquerque, New Mexico; and the Lt. William Johnson Scott Memorial Scholarship.
References
Axelrod, D.I. 1956. Mio-Pliocene floras from west-central Nevada. University of California Publications in the Geological Sciences 33: 1–321.
Axelrod, D.I. 1985. Miocene floras from the Middlegate Basin west-central Nevada. University of California Publications in the Geological Sciences 129: 1–279.
Badgley, C., Smiley, T.M., and Finarelli, J.A. 2014. Great Basin mammalian diversity in relation to landscape history. Journal of Mammalogy 95 (6): 1090–1106. Crossref
Barbour, E.H. and Schultz, C.B. 1934. A new antilocaprid and a new cervid from the late Tertiary of Nebraska. American Museum Novitates 734: 1–4.
Bärmann, E.V. and Rössner, G.E. 2011. Dental nomenclature in Ruminantia: towards a standard terminological framework. Mammalian Biology 76: 762–768. Crossref
Barrows, K.J. 1971. Geology of the Southern Desatoya Mountains, Churchill and Lander Counties, Nevada. 349 pp. Ph.D. Dissertation, University of California, Los Angeles.
Bromm, G.E. and Hilton, R.P. 2011. The early Barstovian Yellow Hills fauna of northwestern Nebraska. In: CalPaleo 2011 Abstracts with Programs, Sierra College, Rocklin, CA, 3. Sierra College, Rocklin.
Bryant, J.D. 1991. New early Barstovian (middle Miocene) vertebrates from the upper Torreya Formation, eastern Florida Panhandle, Journal of Vertebrate Paleontology 11: 472–489. Crossref
Bryant, J.D., MacFadden, B.J., and Mueller, P.A. 1992. Improved chronologic resolution of the Hawthorn and Alum Bluff Groups in northern Florida: Implications for Miocene chronostratigraphy. Geological Society of America Bulletin 104: 208–218. Crossref
Cifelli, R.L., Madsen, S.K., and Larson, E.M. 1996. Screenwashing and associated techniques for the recovery of microvertebrate fossils. In: R.L. Cifelli (ed.), Techniques for Recovery and Preparation of Microvertebrate Fossils. Oklahoma Geological Survey Special Publication 96 (4): 1–22.
Coombs, M.C. 1978. Reevaluation of early Miocene North American Moropus (Perissodactyla, Chalicotheriidae, Schizotheriinae). Bulletin of the Carnegie Museum of Natural History 4: 1–62.
Coombs, M.C. 1979. Tylocephalonyx, a new genus of North American dome-skulled chalicotheres (Mammalia, Perissodactyla). Bulletin of the American Museum of Natural History 164: 1–64.
Coombs, M.C. 1983. Large mammalian clawed herbivores: a comparative study. Transactions of the American Philosophical Society 73 (7): 1–96. Crossref
Coombs, M.C. 1998. Chalicotherioidea. In: C.M. Janis, K.M. Scott, and L.L. Jacobs (eds.), Evolution of Tertiary Mammals of North America. Vol. 1, Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals, 560–568. Cambridge University Press, Cambridge. Crossref
Coombs, M.C. 2004. Moropus merriami in the early Barstovian Lower Snake Creek fauna of Nebraska, with comments on biogeography of North American chalicotheres. Bulletin of the American Museum of Natural History 285: 191–208. Crossref
Coombs, M.C. and Coombs, W.P., Jr. 1997. Analysis of the geology, fauna, and taphonomy of Morava Ranch Quarry, early Miocene of northwest Nebraska. Palaios 12: 165–187. Crossref
Coombs, M.C. and Reynolds, R.E. 2015. Chalicothere material (Perissodactyla, Chalicotheriidae, Schizotheriinae) from late Hemingfordian and early Barstovian faunas of the Cajon Valley Formation in the Mojave Desert Province of southern California. In: R.E. Reynolds (ed.), Mojave Miocene: The 2015 Desert Symposium Field Guide and Proceedings, 259–273. California State University, Desert Studies Center, Baker.
Coombs, M.C., Hunt, R.M., Jr., Stepleton, E., Albright, L.B., III, and Fremd, T.J. 2001. Stratigraphy, chronology, biogeography, and taxonomy of early Miocene small chalicotheres in North America. Journal of Vertebrate Paleontology 21: 607–620. Crossref
Cope, E.D. 1878. Descriptions of new extinct Vertebrata from the upper Tertiary and Dakota formations. Bulletin of the United States Geological and Geographical Surveys of the Territories 4: 379–396.
