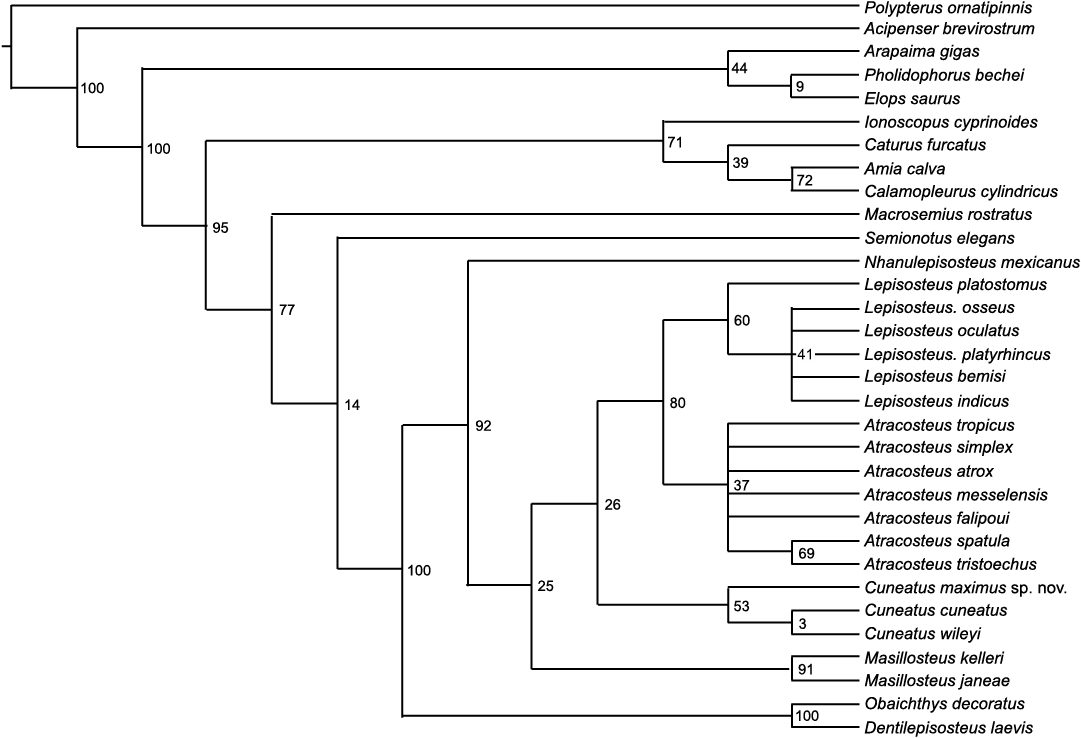Unappreciated Cenozoic ecomorphological diversification of stem gars revealed by a new large species
CHASE DORAN BROWNSTEIN
Brownstein, C.D. 2022. Unappreciated Cenozoic ecomorphological diversification of stem gars revealed by a new large species. Acta Palaeontologica Polonica 67 (3): 559–568.
The evolutionary history of gars, an ancient group of ray-finned fishes, is excellently documented in the fossil record. These fishes are notable for how little the anatomy of extant species differs from that of their earliest known relatives from over 150 million years ago. As such, the low species richness of the gar crown group is thought to reflect the diversity of this clade over most of their history. Here, I describe the skeleton of a new gar species from the Eocene Willwood Formation of Wyoming, USA. Numerous features, including a shortened skull, ornamented external cranial bones, and microteeth ally the new species with Cuneatini, an obscure clade of gars restricted to the Eocene of southwestern North America. Yet, Cuneatus maximus sp. nov. is more than twice as large as its closest relatives. The holotype of the new species preserves a partial palate, providing new information about the anatomy of this poorly known cranial region in cuneatins. Phylogenetic analysis of gars with the new species C. maximus included implies a diversification of cuneatins in North America following the Cretaceous/Paleogene extinction. The presence of large-bodied stem-gars in the Eocene Willwood Formation also suggests that the fish fauna of this region was reminiscent of present-day ones from the American southeast. The discovery of C. maximus emphasizes the propensity of the fossil record to significantly increase the diversity and biogeographic range of even the most depauperate lineages.
Key words: Actinopterygii, Osteichthyes, Cuneatus, lepisosteid, Eocene, USA.
Chase D. Brownstein [chase.brownstein@yale.edu], Department of Ecology and Evolutionary Biology, Yale University, 21 Sachem Street, New Haven, CT 06511, USA; Stamford Museum and Nature Center, Stamford, USA.
Received 3 November 2021, accepted 15 December 2021, available online 20 May 2022.
