
Redescription of the type specimens for the Late Jurassic rhynchocephalian Opisthias rarus and a new specimen of Theretairus antiquus from Quarry 9, Morrison Formation, Wyoming, USA
JORGE A. HERRERA-FLORES, THOMAS L. STUBBS, and FRANCISCO SOUR-TOVAR
Herrera-Flores, J.A., Stubbs, T.L., and Sour-Tovar, F. 2022. Redescription of the type specimens for the Late Jurassic rhynchocephalian Opisthias rarus and a new specimen of Theretairus antiquus from Quarry 9, Morrison Formation, Wyoming, USA. Acta Palaeontologica Polonica 67 (3): 623–630.
We reexamine and redescribe the type specimens and other associated material of the Late Jurassic rhynchocephalian Opisthias rarus, from Quarry 9 of the Morrison Formation. We rediscover and describe a fragment of rock matrix belonging to the holotype that is presented for the first time, and we also comment on undescribed material from Quarry 9 that possibly belongs to an unnamed species of Opisthias. In addition, we describe a new specimen of Theretairus antiquus that helps to reject its proposed status as a junior synonym of O. rarus, and contributes to increase the knowledge of the microvertebrate fauna of the Late Jurassic of North America.
Key words: Lepidosauria, Rhynchocephalia, Morrison Formation, Jurassic, Wyoming, USA.
Jorge A. Herrera-Flores [jorge.herrera@ciencias.unam.mx] and Francisco Sour-Tovar [fcosour@ciencias.unam.mx], Museo de Paleontología, Facultad de Ciencias, Universidad Nacional Autónoma de México, Ciudad Universitaria, 04510, Ciudad de México, Mexico.
Thomas L. Stubbs [tom.stubbs@bristol.ac.uk], School of Earth Sciences, University of Bristol, Queens Road, Bristol, BS8 1RJ, UK.
Received 24 July 2021, accepted 30 December 2021, available online 15 June 2022.
Copyright © 2022 J.A. Herrera-Flores et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Rhynchocephalia is a group of reptiles that were very successful during most of the Mesozoic, and their origins can be traced back to the Early or Middle Triassic (Jones et al. 2013). It is well known that rhynchocephalians reached their highest taxonomic diversity by the Jurassic and then declined through the Cretaceous to their near extinction by the early Paleocene (Apesteguía et al. 2014). Before this decline, rhynchocephalians showed very high morphological disparity that allowed them to adapt to a great variety of ecological niches and diets (Jones 2008; Meloro and Jones 2012; Herrera-Flores et al. 2017). Between the Late Triassic and Late Jurassic, rhynchocephalians were a very important component of the terrestrial vertebrate faunas, being especially abundant and diverse if compared with other small vertebrates such as lizards and early mammals.
The Morrison Formation includes many of the richest fossil sites from the Late Jurassic of North America. For over a hundred years, the Morrison Formation has provided a great number of fossil vertebrates, mostly represented by terrestrial macrovertebrates such as crocodylomorphs and dinosaurs (Foster 2003). On the other hand, the microvertebrate fauna of Morrison Formation is primarily composed by amphibians, basal mammaliforms, turtles, and lepidosaurs. Among the Late Jurassic lepidosaurs of North America, lizards and snakes have a relatively good but fragmentary fossil record (Evans and Chure 1998, 1999; Nydam 2013). In contrast, rhynchocephalians are only represented by three species: Opisthias rarus, Theretairus antiquus, and Eilenodon robustus. Opisthias rarus was the first rhynchocephalian described from the Late Jurassic Morrison Formation about a century ago, while E. robustus was the latest one described, four decades ago (Gilmore 1909; Simpson 1926; Rasmussen and Callison 1981). It is notable that Opisthias is a genus that is frequently cited in several fossil sites of the Late Jurassic of North America (e.g., Foster 2003), but also includes records from the Late Jurassic of Portugal (Ortega et al. 2009), and the Early Cretaceous of England (Evans and Fraser 1992); however, the taxonomy and actual diversity of the genus is poorly understood. For a long time, the genus Opisthias has been recognized as monospecific, but recent research suggests that fossil material previously referred to this taxon could represent a number of undescribed species (e.g., Maldonado et al. 2013; Maldonado and Bertog 2015; Herrera-Flores and Stubbs 2018). Indeed, recent research has acknowledged that an Opisthias specimen from England is a different species to O. rarus (Herrera-Flores 2020); therefore, all other available material of Opisthias must be reassessed, including a complete redescription and rediagnosis of the type specimens.
We present herein a redescription of the type specimens of O. rarus and provide an emended diagnosis for the species. We comment on undescribed material of O. rarus from Quarry 9 of the Morrison Formation, including a specimen with a clearly distinctive tooth morphology that might represent an unnamed species. Additionally, we describe a new specimen of the enigmatic sphenodontid Theretairus antiquus that helps to confirm its taxonomical validity.
