
A new euarthropod from the Cambrian Stage 4 Guanshan Biota of South China
DE-GUANG JIAO and KUN-SHENG DU
Jiao, D.-G. and Du, K.-S. 2022. A new euarthropod from the Cambrian Stage 4 Guanshan Biota of South China. Acta Palaeontologica Polonica 67 (4): 969–974.
A new small euarthropod Astutuscaris bispinifer gen. et sp. nov. is described from the early Cambrian Stage 4 Guanshan Biota in Yunnan, China. This new euarthropod possesses a wide head shield, a pair of possible eyes, paired frontalmost appendages located antero-medially, 11 imbricated tergites most of which have backward-directed tergopleura ending in almost posteriorly oriented spines, and two well-marked wide spines. The affinity of Astutuscaris among euarthropods is uncertain because of the undefined nature of its frontalmost appendages, the incomplete head shield, the anterior trunk tergites, and the limbs. There are about 24 species of non-trilobite euarthropods reported from the Guanshan Biota to date, the documentation of this new taxon expands the biological diversity of euarthropods from this important biota in Yunnan, China.
Key words: Arthropoda, evolution, Cambrian, Guanshan Biota.
De-Guang Jiao [dgjiao@yxnu.edu.cn] and Kun-Sheng Du [kunshengdu@qq.com] (corresponding author), Research Center of Paleobiology, Yuxi Normal University, 134 Phoenix Road, Yuxi, Yunnan, 653100, China; Institute of Paleontology, Yunnan University (Chenggong), Kunming, 650091, China.
Received 21 August 2021, accepted 16 June 2022, available online 21 October 2022.
Copyright © 2022 D.-G. Jiao and K.-S. Du. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The knowledge on Cambrian arthropods is essential for resolving the phylogenetic relationships and evolutionary history of extinct and extant arthropods (Yang et al. 2013; Daley et al. 2018; Giribet and Edgecombe 2019). Euarthropods are the most diverse group among the typical Burgess Shale-type Cambrian-aged Guanshan Biota (Hu et al. 2010, 2013). There are many euarthropods (e.g., trilobites, radiodonts, and bivalved euarthropods) described from this famous biota (Hu et al. 2010, 2013; Liu et al. 2012; Chen et al. 2019; Jiao et al. 2021a, b). Except the above mentioned groups there are only a few other taxa reported (Hu et al. 2010, 2013; Zhao et al. 2020; Jiao et al. 2021a, 2022), i.e., Guangweicaris spinatus, Panlongia tetranodusa, Longquania bispinosa, Leanchoilia sp., Sinoburius sp., the unnamed naraoiid, Alacaris? sp., Lihuacaris ferox, and Bailongia longicaudata. In this contribution we describe a new and rare euarthropod with an elongated exoskeleton from the Cambrian Stage 4 Guanshan Konservat-Lagerstätte.
Institutional abbreviations.—RCP, Research Center of Paleobiology, Yuxi Normal University, Yuxi, China.
Other abbreviations.—en, endopod; ex, exopod; ey, eye; fa, frontal appendage; gut, digestive tract; hs, head shield; lim, limb; p1–p5, podomeres 1–5; st, stalk; T1–T13, tergites 1–13; ts, terminal spine.
Nomenclatural acts.—This work and the nomenclatural acts it contains, have been registered in ZooBank: urn:lsid: zoobank.org:pub:7396B74C-AE35-450C-90FA-27A9A609 DAAD
Material and methods
The sole and articulated specimen (RCP 0002) of Astutuscaris bispinifer gen. sp. nov. and two specimens (RCP 0003 and RCP 0004) belonging to Guangweicaris spinatus were collected from Yiliang County which already provided exquisitely preserved fossils in the past (Jiao et al. 2016, 2021b). The fossils are housed at the Research Center of Paleobiology, Yuxi Normal University.
All the specimens were photographed with a LEICA DFC 500 digital camera mounted on a Stereoscope LEICA M205 C. The single specimen of Astutuscaris bispinifer gen. sp. nov. was photographed with a LEICA DFC7000 T monochrome digital camera attached to a LEICA M205 FA fluorescence stereomicroscope, and also scanned with a Zeiss Xradia 520 Versa X-ray Microscope.
