
The odonatan insects from the Paleocene of Menat, central France
ANDRÉ NEL and CORENTIN JOUAULT
Nel, A. and Jouault, C. 2022. The odonatan insects from the Paleocene of Menat, central France. Acta Palaeontologica Polonica 67 (3): 631–648.
The current knowledge on the Paleocene Odonata is rather limited despite the fact that it is a crucial period for the history of this order. An overview of the fossil odonatans from the Paleocene of Menat (France) is provided. We describe the anisopteran Macrogomphus menatensis sp. nov., first fossil representative of the family Epigomphidae, together with two zygopteran, viz. the dysagrionid Menatagrion hervetae gen. et sp. nov., and the new family Menatlestidae fam. nov., with its type species Menatlestes palaeocenicus gen. et sp. nov. The genus Menatagrion gen. nov. is the first Paleocene record of the Dysagrionidae, otherwise known by a putative Cretaceous genus and several Eocene to Miocene genera. Menatlestes gen. nov., putatively attributed to the stem-group of the Lestinoidea (Megalestidae and Lestidae), would correspond to the oldest record of this clade. With these three new taxa, and the previously described Thanetophilosina menatensis, Valerea multicellulata, “Lestes” zalesskyi, and an Aeshna species indet., the total number of Odonata from Menat goes up to seven species in total; two Anisoptera and five Zygoptera. Furthermore, we propose new evidences showing that the head characters defining the putative suborder Cephalozygoptera are due to deformations, very frequent among the fossil Odonatoptera. We treat the Cephalozygoptera as a junior synonym of Zygoptera.
Key words: Insecta, Anisoptera, Cephalozygoptera, Zygoptera, head morphology, Auvergne, France.
André Nel [anel@mnhn.fr], Institut Systématique Evolution Biodiversité (ISYEB), Muséum national d’Histoire naturelle, CNRS, Sorbonne Université, Université des Antilles, EPHE, 57 rue Cuvier, CP 50, 75005 Paris, France.
Corentin Jouault [jouaultc0@gmail.com], Institut Systématique Evolution Biodiversité (ISYEB), Muséum national d’Histoire naturelle, CNRS, Sorbonne Université, Université des Antilles, EPHE, 57 rue Cuvier, CP 50, 75005 Paris, France; and Université Rennes, CNRS, Géosciences Rennes, UMR 6118, F-35000, Rennes, France; CNRS, Institut des Sciences de l’Évolution de Montpellier, Place Eugène Bataillon, UMR 5554 34095, Montpellier, France.
Received 12 November 2021, accepted 11 January 2022, available online 31 May 2022.
Copyright © 2021 A. Nel and C. Jouault. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Despite the very important recent discoveries of Odonata of the early Late Cretaceous age, especially in the Burmese amber (Zheng 2020), the Cretaceous/Paleocene transition remains a crucial period for the understanding of the ecological conditions and timing of the diversification of the modern odonatan fauna. The current knowledge on the Turonian–Maastrichtian Odonata is very fragmentary with less than 10 species (Fossilworks Database at http://fossilworks.org/; Nel et al. 2010b), but the recent unexpected discoveries of Maastrichtian Aeschnidiidae confirms the importance of this period for the turnover between the Mesozoic and a more modern fauna (Nel 2021). With 18 described species, the Paleocene Odonata are better known but the majority of taxa have been found in upper Paleocene deposits: Wighton and Wilson (1986) described some Gomphaeschnidae from the Paskapoo Formation (ca. 58 Ma, Alberta, Canada). Lewis (1988) described also a gomphaeschnid from the Tongue River Formation (North Dakota, USA), ca. 60 Ma old (Erickson 1999). The younger (late Paleocene–early Eocene) Maíz Gordo Formation (Argentina) and Fur Formation (Mo clay) of Denmark, have given very diverse odonatan faunas (e.g., Madsen and Nel 1997; Petrulevičius and Nel 2002a, b, 2003a, b, 2004a, b, 2005, 2007, 2009; Bechly and Rasmussen 2019; Petrulevičius 2020). With 66 Oligocene species, and 121 Miocene ones, younger odonatans are better known from fossils found in China, Europe, North and South Americas.
The lacustrine Lagerstätte of Menat (Puy-de-Dôme, France) is among the oldest Paleocene insect outcrops, with an estimated age between 59 and 61 Ma (see Material and methods). It is well known for its very rich and diverse flora (e.g., Laurent 1912; Piton 1940) and fauna (vertebrates and insects). Piton (1940) was first to extensively study this Lagerstätte. Since then, several works have been done on the entomofauna, mainly undertaken during the last twenty years by the team of MNHN (e.g., Nel and Roy 1996; Nel et al. 1997, 2010a; Nel and Auvray 2006; Nel 2007; Wedmann et al. 2018; Garrouste et al. 2017; Schubnel and Nel 2019; Schubnel et al. 2020). The insect fauna is quite diverse, but beetles are the most diverse and abundant group, followed by Dictyoptera and Hemiptera. Despite the fact that Menat is clearly of lacustrine origin, aquatic insects are uncommon, with only two species known so far, a Gyrinidae or whirligig beetles and a Gerridae or water striders, both having an aerial respiration) (Nel 1989; Hartung et al. 2016). The fish fauna is abundant but rather restricted in diversity (Wedmann et al. 2018). No odonatan nymph has been found in Menat to date. Thus the water quality was likely inappropriate for their development and the first stages of the Odonata were probably thriving in the rivers around the lake. The adult odonatans are also extremely rare, with less than 13 known specimens in a collection of more than 4000 fossil insects. Piton (1940) described “Lestes” zalesskyi and an Aeshna species indet. (incomplete abdomen); both are of very uncertain affinities (Nel et al. 1994; Nel and Paicheler 1994). Nel et al. (1997) described the third odonatan from Menat, the megapodagrionid Thanetophilosina menatensis, and Garrouste et al. (2017) described a further damselfly, Valerea multicellulata belonging to the zygopteran Amphipterygidae or Devadattidae.
Here we describe a new anisopteran and two new zygopterans, respectively belonging to the Dysagrionidae and to a new family of Lestinoidea Calvert, 1901.
Note that Archibald et al. (2021) tentatively put Valerea and Thanetophilosina in the Dysagrionidae. Nel and Zheng (2021), restored Valerea in either the families Amphipterygidae Tillyard, 1917, or Devadattidae Dijkstra, Kalkman, Dow, Stokvis, and Van Tol, 2014, on the basis of arguments excluding it from the Dysagrionidae; Archibald and Cannings (2021) maintained it in the Dysagrionidae “for reasons given by Archibald et al. (2021, pages 20 and 42)”, but without any discussion on the arguments given by Nel and Zheng (2021). Therefore we restore it again in either the families Amphipterygidae or Devadattidae on the basis of the arguments that were not discussed by Archibald and Cannings (2021). We also discuss again the head characters supporting the putative suborder Cephalozygoptera.
Institutional abbreviations.—MNHN, Muséum national d’Histoire naturelle, Paris, France; MNT, Musée de Paléontologie de Menat¸ Menat, Puy-de-Dôme, France; PNRL, Parc Naturel régional du Lubéron, Apt, France.
Other abbreviations.—AA/P, anal anterior/posterior; Ax, primary antenodal crossvein; C, costa; CuA/P, cubitus anterior/posterior; d, discoidal cell; h, hypertriangle; IR, intercalary longitudinal veins between main branches of RP; MA/P, median anterior/posterior; N, nodus; Pt, pterostigma; RA/P radius anterior/posterior; ScP, subcostal posterior; t, discoidal triangle.
Nomenclatural acts.—Published work and nomenclatural acts are registered in ZooBank (http://zoobank.org/), with the following LSID: urn:lsid:zoobank.org:pub:074F26DC-1D61-4362-A213-F986D7865FF9.
Material and methods
The middle Paleocene Menat site, near the south of the village of Menat (46°06’ N; 2°54’ E, Menat Basin, Puy-de-Dôme, France), is a volcanic maar corresponding to a rather small palaeolake (ca. 1 km in diameter), filled with spongo-diatomites. This palaeolake contains numerous remains of diverse aquatic and terrestrial flora and fauna (Piton 1940; Nel 1989, 2007; Nel and Roy 1996). The composition of faunal and floral remains suggests that this lake was surrounded by a forest and that the palaeoenvironment was warm and humid (Wedmann et al. 2018). Following the pollen, mammalian stratigraphy, and radiometric K/Ar analyses, its age was estimated as 59 Ma (Kedves and Russel 1982; Nel 2007); vs. an age within 60–61 Ma, based on macroflora (Wappler et al. 2009). Odonatans were already diverse at the time but uncommon in Menat deposit, representing only 0.2% of the very rich and diverse entomofauna, dominated by Coleoptera (78%).
