
Triassic coleoid beaks and other structures from the Calcareous Alps revisited
LARISA A. DOGUZHAEVA, HERBERT SUMMESBERGER, FRANZ BRANDSTAETTER, DANIELA GRUBER, and ANDREA TINTORI
Doguzhaeva, L.A., Summesberger, H., Brandstaetter, F., Gruber, D., and Tintori, A. 2022. Triassic coleoid beaks and other structures from the Calcareous Alps. Acta Palaeontologica Polonica 67 (3): 655–666.
We performed comprehensive study of seven Carnian, Late Triassic specimens of a coleoid cephalopod Phragmoteuthis bisinuata, on which Suess based his hypothesis on “beaks of P. bisinuata”. Using SEM/EDS, we found that “beaks of P. bisinuata” consist of a micro-granular carbonized matrix containing ~4–30 μm diameter and ~50–200 μm visible length, dense calcified bone-like micro-structures. This strongly suggests that these objects are vertebrate bone-inducing cartilages in which the matrix was post-mortem reworked by carbon-accumulating bacteria and substituted by nano-particles of carbon accumulated in micro-granules. Hence, the presumed “beaks of P. bisinuata” are cartilaginous remains of a prey, presumably juvenile fish. This data dismissed the entire hypothesis of Seuss. A small spatula-shape plate with a rachis-like process in an association with 10 or so imprints around (arm crown), found in front of a proostracum of P. bisinuata evidences an unknown Late Triassic juvenile teuthid which possessed a gladius resembling that of the early Permian Glochinomorpha stifeli. It inhabited the open sea area of the northwestern Tethys Ocean, and was, along with juvenile fishes, in the diet of P. bisinuata. The first identified Anisian (Middle Triassic) coleoid beak is represented by an isolated specimen from the Gardena Valley, NE Italy. It has a typical composition and morphology of coleoid upper beak: chitinous, wide-oval lateral walls, short wings, and pointed hook-like rostrum. This suggests similar upper beak structure in the Carnian P. bisinuata in which the lower beaks were apparently similar to that of the co-occurring Lunzoteuthis schindelbergensis and had a widely open outer lamella with posteriorly elongated paired wings joined into a pointed rostrum in the anterior portion.
Key words: Cephalopoda, Coleoidea, Phragmoteuthis, beaks, vertebrate and invertebrate prey, Triassic, Alps.
Larisa A. Doguzhaeva [larisa.doguzhaeva@gmail.com], Department of Palaeobiology, Swedish Museum of Natural History, P.O. Box 50007, Stockholm SE-104 05, Sweden.
Herbert Summesberger [herbert.summesberger@nhm-wien.ac.at], Department of Geology and Palaeontology, Museum of Natural History, Vienna, Burgring 7, Austria.
Franz Brandstaetter [franz.brandstaetter@nhm-wien.ac.at], Mineralogical Department, Museum of Natural History, Vienna, Austria; Vienna, Burgring 7, Austria.
Daniela Gruber [daniela.gruber@univie.ac.at], Core Facility of Cell Imaging and Ultrastructure Research, Life Sciences Faculty, University of Vienna, 1090 Wien Althanstraße 14, Austria.
Andrea Tintori [paleo.tintori@outlook.it], Triassica, Institute for Triassic Lagerstätte, 23828 Periedo (LC), Via al Verder 6, Italy.
Received 21 October 2021, accepted 11 February 2022, available online 22 June 2022.
Copyright © 2022 L.A. Doguzhaeva et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
All known specimens of the Carnian (Late Triassic) proostracum-bearing coleoid cephalopod Phragmoteuthis bisinuata (Bronn, 1859) were discovered during the mid-19th century in two localities in the Calcareous Alps. These are the lower Carnian (Upper Triassic) fish beds in Cave del Predil (NE Italy; formerly Raibl of Austria) and, ~200 km away to northeast, the Raingraben Shale of a coal mining area in Schindelberg (Lunz, Lower Austria) (Bronn 1859; Suess 1865; Mojsisovics 1882) (Fig. 1). The Raingraben Shale originated in paralic coal swamps that led to a deposition of coal beds containing fossils of Carnian plants (Dobruskina 1998).
Fig. 1. Sketch map showing the position of the localities Cave del Predil, NE Italy, and Schindelberg, Lower Austria (asterisks).
Since the first description of Phragmoteuthis Mojsisovics, 1882 (= Belemnoteutis in Bronn 1859; = Acanthoteuthis in Suess 1865) available proostraca from the above two sites were similar to each other, except for a short broadly acute proostracum imprinted on a small fragmentarily preserved breviconic phragmocone of Lunzoteuthis schindelbergensis Doguzhaeva, Summesberger, and Mutvei, 2006 (Doguzhaeva et al. 2006; Doguzhaeva and Summesberger 2012).
Museum specimens of P. bisinuata (Bronn, 1859) are embedded on ~75–150 mm long slabs (Figs. 2A, 3A) exhibiting proostraca, phragmocones, non-biomineralized, postmortem carbonized, belemnoid-type black structures such as ink sacs, arm hooks, and mantle sheets on top of conchs (Bronn 1859; Suess 1865; Mojsisovics 1882). In addition, the slabs contain structures which since the mid-19th century (Suess 1865: pl. 1: 1a, b; pl. 2: 1; Mojsisovics 1882: figs. 1, 4–6; Jeletzky 1966; Rieber 1970: figs. 4.1, 4.2) have been considered “beaks of P. bisinuata”. In the description of P. bisinuata, Mojsisovics (1882: 307) writes: “The beaks are also preserved commonly in a distance from the aperture (Figs. 1, 4–6). They consist of a compact enamel-like glossy black substance. The best preserved specimen shows (Fig. 1) two equally shaped halves with two pointing upwards tubercle-like processes.” This “best preserved specimen” is illustrated by Suess (1865: fig. 1a, b), Rieber (1970: figs. 4.1, 4.2), and herein in Figs. 2C, 3C, 4. Similar beaks have not been reported since their first description by Suess (1865). However, high evolutionary stability of coleoid beaks, in respect to the development of lateral wing-like structures and hook-shape rostrum, has recently become known due to a revealed similarity between a Visean (early Carboniferous) bactritoid-like coleoid from Arkansas, USA, and the present-day Vampyroteuthis Chun, 1903 (Doguzhaeva and Mapes 2017). A broad lower beak in the Carnian (Late Triassic) Lunzoteuthis schindelbergensis from Lunz, Lower Austria, shows the inner and outer lamellae and a fractured rostrum (Doguzhaeva et al. 2006; Doguzhaeva and Summesberger 2012). The latest Anisian (Middle Triassic) isolated upper beak (Fig. 5) from the basal part of the Buchenstein Formation outcrop (Brack et al. 2000; Wotzlaw et. al. 2017) at the top of Secede Mountain in the Gardena valley, NE Italy reported herein, supports the idea of high evolutionary stability of the beak with respect to the presence of lateral wing-like structures and non-biomineralized hook-shaped rostrum in coleoid cephalopods.
