
Mollusks from Miocene hydrocarbon-seep deposits in the Ilocos-Central Luzon Basin, Luzon Island, Philippines
STEFFEN KIEL, ALLAN GIL S. FERNANDO, CLARENCE Y. MAGTOTO, and TOMOKI KASE
Kiel, S., Fernando, A.G.S., Magtoto, C.Y., and Kase, T. 2022. Mollusks from Miocene hydrocarbon-seep deposits in the Ilocos-Central Luzon Basin, Luzon Island, Philippines. Acta Palaeontologica Polonica 67 (4): 917–947.
We report 35 molluscan species from Late Miocene cold-seep carbonates from the Amlang Formation in the Ilocos-Central Luzon Basin in Luzon Island, Philippines, collected in a large quarry in the province of Pangasinan. The 19 bivalve species are largely representatives of chemosymbiotic families; the six new species are the nuculid Acila (Truncacila) interferencia sp. nov., the mytilid Bathymodiolus labayugensis sp. nov., the thyasirid Conchocele pangasinanensis sp. nov., the lucinid Megaxinus gorrospei sp. nov., the vesicomyid Pliocardia ballesterosi sp. nov., and Sisonia frijellanae gen. et sp. nov., of uncertain taxonomic affinity. The 16 gastropods include one species restricted to seep deposits, the neritid species Thalassonerita hagai sp. nov.; the buccinid Enigmaticolus semisulcata represents the first fossil record of its genus. Biogeographically, the Pangasinan seep fauna shows several links to Neogene seep faunas in other tropical/subtropical areas, namely the Mediterranean and Caribbean regions. In contrast, shared taxa with nearby but extratropical Japan are few, as are shared taxa with Miocene seep deposits in New Zealand.
Key words: Gastropoda, Bivalvia, chemosynthesis-based ecosystem, deep sea, hydrocarbon seep, Miocene, Philippines.
Steffen Kiel [steffen.kiel@nrm.se], Swedish Museum of Natural History, Department of Palaeobiology, Box 50007, 10405 Stockholm, Sweden.
Allan Gil S. Fernando [asfernando@up.edu.ph] and Clarence Y. Magtoto [renz_y_magtoto@yahoo.com], National Institute of Geological Sciences, University of the Philippines Diliman, Quezon City 1101, Philippines.
Tomoki Kase [neritopsis@gmail.com], National Museum of Nature and Science, Department of Geology and Paleontology, Tsukuba, Ibaraki 305-0005, Japan.
Received 25 January 2022, accepted 12 May 2022, available online 09 September 2022.
Copyright © 2022 S. Kiel et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Methane seeps in the deep sea are inhabited by a distinct fauna dominated by chemosymbiotic animals (Paull et al. 1984; Fisher 1995). The high degree of endemism in these habitats, many families of annelids, crustaceans, and mollusks are known exclusively from methane seeps, hydrothermal vents, and sulfide-rich habitats around sunken whale carcasses and wood, and their specialized physiology spurred the idea that these faunas have a long and distinct evolutionary history (Tunnicliffe 1991; Distel et al. 2000; Vrijenhoek 2013). The fossil record provides the primary evidence for the history of life and indeed has provided unique insights into the questions of antiquity and evolutionary origin of these faunas (Campbell and Bottjer 1995; Little et al. 1997; Kiel and Little 2006; Kiel 2015). Especially the Cenozoic record of methane-seep communities is increasingly well documented (Majima et al. 2005; Campbell 2006; Kiel 2010b; Saether et al. 2010a; Kiel and Hansen 2015; Amano et al. 2019b; Kiel et al. 2020b; Hryniewicz et al. 2021; and references therein); however, the most species-rich area of the world’s oceans, the central Indo-Pacific Ocean including the Malayan and Philippine archipelagos, is only beginning to be explored for such faunas (Beets 1953; Majima et al. 2007, 2010; Kase et al. 2019; Kiel et al. 2020a).
The purpose of this study is to document the mollusk fauna of newly discovered Late Miocene methane-seep deposits in the Ilocos-Central Luzon Basin in Luzon Island, Philippines. The Philippine island arc system is an assemblage of allochthonous arcs, ophiolite complexes, continental and oceanic rocks, and lies at the boundary of the Eurasian and the Philippine Sea plates (Karig 1983; David et al. 1997; Hall 2002; Galgana et al. 2007; Yumul et al. 2008; Aurelio et al. 2013). The north-south oriented Ilocos-Central Luzon Basin is one of 16 recognized major sedimentary basins in the Philippines, and stretches from the vicinity of the Manila metropolitan area in the south to Ilocos Norte in the north (Pinet and Stephan 1990a; Peña 2008; MGB 2010). The about 8000 m thick sedimentary sequence filling the basin reflects a wide diversity of depositional environments; source of the clastic sediments are the Luzon Central Cordillera on the eastern side and the Zambales Mountains on the western side of the basin (Saldivar-Sali 1978; Pinet and Stephan 1990a, b).
Both shallow and deep marine deposits of the Ilocos-Central Luzon Basin are exposed in Barangay Labayug in the Municipality of Sison, Pangasinan province, in a large quarry at 16°10’N, 120°34’E (Fig. 1), owned by the Northern Cement Corporation. These deposits include a bedded limestone unit with common reefal fossils, named Labayug Limestone, and alternating sequences of limestone and mudstone beds mapped as part of the Amlang Formation (Lorentz 1984). The seep deposits reported here are from the Amlang Formation, found as floats in the lower part of the quarry, locally known as the “Shale Quarry” (Magtoto and Fernando 2017).
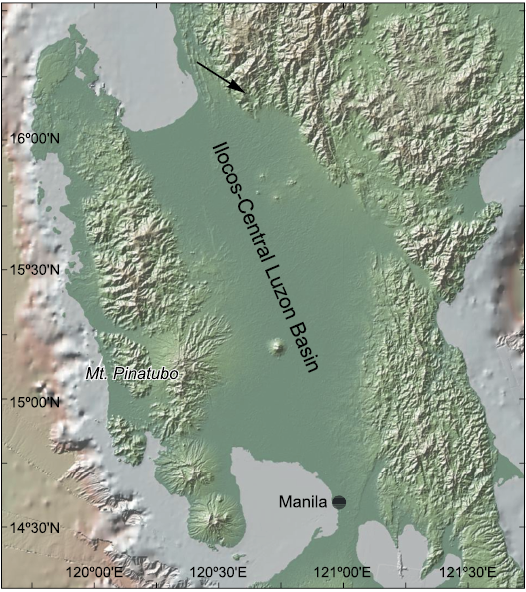
Fig. 1. Map showing the location of the Northern Cement Corporation (NCC) quarry in the province of Pangasinan, Luzon, Philippines (arrow). Modified from www.geomapapp.org (CC-BY) and Ryan et al. (2009).
Institutional abbreviations.—NIGSPAL (NIGS-UPGAA Museum) National Institute of Geological Sciences, University of the Philippines, Diliman, Quezon City, Philippines; NRM, Swedish Museum of Natural History, Stockholm, Sweden.
Other abbreviations.—aams, anterior adductor muscle scar; aprs, anterior pedal retractor scar; H, height; L, length; LV, left valve; pams, posterior adductor muscle scar; RV, right valve; W, width. The width of bivalves always refers to specimens with two conjoined valves, measurements on single valves are marked by “(single)”; the plus sign after a measurement indicates that the specimen was incomplete.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in ZooBank: urn:lsid:zoobank.org:pub:0A7EA08E-1FB4-48D1-833A-A11B3DEE6DC9
Material and methods
The specimens were collected during two field seasons in 2015 and 2019. Four blocks of seep carbonate were sampled in 2015, referred to as blocks A–D; in 2019 we sampled block B again, and we sampled six additional blocks, referred to as blocks E–J. Further specimens were given to us by the staff of the Northern Cement Corporation; they are referred to as coming from the “NCC collection”. Except possibly for block F, all blocks from which we report mollusks were found as floats. The blocks had most likely been left aside or moved away while quarrying the shale. Planktic foraminifera of biostratigraphic relevance extracted from Block B include Trilobatus sacculifer, Trilobatus trilobus, Dentoglobigerina altispira altispira, and Globorotalia plesiotumida, among others (Hiroki Hayashi, Shimane University, Japan, personal communication 2020). Based on the occurrence of G. plesiotumida, which has its first occurrence at the base of zone N17 (cf., Blow 1969), this block is most likely of Late Miocene age. This is consistent with the nannofossil age dating of the Amlang Formation (Lorentz 1984; Magtoto and Fernando 2017). In the lab, specimens were mechanically extracted from the carbonate matrix using pneumatic chisels and needles. All specimens were coated with ammonium chloride prior to photography. Most holotypes and the specimens of the NCC collection are housed at NIGS, the remaining material is housed in the Department of Palaeobiology at NRM.
Discussion
Faunal remarks.—Amano and Ando (2011) noted that the fossil record of very large solemyids in Japan is confined to deep-marine deposits of Early and Middle Miocene age, with specimens of Acharax yokosukensis and A. gigas reaching almost 300 mm in length. To this we can add the large shells of Acharax aff. bartschii reaching 245 mm in length from the Late Miocene-aged deep-marine cold-seep deposits of Luzon, Philippines. But why solemyids reached so much larger sizes in the tropical northwestern Pacific Ocean in (or since) the Miocene, while typically not exceeding 100 mm in length elsewhere, both in the Cenozoic fossil record (Goedert and Squires 1990; Gill et al. 2005; Kiel 2010b; Taviani et al. 2011; Kiel and Hansen 2015; Pérez-Barría and Nielsen 2020; Kiel et al. 2020b; Hryniewicz et al. 2020) and in the oceans today (Abbott 1974; Coan et al. 2000; Taylor et al. 2008; Kamenev 2009) remains unclear.
The nuculid subgenus Truncacila is common at seeps since the Cretaceous (Amano 2003; Amano and Jenkins 2007; Jenkins et al. 2007b; Kiel et al. 2008, 2020b; Amano et al. 2010, Nesbitt et al. 2013; Miyajima et al. 2018) and is most likely a vagrant taking advantage of the abundant organic matter at seeps (Kiel 2010b). Among the bathymodiolins, the genus Gigantidas appears to have been established in the western Pacific Ocean by Middle to Late Miocene time (Saether et al. 2010b). The large thyasirid genus Conchocele is known from seep deposits since the Late Cretaceous, though mostly from the Pacific Ocean (Hryniewicz et al. 2017). The Miocene records from the Philippines documented here (Conchocele pangasinanensis and Conchocele aff. quadrata) add to the tropical record of the genus (Okutani 2002; Kiel and Hansen 2015; Samadi et al. 2015; Kiel et al. 2020a).
The lucinid Megaxinus gorrospei is the first record of this genus from the central Indo-Pacific Ocean and also its easternmost occurrence. Interestingly, no species of Megaxinus has been found in the recently described Pliocene and Pleistocene seep deposits in the Philippines, whereas it dominates a Pliocene seep deposit in Italy (Kiel and Taviani 2018). The large Meganodontia reached a circum-tropical distribution during the Miocene, reaching from the Tethyan/Mediterranean Sea via Taiwan and the Philippines to the Caribbean Sea (Kiel and Hansen 2015; Kiel and Taviani 2017), with Meganodontia haunuiensis from the Early Miocene of New Zealand being the only extra-tropical record (Amano et al. 2018). Also the two species of Lucinoma present in the Pangasinan seep deposits show links to extratropical areas: L. acutilineata is a widespread North Pacific species (Hirayama 1954; Moore 1963; Majima et al. 2005; Kiel 2010a) and L. saetheri occurs in Early to Middle Miocene-aged seep deposits in New Zealand (Amano et al. 2018).
Among the vesicomyids, Pliocardia ballesterosi is similar to the Japanese P. crenulomarginata and P. kuroshimana, and hence to the actual genus Pliocardia rather than the “cordata group” (cf., Johnson et al. 2017). Although not perfectly preserved, the Pangasinan seep deposits produced the as-yet earliest record of Wareniconcha, a possible record of Archivesica, and two small-sized vesicomyid species of uncertain affinity. The enigmatic Sisonia frijellanae remains difficult to place among extant and fossil bivalve families, having the shell shape, but definitely not the size, of some Galeommatoidea, and also falls outside the known shell shapes and hinge characteristics of extant and fossil Vesicomyidae. It might have a congener in the Miocene of New Zealand and certainly in the Pleistocene of Taiwan.
Gastropods are neither well preserved nor abundant in the Pangasinan seep deposits. Noteworthy is Thalassonerita hagai, representing the first record of this genus from the central Indo-West Pacific Ocean and thus indicating a former widespread occurrence of this genus (Moroni 1966; Taviani 1994; Squires and Goedert 1996; Gill et al. 2005; Hybertsen and Kiel 2018), which is today restricted to a single species, Thalassonerita naticoides, in the Gulf of Mexico and Caribbean Sea (Clarke 1989; Olu et al. 1996; Warén and Bouchet 2001). Another biogeographic link to the Caribbean Sea is a species of the seguenziid Cataegis, a genus which has persisted in both the Caribbean Sea and the central Indo-West Pacific at least since the Miocene (McLean and Quinn 1987; Gill et al. 2005; Kiel and Hansen 2015; Vilvens 2016; Kiel et al. 2020a). The buccinid Enigmaticolus semisulcata comb. nov. is the first fossil record of this genus, whose extant members live in deep waters of the Indian and West Pacific Oceans. The cephalaspidean Volvulella aff. minoensis, is one of the few gastropod species showing affinities to the Japanese Miocene deep-water fauna.
Biogeographic remarks.—Although composed of clades typically found at Miocene methane seeps, such as solemyids, bathymodiolins, thyasirids, lucinids, and vesicomyids, the fauna of the Pangasinan seep fauna has a quite distinctive biogeographic character. It shows close links to seep faunas in tropical/subtropical regions, such as Meganodontia and Thalassonerita, which occur in the Miocene of both the Mediterranean and the Caribbean regions (Taviani 1994; Kiel and Hansen 2015; Kiel and Taviani 2017). Another link to the Caribbean is the gastropod Cataegis (Gill et al. 2005; Kiel and Hansen 2015), whereas Megaxinus is common in a Pliocene seep deposit in Italy (Kiel and Taviani 2018).