These new discoveries were possible thanks to a very careful examination of all the pieces of rocks during the field and laboratory researches, because the insects are frequently very hard to detect on these rocks. The specimens were prepared with a very sharp pneumatic needle and studied using a stereomicroscope Olympus SCX9 in MNHN. Photographs were taken with a Nikon SMZ 25 stereomicroscope with an attached Nikon D800 camera or using a Canon 50D with a 65 MPE camera lens mounted on an automated stacking rail (StackShot). All images are digitally stacked photomicrographic composites of several individual focal planes, which were obtained using Helicon Focus. The figures were composed with Adobe Illustrator CC2019 and Adobe Photoshop CC2019 software. We follow the wing venation nomenclature of Riek and Kukalová-Peck (1984), modified by Nel et al. (1993), and Jacquelin et al. (2018).
Systematic palaeontology
Odonata Fabricius, 1793
Anisoptera Selys in Selys and Hagen, 1854
Epigomphidae Fraser, 1934 sensu Bechly (2016)
Genus Macrogomphus Selys, 1857
Type species: Macrogomphus annulatus (Sélys-Longchamps, 1854) in Sélys-Longchamps and Hagen (1854); Recent, India, Sri Lanka.
Macrogomphus menatensis sp. nov.
Figs. 1, 2.
ZooBank LSID: urn:lsid:zoobank.org:act:6223E118-CBE0-46CE-AE 4E-BE6562ACB0BA
Etymology: Named after the type locality Menat. The specific epithet is to be treated as an adjective.
Holotype: MNT Nel 4173 (two forewings and a hind wing superposed and attached to fragments of thorax, fragments of abdomen poorly preserved).
Type locality: New quarry, Menat, Puy-de-Dôme, France.
Type horizon: Middle Paleocene, Menat Basin.
Material.—Holotype only.
Diagnosis.—Forewing nodal index 8–13, wings relatively small, hind wing 23.4 mm long.
Description.—Wings hyaline; forewing 25.7 mm long, ca. 5.4 mm wide; distance from base to arculus 3.5 mm, from arculus to nodus 10.1 mm, from nodus to pterostigma 8.0 mm, from pterostigma to wing apex 2.8 mm, from arculus to Ax1 1.1 mm, from Ax1 to Ax2 5.5 mm; Ax2 slightly distal to distal angle of discoidal triangle; four antenodal crossveins between Ax1 and Ax2, seven antenodal crossveins distal of Ax2; eight postnodals; eight crossveins between RA and RP basal of RP3/4; four crossveins between RP and MAa basal of RP3/4; four Bqs veins; oblique vein “O” two cells distal of subnodus; no secondary branch of IR2; pterostigma 2.7 mm long, 0.8 mm wide, covering five cells, with costal margin and RA widened along it; pterostigmal brace present but weak; hypertriangle free, 3.5 mm long; discoidal triangle free isosceles, with costal side 1.5 mm long, basal side 1.5 mm long, distal side MAb 2.1 mm long, MAb without angle; no distinct tspl; RP3/4 and MAa weakly curved and parallel in preserved parts, with one row of cells in-between; two rows of cells in basal part of postdiscoidal area, broadened distally; no posterior branch of MAa; median area free; CuP not visible; subtriangle free as large as discoidal triangle.
Hind wing more poorly preserved than forewings, 23.4 mm long, width unknown because the wing is wrapped; distance from wing base to nodus 9.2 mm, from nodus to pterostigma 9.3 mm, pterostigma 2.4 mm long, 0.7 mm wide.

Fig. 1. Anisopteran odonatan Macrogomphus menatensis sp. nov., holotype MNT Nel 4173; from Paleocene, Menat, France. Imprint (A1), counterimprint (A2), details of pterostigmata (A3, A4). Scale bars 1 mm.
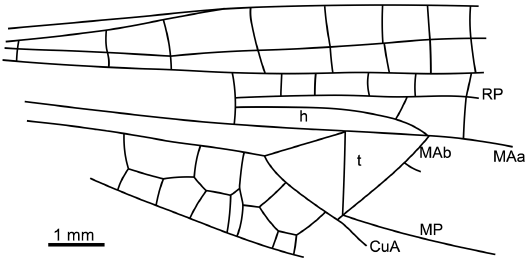
Fig. 2. Reconstruction of forewing base of anisopteran odonatan Macrogomphus menatensis sp. nov., holotype MNT Nel 4173; from Paleocene, Menat, France.
Remarks.—The forewing discoidal triangle equilateral and one-celled excludes affinities with the Aeshnoptera and the Petalurida. The absence of the libellulid gap (basal part of postsubnodal area free of crossveins) and of the cordulegastrid gap (most distal part of the antesubnodal area between RA and RP free of crossveins) exclude affinities with the Cavilabiata Bechly, 1996. The attribution to the clade Gomphida Bechly, 1996, is supported by the discoidal triangles secondarily unicellular in both pairs of wings.
The new fossil has a reduced pterostigmal brace, very weakly oblique, a character of the Epigomphidae. Also, the presence of six crossveins between arculus and fork of RP in the area between RA and RP is a character present in several Epigomphidae, and in the Hageniinae (Gomphidae). The new fossil has a discoidal triangle without a distinct tspl, excluding affinities with the Hageniinae or with the Lindeniinae. The Hageniinae have a costal margin and RA not widened along the pterostigmata, and the discoidal triangles distinctly longitudinally elongate in both pairs of wings unlike the new fossil. The Lindeniinae have a very distinct secondary branch of IR2, therefore IR2 appears to be dichotomously forked distad the lestine oblique vein, unlike in the new fossil.
The status and limits of the Epigomphidae (or Epigomphinae sensu Ware et al. 2017) remains unclear. Nevertheless, within the Epigomphidae sensu Bechly (2016), the new fossil differs from the species of Archaeogomphus Williamson, 1919, Agriogomphus Selys, 1869, Cyanogomphus Selys, 1873, and Lestinogomphus Martin, 1911 (all in Austrogomphinae sensu Carle 1986), Epigomphus Selys, 1854, Eugomphus Kennedy, 1947, and Heliogomphus Laidlaw, 1922, owing to the more numerous crossveins between arculus and RP fork in radial area, larger discoidal triangles and subtriangles (Martin 1911; Williamson 1916, 1919; Fraser 1934, 1943; Kennedy 1936, 1947; Gambles 1968; Legrand and Lachaise 2001). Species of Microgomphus Selys, 1857 (= Africogomphus Fraser, 1936, after Legrand 1993) have a different shape of the forewing discoidal triangle, and the pterostigmata covering only three-four cells, vs. five cells in the new fossil (Fraser 1934: fig. 112, 1936). Species of Leptogomphus Selys, 1878, mainly differ from the new fossil in the absence of a pterostigmal brace (Carle 1986; Dow et al. 2017: figs 130–132). The new fossil better fits with the genus Macrogomphus Selys, 1857, in the shape of the antenodal crossveins, discoidal triangle, supratriangle, subtriangle, number of crossveins basad fork of RP, and pterostigmata (Fraser 1934: fig. 106; Kosterin 2019). We tentatively place it in this genus until discovery of a fossil preserving genital appendages (body parts ensuring an accurate placement).
The nodal index of the new fossil is 8 postnodals–13 antenodals, less than in the extant species of Macrogomphus (9–16 in M. annulatus; 14–20, M. wynaadicus; 12–18, M. montanus; 19–13, M. robustus; 12–15, M. seductus; 10–16, M. lankanensis; 13–18, M. albardae; 11–20, M. rivularis; 9–16, M. borikhanensis; 11–17, M. phalantus) (Fraser 1934; Kosterin 2019). It is also smaller than in the extant species, with a hind wing 23.4 mm long while they range between 33 and 45 mm in the extant species.
Stratigraphic and geographic range.—Type locality and horizon only.
Zygoptera Selys in Selys and Hagen, 1854
Dysagrionidae Cockerell, 1908
Genus Menatagrion nov.
ZooBank LSID: urn:lsid:zoobank.org:act:6EAF1BD6-07B6-4505-B68C-5D15D83A3BE9
Type species: Menatagrion hervetae sp. nov.; see below.
Etymology: Named after the locality Menat and agrion, suffix frequently used for damselflies. Gender masculine.