Dawson, M.R. 2008. Lagomorpha. In: C.M. Janis, G.F. Gunnell, and M.D. Uhen (eds.), Evolution of Tertiary Mammals of North America. Vol. 2, Small Mammals, Xenarthrans, and Marine Mammals, 293–310. Cambridge University Press, Cambridge. Crossref
Dollo, L. 1885. Rhinocéros vivants et fossils. Revue des Questions Scientifiques 17: 293–300.
Feranec, R.S. and MacFadden, B.J. 2006. Isotopic discrimination of resource partitioning among ungulates in C3-dominated communities from the Miocene of Florida and California. Paleobiology 32: 191–205. Crossref
Flower, B.P. and Kennett, J.P. 1994. The middle Miocene climatic transition: East Antarctic ice sheet development, deep ocean circulation and global carbon cycling. Palaeogeography, Palaeoclimatology, Palaeoecology 108: 537–555. Crossref
Flynn, L.J. and Jacobs, L.L. 2008a. Aplodontoidea. In: C.M. Janis, G.F. Gunnell, and M.D. Uhen (eds.), Evolution of Tertiary Mammals of North America. Vol. 2, Small Mammals, Xenarthrans, and Marine Mammals, 377–390. Cambridge University Press, Cambridge. Crossref
Flynn, L.J. and Jacobs, L.L. 2008b. Castoroidea. In: C.M. Janis, G.F. Gunnell, and M.D. Uhen (eds.), Evolution of Tertiary Mammals of North America: Small Mammals, Xenarthrans, and Marine Mammals, Vol. 2, 391–405. Cambridge University Press, Cambridge. Crossref
Frick, C. 1937. Horned ruminants of North America. Bulletin of the American Museum of Natural History 69: 1–669.
Galbreath, E.C. 1953. A contribution to the Tertiary geology and paleontology of northwestern Colorado. University of Kansas Paleontological Contributions Vertebrata 4: 1–120.
Gill, T. 1872. Arrangement of the families of mammals with analytical tables. Smithsonian Miscellaneous Collections 11: i–vi, 1–98. Crossref
Graham, A. 1999. Late Cretaceous and Cenozoic History of North American Vegetation North of Mexico. 349 pp. Oxford University Press, Oxford. Crossref
Gray, J.E. 1821. On the natural arrangement of vertebrose animals. London Medical Repository Review 15: 296–310.
Green, M. 1985. Micromammals from the Miocene Bijou Hills local fauna. South Dakota: fossiliferous Cenozoic deposits of eestern South Dakota and Northwestern Nebraska. Dakoterra 2: 141–154.
Green, M. and Holman, J.A. 1977. A later Tertiary stream channel fauna from South Bijou Hill, South Dakota. Journal of Vertebrate Paleontology 51: 543–547.
Gunnell, G.F., Bown, T.M., Hutchison, J.H., and Bloch, J.I. 2008. Lipotyphla. In: C.M. Janis, G.F. Gunnell, and M.D. Uhen (eds.), Evolution of Tertiary Mammals of North America. Vol. 2, Small Mammals, Xenarthrans, and Marine Mammals, 89–125. Cambridge University Press, Cambridge. Crossref
Hatcher, J.B. 1894. A median horned rhinoceros from the Loup Fork Beds of Nebraska. American Geologists 13: 149–150.
Hesse, C.J. 1943. A preliminary report on the Miocene vertebrate fauna of southeast Texas. Proceedings and Transcripts Texas Academy of Science 26: 157–179.
