
New records of Late Triassic wood from Argentina and their biostratigraphic, paleoclimatic, and paleoecological implications
LAURA VALLEJOS LEIZ, ALEXANDRA CRISAFULLI, and SILVIA GNAEDINGER
Vallejos Leiz, L., Crisafulli A., and Gnaedinger, S. 2022. New records of Late Triassic wood from Argentina and their biostratigraphic, paleoclimatic, and paleoecological implications. Acta Palaeontologica Polonica 67 (2): 329–340.
We report gymnospermous wood found in sandstone and siltstone beds of the Upper Triassic Hilario Formation, Sorocayense Group at Hilario Creek located in San Juan province, Argentina. The identified xylotaphoflora comprises Baieroxylon cicatricum (Ginkgoales) and a new species of Protophyllocladoxylon (Coniferales), it constitutes the first reports of these taxa from the Triassic in Argentina. Protophyllocladoxylon hilarioense sp. nov. differs from the other species by the following combination of anatomical characters: radial pits araucarian, mixed and some with abietinian tendency, uni-biseriate; contiguous, separated; tangential pits uni-biseriate; cross-field pits are simple elliptic, oblique, one to two in number and low uni-biseriate rays. The growth rings in the reported woods show a gradual transition from earlywood to latewood, suggesting little change in the climatic conditions experienced during their growth. The type of growth rings observed is consistent with a humid but seasonally dry subtropical climate. These woods are representatives of the arboreal stratum of a mesophytic association.
Key words: Ginkgoales, Coniferales, Pinales, Baieroxylon, Protophyllocladoxylon, Triassic, Hilario Formation, Argentina, San Juan.
Laura Vallejos Leiz [lauravallejosleiz@gmail.com], Alexandra Crisafulli [alexandracrisafulli@hotmail.com], and Silvia Gnaedinger [scgnaed@hotmail.com], Centro de Ecología Aplicada del Litoral (CECOAL-CONICET-UNNE), Ruta 5 km 2,5, (3400), Corrientes, Argentina.
Received 30 August 2021, accepted 5 November 2021, available online 29 December 2021.
Copyright © 2021 L. Vallejos Leiz et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The wood records from the Cuyana Basin of Argentina are scarce and poorly known. This basin is filled up with sediments of two groups: Sorocayense Group and Rincón Blanco Group, and it is located in the southwest of the San Juan province. These sedimentary units yielded rich taphofloras composed of Bryophyta, Lycophyta, Sphenophyta, Corystospermales, Peltaspermales, Cycadales, Coniferales, and Gnetales (Artabe et al. 2001, 2007a, b; Gnaedinger and Lutz 2008; Bodnar et al. 2018, 2019; Drovandi et al. 2020). The Upper Triassic Hilario Formation of the Sorocayense Group has yielded petrified wood that is an important addition to the Triassic lignoflora of Gondwana and the importance of this finding is discussed in this paper, both in systematic and palaeoenvironmental aspects.
Institutional abbreviations.—CTES-PB, Paleontological Collections of the National University of the Northeast Dr. Rafael Herbst at the CECOAL-CONICET-UNNE, Corrientes, Argentina.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in Plant Fossil Names Registry (PFNR): urn:lsid:plantfossilnames.org: ref:967
Geological setting
The Cuyana, Ischigualasto-Villa Unión, and Marayes-El Carrizal basins are part of a series of basins that extended along the western margin of Gondwana in the early Mesozoic. They represent a continuous record of the Triassic in Argentina exhibiting several common paleofloristic elements (Drovandi et al. 2020). The Triassic sediments of the Cuyana Basin expose diverse taphofloras and are located from the southwest region of San Juan to the north of Mendoza. The taphofloras found in San Juan are located in two regions: Barreal-Calingasta, along the eastern flank of Los Patos river valley, and in Rincón Blanco in the Sierra del Tontal (Kokogian et al. 1999) (Fig. 1). The Barreal-Calingasta successions are assigned to the Sorocayense Group (Groeber and Stipanicic 1953; Baraldo and Guerstein 1984; Spalletti 1995; Barredo et al. 2016). This group is divided into the northern and the southern regions. Stratigraphically, the northern region is composed of the formations Agua de los Pajaritos, Monina, Hilario, and El Alcázar; in the Hilario area (Stipanicic 1979; Groeber and Stipanicic 1953; Baraldo and Guerstein 1984; Baraldo et al. 1990; Barredo 2012; Barredo et al. 2016; Table 1) while in the southern región of the Sorocayense Group, the Barreal, Cortaderita and Cepeda formations crop out in the Barreal area (Bodnar et al. 2019).
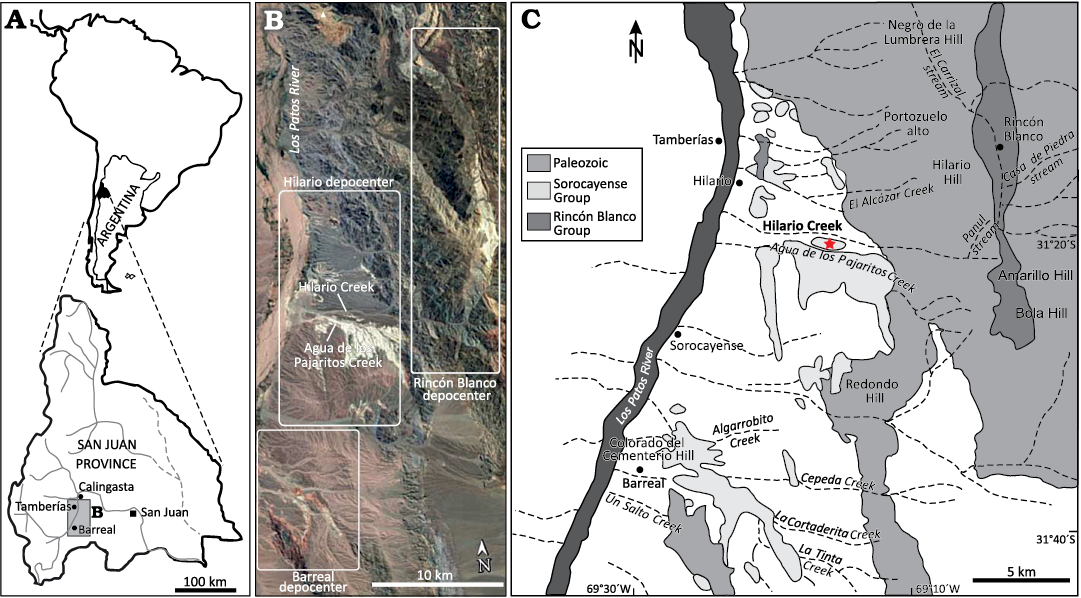
Fig. 1. Location of petrified woods. A. Location of the study area at San Juan province, Argentina. B. Hilario (northern), Barreal (southern) and Rincón Blanco depocenters of Cuyana Basin. The satellite image taken from Google Earth Pro. C. Location map of the Hilario Creek, in the Hilario (northern) depocenter. Modified from Ruiz and Bodnar (2019).
The stratigraphic framework of the Sorocayense Group in the Barreal (southern) area is overlapping on the discordance of Paleozoic sediments and Devonian diabases, and Hilario on “schists”, graywacke and diabases of the Lower Paleozoic (Stipanicic 1972; Barredo et al. 2016).
The Triassic sequence in the Hilario depocenter (northern) of the Sorocayense Group begins with the Agua de los Pajaritos Formation, which represents the onset of the basin filling with fluvial-sandy elements and abundant pyroclastic input followed by levels of bituminous pellets from the Monina Formation (Baraldo and Guerstein 1984).