Diagnosis.—As for the type species by monotypy.
Menatagrion hervetae sp. nov.
Figs. 3–6.
ZooBank LSID: urn:lsid:zoobank.org:act:0545DECE-66A0-49E3-84 03-97668D054455
Etymology: Named after Sophie Hervet, curator of the collections of fossils at the Musée of Menat town. The specific epithet is to be treated as a noun in the genitive case.
Holotype: MNT Nel Odo 1 (imprint and counterimprint).
Type locality: New quarry, Menat, Puy-de-Dôme, France.
Type horizon: Middle Paleocene, Menat Basin.
Material.—Holotype and another specimen MNHN ‘4/10/ 1973, Menat 63/64) imprint and counterimprint of a wing apex (showing the same pattern of coloration and same pterostigma and areas between C and RA and between RA and RP1).
Diagnosis.—Wing venation characters only. Arculus opposite Ax2; three secondary antenodals between C and RA distal of Ax2; base of RP3/4 at basal fourth of space between arculus and nodus; base of IR2 midway between arculus and nodus; two-three rows of cells between RA and RP1 below pterostigma; no pterostigmal brace; one-two rows of cells between C and RA distal of pterostigma; two rows of cells between CuA and posterior wing margin; CuA nearly straight and elongate; no oblique vein “O”; postdiscoidal area greatly widened distally, with two secondary longitudinal veins between MAa and RP3/4.
Description.—Head and legs missing; meso-metathorax and abdomen only partly preserved; one forewing nearly complete with distal half darkened and basal half hyaline; other wings only partly preserved; wing ca. 27.0 mm long, 8.1 mm wide; distance for base to arculus 4.7 mm, from arculus to nodus 5.8 mm, from nodus to pterostigma ca. 16.7 mm, from pterostigma to wing apex 4.1 mm; distance between Ax1 and Ax2 1.4 mm; Ax2 opposite arculus; three secondary antenodals distal of Ax2 between C and RA; nodus of normal shape, without ScP prolonged distad nodus; nodal crossvein and subnodus aligned and of distinct normal obliquity; base of RP3/4 1.4 mm of arculus, at basal fourth of distance between arculus and nodus; base of IR2 midway between arculus and nodus; numerous postnodal crossveins (ca. 25–30), not aligned with postsubnodals; pterostigma elongate, 3.5 mm long, 1.0 mm wide, with basal side with a normal obliquity and a distal side curved, covering seven cells, disposed in two irregular rows, no pterostigmal brace; one-two row of cells between C and RA distal of pterostigma; two-three rows of cells between RA and RP1 for a distance of 8.3 mm basad pterostigma to wing margin; base of RP2 three cells, 2.0 mm distad subnodus; no oblique vein “O”; base of IR1 three cells distad that of RP2; two rows of cells between RP1 and IR1, and between IR1 and RP2; all main veins nearly straight basally and smoothly curved distally; two zigzagged intercalary longitudinal veins between RP2 and IR2, three between IR2 and RP3/4; two between RP3/4 and MAa; one row of cells in basal half of postdiscoidal area and eight along posterior wing margin; one row of cells between MP and CuA; two rows of cells between CuA and posterior wing margin; CuP just distad base of AA; petiole 3.5 mm long, 1.8 mm wide; forewing discoidal cell with basal side 0.4 mm long, anterior side 0.8 mm long, distal side MAb 0.8 mm long, posterior side 1.3 mm long; hind wing discoidal cell with basal side 0.5 mm long, anterior side 1.0 mm long, distal side MAb 0.8 mm long, posterior side 1.0 mm long.

Fig. 3. Zygopteran odonatan Menatagrion hervetae gen. et sp. nov., holotype MNT Nel Odo 1; from Paleocene, Menat, France. Imprint (A1, A2), counterimprint (A3). Scale bars 1 mm.

Fig. 4. Wing of zygopteran odonatan Menatagrion hervetae gen. et sp. nov., holotype MNT Nel Odo 1; from Paleocene, Menat, France. Counterimprint (A1), wing apex, imprint (A2). Scale bars 2 mm.
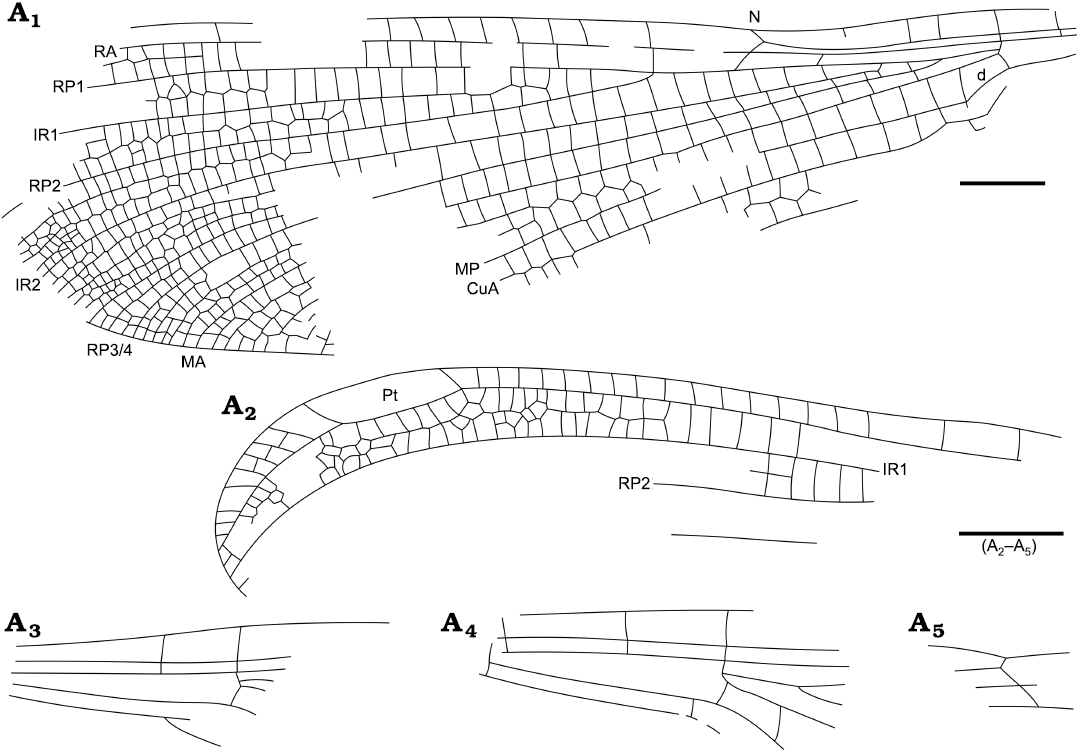
Fig. 5. Reconstructions of wing of zygopteran odonatan Menatagrion hervetae gen. et sp. nov., holotype MNT Nel Odo 1; from Paleocene, Menat, France. Wing (A1), apex (A2), base (A3), base (A4), and nodus (A5) region. Scale bars 2 mm.

Fig. 6. Zygopteran odonatan Menatagrion hervetae gen. et sp. nov., MNHN ‘4/10/1973, Menat 63/64; from Paleocene, Menat, France. Imprint (A1), counterimprint (A2). Scale bars 2 mm.
Remarks.—This new zygopteran differs from Valerea multicellulata in the presence of only one row of cells between C and RA distad the pterostigma and the absence of crossveins ending into the basal side of the pterostigma (Garrouste et al. 2017). It also differs from Thanetophilosina menatensis in the positions of the bases of IR2 and RP3/4, basad midway between arculus and nodus, vs. well distad this point (Nel et al. 1997).
The new Zygoptera has a very broad discoidal cell, a character present in the species of the fossil families Dysagrionidae, Sieblosiidae, Whetwhetaksidae, and Frenguelliidae, known between the Cretaceous and the early Miocene, with a maximum diversity during the Paleogene.
The basal positions of the bases of RP3/4 and IR2 are present in the Sieblosiidae. The new fossil differs from the representatives of this family, except the species of the Oligolestes, in possessing a subnodus of normal obliquity and ScP not passing through the nodus. It is possible that Oligolestes belongs to the stem-group of the Sieblosiidae, as the sieblosiid Paraoligolestes has a wing venation very similar to that of Oligolestes, with the only difference in the structure of the nodal veins (Nel and Escuillié 1993). Other differences between the new fossil and all the Sieblosiidae (including Oligolestes) are the vein CuA nearly straight (vs. distinctly curved), and the absence of the oblique vein “O”.