Systematic palaeontology
Clade Euarthropoda Lankester, 1904
Class, order, and family uncertain
Genus Astutuscaris nov.
Zoobank LCID: urn:lsid:zoobank.org:act:8B855A7C-119F-4DDC-BE27-9AECF99CC735
Etymology: From latin astutus, flexible; and caris, shrimp; referring to its arthropod affinities.
Type species: Astutuscaris bispinifer sp. nov. by monotypy, see below.
Diagnosis.—As for the type species.
Astutuscaris bispinifer sp. nov.
Figs. 1, 2A1, A2, B1–B3.
Zoobank LCID: urn:lsid:zoobank.org:act:D09B7A08-1422-484C-A6AD-0204E2E9F55C
Etymology: From Latin bispinifer, two-spined; referring to its two terminal spines on the end of the body.
Holotype: RCP 0002, an articulated specimen (Fig. 1).
Type locality: The outcrop near Lihuazhuang village (see Jiao et al. 2021b: fig. 1), Yiliang, Kunming, Yunnan, China.
Type horizon: Lower part of Wulongqing Formation, Palaeolenus Biozone, Cambrian Series 2, Stage 4 (Hu et al. 2010; Jiao et al. 2021b).
Material.—Type material only.
Diagnosis.—Small elongate euarthropod with a pair of prominent and bull-horn-like first appendages without sturdy spines and elbow articulations, a wide head shield potentially possessing a pair of antero-median eyes, an elongated thorax consisting of eleven imbricated and homonomous tergites, second to eighth thoracic tergites having lateral doublures and pleural spines, and two long separate spines attached to the last tergite.
Description.—The single nearly complete specimen, dorsoventrally compressed, has a length of 8.6 mm (excluding the first appendage pair), and a width of 2.2 mm (Fig. 1A1–A3, B1–B4). The body is about four times as long as wide in dorsal view. The dorsal exoskeleton consists of a wide head shield and 11 imbricated tergites, and terminates with two sturdy separate spines which are maybe modified, posteriorly directed appendages of the terminal somite (Figs. 1A1, A2, B1–B 4, 2A1, A2, B1–B3). The wide cephalon is incomplete, but it may have a semi-elliptical outline (Figs. 1A1–A4, B1–B4, 2A1, A2, B1–B3). The black carbonaceous structure preserved as a circular shape and located in the anterior and middle part of head is here interpreted as an uncertain structure given that it locates on a different level than the rest of the structures (Figs. 1B1, B2, B6, 2A1, A2, B1–B3). The left frontalmost appendage seems to be an antenna although having no distinct segments, while the right one is likely to be a great appendage for some small and medial spines on its possible last podomere (Fig. 1A1–A4, B1–B4). Considering that the anterior sclerite of fuxianhuiids and some bivalved euarthropods is a complete structure (Fu et al. 2018; Jin et al. 2021), two disjunct, elliptical structures located anterior to the proximal articles of frontalmost appendages are likely to be the eyes for the oval structures and elongate structures which here are respectively interpreted as possible eyes and stalks (Fig. 1A1–A4, B1–B4).