In contrast, the Pangasinan seep fauna shows remarkably few direct links to the well-known Miocene seep faunas of nearby Japan, with which they share only the widespread species Lucinoma acutilineata (Majima et al. 2005) and possibly Conchocele aff. quadrata and Volvulella aff. minoensis. This is a remarkable difference to the Pliocene to Recent situation, where numerous links between the Philippine and the Japanese seep faunas exist (Kiel et al. 2020a). Other links to extratropical areas are Gigantidas coseli and Lucinoma saetheri, two species known from Miocene seep deposits in New Zealand (Saether et al. 2010b; Amano et al. 2018).
Conclusions
The mollusks reported here from the Pangasinan seep deposits in Luzon, Philippines, provide new insights into the taxonomic composition and biogeographic relationships of seep faunas from the Neogene central Indo-West Pacific Ocean. The 19 bivalve species are represented largely by chemosymbiotic taxa, many of which are known largely or exclusively from seep and vent environments. The 16 gastropods include only few taxa that are restricted to seep and vent environments; the others are vagrants from the background fauna that took advantage of the abundant organic matter around the seeps. The fauna shows biogeographic links to other tropical seep faunas, especially in Italy and the Caribbean regions. Only few shared species were found with the well-known Miocene seep fauna of Japan, in contrast to the Pliocene–Pleistocene situation, when such links were more common.
Systematic palaeontology
Class Bivalvia Linnaeus, 1758
Subclass Protobranchia Pelseneer, 1889
Family Solemyidae Gray, 1840
Genus Acharax Dall, 1908b
Type species: Solemya johnsoni Dall, 1891, by original designation; Recent, Pacific Ocean.
Acharax aff. bartschii (Dall, 1908a)
Fig. 2A–C.
Material.—One specimen from block I (NRM Mo 192667, L = 190.0 mm), two specimens from the NCC collection (NIGSPAL-003, L = 240.0 mm; NIGSPAL-033, L = 160.0 mm). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
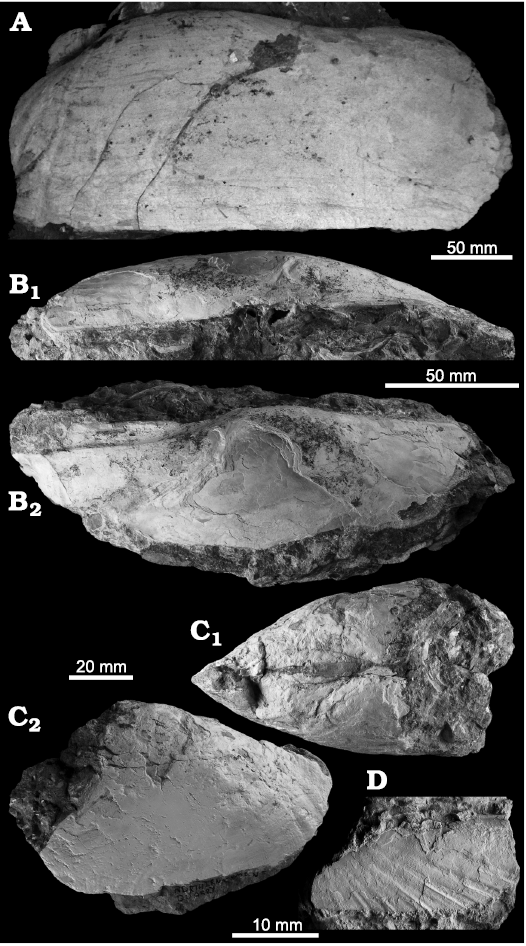
Fig. 2. Solemyid bivalves from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. A–C. Acharax aff. bartschii (Dall, 1908). A. NIGSPAL-003, large RV, view on outer side of shell. B. NRM Mo 192667, large RV in dorsal view (B1) and view on outer side of LV (B2). C. NIGSPAL-033, large articulated specimen in dorsal view (C1) and view on outer side of LV (C2); note that the specimen is gaping so in dorsal view it appears wider than it is. D. Acharax sp. with only the anterior portion preserved (NRM Mo 192660).
Remarks.—The extant Acharax bartschii is a large rare species known from central Philippine waters (Dall 1908a). It agrees well in overall shape with the Miocene specimens reported here, though the radial ribs on the posterior side are directed postero-ventrally in the extant specimens, whereas in the Miocene specimen they are much more ventrally oriented (see Fig. 2A). Hence, we assign the Miocene specimens only hesitantly to A. bartschii. The Early to Middle Miocene Acharax yokosukensis Kanie and Kuramochi, 1995, from central Honshu, Japan, has a more slender shell, and Acharax gigas (Kanno, 1960) from the Miocene Hiranita and Haratajino formations in central Honshu, Japan, has a much shorter posterior side of shell (Kanno 1960; Kanie and Kuramochi 1995, 2002; Amano and Ando 2011: pl. 5: 3).
Acharax sp.
Fig. 2D.
Material.—One specimen from block I (NRM Mo 192660, L = 26.0 mm), one specimen from block J (NRM Mo 192534, L = 20.0 mm). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Remarks.—The two small available fragments seem to have parallel dorsal and ventral shell margins, unlike A. aff. bartschii, and their radial ribs are broad and flat, unlike the corrugated ribs of A. aff. bartschii.
Family Nuculidae Gray, 1824
Genus Acila Adams and Adams, 1858
Type species: Nucula divaricata Hinds, 1843, by subsequent designation; Recent, temperate northwestern Pacific Ocean.
Subgenus Truncacila Grant and Gale, 1931
Type species: Nucula castrensis Hinds, 1843, by original designation; Pliocene to Recent, northeastern Pacific Ocean.
Acila (Truncacila) interferencia sp. nov.
Fig. 3.
Zoobank LCID: urn:lsid:zoobank.org:act:215BA2D8-5F8A-42D1-A54C-D2E66CE97C37
Etymology: For the sculpture on the anterior shell portion, where the typical divaricate pattern in Truncacila turns into some interference-like pattern.
Type material: Holotype: NIGSPAL-004, articulated specimen with preserved shell material, from block I. Paratype: NRM Mo 192694, articulated specimen with shell partially broken off, from block G from the type locality and horizon.
Type locality: The “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Type horizon: Seep carbonate blocks enclosed in the Amlang Formation (Upper Miocene).
Material.—Five specimens from block B (NRM Mo 192589–192593), one specimen from block G (paratype NRM Mo 192694), two specimens from block I (holotype NIGSPAL-004, NRM Mo 192659). All from the type locality and horizon.
Dimensions (in mm).—Holotype NIGSPAL-004: L = 23.7, H = 16.0, W = 11.7; NRM Mo 192659: L = 23.4, H = 15.0, W = 12.0; NRM Mo 192592: L = 21.4, H = 13.6; NRM Mo 192694: L = 27.0, H = 15.0, W = 13.0.
Diagnosis.—Average sized shell for the genus; dorsal and ventral margins evenly convex, subparallel; rostral sinus indistinct; escutcheon broad, wide, oval, lunule very narrow; line of primary divarication displaced toward anterordorsal shell margin, divaricate sculpture disintegrates in anteroventral third of shell.
Description.—Shell subquadrate to elliptical; height/length ratio ~0.65, width/height ratio ~0.8. Anterodorsal shell margin evenly convex, anterior margin acutely rounded, ventral margin convex, posterior margin concave; escutcheon broadly oval, with distinct groove and ridge subparallel to margin, ridge with nodular, commarginal ribs, inner part of escutcheon elevated toward ventral side. Line of primary divarication of external sculpture displaced toward anterordorsal shell margin; two secondary lines of bifurcation, one close and subparallel to anterordorsal margin, one posterior at about one third of shell length, divaricate sculpture becomes irregular on anterior third of shell. Anterior adductor muscle scar near anterior end of dorsal margin, nearly round, bounded posteriorly by strong ridge; posterior adductor scar at posterior end of escutcheon, elongate-oval, bounded anteriorly by distinct ridge; pallial line distinct, entire, parallel to ventral shell margin; inner ventral shell margin crenulated. Hinge with up to 20 anterior teeth and nine posterior teeth.

Fig. 3. The nuculid bivalve Acila (Truncacila) interferencia sp. nov. from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. A. NIGSPAL-004, holotype, an articulated specimen showing external sculpture; view on RV (A1), dorsal view on anterodorsal margin (A2) and on posterior margin (A3). B. NRM Mo 192694, paratype, specimen with most of the shell material removed, revealing internal features; view on LV (B1), dorsal view on anterodorsal margin (B2) and on posterior margin (B3). Abbreviations: aams, anterior adductor muscle scar; pams, posterior adductor muscle scar.
Remarks.—With its elongated shell with nearly parallel dorsal and ventral margins, a very indistinct rostral sinus, and rather unordered sculpture on the anterior half of the shell, this species is quite distinct from other species of Acila and Truncacila. Similar regarding the elongation is a specimen reported as “Acila divaricata (Hinds)” from the Miocene Kabeya Formation in Fukushima prefecture, Japan (Aoki 1954: pl. 1: 17), but that specimen widens anteriorly (rather than narrowing as in A. [T.] interferencia) and has a deeper rostral sinus than A. (T.) interferencia. The late Oligocene to Early Miocene Acila vigilia var. elongata Nagao and Huzioka, 1941, from Hokkaido, Japan, has a more distinct and broader escutcheon and a better defined rostrum than A. (T.) interferencia, and has finer sculpture without the interference-like pattern (Nagao and Huzioka 1941).
Also similar regarding elongation are several specimens reported as A. (T.) nehalemensis Hanna, 1924, from the Eocene Keasey Formation in Oregon (Schenck 1936: pl. 5: 6, 8), and as A. (T.) conradi (Meek, 1864) from the Early Miocene Astoria Formation in Oregon and Washington (Schenck 1936: pl. 8: 2, 3, 5–10, 12, 13), but the ribs on the anterior side of these shell radiate in an orderly way toward the shell margin, quite distinct from the granular pattern of A. (T.) interferencia. All other species reported by Schenck (1936) are shorter and/or more oval, or have a more distinct posterior projection, and more ordered divaricate sculpture.
Stratigraphic and geographic range.—Upper Miocene of Luzon, Philippines.
Family Mytilidae Rafinesque, 1815
Genus Gigantidas Cosel and Marshall, 2003
Type species: Gigantidas gladius Cosel and Marshall, 2003, Recent from the southern Kermadec Ridge in 360–750 m depth.
Gigantidas coseli Saether, Little, Campbell, Marshall, Collins, and Alfaro, 2010b
Fig. 4.
2010 Gigantidas coseli sp. nov; Saether et al. 2010b: 21, figs. 4B, C, 8–10, 17.
2016 Gigantidas coseli Saether et al. 2010; Saether et al. 2016: 7, fig. 4.
Material.—One specimen from block A (NIGSPAL-039), seven specimens from block E (NRM Mo 192454–192462), two specimens from block I (NRM Mo 192668, 192669). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Dimensions (in mm).—NRM Mo 192454, L = 113.0, H = 28.3, W = 27.3; NRM Mo 192455, L = 60.7, H = 16.2, W = 12.4; NRM Mo 192456, L = 43.0, H = 13.8, W (single) = 0.4; NRM Mo 192668, L = 26.0+, H = 12.8, W (single) = 7.6; NRM Mo 192669, L = 45.8, H = 18.4, W (single) = 7.5.

Fig. 4. The bathymodiolin bivalve Gigantidas coseli Saether, Little, Campbell, Marshall, Collins, and Alfaro, 2010, from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. A. NRM Mo 192455, articulated specimen, view on LV (A1) and dorsal view (A2). B. NRM Mo 192456, internal mold, view on LV showing posterior adductor muscle scar (pams). C. NRM Mo 192668, isolated LV, view on exterior (C1) and on dorsal side (C2). D. NRM Mo 192669, isolated LV, view on exterior. E. NRM Mo 192454, large, articulated, and slightly distorted specimen, view on RV.
Remarks.—The New Zealand specimens of Gigantidas coseli show some variation regarding the expansion of the posterior shell portion, ranging from being almost straight to bending and expanding ventrally (Saether et al. 2010b), which is also seen among the specimens from the Northern Cement Corporation quarry.
Stratigraphic and geographic range.—Middle Miocene of New Zealand (North Island) and Upper Miocene of Luzon, Philippines.
Genus Bathymodiolus Kenk and Wilson, 1985
Type species: Bathymodiolus thermophilus Kenk and Wilson, 1985, by original designation; Recent, Galapagos Ridge.
Bathymodiolus labayugensis sp. nov.
Fig. 5.
Zoobank LCID: urn:lsid:zoobank.org:act:5E3EB20E-A4A9-4E9F-BF8E-0295FF7A96BB
Etymology: For the Barangay Labayug, where the Northern Cement Corporation Quarry is located.
Type material: Holotype NIGSPAL-005, isolated LV with shell partially broken off. Paratypes: NRM Mo 192523–192532, mostly isolated valves embedded in rock matrix; all from block B in the type locality.
Type locality: The “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Type horizon: Seep carbonate blocks enclosed in the Amlang Formation (Upper Miocene).
Material.—65 specimens from block B (NIGSPAL-005, 006, 007, NRM Mo 192472–192533), one specimen from block D (NIGSPAL-008), one specimen from block F (NRM Mo 192541), six fragmentary specimens from block G, five specimens from block I (NRM Mo 192680–192684). All from the type locality and horizon.
Dimensions (in mm).—Holotype NIGSPAL-005: L = 57.5, H = 27.0, W (single) = 10.0; paratypes: NRM Mo 1922523: L = 41.6, H = 22.5; NRM Mo 1922524: 44.5, H = 24.2; NRM Mo 192525: 41.5+, H = 25.0; NRM Mo 192526: L = 42.5, H = 28.3; NRM Mo 192527: L = 42.5, H = 25.8; NRM Mo 192528: L = 33.0, H = 22.4; NRM Mo 192529: L = 37.1, H = 21.7; NRM Mo 192530: L = 45.4, H = 22.0; NRM Mo 192531: L = 30.3, H = 13.8, W = 9.5; NRM Mo 192532: L = 24.5, H = 13.0.
Diagnosis.—Shell small for genus, widening posteriorly; umbo in subterminal position; anterior side broad, evenly rounded, dorsal margin long, gently convex, ventral margin concave; maximum inflation at center of shell.