The filling is completed by the El Alcázar Formation characterized by clastic and pyroclastic sediments of fluvial and lacustrine environments alternating with tuff deposits (Baraldo and Guerstein 1984; Baraldo et al. 1990; Barredo et al. 2016; Table 1).
In the Barreal (southern) depocenter, the Hilario Formation correlates with the Don Raúl member of the Cortaderita Formation, which corresponds to an anastomosed fluvial system and flood plains where temporal ponds or lakes developed (Bodnar et al. 2019).
The Hilario Formation consists of sandstone-pelite deposits, and intercalations of tuffs, corresponding to a lacustrine environment that passes into a fluvial environment with flood plains and a palustrine environment.
The Hilario Formation was first described as a part of the Hilario Triassic Series (Pozzo 1948; Groeber and Stipanicic 1953) and later, within the Hilario Group, as an informal denomination of Stipanicic (1969). According to the nomenclature of the “Comité Argentino de Estratigrafía (1992)” (Argentine Committee of Stratigraphy), the denomination of “Hilario Triassic Series” and “Hilario Series” were not valid due to the use of the same toponym for two entities of different hierarchies such as Formation and Group (Stipanicic 2002; Table 1).
The age of the Hilario Formation would correspond to the Norian–Rhaetian (Upper Triassic), based on the stratigraphic correlations elaborated by Spalletti et al. (1999) and Stipanicic (2002).
Table 1. Table of historical comparison of the different stratigraphic nomenclatures of the Sorocayense Group (modified from Barredo et al. 2016).
|
Pozzo (1948) |
Groeber and Stipanicic (1953) |
Stipanicic (1972) |
Baraldo and Guerstein (1984) |
Barredo et al. (2016) |
|
Hilario Triassic Series |
Hilario Series |
Sorocayense Group (North) |
Sorocayense Group |
Sorocayense Group |
| |
|
|
El Alcázar Formation |
El Alcázar Formation |
|
Tobiferous-sandy Trias |
Strata of Hilario |
Hilario Formation |
Hilario Formation |
Hilario Formation |
|
Monina Formation |
||||
|
Clay Trias |
Strata of El Alcázar |
El Alcázar Formation |
Monina Formation |
|
|
Agua de los Pajaritos Formation |
||||
|
Basal or conglomerate Trias |
Strata of Agua de los Pajaritos |
Agua de los Pajaritos Formation |
Agua de los Pajaritos Formation |
Historical background
Bodnar et al. (2019) suggested that the taphofloras of the Barreal (southern), Hilario (northern), and Rincón Blanco (eastern) depocentres display similarity in plant paleocommunities, in which the main components are Corystospermales: of arboreal species of Zuberia and shrubs Dicroidium, Xylopteris, Peltaspermales (Lepidopteris, Pachydermophyllum), and Cycadales (Pterophyllum and Pseudoctenis). Wood taxa reported from the Sorocayense Group are as follows: Rhexoxylon sp. from the Barreal Formation (Lutz and Herbst 1992); Rhexoxylon sp., Tranquiloxylon sp., and Protocircoporoxylon sp. from the El Alcazar Formation (Ganuza et al. 1998; Drovandi et al. 2016), Rhexoxylon cortaderitaense, Juniperoxylon zamunerae from the Cortaderita Formation, and Protocircoporoxylon sp. from the Monina Formation (Drovandi et al. 2020).
However, the detailed descriptions of fossils from the Hilario Formation are only those of the sphenopsids Neocalamites carrerei, Neocalamites sp. and the conchostracans Estheria sp. (Groeber and Stipanicic 1953).
Material and methods
The studied material consists of silicified, decorticated specimens, with good preservation of cellular elements, which are deposited in the Colecciones Paleontológicas de la Universidad Nacional del Nordeste “Dr Rafael Herbst”, Sección Paleobotánica de Corrientes (CTES-PB). They were collected in various expeditions in the decades 1980–1990 by Rafael Herbst, Alicia Lutz, Susana Morton, Oscar Gallego and collaborators in Hilario Creek (Fig. 1). This site is located 122 m above sea level, and its coordinates are 31°20’S and 69°15’W.
The specimens were processed according to the standard methodology by making cuts in the transverse, radial longitudinal, and tangential longitudinal sections, and polishing with abrasives of different granulometry up to a thickness of 40 μm (Hass and Rowe 1999). The thin sections were studied in detail with a Leica microscope (DM500), the photomicrographs were taken with a digital camera (Leica ICC50). The scanning electron microscope (SEM Jeol 5800LV) from the Universidad Nacional del Nordeste, Corrientes, Argentina, has been used in pertinent cases.
The measurements of the different anatomic elements were obtained after with a minimum of 30 measurements. The mean and, between parent, the minimum and maximum values were recorded in the descriptions.
The terminology used corresponds to the list of microscopic characters for the wood identification of the International Association of Wood Anatomists (Richter et al. 2004) and García Esteban et al. (2002, 2003). Systems of nomenclature and nomenclatural reviews are those of Philippe and Bamford (2008) and Zijlstra and Phillipe (2020).
Systematic palaeontology
Class Spermatopsida Serbet and Rothwell, 1995
Order Ginkgoales Engler, 1897
Genus Baieroxylon Greguss, 1961
Type species: Baieroxylon implexum Greguss, 1961; Löwenstein Formation, Keuper, Upper Triassic of Germany.
Baieroxylon cicatricum Prasad and Lele, 1984
Figs. 2–4.
1984 Baieroxylon cicatricum sp. nov.; Prasad and Lele 1984: 389–391, pl. 1, pl. 2: 2.
1986 Baieroxylon cicatricum Prasad and Lele, 1984; Muralidhar-Rao and Ramanujan 1986: 119–122, text-figs. 1–7, pl. 1: 1–5.
2001 Baieroxylon cicatricum Prasad and Lele, 1984; Crisafulli 2001: 62–63, 65, figs. 2A–G, 4.
2009 Baieroxylon cicatricum Prasad and Lele, 1984; Crisafulli and Herbst 2009: 103–106, figs. 5H, 6A–E.
2009 Baieroxylon cicatricum Prasad and Lele, 1984; Bardola et al. 2009: 142–144, figs. 4–6.
2012 Baieroxylon cicatricum Prasad and Lele, 1984; Leiva Verón et al. 2012: 76, 78, fig. 8.
Material.—CTES-PB 11411, a piece of silicified wood branch from Hilario Creek, San Juan province, southwest Argentina, Upper Triassic.
Description.—Decorticated secondary wood fragment, silicified, dark grey, 10 cm in major diameter and 9 cm in minor diameter, and 3.5 cm in length. Externally, it presents a branch with numerous simple scars (from smaller branches or leaves) that follow a helical disposition (Fig. 2A1). These are ovoid or eye-shaped (sensu Prasad and Lele 1984) in outline and range in diameter from 5 to 7 mm. In transverse section, the transition from earlywood to latewood is gradual. The tracheids of the earlywood are round to quadrangular in shape, with variable size and with intercellular spaces of varying shape and dimensions, which give the wood a disorganized appearance in some sectors. They have an average radial diameter of 48 μm (30–65 μm) and a tangential diameter of 46 μm (26–62 μm). These present an average of 6 tracheids (2–10) between adjacent rays. Latewood forms one or two rows of tracheids (Fig. 2A3). The cell radial diameter is 16 µm (13–18 µm) and the tangential diameter is 23 µm (14–35 µm) (Fig. 2A4). The mean sensitivity is 0.3 (Fritts 1976; Gou et al. 2021). In some sectors, false growth rings are observed.