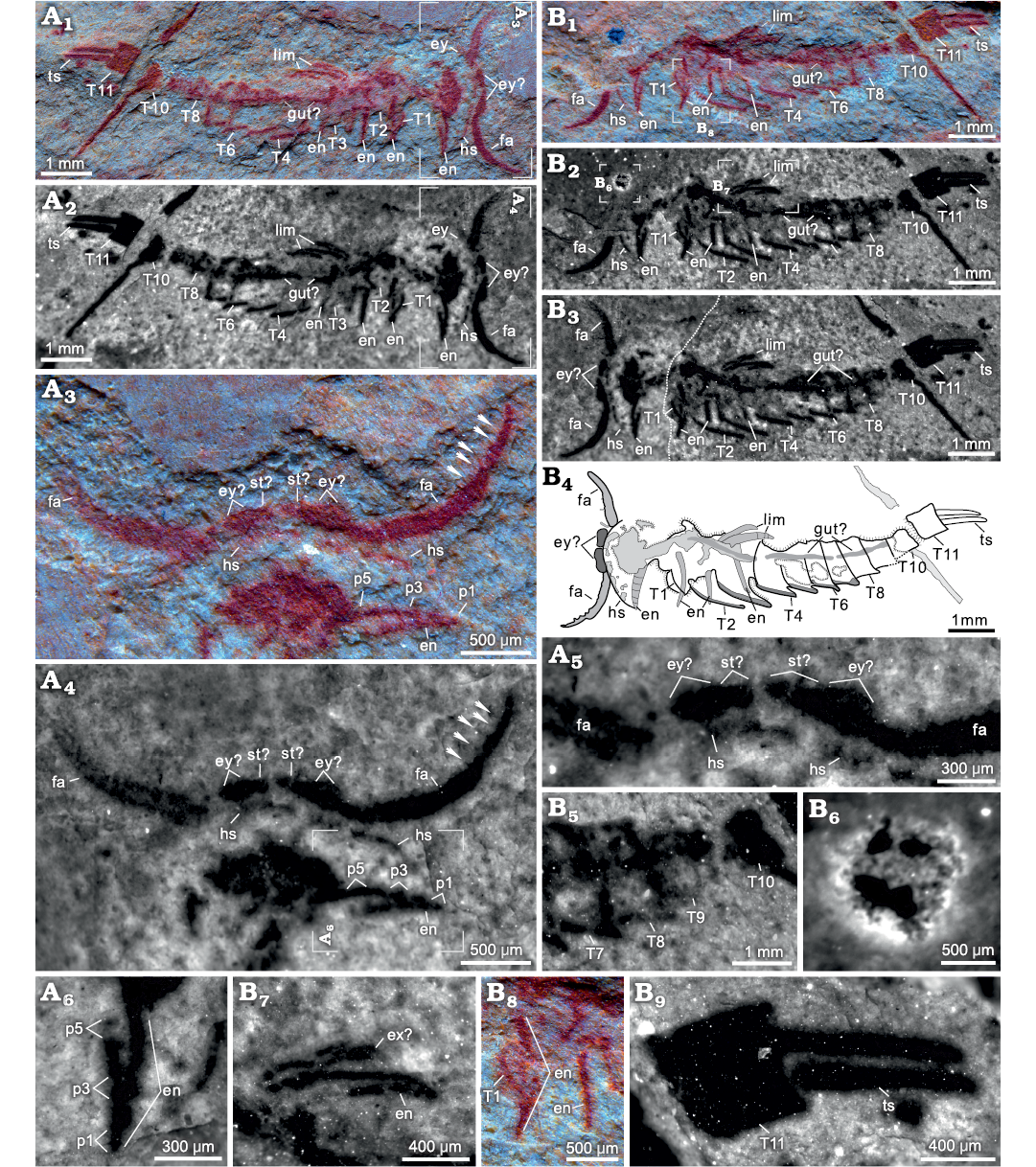
Fig. 1. Euarthropod Astutuscaris bispinifer gen. et sp. nov. from the Cambrian Stage 4 Guanshan Biota, Lihuazhuang village, Yunnan, China. A. RCP 0002a, part of the sole specimen; showing the head shield, 11 imbricated tergites most with pleural spines, and two terminal spines (A1, A2); possible eyes (A3, A4, A5); first appendages with small spines (A3, A4, white arrows); endopod with at least five podomeres (A6). B. RCP 0002b, counterpart of the sole specimen; note the 11 imbricated tergites and trunk limbs (B1, B2); restored specimenby digitally combining the part and counterpart, the dashed line indicating the contact (B3); interpretive drawing of the narrow doublures of thoracic tergites marked in dark grey (B4); T7–T10 and pleural spines (B5); detail of the unknown structure (B6); the limb with an endopod and a possible exopod (B7); single endopod under T1 and T2 respectively (B8); T11 and two separate spines being marked (B9). Natural light photographs (A1, A3, A4, B1, B8); fluorescent photographs (A2, A5–A6, B2, B3, B5–B7, B9). Abbreviations: en, endopod; ex, exopod; ey, eye; fa, frontal appendage; gut, digestive tract; hs, head shield; lim, limb; p1–p5, podomeres 1–5; st, stalk; T1–T11, tergites 1–11; ts, terminal spine.