Description.—Elongate, arched modioliform shell of medium size (up to 57.5 mm), height/length ratio about 0.45, width/height ratio ~0.8. Surface smooth except for fine growth lines. Blunt ridge running from beak toward posteroventral area; area before ridge depressed, area dorsal to ridge broad and flat. Beak low, prosogyrate, in subterminal position. Anterior margin rounded; ventral margin concave; posterodorsal margin long, convex, continuing into subtruncated posterior margin. Internal groove running from umbones to dorsal side of large, D-shaped posterior adductor muscle scar.
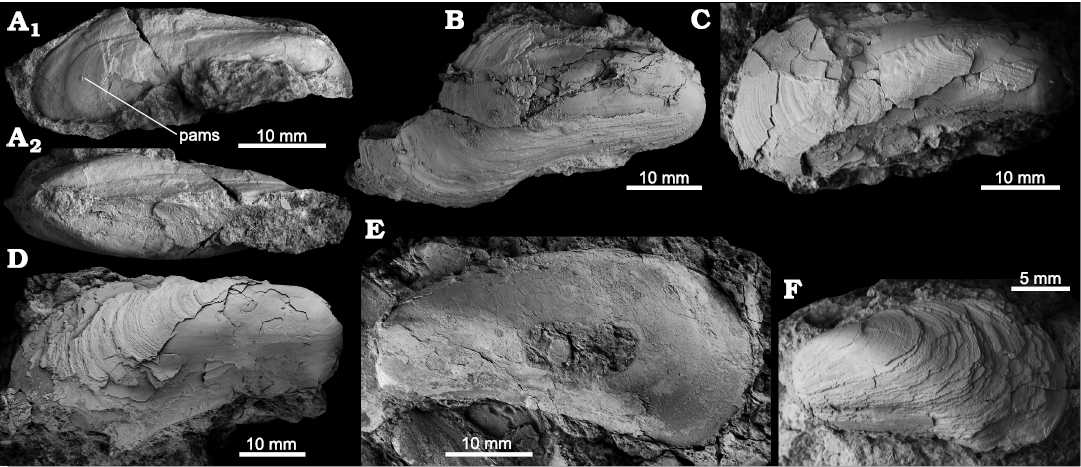
Fig. 5. The bathymodiolin bivalve Bathymodiolus labayugensis sp. nov. from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. A. NRM Mo 192681, internal mold showing posterior adductor muscle scar (A1, pams) and dorsal view showing inflation (A2). B. NRM Mo 192684, external view of RV. C. NRM Mo 192682, external view of RV. D. NRM Mo 192683, external view of RV. E. NIGSPAL-005, holotype, external view of RV. F. NRM Mo 192532, paratype, external view of RV.
Remarks.—Bathymodiolus labayugensis differs from the Miocene Bathymodiolus heretaunga Saether, Little, Campbell, Marshall, Collins, Alfaro, 2010b, from seep deposits in New Zealand by having the umbones in a more anterior position (Saether et al. 2010b). Two fossil Bathymodiolus species have been reported from the fossil record of Japan, the Oligocene Japanese “Bathymodiolus” inouei Amano and Jenkins, 2011 and the Miocene Bathymodiolus akanudaensis (Kuroda, 1931); both have a more angular and more anteriorly situated transition from dorsal to posterior margin compared to B. labayugensis (Kuroda 1931; Amano et al. 2010; Amano and Jenkins 2011). The Middle to Late Miocene “Bathymodiolus” moroniae from the “Calcari a Lucina” seep deposits in Italy (Kiel and Taviani 2017) differs from B. labayugensis by having a straight dorsal margin, in contrast to the curved one of B. labayugensis. The same applies to the Oligocene–Miocene Bathymodiolus palmarensis Kiel, Campbell, and Gaillard, 2010, from the Caribbean region (Kiel et al. 2010, Kiel and Hansen 2015).
Stratigraphic and geographic range.—Upper Miocene of Luzon, Philippines.
Family Thyasiridae Dall, 1901
Genus Conchocele Gabb, 1866
Type species: Thyasira bisecta Conrad, 1849; Miocene, Astoria Formation, Oregon, USA.
Conchocele pangasinanensis sp. nov.
Fig. 6A–D.
Zoobank LCID: urn:lsid:zoobank.org:act:157EFA74-F306-45B6-AE 7B-CDB71E19469B
Etymology: For the province of Pangasinan, where the Northern Cement Corporation Quarry is located.
Type material: Holotype NIGSPAL-009, isolated RV with preserved hinge; paratype NIGSPAL-011 from the type locality and horizon.
Type locality: The “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Type horizon: Seep carbonate blocks enclosed in the Amlang Formation (Upper Miocene).
Material.—20 specimens from block B (NIGSPAL-010, 034, NRM Mo 192566–192582, 192588), one specimen from block D (NIGSPAL-035), 2 specimens from block F (NRM Mo 192544–192545), 19 specimens from block G (NIGSPAL-009, NRM Mo 192685–192702), and 4 specimens from the NCC collection (NIGSPAL-036, 037 [3 specimens on one sample]). All from the type locality and horizon.
Dimensions (in mm).—Holotype (NIGSPAL-009): L = 66.0, H = 64.4, W (single) = 22.0; NRM Mo 192544: L = 59.0, H = 59.0, W = 34.4; NRM Mo 192588: L = 61.3, H = 54.5, W = 37.8; paratype (NIGSPAL-011): L = 76.6, H = 63.0, W (single) = 19.7; NIGSPAL-036: L = 77.0+, H = 63.5+, W (single) = 19.7; NIGSPAL-037: L = 68.0, H = 67.2, W (single) = 21.5.
Diagnosis.—Moderate size for genus, well-inflated, outline rounded-quadrate, sharp posterior ridge with wide posterodorsal area, secondary posterior grove present but indistinct, umbones blunt, strongly prosogyrate.
Description.—Moderately sized, well-inflated shells, W/H ratio 0.58–0.69, rounded-quadrate outline, umbones terminal, blunt, prosogyrate; anterior margin long, concave; anterodorsal area broad, with indistinct internal ridge and groove, bordered by blunt ridge; posterodorsal area wide, bordered by deep sulcus and sharp ridge that form deep indentation at posterior margin, secondary ridge indistinct; ventral margin broadly convex, with slight angulation in some specimens, ending posteriorly at posterior sulcus; outer surface with fine, irregular growth lines. Anterior adductor muscle scar narrow, elongate, detached from, and parallel to, pallial line, length about 2/5th of shell length.
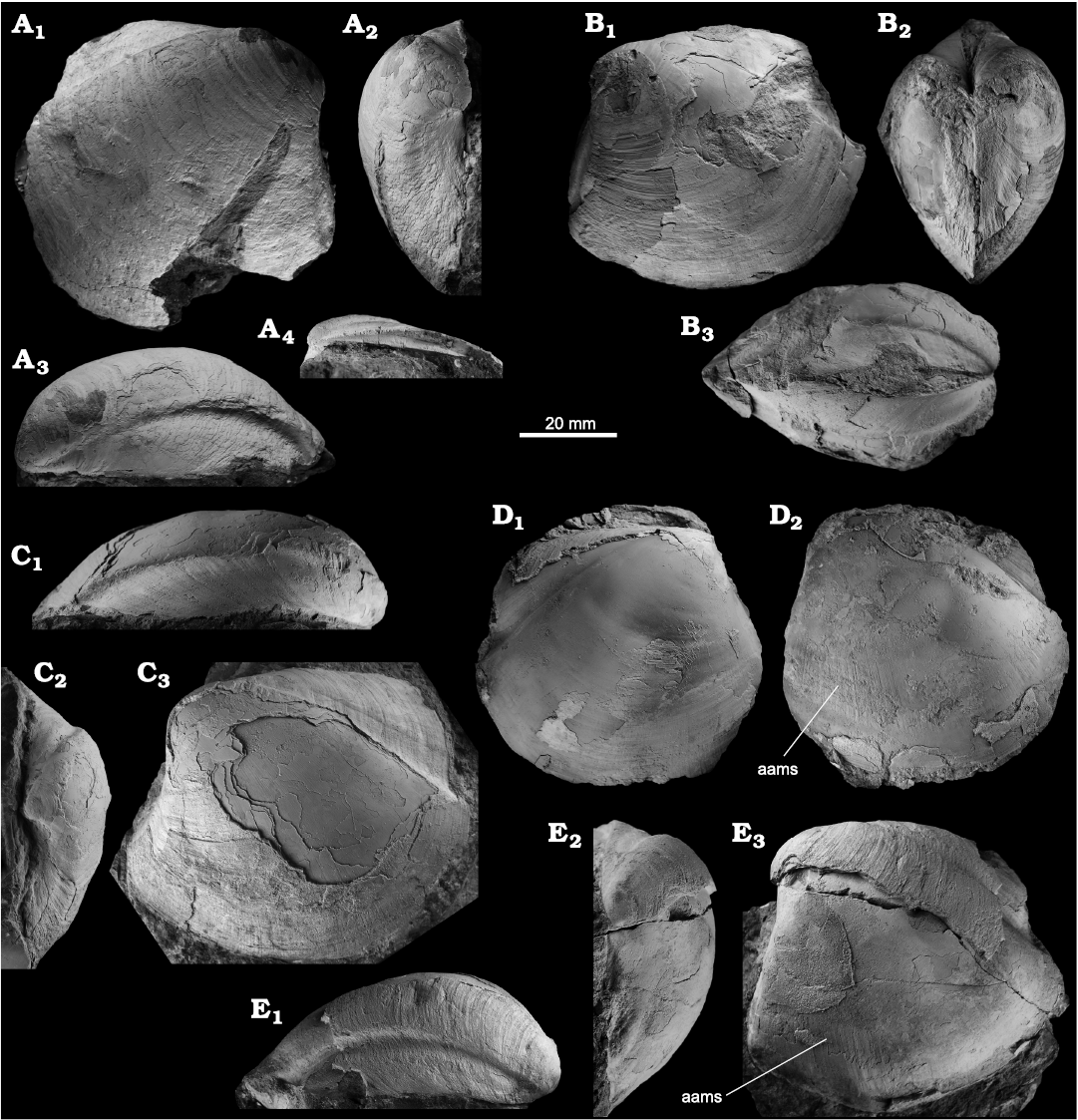
Fig. 6. The thyasirid bivalves Conchocele spp. from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. A–D. Conchocele pangasinanensis sp. nov. A. NIGSPAL-009, holotype, isolated RV, view on outer side (A1), seen from anterior (A2), dorsal view (A3) and close-up on hinge and ligament nymph (A4). B. NRM Mo 192588, articulated specimen, view on LV (B1), seen from anterior (B2), and dorsal view (B3). C. NIGSPAL-011, paratype, large LV, dorsal view (C1), seen from anterior (C2), and lateral view (C3). D. NRM Mo 192544, articulated specimen, view on RV (D1) and on LV, showing elongated aams (D2). E. Conchocele aff. quadrata Yabe and Nomura, 1925, NIGSPAL-012, LV, dorsal view (E1), seen from anterior (E2), and lateral view showing aams (E3). Abbreviation: aams, anterior adductor muscle scar.
Remarks.—Numerous species- and subspecies-level names exist for large, Neogene thyasirids from the western Pacific region, and many specimens have simply been reported as Conchocele bisecta (Conrad, 1849) or C. disjuncta Gabb, 1866 (Yabe and Nomura 1925; Krishtofovich 1936, 1964; Squires and Gring 1996; Majima et al. 2005; Hryniewicz et al. 2017). A revision of these names and taxa is pending. Virtually all of these species, however, differ from Conchocele pangasinanensis by being more elongate. Species with more rounded-quadrate shape as Conchocele pangasinanensis include Conchocele majimai Kiel, Aguilar, and Kase, 2020a, and Conchocele visayaensis Kiel, Aguilar, and Kase, 2020a, two Plio-Pleistocene species from seep deposits in Leyte of the Philippine islands (Kiel et al. 2020a). Conchocele pangasinanensis differs from those two species by its broader umbonal angle (angle between antero- and posterodorsal margins) and its posterodorsal ridge and sulcus do not slope downward as much as in the two species from Leyte. Conchocele adoccasa Van Winkle, 1919, from the Middle Miocene of Trinidad and possibly Oligocene seep deposits of Colombia is flatter and has a narrower posterodorsal area than Conchocele pangasinanensis (Van Winkle 1919; Gill et al. 2005; Kiel and Hansen 2015).
Stratigraphic and geographic range.—Upper Miocene of Luzon, Philippines.
Conchocele aff. quadrata (Yabe and Nomura, 1925)
Fig. 6E.
Material.—One specimen from the NCC collection (NIGSPAL-012, L = 66.0 mm, H = 62.7 mm, W [single] = 21.8 mm). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Remarks.—In overall shape, inflation and especially the narrow posterodorsal area, this specimen resembles Conchocele quadrata Yabe and Nomura, 1925, from an unknown locality in Japan (Yabe and Nomura 1925: pl. 1: 1) though it differs from the Japanese specimen by its slightly less truncate posterior margin and the more convex ventral margin. A specimen illustrated as C. quadrata from the Upper Miocene of Sakhalin Island in eastern Russia (Krishtofovich 1964: pl. 53: 1) is more quadrate and has a wider posterodorsal area than both the Japanese and the Philippine specimens.
Family Lucinidae Fleming, 1828
Genus Megaxinus Brugnone, 1880
Type species: Lucina transversa Bronn, 1831, subsequent designation by Pallary (1904), see Glover and Taylor (1997) for a detailed discussion; Pliocene, Italy.
Megaxinus gorrospei sp. nov.
Fig. 7.
Zoobank LCID: urn:lsid:zoobank.org:act:768EE30D-DE0E-4B20-B6DF-D90B9B0FCC74
Etymology: For Oliver R. Gorrospe, Vice President and General Manager of the Northern Cement Corporation, who aided collections within the quarry.
Type material: Holotype NIGSPAL-015, articulated specimen with most shell material preserved; paratypes: NIGSPAL-013, 014, 016 (all from block A) and NRM Mo 192433 (from block E), small, isolated LV. All from the type locality and horizon.
Type locality: The “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Type horizon: Seep carbonate blocks enclosed in the Amlang Formation (Upper Miocene).
Material.—Type material only.