Morphological dissimilarity between the “beaks of P. bisinuata” suggested by Suess (1865) and the currently known beaks of fossil and extant coleoids (Clarke 1986; Lu and Ickeringill 2002; Tanabe 2012; Nixon 2015) poses, in our opinion, a problem. It has been unclear so far whether P. bisinuata was a highly specialized coleoid with highly modified beaks; or structures not belonging to P. bisinuata were preserved together with it and mistaken for its beaks. We present ultrastructural and geochemical data on the “beaks of P. bisinuata” that help to reconstruct the material of these “beaks” and thus clarify their true nature.
Institutional abbreviations.—GBA, Geological Survey of Austria, Vienna; NHMW, Museum of Natural History in Vienna; MG, Museum of Gherdeina, Ortisei, NE Italy.
Material and methods
The material comprises original specimens of Bronn (1859) housed at the Museum of the Institute für Geowissenschaften, Heidelberg, Germany; specimens of Suess (1865) and Mojsisovics (1882) stored in the Austrian Geological Survey, Vienna; unpublished specimens exhibiting proostraca associated with arm hooks, ink sacs and mantle sheets from the Cave del Predil, NE Italy, collected in the mid-19th century and also stored in the Austrian Geological Survey; and single specimen of latest Anisian isolated coleoid-type upper beak from the basal part of the Buchenstein Formation recently sampled by AT at Seceda Mountain in the Gardena Valley of the northeastern Italy. Additionally, we examined five cartilaginous cranial capsules extracted from extant Loligo vulgaris (Lamarck, 1798) from the Mediterranean and North seas.
The proostraca of Phragmoteuthis bisinuata with the “beaks” (Figs. 2A, C, 3, 4) or assumed gladius and imprints of the arms (Fig. 2B) in front of them were photographed in photomicroscope at the Department of Geology and Paleontology at the Museum of Natural History in Vienna. The Anisian beak (Fig. 5) from the lower part of the Buchenstein Formation was photographed at the Museum of Ghereina, Ortisei, Italy.

Fig. 2. Coleoid cephalopod Phragmoteuthis bisinuata (Bronn, 1859), overview of four specimens three of which (A, C, D) show the previously thought “beaks of P. bisinuata”, and one (B) exhibits a whitish triangular structure with straight shoulders and a rachis-like process preliminary considered as a gladius of unknown Glochinomorpha-like teuthid; all lower Carnian, Upper Triassic, Cave del Predil, NE Italy. A. GBA 2006/011/0021. B. GBA 2006/011/0028. C. GBA 2006/011/0001, part (C1) and counterpart (C2). Abbreviations: ah, arm hook; atg, assumed triangular posterior part of a gladius of a small squid prey of P. bisinuata; hb, hypothesized “beaks of P. bisinuata”; im, discrete imprints arranged in a shape of an oval in front of proostracum; is, ink sac; lf, lateral field of proostracum; m, mantle debris; ph, phragmocone; pro, proostracum.
Three untreated, ~3–5 mm size, pieces of a black substance of the hypothesized “beaks of P. bisinuata” were mounted on the stubs, coated with gold, dried and analyzed using a coupled SEM/EDS method (Figs. 6, 7, 8–11, 12A). Cartilaginous (Fig. 12C) cranial capsules of Loligo vulgaris were treated by boiling in 5–10% solution of hydroxide peroxide for 5–7 minutes, then dried, mounted on stubs, coated with gold (Fig. 12C2). Coupled SEM/EDS analyses were carried out at the Mineralogical Department in the Museum of Natural History in Vienna and the Core Facility of Cell Imaging and Ultrastructure Research, Faculty of Life Sciences, University of Vienna. Geochemical composition of shale, proostracum, shale on a top of a proostracum in a contact with black sheet (= mantle), a black sheet (= mantle) and ink sac content were analyzed by using Jeol JSM 6400 with a KEVEX energy dispersive detector and CamScan Instrument with LINK (Figs. 6, 7).
Ultrastructural and geochemical data on the material of “beaks of P. bisinuata” was compared with an in-life organic composition of co-occurred ink sacs, arm hooks and mantle of P. bisinuata, as well as, the mantle of the ceratitid Austrotrachyceras sp. of Krystyn (1991), and the lower beak of Lunzoteuthis schindelbergensis (Fig. 12B) (see Doguzhaeva et al. 2004, 2006, 2007a, b; Doguzhaeva and Summesberger 2012). Other comparative materials were cartilaginous cranial capsules of Loligo vulgaris (Fig. 12C); cartilaginous intermediate layer in the gladius of a squid Berryteuthis magister (Doguzhaeva 2018: fig. 6A, B); canalicular cartilage of fin-supported structures and dorsal mantle along gladius of the middle Olenekian (Early Triassic) squid Idahoteuthis parisiana Doguzhaeva and Brayard in Doguzhaeva et al., 2018 (Doguzhaeva et al. 2018: figs. 5A–D, 6A–F, 7A, B); and Vampyroteuthis-like lower beak of the Visean (early Carboniferous) bactritoids-like coleoid (Doguzhaeva and Mapes 2017: figs. 2A, B; 3A, B).
The terms applied for the description of the late Anisian (Middle Triassic) isolated three dimensional beak from the Buchenstein Formation, Seceda Mountain, Gardena Valley, NE Italy, follow Clarke (1986) and Tanabe (2012).
Results
“Beaks of Phragmoteuthis bisinuata”
Morphology.—The structures known as “beaks of Phragmoteuthis bisinuata” (see Suess 1865) are black, small, ~10 mm maximum size, variable in shape, paired or grouped in clusters, structures positioned distantly in front of proostraca and surrounded by the arm hooks in some specimens (Figs. 2A1, 2C, 3A1, 4). They exhibit diverse, complex shapes, with broadly rounded convex and concave parts of different configurations. In some clusters, they have long articulating processes with the swollen rounded ends (Figs. 3, 4) and relatively large rounded “holes” (Fig. 3B), or look like “spirally twisted” (Fig. 3), or are articulated in such a way that one of them enters another (Fig. 4).