There are 11 imbricated tergites (Tn) behind the head (Figs. 1A1, A2, B1–B4, 2A1, A2, B1–B3). Incomplete anterior margin of T1, having a similar curvature to that of the successive tergites, is preserved on both part and counterpart (Fig. 1A1, A 2, B1–B4, B8). Anterior margin of T3 is straight, and posterior margins of T5–T9 and T11 are straight (Fig. 1A1, A2, B1–B4). The tergopleura of T3–T8 curve backwards, and their pleural spines are almost posteriorly oriented (Fig. 1A1, A2, B1–B5). The tergopleura of T2–T8 have narrow doublures (Fig. 1A1, A2, B1–B5). T2–T10 have a similar length. T11 is slightly longer than the preceding tergites and lacks pleural spines (Figs. 1A1, A2, B1–B4, B9, 2A1, A2, B1–B3). One robust endopod underneath the head shield preserves at least five conical podomeres (Fig. 1A3, A4, A6). Four elongate, red structures (“en” in Figs. 1A1, A2, B1–B4, B7, B8, 2A1, A2, B1–B3), represent trunk endopods, although they do not display obvious segments. These endopods, one located underneath T1 and T2, and two beneath T3 (Fig. 2A1, A2, B1–B3), indicate there is one pair limb belonging to each of these three tergites. The smaller structure, relative to the endopod, of the limb beneath T3 is tentatively interpreted as exopod (Figs. 1B7, 2A2, B2, B3), further clarification of its morphology is contingent on the discovery of additional specimens. The irregular line passing along the entire length of the body may represent the gut (Fig. 1A1, A 2, B1–B4).
Remarks.—The elongate exoskeleton, the possible eyes and the first pair of appendages located antero-medially on the head, and the curvature of the first appendages are also observed in the great-appendage euarthropods (Haug et al. 2012b; Aria et al. 2020; Liu et al. 2020). The morphology of the frontalmost appendages, 11 trunk tergites, the narrow doublures of tergites, and the two terminal spines of Astutuscaris contribute to distinguish this new euarthropod genus from Yohoia (Haug et al. 2012b). The morphology of the first appendage of Astutuscaris and the pair of posterior terminal spines differ from those of other megacheirans, e.g., leanchoiliids and jianfengiids (Haug et al. 2012a, b; Aria et al. 2020; Liu et al. 2020)
Astutuscaris is distinguishable from Guangweicaris by its curving frontalmost appendages, and elongate exoskeleton without medial axial spines and distinct sections (e.g., the abruptly reduced abdomen). Guangweicaris possesses a wide head sheild with a pair of long antennae, a thorax composed of three small tergites and five normal tergites, and an abruptly reduced abdomen (Fig. 2C, D) consisting of seven tergites all bearing a axial spine (Luo et al. 2007; Yang et al. 2008; Wu and Liu 2019; Chen et al. 2020). The curving first appendages of Astutuscaris are different from the long antennae (at least 21 podomeres) of Guangweicaris (Yang et al. 2008; Chen et al. 2020). Unlike the trunk of Guangweicaris including a suddenly reduced abdomen (Yang et al. 2008; Wu and Liu 2019; Chen et al. 2020), Astutuscaris has a trunk gradually tapering backward. The last tergite of Guangweicaris bears an axial spine (Yang et al. 2008; Chen et al. 2020), while, Astutuscaris has no axial spines. The posterior end of Astutuscaris composes of two isolated spines, and the tail of Guangweicaris possesses a ventral medial extension and two lateral processes (Chen et al. 2020).