In longitudinal radial section, the tracheids present uniseriate araucarian pitting (50%), biseriate (25%) and uniseriate partially biseriate (15%) and mixed (10%) pitting with fine spiral thickenings inclined at an angle of 52° in a clockwise direction. Uniseriate pits are circular, contiguous, or widely spaced. Their average measurements are 10 μm wide and 12 μm high, with a flattening coefficient of 0.83. Biseriate pits are circular, contiguous, alternate, opposite, and some of them are spaced. (Figs. 2A5, A7, 3A1). They measure an average of 9 μm wide by 9 μm high. The opening of the areola measures 7 µm on average. Some cross-fields have up to 5 cupressoid pits, with an araucarioid arrangement, others with 3–4 pits grouped in a cluster measure 5 μm on average (Figs. 2A6–A8, 3A2).
In longitudinal tangential section, the rays are homocellular and uniseriate. The height of the rays is 1–12 cells; the average height is medium (6 cells). The average diameter of the cells is 23 µm high by 22 µm wide (Fig. 2A9). The density of rays is 23 per mm2. In this section, it is possible to visualize a rameal trace that has preserved a homogeneous medulla composed of parenchymatic cells with an oval to circular contour surrounded by the rays. Their average diameter is 13 µm (7–23 µm) (Fig. 2A10).

Fig. 2. Ginkgoalean wood Baieroxylon cicatricum Prasad and Lele, 1984 (CTES-PB 14411) from Hilario Creek, San Juan province, Argentina, Upper Triassic. General aspect of the wood showing the “eye-shaped” traces (A1). Detail of the “eye-shaped” trace (A2). Transverse sections of the secondary wood showing growth rings (white arrow) and “shearing zones” (black arrows) (A3) and tracheids of unequal size (A4). Radial longitudinal sections showing partially biseriate uniseriate pits, opposite biseriate pits (arrow) (A5, A7) and cupressoid cross-fields and others with the pits grouped in a cluster (A6, A8). Radial longitudinal sections showing rays (A9) and trace detail (A10).

Fig. 3. Ginkgoalean wood. Baieroxylon cicatricum Prasad and Lele, 1984 (CTES-PB 14411) from Hilario Creek, San Juan province, Argentina, Upper Triassic. Longitudinal radial section of tracheids with biseriate pits (A1), Pits in the cross-fields (A2, arrows).
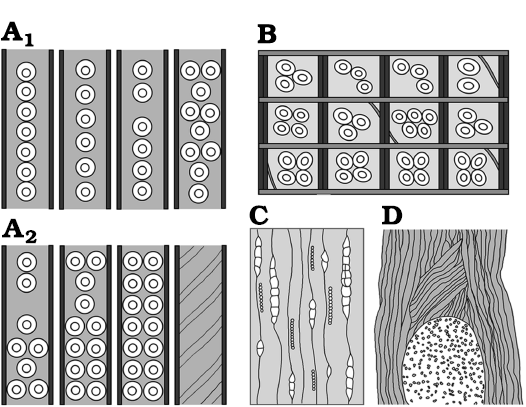
Fig. 4. Schemes showing anatomical characters of Baieroxylon cicatricum. A. Tracheid radial pitting patterns (A1, A2). B. Cross field pitting. C. Radial system. D. Detail of scar.
Remarks.—The anatomical characters of the wood branch under description allow to assign it to Baieroxylon, which is characterized by the presence on the radial walls of the tracheids of uni-biseriate pits, often araucarian, flattened, mixed type pitting with helical thickenings; and rays 1–15 cells high, mostly uniseriate (Greguss 1961; Philippe and Bamford 2008). Based on the radial pitting, Baieroxylon unites a group of heterogeneous woods; some species show an araucarioid type and others a mixed type (Gnaedinger 2012).
Baieroxylon has a wide stratigraphic distribution. It is present in sediments from the Permian to the Cretaceous (Crisafulli 2001; Gnaedinger 2012), and includes seven species: Baieroxylon multiseriale Prasad, 1982, from Kamthi Formation from upper Permian of India; Baieroxylon implexum Greguss, 1961 from Permian of Hungary and Upper Triassic of Germany; Baieroxylon graminovillae Prasad and Lele, 1984, from Keuper (Upper Triassic of Germany); Baieroxylon cambodiense Serra, 1966a, from Mesozoic of Cambodia; Baieroxylon lindicianum Philippe, 1995, from Liassic (Lower Jurassic) of France and Hungary; Baieroxylon chilense Torres and Philippe, 2002, from Upper Triassic of Chile and Argentina; Baieroxylon rocablanquense Gnaedinger, 2012, from Lower Jurassic of Argentina.
According to the comparison made between the species of Baieroxylon (SOM: table 1, Supplementary Online Material available at http://app.pan.pl/SOM/app67-VallejosLeiz_etal_SOM.pdf), the studied specimen of the Hilario Formation fits the diagnosis of B. cicatricum due to a similarity in types, size, shape, and seriation of radial pits, cross-fields, and uniseriate woody rays in the secondary wood and numerous “eye-shaped scars”.
The analyzed species is similar to B. graminovillae and B. cambodiense, due to similarities in radial pitting of tracheids and cross-fields (pits grouped in a cluster). However, B. graminovillae has tracheids with some triseriate pits and B. cambodiense has crassulae in the pits and a greater number of them in the cross-fields and also the presence of biseriate rays.
Our material supports the assignment of Baieroxylon to the woods of Ginkgoales due to the following anatomical characters: intercellular spaces between tracheids pointed ends in recurvate tracheids, simple or cupressoid pits in cross-fields, low rays and the variable size of the secondary wood tracheids (compare Greguss 1955; Prasad and Lele 1984; Crisafulli 2001; Leiva Verón et al. 2012; Gnaedinger 2012). It is also worth mentioning that Prasad and Lele (1984) found impressions of Baiera digitata (Brongniart 1828) Heer, 1876, from the Permian of Hungary, in the same outcrops as Baieroxylon sp.
Stratigraphic and geographic range.—Recognised from the Late Permian of Paraguay, Uruguay, and Africa, Upper Triassic of Argentina (this paper), Brazil and India, and Cretaceous of India.
Order Pinales Dumortier, 1829
(= Coniferales Gorozhankin, 1904)
Family Podocarpaceae Endlicher, 1847
Genus Protophyllocladoxylon (Kräusel, 1939) Mussa, 1958
Type species: Protophyllocladoxylon dolianitii Mussa, 1958; Tubarão Series, Guatá Group, Rio Bonito Formation, Santa Catarina System of Santa Catarina, Brazil, lower Permian; see Zijlstra and Philippe (2020).
Protophyllocladoxylon hilarioense sp. nov.
Figs. 5–7.
PFNR: PFN002690.
Etymology: In reference to the location where it was found.
Type material: Holotype CTES-PB 11409. Paratype CTES-PB 11406; both silicified wood fragments from the type locality and horizon.
Type locality: Hilario Creek, Cuyana Basin, San Juan province, Argentina.
Type horizon: Hilario Formation, Upper Triassic.
Diagnosis.—Secondary pycnoxylic wood with growth rings slightly marked. Radial tracheid pitting of the earlywood uni-biseriate. Radial tracheid pitting of the latewood uniseriate. Uniseriate pits are circular, contiguous, flattened, and spaced with a tendency to abietinoid type. Biseriate pits circular, opposite and alternate, contiguous, and flattened. Cross-fields with 1–2 simple oblique to horizontal phyllocladoid oopores. Very low-medium uniseriate rays and some uniseriate rays with a short biseriate portion, 1–8 cells high. Tangential walls of tracheids with contiguous, circular and uniseriate pits.
Description.—Decorticated, silicified, dark brown wood fragments. CTES-PB 11409 (Fig. 5A1) has been selected as the holotype, and CTES-PB 11406, as the paratype. The holotype measures 3.5 cm in length, 5.5 cm in the largest diameter and 3.5 cm in the smallest diameter.