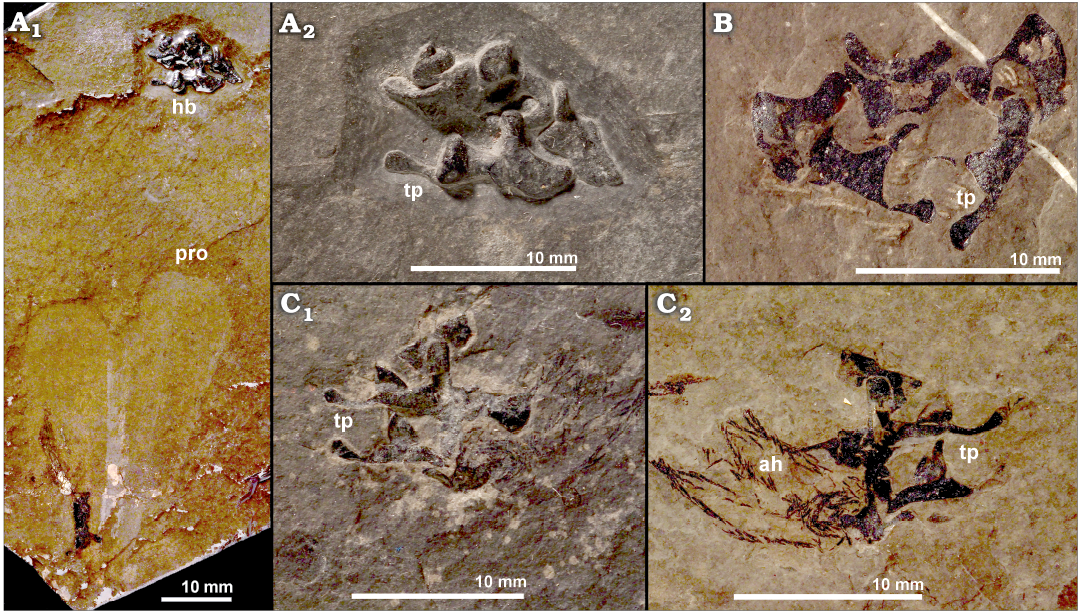
Fig. 3. Coleoid cephalopod Phragmoteuthis bisinuata (Bronn, 1859), overview of the hypothesized “beaks of P. bisinuata”; lower Carnian, Upper Triassic, Cave del Predil, NE Italy. A. GBW 2006/011/0009, an imprint of a proostracum, ink sac and “beaks” (A1), enlarged detail of the “beaks” (A2). B. GBA 2006/011/0041; isolated “beaks”. C. GBW 2006/011/0001, “beaks” associated with the arm hooks (enlarged details of Fig. 2C), part (C1) and counter-part (C2). Abbreviations: ah, arm hook; hb, hypothesized “beaks of P. bisinuata”; pro, proostracum; tp, “tubercle-like processes” of the hypothesized “beak of P. bisinuata”. The “beak” strutures are interpreted herein as the cartilagenous vertebrate remains most likely of juvenile fish that was a prey of P. bisinuata.

Fig. 4. Coleoid cephalopod Phragmoteuthis bisinuata (Bronn, 1859); lower Carnian, Upper Triassic, Cave del Predil, NE Italy; GBW 2006/011/0001; overview of the “beaks” and arm hooks (enlargement of Fig. 2C2). Abbreviations: ah, arm hook; tp, “tubercle-like processes” of the hypothesized “beak of P. bisinuata. The “beaks” structures are interpreted herein as the cartilagenous vertebrate remains of a prey of P. bisinuata which most likely belonged to juvenile fish.

Fig. 5. The isolated upper beak and arm hooks of unknown coleoid (GM-P393) from the Buchenstein Formation, uppermost Anisian, Middle Triassic, Seceda Mountain, Gardena Valley, NE Italy. A1, an overview of the specimen; A2, enlarged arm hook; A3, the anterior part of the upper beak showing a pointed hook-shape rostrum and a deep U-shape inter-space between the lateral walls; A4, S-shape boundary between a rostrum and lateral wall. Abbreviations: ah, arm hook; bl, the curved broken lines consisted from the hook-like structures in the vicinity of the isolated upper beak from the NE Italy; lw, lateral wall of the upper beak; r, rostrum of the upper (dorsal) beak; sah, transversely sectioned arm hook; w, wing of the beak.
Ultrastructure and composition.—Fracturing of the “beaks of P. bisinuata” revealed structureless material with anthracitic luster (Fig. 4), interpreted as representing an in-life organic composition. Coupled SEM/EDS analyses revealed a carbonized, irregularly micro-laminated, micro-granular matrix with a dimpled outer surface, containing numerous non-oriented, complex, ~4–30 µm diameter and ~100–200 µm visible length, bone-like nanostructures of different shapes and sizes, with smooth or papillose surfaces (Figs. 6, 8–11, 12A). This implies a cartilage acted as an inducer of bone formation. Fractured surfaces of the matrix also show numerous imprints of bone-like nanostructures, exhibiting a “porosity” left by papillae on the outer surfaces of the removed bone-like nanostructures.
Fossilization.—The “beaks of P. bisinuata” (Figs. 2A, C, 3A1) similarly to ink sacs, arm hooks and mantle sheets associated with the proostraca of P. bisinuata, were fossilized due to their post-mortem carbonization. It was provided by C-accumulating bacteria which reworked organic remains and thus substituted non-biomineralized structures with nano-granules of carbon accumulated in larger agglomerates (Figs. 4, 6; Doguzhaeva et al. 2007b).
Taphonomy.—The analyzed Carnian “beaks of P. bisinuata” (Figs. 2A, C, 3A1) associated with the proostraca of the P. bisinuata are known from seven specimens, all from the Cave del Predil while the other non-biomineralized structures, ink sacs, arm hooks, mantle debris, are present both in Cave del Predil, NE Italy and Schindelberg, Lower Austria localities. The fish beds of Cave del Predil yielding the “beaks of P. bisinuata” were deposited distantly southwards from the source area in an intra-carbonate platform basin (Fig. 6A3). The Raingraben Shale was deposited at the northern margin of the Tethys near the European continent in a shelf (Fig. 7A; Doguzhaeva et al. 2007b). These relatively shallow-water deposits yielded rare beaks of the ceratitid ammonoids (Trauth 1935; Doguzhaeva et al. 2007a: figs. 11.1A–C, 11.8A–D) and a single, small and flattened beak of coleoid Lunzoteuthis schindelbergensis (Fig. 13; Doguzhaeva et al. 2006: figs. 2, 7A–C). The occurrence of the cartilaginous “beaks P. bisinuata” in the Cave del Predil locality was evidently restricted by the open-sea areal of small fishes which were an easy prey for P. bisinuata. This explains the occurrence of ink sacs, arm hooks and mantle debris of P. bisinuata in both above localities.
Non-biomineralized structures.—Arm hooks, ink sacs, and mantle sheaths were preserved either due to a compact microlaminated ultrastructure of chitin (arm hooks), or fast solidification of ink substituted by the structureless agglomerates of nanoparticles of carbon under protection of broad proostraca (ink sacs), or due to a tight fitting to proostraca.