The new fossil differs from the Whetwhetaksidae owing to the arculus opposite Ax2 (vs. nearly opposite Ax1), and the presence of secondary antenodals. However, they share the bases of RP3/4 and IR2 closer to arculus than to nodus, the absence of the oblique vein “O”, and CuA less curved than in the Sieblosiidae. The Frenguelliidae have an oblique vein “O”, a vertical subnodus, and the base of IR2 opposite the subnodus, configuration not found in the new fossil.
The new fossil preserves several diagnostic characters of the Dysagrionidae, as proposed by Archibald et al. (2021: 16–17): oblique crossvein “O” absent; arculus at or immediately proximad Ax2 (here opposite Ax2); discoidal quadrangle broad, with distal side longer than proximal side, posterior side longer than anterior, distal-posterior angle oblique, proximal-anterior angle usually about 90°; nodus positioned at least a quarter wing length, usually more (here nearly at a third); vein AA separating from AP briefly before joining CuP (a character also present in many Lestoidea); CuA–A space expanded in middle to at least two cells wide, often more (here two rows of cells); CuA long, terminating on posterior margin at mid-wing or longer. Nevertheless, the new fossil differs from the species of Dysagrionidae in an important character also present in the species of Sieblosiidae, viz. RP3–4 originating at one quarter length from arculus to subnodus, vs. roughly between one third to two thirds (usually about two thirds). Also all of the species of Dysagrionidae, except Congqingia rhora and the Petrolestes spp., have the base of IR2 below the subnodus. In the two latter genera and the new fossil, the base of IR2 is midway between the arculus and the nodus. The new fossil and Congqingia rhora differ from the species of Petrolestes in the cubital area with only two rows of cells. The new fossil differs from both Congqingia rhora and the species of Petrolestes in the postdiscoidal area greatly widened distally, with two secondary longitudinal veins between MAa and RP3/4 (vs. with one-two rows of cells in-between). Such a widening of the posdiscoidal area is also found in the other dysagrionid genera.
Archibald et al. (2021: 17) listed the absence of secondary antenodals as a diagnostic characters of the Dysagrionidae. But they also included in this family the two genera Phenacolestes and Electrophenacolestes that have three secondary antenodal crossveins distal of Ax2, between C and RA. This character is also present in the new fossil. It differs from Phenacolestes and Electrophenacolestes in the narrower cubital area with only two rows of cells (vs. three-four rows of cells), and the base of IR2 midway between arculus and nodus (vs. below subnodus).
In conclusion, we tentatively place the new fossil in the Dysagrionidae, even if it clearly differs from all the genera and species currently included in this family. As it also shares some characters with the Sieblosiidae, it is only after a new phylogenetic analysis of all these damselflies that its relationships will be clarified.
Stratigraphic and geographic range.—Type locality and horizon only.
Lestinoidea Calvert, 1901 (sensu Bechly 2016)
Family Menatlestidae nov.
ZooBank LSID: urn:lsid:zoobank.org:act:1F1AB163-E015-449B-862D-FF08A8233D56
Type genus: Menatlestes gen. nov.
Genera included: Type genus only.
Diagnosis.—Wing venation characters. Discoidal cell narrow, with elongate anterior and distal sides, and a very acute postero-distal angle; subdiscoidal cell very long and narrow, posteriorly closed; base of RP3/4 midway between arculus and nodus; base of IR2 at distal third between arculus and nodus, well basal of subnodus; Ax2 opposite arculus; three secondary antenodals distal of Ax2; numerous postnodals, not aligned with postsubnodals; pterostigma elongate, with an oblique pterostigmal brace; cubital area very narrow; no oblique vein “O”.
Genus Menatlestes nov.
ZooBank LSID: urn:lsid:zoobank.org:act:618A75FA-3552-49D2-97 EC-7067F5DA3442
Type species: Menatlestes palaeocenicus sp. nov.; see below.
Etymology: Named after the type locality Menat, and the genus name Lestes. Gender feminine.
Diagnosis.—As for the type species by monotypy.
Remarks.—The presence of a very oblique distal side MAb of the discoidal cell excludes an attribution to the Dysagrionidae, Sieblosiidae, and Whetwhetaksidae. The absence of an oblique crossvein “O” further excludes affinities with the Sieblosiidae. The base of vein RP3/4 basad midway between arculus and nodus, and that of IR2 just distad the latter, plus the elongate pterostigma exclude affinities with the Eocene Frenguelliidae and with the majority of the Coenagrionomorpha (Petrulevičius and Nel 2003b, 2007; Bechly 2016). These characters are present in some Caloptera and Lestomorpha. In the Coenagrionomorpha, only the Hypolestidae have rather elongate pterostigma as in Menatlestes gen. nov. In this family, only the Hypolestinae share with Menatlestes gen. nov. the bases of IR2 and RP3/4 midway between arculus and nodus, unlike Heteragrion and Philogenia (Münz 1919). The genus Philosina is excluded because it has a broad cubital area, and base of IR2 below subnodus (Ris 1917). Rhipidolestes and Lestomima have a base of IR2 well basad the subnodus, but both differ from Menatlestes gen. nov. in the basal side of the pterostigma strongly oblique, a very long anterior side of the discoidal cell, quite longer than MAb, and a broader cubital area (Münz 1919; May 1933). The Eocene Anglohypolestes and Eohypolestes, Oligocene Prohypolestes, and the extant Hypolestes share with Menatlestes gen. nov. the bases of IR2 and RP3/4 midway between arculus and nodus, plus general patterns of venation, including a narrow cubital area (unknown in Eophypolestes), but all differ from the new fossil in having a shorter pterostigma and a very long anterior side of the discoidal cell, quite longer than MAb (Calvert 1893; Münz 1919; Nel and Paicheler 1994; Nel and Fleck 2014).
Affinities with nearly all the Caloptera could be excluded because of the presence of a clearly oblique pterostigmal brace and long petiole in Menatlestes gen. nov. Only the Pseudolestidae show similarities with Menatlestes gen. nov. in the presence of a long petiole, bases of IR2 and RP3/4 between arculus and nodus, and narrow cubital area. However, Pseudolestes has the bases of IR2 and RP3/4 very close to arculus and a MAb short and not strongly oblique, unlike Menatlestes gen. nov.
Indeed, a strongly oblique MAb is only found in some Lestomorpha, supporting an attribution of Menatlestes gen. nov. to this clade. The Cretaceous Protohemiphlebiidae and the Hemiphlebiidae are excluded because they have RP1 kinked at the insertion of the pterostigmal brace vein and all intercalary veins suppressed, unlike Menatlestes gen. nov. (Zheng et al. 2021). Nevertheless the Cretaceous Cretahemiphlebia has a discoidal cell with an elongate and strongly oblique MAb and a very narrow subdiscoidal cell very similar to those of Menatlestes gen. nov. (Jarzembowski et al. 1998). The Cretaceous Cretacoenagrionidae (Cretacoenagrion Jarzembowski, 1990) have the base of IR2 below the arculus. The Cretaceous Lestoidea of uncertain affinities Cretalestes Jarzembowski, Martinez-Declos, Bechly, Nel, Coram, and Escullié, 1998, has also a very narrow subdiscoidal cell and a strongly oblique MAb, but its anterior side of discoidal cell is very short and the base of RP2 is aligned with the subnodus, unlike Menatlestes gen. nov. (Jarzembowski et al. 1998). The Eolestidae (Eocene Eolestes) have a discoidal cell very similar to that of Cretalestes. An additional difference with Menatlestes gen. nov. is the broad area between MP and CuA with four rows of cells, instead of one in Menatlestes gen. nov. (Greenwalt and Bechly 2014). The Eocene Lutetialestes Greenwalt and Bechly, 2014, also differs from Menatlestes gen. nov. owing to a similar character (two rows of cells between MP and CuA, even if some fossil and extant Lestes Leach, 1815, have also two rows, while others have only one row of cells). It also differs from Menatlestes gen. nov. in having a shorter anterior side of the discoidal cell. The Eocene Austroperilestidae share with Menatlestes gen. nov. a very long and strongly oblique MAb and a very long pterostigma, but the anterior side of its discoidal cell is quite short; also the base of IR2 is below subnodus and it has two rows of cells in cubital area (Petrulevičius and Nel 2005).