Dimensions (in mm).—Holotype (NIGSPAL-015): L = 31.4, H = 35.1, W = 18.1; paratypes: NIGSPAL-013: L = 31.6, H = 28.2, W = 15.5; NIGSPAL-016: L = 38.3, H = 37.2, W = 22.5; NRM Mo 192433: L = 27.8, H = 25.4.
Diagnosis.—Subquadrate to broadly triangular, moderately inflated shell; anterodorsal shell margin straight; no lunule; hinge plate short, moderately broad, edentulous; blunt posterior ridge close to dorsal shell margin, posterodorsal area narrow; anterior adductor muscle scar narrow, elongate, length 2/5 of shell length, close to pallial line.
Description.—Shell of medium size for genus, ontogenetic change in shell shape from subquadrate in smaller specimens to broadly triangular in adults. Posterior ridge more pronounced in adults, close to dorsal margin, corresponding posterodorsal area narrow; surface sculpture of commarginal growth lines only: umbones elevated above hinge line, pointed, prosogyrate; posterodorsal and ventral shell margins convex, anterodorsoal margin nearly straight. Lunule large, elongate. Anterior adductor muscle scar elongate, narrow, close and parallel to ventral shell margin, length about 2/5 of shell length; posterior adductor muscle scar just above mid-height of shell, diamond-shaped; pallial line entire, very close to ventral shell margin. Hinge plate broad, short, edentulous, with weak central undulation.
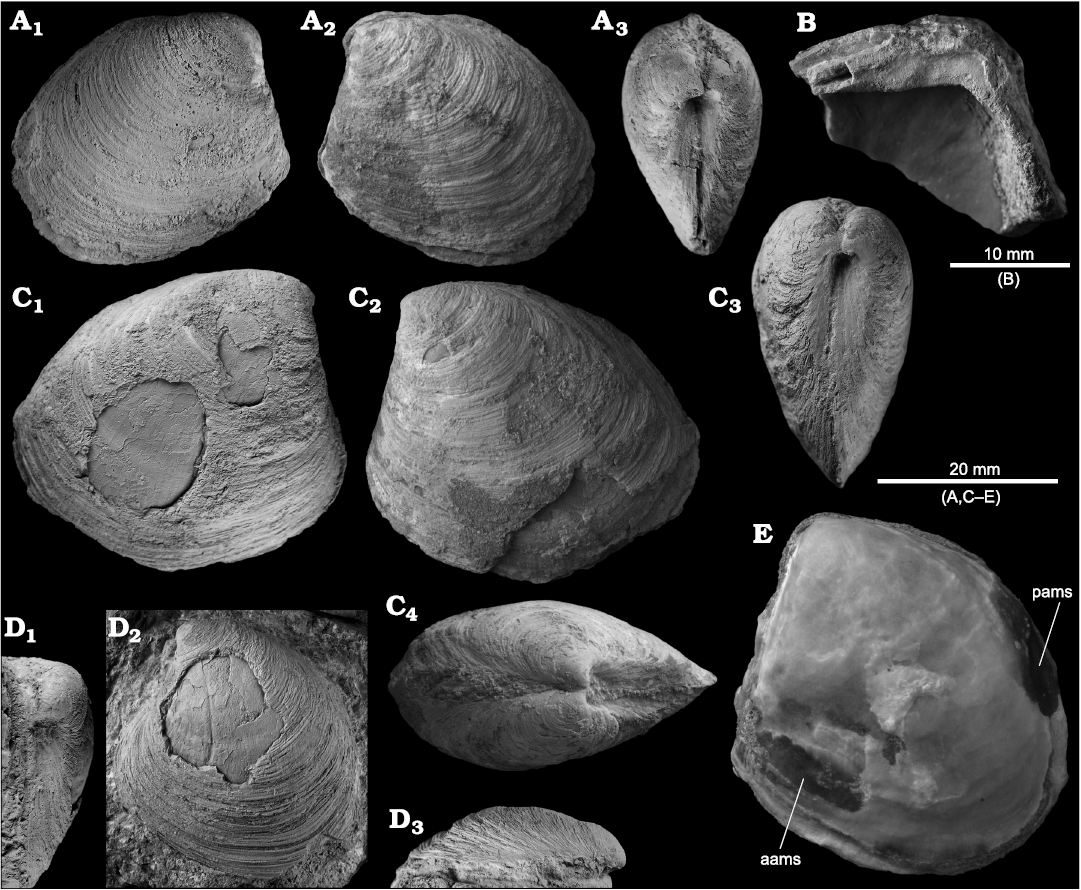
Fig. 7. The lucinid bivalve Megaxinus gorrospei sp. nov. from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. A. NIGSPAL-013, paratype, articulated, medium-sized specimen, view on RV (A1), LV (A2), and anterior margin (A3). B. NIGSPAL-014, paratype, fragment of LV showing the hinge. C. NIGSPAL-015, holotype, large, articulated specimen, views on RV (C1), LV (C2), anterior margin (C3), and dorsal margin (C4). D. NRM Mo 192433, paratype, an isolated LV, views on anterior side (D1), the outer shell (D2), and the posterodorsal side (D3). E. NIGSPAL-016, paratype, internal mold showing pallial line and adductor muscle scars (aams, anterior adductor muscle scar; pams, posterior adductor muscle scar).
Remarks.—Species of Megaxinus show an ontogenetic change in shell shape, from being circular when small, whereas larger shells are higher with a more quadrate anterior (Glover and Taylor 1997: 8). Megaxinus gorrospei sp. nov. resembles those more angular, large species of Megaxinus, and is hence placed in this genus. The extant genus Rasta Taylor and Glover, 2000, includes shells with very similar shell outline and hinge (Taylor and Glover 1997, 2000; Glover and Taylor 1997; Taylor et al. 2005), but Rasta species are much less inflated than Megaxinus gorrospei and smaller.
Externally, shells of Megaxinus gorrospei are very similar to large specimens of Megaxinus ellipticus (Borson, 1825) from the Pliocene of Italy (Kiel and Taviani 2018), and its anterior adductor muscle scar (aams) is very close to the pallial line, as in Megaxinus ellipticus as illustrated by Sacco (1901: pl. 17: 13) and M. stironensis Kiel and Taviani, 2018. In most extant species of Megaxinus illustrated by Glover and Taylor (1997), however, the aams appears to deviate from the pallial line at a somewhat larger angle, especially at its ventral end, than in the Miocene Philippine Megaxinus gorrospei. Most extant species of Megaxinus have a bulge or convexity in the anterodorsal shell margin (Glover and Taylor 1997), absent in Megaxinus gorrospei.
Megaxinus gorrospei differs from Megaxinus ellipticus by its longer and more acutely rounded posterior part of the shell, lacking a distinct lunule, and its narrower hinge plate. The Pliocene Megaxinus stironensis from Italy has a more roundish outline than Megaxinus gorrospei, even at large size, and unlike Megaxinus gorrospei, it bears a lunule (Kiel and Taviani 2018).
Stratigraphic and geographic range.—Upper Miocene of Luzon, Philippines.
Genus Meganodontia Bouchet and Cosel, 2004
Type species: Meganodontia acetabulum Bouchet and Cosel, 2004, by original designation; Recent, Taiwan.
Meganodontia hoernea (Des Moulins, 1868)
Fig. 8.
1868 Lucina hoernea sp. nov.; Des Moulins 1898: 368.
2017 Meganodontia hoernea (Des Moulins, 1868); Kiel and Taviani 2017: 450, figs. 6, 7.
Material.—Three specimens from block A (NIGSPAL-017– 019), 6 specimens from block H (NRM Mo 192403–192409), one from block I (NRM Mo 192661). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Dimensions (in mm).—NRM Mo 192403: L = 87.0, H = 85.0; NRM Mo 192404: L = 80.0, H = 75.5; NRM Mo 192405: L > 100.0; NIGSPAL-017: L = 105.2, H = 103.0, W = 66.6; NIGSPAL-018: L = 90.4, H = 84.4, W = 49.1; NIGSPAL-019: L = 84.1, W = 46.1.
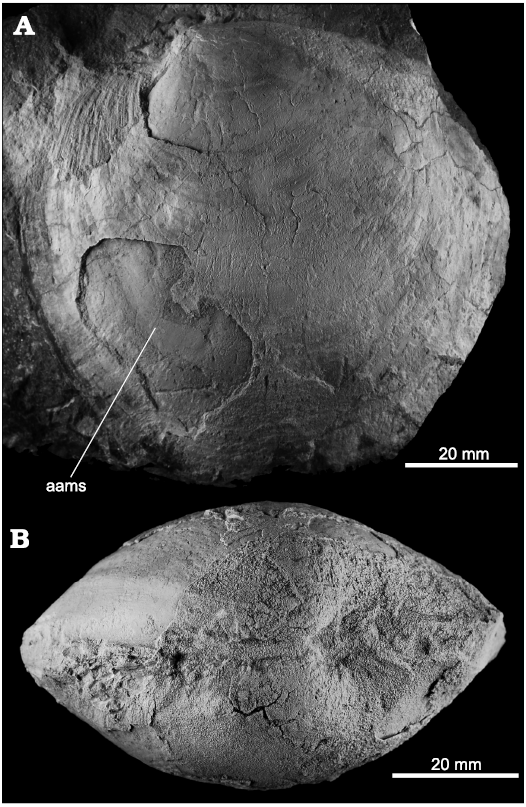
Fig. 8. The lucinid bivalve Meganodontia hoernea (Des Moulins, 1868) from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. A. NRM Mo 192403, isolated LV showing anterior adductor muscle scar (aams). B. NRM Mo 192404, small, articulated specimens in dorsal view.
Remarks.—Meganodontia hoernea is widespread and frequent in Middle and Late Miocene-aged seep deposits in northern Italy (Taviani 1994; Kiel and Taviani 2017), and the Late Miocene specimens reported here from the Philippines are virtually identical to well-preserved specimens of M. hoernea illustrated in Kiel and Taviani (2017: fig. 7).
The extant and Pleistocene Meganodontia acetabulum differs from M. hoernea by its more strongly convex posterodorsal margin and narrower posterodorsal area (Bouchet and Cosel 2004; Kiel and Taviani 2017; Kiel et al. 2020a). An Early Miocene Meganodontia sp. from Cuba (Kiel and Hansen 2015: fig. 7A–C) is more strongly inflated and has a less distinct posterior sulcus. The medium-sized M. haunuiensis Amano, Little, and Campbell, 2018, from Early Miocene seep deposits in New Zealand differs mainly by having a shorter anterior adductor muscle scar (see Amano et al. 2018: fig. 3F2).
Two large Neogene lucinids described from Taiwan most likely also belong to Meganodontia. Among them, the Pliocene Loripes goliath Yokoyama, 1928, differs from the Philippine specimens of Meganodontia hoernea by having stronger, more elevated umbones (Yokoyama 1928: 86, pl. 9: 8, pl. 10: 1). The same applies to Loripes pregoliath Hu and Tseng in Hu, 1995, from the Miocene Nanchuang Formation in Taiwan, which is also smaller (L = 75.0 mm vs. L > 100.0 mm in the Philippine M. hoernea) and more inflated than M. hoernea from the Northern Cement Corporation quarry (Hu 1995: 1995–1996, pl. 349: 10, 13; pl. 350: 4, 6, 7).
Stratigraphic and geographic range.—Middle and Upper Miocene strata of Italy and the Philippines.
Genus Lucinoma Dall, 1901
Type species: Lucina filosa Stimpson, 1851, by original designation; Recent, North Atlantic Ocean.
Lucinoma saetheri Amano, Little, and Campbell, 2018
Fig. 9.
Material.—Two specimens from block A (NIGSPAL-020, 021), three specimens from block B (NRM Mo 192623–192625), one specimen from block E (NRM Mo 192434). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Dimensions (in mm).—NIGSPAL-020: L = 44.0, H = 43.5; NIGSPAL-021: L = 52.1, H = 45.1, W = 29.0.
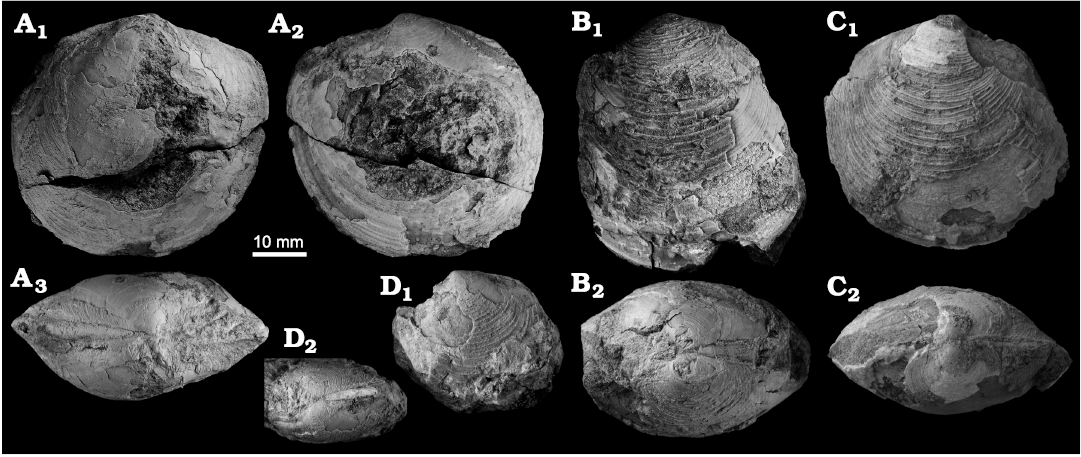
Fig. 9. The lucinid bivalve Lucinoma saetheri Amano, Little, and Campbell, 2018, from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. A. NRM Mo 192623, articulated specimen, view RV (A1), LV (A2) and on dorsal side (A3). B. NRM Mo 192624, damaged, articulated specimen, view on LV showing fine concentric ribbing (B1) and on dorsal side showing inflation and ligament (B2). C. NIGSPAL-020, articulated specimen with well-preserved concentric ribs, view on RV (C1) and on dorsal side (C2). D. NRM Mo 192625, small articulated specimen, view on LV (D1) and on dorsal side (D2).
Remarks.—A similar species that should be re-assigned to Lucinoma is Lucina (Myrtea) butonensis Beets (1942: pl. 30: 143–146); the anterodorsal margin of Lucinoma saetheri is more acutely rounded than in L. butonensis.