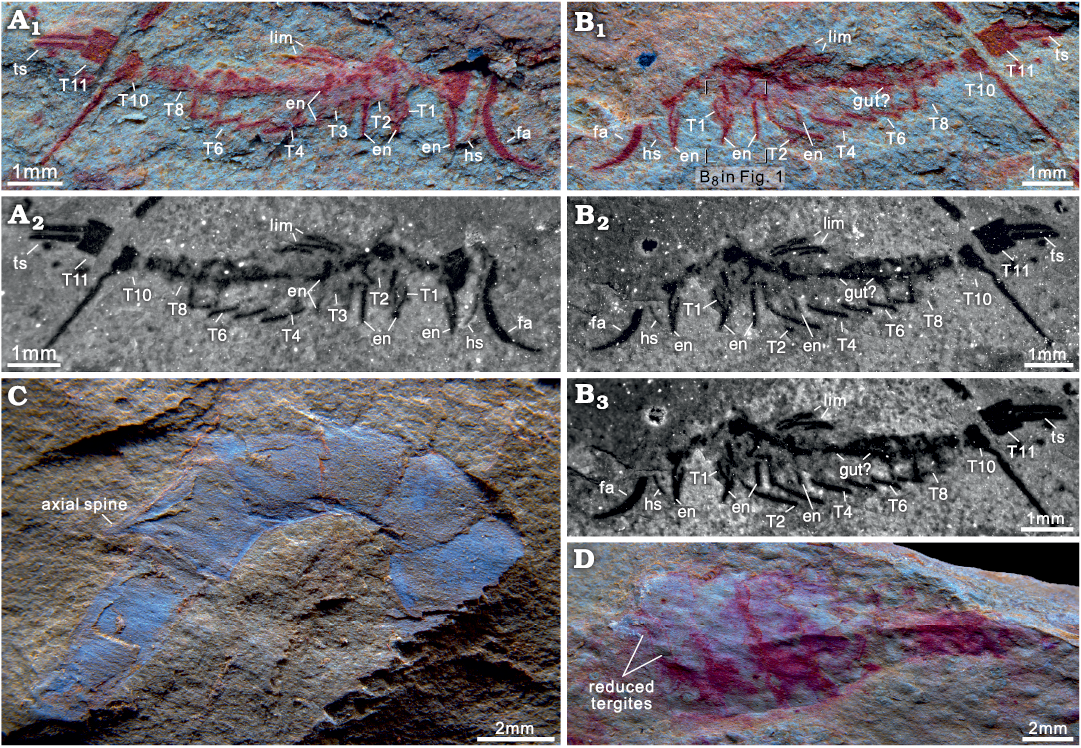
Fig. 2. Euarthropods Astutuscaris bispinifer gen. et sp. nov. (A, B) and Guangweicaris spinatus Luo, Fu, and Hu, 2007 (C, D) collected from Cambrian Guanshan Biota, Lihuazhuang village, Yunnan, China. A. RCP 0002a, specimen before preparation. B. RCP 0002b, specimen before preparation (B1, B2) and after preparation (B3). C. RCP 0003, a small and incomplete specimen about 16 mm in length. D. RCP 0004, a small specimen about 17 mm in length. Natural light photographs (A1, B1, C, D); fluorescent photographs (A2, B2, B3,). Abbreviations: en, endopod; ex, exopod; ey, eye; fa, frontal appendage; gut, digestive tract; hs, head shield; lim, limb; p1–p5, podomeres 1–5; st, stalk; T1–T11, tergites 1–11; ts, terminal spine.
Astutuscaris bispinifer gen. et sp. nov. is different from the stages 8 and 9 of Fuxianhuia protensa Hou, 1987, by its at least 11 trunk tergites, one pair endopod under normal tergites two and three respectively, and last tergite carrying two long and separate spines. The juveniles (stages 8 and 9) of F. protensa are about 11 mm to 15 mm long, and consist of eight or nine trunk tergites which have a pattern of segmental mismatch in thoracic region and a terminal telson with paired caudal flukes (Fu et al. 2018).