The Chorismagrionidae (Chorismagrion) and the Perilestidae (Perilestes, Nubiolestes, Perissolestes) have a posteriorly opened subdiscoidal cell, a short pterostigma, and the base of RP3/4 below or distad the subnodus (Münz 1919; Kennedy 1941; Schmidt 1942). The Synlestidae have also posterior margin of the subdiscoidal cell mostly fused to the posterior margin of wing. Synlestes Sélys-Longchamp, 1869, has a discoidal cell very similar to that of Menatlestes gen. nov., but the base of its RP3/4 is located below the nodus. The Lestinoidea (Megalestidae and Lestidae) share with Menatlestes gen. nov. the midfork basally recessed midway between arculus and nodus. The Lestidae and the enigmatic Eocene genus Promegalestes have an oblique vein “O”, unlike Menatlestes gen. nov. and Megalestes Sélys-Longchamp, 1862 (Münz 1919; Petrulevičius and Nel 2004b). Also, all Megalestidae and Lestidae have a short anterior side of the discoidal cell, unlike Menatlestes gen. nov.
The Eocene Latibasalidae share with Menatlestes gen. nov. the bases of RP3/4 and IR2 midway between arculus and nodus, but they have a MAb of inverted obliquity and a broad cubital area, unlike Menatlestes gen. nov. (Petrulevičius and Nel 2004a, 2007).
As the new fossil cannot fit in any damselfly family, we propose a new family, genus and species to accommodate its particular wing venation. Owing to the differences mentioned above, this new family likely belongs to the stem-group of the Lestinoidea.
Menatlestes palaeocenicus sp. nov.
Figs. 7–10.
ZooBank LSID: urn:lsid:zoobank.org:act:C7C9B602-ACF8-45D2-89 A5-FB1849B2B034
Etymology: Named after the Paleocene age of the type species. The specific epithet is to be treated as an adjective.
Holotype: MNT Nel Odo 2 (imprint and counterimprint of a nearly complete adult).
Type locality: New quarry, Menat, Puy-de-Dôme, France.
Type horizon: Middle Paleocene, Menat Basin.
Material.—Holotype only.
Diagnosis.—Wing venation characters. Areas between MP and CuA, MP and MAa, MAa and RP3/4 with one row of cells till to close of posterior wing margin; base of RP2 far distal to subnodus, that of IR1 close to that of RP2; IR1 very long; all main veins straight in their main parts.
Description.—Head deformed by the forward projection of the mouthparts, eyes 1.0 mm wide, 0.8 mm apart; head width across eyes 3.5 mm, length from anterior margin of antefrons to posterior margin of occiput ca. 2.0 mm; thorax fossilized in lateral position, meso-metathorax, 5.0 mm long; abdomen 42.0 mm long, with female genital appendages well visible, a well-developed ovipositor (resembling that of a Lestidae, thus probably adapted to endophytic oviposition); three legs partly preserved; wings apparently hyaline; fore- and hind wings nearly identical, 28.7 mm long, 8.1 mm wide; distance from base to arculus 2.8 mm, from arculus to nodus 4.0 mm, from nodus to pterostigma ca. 14.2 mm, from pterostigma to wing apex 2.9 mm; distance between Ax1 and Ax2 0.5 mm; Ax2 opposite arculus; three secondary antenodals distad Ax2; nodus of normal shape, without ScP prolonged distad nodus; nodal crossvein and subnodus aligned and of distinct normal obliquity; base of RP3/4 2.2 mm of arculus, nearly midway between arculus and nodus; base of IR2 at distal third between arculus and nodus, 2.9 mm from arculus; 20 postnodal crossveins, not aligned with postsubnodals; pterostigma elongate, 2.3 mm long, 0.6 mm wide, with basal and distal sides with a normal obliquity, covering six cells, an oblique pterostigmal brace; one row of cells between C and RA distad pterostigma; base of RP2 eight cells, 5.9 mm distad subnodus; no oblique vein “O”; base of IR1 three cells distad that of RP2; one row of cells between RP1 and IR1, three rows of cells between IR1 and RP2; all main veins nearly straight basally and curved near their apices; one row of cells between RP2 and IR2 in preserved part, but area widened distally, two between IR2 and RP3/4 with a longitudinal intercalary vein in-between; one row of cells between RP3/4 and MAa; one row of cells in preserved part of postdiscoidal area; one row of cells between MP and CuA; one row of cells between CuA and posterior wing margin, cubital area very narrow; CuP just distad base of AA; petiole 1.7 mm long, 0.8 mm wide; hind wing discoidal cell with basal side 0.3 mm long, anterior side elongate, 0.65 mm long, distal MAb side strongly oblique and elongate, 0.8 mm long, posterior side 1.2 mm long; subdiscoidal cell very long and narrow.

Fig. 7. Zygopteran odonatan Menatlestes palaeocenicus gen. et sp. nov., holotype MNT Nel Odo 2; from Paleocene, Menat, France. Habitus, imprint.

Fig. 8. Zygopteran odonatan Menatlestes palaeocenicus gen. et sp. nov., holotype MNT Nel Odo 2; from Paleocene, Menat, France. Habitus, counterimprint.

Fig. 9. Zygopteran odonatan Menatlestes palaeocenicus gen. et sp. nov., holotype MNT Nel Odo 2; from Paleocene, Menat, France. Head and thorax (A1), pterostigma (A2) . Scale bars 1 mm.
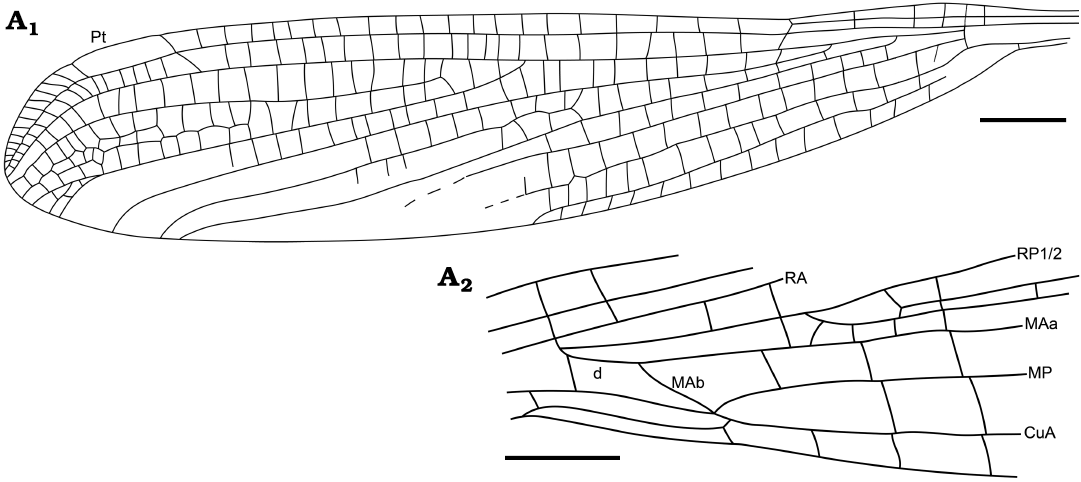
Fig. 10. Zygopteran odonatan Menatlestes palaeocenicus gen. et sp. nov., holotype MNT Nel Odo 2; from Paleocene, Menat, France. Reconstructions. Forewing (A1), hind wing base (A2). Scale bars 1 mm.
Remarks.—This new fossil differs from V. multicellulata in possessing only one row of cells between C and RA distad the pterostigma and the absence of crossveins ending into the basal side of the pterostigma (Garrouste et al. 2017). It also differs from Thanetophilosina menatensis in the positions of the base of RP3/4, basad midway between arculus and nodus (vs. well distal to this point) (Nel et al. 1997). It differs from Menatagrion hervetae gen. et sp. nov. owing to the presence of a pterostimal brace and only one row of cells between RA and RP1 and a very different shape of the discoidal cell. It differs from the poorly known “Lestes” zalesskyi Piton, 1940, in having, at least, longer wings, much more postnodal crossveins, a zigzagged IR1 with two rows of cells between it and RP1, a zigzagged CuA (Piton 1940: fig. 24). Nel and Paicheler (1994) indicated that the type of “Lestes” zalesskyi is lost and its affinities very uncertain.
Stratigraphic and geographic range.—Type locality and horizon only.