Copyright © 2022 C.D. Brownstein. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Holostei is one of four major branches along the evolutionary tree of ray-finned fishes (e.g., Grande 2010; Near et al. 2012; Wright et al. 2012; Clarke et al. 2016; Hughes et al. 2018). Yet, only a handful of species in two clades comprise the whole of surviving holostean diversity and disparity. The most diverse extant holosteans are gars, which originated in the middle Mesozoic and have since become restricted to North America (e.g., Grande 2010; Wright et al. 2012; Echelle and Grande 2014; Brito et al. 2017). The fossil record of gars has revealed that the gross anatomy of this lineage has been conserved over their long evolutionary history (e.g., Wiley and Schultze 1984; Grande 1985, 2010; Echelle and Grande 2014; Brito et al. 2017).
The past decade of research into early Cenozoic gar diversity has recognized the presence of at least two extinct clades in the gar family Lepisosteidae with cranial anatomies that differ substantially from extant species (e.g., Grande 2010; Echelle and Grande 2014). These coexisted with early members of the living gar Atractosteus and Lepisosteus in the system of massive lakes that are preserved as the Green River Formation (Wiley and Schultze 1984; Grande 1985, 2010; Echelle and Grande 2014). Unfortunately, the restricted set of fossils known for the short-snouted gars in the clade Cuneatini means that the anatomy and biogeography of these fishes are poorly understood.
Here, I describe the nearly complete skeleton of a large gar from the Willwood Formation of Wyoming, USA. The specimen includes a complete skull that preserves nearly all of the key specializations of cuneatins. These include the presence of packed “microteeth” in place of the large fangs found in extant species, a shortened rostrum, and heavily ornamented dermal skull roof bones (Grande 2010). The specimen is assignable to a new cuneatin that grew to more than twice the size of previously known species, demonstrating that the body size disparity of this clade is undersampled. In addition to expanding the range of this clade by nearly 1000 km, Cuneatus maximus sp. nov. reveals a diversification of cuneatin gars that likely postdated the K/Pg boundary.
Institutional abbreviations.—FMNH, Field Museum of Natural History, Chicago, USA; YPM VPPU, Yale Peabody Museum Vertebrate Paleontology, collections from Princeton University, New Haven, CT, USA.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:093495E9-4517-4713-83C6-BBF7AB96F5D0
Material and methods
The specimen described in this contribution (YPM VPPU. 018063) was recovered from Willwood Formation exposures during a 1962 expedition to Wyoming by Roger C. Wood and includes portions of the surrounding sandstone matrix. In the Yale Peabody Museum collections, the specimen was identified as an individual of Lepisosteus sp. indet.
The new specimen was included in the phylogenetic analysis of Grande (2010) as updated by Brito et al. (2017) to assess its position among ginglymodian fishes (Appendix 1). The phylogenetic matrix to which YPM VPPU.018063 was added included 31 taxa coded for 105 morphological characteristics. I entered the resulting matrix into the program TNT v.1.5 (Goloboff and Catalano 2016) for analysis. An initial Wagner search with space for 1000 trees and default settings for ratchet, tree fuse, tree drift, and sectorial search was employed and followed by traditional bisection-reconnection (TBR) branch-swapping over 100 000 trees to further explore tree islands. The resulting most parsimonious trees were resampled over 100 replicates to generate bootstrap supports for each estimated node (Appendix 2). Finally, a strict consensus tree was used to summarize the most parsimonious topologies. All tree files and morphological matrices used in this study are in the Supplementary Online Material available at http://app.pan.pl/SOM/app67-Brownstein_SOM.pdf.
Geological setting
The Willwood Formation of the Bighorn Basin in northwestern Wyoming represents an ecosystem of flood basins and channels that existed during the latest Paleocene to earliest Eocene epochs of the Paleogene period (e.g., Neasham and Vondra 1972; Bown and Kraus 1981; Smith et al. 2008; Chew 2009). This unit consists of mudstone and fine-grained sandstone rich in quartz, feldspar, and chert (Neasham and Vondra 1972). The red, purple, and brown mudstone of the unit are considered to be overbank paleosols (Kraus and Gwinn 1997).
The Willwood Formation is widely known for its extremely fossiliferous alluvial horizons, which have produced mammal fossils in particularly high quantities (e.g., Rose 1990, 2001; Bown et al. 1994; Chew 2009) and have allowed for several hypotheses regarding the impact of rather short-term climactic and other environmental shifts on faunal change in Paleocene–Eocene mammalian assemblages (Bown et al. 1994; Bao et al. 1999; Wing et al. 2000; Clyde 2001; Gingerich and Clyde 2001; Clyde et al. 2007; Chew 2009). Studies of paleosols in the Willwood Formation suggest that the majority of these fossils likely experienced minimal transport and represent a local vertebrate assemblage (Bown 1979; Bown and Kraus 1981; Bown and Beard 1990; Chew 2009).
Systematic palaeontology
Actinopterygii Cope, 1887
Neopterygii Regan, 1923
Holostei Müller, 1844
Ginglymodi Cope, 1872
Lepisosteiformes Hay, 1929
Lepisosteidae Cuvier, 1825
Cuneatini Grande, 2010
Genus Cuneatus Grande, 2010
Type species: Cuneatus cuneatus Cope, 1878; upper Paleocene–lower Eocene units in Utah, USA.