Institutional abbreviations.—DINO, Dinosaur National Monument, Colorado, USA; DORCM, Dorset County Museum, Dorset, UK; USNM, National Museum of Natural History, Smithsonian Institution, Washington D.C., USA; YPM, Yale Peabody Museum of Natural History, New Haven, USA.
Material and methods
The studied fossils used in this research were collected between 1883 and 1888 at Quarry 9, Como Bluff, Wyoming by fossil diggers Edward Kennedy, Fred Brown, and W. Beck. All fossil material is housed in the paleontological collections at the National Museum of Natural History, Smithsonian Institution and was studied with the aid of a stereoscopic microscope. All measurements are expressed in millimeters and were taken with a digital caliper. For further anatomical comparisons, we examined the Early Cretaceous specimen of Opisthias (DORCM G. 10831) from the collection of Dorset County Museum, UK.
Systematic palaeontology
Order Rhynchocephalia Günther, 1867
Suborder Sphenodontia Williston, 1925
Infraorder Eusphenodontia Herrera-Flores, Stubbs, Elsler, and Benton, 2018
Clade Neosphenodontia Herrera-Flores, Stubbs, Elsler, and Benton, 2018
Clade Opisthodontia Apesteguía and Novas, 2003
Genus Opisthias Gilmore, 1909
Type species: Opisthias rarus (Gilmore, 1909), Upper Jurassic, Quarry 9, Como Bluff, Wyoming, USA.
Emended diagnosis.—Taxon with moderate body size, a deep, robust and slightly curved mandible and high and well-developed coronoid process. Robust dentary dorsoventrally curved with acrodont teeth implantation and three generations of teeth (an enlarged successional tooth, hatchling and additional teeth). Additional teeth with a distinctly pyramidal shape, slightly curved with a marked quadrangular shape in occlusal view. Dentary with a broad symphysis and with a well-developed pointed and curved ventral projection. Maxillary teeth conical, slightly curved and strongly striated.
Opisthias rarus Gilmore, 1909
Fig. 1.
Type material: Holotype: USNM 2860, nearly complete left dentary (Fig. 1A, C, D). Paratype: USNM 2858, partial left dentary (Fig. 1B), from the type locality and horizon.
Type locality: Quarry 9, Como Bluff, Wyoming, USA.
Type horizon: Upper Kimmeridgian/lower Tithonian (Upper Jurassic).
Material.—Type material and USNM 6126–6132, USNM 26077–26089, USNM 508532 (see comments below on the true taxonomic identity of some of these fossils).
Emended diagnosis.—Species with a high coronoid process with regard to dentary height, with a pronounced triangle-like shape. Dentary notably tall and about three or four times the height of the additional teeth above it. Additional teeth of the dentary slightly curved, strongly striated with a distinguishable groove on the labial and lingual surfaces, the groove extends along the height of the crown and are more marked in labial view. Additional teeth in lingual view present rounded lateral flanges. Additional teeth of maxilla, conical, curved and notably striated.
Description.—The holotype (USNM 2860) of Opisthias rarus is a nearly complete left dentary from an adult individual (Fig. 1A). The coronoid process is broken, but the preserved fragment is high, suggesting that the complete shape of the coronoid process is tall, likely similar to a triangle (Fig. 1A1, A2). In labial view the dentary shows a marked ridge formed by a secondary bony skirt. Extending below the ridge, there is a series of six or seven small mental foramina. In lingual view, the Meckelian groove extends along the midline of the jaw and, as in other derived sphenodontians, the Meckelian groove is open and deep, considerably wider at the level of the coronoid process, but it narrows gradually until the posterior part of the jaw (Fig. 1A2). The mandibular symphysis is wide with a marked mentonian process, pointy and slightly curved posteriorly. The dentary includes three series of teeth, most of them relatively well preserved but some are broken or heavily worn. There are twelve additional teeth; almost all of them show notable signs of wear on the top of the dental crown, especially the two mesialmost teeth that are clearly damaged. The shape of additional teeth is pyramidal, slightly curved and strongly striated. In labial view, the additional teeth show a distinguishable deep groove that extends along the height of the crown (Fig. 1A4). In lingual view, smaller groves are also present, but the teeth also show strong and rounded lateral flanges (Fig. 1A5). Hatchling teeth are completely worn away, giving the impression of an edentulous zone between the additional teeth and the enlarged successional tooth. The enlarged successional tooth is broken at the base of the tooth crown, leaving the tooth neck exposed which has a clearly cylindrical shape (Fig. 1A6).