In transverse section, the growth rings are slightly marked (Fig. 5A2, A3). The tracheids are circular, oval to quadrangular, and with wide lumens. The average tangential is 33 µm, and the radial diameter is 34 µm. The thickness of the tracheal wall is 5 µm. It presents “shearing zones” sensu Erasmus (1976). It shows false growth rings.
In longitudinal radial section, the walls of the tracheids present uniseriate (70%) and biseriate (30%) araucarian pitting. When uniseriately arranged the pits are circular, contiguous (araucarioid type), and mixed, others are spaced (abietinoid type) and some are flattened (Fig. 5A4, A5, A7, B). It measures an average of 18 μm wide × 16 μm long, with a flattening coefficient of 0.89. When biseriately arranged the pits are circular, sub-opposite, contiguous, and flattened, some are spaced. They have an average width of 14 µm × 16 µm long (Figs. 5A6–A8, 6A1, A2). The aperture of the areole measures 8 µm on average. The cross-fields present 1–2 simple oblique to horizontal elliptical phyllocladoid-type oopores, with an average aperture of 14 μm (Figs. 5A9, A10, 6A3).
In longitudinal tangential section, the radial system is homogeneous. Some rays are uniseriate and some are uniseriate with a short biseriate portion. The rays are very low and medium; the average height is 4 (1–8) cells. Cells average is 26 μm high × 23 μm wide. The density of rays is 22 per mm2. The cells of the rays are oval in the central portion and triangular to ellipsoidal at the ends. The tangential walls of the tracheids have contiguous, circular, uniseriate areolate pits (Fig. 5A11, A12).

Fig. 5. Podocarpacean wood, Protophyllocladoxylon hilarioense sp. nov. from Upper Triassic, Hilario Creek, San Juan province, Argentina. Transverse sections (A1–A3); radial longitudinal sections of earlywood (A4–A10, B); tangential longitudinal sections (A11, A12). A. CTES-PB 14409, the general aspect of the wood (A1), secondary xylem with growth rings (A2), tracheids of late wood (arrow, A3); uniseriate pits and flattened pits (A4, A5), biseriate and alternate pits (A6), uniseriate flattened pits (black arrow) and biseriate opposite pits (white arrow) (A7), opposite pits (black arrow) and subopposite pits (white arrows) (A8), cross-fields with oblique to horizontal phyllocladoid oopores (A9, A10), uniseriate rays (A11, A12). B. CTES-PB 14406, uniseriate pits and flattened pits (B1), mixed pits with a tendency to abietinoid (B2), uniseriate separate pits (B3).
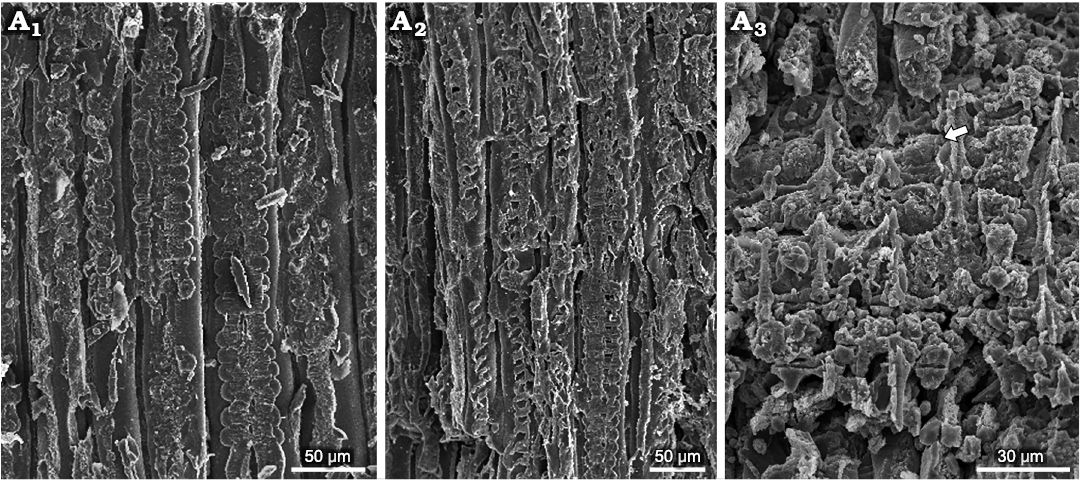
Fig. 6. Podocarpacean wood Protophyllocladoxylon hilarioense sp. nov. (CTES-PB 14409) from Upper Triassic, Hilario Creek, San Juan province, Argentina. Longitudinal radial section. Flattened biseriate and uniseriate pits (A1); flattened uniseriate and biseriate pits (A2); cross fields with phyllocladoid oopores (A3, arrow).
Remarks.—The specimens present araucarioid, abietinoid and mixed pitting on the radial walls of the tracheids and simple phyllocladoid pits in the cross-fields, consistent with the diagnosis of the genus Protophyllocladoxylon (Kraüsel, 1939) Mussa, 1958. Vozenin-Serra (1970) classified Protophyllocladoxylon species according to four characters: types of radial pitting, various aspects of pits in cross-fields, presence or absence of axial parenchyma and seriation of rays. Pant and Sing (1987) include it in the group of araucarioid woods for the Paleozoic and Bamford and Philippe (2001) in the mixed type for Mesozoic woods. Gnaedinger (2007) and Philippe and Bamford (2008) according to the described species show that Protophyllocladoxylon is characterized by species with an araucarioid woody plan and others with a mixed woody plant. Therefore, the key of the latter authors includes Protophyllocladoxylon in Group B (araucarian or xenoxylean radial pitting) and in Group D (with mixed type radial pitting). The described specimens of the Hilario Formation correspond to the mixed type or group D.
Wan et al. (2019) point out that Protophyllocladoxylon species can have three types of pits in the cross-fields: phyllocladoid type (Philippe and Bamford 2008), window-type (Richter et al. 2004) and others with small to medium-sized oval/circular pits, being all simple without borders. Some species have two types such as Protophyllocladoxylon owensii Fletcher, Cantrill, Moss, and Salisbury, 2014, showing phyllocladoid oopores and less circular pits. In the Hilario Formation specimens, the cross fields have only the phyllocladoid type of pits.
For the specific determination of the wood, the comparisons were established with species from the Triassic (SOM: table 2). It is worth mentioning that this specimen has similarities with P. korubaense Serra, 1966b, and with P. xenoxyloides Serra, 1966b, both from Cambodia, regarding the pitting of the radial walls of the tracheids, and the cross-field pits. However, it differs from P. korubaense because the specimen from Argentina does not have axial parenchyma, nor tyloses, and presents flattened pits, a greater number of tangential pits and the pits of the cross-fields with elliptical oopores. Similarly, P. xenoxyloides is distinguished from the Hilario Formation specimen by presenting axial parenchyma, tyloses, spaced pits and the percentage of flattened pits is different from the specimens analyzed here. However, Boura et al. (2013) synonymise the species P. korubaense and P. thylloides (from Upper Triassic of Vietnam) with P. xenoxyloides Serra, 1966a (Triassic–Jurassic) based on a reassessment of quantitative anatomical parameters and the presence of tyloses.
According to the indicated differences with the two closest species shown in SOM: table 2, it is proposed to assign the materials from the Hilario outcrop to a new species Protophyllocladoxylon hilarioense sp. nov.