Concluding remarks
The curvature and insertion of first appendages, the location of the possible eyes, and the elongate body of Astutuscaris bispinifer gen. et sp. nov. are similar to the Cambrian megacheiran, such as species of Fortiforceps, Jianfengia, Sklerolibyon, and Yohoia. Similarities of A. bispinifer and Yohoia tenuis from the Cambrian Burgess Shale are particularly marked for the elongate exoskeleton and the tergopleura with pleural spines (Haug et al. 2012b; Aria et al. 2020). The smallest Y. tenuis is about 6 mm in length, and bears thirteen trunk tergites and a paddle-like telson with eight marginal spines (Haug et al. 2012b), while A. bispinifer is about 8.6 mm in length, possesses eleven trunk tergites and two terminal spines. The thorax with eleven tergites of A. bispinifer is the same as that of the leanchoiliid megacheiran arthropods, like species of Leanchoilia and Alalcomenaeus (Edgecombe et al. 2011; Hu et al. 2013; Aria et al. 2020; Liu et al. 2020). Evidently, the frontalmost appendage without sturdy spines of this new taxon differs quite considerably from the multisegmented sub-chelate appendages of megacheirans. The pair of terminal spines are another character differentiating A. bispinifer from typical megacheirans (Edgecombe et al. 2011; Hu et al. 2013; Aria et al. 2020; Liu et al. 2020). The prominent and curving frontalmost appendages are also found in some bivalved euarthropods (Fu et al. 2014; Yang et al. 2016; Hou et al. 2017; Jin et al. 2021), e.g., Pectocaris inopinata, Isoxys inopinata, and Clypecaris serrata.
The incompletely preserved walking leg with conical podomeres under the head resembles the limbs of Chengjiangocaris kunmingensis from the Xiaoshiba Biota (see Yang et al. 2013: fig. 1f), but, one pair of limbs beneath each of the first three tergites and the two long terminal spines of Astutuscaris bispinifer gen. et sp. nov. support it is not a fuxianhuiid which have a thorax with ventral segmental mismatch and a terminal tailspine with paired caudal flukes (Fu et al. 2018).
The two sturdy separate spines of Astutuscaris bispinifer gen. et sp. nov. are possibly the modified, posteriorly directed appendages of the terminal somite that is a common character of vicissicaudates a major clade of artiopods (Van Roy 2005; Lerosey-Aubril et al. 2017). If this interpretation is correct, then A. bispinifer might be a vicissicaudate-like euarthropod. Nevertheless, its curving frontalmost appendages and possible anterior and middle eyes still separate it from the vicissicaudates.
The nature of the first curving appendage without obvious segments of Astutuscaris bispinifer gen. et sp. nov. is contingent on the discovery of new specimens, although it’s possible to be an antenna for lacking the typical elbow joints or articulations. Together with the incomplete head, anterior trunk tergites, and limbs, the affinity of A. bispinifer among euarthropods remains unclear until new material that could provide more key taxonomic characters is found. So far, about 24 non-trilobite euarthropod species were reported from Guanshan Biota, i.e., four radiodont species (Jiao et al. 2021b), two fuxianhuiid species (Jiao et al. 2021a), 11 species of bivalved euarthropods (Hu et al. 2013; Wu and Liu 2022), five non-trilobite artiopod species (Hu et al. 2013; Zhao et al. 2020; Jiao et al. 2022), Leanchoilia sp. (Hu et al. 2013), and Lihuacaris ferox (Jiao et al. 2021a). Accordingly, this new taxon increases the species diversity of the typical Burgess Shale-type Guanshan Biota, especially that of euarthropods.
Acknowledgments
We thank Peter Van Roy (Ghent University, Belgium) and one anonymous reviewer for their critical reviews. Special acknowledgements are given to Xi-Guang Zhang, Jie Yang, Tian Lan, Jin-Bo Hou, and Jia-Lin Du (all Yunnan University, Kunming, China) for assistance in fieldwork. This work was supported by the National Natural Science Foundation of China (41730318) and the Innovative Research Fund of Yuxi Normal University (KSD, grant number 202111390020 and 2020B49).