Order Cephalozygoptera Archibald, Cannings, Erickson, Bybee, and Mathewes, 2021
Remarks.—Archibald et al. (2021) grouped the two families Dysagrionidae + Sieblosiidae, and possibly Whetwhetaksidae (in which the head characters are unknown), in a new suborder Cephalozygoptera on the basis of a particular shape of the head. Nel and Zheng (2021) discussed this taxonomic treatment and considered that the particular shape of the head results from deformations due to the compression and overall a frontal projection of the head. Archibald and Cannings (2021) refuted the arguments of Nel and Zheng (2021), on the basis of the following arguments: the compound eyes of the specimen of Lestes ceresti chosen by Nel and Zheng (2021) would be absent, while after reexamination of the specimen it is not the case; they argued that if a distortion of the head occurs in Cephalozygoptera, the thorax has also to be deformed, and they considered the latter as normal. However, they did not manage to notice that also the thorax of the cephalozygopteran fossils are distorted and compressed. The major point to keep in mind is that the thorax deformation appears to be less pronounced due to its robustness and its sclerotization more important than in the head (partly due to the presence of the eyes creating huge soft areas).
Archibald and Cannings (2021) ignored the discussion in Nel and Zheng (2021) about Oligolestes stoeffelensis Nel, Poschmann, and Wedmann, 2020, belonging to the stem-group of the Sieblosiidae and with a non-deformed head, of zygopteran type (due to its fossilization in lateral position). If we take into account their arguments O. stoeffelensis should have a head similar to that of Cephalozygoptera, which is not the case. They also ignored the important argument about the fact that the “cephalozygopteran” type of head is also present in the Epiproctophora and several more inclusive taxa, rendering this head pattern (even if not corresponding to deformations) a plesiomorphy, not sufficient to support a new suborder.
Here we repeat the list of characters that would be diagnostic of the Cephalozygoptera, after Archibald et al. (2021): head width across eyes about twice the length from anterior margin of antefrons to posterior of occiput (Zygoptera: usually about three to five times as wide); compound eyes more or less adpressed to head, posterolateral corners protruding posteriorly to varying degrees, more or less rounded, sometimes acutely (Zygoptera: distinctly bulging laterally from head capsule); compound eyes separated by about one eye’s width dorsally (ratio of width between eyes/width eye has a range of 0.8–1.9 and a mean 1.0), n = 11 (Zygoptera: usually more than twice an eye’s width).
In the holotype of the lestomorphan Menatlestes palaeocenicus gen. et sp. nov., the head width across compound eyes is less than twice the length from anterior margin of antefrons to posterior of occiput, and the compound eyes are separated by less than one eye’s width dorsally (Fig. 9A1). These dimensions are clearly due to the deformation of the head, more important than that of the thorax that is simply compressed. Thus this taxon would fall in the Caphalozygoptera, while it clearly belongs to a different group.
The heads of several specimens of Lestes ceresti Nel and Papazian, 1985, from the Oligocene of Céreste (Lubéron, France), are also strongly deformed while the thorax are simply compressed and projected forward. In specimen PNRL 2019 (Fig. 11A), the head width across eyes is 2.7 the length from anterior margin of antefrons to posterior of occiput; the compound eyes are oval, with posterolateral corners protruding posteriorly, and the ratio of width between eyes/width eye = 1.5. In specimen PNRL 2020 (Fig. 11B), the head width across eyes is twice the length from anterior margin of antefrons to posterior of occiput; the compound eyes are oval, with posterolateral corners protruding posteriorly, and the ratio of width between eyes/width eye = 1.1. In specimen PNRL 2021 (Fig. 12A2, A3), the head width across eyes is 2.4 the length from anterior margin of antefrons to posterior of occiput; the compound eyes are oval, with posterolateral corners protruding posteriorly, and the ratio of width between eyes/width eye = 2.
Indeed, it is clear that the heads of these specimens of Lestes ceresti are deformed, with the mouthparts anteriorly projected, while they should be below the head. Their thorax are clearly less deformed, even their prothorax and meso-metathorax are also frontally projected. Therefore we keep considering that the characters used to define the Cephalozygoptera are insufficient. Until a proper justification (allowing to define at least one clear apomorphy) of the suborder status, the Cephalozygoptera are treated as a synonym of the Zygoptera.

Fig. 11. Zygopteran odonatan Lestes ceresti Nel and Papazian, 1985, from Oligocene, Céreste, France. A. PNRL 2019, imprint (A1) and counterimprint (A2) of head and thorax. B. PNRL 2020, imprint of head and thorax. Scale bars 2 mm.

Fig. 12. Zygopteran odonatan Lestes ceresti Nel and Papazian, 1985, PNRL 2021; from Oligocene, Céreste, France. Imprint (A1, A2), counterimprint (A3). Scale bars 2 mm.
Discussion
The extant species of Macrogomphus and Leptogomphus are distributed in the Indo-Malaysian region (Tibet, India, Indochina, Borneo, Java, and Sumatra). The presence of a species with major similarities with these two genera in the Paleocene of Menat is not so surprising, as the climate and paleoenvironment were warm and humid in Menat at that time (Wappler et al. 2009). The vertebrate fauna and the entomofauna comprise numerous taxa with tropical affinities, e.g., the mantodean Arvernineura insignis Piton, 1940, belonging to the Neotropical family Chaeteessidae the orthopteran Paleochina duvergeri Schubnel, Desutter-Grandcolas, Garrouste, Hervet, and Nel, 2020, belonging the tropical African and Asian family Chorotypidae, and a high diversity of large Blattodea, Hemiptera: Fulgoroidea, Coleoptera: Buprestidae and Cerambycidae (AN unpublished data; Piton 1940; Schubnel and Nel 2019; Schubnel et al. 2020).
Menatagrion hervetae gen. et sp. nov. is the first representative of the Dysagrionidae in the Paleocene. This family was previously known by one Cretaceous taxon Congqingia rhora Zhang, 1992, whose placement need to be reconsidered. Otherwise, all the other Dysagrionidae are Eocene to Miocene (Archibald et al. 2021), if we exclude the dubious placements of Thanetophilosina menatensis and Valerea multicellulata in this family.
The discovery of the new family Menatlestidae fam. nov. shows that the stem-group of the clade (Megalestidae + Lestidae) was present during the Paleocene and probably well before, during the Late Cretaceous, even if no representative is known for this period. The oldest known Lestidae are early Eocene (undescribed specimen from Tibet, Huang Diying personal communication, 2021), and middle Eocene (Greenwalt and Bechly 2014; Xia et al. 2021).
Conclusions
The present descriptions of three new species of Odonata increase our knowledge on this order for the paleolake of Menat, and more generally for the Paleocene. Their diversity and morphological disparity show that this period was probably crucial for the understanding of the diversification of the extant odonatan faunas. Better documenting and taking advantage of the set of fossils at our disposal will also allow to highlight the beginning of the radiation of many modern families that have their estimated divergence times at the end of the Cretaceous or during the Paleocene (e.g., Kohli et al. 2021; Suvorov et al. 2021). Additionally, the fact that all the known species from Menat are based on one (max. two) specimen(s) suggests that much more taxa should be present in the outcrop.
Acknowledgements
We sincerely thank Günter Bechly (Biologic Institute, Redmond, USA) and an anonymous referees for their useful remarks on the first version of the paper. We had the pleasure to receive generous assistance from Bernard Duverger (President of the Communauté de Communes of Menat region), Clotilde Berger-Pompili and Mathilde Leygnac (directors of the EHPAD du pays de Menat), for their kind help and authorizations to collect fossil insects in a small but rich outcrop near the village of Menat (Puy-de-Dôme, France).
References
Archibald, S.B., Cannings, R.A., Erickson, R.J., Bybee, S.M., and Mathewes, R.W. 2021. The Cephalozygoptera, a new, extinct suborder of Odonata with new taxa from the early Eocene Okanagan Highlands, western North America. Zootaxa 4934: 1–133. Crossref
Archibald, S.B. and Cannings, R.A. 2021. The head of Cephalozygoptera (Odonata). Zootaxa 5047: 97–100. Crossref
Bechly, G. 2016. Phylogenetic Systematics of Odonata. Available at https://bechly.lima-city.de/phylosys.htm
Bechly, G. and Rasmussen, J.A. 2019. A new genus of hawker dragonfly (Odonata: Anisoptera: Aeshnidae) from the Early Eocene Fur Formation of Denmark. Zootaxa 4550 (1): 123–128. Crossref
Calvert, P.P. 1893. The odonate genus Ortholestes. Proceedings of the Academy of Natural Sciences of Philadelphia 45: 377–382.
Calvert, P.P. 1901 Odonata. In: A.E. Eaton (eds.), Biologia Centrali-Americana. Zoology, Botany and Archaeology. Insecta. Neuroptera, 17–410. Published by R.H. Porter, London.
Carle, F.L. 1986. The classification, phylogeny and biogeography of the Gomphidae (Anisoptera). I. Classification. Odonatologica 15: 275–326.