Cuneatus maximus sp. nov.
Figs. 1–4.
ZooBank LSID: urn:lsid:zoobank.org:pub:093495E9-4517-4713-83C6- BBF7AB96F5D0
Etymology: From Latin maximus, superlative of large, referencing the size of the new species relative to other cuneatins.
Holotype: YPM VPPU.018063, the skeleton of a large-bodied lepisosteid gar consisting of the skull, thorax, and portions of the fins.
Type locality: ~27 km west of Powell, Park County, Wyoming, USA.
Type horizon: Willwood Formation, upper Paleocene to lower Eocene (Bown 1980).
Diagnosis.—Distinguished from other Cuneatus species by: large size (Figs. 1, 4; head length = 181.5 mm; 60–112 mm in Cuneatus cuneatus Cope, 1878 and Cuneatus wileyi Grande, 2010); skull subtriangular in dorsal view, with confluent lateral margins of the preorbital and postorbital portions (Fig. 2); palatal teeth cover the ectopterygoid, vomer, and dermopalatine, with especially large crowns on the lattermost element (Fig. 3); dermal skull roof ornamentation includes extensive pitting (Figs. 1, 4).
Description.—YPM VPPU 0.018063 is the partial skeleton of a large-bodied lepisosteid that has been three-dimensionally preserved in deposits of the Willwood Formation (Fig. 1). The skull of the specimen is still partially encapsulated within the matrix. The size of the skull of the specimen relative to its body length as indicated by the presence of the base of fins at the posterior end of the ventral margin of the specimen shows that the skull/body length ratio was high.

Fig. 1. Holotype of the cuneatin fish Cuneatus maximus sp. nov. (YPM VPPU.018063), from Upper Paleocene to lower Eocene Willwood Formation of Wyoming, USA; dorsal (A1) and ventral (A2) views. Note the ganoid scales characteristic of lepisosteiform fishes and relatives.
The skull roof of Cuneatus maximus sp. nov. is formed by the extrascapulars, frontals, nasals, premaxillae, parietals, and dermopterotics (Fig. 2A1). The supraoccipital bone is absent as in other holosteans. The external surfaces of the posterior frontals, parietals, and extrascapulars show the developed ornamentation characteristic of cuneatin gars (Fig. 2A1, A2). This ornamentation includes extensive pitting, which differs from other species of cuneatins (Fig. 4), the stem-lepisosteid Masillosteus spp., Atractosteus messelensis Grande, 2010, and Atractosteus tropicus (Gill, 1863).
Anteriorly, the premaxillae bear the large marginal pores (Fig. 2A2) characteristic of cuneatin gars (Grande 2010). The premaxillae themselves are shortened elements tightly sutured to the rest of the skull. The olfactory foramina are prominent and positioned anterodorsally. The nasals and frontals are both poorly preserved and obscured by tough matrix. The parietals are widened and subrectangular, contacting the frontals anteriorly and the dermopterotics laterally (Fig. 2A1). These paired elements contact the posterior edge of the skull roof and sit atop the posterior portion of the braincase. The dermopterotics are only partially represented and are the lateral borders of the posterior skull roof. These bones flare outward laterally at their posterior ends and seem to have been similarly rugose in surface texture. The parietals and dermopterotics form a line of bone sitting between the frontals and the posteriormost elements of the skull roof. All these bones are extensively ornamented and form the tight sutural connections characteristic of lepisosteids (e.g., Wiley and Schultze 1984; Grande 1985, 2010; Brito et al. 2017). Posterior to the dermopterotics and parietals, three extrascapulars compose the posterior section of the skull roof (Fig. 2A1). Three extrascapulars are also present in C. wileyi, C. cuneatus, and other lepisosteid species (Grande 2010). All skull roof bones contribute to a subtriangular skull shape in dorsal view with anteromedially tapering lateral margins. In other species of Cuneatus, Atractosteus, and Lepisosteus, the rostrum of the skull is medially offset from the postorbital region in dorsal view (Grande 1985, 2010; Scherrer et al. 2017: fig. 4A).

Fig. 2. Cranial anatomy of the skull of the cuneatin fish Cuneatus maximus sp. nov. (YPM VPPU.018063), from Upper Paleocene to lower Eocene Willwood Formation of Wyoming, USA; dorsal view (A1), detail of the dermal skull roof ornamentation (A2), left (A3) and right (A4) lateral views, detail of the anterior rostrum and dentition in left lateral view (A5), anterior view (A6).