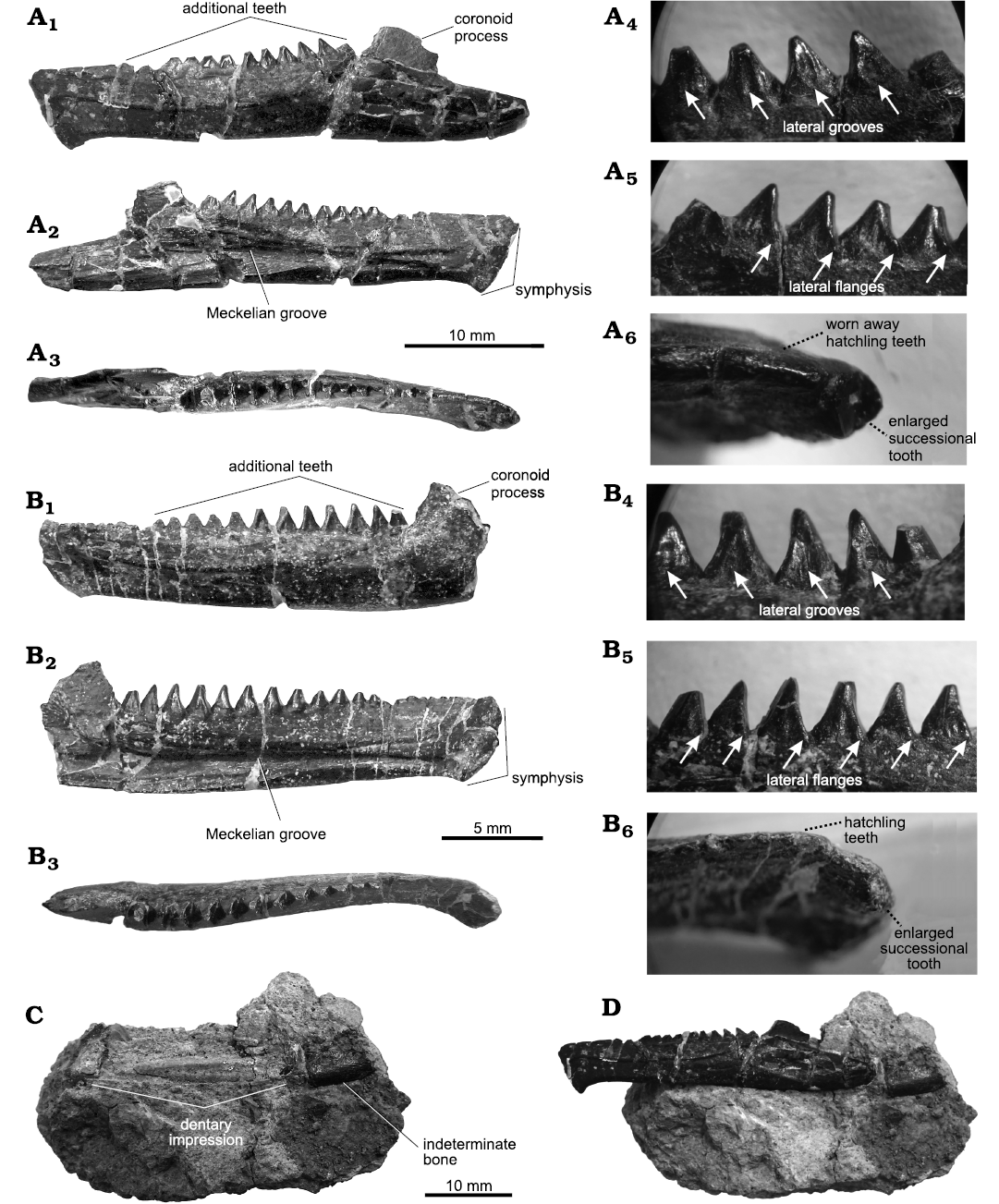
Fig. 1. Teeth in the type specimens of the rhynchocephalian Opisthias rarus Gilmore, 1909, Quarry 9, Como Bluff, Wyoming, USA, Upper Jurassic. A. USNM 2860 (holotype), nearly complete left dentary, in labial (A1, A4), lingual (A2, A5), and occlusal (A3, A6) views. B. USNM 2858 (paratype), partial left dentary, in labial (B1, B4), lingual (B2, B5), and occlusal (B3, B6) views. C. Fragment of rock matrix belonging to the holotype. D. Holotype when it is superimposed to the rock matrix.
When reviewing fossil material of rhynchocephalians from the collection of the Smithsonian Institution, we found an unnumbered specimen which consists of a fragment of rock matrix, that includes a very well-preserved impression of a left dentary and a small fragment of bone attached (Fig. 1C). The dentary impression has a very similar form to the holotype of O. rarus, and also fits perfectly when the holotype is superimposed to the matrix impression. Also, the small bone attached to the rock is relatively large and it seems to be the posterior process of another dentary (Fig. 1D). We reviewed the accession notes from the collections, and they indicate that the holotype of O. rarus also has an impression in rock matrix. It must be noted that Gilmore (1909) vaguely commented on an impression in rock matrix, however, this element is not illustrated or photographed in the original description. Thus, this is the first time the rock matrix is presented and described. The length of the holotype as preserved is 35 mm.