This new taxon is added to others from South America such as Protophyllocladoxylon dolianitii Mussa, 1958, Protophyllocladoxylon derby (Olivera, 1936) Maheshwari, 1972, and Protophyllocladoxylon rosablancaense Pons, 1971. Protophyllocladoxylon derbyi comes from the upper Carboniferous of Brazil. Protophyllocladoxylon dolianitii is recorded in the Permian formations of Brazil, Paraguay, Antarctica, and Australia (Mussa 1958; Crisafulli and Herbst 2009; Maheshwari 1972; Wan et al. 2020, respectively) and P. rosablancaense was found in Rosa Blanca Formation (Middle Valley of Rio Magdalena, Colombia) in Cretaceous sediments (Pons 1971). The SOM: table 3 compares the anatomical characters of these three taxa with the Hilario specimen. Protophyllocladoxylon rosablancaense is the one that shows the most similarity with the material studied herein; although P. rosablancaense does not show spaced radial pitting or tangential pitting and the height of the rays is high.
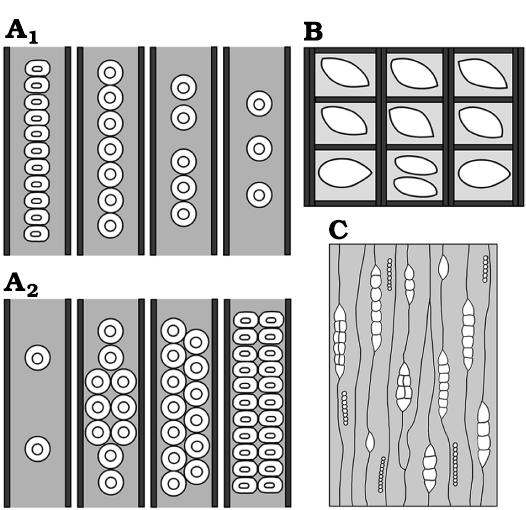
Fig. 7. Schemes showing anatomical characters of Protophyllocladoxylon hilarioense. A. Tracheid radial pitting patterns (A1, A2). B. Cross field pitting. C. Radial system.
Following the criteria of Gnaedinger (2007), Zhang et al. (2010), and Pujana et al. (2014, 2015), Protophyllocladoxylon is most likely related to the basal forms of Podocarpaceae, due to the presence of cross-field pits of phyllocladoid-type, characteristic of some current members of Pinaceae and Podocarpaceae (Ritcher et al. 2004) and for presenting araucarian and mixed pitting on the radial walls of the tracheids. Likewise, in the Triassic, this type of woody axis was found close to Rissikia sp. (Podocarpaceae) leaves with female (Rissikistrobus sp.) and male (Rissikianthus sp.) cones (Townrow 1967; Anderson and Anderson 2003; Gnaedinger and Herbst 2008; Gnaedinger 2010; Holmes and Anderson 2013; Gnaedinger and Zavattieri 2017).
Stratigraphic and geographic range.—Upper Triassic, Hilario Formation, Hilario Creek, Cuyana Basin, San Juan province, southwest Argentina.
Discussion
Biostratigraphic implications.—Baieroxylon cicatricum is known from the upper Permian of the Tacuary Formation in the localities of Arroyo Vino (Crisafulli and Herbst 2009) and Guavirá in Paraguay (Leiva Verón et al. 2012), the Yaguarí Formation in Uruguay (Crisafulli 2001), and the Ecca Group in Africa (Crisafulli and Herbst 2010). It has also been found in the Upper Triassic of the Tiki Formation (Prasad and Lele 1984) and in the Cretaceous of the Gangapur Formation (Muralidhar-Rao and Ramanujan 1986), both in India, and in Rio Grande do Sul, Brazil (Bardola et al. 2009). Here we present the first record from the Triassic of Argentina. The presence of Ginkgoales in the Sorocayense Group is furthermore corroborated by the record of leaf impressions.
Ginkgophyta had its acme in the Mesozoic times. Then, the group declined during the Cenozoic, with only Ginkgo biloba persisting to the present times (Taylor et al. 2009). According to the considerations made by Gnaedinger (2012), Ginkgoales fossils present secondary xylem of the araucarian or mixed type from the Permian to the Cretaceous and an abietinian type from the Late Cretaceous to the Tertiary (similar to the living species of Ginkgo biloba). Moreover, the stratigraphic and paleogeographic distributions of these woods in terms of anatomical changes (from mixed type to abietinian type) are concurrent with the morphological changes in leaves (lamina and petiole differentiation) and reproductive structures (reduction of numbers of seeds and increase in their size) reported in Zhou and Wu (2006) and Zhou (2009).
This group is recorded throughout Gondwana, presenting a large number of endemic forms in the southwest. Meanwhile, the greatest biodiversity is recorded in the high paleolatitudes (around 60° south paleolatitude), in the Karoo Basin (South Africa) and El Tranquilo Group (Patagonia, Argentina).
Protophyllocladoxylon is a cosmopolitan taxon. Its oldest record is from the Carboniferous. Zhang et al. (2010) presented a comparative table that includes most of the known Paleozoic and Mesozoic species. Then Boura et al. (2013), Fletcher et al. (2014), Pujana et al. (2014, 2015), Iamandei et al. (2018), Wan et al. (2019), and Gou et al. (2021) detailed other species of Protophyllocladoxylon and extended their range to the Eocene.
Protophyllocladoxylon hilarioense sp. nov. is the thirty-sixth species of the genus and its first record from the Triassic of Argentina.
Paleoclimatic and paleoecological implications.—Baieroxylon and Protophyllocladoxylon species occur in various paleoclimates, both in warm and humid and temperate to cold conditions (Crisafulli and Herbst 2009; Zhang et al. 2010; Gnaedinger 2012).
The presence of variations in the growth rings of the analyzed woods may indicate slight seasonal changes characterized by the alternation of dry and humid periods in response to variables such as rainfall, temperature and light availability (Creber and Chaloner 1984; Taylor and Ryberg 2007; Pires and Guerra Sommer 2011; Yang et al. 2013).
The rings analyzed correspond to type “E” of the classification proposed by Creber and Chaloner (seen in Brison et al. 2001). It means that these plants went through relatively uniform growing seasons but each one with a terminal event represents a cessation or delay of cambium activity.
The presence of narrow latewoods, such as in these specimens (composed of 2–5 radially compressed cells), probably reflects a rapid onset of unfavourable growth conditions and/or the result of a water deficit during the summer. However, it is not ruled out that it could be associated with the genetic makeup load of the species (Creber and Chaloner 1984; Brea et al. 2005).
A value of 0.3 of mean sensitivity (SM) was obtained for Baieroxylon cicatricum, which corresponds to a complacent wood, equivalent to little marked climatic changes (Brea et al. 2005; Pires and Guerra Sommer 2011).
The presence of false growth rings observed in these woods could be attributed to a cold season, or also by drought, or by defoliation caused by insects (Jefferson 1982; Zamuner 1986). Another anatomical feature with paleoenvironmental significance is the value of the flattening coefficients of the pits. The numbers obtained here indicate openings of considerable size and tracheids with a wide lumen, which denotes that they grew in subtropical conditions with little marked seasons (Creber 1977).
According to these data, it can be inferred that these trees grew in a humid but seasonally dry subtropical climate, in an environment that corresponds to a fluvial system with flood plains and paludal environments. This is suggested by the sandy-limolithic sediments of the Hilario Formation inferred by Baraldo and Guerstein (1984) and Barredo et al. (2016) that are the hosts of the woods of this xylotaphoflora.
Conclusions
The first wood species found in the Hilario Formation are described and assigned to Baieroxylon cicatricum (Ginkgoales) and Protophyllocladoxylon hilarioense sp. nov. (Coniferales: Podocarpaceae). The new specie contributes to the diversity and confirms the presence of the Podocarpaceae.