The lateral surface of the skull preserves portions of the premaxillae, the lacrimomaxillae, supraorbitals, and post- and subinfraorbitals (Fig. 2A3–A5). The lacrimomaxillae are large and few in number, contrasting with the condition in crown lepisosteids (Grande 2010). The orbital region is poorly preserved, but several portions of bone below the orbit are identifiable as the strip-like subinfraorbitals. Two ornamented lacrimals are placed anterodorsal to the orbit. Posterior to the orbit, the suborbitals appear as distinct rectangular elements anterior to the opercle. The whole lateral profile of the skull lacks the wedge shape present in C. cuneatus, but whether the posterior mandible was expanded as in both C. cuneatus and C. wileyi is unclear (Grande 2010).
The palate of C. maximus sp. nov. is mostly complete and includes detailed surface texture for many of the anterior bones (Fig. 3). The palatal anatomy of cuneatin gars is somewhat obscure (Grande 2010) so the preservation of these region in YPM VPPU.018063 is particularly important. The anterior palate is formed by the slender vomers medially and the dermopalatines laterally. The anterior half of the ectopterygoid invades the space between these two elements. The expansion of the ectopterygoid to form a large portion of the palate is a distinguishing feature of Lepisosteidae (Grande 2010). All these bones are lined with developed plates of palatal teeth. The palatal tooth crowns appear as clustered papules and are especially large on the vomers and dermopalatines (Fig. 3A2, A3). Similarly extensive palatal tooth plates are known for “Lepisosteus” indicus Woodward, 1890, among lepisosteids. Extant species of Lepisosteus show reduced palatal tooth rows that tend to disappear towards the posterior of the palate (Grande 2010). Given that the palates of obaichthyids, the oldest lepisosteid Nhanulepisosteus mexicanus Brito, Alvarado-Ortega, and Meunier, 2017, and the Cretaceous form Herreraichthys coahuilensis Alvaro-Ortega, Brito, Porras-Múzquiz, and Mújica-Monroy, 2016; are poorly known (Grande 2010; Alvaro-Ortega et al. 2016; Brito et al. 2016, 2017), the polarity of developed palatal tooth rows as a character is unclear. Because previously recognized species of Cuneatus lack any clear palatal teeth (Grande 2010), we consider the condition in YPM VPPU.018063 an autapomorphy of C. maximus.
The lower jaw is very poorly preserved in the holotype of C. maximus sp. nov. Although it is clear both dentaries made up less than 36% of the total skull length (Fig. 3A1) and had medially curved symphyses (Fig. 2A5) as in other Cuneatus species (Grande 2010), not much more can be said about this region. The preserved opercular series consists of the preopercle, the subopercle, and the opercle. As in other lepisosteids with the exception of Atractosteus falipoui (Cavin, Boudad, Tong, Läng, Tabouelle, and Vullo, 2015), there is no interopercle (Grande 2010). The opercular series is eroded in C. maximus, and so the presence of extensive ornamentation could not be assessed. However, it is clear the long anterior ramus (horizontal arm of Grande 2010) characteristic of lepisosteids is present. Posteriorly, the plate-like preopercle articulates with the subopercle, which possesses the large anterodorsal ramus diagnostic for species of Cuneatus (Grande 2010). The opercle appears as a subrectangular plate.

Fig. 3. Palatal and mandibular anatomy of the cuneatin fish Cuneatus maximus sp. nov. (YPM VPPU.018063), from Upper Paleocene to lower Eocene Willwood Formation of Wyoming, USA; ventral view (A1), detail of the anterior palate (A2), toothed ectopterygoid (A3).

Fig. 4. Comparative anatomy of cuneatin skulls. A. Cuneatus maximus sp. nov., YPM VPPU.018063, from Upper Paleocene to lower Eocene Willwood Formation of Wyoming, USA; dorsal view. B, C. An uncatalogued portion of the Cuneatus wileyi Grande, 2010, mass death block (FMNH PF15202) discovered in the YPM collections where it is currently under transfer from FMNH.
The primary dentition consists of the distinctly striated teeth that characterize the family Lepisosteidae (Fig. 2A5; Grande 2010; Echelle and Grande 2014). The folded dentine present in the teeth of gars is known as plicidentine (e.g., Wiley 1976; Grande 2010; Echelle and Grande 2014). The external dentition consists of the numerous small crowns (= microteeth of Grande 2010) characteristic of cuneatins. The three-dimensional preservation of the skull reveals that the rostrum of cuneatins appears mediolaterally widened relative to crown-group gars in anterior view (Fig. 2A5). The anterior rostrum presents an arcade of microteeth, some of which appear very slightly recurved. The dentition of the primary tooth rows of C. maximus is larger than the palatal teeth.
The preserved portion of the body includes numerous ganoid scales (Fig. 1; e.g., Grande 1985, 2010; Echelle and Grande 2014). These are arranged in neat rows along the thorax of the specimen. Each scale is trapezoidal, with their long axes oriented anteroposteriorly as in other lepisosteid gars (Grande 2010; Echelle and Grande 2014). The portions of the fins in C. maximus that are preserved are small, but thy clearly show that as preserved the specimen includes the nearly complete thorax.