Dow, R.A., Stokvis, F., and Ngiam, R.W.J. 2017. Revision of the genus Leptogomphus Selys in Borneo, including gene trees and a two marker molecular phylogeny (Odonata: Anisoptera: Gomphidae). Zootaxa 4358: 201–257. Crossref
Erickson, B.R. 1999. Fossil lake Wannagan (Paleocene: Tiffanian) Billings County, North Dakota. North Dakota Geological Survey 87: i–iv + 1–9.
Fraser, F.C. 1934. Odonata. In: J. Stephenson (ed.), The Fauna of British India Including Ceylon and Burma. Vol. 2, 398 pp. Taylor and Francis, London.
Fraser, F.C. 1936. A note on the identification of some obscure genera and species of the family Gomphidae. Proceedings of the Royal Entomological Society B 5: 137–143. Crossref
Fraser, F.C. 1943. Notes on the genera Agriogomphus Selys and Ischnogomphus Williamson with the description of the male of Agriogomphus sylvicola Selys (Odonata). Proceedings of the Royal Entomological Society B 12: 161–166.
Gambles, R.M. 1968. A new species of Lestinogomphus (Odonata) and the hitherto undescribed male of Microgomphus camerunensis. The Entomologist 101: 281–288.
Garrouste, R., Wedmann, S., Pouillon, J.-M., and Nel, A. 2017. The oldest “amphipterygid” damselfly of tropical affinities in the Paleocene of Menat (Zygoptera: Eucaloptera). Historical Biology 29: 818–821. Crossref
Greenwalt, D.E. and Bechly, G. 2014. A re-description of the fossil damselfly Eolestes syntheticus Cockerell, 1940 (Odonata: Zygoptera: Eolestidae n. fam.) with description of new taxa from the Eocene of North America. Zootaxa 3887: 138–156. Crossref
Hartung, V., Garrouste, R., Pouillon, J.-M., and Nel, A. 2016. First fossil of Cylindrostethinae (Heteroptera: Gerromorpha: Gerridae) in the Paleocene of Menat, France. Palaeontologia Electronica 19.3.33A: 1–10. Crossref
Jacquelin, L., Desutter-Grandcolas, L., Chintauan-Marquier, I., Boistel, R., Zheng, D.R., Prokop, J., and Nel, A. 2018. New insights on basivenal sclerites using 3D tools and homology of wing veins in Odonatoptera (Insecta). Scientific Reports 8: 1–7. Crossref
Jarzembowski, E.A. 1990. Early Cretaceous zygopteroids of Southern-England with the description of Cretacoenagrion alleni, gen. nov., spec. nov. (Zygoptera, Coenagrionidae, “Anisozygoptera”: Tarsophlebiidae, Euthemistidae). Odonatologica 19: 27–37.
Jarzembowski, E.A., Martinez-Declos, X.,
Bechly, G., Nel, A., Coram, R., and Escullié, F. 1998. The Mesozoic
non-calopterygoid Zygoptera: description of new genera and species
from the Lower Cretaceous of England and Brazil and their phylogenetic
significance (Odonata, Zygoptera, Coenagrionoidea, Hemiphleboidea,
Lestoidea). Cretaceous Research
19: 403–444. Crossref
Kedves, M. and Russell, D.E. 1982. Palynology of the Thanetian layers of Menat. The geology of the Menat Basin, France. Palaeontographica B 182 (4–6): 87–150.
Kennedy, C.H. 1936. Epigomphus crepidus a new dragonfly (Odonata: Gomphidae) from Nayarit, Mexico with notes on the genus. Annals of the Entomological Society of America 29: 126–137. Crossref
Kennedy, C.H. 1941. Perilestinae in Ecuador and Peru: revisional notes and descriptions (Lestidae: Odonata). Annals of the Entomological Society of America 34: 658–688. Crossref
Kennedy, C.H. 1947. Epigomphus subquadrices, a new dragonfly (Gomphidae) from Panama with notes on A. quadrices and Eugomphus n. subgenus. Annals of the Entomological Society of America 39: 662–666. Crossref
Kohli, M., Letsch, H., Greve, C., Béthoux, O., Deregnaucourt, I., Liu, S., Zhou, X., Donath, A., Mayer, C., Podsiadlowski, L., Gunkel, S., Machida, R., Niehuis, O., Rust, J., Wappler, T., Yu, X., Misof, B., and Ware, J. 2021. Evolutionary history and divergence times of Odonata (dragonflies and damselflies) revealed through transcriptomics. IScience 103324. [published online, https://doi.org/10.1016/j.isci.2021.103324] Crossref
Kosterin, O.E. 2019. New synonyms and a new subspecies of Macrogomphus Selys, 1858 (Odonata: Gomphidae) from continental south-east Asia. Zootaxa 4615: 57–90. Crossref
Laurent, L. 1912. Flore fossile des schistes de Menat (Puy-de-Dôme). Annales du Musée d’Histoire Naturelle de Marseille 14: 1–240.
Leach, W.E. 1815. Entomology. In: D. Brewster (ed.), The Edinburgh Encyclopaedia, Vol. 9, 57–172. William Blackwood, Edinburgh.
Legrand, J. 1993. Sur l’identité des genres Microgomphus Sélys et Africogomphus Fraser (Odonata, Anisoptera, Gomphidae). Revue Française d’Entomologie (N.S.) 15: 107–108.
Legrand, J. and Lachaise, M. 2001. Un nouveau Gomphidae afrotropical, Lestinogomphus matilei n. sp. (Odonata, Anisoptera). Revue Française d’Entomologie (N.S.) 23: 1–8.
Lewis, S.E. 1988. A dragon-fly (Odonata: Aeschnidae) from the Paleocene of North Dakota. Occasional Papers in Paleobiology of the St Cloud State University 2: 1–7.
Madsen, H.O.R. and Nel, A. 1997. Two new fossil species of Gomphaeschna Selys, 1871 in the Paleocene/Eocene of Denmark. Annales de la Société Entomologique de France (N.S.) 33: 285–293.
Martin, R. 1911. Notes sur les Gomphines d’Afrique. Annales de la Société Entomologique de France 80: 480–486.
May, E. 1933. Zwei neue Genera der Legion Podagrion (Agriomorpha und Lestomima). Senckenbergiana 15: 340–345.
Münz, P.A.
1919. A venational study of the suborder Zygoptera (Odonata) with keys
for the identification of genera. Memoirs
of the Entomological Society 3: 1–78. Crossref
Nel, A. 1989. Les Gyrinidae fossiles de France (Coleoptera). Annales de la Société Entomologique de France (N.S.) 25: 321–330.
Nel, A. 2007. Les Bibionidae du Paléocène de Menat (Puy-de-Dôme, France) (Diptera). Bulletin de la Société Entomologique de France 112: 369–370. Crossref
Nel, A. 2021. Maastrichtian
representatives of the dragonfly family Aeschnidiidae question the
entomofaunal turnover of the early Late Cretaceous. Palaeoentomology 4 (3):
209–212. Crossref
Nel, A. and Auvray, F. 2006. The oldest Vespinae from the Paleocene of Menat (France) (Hymenoptera: Vespidae). Zootaxa 1344: 59–62. Crossref
Nel, A. and Escuillié, F. 1993. Découverte d’un nouveau genre de Sieblosiidae dans le Miocène supérieur de l’Ardèche (France) (Odonata, Zygoptera, Lestoidea, Sieblosiidae). Nouvelle Revue d’Entomologie (N.S.) 10 (3): 233–242.
Nel, A. and Fleck, G. 2014. Dragonflies and damselflies (Insecta: Odonata) from the Late Eocene of the Isle of Wight. Earth and Environmental Science, Transactions of the Royal Society of Edinburgh 104: 283–306. Crossref
Nel, A. and Paicheler, J.-C. 1994. Les Lestoidea (Odonata, Zygoptera) fossiles: un inventaire critique. Annales de Paléontologie 80: 1–59.
Nel, A. and Roy, R. 1996. Revision of the fossil “mantid” and “ephemerid” species described by Piton from the Palaeocene of Menat (France) (Mantodea: Chaeteessidae, Mantidae; Ensifera: Tettigonioidea). European Journal of Entomology 93: 223–234.