The paratype (USNM 2858) of O. rarus is a left partial dentary fragment of a young adult individual (Fig. 1B1, B3, B4, B6). The bone is broken behind the coronoid process, thus, the complete dentary posterior process is lost. The coronoid process is broken dorsally and posteriorly, but it shows similar morphology as the holotype. The medial and lateral sides of dentary have the same characteristics of the type specimen with an open and deep Meckelian groove that narrows close to the mandibular symphysis, and a marked ridge on the labial side formed by the secondary bone skirt (Fig. 1B1, B2). The mandibular symphysis is wide, and with a pointy and curved ventral projection. The dentary includes three generations of teeth, there are twelve additional teeth which do not show signs of heavy wear, but most of them are broken at the top of the crown. The additional teeth show deep and well-marked grooves on their labial and lingual surfaces similar to those of the type specimen (Fig. 1B4). Mesiolingual flanges are also visible on the lingual surface of the additional teeth; however, these flanges are smaller than those showed by the type specimen (Fig. 1B5). There are eight very small and heavily worn hatchling teeth; the enlarged successional tooth is broken but apparent remains of the base of the dental crown are visible in occlusal and lingual views (Fig. 1B6). Length as preserved for the paratype is 22.3 mm.
Additionally, there are twenty two specimens assigned to O. rarus (USNM 6126–6132, 7767, 26077–26089, 508532) that were not described by Gilmore (1909) or other authors. It must be noted that these additional specimens come from the type locality of O. rarus, and all of them were also collected by Kennedy and colleagues between 1883 and 1888. Most of these specimens are very fragmentary, including fragments of dentaries, a few maxillae and other bone fragments, some of them embedded in rock matrix. Among the undescribed O. rarus material from Quarry 9, there are some fossils that cannot be referred to Opisthias or other rhynchocephalians, but they may belong to other small-sized reptiles such as Cteniogenys, squamates or turtles (e.g., USNM 6128, rock matrix with a fragment of a right dentary and a small fragment of maxilla; USNM 26081, a possibly lumbar vertebrae; USNM 26082, right dentary of an indeterminate small vertebrate; USNM 26085, semi-complete left scapula of an indeterminate small vertebrate). Therefore, we only comment on those materials we are confident that belong to rhynchocephalians and that we believe would contribute to the understanding of Opisthias morphology. USNM 6126 is a fragment of the posterior part of a right dentary from an adult individual with two fragmentary additional teeth (Fig. 2A). This fragment has a complete and well-preserved posterior process of dentary; the coronoid process is incomplete, lacking portions of its dorsal and posterior margins. The relative completeness of the coronoid process makes it even clearer that the shape of this element is quite similar to a triangle as was noted for the holotype and paratype. The multiple specimens cataloged under USNM 26080 include several fragmentary maxillae, dentaries and a palatine with teeth. The best-preserved fragment is a small bone that includes four palatine teeth (Fig. 2B). Three of the preserved teeth are broken, but one is complete and well-preserved. This tooth is conical, very pointed, slightly curved with some striations. USNM 508532 is a rock fragment that includes two partial maxillae and a small indeterminate bone (Fig. 2C1). The largest visible element is a right maxilla, the element is robust with a high frontal process and a relatively short and broad posterior process (Fig. 2C2), the overall shape of this element is quite similar to specimen DINO 15911, a right maxilla referred to Opisthias by Fraser and Wu (1998: fig. 2) from the Dinosaur National Monument, Brushy Basin member of the Morrison Formation, Colorado, USA. The hatchling teeth on the larger preserved piece of the maxilla are difficult to observe, but the two additional teeth are well preserved, both are conical and curved with apparent striations (Fig. 2C2). The second fragment of maxilla is fixed in the rock matrix, however, teeth of this element are clearly exposed (Fig. 2C3). Three small and conical hatchling teeth are preserved along with two complete and well preserved additional teeth, which are robust, conical, slightly curved and highly striated.

Fig. 2. Teeth of rhynchocephalians Opisthias sp. (A–D) and Theretairus antiquus Simpson, 1926 (E) from Quarry 9, Como Bluff, Wyoming, USA, Upper Jurassic. A. USNM 6126, right dentary fragment with a relatively complete coronoid process in labial view. B. USNM 26080, fragment of palatine in ?lingual view with four teeth attached. C. USNM 508532, rock matrix with fragments of right maxilla in lingual view (C1), close-up detail (C2), maxillary teeth (C3). D. USNM 7767 and USNM 26086, fragments of a right dentary, in lingual (D1), labial (D2), and occlusal (D3) views, close-up detail of the four distalmost additional teeth (D3), close-up detail of the four distalmost additional teeth in labial (D4) and lingual (D5) views. E. USNM 26088, anterior fragment of a left dentary, in labial (E1), lingual (E2), and occlusal (E3) views.