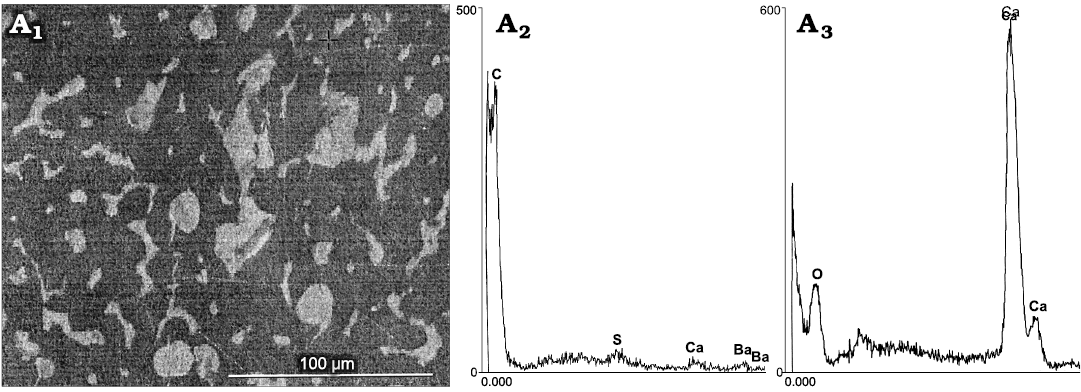
Fig. 6. Hypothesized “beak” of the coleoid cephalopod Phragmoteuthis bisinuata (Bronn, 1859) (GBW 2006/011/0009), lower Carnian, Upper Triassic, Cave del Predil, NE Italy. SEM/EDS data on ultrastructure and geochemical composition of the hypothesized “beak”. A1, a fracture of the “beak” showing a black structureless matrix containing whitish bone-like nanostructures; A2. high peak of C and lower peaks of S, Ca, and Ba, indicative a carbonized matrix (dark grey areas in A1); A3, high peak of Ca, indicative a calcified material of nanostructures (whitish areas in A1).
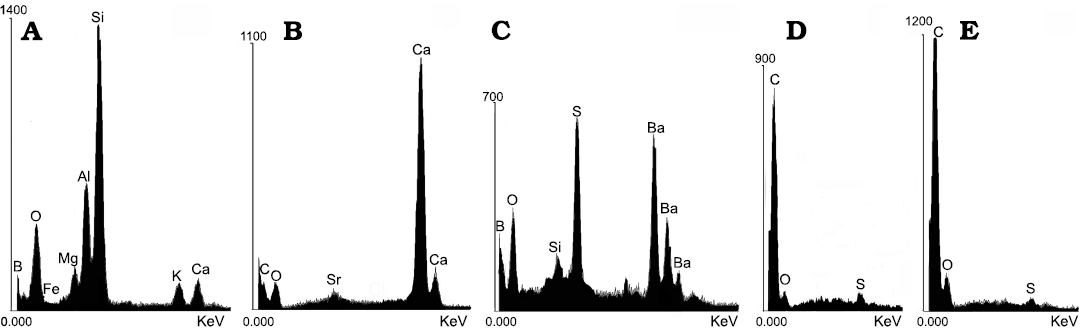
Fig. 7. Coleoid cephalopod Phragmoteuthis bisinuata (Bronn, 1859), lower Carnian, Upper Triassic, Schindelberg, Lower Austria. EDS data on geochemical composition of shale and fossilized remains. A. NHMW 2006z0235/0007, silicified shale. B. NHMW 2006z0235/0012, calcified proostracum. C. NHMW 2006z0235/0010, high content of Ba and S in the shale in intimate contact with black sheet on a top of proostracum, indicative a life time organic material. D. NHMW 2006z0235/0010, carbonized black sheet (mantle) on a top of proostracum. E. NHMW 2006z0235/0007, carbonized ink sac content.

Fig. 8. Hypothesized “beak” of the coleoid cephalopod Phragmoteuthis bisinuata (Bronn, 1859) (GBW 2006/011/0009), lower Carnian, Upper Triassic, Cave del Predil, NE Italy. A fracture of the “beak”. A1, a dimpled outer surface of the “beak” coated by a shale; A2, “glassy” structureless matrix and whitish fractured compressed bone-like nanostructure; A3, dense wall of bone-like nanostructure; A4, a dimpled outer surface of the wall of the bone-like nanostructure.

Fig. 9. A fracture of the hypothesized “beak” of the coleoid cephalopod Phragmoteuthis bisinuata (Bronn, 1859) (GBW 2006/011/0009); lower Carnian, Upper Triassic, Cave del Predil, NE Italy. A1, “nano-porosity” of the matrix of the hypothesized “beak”; A2, A3, bone-like structures (light grey) in the matrix (dark grey) (enlargements of Fig. 8A1, A2); A4, enlarged detail of Fig. 8A3 to show a smooth surface of dense bone-like structure.
Teuthoid gladius
In a single specimen of Phragmoteuthis bisinuata, there is a small, whitish, object located at a place occupied by the “beaks of P. bisinuata” in other specimens (Figs. 2B, 14). It is spatula-like, triangular, pointed posteriorly, having straight shoulders anteriorly and a relatively wide but short rachis-like process with a median notch. This seems to show the inner surface with fine indistinct uneven longitudinal ridges of the discussed object (Fig. 14). This structure is surrounded by at least 10 whitish imprints supposedly left by the arms of the same individual whom it belonged to. The outer surface is unknown.
The spatula-shaped plate and rachis-like process emphasize the similarity between this structure and the gladii of early Permian Glochinomorpha stifeli Gordon, 1971, from the Meade Peak Phosphatic Shale Member of the Phosphoria Formation in the northwest Utah, USA (compare Fig. 14 and Doguzhaeva and Mapes 2015: figs. 1A–H, 2–4). We interpret this specimen to be a juvenile gladius of as yet undescribed Carnian (Late Triassic) Glochinomorpha-like teuthid inhabiting the open sea area of the northwestern Tethys Ocean. The assumed gladius strongly differs from the thin, pointed and slender gladius with rachis, narrow median and lateral fields and breviconic conus of the middle Olenekian (Early Triasssic) Idahoteuthis parisiana Doguzhaeva and Brayard in Doguzhaeva et al., 2018 (Myopsida: Decabrachia) (Doguzhaeva et al. 2018).
Isolated beak from the Gardena Valley
Single late Anisian (Middle Triassic) isolated three dimensional object from the Buchenstein Formation at the Seceda Mountain in Gardena Valley, NE Italy, is here identified as an upper beak of a coleoid. This is small, ~18 mm long and ~8 mm broad beak exposing its left side and the right side is embedded in the rock matrix (Fig. 5A1, A3, A4). Well exposed lateral wall is broadly oval, narrowed posteriorly; width/length ratio is ~0.5; the maximum width is in the posterior one/third of its length; it shows fine fibrous pattern evidencing chitinous composition (Fig. 5A3, A4). Also, a small uppermost part of the right lateral wall is exposed from inside. The exposed wing is short and bordered by a gently curved S-shaped contour (Fig. 5A1, A3, A4). Two lateral walls are dorsally separated by a narrow U-shaped interval. A massive, curved, pointed rostrum is ~7 mm long. A solo, ~2.5 mm arm hook, is positioned near the tip of the rostrum on its lateral side (Fig. 5A2–A4). It is pointed and gently curved into a short stout hollow shaft; the basement is ~0.6 mm. A few small black ring-like structures are exposed on the fractured surface of the sediment and lateral wall of the beak. Judging on rounded one side where the rings are thicker, and the opposite acute side where the rings are thinner, these are transversely fractured arm hooks. There are less distinct arm hooks like structures (supposed arm hooks) arranged along a gently curved line situated at the arm hook near the rostrum (Fig. 5A2–A4).