Stratigraphic and geographic range.—Lower and Middle Miocene of New Zealand (North Island), Upper Miocene of Luzon, Philippines.
Lucinoma acutilineata (Conrad, 1849)
Fig. 10.
1849 Lucina acutilineata sp. nov; Conrad 1849: 725, pl 18: 2.
1962 Lucinoma acutilineatum (Conrad): Kamada 1962: 95, pl. 9: 10–16, pl. 10: 1–3.
1963 Lucinoma acutilineata (Conrad), Moore 1963: 70, pl. 15: 7–10, 12.
Material.—Four specimens from block A (NIGSPAL-022– 024), 1 specimen from block B (NRM Mo 192584), 3 specimens from block E (NRM Mo 192428–192430), 2 specimens from block I (NRM Mo 192634, 192635). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Dimensions (in mm).—NIGSPAL-023: L = 49.5, H = 49.9, W = 22.1; NIGSPAL-024: L = 36.2, H = 33.3, W = 13.4.

Fig. 10. The lucinid bivalve Lucinoma acutilineata (Conrad, 1947) from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. A. NIGSPAL-023, large articulated specimen, view on RV. B. NIGAPAL-024, medium-sized, articulated specimen, view on RV showing outline and widely spaced commarginal ribs (B1), and dorsal view showing inflation, lunule and escutcheon. C. NRM Mo 192429, articulated specimen with most shell material detached, showing muscle scars and pallial line, view on RV (C1), LV (C2) and on dorsal side (C3). D. NRM Mo 192584, specimen with broken apical portion, view on LV. E. NRM Mo 192430, small specimen, RV mostly an internal mold showing muscle scars and pallial line (E1), LV with preserved shell (E2), and view on dorsal side showing ligament (E3). F. NRM Mo 192428, small RV showing external sculpture. G. NRM Mo 192634, small LV showing external sculpture.
Remarks.—The Miocene specimens of Lucinoma acutilineata from western North America and Japan show a range of outlines from roundish to pentangular shells (Hirayama 1954; Kamada 1962; Moore 1963). The roundish Philippine specimens documented here are quite similar to the holotype, which was re-illustrated in Moore (1963: pl. 15: 9). Similarly flat Lucinoma species from the Neogene of the Philippines include Lucinoma aff. kastoroae Cosel and Bouchet, 2008, from the Pliocene Liog-Liog Point seep deposit, Leyte, Philippines and Lucinoma kosatorea Kiel, Aguilar, and Kase, 2020, from the Pleistocene Cambantug Point seep deposit, Leyte. Both species have a more acutely rounded anterior shell margin than L. acutilineata.
Stratigraphic and geographic range.—Miocene; Philippines, Japan, northwestern North America.
Genus Elliptiolucina Cosel and Bouchet, 2008
Type species: Elliptiolucina magnifica Cosel and Bouchet, 2008, by original designation; Recent, Philippines, 520–550 m depth.
Elliptiolucina sp.
Fig. 11.
Material.—Six specimens from block E (NRM Mo 192466–192471), 2 specimens from block H (NRM Mo 192411–192412), 1 specimen from block I (NRM Mo 192664). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Dimensions (in mm).—NRM Mo 192470: L = 21.0, H = 18.8, W = 9.0; NRM Mo 192469: LV L = 17.2, H = 12.0; NRM Mo 192412: LV L = 15.0, H = 14.4; NRM Mo 192411: RV L = 14.3+, H = 11.7; NRM Mo 192664, L = 21.7+, H = 21.8.
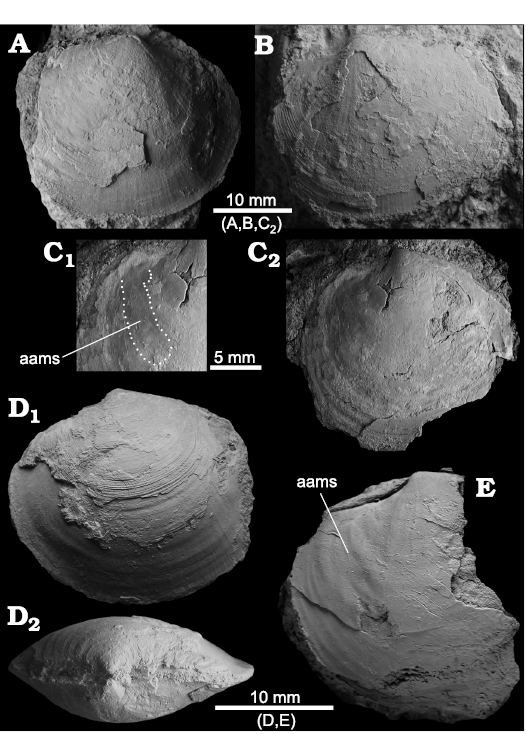
Fig. 11. The lucinid bivalve Elliptiolucina sp. from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. A. NRM Mo 192411, small RV. B. NRM Mo 192412, small LV. C. NRM Mo 192412, small LV, showing aams (C1) and in general view (C2). D. NRM Mo 192470, larger, articulated specimen, view on LV (D1) and in dorsal view (D2). E. NRM Mo 192664, large internal mold of LV. Abbreviation: aams, anterior adductor muscle scar.
Remarks.—Species of Elliptiolucina typically have lengths between 70.0 and 140.0 mm (Cosel and Bouchet 2008; Okutani 2011; Kiel 2013; Glover and Taylor 2016; Kiel et al. 2020a). This applies also to the two species of Elliptiolucina reported from Pliocene seep deposits in Leyte, Philippines (E. ingens Okutani, 2011, and E. fernandoi Kiel, Aguilar, and Kase, 2020a), which further differ from the species reported here by having less convex ventral margins (Kiel et al. 2020a). An exception is the Middle Miocene Elliptiolucina neozelandica Amano, Little, and Campbell, 2018, which reaches only about 35.0 mm in length, but differs from the species reported here by being more elongated and having a less convex ventral margin (Amano et al. 2018). We consider the small specimens reported here as juveniles and refrain from assigning them to any described or new species.
Family Vesicomyidae Dall and Simpson, 1901
Genus Pliocardia Woodring, 1925
Type species: Anomalocardia bowdeniana Dall, 1903, by original designation; Pliocene, Bowden Formation, Jamaica.
Pliocardia ballesterosi sp. nov.
Fig. 12.
Zoobank LCID: urn:lsid:zoobank.org:act:67677ED9-BF0C-4BD1-B 194-A6BFCFC836DF
Etymology: For Alfredo D. Ballesteros, Jr., Sustainable Development and Community Relations Department Head of the Northern Cement Corporation, who aided collecting at the quarry.
Type material: Holotype NIGSPAL-025, isolated RV with preserved shell. Paratypes: NRM Mo 192546–192552, articulated and disarticulated specimens mostly with preserved shell material, from the type locality and horizon.
Type locality: The “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Type horizon: Seep carbonate blocks enclosed in the Amlang Formation (Upper Miocene).
Material.—Nine specimens from block B (NRM Mo 192613–192618, NIGSPAL-025, 027), 20 specimens from block F (NRM Mo 192546–192565). All from the type locality and horizon.
Dimensions (in mm).—NIGSPAL-025: L = 31.0, H = 24.0, W (single) = 11.0; NRM Mo 192552: L = 32.4, H = 24.7, W = 20.0; NRM Mo 192550: L = 29.7, H = 22.3, W = 16.8; NRM Mo 192546: L = 30.0, H = 2.1, W (single) = 8.0; NRM Mo 192551: L = 35.0, H = 25.7, W (single) = 10.0; NRM Mo 192547: L = 30.6, H = 25.2, W (single) = 7.8); NIGSPAL-026: L = 26.1, H = 20.1, W = 16.0.
Diagnosis.—Large, well inflated, oval to subquadrate shell, posterodorsal sulcus poorly developed; blunt, elevated, prosogyrate umbones in very anterior position; lunular incision well-defined; small, shallow pallial sinus.
Description.—Shell large compared to the type species of Pliocardia, but small compared to members of the “cordata group”, reaching 32.4 mm in length; outline oval to subquadrate, posterior margin subtruncated to acutely rounded, ventral and posterodorsal margin convex, anterior margin narrow, more-or-less acutely rounded; posterior ridge and groove poorly developed on most specimens; lunular incision deep, outline broadly lenticular; escutcheon narrow, poorly defined; no external sculpture except for numerous fine growth increments. Hinge plate moderately broad, rather short, three cardinal teeth in each valve; in RV, cardinal strong, pointing anteroventrally, cardinal 3a short, parallel to shell margin, 3b elongate, bifurcated, raised edges, subparallel to shell margin, posterior edge about three time the length as anterior edge; in LV, cardinal 2a strong, pointing anteriorly, cardinal 2b blunt, broadly trapezoid, with two short raised edges, cardinal 4b moderately long, parallel to shell margin. Anterior adductor muscle scar oval, with narrow connection to small pedal retractor scar; posterior adductor muscle scar round to slightly oval, distinct but not deep; pallial line entire, with shallow pallial sinus.
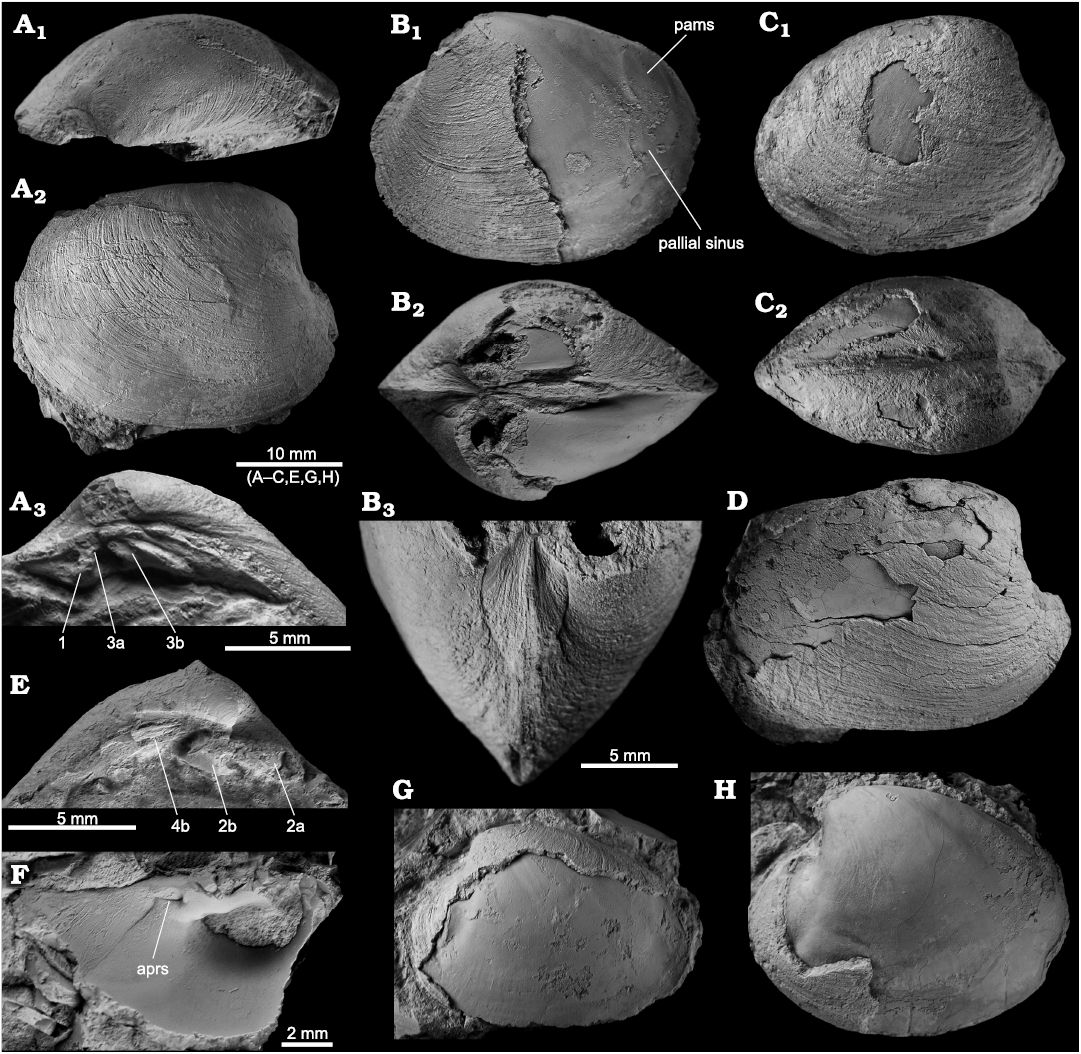
Fig. 12. The vesicomyid bivalve Pliocardia ballesterosi sp. nov. from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. A. NIGSPAL-025, holotype, an isolated RV with preserved hinge, in apical view showing lunular incision (A1), view on outer shell showing sculpture and outline (A2), and close-up on hinge with cardinal teeth numbers indicated (A3). B. NRM Mo 192552, paratype, articulated specimen with partially removed shell, view on LV showing pallial sinus and pams (B1), in apical view showing strong inflation (B2), close-up on anterior side showing lunular incision (B3). C. NRM Mo 192550, paratype, articulated shell with only moderate inflation, view on RV (C1) and apical view (C2). D. NRM Mo 192549, paratype, outer side of large RV. E. NRM Mo 192546, paratype, hinge of LV with cardinal teeth numbers indicated. F. NRM Mo 192548, paratype, fragment of interior of RV showing anterior muscle scars. G. NRM Mo 192551, paratype, isolated LV showing pallial line. H. NRM Mo 192547, paratype, large, isolated LV showing pallial line. Abbreviations: aprs, anterior pedal retractor scar; pams, posterior adductor muscle scar.
Remarks.—Most similar to P. ballesterosi in overall shell shape is the extant Pliocardia kuroshimana Okutani, Fujikura, and Kojima, 2000, which occurs also in a Pleistocene seep deposit in the central Philippines (Kiel et al. 2000). Pliocardia ballesterosi differs from P. kuroshimana mainly by its smaller and less elevated umbones, with P. kuroshimana having the point of maximum inflation in the shell’s center whereas it is displaced toward the anterior in P. ballesterosi, and by having a narrower but longer ligament nymph. Another similar species is “Pliocardia” ticaonica Dall, 1908a, extant off Ticao island (central Philippines) and also found in the early Pleistocene Cambantug Point seep deposit in Leyte, Philippines (Kiel et al. 2020a). Pliocardia ballesterosi differs from “P.” ticaonica by having distinct lunular incision, which is only very indistinct in “P.” ticaonica, by its slightly less prosogyrate and prominent umbones, and by having a less distinct posterodorsal ridge than “P.” ticaonica.