Remarks.—YPM VPPU.018063 can be confidently referred to the genus Cuneatus based on the following features: densely ornamented skull bones; numerous small microteeth in the place of fangs; robust anterodorsal process of subopercle; large series of marginal pores on premaxillae; mandibular length is 35–41% of head length (~36%); short, deep lacrimomaxillae.
Stratigraphic and geographic range.—Upper Paleocene to lower Eocene Willwood Formation of Wyoming, USA.
Phylogenetic results
Phylogenetic analysis resolved the new species alongside other lepisosteid gars as expected based on general morphological comparisons. The strict consensus topology of 15 most parsimonious trees (Fig. 5; length 184, consistency index 0.674, retention index 0.901) resolved Cuneatus maximus sp. nov. within a polytomy consisting of all Cuneatus species. The high consistency and retention indices imply that this result is unlikely to derive from morphological homoplasy. Bootstrap values indicate moderate support (53) for a monophyletic Cuneatini including C. maximus and weak support (3) for an ingroup containing only C. wileyi and C. cuneatus. All other major lepisosteid relationships remained consistent with previous studies of this clade (e.g., Grande 2010; Brito et al. 2017). Cuneatini was united by characters 43 (1, microteeth in marginal upper jaw rows) and 66 (2, dermopalatine teeth lost). No autapomorphies were found for C. wileyi, whereas C. maximus was found to have the autapomorphic condition of 66:1 (dermopalatine teeth present but reduced) and C. cuneatus was found to have 88:2 (number of principal caudal fin rays in adults less than 12) as an autapomorphy.
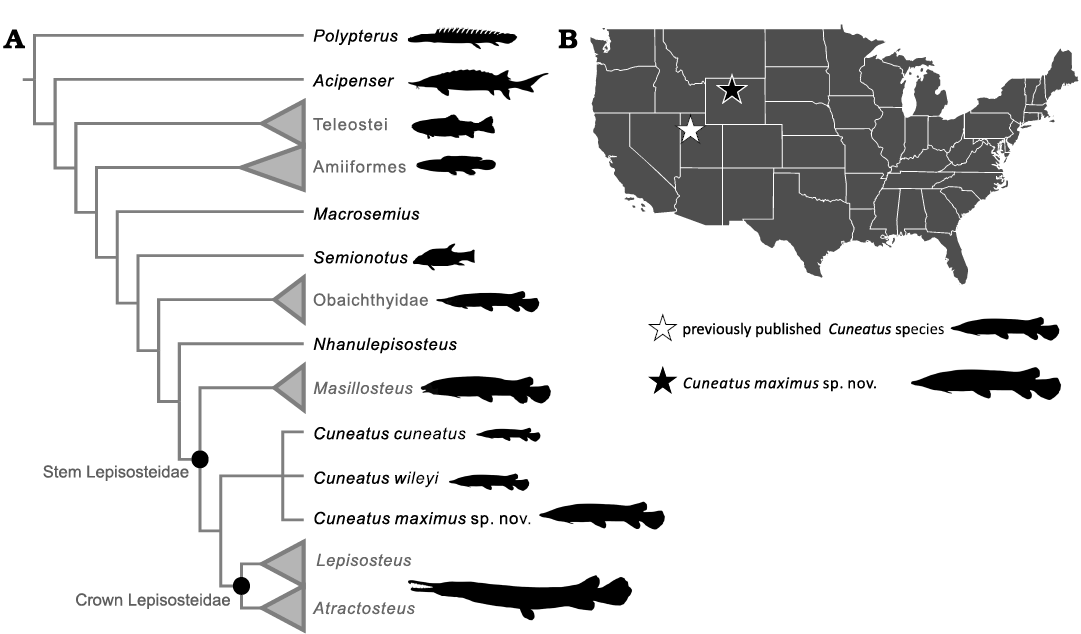
Fig. 5. Phylogeny and biogeography of Cuneatus maximus sp. nov. A. Strict consensus phylogeny generated from parsimony analysis of the 105 character, 32 taxon matrix (Grande 2010; Brito et al. 2017) in TNT v. 1.5 (Goloboff and Catalano 2016). B. Occurrences of Cuneatus maximus sp. nov. and the other two named cuneatins. Grey labels denote collapsed clades that include additional species (see the Appendix 2)
Discussion
Cuneatus maximus sp. nov. is the largest species of cuneatin currently known. Despite clearly belonging to this obscure clade of short-snouted gars, C. maximus possesses features unique among cuneatins that have been used to separate whole genera (i.e., Amia and Cyclurus) and families (Obaichthyidae and Lepisosteidae) in Holostei (Grande 1985; Grande and Bemis 1998; Grande 2010; Echelle and Grande 2014; Brito et al. 2016, 2017). These include the texture of the dermal bones and scales and the presence of numerous rows of palatine teeth (Grande and Bemis 1998; Brito et al. 2000, 2016; Grande 2010). The anatomy of C. maximus therefore implies a relatively high level of unsampled morphological variability in cuneatin gars. Although further large-scale morphological phylogenetic analyses will be needed to validate this hypothesis, the palatal anatomy of C. maximus implies this species is the most basal cuneatin. This interpretation is weakly supported by our majority-rule tree, wherein the southern species C. wileyi and C. cuneatus form a clade.