References
Aria, C., Zhao, F.C., Zeng, H., Guo, J., and Zhu, M.Y. 2020. Fossils from South China redefine the ancestral euarthropod body plan. BMC Evolutionary Biology 20: 4. Crossref
Chen, F.Y., Zhang, Z.F., Betts, M.J., Zhang, Z.L., and Liu, F. 2019. First report on Guanshan Biota (Cambrian Stage 4) at the stratotype area of Wulongqing Formation in Malong County, Eastern Yunnan, China. Geoscience Frontiers 10: 1459–1476. Crossref
Chen, H., Legg, D., Liu, Y., and Hou, X.G. 2020. New data on the anatomy of fuxianhuiid arthropod Guangweicaris spinatus from the lower Cambrian Guanshan Biota, Yunnan, China. Acta Palaeontologica Polonica 65: 139–148. Crossref
Daley, A.C., Antcliffe, J.B., Drage, H.B., and Pates, S. 2018. Early fossil record of Euarthropoda and the Cambrian Explosion. Proceedings of the National Academy of Sciences of the United States of America 115: 5323–5331. Crossref
Edgecombe, G.D., García-Bellido, D.C., and Paterson, J.R. 2011. A new leanchoiliid megacheiran arthropod from the lower Cambrian Emu Bay Shale, South Australia. Acta Palaeontologica Polonica 56: 385–400. Crossref
Fu, D.J., Ortega-Hernández, J., Daley, A.C., Zhang, X.L., and Shu, D.G. 2018. Anamorphic development and extended parental care in a 520 million-year-old stem-group euarthropod from China. BMC Evolutionary Biology 18: 147. Crossref
Fu, D.J., Zhang, X.L., Budd, G.E., Liu, W., and Pan, X.Y. 2014. Ontogeny and dimorphism of Isoxys auritus (Arthropoda) from the Early Cambrian Chengjiang biota, South China. Gondwana Research 25: 975–982. Crossref
Giribet, G. and Edgecombe, G.D. 2019. The phylogeny and evolutionary history of arthropods. Current Biology 29: R592–R602. Crossref
Haug, J.T., Briggs, D.E., and Haug, C. 2012a. Morphology and function in the Cambrian Burgess Shale megacheiran arthropod Leanchoilia superlata and the application of a descriptive matrix. BMC Evolutionary Biology 12: 162. Crossref
Haug, J.T., Waloszek, D., Maas, A., Liu, Y., and Haug, C. 2012b. Functional morphology, ontogeny and evolution of mantis shrimp-like predators in the Cambrian. Palaeontology 55: 369–399. Crossref
Hou, X.G., Siveter, D.J., Siveter, D.J., Aldridge, R.J., Cong, P.Y., Gabbott, S.E., Ma, X.Y., Purnell, M.A., and Williams, M. 2017. The Cambrian Fossils of Chengjiang, China: The Flowering of Early Animal life. John Wiley & Sons, Kunming. Crossref
Hu, S.X., Zhu, M.Y., Luo, H.L., Steiner, M., Zhao, F.C., Li, G.X., Liu, Q., and Zhang, Z.F. 2013. The Guanshan Biota [in Chinese, with English abstract]. 204 pp. Yunnan Science and Technology Press, Kunming.