Hilton, R.P. and Hausback, B.P. 2015. Constraining paleofaunas near High Rock Lake, northwest Nevada. In: J. Sankey and J. Biewer (eds.), Western Association of Vertebrate Paleontology Annual Meeting, Program with Abstracts. PaleoBios 32 (1) Supplement: 8–9. Crossref
Holland, W.J. and Peterson, O.A. 1914. The osteology of the Chalicotheroidea with special reference to a mounted skeleton of Moropus elatus Marsh, now installed in the Carnegie Museum. Memoirs of Carnegie Museum 3: 189–406. Crossref
Hopkins, S.B. 2019. Phylogeny, systematics, and evolution of hypsodonty in the Aplodontiinae (Mammalia, Rodentia, Aplodontiidae), with the description of several new species. Journal of Vertebrate Paleontology 39: 4. Crossref
Hulbert, R.C., Jr. 1988. Calippus and Protohippus (Mammalia, Perissodactyla, Equidae) from the Miocene (Barstovian–early Hemphillian of the Gulf Coastal Plain). Bulletin of the Florida State Museum, Biological Sciences 22: 221–340.
Hulbert, R.C., Jr. and MacFadden, B.J. 1991. Morphological transformation and cladogenesis at the base of the adaptive radiation of Miocene hypsodont horses. American Museum Novitates 3000: 1–61.
Hunt, R.M., Jr. 1978. Depositional setting of a Miocene mammal assemblage, Sioux County, Nebraska (U.S.A.). Palaeogeography, Palaeoclimatology, Palaeoecology 24: 1–52. Crossref
Hunt, R.M., Jr. 1981. Geology and paleontology of the Agate Fossil Beds National Monument and surrounding region, Sioux County, Nebraska (1972–1978). National Geographic Society Research Reports 13: 263–285.
Hunt, R.M., Jr. 1990. Taphonomy and sedimentology of Arikareean (lower Miocene) fluvial, eolian, and lacustrine paleoenvironments, Nebraska and Wyoming; a paleobiota entombed in fine-grained volcaniclastic rocks. In: M.J. Lockley and A. Rice (eds.), Volcanism and Fossil Biotas, Geological Society of America, Special Paper 244: 69–111. Crossref
Hunt, R.M., Jr. 2002. New amphicyonid carnivorans (Mammalia, Daphoeninae) from the early Miocene of southeastern Wyoming. American Museum Novitates 3385: 1–41. Crossref
Janis, C.M. and Manning, E. 1998. Dromomerycidae. In: C.M. Janis, K.M. Scott, and L.L. Jacobs (eds.), Evolution of Tertiary Mammals of North America. Vol. 1, Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals, 477–490. Cambridge University Press, Cambridge. Crossref
Janis, C.M. and Scott, K.M. 1987. The interrelationships of higher ruminant families with special emphasis on the members of Cervoidea. American Museum Novitates 2893: 1–85.
Janis, C.M., Gunnell, G.F., and Uhen, M.D. (eds.) 2008. Evolution of Tertiary Mammals: Small Mammals, Xenarthrans, and Marine Mammals, Vol. 2. vii + 795 pp. Cambridge University Press, Cambridge. Crossref
Lander, B. 1998. Oreodontoidea. In: C.M. Janis, K.M. Scott, and L.L. Jacobs (eds.), Evolution of Tertiary Mammals of North America. Vol. 1, Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals, 402–420. Cambridge University Press, Cambridge. Crossref
Leidy, J. 1873. Contributions to the Extinct Vertebrate Fauna of the Western Territories. Report of the United States Geological Survey of the Territories, Government Printing Office, Washington DC, Vol. 1. Fossil Vertebrates. 358 pp. US Government Printing Office, Washington, DC.
Lindsay, E.H. 2008. Cricetidae. In: C.M. Janis, G.F. Gunnell, and M.D. Uhen (eds.), Evolution of Tertiary Mammals of North America. Vol. 2, Small Mammals, Xenarthrans, and Marine Mammals, 456–479. Cambridge University Press, Cambridge. Crossref
Lydekker, R. 1883. Synopsis of the fossil Vertebrata of India. Records of the Geological Survey of India 16: 61–69.
MacFadden, B.J. 1998. Equidae. In: C.M. Janis, K.M. Scott, and L.L. Jacobs (eds.), Evolution of Tertiary Mammals of North America. Vol. 1, Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals, 537–559. Cambridge University Press, Cambridge. Crossref
Marsh, O.C. 1877. Notice of some new vertebrate fossils. American Journal of Science, Series 3 14: 249–256. Crossref
Martin, L.D. 1998. Nimravidae. In: C.M. Janis, K.M. Scott, and L.L. Jacobs (eds.), Evolution of Tertiary Mammals of North America. Vol. 1, Terrestrial Carnivores, Ungulates, and Ungulate-like Mammals, 228–242. Cambridge University Press, Cambridge. Crossref
Matthew, W.D. 1929. Critical observations upon Siwalik mammals. Bulletin of the American Museum of Natural History 55: 437–560.