The discovery of C. maximus sp. nov. is also important for illuminating cuneatin biogeography. Cuneatini is one of three major branches in Lepisosteidae along with the Masillosteinae and crown Lepisosteidae (= Lepisosteini of Grande 2010) yet is restricted to a small region in the western United States (Grande 2010, this paper). This makes cuneatin gars the most regionalized major branch in the gar tree of life. Although the limited distribution of cuneatins might be a result of incomplete sampling, the excellent Cretaceous–Paleogene fossil record of lepisosteids in the northern hemisphere does not include any species identifiable as a cuneatin before the middle Eocene (e.g., Grande 1985, 2010; Echelle and Grande 2014; Cavin et al. 2015; Alvarado-Ortega et al. 2016; Brito et al. 2016; Szabó et al. 2016; Cooper et al. 2021). Thus, it is reasonable to infer that cuneatins were North American endemics.
The fauna of the Willwood Formation features heavily in discussions of vertebrate biogeography during the Eocene Thermal Maximum (ETM; e.g., Smith 2009, 2011; Bourque et al. 2015; Mayr 2016; Scarpetta 2020). Particularly, the Eocene herpetological faunas of this formation have been used to support the hypothesis that the ETM induced large-scale shifts in the distributions of major vertebrate clades (Smith 2009, 2011; Bourque et al. 2015). The presence of cuneatin gars in Wyoming adds to the evidence that the fauna of this region was “megathermal-adapted”, as lepisosteids and members of the sister lineage Obaichthyidae are almost exclusively known from warm environments (e.g., Grande 1985, 2010; Echelle and Grande 2014; Brito et al. 2017). To my knowledge, C. maximus sp. nov. is the only species of fish known only from the Willwood Formation and one of the only articulated fish fossils collected from this unit. Along with the holotype of this new species of gar, I have also examined several specimens referable to medium-sized members of the clade Amiidae (bowfins) from the Willwood Formation. The recognition of this new cuneatin suggests that the poorly known ichthyofauna of the Willwood Formation could provide further support for proposed patterns of migration and endemicity in ETM terrestrial vertebrate faunas.
Cuneatus maximus sp. nov. evidences that lepisosteid disparity in the northern hemisphere remains considerably undersampled. This species was penecontemporaneous with the Green River gar fauna, which includes members of at least five species represented by numerous complete skeletons and a smattering of isolated material (Grande 1985, 2010; Echelle and Grande 2014). This specimen, initially labeled as Lepisosteus sp., illustrates how critical reassessment of previously identified material can drastically change our knowledge of the diversity of depauperate clades.
Acknowledgements
I thank Daniel Brinkman and Vanessa Rhue (both YPM) for access to the collections of the Yale Peabody Museum. Silhouettes in Fig. 5 are taken from phylopic.org. I thank members of the Near Lab for conversations about gar evolution. Finally, I thank the editor and reviewers, Lance Grande (Field Museum of Natural History, Chicago, IL, USA) and Paulo M. Brito (Rio de Janeiro State University, Brazil), for their comments on this manuscript, which greatly improved the final product.
References
Alvarado-Ortega, J., Brito, P.M., Porras-Múzquiz, H.G., and Mújica-Monroy, I.H. 2016. A Late Cretaceous marine long snout “pejelagarto” fish (Lepisosteidae, Lepisosteini) from Múzquiz, Coahuila, northeastern Mexico. Cretaceous Research 57: 19–28. Crossref
Bao, H., Koch, P.L., and Rumble, D. 1999. Paleocene–Eocene climatic variation in western North America: Evidence from the deltals δ18Ο of pedogenic hematite. Bulletin of the Geological Society of America 111: 1405–1415. Crossref
Bourque, J.R., Hutchison, J.H., Holroyd, P.A., and Bloch, J.I. 2015. A new dermatemydid (Testudines, Kinosternoidea) from the Paleocene–Eocene Thermal Maximum, Willwood Formation, southeastern Bighorn Basin, Wyoming. Journal of Vertebrate Paleontology 35: e905481. Crossref
Bown, T.M. 1979. Geology and Mammalian Paleontology of the Sand Creek Facies, Lower Willwood Formation (Lower Eocene), Washakie County, Wyoming. 151 pp. Geological Survey of Wyoming, Laramie.
Bown, T.M. 1980. Summary of latest Cretaceous and Cenozoic sedimentary, tectonic, and erosional events, Bighorn Basin, Wyoming. In: P.D. Gingerich (ed.), Early Cenozoic Paleontology and Stratigraphy of the Bighorn Basin, Wyoming. University of Michigan Papers on Paleontology 24: 25–32.