In modern coleoids the lower beak is usually broader than the upper one (Clarke 1986). A narrow U-shaped interval between the lateral sides shows that the beak is narrow along its total length with nearly parallel lateral walls and therefore is interpreted as an upper (dorsal) one. For comparison, the beak of the Carnian (Late Triassic) Lunzoteuthis schindelbergensis is broad, spoon-like (Fig. 13) and hence interpreted as a lower beak (Doguzhaeva et al. 2006).
Discussion
The interpretation of black structures associated with proostraca of Phragmoteuthis bisinuata as the “beaks” by Suess (1865), Mojsisovics (1882), Jeletzky (1966), and Rieber (1970), was based on their black color (served as an indicator of their in-life organic composition) and their distribution approximately constant intervals and equally distant from the anterior edge of proostraca, which may have resulted from postmortem forward displacement of presumed beaks (Figs. 2A, C, 3A1). However, the ultrastructural and geochemical analyses, here performed for the first time for these structures, have shown that the material of the “beaks of P. bisinuata” is a carbonized substance including ~4–30 μm in diameter and ~50–200 μm in length bone-like micro-structures at the initial stage of bone formation (Figs. 6, 8, 11, 12A). This means a vertebrate cartilage, which acted as an inducer of bone formation (Scott-Savage and Hall 1980; Fig. 12C) and not an invertebrate cartilage (Cole and Hall 2004; Fig. 12C), or chitin of cephalopod beaks (Fig. 12B). Hence, in terms of ultrastructure and composition, the hypothesized “beaks of P. bisinuata” differ from coleoid beaks which consist of ß-chitin-protein complex (Hunt and Nixon 1981) and have dense micro-laminated ultrastructure (Fig. 12B). In fossil coleoids, the micro-laminated ultrastructure is an indicator of the vestigial strong chitinous material of beaks, arm hooks and gladii (Doguzhaeva and Mutvei 2003, 2006; Doguzhaeva et al. 2003, 2006, 2007b, 2018; Doguzhaeva 2012).
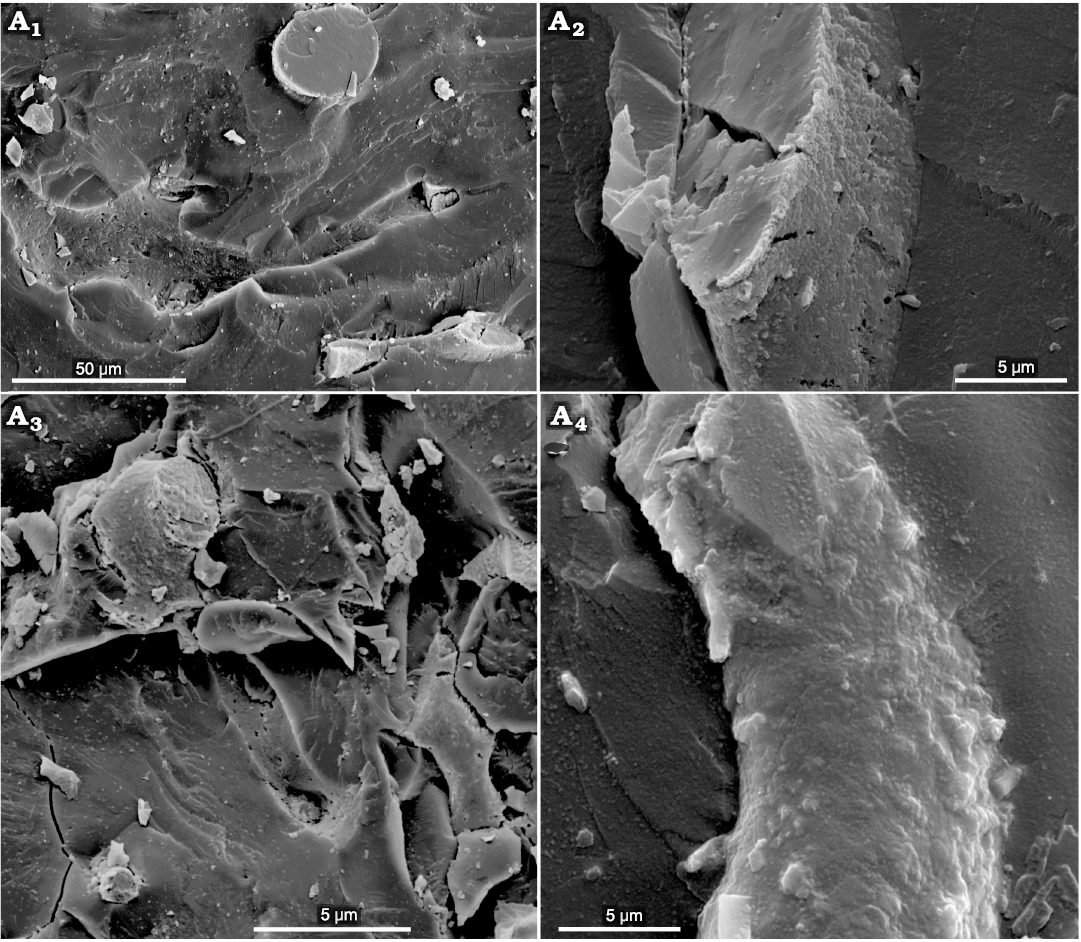
Fig. 10. The ultrastructure of the hypothesized “beak” of the coleoid cephalopod Phragmoteuthis bisinuata (Bronn, 1859) (GBW 2006/011/0009); lower Carnian, Upper Triassic, Cave del Predil, NE Italy. A1, A2, a fracture of the structureless matrix containing bone-like nanostructures or their imprints; A3, a complex bone-like structure within the matrix; A4, a warty outer surface of the bone-like structure; the longest warts leave a “micro-porosity” of the imprints left by bone-like nanostructures.