The Early to Middle Miocene Pliocardia kawadai (Aoki, 1954) from Japan has a deeper posterior groove and associated wider posterodorsal area, and its pallial sinus reaches up to the anterior margin of the posterior adductor muscle scar (see Amano and Kiel 2012: figs. 7 and 12), whereas in Pliocardia ballesterosi the pallial sinus is entirely below the posterior muscle scar, and the pallial line reaches the scar on its ventral side. The Middle Miocene Pliocardia? tanakai Miyajima, Nobuhara, and Koike, 2017, from Japan differs from P. ballesterosi by its more elongate-oval shell outline, smaller and more pointed umbones. Pliocardia? tanakai is also less inflated and thus has a narrower and more elongate lunular area, and lacks a pallial sinus (Miyajima et al. 2017).
Stratigraphic and geographic range.—Upper Miocene of Luzon, Philippines.
Genus Archivesica Dall, 1908
Type species: Callocardia gigas Dall, 1896, by original designation; Recent, Gulf of California.
Remarks.—Archivesica is used here in the sense of Amano and Kiel (2010) and Amano et al. (2019a); in molecular studies, this genus is of often referred to as “gigas/kilmeri complex” or “gigas group” (Audzijonyte et al. 2012; Valdés et al. 2013; Johnson et al. 2017).
Archivesica? sp. 1
Fig. 13.
Material.—One specimen from block B (NIGSPAL-029), 17 specimens from block C (NIGSPAL-028), 3 specimens from block E (NRM Mo 192418–192420), 3 specimens from block I (NRM Mo 192670–192672). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Dimensions (in mm).—NRM Mo 192670: L = 34.5, H = 18.7; NRM Mo 192671: L = 68.3, H = 38.4; NRM Mo 192672: L = 16.5, H = 9.3.
Description.—Elongate, oval shell, umbones positioned at anterior 20% of shell; external surface smooth, internal surface with radial striation; hinge plate short, moderately broad; RV hinge with two radiating, strong cardinal teeth, cardinal 1 pointing downward, cardinal 3b pointing posteriorly; LV hinge with three radiating, strong cardinals, 2a and 2b at almost right angles to each other, 4b short, subparallel to dorsal margin.

Fig. 13. The vesicomyid bivalve Archivesica? sp. 1 from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. A. NRM Mo 192671, views on hinges of RV (A1) and LV (A2), cardinal teeth numbers indicated; and view on outer side of LV showing interior radial striations (A3). B. NRM Mo 192418, small, articulated specimen, in dorsal view showing absence of lunule or lunular incision (B1), and view on RV (B2). C. NRM Mo 192419, very small specimen showing pallial line and posterior adductor muscle scar. D. NRM Mo 192672, small, isolated LV. E. NRM Mo 192670, medium-sized, isolated RV showing internal radial striation.
Remarks.—These specimens have the general outline of other elongate, fossil Archivesica, and also the hinge dentition resembles that of other Archivesica species. However, they are only hesitantly assigned to Archivesica because the available material does not show the taxonomically important pallial sinus. The internal radial striation seen on some of the specimens is uncommon among vesicomyids, but has for example been documented from the elongate, extant West African vesicomyid Christineconcha regab (Cosel and Olu, 2009) (Krylova and Cosel 2011).
Archivesica? sp. 2
Fig. 14.
Material.—Four specimens from block I (NRM Mo 192676–192679). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Dimensions (in mm).—NRM Mo 192676: L = 51.8, H = 43.2, W = 23.0; NRM Mo 192677: L = 50.6, H = 41.0, W = 24.1; NRM Mo 192678: L = 38.2, H = 31.4, W = 13.7; NRM Mo 192679: L = 17.2, H = 12.3, W = 7.2.
Description.—Short, oval shell, umbones elevated, prosogyrate, positioned at anterior 27–31% of total shell length; anterior margin acutely rounded, posterior margin wide, broadly rounded. External surface with fine growth increments, internal surface with radial striation; no lunular incision; hinge plate narrow, RV with three thick, short cardinal teeth: cardinal 1 displaced anteriorly, 3a just anterior to beak, pointing anteriorly, 3b just posterior to beak, pointing posteriorly; no subumbonal pit.

Fig. 14. The vesicomyid bivalve Archivesica? sp. 2 from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. A. NRM Mo 192676, large articulated specimen, view on LV showing interior posterior ridge (A1), dorsal view showing moderate inflation and lunule (A2). B. NRM Mo 192678, articulated specimen, view on LV showing general outline. C. NRM Mo 192677, isolated RV, view on hinge.
Remarks.—These specimens show an unusual combination of shell characters. The overall shape (short, oval, little inflation) resembles that of species of Wareniconcha or Calyptogena (see Krylova and Sahling 2006; Cosel and Olu 2009). But the hinge with its strong, thick, radiating teeth is very similar to those of Archivesica nankaiensis (Okutani, Kojima, and Ashi, 1996), A. soyoae (Okutani, 1957), A. solidissima (Okutani, Hashimoto, and Fujikura, 1992a) and A. okutanii (Kojima and Ohta, 1997), all of which, however, are much more elongated and inflated (Okutani 1957; Okutani et al. 1992a; Kojima and Ohta 1997; Okutani et al. 1997; Sasaki et al. 2005). Furthermore, species of Archivesica possess a subumbonal pit, lacking in the present specimens. Therefore, we only hesitantly assign this species to Archivesica.
Genus Wareniconcha Cosel and Olu, 2009
Type species: Vesicomya guineensis Thiele in Thiele and Jaeckel, 1931, by original designation; Recent, central eastern Atlantic Ocean.
Wareniconcha sp.
Fig. 15.
Material.—Three specimens from block F (NRM Mo 192537–192539). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Dimensions (in mm).—NRM Mo 192539: L = 49.0, H = 34.0, W (single) = 13.0; NRM Mo 192538: L = 40.0, H = 33.0, W = 22.0; NRM Mo 192537: L = 44.0+, H = 41.0, W (single) = 15.0.
Description.—Medium-sized, moderately inflated, oval shell; umbones elevated, prosogyrate; blunt posterior ridge creating broad posterodorsal area, no escutcheon and lunule; hinge plate narrow, RV with three thick cardinal teeth arranged more-or-less parallel to dorsal shell margin.

Fig. 15. The vesicomyid bivalve Wareniconcha sp. from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. A. NRM Mo 192537, large, slightly disarticulated specimen, view on RV (A1), view on LV and RV hinge plate (A2), and close-up on RV hinge with cardinal teeth numbers indicated (A3). B. NRM Mo 192538, articulated, medium-sized specimen, view on LV (B1) and on dorsal side showing inflation and lack of lunular incision (B2).
Remarks.—The available specimens are too poorly preserved for species-level identification.
Vesicomyidae gen. et sp. indet. 1
Fig. 16A.
Material.—15 specimens from block E (NRM Mo 192438–192453), many specimens on a rock sample from the NCC collection. Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Dimensions (in mm).—NRM Mo 192439: L = 15.6, H = 13.0, W = 8.9; NRM Mo 192440: L = 20.8, H = 15.7; NRM Mo 192442: L = 23.2, H = 19.4; NRM Mo 192444: L = 22.3, H = 17.3, W = 12.4; NRM Mo 192446: L = 20.0, H = 14.6, W = 11.3.

Fig. 16. Small vesicomyid bivalves from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. A. Vesicomyidae gen. et sp. indet. 1, NRM Mo 192444, articulated specimen, view on LV (A1) and view on dorsal side showing lunular incision and ligament (A2). B. Vesicomyidae gen. et sp. indet. 2, NRM Mo 192426, articulated specimen, view on LV (B1) and view on dorsal side showing lunular incision and escutcheon (B2).
Remarks.—This locally very abundant, small-sized vesicomyid defies a more precise classification because the available material does not show internal characters such as hinge, muscle scars and pallial line. It is smaller and more elongate than Pliocardia ballesterosi. Its overall shell outline is similar to that of Isorropodon cf. perplexum Sturany, 1896, from Pliocene seep deposits in Leyte Island, Philippines (Kiel et al. 2020a), but that species lacks a lunular incision, clearly visible in vesicomyid sp. 1 reported here.
Vesicomyidae gen. et sp. indet. 2
Fig. 16B.
Material.—Seven specimens from block E (NRM Mo 192422–192427, 192435). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Dimensions (in mm).—NRM Mo 192424: L = 11.7, H = 9.5; NRM Mo 192425: L = 14.7, H = 12.0; NRM Mo 192426: L = 10.8, H = 9.8, W = 7.0; NRM Mo 192427: L = 11.8, H = 10.8, W = 8.3; NRM Mo 192435: L = 10.2, H = 8.4.
Remarks.—This small species is shorter and more inflated than vesicomyid sp. 1 reported above. It resembles the smaller of the specimens reported as Vesicomya margotae Beets, 1953, from the Pliocene of Leyte Island, Philippines (Kiel et al. 2020a: fig. 18A), and is also similar to Vesicomya nakaii Okutani, 1962, for example as illustrated from the Pliocene of Leyte Island, Philippines (Kiel et al. 2020a: fig. 19A). But as internal characters cannot be assessed, we prefer to keep these specimens in open nomenclature.
Family uncertain
Genus Sisonia nov.
Type species: Sisonia frijellanae sp. nov., see below.
Species included: Possibly belonging to Sisonia are three specimens identified as “Solemyidae gen. indet.” from the Miocene Moonlight North seep deposit in New Zealand (Saether et al. 2016: fig. 3B). The figured specimen shows a similar overall outline and muscle scar pattern, though it is much less inflated than Sisonia frijellanae. Further specimens most likely belonging to Sisonia have been found in a Pleistocene seep deposit in Taiwan (SK unpublished data).
Etymology: For the Municipality of Sison, where the type locality is located in Pangasinan province.
Diagnosis.—As for the type species.
Remarks.—This genus is difficult to place among the heterodont bivalves. Its overall outline and hinge resembles some vesicomyids, though the strong posterior inflation is uncommon in that family. The most distinctive difference of Sisonia frijellanae from vesicomyids is the rather indistinct and elongate, pear-shaped anterior adductor muscle scar. Vesicomyids of this size typically have a distinct, deeply impressed anterior adductor muscle scar bound by a strong ridge on the posterior side. Trapeziidae have a somewhat similar outline and also lack a pallial sinus, but their umbones is more prosogyrate, and they possess posterior lateral teeth, absent in Sisonia frijellanae. Remarkably similar shell shapes, both internally and externally, can be found among the galeommatoids (Morton and Scott 1989), for example the lasaeid Neaeromya rugifera (Carpenter, 1864) or Bornia sebetia (Costa, 1830). However, galeommatoids rarely reach sizes beyond 10.0 mm and are thus much smaller than the almost 70.0 mm long Sisonia frijellanae. Interestingly, at least one galeommatoid species, the saxicavellin Atopomya dolobrata Oliver, 2013, harbors symbiotic bacteria in its gills (Oliver 2013).
Sisonia frijellanae sp. nov.
Fig. 17.
Zoobank LCID: urn:lsid:zoobank.org:act:08086988-4440-4724-B293-0C12A48F2B82
Etymology: For Myra Vanessa A. Frijellana, Environmental Health and Safety Staff of the Northern Cement Corporation, who aided collecting at the quarry.
Type material: Holotype NIGSPAL-030, articulated specimen with exposed RV hinge. Paratypes: NRM Mo 192594–192596, 192598 (block B), NRM Mo 192707 (block G), NRM Mo 192673, 192674 (block I), articulated specimen with partially preserved shell, from the type locality and horizon.
Type locality: The “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Type horizon: Seep carbonate blocks enclosed in the Amlang Formation (Upper Miocene).
Material.—25 specimens from block B (NIGSPAL-031 [for a lot of 8 specimens]; NRM Mo 192594–192599, 192601–192611), 14 specimens from block G (NRM Mo 192703–192714), 3 specimens from block I (NRM Mo 192673–192675), one specimen from block J (NRM Mo 192536). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Dimensions (in mm).—NIGSPAL-030: L = 39.6, H = 25.6, W = 20.7; NRM Mo 192595: L = 37.4, H = 24.7, W = 19.0; NRM Mo 192707: L = 32.4, H = 22.4, W = 13.5; NRM Mo 192674: L = 68.7, H = 40.0, W = 31.8; NRM Mo 192673: L = 48.5, H = 31.3, W (single) = 10.5.
Diagnosis.—Shell trapezoid to short modioliform in outline, strongly inflated, umbones in anterior third of shell, slightly elevated above hinge line, nearly orthogyrate; blunt ridges running from umbo to antero- and posterodorsal margins, area in between depressed; no lunule or escutcheon, ligament external; sculpture of fine growth increments only, internally with radiating striations; adductor muscle scars indistinct, pallial line entire, apparently no sinus; hinge with two strong, radiating cardinals in each valve.
Description.—Shell reaching 68.7 mm in length, strongly inflated, widest in posterior third of shell, outline short modioliform, dorsal and ventral margins subparallel, posterior margin truncate, anterior margin rounded to acutely rounded; ventral margin concave. Umbones broad, slightly elevated above hinge line, in anterior third of shell; ridge from umbo to posteroventral corner broad but distinct, ridge from umbo to anteroventral corner broad, indistinct; area between ridges deeply depressed, causing concavity of ventral margin; no sculpture except fine growth increments. Ligament short, resting on deeply incised nymphs; no lunule and escutcheon. Anterior adductor muscle scar indistinct, elongated pear-shaped, broadening ventrally, situated about midline of shell; pallial line starting just posterior of its ventral base; pallial line faintly impressed, moderately distant from shell margin, undulating, entire, apparently no sinus; anterior pedal retractor scar elongate, connected to adductor scar; posterior adductor muscle scar indistinct, shaped like an inverted comma, situated dorsally to groove running from umbo to posterior side of shell. Hinge plate narrow, subumbonal pit and two strong, short, radiating cardinals in each valve, RV with cardinal 1 and 3b, 3a apparently reduced, LV with cardinal 2a and 2b, cardinal 4 may be reduced.