Merriam, J.C. 1911. Tertiary mammal beds of Virgin Valley and Thousand Creek in northwestern Nevada. Part 2. Vertebrate faunas. University of California Publications, Bulletin of the Department of Geology 6: 199–304.
Morea, M.F. 1981. The Massacre Lake Local Fauna (Mammalia, Hemingfordian) from Northwestern Washoe County, Nevada. 247 pp. Ph.D. Dissertation, University of California, Riverside.
Morgan, G.S. 2015. Oligocene and Miocene Vertebrates of New Mexico. In: S.G. Lucas and R.M. Sullivan (eds.), Fossil Vertebrates in New Mexico, New Mexico Museum of Natural History and Science Bulletin 68: 159–232.
Munthe, K. 1998. Canidae. In: C.M. Janis, K.M. Scott, and L.L. Jacobs (eds.), Evolution of Tertiary Mammals of North America. Vol. 1, Terrestrial Carnivores, Ungulates, and Ungulate-like Mammals, 124–143. Cambridge University Press, Cambridge. Crossref
Noble, D.C. 1972. Some observations of the Cenozoic volcano-tectonic evolution of the Great Basin, western United States. Earth and Planetary Sciences Letters 17: 142–150. Crossref
Osborn, H.F. 1919. Seventeen skeletons of Moropus: probable habits of this animal. Proceedings of the National Academy of Science USA 5: 250–252. Crossref
Owen, R. 1848. Description of teeth and proportion of jaws of two extinct Anthracotherioid quadrupeds (Hyopotamus vectianus and Hyopotamus bovinus) discovered by the Marchioness of Hastings in the Eocene deposits on the NW coast of the Isle of Wight: with an attempt to develop Cuvier’s idea of the classification of pachyderms by the number of their toes. Quarterly Journal of the Geological Society of London 4: 103–141. Crossref
Pagnac, D.C. and Reynolds, R.E. 2010. A juvenile specimen of Archaeohippus mourningi (Perissodactyla: Equidae) from the Cajon Valley Formation (middle Miocene) of California. In: 2010 Desert Symposium Volume, California State University, Desert Studies Consortium, 241–247. California State University, Fullerton.
Patton, T.H. 1969. Miocene and Pliocene artiodactyls, Texas Gulf Coastal Plain. Bulletin of the Florida State Museum Biological Sciences 14: 115–226.
Prothero, D.R. 1998. Rhinocerotidae. In: C.M. Janis, K.M. Scott, and L.L. Jacobs (eds.), Evolution of Tertiary Mammals of North America. Vol. 1, Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals, 595–605. Cambridge University Press, Cambridge. Crossref
Prothero, D.R. 2005. The Evolution of North American Rhinoceroses. 218 pp. Cambridge University Press, Cambridge.
Prothero, D.R. 2008. Systematics of the musk deer (Artiodactyla: Moschidae: Blastomerycinae) from the Miocene of North America. New Mexico Museum of Natural History and Science Bulletin 44: 207–224.
Prothero, D.R. and Liter, M.R. 2007. Family Palaeomerycidae. In: D.R. Prothero and S.E. Foss (eds.), The Evolution of Artiodactyls, 241–248. Johns Hopkins University Press, Baltimore.
Prothero, D.R. and Liter, M.R. 2008. Systematics of the Dromomerycines and Aletomerycines (Artiodactyla: Palaeomerycidae) from the Miocene and Pliocene of North America. New Mexico Museum of Natural History and Science Bulletin 44: 273–298.
Prothero, D.R. and Sereno, P.C. 1980. Allometry and paleoecology of medial Miocene dwarf rhinoceroses from the Texas Gulf Coastal Plain. Paleobiology 8: 16–30. Crossref
Prothero, D.R., Campbell, K.E., Jr., Beatty, B.L., and Frailey, C.D. 2014. New late Miocene dromomerycine artiodactyl from the Amazon Basin: implications for interchange dynamics. Journal of Paleontology 88: 434–443. Crossref
Prothero, D.R., Davis, E.B., and Hopkins, S.S.B. 2008. Magnetic stratigraphy of the Massacre Lake Beds (Late Hemingfordian, Early Miocene), northwest Nevada and the age of the “proboscidean datum” in North America. New Mexico Museum of Natural History and Science Bulletin 44: 239–246.