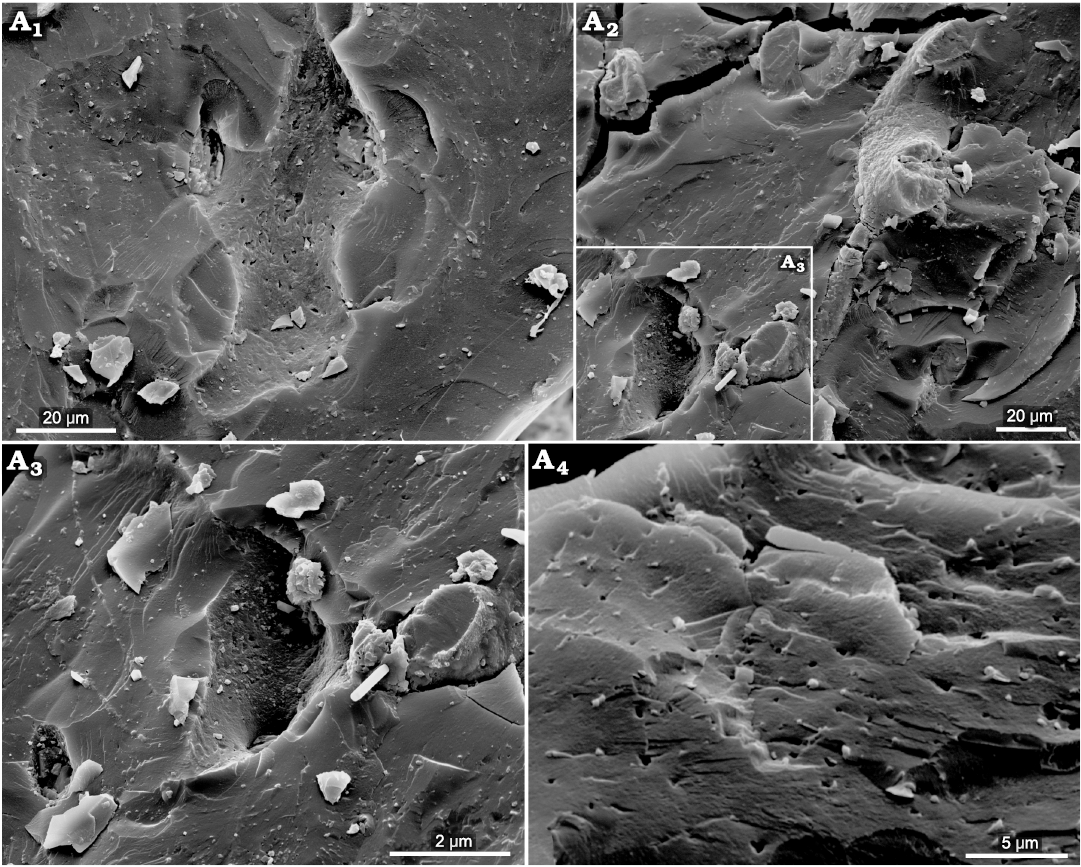
Fig. 11. The ultrastructure of the hypothesized “beak” of the coleoid cephalopod Phragmoteuthis bisinuata (Bronn, 1859) (GBW 2006/011/0009); lower Carnian, Upper Triassic, Cave del Predil, NE Italy. A1, an imprint of a bone-like nanostructure showing “nano-porosity” and three rounded openings left by missing processes (enlarged detail of Fig. 10A1); A2, a semi-ring structure with thin processes ended by swollen rounded part; A3, a deep imprint of a swollen bone-like nano-structure which left two openings of the assumed processes (enlarged detail of A2 marked in the left bottom corner); A4, a mangy skin like micro-lamination and micro-porosity of the matrix.
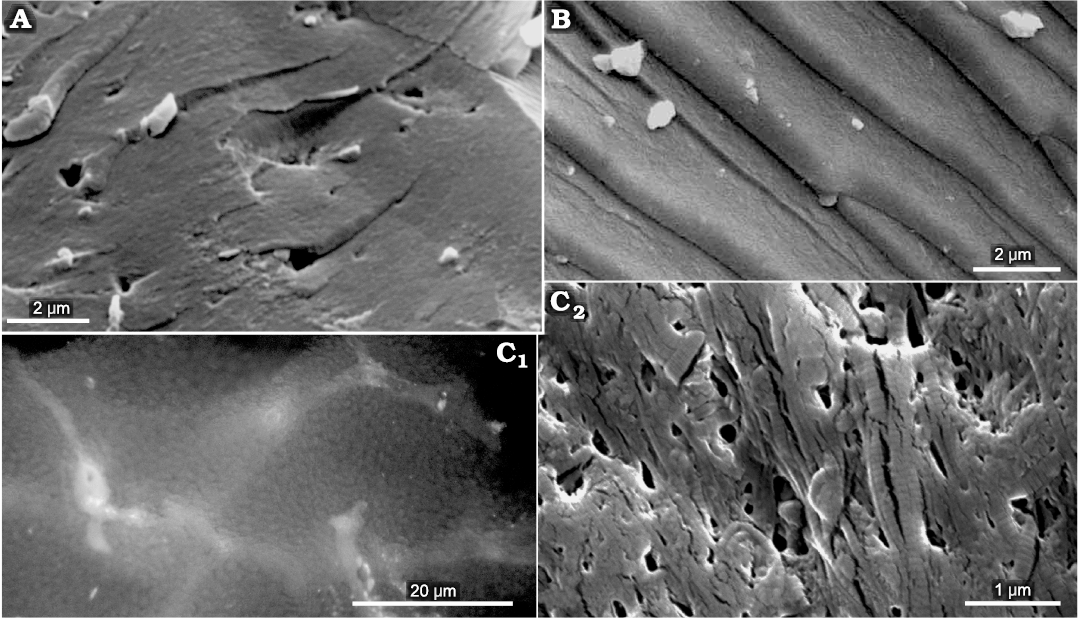
Fig. 12. The comparative ultrastructural data. A. The hypothesized “beaks of Phragmoteuthis bisinuata” (GBW 2006/011/0009, lower Carnian, Upper Triassic; Cave del Predil, NE Italy). B. Chitin of the lower beak of Lunzoteuthis schindelbergensis Doguzhaeva, Summesberger, and Mutvei, 2006 (NHMW 2005z0005/0001, lower Carnian, Upper Triassic; Schindelberg, Lower Austria). C. A cartilage of the cranial capsule of Loligo vulgaris (Lamarck, 1798) (Recent; North Sea); C1, C2, a matrix containing the collagen fibers showing a banding pattern. SEM images, except C1 which is taken with photomicroscope.

Fig. 13. Coleoid cephalopod Lunzoteuthis schindelbergensis Doguzhaeva, Summesberger, and Mutvei, 2006 (holotype, NHMW 2005z0005/0001); lower Carnian, Upper Triassic; Schindelberg, Lower Austria. SEM image of a fractured lower beak in a contact with the phragmocone. Abbreviations: il, inner lamella of the lower beak; ol, outer lamella of the lower beak; ph, phragmocone; r, rostrum of the lower (ventral) beak; w, wing of the beak.

Fig. 14. A small, triangular, pointed posteriorly, gladius, indicative a coleoid prey of Phragmoteuthis bisinuata (Bronn, 1859) (GBA 2006/011/0028); lower Carnian, Upper Triassic, Cave del Predi, NE Italy (enlarged detail of Fig. 2B).

Fig. 15. Coleoid cephalopod Phragmoteuthis bisinuata (Bronn, 1859) (GBW 2006/011/0001); lower Carnian, Upper Triassic, Cave del Predil, NE Italy. Arm hooks with basal parts longer than shafts saving in-life paired arrangement along the arms.
The oldest recovered chitin-compatible material of a marine animal was detected in the late Eocene (~ 34.36 my) cuttlebone of the coleoid Missisaepia missisippiensis Weaver, Dockery III, and Ciampaglio, 2010, from the Mississippi, USA (Weaver et al. 2011). The fin-supporting cartilages of the middle Olenekian (early Triassic) squid of Idahoteuthis parisiana (Decabrachia: Myopsida) from Idaho, USA, are ones of the as yet known oldest fossilized cartilaginous structures (Doguzhaeva et al. 2018).