Among the undescribed rhynchocephalian material of Quarry 9, we found a couple of specimens that clearly do not belong to O. rarus. The specimens USNM 7767 and USNM 26086 are dentary fragments with clearly distinguished tooth morphology that refers to a possibly distinct species of Opisthias. A detailed description of both specimens is given here below.
Stratigraphic and geographic range.—Upper Kimmeridgian–lower Tithonian (Upper Jurassic), Quarry 9, Como Bluff, Wyoming, USA.
Opisthias sp.
Fig. 2D.
Material.—USNM 7767, USNM 26086, a fragment of a right dentary with eight additional teeth from Quarry 9, Como Bluff, Wyoming, USA; upper Kimmeridgian/lower Tithonian (Upper Jurassic).
Description.—USNM 7767 and USNM 26086 are broken fragments of a right dentary that were cataloged as separate specimens but clearly belong to a single individual, the preserved specimen lacks the coronoid and symphyseal regions (Fig. 2D1, D2). This fragment belongs to an adult individual and includes eight additional teeth; the six distalmost teeth are complete and well-preserved with the distinctive pyramidal shape of Opisthias. The preserved region shows, in lingual view, an open and broad Meckelian groove that narrows interiorly, very similar to that of O. rarus but slightly deeper. The labial surface of the dentary presents a notably marked ridge formed by a secondary bony skirt and a margin formed by the depth of the Meckelian groove. Additional teeth of the dentary show very light signs of wear; all teeth are closely packed and in labial view, teeth are flat and smooth without marked striations or grooves (Fig. 2D4). In lingual view, the six distalmost teeth are robust, rounded and smooth without marked striations and lateral flanges (Fig. 2D5).
Family Sphenodontidae Cope, 1871
Subfamily Sphenodontinae Cope, 1871
Genus Theretairus Simpson, 1926
Type species: Theretairus antiquus (Simpson, 1926), Quarry 9, Como Bluff, Wyoming, USA, Upper Jurassic.
Theretairus antiquus Simpson, 1926
Fig. 2E.
Material.—USNM 26088, a fragment of a left dentary with remains of two caniniform teeth, one of them complete from Quarry 9, Como Bluff, Wyoming, USA; upper Kimmeridgian/lower Tithonian (Upper Jurassic).
Description.—USNM 26088 is a fragment of the symphyseal region of a left dentary, that preserves remains of two large caniniform teeth and four or five smaller alternate hatchling teeth. The mesialmost caniniform tooth is broken at the base of the tooth crown, but remains of the tooth neck are well preserved and show an oval and slightly curved shape (Fig. 2E). The distalmost caniniform is complete and with a marked conical blade-like shape; it is a high, pointy, slightly striated and notably curved element, with a curvature of about 70°. Serrations on the mesial and distal cutting edges of the caniniform tooth are not present. Alternate teeth are small, conical and all of them are broken at the top of the crown. The mandibular symphysis is robust, curved and with a marked ventral projection, pointy and curved. In labial view, five mental foramens are present, the three foramens closer to the mandibular symphysis are small and rounded, and the other two are larger and oval. In lingual view, the Meckelian groove is open and notably wide. Length as preserved is 5.7 mm.
Stratigraphic and geographic range.—Upper Kimmeridgian–lower Tithonian (Upper Jurassic), Quarry 9, Como Bluff, Wyoming, USA
Discussion
Since originally described, Opisthias rarus has been an enigmatic and problematic taxon, mainly because of its poorly known taxonomy. Gilmore’s (1909) original work offered a good description of the holotype, however, the description of the paratype is vague, and all the additional material collected by Kennedy and colleagues was not even mentioned, excluding important information of the jaw, maxillae, and palatine. Additionally, the pictures and drawings of the holotype, while outstanding for the time, do not allow detailed observations for some important aspects of the anatomy for the species. Another situation that led to confusion about the taxonomy of O. rarus, was the description of a smaller rhynchocephalian which also comes from Quarry 9, the same type locality of O. rarus. This species was named Theretairus antiquus by Simpson (1926). In his work Simpson (1926) frequently compared T. antiquus with O. rarus, finding many similarities between both; though, one of the main differences between the dentaries of O. rarus and T. antiquus is the presence of two caniniforms and alternate teeth in the latter. Additionally, Simpson (1926) described two specimens of a dentary and maxilla that he referred to O. rarus, however, the description of those materials did not really improve the knowledge about the species, and only created more misconceptions that led other authors to suggest that T. antiquus was a juvenile of O. rarus (e.g., Hoffstetter 1953). A much later work on rhynchocephalians from Morrison Formation, Fraser and Wu (1998) described new specimens that include cranial and postcranial material collected at Dinosaur National Monument. In their work, Fraser and Wu (1998) discus the difficulties in referring the fossil material to O. rarus, or another species, due to the incompleteness of the type specimens and the limitations that the original description of O. rarus has, and the lack of other taxonomical studies that provide clearer insights on the taxonomy of the species. However, even if the authors had difficulties accurately assigning the specimens to a species of Opisthias, or any other genus (e.g., Homoeosaurus), they discarded the idea of T. antiquus as a junior synonym of O. rarus.