Savage, D.E. and Russell, D.E. 1983. Mammalian Paleofaunas of the World. 432 pp. Addison-Wesley, Reading.
Schultz, C.B. and Falkenbach, C.H. 1941. Ticholeptinae a new subfamily of oreodonts. Bulletin of the American Museum of Natural History 79: 1–105.
Semprebon, G.M., Rivals, F., and Janis, C.M. 2019. The role of grass vs. exogenous abrasives in the paleodietary patterns of North American ungulates. Frontiers in Ecology and Evolution 7 (65): 1–23. Crossref
Semprebon, G.M., Sise, P.J., and Coombs, M.C. 2011. Potential bark and fruit browsing as revealed by stereomicrowear analysis of the peculiar clawed herbivores known as chalicotheres. Journal of Mammalian Evolution 18: 33–55. Crossref
Shotwell, J.A. 1968. Miocene Mammals of southeast Oregon. Bulletin of the Museum of Natural History, University of Oregon 14: 1–67.
Skinner, M.F., Skinner, S.M., and Gooris, R.J. 1977. Stratigraphy and biostratigraphy of late Cenozoic deposits in central Sioux County, western Nebraska. Bulletin of the American Museum of Natural History 158: art. 5.
Smith, K.S. 2002. Mammalian Paleontology of the Monarch Mill Formation at Eastgate, Churchill County, Nevada. 711 pp. Ph.D. Dissertation, University of Oklahoma, Norman.
Smith, K.S., Cifelli, R.L., and Czaplewski, N.J. 2006. A new genus of eomyid rodent from the Miocene of Nevada. Acta Palaeontologica Polonica 51: 385–392.
Smith, K.S., Czaplewski, N.J., and Cifelli, R.L. 2016. Middle Miocene carnivorans from the Monarch Mill Formation, Nevada. Acta Palaeontologica Polonica 61: 231–252.
Steinthorsdottir, M., Coxall, H.K., de Boer, A.M., Huber, M., Barbolini, N., Bradshaw, C.D., Burls, N.J., Feakins, S.J., Gasson, E., Henderiks, J., Holbourn, A.E., Kiel, S., Kohn, M.J., Knorr, G., Kürschner, W.M., Lear, C.H., Liebrand, D., Lunt, D.J., Mörs, T., Pearson, P.N., Pound, M.J., Stoll, H., and Strömberg, C.A.E. 2020. The Miocene: the future of the past. Paleoceanography and Paleoclimatology 36: e2020PA004037. Crossref
Stewart, J.H., Sarna-Wojcicki, A.M., Meyer, C.E., and Wan, E. 1999. Stratigraphy, tephrochronology, and structural setting of Miocene sedimentary rocks in the Cobble Cuesta area, west-central Nevada (Report 99-352). 21 pp. U.S. Geological Survey, Menlo Park. Crossref
Stirton, R.A. 1940. The Nevada Miocene and Pliocene mammalian faunas as faunal units. Proceedings of the Sixth Pacific Science Congress 2: 627–640.
Storer, C.C. 1975. Tertiary mammals of the Saskatchewan. Part III: The Miocene fauna. Life Sciences Contributions, Royal Ontario Museum 103: 1–134.
Storer, C.C. 1978. Tertiary sands and gravels in Saskatchewan and Alberta; Correlation of mammalian faunas. In: C.R. Steleck and Challerton, B.D.E. (eds.), Western and Arctic Canadian Biostratigraphy. Geological Association of Canada Special Paper 18: 595–602.
Storer, C.C. 1993. Additions to the mammalian paleofauna of Saskatchewan. Modern Geology 18: 475–487.
Swisher, C.C. 1992. 40Ar/39Ar Dating and Its Application to the Calibration of the North American Land Mammal Ages. 239 pp. Ph.D. Dissertation, University of California, Berkeley.