These taxa are added to the impressions of Neocalamites carrerei and Neocalamites sp. found in this Upper Triassic formation of San Juan province, described by Groeber and Stipanicic (1953). The wood of Baieroxylon cicatricum found in the Hilario Formation constitutes its first occurrence in the Triassic of Argentina, thus adding a new Gondwanan occurrence to those already known from Brazil and India. The species has wide both stratigraphic and geographic ranges occurring from the Permian in sediments of Paraguay, Uruguay, and Namibia to the Cretaceous of India.
From the paleoenvironmental point of view, the type of growth rings observed is consistent with a humid but seasonally dry subtropical climate. False growth rings were probably formed due to drought during the growing season, and/or arthropod damage during the life of these trees. These woods are probably representatives of the arboreal stratum of a mesophytic association where the Sphenophytes found would represent the understory. The described taxa extend the paleobotanical spectrum of the Sorocayense Group, composed of Corystospermales, Gingkoales and Coniferales (Podocarpaceae, Cupressaceae).
Acknowledgements
We extend our thanks to Marion Bamford (University of the Witwatersrand, South Africa) and Marc Philippe (Université Claude Bernard Lyon, France) for their insightful reviews. This work was partially funded by the PI 2018/F013 SGCyT-UNNE. 2018-2022. “Paleobiodiversity of the Carboniferous/Permian–Triassic/Jurassic and Neogene of South America”.
References
Anderson, J.M. and Anderson, H.M. 2003. Heyday of the Gymnosperms: Systematics and Biodiversity of the Late Triassic Molteno Fructifications. Strelitzia 15: 1–398.
Artabe, A., Morel, E., and Spalletti, L. 2001. Paleoecología de las floras triásicas argentinas. In: A.E. Artabe, E. Morel, and A.B. Zamuner (eds.), El Sistema Triásico de Argentina, 199–225. Fundación Museo de La Plata “Francisco Pascasio Moreno”, La Plata.
Artabe, A., Morel, E., and Ganuza, D. 2007a. Las floras triásicas de La Argentina. Ameghiniana 50: 75–86.
Artabe, A., Morel, E., Ganuza, D., Zavattieri, A., and Spalletti, L. 2007b. La paleoflora triásica de Potrerillos, provincia de Mendoza, Argentina. Ameghiniana 44: 279–301.
Bamford, M. and Philippe, M. 2001. Jurassic–Early Cretaceous Gondwanan homoxylous woods: a nomenclatural revision of the genera with taxonomic notes. Review of Palaeobotany and Palynology 113: 287–297. Crossref
Baraldo, J. and Guerstein, P. 1984. Nuevo ordenamiento estratigráfico para el Triásico de Hilario (Calingasta, San Juan). IX Congreso Geológico Argentino, Actas 1: 79–94.
Baraldo, J., Monetta, A., and Soechting, W. 1990. Triásico de San Juan. In: O. Bordonaro (ed.), Geología y Recursos Naturales de la Provincia de San Juan. XI Congreso Geológico Argentino, Relatorio, 124–138. San Juan.
Bardola, P.T., Degani Schmidt, I., Guerra Sommer, M., and Schultz, C. 2009. Lenhos de Ginkgophyta em florestas petrificadas no Triássico Superior Sul-Rio- Grandense, Brasil. Revista Brasileira de Paleontologia 12: 139–148. Crossref
Barredo, S.P. 2012. Geodynamic and tectonostratigrafic study of a continental rift: the Triassic Cuyana Basin, Argentina. In: E. Sharkov (ed.), Tectonics, 99–130. Institute of Geology of Deposits, Petrography, Mineralogy and Geochemistry RAS, Moscow.
Barredo, S., Abarzúa, F., and Banchig, A. 2016. Nueva propuesta estratigráfica para el Triásico del depocentro Agua de los Pajaritos, Precordillera Occidental. Provincia De San Juan. Acta Geológica Lilloana 28: 52–57.
Bodnar, J., Drovandi, J.M., Morel. E., and Ganuza, D.G. 2018. Middle Triassic dipterid ferns from west-central Argentina and their relationship to palaeoclimatic changes. Acta Palaeontologica Polonica 63: 397–416. Crossref
Bodnar, J., Iglesias, A., Colombi, C., and Drovandi, J. 2019. Stratigraphical, sedimentological and palaeofloristic characterization of the Sorocayense Group (Triassic) in Barreal depocenter, San Juan province, Argentina. Andean Geology 46: 567–603. Crossref
Boura, A., Pons, D., Vozenin-Serra, C., and Phú My, B. 2013. Mesozoic fossil wood of Kiên Giang Province, southwestern Vietnam. Palaeontographica B 290: 11–40. Crossref
Brea, M., Matheos, S., Zamuner, A., and Ganuza, D. 2005. Análisis de los anillos de crecimiento del bosque fósil de Víctor Szlápelis, Terciario inferior del Chubut, Argentina. Ameghiniana 42: 407–418.
Brison, A.L., Philippe, M., and Thevenard, F. 2001. Are Mesozoic wood growth rings climate-induced? Paleobiology 27: 531–538. Crossref
Brongniart, A. 1828. Histoire des végétaux fossiles, ou recherches botaniques et géologiques sur les végétaux renfermés dans les diverses couches du globe. 488 pp. Crochard et Comp., Paris. Crossref
Comité Argentino de Estratigrafía 1992. Código Argentino de Estratigrafía. Asociación Geológica Argentina Serie B (Didáctica y Complementaria) 20: 1–64.
Creber, G.T. 1977. Tree rings: a natural data storage system. Biological Review 52: 349–383. Crossref
Creber, G.T. and Chaloner, W.G. 1984. Influence of environmental factors on the wood structure of living and fossil trees. Botanical Review 50: 357–448. Crossref
Crisafulli, A. 2001. Leños permicos de la Formación Yaguarí, Republica Oriental del Uruguay. Ameghiniana 38: 61–72.
Crisafulli, A. and Herbst, R. 2009. Gymnospermous wood (Coniferales, Taxales and Ginkgoales) from the Upper Permian Tacuary Formation, Eastern Paraguay. Paleobiodiversity and Palaeoenvironment 89: 95–109. Crossref
Crisafulli, A. and Herbst, R. 2010. Revisión de algunas lignofloras pérmicas de Namibia, África. In: X Congreso Argentino de Paleontología y Bioestratigrafía y VII Congreso Latinoamericano de Paleontología, La Plata, Argentina. Resúmenes, 151. La Plata.
Drovandi, J.M., Colombi, C.E., Bodnar, J., Ejarque, Y., García, G., Santi Malnis, P., Morel. E., and Alcober, O. 2016. Evidencias preliminares de un bosque destruido por un evento volcaniclástico en el Triásico de Formación El Alcázar, Cuenca Cuyana, San Juan, Argentina. Jornadas de Geología de Precordillera 3, Acta Geológica Lilloana 28 (Suplemento): 94–100.
Drovandi, J.M., Correa, G.A., Bodnar, J., Colombi, C.E., Coturel, E.P., and Morel, E.M. 2020. A new paleofloristic assemblage from the Cuyana Basin (Agua de los Pajaritos depocenter), Argentina and its paleobiogeographic and paleoenvironmental implications. Journal of South American Earth Sciences 104: 102819. Crossref
Dumortier, B.C.J. 1829. Analyse des familles des plantes, avec l’indication des principaux genres qui s’y rattachent. 104 pp. Casterman, Tournay. Crossref
Endlicher, S. 1847. Podocarpeae. In: Synopsis Coniferarum II, 201–228. Schleitlin and Zollikofer, Saint Gallen. Crossref
Engler, A. 1897. Coniferales. In: A. Engler and K. Prantl (eds.), Die natürlichen Pflanzenfamilien II–IV, 144–149. Engelmann, Leipzig.