When studying the additional material of Opisthias from the Smithsonian Institution, we immediately noted that specimen USNM 26088 was clearly distinct from the other specimens referred to O. rarus (Fig. 2E). A closer examination of the specimen allowed us to observe that it has two relatively large caniniform teeth close the border of the mandibular symphysis, the more mesial one is broken at the base but the more distal tooth is complete, also the specimen has alternate teeth on the rest of the preserved dentary, these characters lead us to refer specimen USNM 26088 to T. antiquus. As far as we know, USNM 26088 would represent the third known and the second formally described specimen of T. antiquus, considering that in his review of the vertebrate fauna of the Morrison Formation, Foster (2003: table 5) mentioned the existence of another specimen from Quarry 9 apart of the holotype (YPM 13764), however, to our knowledge, that supposed additional specimen of T. antiquus has never been described and it is unknown where is it housed. On the other hand, our study of USNM 26088 gives us enough confidence to reject the old hypothesis that T. antiquus might be a junior synonym of O. rarus, allowing us to confirm that in the Upper Jurassic Morrison Formation at least three well distinguishable genera coexisted, namely Opisthias, Eilenodon, and Theretairus. It must be noted that the presence of three highly derived taxa indicates a remarkable ecomorphological diversification in the rhynchocephalians of the Morrison Formation, considering that Opisthias and Eilenodon belong to the Opisthodontia, a group of specialized herbivorous distinguished by the complexity of their dentition (Apesteguía and Novas 2003; Jones et al. 2018; LeBlanc et al. 2020), while Theretairus is likely an insectivorous or carnivorous taxon in view of the blade-like shape of its two caniniforms (Jones 2006, 2009; Rauhut et al. 2012; Martínez et al. 2013). Additionally to their differences in diet, their body size disparity suggest that each rhynchocephalian species of the Morrison Formation was probably adapted to a particular ecological niche, taking into account that Opisthias and Eilenodon had a moderate and large body size respectively, while Theretairus was a considerably smaller taxon; also, the ecological interactions of the rhynchocephalians from the Morrison Formation with squamates and other small vertebrates is a topic that still needs to be carefully explored. Conversely, in recent decades, several new specimens of Opisthias sp. have been collected from many localities of the Morrison Formation and Europe (Evans and Fraser 1992; Fraser and Wu 1998; Ortega et al. 2009; Maldonado et al. 2013), however, the poorly known taxonomy and the lack of a clear diagnosis of O. rarus did not allow unmistakably referral any of those new materials to O. rarus or a new species.
For over one hundred years, the genus Opisthias has been recognized as monospecific, but its underestimated diversity has been a subject of discussion until very recently (Maldonado et al. 2013; Maldonado and Bertog 2015; Herrera-Flores and Stubbs 2018; Herrera-Flores 2020). For example, Maldonado et al. (2013) and Maldonado and Bertog (2015) report dentaries and maxillae of Opisthias sp. from the Aaron Scott Quarry of Central Utah that present notable differences in dentary shape and tooth morphology. This suggests that there is at least one additional species distinct to Opisthias rarus from the Morrison Formation. Unfortunately, a detailed description of these specimens and further discussion about the morphological differences between those specimens from the Aaron Scott Quarry and the type specimens of O. rarus were never published. More recently, Herrera-Flores and Stubbs (2018) and Herrera-Flores (2020), reexamined the O. cf. rarus specimen from the Purbeck Limestone of Dorset (DORCM G. 10831). In their work, the authors suggest that DORCM G. 10831 has clear morphological differences when compared to the holotype and paratype of O. rarus, discarding the notion that the Dorset specimen is identical to O. rarus, as was formerly proposed by Evans and Fraser (1992). When comparing DORCM G. 10831 to the O. rarus specimens from the Smithsonian Institution, we noted that tooth morphology of additional teeth in the Dorset specimen is remarkably different. Additional teeth in DORCM G. 10831 do not show the same grooves and lateral flanges that are present in O. rarus in lingual view, also hatchling teeth are broader. Regarding specimens USNM 7767 and USNM 26086, referred here as Opisthias sp., it is evident that the additional teeth of dentary are more closely packed than in O. rarus (Fig. 2D), a condition that was also noted in the unnamed species reported by Maldonado et al. (2013). Of course, we are aware that the presence of more closely packed additional teeth in USNM 7767 and USNM 26086 do not necessary imply that it is a different species than O. rarus, and that it could be merely an interesting case of intraspecific variation. Nevertheless, the lack of marked grooves, lateral flanges and strong striations in the additional teeth of the specimens (Fig. 2D4, D5), suggest us that it could be a distinct species. We consider that it is still necessary to review many more specimens of Opisthias from other Late Jurassic fossil sites from Morrison Formation, in order to improve the knowledge of the skull and skeleton, as well as its intraspecific variation to clarify if the diversity of the genus has been truly underestimated for the Late Jurassic of North America as has been recently suggested.