We also revealed that the site of the “beaks of P. bisinuata” (at the tip of a proostracum) may be occupied with another structures of similar size. These are the gladius and arm crown imprint (Figs. 2B, 14). They apparently represent the remains of another prey of P. bisinuata, also held by its handle hooks. This find points out an as yet unknown teuthid resembling the early Permian Glochinomorpha stifeli Gordon, 1971 (see Doguzhaeva and Mapes 2015: figs. 1A–H, 2, 3).
Thus, in the light of new ultrastructural and geochemical data on the “beaks of P. bisinuata”, morphological data on the upper beak of the reported Anisian (Middle Triassic) coleoid beak from Italy, and previously observed lower beak of co-occurring Lunzoteuthis schindelbergensis, P. bisinuata had hardly differed by its beaks from other coleoids.
The discussed black structures associated with the proostraca of the early Carnian P. bisinuata, previously considered to be the “beaks of P. bisinuata” (Figs. 2A, 2C, 3A1), are known by seven specimens from the Cave del Predil locality, although the number of specimens with other non-biomineralized structures, like ink sacs and arm hooks, is about the same in both localities. The fish beds of Cave del Predil were deposited distantly southwards from the source area in an intra carbonate platform basin and are characterized by high calcium carbonate content (Fig. 6C). The confinement of “beaks of P. bisinuata” to the Cave del Predil locality was probably determined by the presence of small fish and juvenile teuthids, which dwelled at a distance from the shelf and constituted an easy prey for P. bisinuata. Meanwhile, the Raingraben Shale, which did not yield the specimens exhibiting the “beaks of P. bisinuata” but provided chitinous beaks of the ceratitid ammonoids (Trauth 1935; Doguzhaeva et al. 2007a: figs. 11.1A–C, 11.8A–D) and a small flattened solo uncovered beak of coleoid Lunzoteuthis schindelbergensis (Doguzhaeva et al. 2006: figs. 2, 7A–C), was deposited at the northern margin of the Tethys near the European continent in a shelf with river deltas transporting clastics from northern parts of Europe and dominated by silicates rather than carbonates (Fig. 7A).
Thus, the new evidence suggests that the Carnian (Late Triassic) P. bisinuata as well as other phragmocone-bearing and gladius-bearing coleoids had chitinous beaks consisting of a wide lower beak with a rostrum, similar to that of the Carnian Lunzoteuthis schinderlbergensis, and a smaller upper beak with small wings, wide-oval lateral walls and pointed rostrum, like in the isolated Anisian upper beak from NE Italy described herein.
Conclusions
Our study on ultrastructure and geochemical composition of the “beaks of Phragmoteuthis bisinuata” (Suess 1865; Mojsisovics 1882; Jeletzky 1965, 1966; Rieber 1970) showed that they are made of a vertebrate bone-inducing cartilage in which the matrix was post-mortem reworked by carbon-accumulating bacteria and substituted by nano-particles of carbon accumulated in micro-granules. Hence, the “beaks of P. bisinuata” are cartilaginous remains of a prey, evidently juvenile fish, that dismissed the entire hypothesis of Seuss (1865).
Along with small size vertebrates, the diet of P. bisinuata included juvenile teuthids; this is evidenced by a discovery of an as yet unnamed small spatula-shape gladius, associated with an arm crown, located in front of a proostracum of P. bisinuata; it resembles the early Permian Glochinomorpha stifeli Gordon, 1971.
Here reported, first identified Middle Triassic (Anisian) coleoid beak from the Gardena Valley, NE Italy, demonstrates typical coleoid upper beak morphology and composition (chitinous, wide-oval lateral walls, short wings, and pointed hook-like rostrum), that suggests similar upper beak structure for the Late Triassic P. bisinuata. The lower beak of the latter was apparently similar to that of the co-occurring Lunzoteuthis schindelbergensis.
Acknowledgements
Irene Zorn (Austrian Geological Survey, Vienna, Austria) and Katrine Eck (Museum of the Institute für Geowissenschaften, Heidelberg, Germany) granted the access to the original specimens studied by Bronn (1859), Suess (1865), and Mojsisovics (1882). Patricia Weaver (North Carolina Museum of Natural Sciences, Raleigh, USA) and our journal editor Andrzej Kaim made stylistic improvements to the earlier drafts of the manuscript, and Kazushige Tanabe (University of Tokyo Museum, Japan) and Dirk Fuchs (Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany) reviewed the paper and made useful comments. We are deeply indebted to the above organizations and all these people. This study was carried out thanks to financial support of the Austrian and Swedish Academies of Sciences, Department of Geology and Paleontology of the Museum of Natural History in Vienna and the Society “Friends of the Museum of Natural History, Vienna”, Department of Palaeobiology of the Swedish Museum of Natural History, and Museo de Ghereina, Italy.
References
Brack, P., Schlager, W., Stefani, M., Maurer, F., and Kenter, J. 2000. The Seceda drill hole in the Middle Triassic Buchenstein Beds (Livinallongo Formation, Dolomites, Northern Italy). A Progress Report. Rivista Italiana di Paleontologia e Stratigrafia 106: 283–292.
Bronn, H.G. 1859. Nachtrag über die Trias-Fauna von Raibl. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde 1859: 39–45.
Chun, C. 1902–1903. Aus den Tiefen des Weltmeeres. Schilderungen von der DeutschenTiefsee-Expedition. Mit 6 Chromolithographien, 8 Heliogravüren, 32 als Tafelngedruckten Vollbildern, 3 Karten und 482 Abbiludingenim Text. Zweiteumgearbeitete und stark vermehrte Auflage. ix + 592 pp., 46 pls. G. Fischer, Jena.
Clarke, M.R. 1986. A Handbook for the Identification of Cephalopod Beaks. 273 pp. Clarendon Press, Oxford.
Cole, A.G. and Hall, B.K. 2004. The nature and significance of invertebrate cartilages revisited: distribution, and histology of cartilage and cartilage-like tissues within the Metazoa. Zoology 107: 261–273.
Dobruskina, I.A. 1998. Lunz flora in the Austrian Alps—a standard for Carnian floras. Palaeogeography, Palaeoclymatology, Palaeoecology 143: 307–345.
Doguzhaeva, L.A. 2012. The original composition of the pro-ostracum of an Early Sinemurian belemnite from Belgium deduced from mode of fossilization and ultrastructure. Palaeontology 55 (2): 249–260.
Doguzhaeva, L.A. 2018. An early Eocene Antarcticeras nordenskjoeldi: the analysis of the “oegopsid coleoid” hypothesis. Historical Biology 32 (3): 339–345.
Doguzhaeva, L.A. and Mapes, R.H. 2015. Arm hooks and structural features in the Early Permian Glochinomorpha Gordon 1971, indicative of its coleoid affiliation. Lethaia 48: 100–114.
Doguzhaeva, L.A. and Mapes, R.H. 2017. Beak from the body chamber of the Early Carboniferous (Visean) shelled longiconic coleoid (Cephalopoda) from Arkansas, USA. Lethaia 50: 540–547.