Nel, A. and Zheng, D.-R. 2021. The recently proposed odonatan “suborder” Cephalozygoptera: fact or fiction. Palaeoentomology 4: 165–170. Crossref
Nel, A., Azar, D., and Hervet, S. 2010a. A new rhopalosomatid wasp in the Paleocene of France (Hymenoptera). Annales de la Société Entomologique de France (N.S.) 46: 211–215. Crossref
Nel, A., DePalma, R.A., and Engel, M.S. 2010b. A possible hemiphlebiid damselfly in Late Cretaceous amber from South Dakota (Odonata: Zygoptera). Transactions of the Kansas Academy of Science 113: 231–234. Crossref
Nel, A., Martínez-Delclòs, X., Escuillié, F., and Brisac, P. 1994. Les Aeshnidae fossiles: Etat actuel des connaissances (Odonata, Anisoptera). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 194: 143–186. Crossref
Nel, A., Martínez-Delclòs, X., Paicheler, J.-C., and Henrotay, M. 1993. Les “Anisozygoptera” fossiles. Phylogénie et classification (Odonata). Martinia Numéro Hors Série 3: 1–311.
Nel. A., Martínez-Delclòs, X., Papier, F., and Oudard, J. 1997. New Tertiary fossil Odonata from France. (Sieblosiidae, Lestidae, Coenagrioniidae, Megapodagrionidae, Libellulidae). Deutsche Entomologische Zeitschrift 44: 231–258. Crossref
Petrulevičius, J.F. 2020. First argiolestid damselfly (Odonata: Zygoptera) from the late Palaeocene of Northwest Argentina. Palaeoentomology 3: 541–545. Crossref
Petrulevičius, J.F. and Nel, A. 2002a. A new libelluloid dragonfly from late Paleocene deposits in Argentina (Odonata: Italoansida). European Journal of Entomology 99: 485–489. Crossref
Petrulevičius, J.F. and Nel, A. 2002b. New palaeomacromiid dragonflies from the Upper Paleocene of Argentina. Palaeontology 45: 751–758. Crossref
Petrulevičius, J.F. and Nel, A. 2003a. A new libelluloid dragonfly (Insecta: Odonata: Italoansida) from the late Paleocene of Argentina. Geobios 36: 401–406. Crossref
Petrulevičius, J.F. and Nel, A. 2003b. Frenguelliidae, a new family of dragonflies from the earliest Eocene of Argentina (Insecta: Odonata): phylogenetic relationships within Odonata. Journal of Natural History 37: 2909–2917. Crossref
Petrulevičius, J.F. and Nel, A. 2004a. A new damselfly family from the late Paleocene of Argentina (Insecta: Odonata: Zygoptera). Palaeontology 47: 109–116. Crossref
Petrulevičius, J.F. and Nel, A. 2004b. Recognition of the first fossil lestoid damselfly (Insecta: Zygoptera) in South America: biogeographic and phylogenetic remarks. Journal of Paleontology 78: 798–801. Crossref
Petrulevičius, J.F. and Nel, A. 2005. Austroperilestidae, a new family of damselflies from the earliest Eocene of Argentina (Insecta: Odonata). Phylogenetic relationships within Odonata. Journal of Paleontology 79: 658–662. Crossref
Petrulevičius, J.F. and Nel, A. 2007. Enigmatic and little known Odonata (Insecta) from the Paleogene of Patagonia and northwest Argentina. Annales de la Société Entomologique de France (N.S.) 43: 341–347. Crossref
Petrulevičius, J.F. and Nel, A. 2009. First Cordulephyidae dragonfly in America: a new genus and species from the Paleogene of Argentina. CR Palevol 8: 385–388. Crossref
Piton, L. 1940. Paléontologie du gisement éocène de Menat (Puy-de-Dôme), flore et faune. Mémoire de la Société d’Histoire Naturelle d’Auvergne 1: 1–303.
Riek, E.F. and Kukalová-Peck, J. 1984. A new interpretation of dragonfly wing venation based upon Early Carboniferous fossils from Argentina (Insecta: Odonatoidea) and basic characters states in pterygote wings. Canadian Journal of Zoology 62: 1150–1166. Crossref
Ris, F. 1917. Eine neue Agrioniden-Gattung der “Légion Podagrion” (Odonata) aus China. Tijdschrift voor Entomologie 60: 185–191.
Sélys-Longchamps, E. de 1862. Synopsis des Agrionines. Troisième légion: Podagrion. Bulletin de l’Académie Royale des Sciences, des Lettres et des Beaux-Arts de Belgique (Série 2) 14: 5–44.
Sélys-Longchamps, E. de 1869. Les névroptères de Port Denison. Annales de la Société Entomologique de Belgique (Comptes-Rendus) 11: lxxvi–lxxvii. Crossref
Sélys-Longchamps, E. de and Hagen, H. 1854. Synopsis des Gomphines. Bulletin de l’Académie Royale des sciences et belles-lettres de Bruxelles 21 (2): 23–114.
Schmidt, E. 1942. Bemerkungen über Lestiden. 2. Eine neue Gattung und Art aus Kamerun (Eolestes diotima). Mitteilungen der Deutsche Entomologische Gesellschaft 11: 102–111.
Schubnel, T. and Nel, A. 2019. New Paleogene mantises from the Oise amber and their evolutionary importance. Acta Palaeontologica Polonica 64: 779–786. Crossref
Schubnel, T., Desutter-Grandcolas, L., Garrouste, R., Hervet, S., and Nel, A. 2020. Paleocene of Menat Formation, France, reveals an extraordinary diversity of orthopterans and the last known survivor of a Mesozoic Elcanidae. Acta Palaeontologica Polonica 65: 371–385. Crossref
Suvorov, A., Scornavacca, C., Fujimoto, M.S., Bodily, P., Clement, M., Crandall, K.A., Whiting, M.F., Schrider, D.R., and Bybee, S.M., 2021. Deep ancestral introgression shapes evolutionary history of dragonflies and damselflies. Systematic Biology [published online, https://doi.org/10.1093/sysbio/syab063] Crossref
Wappler, T., Currano, E.D., Wilf, P., Rust, J., and Labandeira, C.C. 2009. No post-Cretaceous ecosystem depression in European forests? Rich insect-feeding damage on diverse middle Palaeocene plants, Menat, France. Proceedings of the Royal Society B 276: 4271–4277. Crossref
Ware, J.L., Pilgrim, E., May, M.L., Donnely, T.W., and Tennessen, K. 2017. Phylogenetic relationships of North American Gomphidae and their close relatives. Systematic Entomology 42: 347–358. Crossref
Wedmann, S., Pouillon, J.-M.,
and Nel, A. 2014. New Palaeogene horntail wasps (Hymenoptera,
Siricidae) and a discussion of their fossil record. Zootaxa
3869 (1): 33–43. Crossref
Wedmann, S., Uhl, D., Lehman, T., Garrouste, R., Nel, A., Gomez, B., Smith, K., and Schaal, S.F.K. 2018. The Konservat-Lagerstätte Menat (Paleocene; France)—an overview and new insights. Geologica Acta 16: 1–31.
Wighton, D.C. and Wilson, M.V.H. 1986. The Gomphaeschninae (Odonata: Aeshnidae): new fossil genus, reconstructed phylogeny, and geographical history. Systematic Entomology 11: 505–522. Crossref
Williamson, E.B. 1916. A new Cyanogomphus (Odonata). Entomological News 27: 167–172.
Williamson, E.B. 1919. Results of the University of Michigan-Williamson Expedition to Colombia, 1916–1917, III: Archaeogomphus, a new genus of dragonflies (Odonata). Occasional Papers of the Museum of Zoology, University of Michigan 63: 1–8.
Xia, G., Zheng, D., Krieg-Jacquier, R., Fan, Q., Chen, Y., and Nel, A. 2021. The oldest-known Lestidae (Odonata) from the late Eocene of Tibet: palaeoclimatic implications. Geological Magazine 159 (4): 511–518. Crossref
Zhang, J.-F. 1992. Congqingia rhora gen. nov., spec. nov.—a new dragonfly from the Upper Jurassic of Eastern China (Anisozygoptera, Congquingiidae fam. nov.). Odonatologica 21: 375–383.
Zheng, D.-R. 2020. Odonatans in lowermost Cenomanian Kachin amber: updated review and a new hemiphlebiid damselfly. Cretaceous Research 118 (104640): 1–7. Crossref
Zheng, D.-R., Jarzembowski, E.A., Zhuo, D., and Nel, A. 2021. Protohemiphlebiidae fam. nov., a stem hemiphlebioid damselfly from Cretaceous amber in Kachin. Geological Society of London Special Publication 521 [published online, https://doi.org/10.1144/SP521-2020-249] Crossref
Acta Palaeontol. Pol. 67 (3): 631–648, 2022
https://doi.org/10.4202/app.00960.2021