Concluding remarks
We consider the redescription of the type specimens of Opisthias rarus necessary as a first step to elucidate the real taxonomic diversity of the genus Opisthias, and current evidence suggests that it could be underestimated. However, more taxonomic work is still necessary for a better understating of this taxon and special attention should be taken on the skull and the rarely studied postcranium. The description of the new specimen of Theretairus antiquus allows us to confirm the validity of the species, but also contributes to our knowledge of the Late Jurassic microvertebrate fauna of North America.
Acknowledgements
We thank Amanda Millhouse (USNM) and Paul Tomlinson (DORCM) for the access provided to the specimens used for this study. We thank Annie Schmaltz-Hsiou (Universidade de Sao Paulo, Brasil) and Hans-Dieter Sues (USNM) for their comments on an earlier version of the manuscript. We thank Sebastián Apesteguía (Universidad Maimónides, Buenos Aires, Argentina) and an anonymous reviewer for their helpful comments to improve the manuscript. The research of JAH-F is funded by Beca Posdoctoral 2020 of the Dirección General de Asuntos de Personal Académico (DGAPA) of UNAM, and the Arthur J. Boucot Award 2018 from the Paleontological Society.
References
Apesteguía, S. and Novas, F.E. 2003. Large Cretaceous sphenodontian from Patagonia provides insight into lepidosaur evolution in Gondwana. Nature 425: 609–612. Crossref
Apesteguía, S., Gómez, R.O., and Rougier, G.W. 2014. The youngest South American rhynchocephalian, a survivor of the K/Pg extinction. Proceedings of the Royal Society B 281: 20140811. Crossref
Cope, E.D. 1871. On the homologies of some cranial bones of the Reptilia and the systematic arrangement of the class. Proceedings of the American Association for the Advancement of Science 19: 194–211.
Evans, S.E. and Chure, D.C. 1998. Paramacellodid lizard skulls from the Jurassic Morrison Formation at Dinosaur national monument, Utah. Journal of Vertebrate Paleontology 18: 99–114. Crossref
Evans, S.E. and Chure, D.J. 1999. Upper Jurassic lizards from the Morrison Formation of Dinosaur National Monument, Utah. Vertebrate Paleontology in Utah 99: 151. Crossref
Evans, S.E. and Fraser, N.C. 1992. A sphenodontid jaw (Reptilia: Lepidosauria) from the Upper Jurassic of Dorset. Proceedings of the Dorset Natural History Society 1992: 199–200.
Foster, J.R. 2003. Paleoecological analysis of the vertebrate fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, USA. New Mexico Museum of Natural History and Science Bulletin 23: 1–95.
Fraser, N.C. and Wu, X.C. 1998. Sphenodontians from the Brushy Basin Member of the Morrison Formation in Dinosaur National Monument. Modern Geology 23: 17–34.
Gilmore, C.W. 1909. A new rhynchocephalian reptile from the Jurassic of Wyoming, with notes on the fauna of Quarry 9. Proceedings of the US National Museum 37: 35–42. Crossref
Günther, A. 1867. Contribution to the anatomy of Hatteria (Rhynchocephalus, Owen). Philosophical Transactions of the Royal Society of London 157: 595–629. Crossref
Herrera-Flores, J.A. 2020. New insights on an enigmatic sphenodontian jaw from Dorset. Proceedings of the Dorset Natural History and Archaeological Society 141: 41–43.
Herrera-Flores, J.A. and Stubbs, T.L. 2018. A re-evaluation of the taxonomy and diversity of the Late Jurassic sphenodontian Opisthias rarus Gilmore 1909 (Lepidosauria: Rhynchocephalia). In: T. Martin, R. Schellhorn, and J.A. Schultz (eds.), 13th Symposium on Mesozoic Terrestrial Ecosystems and Biota, 42–44. GeoUnion Alfred-Wegener-Stiftung, Bonn.