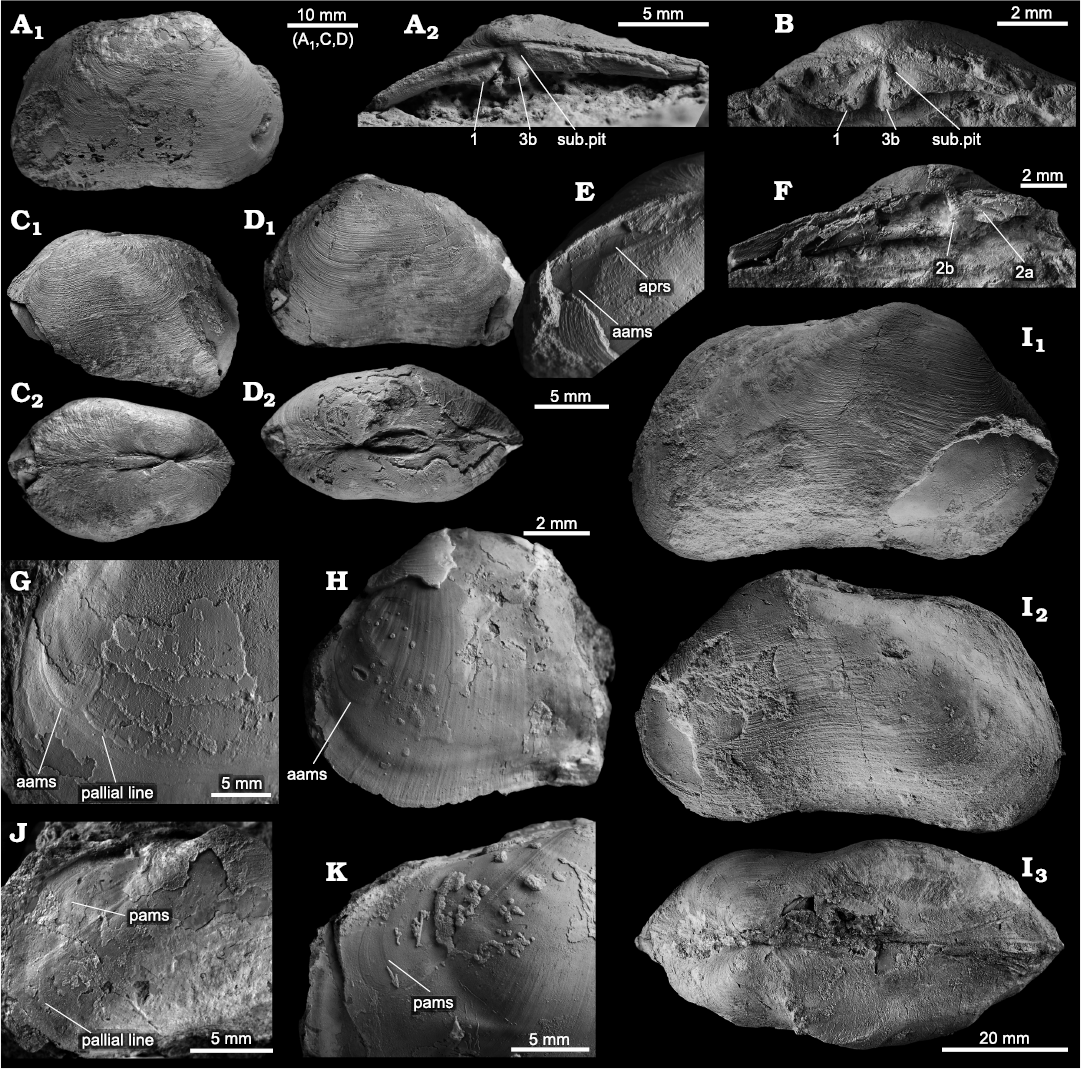
Fig. 17. The bivalve Sisonia frijellanae gen. et sp. nov. from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. A. NIGSPAL-030, holotype, articulated specimen with exposed RV hinge; view on exterior of RV (A1), close-up on hinge of RV (A2). B. NRM Mo 192594, hinge of isolated right valve. C. NRM Mo 192707, articulated specimen with ventral side missing, view on left valve (C1) and dorsal view showing inflation and short ligament (C2). D. NRM Mo 192595, paratype, articulated specimen, view on left valve (D1) and dorsal view (D2). E. NRM Mo 192596, paratype, close-up on anterior side of left valve of an articulated specimen, showing aams and aprs. F. NRM Mo 192598, paratype, hinge of isolated left valve. G. NRM Mo 192673, close-up on anterior side of isolated internal mold of left valve, showing aams and pallial line. H. NRM Mo 192596, paratype, close-up on anterior side of isolated internal mold of left valve, showing aams and internal, radial striation. I. NRM Mo 192674, paratype, very large specimen with mostly intact shell, views on right valve (I1), left valve (I2) and on dorsal side showing median sulcus and posterior ridge (I3). J. NRM Mo 192703, close-up on posterior side of RV showing pams and pallial line. K. NRM Mo 192704, close-up on posterior side of RV showing pams. Abbreviations: aams, anterior adductor muscle scar; aprs, anterior pedal retractor scar; pams, posterior adductor muscle scar; sub.pit, subumbonal pit; cardinal teeth numbers indicated in images A2, B and F.
Stratigraphic and geographic range.—Upper Miocene of Luzon, Philippines.
Class Gastropoda Cuvier, 1795
Unidentified limpet gastropod
Fig. 18A, B.
Material.—One poorly preserved specimen from block B (NRM Mo 192585), 1 specimen from block F (NRM Mo 192421), 1 specimen from block G (NRM Mo 192715), 1 large specimen from block I (NRM Mo 192656). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Dimensions (in mm).—NRM Mo 192421: L = 9.0, W = 7.5, H = 4.6; NRM Mo 192715: L = 6.2, W = 4.5, H = 2.5; NRM Mo 192656: L = 15.2, W = 11.8, H = 7.5.
Remarks.—Shape and the cancellate ornament of these specimens are very similar to species typically assigned to Bathyacmaea (Okutani et al. 1992b; Jenkins et al. 2007a; Chen et al. 2019).
Order Neomphalina McLean, 1981
Family Neomphalidae McLean, 1981
Genus Retiskenea Warén and Bouchet, 2001
Type species: Retiskenea diploura Warén and Bouchet, 2001, by original designation; Recent, North Pacific Ocean.
Retiskenea sp.
Fig. 18C.
Material.—One specimen from block F (NRM Mo 192540, W = 1.0 mm). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Remarks.—The single small specimen is embedded in matrix, blocking the view on the aperture and hence not allowing an identification to species level.
Order Seguenziida Haszprunar, 1986
Family Cataegidae McLean and Quinn, 1987
Genus Cataegis McLean and Quinn, 1987
Type species: Cataegis toreuta McLean and Quinn, 1987, by original designation; Recent, Gulf of Mexico and Caribbean Sea.
Cataegis sp.
Fig. 18D, E.
Material.—One specimen from block F (NRM Mo 192543), 2 specimens from block I (NRM Mo 192665, 192666). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Dimensions (in mm).—NRM Mo 192543: H = 11.5, W = 10.0; NRM Mo 192665: W = 13.0, H = 11.6; NRM Mo 192666: W = 10.7, H = 9.5.
Description.—Low turbiniform shell, ca. 3.5 evenly convex whorls bearing four strong, equally spaced spiral keels, lowest ridge forming basal margin, surface of keels with fine nodular ornament; growth increments fine, strongly opisthocline; base with three further keels.
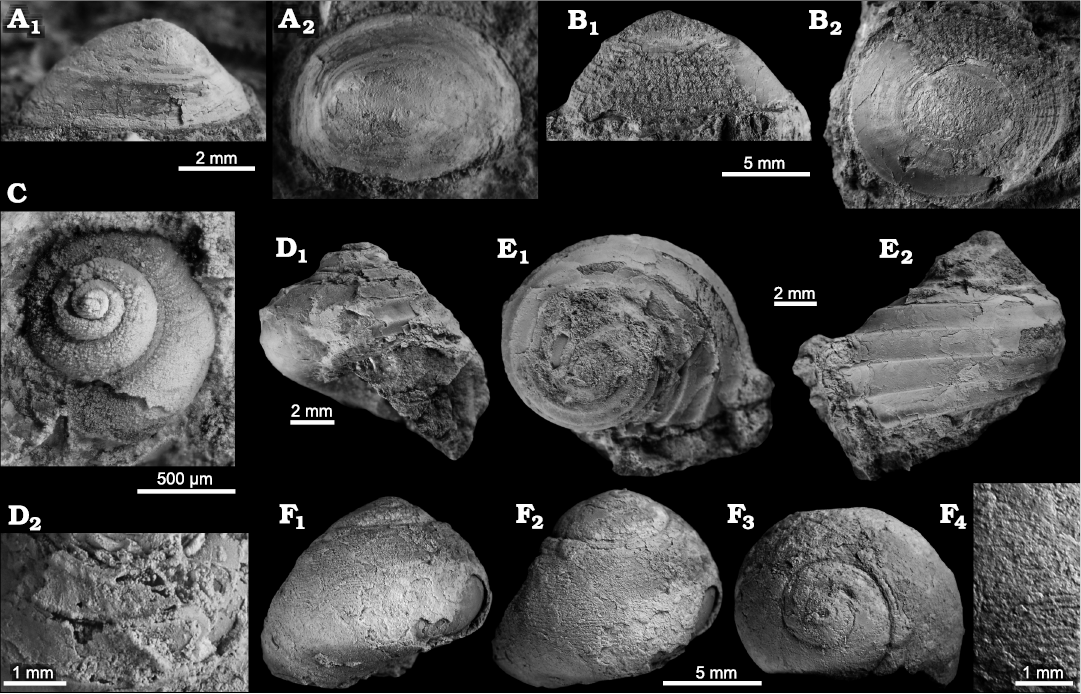
Fig. 18. Limpets, neomphalids, and vetigastropods from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. A, B. Unidentified limpet gastropod. A. NRM Mo 192715, small specimen, mostly an internal mold. B. NRM Mo 192656, large specimen with sculpture. C. The neomphalin Retiskenea sp., NRM Mo 192540, apical view on specimen embedded in rock matrix. D, E. The seguenzoid Cataegis sp. D. NRM Mo 192666; note sculpture on D2. E. NRM Mo 192665, specimen showing the strong spiral ribs, in apical (E1) and abapertual (E2) views. F. The colloniid Homalopoma sp., NRM Mo 192583, views on entire shell (F1–F3) and close-up on spiral sculpture (F4).
Remarks.—Due to the poor preservation of the available specimens, we are unable to assign Cataegis sp. to any existing or new species. Apart from being slightly lower spired, Cataegis sp. is similar to the extant Cataegis celebesensis McLean and Quinn, 1987, from Indonesia, which also has nearly evenly spaced spiral ribs; other species of Cataegis tend to have the keels more closely spaced on the whorl’s shoulder and more widely spaced on the flanks (McLean and Quinn 1987; Vilvens 2016). This applies also to Cataegis ramosi Kiel, Aguilar, and Kase, 2020a, from late Pliocene-aged Liog-Liog Point seep deposit in Leyte, Philippines, which has a more distinctive shoulder than Cataegis sp. reported here. The Late Miocene Cataegis godineauensis (Van Winkle, 1919) from seep deposits in Trinidad (see Kiel and Hansen 2015: fig. 14E) has very similar sculpture but a lower spire than Cataegis sp.
The related cataegid Kanoia myronfeinbergi Warén and Rouse, 2016, has a much more angular whorl profile than the specimens reported here (Warén and Rouse 2016). Members of the genus Carinotrochus Zhang, Zhang, and Zhang, 2020a, from seamounts in the tropical western Pacific (Zhang et al. 2020a) are superficially similar to Cataegis sp. but have nearly smooth spiral ridges lacking the nodular sculpture of Cataegis.
Order Trochida
Family Colloniidae Cossmann in Cossmann and Peyrot, 1917
Genus Homalopoma Carpenter, 1864
Type species: Turbo sanguineus Linnaeus, 1758, by monotypy; Recent, Mediterranean Sea.
Homalopoma? sp.
Fig. 18F.
Material.—One specimen from block B (NRM Mo 192583: W = 11.8 mm, H = 9.6 mm).
Remarks.—Homalopoma has been reported from fossil seeps of Cretaceous age onwards. The specimen reported here has much finer and more numerous spiral cords compared to both Homalopoma abeshinaiensis Kaim, Jenkins, and Hikida, 2009, from the Upper Cretaceous of Hokkaido, Japan (Hikida et al. 2003; Kaim et al. 2009), and Homalopoma domeniconii Moroni, 1966, from Middle–Late Miocene seep deposits in Italy (Moroni 1966). Species with similar spiral ornament but taller shells and a more convex whorl profile include the middle to late Eocene Homalopoma? sp. from the Humptulips seep deposit in Washington state, USA (Goedert and Squires 1990), Homalopoma wattsi (Dickerson, 1916) from a middle Eocene wood fall in Washington (Kiel 2008), Homalopoma sp. from fossil seeps of somewhat uncertain Cenozoic age in Barbados (Gill et al. 2005), and the extant Homalopoma bicolor Okutani, 2001, from the northern Mariana Ridge (Okutani 2001). Homalopoma tosaense Habe, 1953, reported from the Plio-Pleistocene of Luzon Island, Philippines (Helwerda et al. 2014), is smaller, taller, and has very distinct spiral keels rather than fine spiral cords has the specimen reported here from the Northern Cement quarry.
Of very similar overall shape are species of Phanerolepida, including Phanerolepida rehderi MacNeil, 1960, from the Plio-Pleistocene of Okinawa, southern Japan (Noda 1988: pl. 5: 3) but the Philippine specimen illustrated here clearly has spiral sculpture instead of the net-like rhombohedral pattern of Phanerolepida (Hickman 1972).
Subclass Neritimorpha Golikov and Starobogatov, 1975
Family Phenacolepadidae Pilsbry, 1895
Genus Thalassonerita Moroni, 1966
Type species: Nerita (Thalassonerita) megastoma Moroni, 1966, by original designation; Miocene, Italy.
Thalassonerita hagai sp. nov.
Fig. 19.