Hu, S.X., Zhu, M.Y., Steiner, M., Luo, H.L., Zhao, F.C., and Liu, Q. 2010. Biodiversity and taphonomy of the Early Cambrian Guanshan biota, eastern Yunnan. Science China Earth Sciences 53: 1765–1773. Crossref
Jiao, D.G., Du, K.S., Zhang, X.G., Yang, J., and Eggink, D. 2022. A new small soft-bodied non-Trilobite artiopod from the Cambrian Stage 4 Guanshan Biota. Geological Magazine 159: 730–734. Crossref
Jiao, D.G., Pates, S., Lerosey-Aubril, R., Ortega-Hernández, J., Yang, J., Lan, T., and Zhang, X.G. 2021a. New multipodomerous appendages of stem-group euarthropods from the Cambrian (Stage 4) Guanshan Konservat-Lagerstätte. Royal Society Open Science 8: 211134. Crossref
Jiao, D.G., Pates, S., Lerosey-Aubril, R., Ortega-Hernández, J., Yang, J., Lan, T., and Zhang, X.G. 2021b. The endemic radiodonts of the Cambrian Stage 4 Guanshan biota of South China. Acta Palaeontologica Polonica 66: 255–274. Crossref
Jiao, D.G., Yang, J., and Zhang, X.G. 2016. A superarmoured lobopodian from the Cambrian Stage 4 of southern China. Science Bulletin 61: 1372–1376. Crossref
Jin, C.F., Mai, H.J., Chen, H., Liu, Y., Hou, X.G., Wen, R.L., and Zhai, D.Y. 2021. A new species of the Cambrian bivalved euarthropod Pectocaris with axially differentiated enditic armatures. Papers in Palaeontology 7: 1781–1792. Crossref
Lerosey-Aubril, R., Zhu, X.J., and Ortega-Hernández, J. 2017. The Vicissicaudata revisited—Insights from a new aglaspidid arthropod with caudal appendages from the Furongian of China. Scientific Reports 7: 11117. Crossref
Liu, J.N., Ou, Q., Han, J., Zhang, Z.F., He, T.J., Yao, X.Y., Fu, D.J., and Shu, D.G. 2012. New occurrence of the Cambrian (stage 4, series 2) Guanshan Biota in Huize, Yunnan, South China. Bulletin of Geosciences 87: 125–132. Crossref
Liu, Y., Ortega-Hernández, J., Zhai, D.Y., and Hou, X.G. 2020. A reduced labrum in a Cambrian great-appendage euarthropod. Current Biology 30: 3057–3061. Crossref
Luo, H.L., Fu, X.P., Hu, S.X., Li, Y., Hou, S.G., You, T., Pang, J.Y., and Liu, Q. 2007. A new arthropod, Guangweicaris Luo, Fu et Hu gen. nov. from the Early Cambrian Guanshan Fauna, Kunming, China. Acta Geologica Sinica-English Edition 87: 1–7.
Van Roy, P. 2005. An aglaspidid arthropod from the Upper Ordovician of Morocco with remarks on the affinities and limitations of Aglaspidida. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 96: 327–350. Crossref
Wu, Y.C. and Liu, J.N. 2019. Anatomy and relationships of the fuxianhuiid euarthropod Guangweicaris from the early Cambrian Guanshan Biota in Kunming, Yunnan, Southwest China revisited. Acta Palaeontologica Polonica 64: 543–548. Crossref
Wu, Y.C. and Liu, J.N. 2022. New data on the bivalved arthropod Tuzoia from the Cambrian ( Series 2 , Stage 4 ) Guanshan Biota in Kunming, Yunnan, Southwest China. Frontiers in Earth Science 10:862679. Crossref
Yang, J., Hou, X.G., and Dong, W. 2008. Restudy of Guangweicaris Luo, Fu et Hu, 2007 from the lower Cambrian Canglangpu formation in Kunming area. Acta Palaeontologica Sinica 47: 115–122.
Yang, J., Ortega-Hernández, J., Butterfield, N.J., and Zhang, X.G. 2013. Specialized appendages in fuxianhuiids and the head organization of early euarthropods. Nature 494: 468–471. Crossref
Yang, J., Ortega-Hernández, J., Lan, T., Hou, J.B., and Zhang, X.G. 2016. A predatory bivalved euarthropod from the Cambrian (Stage 3) Xiaoshiba Lagerstätte, South China. Scientific Reports 6: 27709. Crossref
Zhao, J., Li, Y.J., Selden, P.A., and Cong, P.Y. 2020. New occurrence of the Guanshan Lagerstätte (Cambrian Series 2, Stage 4) in the Kunming area, Yunnan, southwest China, with records of new taxa. Alcheringa 44: 343–355. Crossref
Acta Palaeontol. Pol. 67 (4): 969–974, 2022
https://doi.org/10.4202/app.00937.2021