Herrera‐Flores, J.A., Stubbs, T.L., and Benton, M.J. 2017. Macroevolutionary patterns in Rhynchocephalia: is the tuatara (Sphenodon punctatus) a living fossil? Palaeontology 60: 319–328. Crossref
Herrera-Flores, J.A., Stubbs, T.L., Elsler, A., and Benton, M.J. 2018. Taxonomic reassessment of Clevosaurus latidens Fraser, 1993 (Lepidosauria, Rhynchocephalia) and rhynchocephalian phylogeny based on parsimony and Bayesian inference. Journal of Paleontology 92: 734–742. Crossref
Hoffstetter, R. 1953. Caractères dentaires juvéniles chez des Rhynchocéphales Jurassiques, Theretairus et Leptosaurus. Bulletin du Museum National d’Histoire Naturelle 25: 213–220.
Jones, M.E.H. 2006. Tooth diversity and function in the Rhynchocephalia (Diapsida: Lepidosauria). In: P.M. Barrett and S.E. Evans (eds.), Ninth International Symposium on Mesozoic Terrestrial and Biota, 55–58. Natural History Museum, London.
Jones, M.E.H. 2008. Skull shape and feeding strategy in Sphenodon and other Rhynchocephalia (Diapsida: Lepidosauria). Journal of Morphology 269: 945–966. Crossref
Jones, M.E.H. 2009. Dentary tooth shape in Sphenodon and its fossil relatives (Diapsida: Lepidosauria: Rhynchocephalia). Comparative Dental Morphology 13: 9–15. Crossref
Jones, M.E.H., Anderson., C.L., Hipsley, C.A., Müller, J., Evans, S.E., and Schoch, R.R. 2013. Integration of molecules and new fossils supports a Triassic origin for Lepidosauria (lizards, snakes, and tuatara). BMC Evolutionary Biology 13: 208. Crossref
Jones, M.E.H, Lucas, P.W., Tucker, A.S., Watson, A.P., Sertich, J.J.W., Foster, J.R., Williams, R., Garbe, U., Bevitt, J.J., and Salvemini, F. 2018. Neutron scanning reveals unexpected complexity in the enamel thickness of an herbivorous Jurassic reptile. Journal of The Royal Society Interface 15: 20180039. Crossref
LeBlanc, A.R.H., Apesteguía, S., Larsson, H.C.E., and Caldwell, M.W. 2020. Unique tooth morphology and prismatic enamel in Late Cretaceous sphenodontians from Argentina. Current Biology 30: 1755–1761. Crossref
Maldonado, J. and Bertog, J. 2015. New species of Opisthias (sphenodontidae) from the Aaron Scott Quarry in the Brushy Basin Member of the Morrison Formation in Central Utah. Journal of Vertebrate Paleontology 35: 173.
Maldonado, J., Bertog, J., and Kintzel, E. 2013. Opisthias (Sphenodontia) From the Aaron Scott Quarry (Morrison Formation, Jurassic Period) of Central Utah. 62nd Annual Meeting of the Southeastern Section of the Geological Society America, Abstracts with Programs 45 (2): 64.
Martínez, R.N., Apaldetti, C., Colombi, C.E., Praderio, A., Fernandez, E., Malnis, P.S., Correa, G.A., Abelin, D., and Alcober, O. 2013. A new sphenodontian (Lepidosauria: Rhynchocephalia) from the Late Triassic of Argentina and the early origin of the herbivore opisthodontians. Proceedings of the Royal Society of London B 280: 20132057. Crossref
Meloro, C. and Jones, M.E.H. 2012. Tooth and cranial disparity in the fossil relatives of Sphenodon (Rhynchocephalia) dispute the persistent “living fossil” label. Journal of Evolutionary Biology 25: 2194–2209. Crossref
Nydam, R.L. 2013. Squamates from the Jurassic and Cretaceous of North America. Palaeobiodiversity and Palaeoenvironments 93: 535–565. Crossref
Ortega, F., Malafaia, E., Escaso, F., Pérez-García, A., and Dantas, P. 2009. Faunas de répteis do Jurássico Superior de Portugal. Paleolusitana 1: 43–56.
Rasmussen, T.E. and Callison, G. 1981. A new herbivorous sphenodontid (Rhynchocephalia: Reptilia) from the Jurassic of Colorado. Journal of Paleontology 55: 1109–1116.
Rauhut, O.W.M., Heyng, A.M., López-Arbarello, A., and Hecker, A. 2012. A new rhynchocephalian from the Late Jurassic of Germany with a dentition that is unique amongst tetrapods. PLoS One 7: e46839. Crossref
Simpson, G.G. 1926. American terrestrial Rhynchocephalia. American Journal of Science 67: 12–16. Crossref
Williston, S.W. 1925. Osteology of the Reptiles. Society for the Study of Amphibians and Reptiles. 300 pp. Harvard University Press, Cambridge. Crossref
Acta Palaeontol. Pol. 67 (3): 623–630, 2022
https://doi.org/10.4202/app.00929.2021