Bown, T.M. and Beard, K.C. 1990. Systematic lateral variation in the distribution of fossil mammals in alluvial paleosols, lower Eocene Willwood Formation, Wyoming. Geological Society of America Special Paper 243: 135–151. Crossref
Bown, T.M. and Kraus, M.J. 1981. Lower Eocene alluvial paleosols (Willwood Formation, northwest Wyoming, USA) and their significance for paleoecology, paleoclimatology, and basin analysis. Palaeogeography, Palaeoclimatology, Palaeoecology 34: 1–30.
Bown, T.M., Rose, K.D., Simons, E.L., and Wing, S.L. 1994. Distribution and stratigraphic correlation of upper Paleocene and lower Eocene fossil mammal and plant localities of the Fort Union, Willwood, and Tatman formations, southern Bighorn Basin, Wyoming. U.S. Geological Survey Professional Paper 1540: 1–103. Crossref
Brito, P.M., Alvarado-Ortega, J., and Meunier, F.J. 2017. Earliest known lepisosteoid extends the range of anatomically modern gars to the Late Jurassic. Scientific Reports 7 (1): 1–8. Crossref
Brito, P.M., Lindoso, R., Carvalho, I.S., and Machado, G.P. 2016. Discovery of †Obaichthyid gars (Holostei, Ginglymodi, Lepisosteiformes) in the Aptian Codó Formation of the Parnaíba Basin: remarks on paleobiogeographical and temporal range. Cretaceous Research 57: 10–17. Crossref
Brito, P.M., Meunier, F., and Gayet, M. 2000. The morphology and histology of the scales of the Cretaceous gar Obaichthys (Actinopterygii: Lepisosteidae): phylogenetic implications. Comptes Rendus de l’Académie des Sciences 331: 823–829. Crossref
Cavin, L., Boudad, L., Tong, H., Läng, E., Tabouelle, J., and Vullo, R. 2015. Taxonomic composition and trophic structure of the continental bony fish assemblage from the Early Late Cretaceous of southeastern Morocco. PLoS ONE 10 (5): e0125786. Crossref
Chew, A.E. 2009. Paleoecology of the early Eocene Willwood mammal fauna from the central Bighorn Basin, Wyoming. Paleobiology 35 (1): 13–31. Crossref
Clarke, J.T., Lloyd, G.T., and Friedman, M. 2016. Little evidence for enhanced phenotypic evolution in early teleosts relative to their living fossil sister group. Proceedings of the National Academy of Sciences 113: 11531–11536. Crossref
Clyde, W.C. 2001. Mammalian biostratigraphy of the McCullough Peaks area in the northern Bighorn Basin. University of Michigan Papers on Paleontology 33: 109–126.
Clyde, W.C., Hamzi, W., Finarelli, J.A., Wing, S.L., Schankler, D., and Chew, A.E. 2007. A basin-wide magnetostratigraphic framework for the Bighorn Basin, WY. Bulletin of the Geological Society of America 119: 848–859. Crossref
Cooper, S.L., Martill, D.M., Beevor, T., and Gunn, J. 2021. A large marine gar fish (Ginglymodi, Lepisosteiformes) from the Turonian Akrabou Formation of Asfla, Morocco. Cretaceous Research 125: 104839. Crossref
Echelle, A.A. and Grande, L. 2014. Lepisosteidae: gars. In: M.L. Warren Jr. and B.M. Burr (eds.), Freshwater Fishes of North America, Vol. 1, 243–278. John Hopkins University Press, Baltimore.
Gingerich, P.D. and Clyde, W.C. 2001. Overview of mammalian biostratigraphy in the Paleocene–Eocene Fort Union and Willwood formations of the Bighorn and Clarks Fork basins. University of Michigan Papers on Paleontology 33: 1–14.
Goloboff, P. and Catalano, S. 2016. TNT version 1.5, including full implementation of phylogenetic morphometrics. Cladistics 32 (3): 221–238. Crossref
Grande, L. 1985. Paleontology of the Green River Formation, with a review of the fish fauna. Bulletin of the Geological Survey of Wyoming 63: 1–333.
Grande, L. 2010. An empirical synthetic pattern study of gars (Lepisosteiformes) and closely related species, based mostly on skeletal anatomy. The resurrection of Holostei. Copeia 2010 (2A, Supplement): iii–x, 1–871.