Tedford, R.H., Albright III, L.B., Barnosky, A.D., Ferrusquía-Villafranca, I., Hunt Jr., R.M., Storer, J.E., Swisher III, C.C., Voorhies, M.R., Webb, S.D., and Whistler, D.P. 2004. Mammalian biochronology of the Arikareean through Hemphillian interval (late Oligocene through early Pliocene epochs). In: M.O. Woodburne (ed.), Late Cretaceous and Cenozoic Mammals of North America: Biostratigraphy and Geochronology, 169–231. Columbia University Press, New York. Crossref
Tedford, R.H., Skinner, F.F., Fields, R.W., Rensberger, J.M. Whistler, D.P. Galusha, T., Taylor, B.E., MacDonald, J.R., and Webb, S.D. 1987. Faunal Succession and Biochronology of the Arikareean through Hemphillian Interval (late Oligocene through earliest Pliocene epochs) in North America. In: M.O. Woodburne (ed.), Cenozoic Mammals of North America: Geochronology and Biostratigraphy, 153–210. University of California Press, Berkeley.
Thorpe, M.R. 1923. The primitive and carnivore-like characteristics of the Merycoidodontidae. American Journal of Science, Ser. 5–6 33: 239–246. Crossref
Voorhies, M.R. and Thomasson, J.R. 1979. Fossil grass anthoecia within Miocene rhinoceros skeletons: diet in an extinct species. Science 206: 331–333. Crossref
Webb, S.D. 1977. A history of savanna vertebrates in the New World. Part 1: North America. Annual Reviews of Ecology and Systematics 8: 355–380. Crossref
Webb, S.D. 1998. Hornless ruminants. In: C.M. Janis, K.M. Scott, and L.L. Jacobs (eds.), Evolution of Tertiary Mammals of North America. Vol. 1, Terrestrial Carnivores, Ungulates, and Ungulate-like Mammals, 463–476. Cambridge University Press, Cambridge. Crossref
Webb, S.D. 2008. Revision of the extinct Pseudoceratinae (Artiodactyla: Ruminantia: Gelocidae). Bulletin of the Florida Museum of Natural History 48(2): 17–58.
Westerhold, T., Marwan, N., Drury, A.J., Liebrand, D., Agnini, C., Anagnostou, E., Barnet, J.S.K., Bohaty, S.M., De Vleeschouwer, D., Florindo, F., Frederichs, T., Hodell, D.A., Hollbourn, A.E., Kroon, D., Lauretano, V., Littler, K., Lourens, L.J., Lyle, M., Pälike, H., Röhl, U., Tian, J., Wilkens, R.H., Wilson, P.A., and Zachos, J.C. 2020. An astronomically dated record of Earth’s climate and its predictability over the last 66 million years. Science 369: 1383–1387. Crossref
Wilson, J.A. 1960. Miocene carnivores, Texas coastal plain. Journal of Paleontology 34: 983–1000.
Wolfe, J.A., Schorn, H.E., Forest, C.E., and Molnar, P. 1997. Paleobotanical evidence for high altitudes in Nevada during the Miocene. Science 276: 1672–1675. Crossref
Woodburne, M.O. 2004. Global events and the North American mammalian biochronology. In: M.O. Woodburne (ed.), Late Cretaceous and Cenozoic Mammals of North America: Biostratigraphy and Geochronology, 315–343. Columbia University Press, New York. Crossref
Woodburne, M.O. 2006. Mammal ages. Stratigraphy 3: 229–261.
Woodburne, M.O. and Swisher, C.C. 1995. Land mammal high-resolution geochronology, intercontinental overland dispersals, sea level, climate, and vicariance. In: W.A. Berggren, D.V. Kent, M.P. Aubry, and J. Hardenbol (eds.), Geochronology, Time Scales and Global Stratigraphic Correlation. Society for Sedimentary Geology Special Publication 54: 335–364. Crossref
Zachos, J.C., Dickens, G.R., and Zeebe, R.E. 2008. An early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature 451: 279–283. Crossref
Zapfe, H. 1979. Chalicotherium grande (Blainv.) aus der miozänen Spaltenfüllung von Neudorf an der March (Devinská Nová Ves), Tschechoslowakei. Neue Denksschriften des Naturhistorischen Museums in Wien 2: 1–282.
Acta Palaeontol. Pol. 67 (1): 239–256, 2022
https://doi.org/10.4202/app.00907.2021