Doguzhaeva, L.A. and Mutvei, H. 2003. Gladius composition and ultrastructure in extinct squid-like coleoids: Loligosepia, Trachyteuthis and Teudopsis. Revue de Paléobiologie 22: 877–894.
Doguzhaeva, L.A. and Mutvei, H. 2006. Ultrastructural and chemical comparison between gladii in living coleoids and Aptian coleoids from Central Russia. In: M. Košťák and J. Marek (eds.), Second International Symposium Coleoid Cephalopods Through Time, Prague, 26–29 September 2005. Acta Universitatis Carolinae Geologica 49: 83–93.
Doguzhaeva, L.A. and Summesberger, H. 2012. Pro-ostraca of Triassic belemnoids (Cephalopoda) from northern Calcareous Alps, with observations on their mode of preservation in an environment of northern Tethys that allowed for carbonization of nonbiomineralized structures. Neues Jahrbuch für Geologie und Paläontology Abhandlungen 266: 31–38.
Doguzhaeva, L.A., Brayard, A., Goudemand, N., Krumenacker, L.J., Jenks, J.F., Bylund, K.G., Fara, E., Olivier, N., Vennin, E., and Escarguel, G. 2018. An Early Triassic gladius associated with soft tissue remains from Idaho, USA—a squid-like coleoid cephalopod at the onset of Mesozoic Era. Acta Palaeontologica Polonica 63: 341–355.
Doguzhaeva, L.A., Mapes, R.H., and Mutvei, H. 2003. The shell and ink sac morphology and ultrastructure of the Late Pennsylvanian cephalopod Donovaniconus and its phylogenetic significance. Berliner Paläobiologischen Abhandlungen 3: 61–78.
Doguzhaeva, L.A., Mapes, R.H., Summesberger, H., and Mutvei, H. 2007a. The preservation of body tissues, shell, and mandibles in the ceratitid ammonoid Austrotrachyceras (Late Triassic), Austria. In: N.H. Landman, R.A. Davis, and R.H. Mapes (eds.), Sixth International Symposium, Cephalopods—Present and Past: New Insights and Fresh Perspectives, Chapter 6, 221–238. The Netherlands Springer, Dordrecht.
Doguzhaeva, L.A., Mutvei, H., Summesberger, H., and Dunca, E. 2004. Bituminous soft body tissues in the body chamber of the Late Triassic ceratitid Austrotrachyceras from the Austrian Alps. Mitteilungen aus em Geologisch-Paläontologischen Institut der Universität Hamburg 88: 37–50.
Doguzhaeva, L.A., Summesberger, H., and Mutvei, H. 2006. An unique Upper Triassic coleoid from the Austrian Alps reveals pro-ostracum and mandible ultrastructure. In: M. Košťák and J. Marek (eds.), Second International Symposium Coleoid Cephalopods Through Time, Prague, 26–29 September 2005. Acta Universitatis Carolinae Geologica 49: 69–82.
Doguzhaeva, L.A., Summesberger, H., Mutvei, H., and Brandstaetter, F. 2007b. The mantle, ink sac, ink, arm-hooks and soft body debris associated with the shells in Late Triassic coleoid cephalopod Phragmoteuthis from the Austrian Alps. Palaeoworld 16: 272–284.
Gordon, M., Jr. 1971. Primitive squid gladii from the Permian of Utah. US Geological Survey Professional Paper 750-C: C34–C38.
Hunt, S. and Nixon, M. 1981. A comparative study of protein composition in the chitin-protein complexes of the beaks, pen, sucker discs, radula an oesophageal cuticles of cephalopods. Comparative Biochemistry and Physiology 65B: 535–546.
Krystyn, L. 1991. Die Fossillagerstätten der alpinen Trias. In: D. Nagel and G. Rabider (eds.), Exkursionen im Jungpaläozoikum und Mesozoicum Österreichs, 23–78. Österreich Paläontologischen Gesellschaft, Wien.
Jeletzky, J.A. 1966. Comparative morphology, phylogeny and classification of fossil Coleoidea. University of Kansas Paleontological Contributions, Mollusca 7: 1–162.
Lamarck, J.B. 1798. Prodrome d’une nouvelle classification des coquilles. Memoires de la Société d’Histoire Naturelle de Paris 1: 63–91.
Lu, C.C. and Ickeringill, R. 2002. Cephalopod beak identification and biomass estimation techniques: tools for dietary studies of southern Australian fin fishes. Museum Victoria Science Reports 6: 1–65.
Mojsisovics, E. von 1882. Die Cephalopoden der mediterranen Triasprovinz, III. Dibranchiata. Abhandlungen der Kaiserlich-Königlichen Geologischen Reichsanstalt 10: 295–307.
Nixon, M. 2015. The buccal apparatus of Recent and fossil forms. Treatise Online 69 [published online, https://doi.org/10.17161/to.v0i0.5047].
Rieber, H. 1970. Phragmoteuthis? ticinensis n. sp., ein Coleoidea-Rest aus der Grenzbitumenzone (Mittlere Trias) des Monte San Giorgio (Kt. Tessin, Schweiz). Paläontologische Zeitschrift 44: 32–40.
Scott-Savage, P. and Hall, B.K. 1980. Differentiative ability of the tibial periosteum from the embryonic chick. Acta Anatomy 106: 129–40.
Suess, E. 1865. Über die Cephalopoden-Sippe Acanthoteuthis R. Wagn. Sitzungsberichte der Kaiserlich-Königlichen Akademie der Wissenschaften zu Wien, mathematisch-naturwissenschaftliche Klasse 51: 225–244.
Tanabe, K. 2012. Comparative morphology of modern and fossil coleoid cephalopod jaw apparatuses. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 266: 9–18.
Trauth, F. 1935. Die Aptychen der Trias. Sitzungsberichte der Kaiserlich-Königlichen Akademie der Wissenschaften zu Wien, mathematisch-naturwissenschaftliche Klasse 144: 455–482.
Weaver, P.G., Dockery III, D.T., and Ciampaglio, C.N. 2010. A new genus of coleoid cephalopods from the Jackson Group (Late Eocene), Hinds County, Mississippi. Palaeontographica A 292: 53–65.
Weaver, P.G., Doguzhaeva, L.A., Lawver, D.R., Tacker, R.C., Ciampaglio, C.N., Crate, J.M., and Zheng, W. 2011. Characterization of organics consistent with chitin preserved in the late Eocene cuttlefish Mississaepia mississippiensis. PLoS ONE 6 (11): e28195.
Wotzlaw, J.-F., Brack, P., and Storck, J.-C. 2017. High-resolution stratigraphy and zircon U-Pb geochronology of the Middle Triassic Buchenstein Formation (Dolomites, northern Italy): precession-forcing of hemipelagic carbonate sedimentation and calibration of the Anisian –Ladinian boundary interval. Journal of Geological Society 175 (1): jgs2017-052.
Acta Palaeontol. Pol. 67 (3): 655–666, 2022
https://doi.org/10.4202/app.00953.2021