Erasmus, T. 1976. On the anatomy of Dadoxylon arberi Seward, with some remarks on the phylogenetical tendencies of its tracheids pits. Palaeontologia Africana 19: 127–133. Crossref
Fletcher, T.L., Cantrill, D.J., Moss, P.T., and Salisbury, S.W. 2014. A new species of Protophyllocladoxylon from the Upper Cretaceous (Cenomanian–Turonian) portion of the Winton Formation, central-western Queensland, Australia. Review of Palaeobotany and Palynology 208: 43–49. Crossref
Fritts, H. 1976. Tree Rings and Climate. 571 pp. Academic Press, New York.
Ganuza, D.G., Zamuner, A.B., Artabe, A.E., and Spalletti, L.A. 1998. Sistemática y Paleoecología de la Flora Triásica de Hilario-Agua de Los Pajaritos (Formación El Alcázar), Provincia de San Juan, Argentina. Ameghiniana 35: 271–283.
García Esteban, L., De Palacios de Palacios, P., Guindeo Casasús, A., García Esteban, L., Lázaro Durán I., González Fernández, L., Rodríguez Salvador, Y., Fernández García, S., Bobadilla Maldonado I., and Camacho Atalaya, A. 2002. Anatomía e identificación de maderas de coníferas a nivel de especies. 421 pp. Ediciones Mundi-Prensa. Madrid.
García Esteban, L., Guindeo Casasús, A., Pereza Oramas C., and De Palacios de Palacios, P. 2003. La madera y su anatomía. 327 pp. Ediciones Mundi-Prensa, Madrid.
Gnaedinger, S. 2007. Podocarpaceae woods (Coniferales) from middle Jurassic La Matilde formation, Santa Cruz province, Argentina. Review of Palaeobotany and Palynology 147: 77–93. Crossref
Gnaedinger, S. 2010. Estructuras reproductivas en el Grupo El Tranquilo (Triásico tardío), provincia de Santa Cruz, Argentina. In: X Congreso Argentino de Paleontología y Bioestratigrafía – VII Congreso Latinoamericano de Paleontología Actas 73. La Plata.
Gnaedinger, S. 2012. Ginkgoaleans woods from Jurassic of the Argentina. Taxonomic considerations and palaeobiogeographical distribution. Geobios 45: 187–198 [electronic supplement]. Crossref
Gnaedinger, S. and Herbst, R. 2008. Listado preliminar de estructuras reproductivas del Grupo El Tranquilo, Triásico Superior, Santa Cruz, Argentina. Sesiones de Comunicaciones Científicas y Tecnológicas (SGCYT), RIO-UNNE B-028, Corrientes.
Gnaedinger, S. and Lutz, A. 2008. Comparación del “patrón de venación” de Taeniopteris Brongniart del Triásico Superior de Chile, Argentina y Sud África. Sesiones de Comunicaciones Científicas y Tecnológicas (SGCYT), RIO-UNNE, B-037, Corrientes.
Gnaedinger, S. and Zavattieri, A.M. 2017. First record of voltzialean male cone (Lutanthus) and podocarpacean female cone (Rissikistrobus) from the Late Triassic of Argentina, including new plant remains from the Paso Flores Formation. Ameghiniana 54: 224–246. Crossref
Gorozhankin, I.N. [Gorožankin, I.N.] 1904. Lekcji po morfologji i sistematike arhegonialnyh rastenij. II, Pteridophyta, I, Archispermae. 104 pp. A.I. Mamontov, Moskva.
Gou, X., Wei, H., Guo, Y., Yang S., and Feng, Z. 2021. Leaf phenology, paleoclimatic and paleoenvironmental insights derived from an Agathoxylon stem from the Middle Jurassic of Xinjiang, Northwest China. Review of Palaeobotany and Palynology 289: 104416. Crossref
Greguss, P. 1955. Identification of Gymnosperms on the Basis of Xylotomy. 263 pp. Akadémiai Kiadó, Budapest.
Greguss, P. 1961. Permische fossile Holzer aus Ungarn. Palaeontographica 109: 131–145.
Groeber, P. and Stipanicic, P. 1953. Triásico. In: P.F.C. Groeber (ed.) Mesozoico, Geografía de la República Argentina. Sociedad Argentina de Estudios Geográficos GAEA 2: 13–141.
Gou, X., Wei, H., Guo, Y., Yang S., and Feng, Z. 2021. Leaf phenology, paleoclimatic and paleoenvironmental insights derived from an Agathoxylon stem from the Middle Jurassic of Xinjiang, Northwest China. Review of Palaeobotany and Palynology 289: 104416. Crossref
Hass, H. and Rowe, N.P. 1999. Thin sections and wafering. In: T.P. Jones and N.P. Rowe (eds.), Fossil Plants and Spores: Modern Techniques, 76–81. Geological Society, London.
Heer, O. 1876. Über permische Pflanzen von Fünfkirchen in Ungarn. Mitteilungen aus dem Jahrbuch der königlich ungarischen geologischen Anstalt 5: 3–18.
Holmes, W.B.K. and Anderson, H.M. 2013. The Middle Triassic megafossil flora of the Basin Creek Formation, Nymboida Coal Measures, New South Wales, Australia. Part 9. The genera Heidiphyllum, Voltziopsis, Rissikia and affiliated cones and Yabeiella. Proceedings of the Linnean Society of New South Wales 135: 55–76.
Iamandei, S., Iamandei, E., and Grădinaru, E. 2018. Contributions to the Study of the Early Jurassic petrified forest of Holbav and Cristian Areas (Brașov Region, South Carpathians, Romania). 1st Part. Acta Palaeontologica Romaniae 14: 3–34.
Jefferson, T.H. 1982. Fossil forests from the Lower Cretaceous of Alexander Island, Antarctica. Palaeontology 25: 681–708.
Kokogián, D.A., Spalletti, L.A., Morel, E., Artabe, A.E., Martinez, R.N., Alcóber, O.A., Milana, J.P., Zavattieri, A.M, and Papú, O.H. 1999. Los depósitos continentales triásicos. In: R. Caminos [eds.], Geología Regional Argentina, 377–398. Secretaría de Estado de Minería de la Nación, Buenos Aires.
Kräusel, R. 1939. Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Aegyptens. IV. Die fossilen Flora Aegyptens. Abhandlungen der Bayerische Akademie der Wissenschaften, Mathematisch-Nnaturwissenschaftliche, Neue Folge 47: 1–140.
Leiva Verón, V., Crisafulli, A., Herbst, R., Fillippi Amábile, V., and Molinas, S. 2012. Guavirá, una nueva localidad con maderas fósiles de la Formación Tacuary (Pérmico Superior) de Paraguay. GAEA Journal of Geoscience 8: 67–81. Crossref
Lutz, A. and Herbst, R. 1992. Una nueva especie de Rhexoxylon del Triásico de Barreal, San Juan Argentina. Asociación Paleontológica Argentina, Publicación Especial 2: 73–76.
Maheshwari, H. 1972. Permian wood from Antarctica and revision of some Lower Gondwana wood taxa. Palaeontographica 203: 1–82.
Muralidhar Rao, G. and Ramanujan, K. 1986. Fossil gymnospermous wood with spiral thickening from Gangapur Formation of Andhra Pradesh. In: Proceedings of The Indian Geological Congress, Poona, 119–122. Poona.
Mussa, D. 1958. Conifera fósil do Carbonifero Superior de Santa Catarina. Servicio Grafico do Instituto Brasileiro de Geografia e Estadística. Boletim 182: 1–22.
Oliveira, E. 1936. Dadoxylon derbyi sp. nov. Servicio Geologico e Mineralogico 1: 1–5.