Grande, L. and Bemis, W.E. 1998. A comprehensive phylogenetic study of amiid fishes (Amiidae) based on comparative skeletal anatomy. An empirical search for interconnected patterns of natural history. Journal of Vertebrate Paleontology 18: 1–696. Crossref
Hughes, L.C., Ortí, G., Huang, Y., Sun, Y., Baldwin, C.C., Thompson, A.W., Arcila, D., Betancur-R, R., Li, C., Becker, L., and Bellora, N. 2018. Comprehensive phylogeny of ray-finned fishes (Actinopterygii) based on transcriptomic and genomic data. Proceedings of the National Academy of Sciences 115: 6249–6254. Crossref
Kraus, M.J. and Gwinn, B. 1997. Facies and facies architecture of paleogene floodplain deposits, Willwood formation, Bighorn Basin, Wyoming, USA. Sedimentary Geology 114: 33–54. Crossref
Mayr, G. 2016. On the taxonomy and osteology of the early Eocene North American Geranoididae (Aves, Gruoidea). Swiss Journal of Palaeontology 135: 315–325. Crossref
Near, T.J., Eytan, R.I., Dornburg, A., Kuhn, K.L., Moore, J.A., Davis, M.P., Wainwright, P.C., Friedman, M., and Smith, W.L. 2012. Resolution of ray-finned fish phylogeny and timing of diversification. Proceedings of the National Academy of Sciences 109: 13698–13703. Crossref
Neasham, J.W. and Vondra, C.F. 1972. Stratigraphy and petrology of the lower Eocene Willwood Formation, Bighorn Basin, Wyoming. Bulletin of the Geological Society of America 83: 2167–2180. Crossref
Rose, K.D. 1990. Postcranial skeletal remains and adaptations in early Eocene mammals from the Willwood Formation, Bighorn Basin, Wyoming. Geological Society of America Special Paper 243: 107–133. Crossref
Rose, K.D. 2001. Compendium of Wasatchian mammal postcrania from the Willwood Formation. In: P.D. Gingerich (ed.), Paleocene–Eocene Stratigraphy in the Bighorn and Clark’s Fork Basins, Wyoming. University of Michigan Papers on Paleontology 33: 157–183.
Scarpetta, S.G. 2020. Effects of phylogenetic uncertainty on fossil identification illustrated by a new and enigmatic Eocene iguanian. Scientific Reports 10 (1): 1–10. Crossref
Scherrer, R., Hurtado, A., Machado, E.G., and Debiais-Thibaud, M. 2017. MicroCT survey of larval skeletal mineralization in the Cuban gar Atractosteus tristoechus (Actinopterygii; Lepisosteiformes). MorphoMuseum, Association Palaeovertebrata 3 (3): 1–15. Crossref
Smith, J.J., Hasiotis, S.T., Kraus, M.J., and Woody, D.T. 2008. Relationship of floodplain ichnocoenoses to paleopedology, paleohydrology, and paleoclimate in the Willwood Formation, Wyoming, during the Paleocene–Eocene Thermal Maximum. Palaios 23: 683–699. Crossref
Smith, K.T. 2009. A new lizard assemblage from the earliest Eocene (zone Wa0) of the Bighorn Basin, Wyoming, USA: biogeography during the warmest interval of the Cenozoic. Journal of Systematic Palaeontology 7: 299–358. Crossref
Smith, K.T. 2011. The long-term history of dispersal among lizards in the early Eocene: New evidence from a microvertebrate assemblage in the Bighorn Basin of Wyoming, USA. Palaeontology 54: 1243–1270. Crossref
Szabó, M., Gulyás, P., and Ősi, A. 2016. Late Cretaceous (Santonian) Atractosteus (Actinopterygii, Lepisosteidae) remains from Hungary (Iharkút, Bakony Mountains). Cretaceous Research 60: 239–252. Crossref
Wiley, E.O. 1976. The phylogeny and biogeography of fossil and recent gars (Actinopterygii: Lepisosteidae). University of Kansas Museum of Natural History Miscellaneous Publications 64: 1–111.
Wiley, E.O. and Schultze, H.‐P. 1984. Family Lepisosteida (gars) as living fossils. In: N. Eldredge and S.M. Stanley (eds.), Living Fossils, 60–165. Springer Verlag, New York. Crossref
Wing, S.L., Bao, H., and Koch, P.L. 2000. An early Eocene cool period? Evidence for continental cooling during the warmest part of the Cenozoic. In: B.T. Huber, K.G. MacLeod, and S.L. Wing (eds.), Warm Climates in Earth History, 197–237. Cambridge University Press, Cambridge. Crossref
Wright, J.J., David, S.R., and Near, T.J. 2012. Gene trees, species trees, and morphology converge on a similar phylogeny of living gars (Actinopterygii: Holostei: Lepisosteidae), an ancient clade of ray-finned fishes. Molecular Phylogenetics and Evolution 63: 848–856. Crossref
Acta Palaeontol. Pol. 67 (3): 559–568, 2022
https://doi.org/10.4202/app.00957.2021
Codings for Cuneatus maximus sp. nov. in the morphological matrix of Grande (2010) with Nhanulepisosteus included after Brito et al. (2017).
Cuneatus maximus sp. nov. 00?0???1?0?111?1?111??????0???????1?00?11111????0???0???????11?
11????111????????????????????????????????
Resampled tree with bootstrap supports.