Pant, D. and Singh, V. 1987. Xylotomy of some woods from Raniganj Formation (Permian), Raniganj Coalfield, India. Palaeontographica 203: 1–82.
Philippe, M. 1995. Bois Fossiles du Jurassique de Franche-Comté (NE-France). Palaeontographica 236: 45–103.
Philippe, M. and Bamford, M.K. 2008. A key to morphogenera used for Mesozoic conifer-like woods. Review of Palaeobotany and Palynology 148: 184–207. Crossref
Pires, E.F. and Guerra-Sommer, M. 2011. Growth ring analysis of fossil coniferous woods from Early Cretaceous of Araripe Basin (Brazil). Anais Academia brasilera Ciencias 83: 409–423. Crossref
Pons, D. 1971. Sur un Bois Fossile du Mésozoïque de la Colombie: Protophyllocladoxylon rosablancaense nov. sp. Review of Palaeobotany and Palynology 11: 101–123. Crossref
Pozzo, A. 1948. Estudio geológico, estratigráfico y tectónico de la Precordillera, al este del río de los Patos y al sud de Calingasta (Provincia de San Juan). 101 pp., 12 láms, 3 perf., 1 mapa. Tesis inédita, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad de Buenos Aires, Buenos Aires.
Prasad, M.N.V. 1982. An annotated synopsis of Indian Palaeozoic gymnospermous woods. Review of Palaeobotany and Palynology 38: 119–156. Crossref
Prasad, M. and Lele, K. 1984. Triassic ginkgolean wood from the South Rewa Gondwana Basin, India. Review of Palaeobotany and Palynology 40: 387–397. Crossref
Pujana, R.R., Marenssi, S.A., and Santillana, S.N. 2015. Fossil woods from the Cross Valley Formation (Paleocene of Western Antarctica): Araucariaceae dominated forests. Review of Palaeobotany and Palynology 222: 56–66. Crossref
Pujana, R.R., Santillana, S.N., and Marenssi, S.A. 2014. Conifer fossil woods from the La Meseta Formation (Eocene of Western Antarctica): evidence of Podocarpaceae-dominated forests. Review of Palaeobotany and Palynology 200: 122–137. Crossref
Richter, H.G., Grosser, D., Heinz, I., and Gasson, P.E. 2004. International Association of Wood Anatomists list of microscopic features for softwood identification. IAWA Journal 25: 1–70. Crossref
Ruiz, D.P. and Bodnar, J. 2019. The oldest record of Juniperoxylon, a cupressaceous fossil wood from the Middle Triassic of Argentina. Acta Palaeontologica Polonica 64: 481–488. Crossref
Serbet, R. and Rothwell, G. W. 1995. Functional morphology and homologies of gymnospermous ovules: evidence from a new species of Stephanospermum (Medullosales). Canadian Journal of Botany 73: 650–661. Crossref
Serra, C. 1966a. Étude anatomique et paléogéographique de quelques espèces homoxylées du Sud Viêt-Nam et du Cambodge. Archives Géologique 8: 94–116.
Serra, C. 1966b. Protophyllocladoxylon korubaense nov. sp., bois fossile du Cambodge. Comptes-rendus du Congrès des Sociétés Savantes de Paris et des Départements. Section du Scientifique 91: 199–215.
Spalletti, L. 1995. Los sistemas de acumulación fluviales y lacustres del Triásico de la región occidental de la Precordillera sanjuanina, República Argentina (resumen). Actas II Reunión del Triásico del Cono Sur: 27–28. Bahía Blanca.
Spalletti, L., Artabe, A., Morel, E., and Brea, M. 1999. Biozonación paleoflorística y cronoestratigrafía del Triásico Argentino. Ameghiniana 36: 419–451.
Stipanicic, P.N. 1969. Las sucesiones triásicas argentinas. In: Gondwana Stratigraphy, Proceedings of the 1st International Union of Geological Sciences Gondwana Symposium, 1121–1149. UNESCO, París.
Stipanicic, P.N. 1972. Cuenca Triásica de Barreal. In: A.F. Leanza (ed.), Geología Regional Argentina, 537–566. Academia Nacional de Ciencias, Córdoba.
Stipanicic, P.N. 1979. El Triásico del valle del Río de Los Patos (provincia de San Juan). In: J.C.M. Turner (ed.), Geología Regional Argentina, 695–744. Academia Nacional de Ciencias, Córdoba.
Stipanicic, P.N. 2002. Triásico. In: P. Stipanicic and C. Marsicano (eds.), Léxico Estratigráfico de la Argentina, Vol. VIII. 379 pp. Asociación Geológica Argentina, Buenos Aires.
Taylor, E.L. and Ryberg, P.E. 2007. Tree growth at polar latitudes based on fossil tree ring analysis. Palaeogeography, Palaeoclimatology, Palaeoecology 225: 246–264. Crossref
Taylor, T.N., Taylor, E.L., and Krings, M. 2009. Paleobotany, the Biology and Evolution of Fossil Plants. Second Edition. 1230 pp. Academic Press, Amsterdam.
Torres, T. and Philippe, M. 2002. Nuevas especies de Agathoxylon y Baieroxylon del Lías de La Ligua (Chile) con una evaluación del registro paleoxilológico en el Jurásico de Sudamérica. Revista Geológica de Chile 29: 151–165. Crossref
Townrow, J.A. 1967. On Rissikia and Mataia: podocarpaceous conifers from the lower Mesozoic of southern lands. Papers and Proceedings of the Royal Society of Tasmania 101: 103–131.
Vozenin-Serra, C. 1970. Etude de structures homoxylées à oopores du Mésozoïque vietnamien. Archives Géologiques du Viêtnam 13: 60–71.
Yang, X.J., Wang, Y.D., and Zhang, W. 2013. Occurrences of Early Cretaceous fossil woods in China: implications for paleoclimates. Palaeogeography, Palaeoclimatology, Palaeoecology 385: 213–220. Crossref
Wan, M., Shi, G.R., Luo, M., Lee, S., and Wang, J. 2020. First record of a petrified gymnospermous wood from the Kungurian (late Early Permian) of the southern Sydney Basin, southeastern Australia, and its paleoclimatic implications. Review of Palaeobotany and Palynology 276: 104202. Crossref
Wan, M., Yang, W., and Wang, J. 2019. A new Protophyllocladoxylon wood from the Induan (Lower Triassic) Jiucaiyuan Formation in the Turpan-Hami Basin, southern Bogda Mountains, northwestern China. Review of Palaeobotany and Palynology 267: 62–72. Crossref
Zamuner, A. 1986. Maderas fósiles: Indicadores ambientales? Actas IV Congreso Argentino de Paleontología y Bioestratigrafía 1: 187–194.
Zhang, Y., Wang, J., Liu, L., and Li. N. 2010. Protophyllocladoxylon jingyuanense sp. nov., a gymnospermous wood of the Serpukhovian (Late Mississippian) from Gansu, Northwest China. Acta Geologica Sinica (English Edition) 84: 257–268. Crossref
Zhou, Z.Y. 2009. An overview of the Ginkgoales. Paleoworld 18: 1–22. Crossref
Zhou, Z.Y. and Wu, X.W. 2006. The rise of Ginkgoalean plants in the early Mesozoic: a data analysis. Geological Journal 41: 363–375. Crossref
Zijlstra G. and Philippe, M. 2020. Proposal to conserve the name Protophyllocladoxylon (fossil Coniferophyta: Coniferales) with a conserved type. Taxon 69: 403–413. Crossref
Acta Palaeontol. Pol. 67 (2): 329–340, 2022
https://doi.org/10.4202/app.00939.2021