Zoobank LCID: urn:lsid:zoobank.org:act: 5A82FE2C-D3D5-41FD-B011-9AD5924DE4B9
Etymology: For Takuma Haga (Tsukuba, Japan), who collected the specimens.
Type material: Holotype: NIGSPAL-032, specimen with some shell, aperture obscured by rock matrix. Paratypes: NRM Mo 192657, 192658, partially preserved specimens from the type locality and horizon.
Type locality: The “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Type horizon: Seep carbonate blocks enclosed in the Amlang Formation (Upper Miocene).
Material.—Type material only.
Dimensions (in mm).—NIGSPAL-032: W = 25.0, H = 17.3.
Diagnosis.—Shell large for genus, two whorls, sculpture of 13 widely spaced axial ribs on last whorl.
Description.—Large neritiform shell, up to 25.0 mm wide, at least two whorls, spire apparently flat; sculpture of 13 oblique axial ribs, interspaces between ribs 2–3 times the width of ribs, both ribs and interspaces covered by fine axial lirae.

Fig. 19. The phenacolepadid gastropod Thalassonerita hagai sp. nov., from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. NIGSPAL-032, holotype, in abapertural view (A1) and apical view (A2).
Remarks.—The specimens have a thin outer layer that appears to differ in mineralogy from the remaining shell material, and such a layer has not been observed in other gastropods from the Northern Cement Corporation quarry. It is here interpreted as the calcitic outer layer typical of neritimorph gastropods. This is the first fossil record of Thalassonerita from the central Indo-West Pacific and with a width of 25.0 mm it is by far the largest Thalassonerita reported to date. Both the middle Eocene Thalassonerita eocenica Squires and Goedert, 1996, from the Humptulips Formation in western Washington state, USA (Squires and Goedert 1996; Hybertsen and Kiel 2018), and the type species Thalassonerita megastoma Moroni, 1966, from the Upper Miocene of Italy, also reported from the Miocene of Barbados, reach only 10.0 mm in width (Moroni 1966; Gill et al. 2005). An early Oligocene neritid reported from seep deposits in Peru reaches 8.5 mm in width (Kiel et al. 2020b), and the extant Thalassonerita naticoides (Clarke, 1989) has a maximum width of 12.5 mm (Clarke 1989). Furthermore, all above mentioned species have less distinctive axial ornament than Thalassonerita hagai; the extant T. naticoides is nearly smooth. An Early Cretaceous neritid reported from a seep deposit on the Crimean Peninsula reaches just over 20.0 mm in width, but is nearly smooth and might not have any direct phylogenetic relationships to Thalassonerita (Kiel and Peckmann 2008).
Stratigraphic and geographic range.—Upper Miocene of Luzon, Philippines.
Subclass Caenogastropoda Cox, 1960
Family Naticidae Guilding, 1834
Naticidae gen. et sp. indet.
Fig. 20A.
Material.—One specimen from block B (NRM Mo 192628: H = 21.3 mm, W = 21.2 mm). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Remarks.—Unfortunately, the umbilical area of this specimen is damaged, preventing a generic assignment.
Family Fasciolariidae Gray, 1853
Genus Fusinus Rafinesque, 1815
Type species: Murex colus Linnaeus, 1758, by typification of replaced name; Recent, Indo-West Pacific Ocean.
Fusinus? sp.
Fig. 20B, C.
Material.—Two large specimens from block I (NRM Mo 192662, 192663). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Dimensions (in mm).—NRM Mo 192662: H = 73.2, W = 52.0; NRM Mo 192663: H = 90.8, W = 45.0.
Remarks.—Fusinus is widespread in the Indian and Pacific Oceans today (Okutani and Tsuchiya 2000; Callomon and Snyder 2007, 2019; Couto et al. 2016), but its fossil record from this area is meager at best. Although the preservation of the two Fusinus? sp. specimens is not ideal, they clearly lack the granulate sculpture typical for Granulifusus, a genus common in the Indo-West Pacific and known from several fossils, especially in Japan (Shuto 1958; Hadorn and Fraussen 2005). With their evenly convex whorls, the two Fusinus? sp. specimens are clearly different from those of several deep-water fusinins inhabiting deeper water around the Philippines today, such as species of Latirus Montfort, 1810, and Fusolatirus Kuroda and Habe in Kuroda et al., 1971 (Garcia 2001; Snyder and Bouchet 2006).
Vermeij and Snyder (2018) reviewed large fasciolariid genera (Fusininae) and introduced several genera for tall species with strongly convex whorls and distinct axial ribs resembling Fusinus? sp. reported here. Among these, Apertifusus includes three or four tropical Atlantic species, and Lyonsifusus consists of a few species from the southern Caribbean, including Miocene fossils. Lastly, Ariefusus was introduced for an extant West African species and for the Miocene Fusus prevosti Partsch in Hörnes, 1856, whose whorls are more constricted near the suture (Hörnes 1856) and consequently have somewhat pointed ribs on the center of the whorls, unlike the Philippine specimens.
Family Buccinidae Rafinesque, 1815
Genus Enigmaticolus Fraussen, 2008
Type species: Enigmaticolus monnieri Fraussen, 2008, by original designation; Recent, Madagascar.
Remarks.—Six species and two unnamed records of this genus are known from the Indian and western Pacific Oceans (Fraussen 2008; Fraussen and Stahlschmidt 2016; Zhang et al. 2020b; Chen et al. 2021). Here we transfer a species formerly reported as Siphonalia semisulcata Martin, 1933, and Siphonalia (Pseudoneptunea) semisulcata, from the Upper Miocene of Buton, Indonesia, to Enigmaticolus, report it also from the Upper Miocene of the Philippines, and regard it as the first fossil record of this genus.
Enigmaticolus semisulcata (Martin, 1933) comb. nov.
Fig. 20D–F.
1933 Siphonalia semisulcata sp. nov.; Martin 1933: 25, pl. 3: 24, 25.
1942 Siphonalia (Pseudoneptunea) semisulcata Martin; Beets 1942: 281, pl. 28: 38.
Description.—High-spired fusiform shell, whorls gently convex with fine constriction below suture; sculpture of flat spirals crossed by numerous fine axial (slightly opisthocyrt) growth increments, giving the spirals a beaded appearance; sculpture most prominent on early whorls and base; inner lip with thin callus.

Fig. 20. Caenogastropoda and Opisthobranchia from Upper Miocene seep deposits of the Amlang Formation in Pangasinan province, Luzon, Philippines. A. Naticidae gen. et sp. indet., NRM Mo 192628, imperfect specimen in apical and apertural views. B, C. Very large Fusinus? sp., imperfect specimens in abapertural view. B. NRM Mo 192662. C. NRM Mo 192663. D–F. Enigmaticolus semisulcata (Martin, 1933), imperfect specimens in apertural and abapertural views. D. NRM Mo 192632. E. NRM Mo 192586. F. NRM Mo 192633. G. Enigmaticolus sp., NRM Mo 192640, specimen with broken apertural base, abapertural and apertural view. H. Mitridae gen. et sp. indet., NRM Mo 192639, imperfect specimen, abapertural view. I. Terebridae gen. et sp. indet., NRM Mo 192627, fragment of spire, lateral view. J, K. Acanthodaphne? sp., imperfect specimens in apertural view. J. NRM Mo 192637. K. NRM Mo 192638. L. Gemmula sp., NRM Mo 192417, imperfect specimen, apertural view. M. Xylodiscula? sp., NRM Mo 192619, specimen embedded in rock matrix in apical view. N. Volvulella aff. minoensis Itoigawa, 1958, NRM Mo 192641, nearly complete specimen, apertural view. O. Ringiculidae gen. et sp. indet., NRM Mo 192642, imperfect specimen, apertural view.
Material.—Two specimens from block B (NRM Mo 192586, 192587), three specimens from block I (NRM Mo 192631–192633). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Dimensions (in mm).—NRM Mo 192586: H = 30.0, W = 21.5; NRM Mo 192586: H = 17.7, W = 11.8; NRM Mo 192632: H = 36.8, W = 24.0; NRM Mo 192632: H = 34.0, H = 21.0.
Remarks.—The specimens illustrated by Martin (1933) from the Upper Miocene of Buton, Indonesia, resemble the Philippine specimens reported here in general shape and in having flat, beaded spiral ornament that becomes more prominent on the base. They differ by having more distinctive axial ribbing in the upper part of the whorls, especially on the early whorls (Martin 1933), whereas the Philippine specimens show more distinctive beaded spiral ornament. Even more similar to the Philippine specimens is another specimen from Buton, referred to as ”Siphonalia (Pseudoneptunea) semisulcata Martin” by Beets (1942), due to its lack of the strong axial ribs on the upper part of the whorls.
Species of Siphonalia and Pseudoneptunea have more convex whorls with much stronger axial ornament than “Siphonalia semisulcata Martin, 1933”; hence the species is unlikely to belong to either of these genera (Makiyama 1941; Cernohorsky 1975; Robba et al. 2004; Zhang and Zhang 2018). Rather, the habitus of the shell, the flat spiral ornament on the upper part of the whorls and the base, the indistinct axial ribs, and fine constriction below the suture, indicate affinities to Enigmaticolus, to which this species is herein re-assigned. Among the extant species of Enigmaticolus, the most similar to E. semisulcata is the type species with its slender shell.
Stratigraphic and geographic range.—Upper Miocene of Luzon, Philippines and Buton, Indonesia.
Enigmaticolus sp.
Fig. 20G.
Material.—One specimen from block I (NRM Mo 192640: H = 23.8 mm, W = 12.0 mm). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Remarks.—This specimen differs from Enigmaticolus semisulcata by being smaller and by having more distinct axial ribs on the whorl’s shoulder. In the latter respect it resembles Enigmaticolus nipponensis (Okutani and Fujiwara, 2000) from methane seeps in the South China Sea (Chen et al. 2021).
Family Mitridae Swainson, 1831
Mitridae gen. et sp. indet.
Fig. 20H.
Material.—One specimen from block I (NRM Mo 192639: H = 17.0 mm, W = 9.0 mm). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Remarks.—This poorly preserved specimen consists of 2.5 whorls, has straight axial ribs, fine spirals, and two columellar folds. It certainly belongs to the Mitridae, but a genus- or species-level identification is not possible.
Family Terebridae Mörch, 1852
Terebridae gen. et sp. indet.
Fig. 20I.
Material.—One specimen from block I (NRM Mo 192627: H = 36.6 mm, W = 8.0 mm). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Remarks.—A similar species is Gemmaterebra bicorona (Hutton, 1885) as illustrated in Beu and Maxwell (1990: 269, pl. 30: O) from the Middle–Upper Miocene of New Zealand.
Family Raphitomidae Bellardi, 1875
Genus Acanthodaphne Bonfitto and Morassi, 2006
Type species: Acanthodaphne sabellii Bonfitto and Morassi, 2006, by original designation; Recent, Gulf of Aden.
Remarks.—This genus of deep-water raphitomids is known from a few extant species from the Indian and West Pacific Oceans; its fossil record is considered to include Puha pusula Laws, 1947, from the Lower Miocene of New Zealand, and possibly Pseudoinquisitor? cf. P.? pulchra (Schepman, 1913) reported by MacNeil (1960) from the Middle Miocene of Okinawa, Japan (Bonfitto and Morassi 2006; Morassi and Bonfitto 2010).
Acanthodaphne? sp.
Fig. 20J, K.
Material.—Three specimens from block I (NRM Mo 192636–192638). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Dimensions (in mm).—NRM Mo 192636: H = 17.0, W = 9.8; NRM Mo 192637: H = 16.0, W = 12.8; NRM Mo 192638: H = 22.8, W = 12.0.
Remarks.—These three specimens have the general shell outline and ornament of species of Acathodaphne, but with its protoconch morphology unknown, their assignment to this genus remain somewhat uncertain. The Miocene Pseudoinquisitor? cf. P.? pulchra from the Yunabaru clay on Okinawa (MacNeil 1960: pl. 5: 13) has fewer but more distinct spirals on the lower part of the whorls.
Family Turridae Adams and Adams, 1853
Genus Gemmula Weinkauff, 1875
Type species: Gemmula hindsiana Berry, 1958, by subsequent designation; Recent, eastern Pacific Ocean.
Gemmula sp.
Fig. 20L.
Material.—One specimen from block E (NRM Mo 192417: H = 24.5 mm, W = 14.0 mm). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Remarks.—This fragmentary specimen (3.5 whorls) is quite similar to Gemmula sp. indet. illustrated by Matsubara (2011: pl. 3: 7) from Miocene shallow marine deposits of the Hokutan Group in the Tajima area, Hyôgo Prefecture, southwest Japan. Gemmula osawanoensis Tsuda, 1959, from Middle Miocene deep-water sediments of the Higashibessho Formation in Toyama Prefecture, central Japan (Amano et al. 2004: fig. 5.15), appears to have more distinctive spiral ornament on the base, though the absence of such ornament from the Philippine specimen might be a preservational artifact. The Gemmula species from the Miocene Yunabaru clay on Okinawa illustrated by MacNeil (1960: pl. 5) have fewer but stronger axial ribs on the central spiral cord than the specimens reported here. Further similar fossil species include Gemmula granosus (Helbling, 1779) illustrated in Noda (1988: pl. 13: 9a, b) from the Pliocene of Okinawa, Japan, and Pleurotoma woodwardi Martin, 1883 from Java, Indonesia (Martin 1891–1906: 37, pl. 4: 91–96). A bit more compact are Bathytoma hetzeli Martin, 1933, from the Upper Miocene of Buton, Indonesia (Martin 1933: 21, pl. 3: 16), and Cosmasyrinx monilifera Marwick, 1931 from Miocene bathyal deposits in New Zealand (Beu and Maxwell 1990: 250, pl. 27: S). Among extant taxa, perhaps most similar are Gemmula cosmoi (Sykes, 1830), which occurs from Sagami Bay to Kyushu, in 50–350 m depth, and G. kieneri (Doumet, 1840), which occurs from Choshi (Pacific central Honshu) southward to the Philippines (Hasegawa et al. 2000).
Subclass Heterobranchia Burmeister, 1837
Family Xylodisculidae Warén, 1992
Genus Xylodiscula Marshall, 1988
Type species: Xylodiscula vitrea Marshall, 1988, by original designation; Recent, SE Australia.
Xylodiscula? sp.
Fig. 20M.
Material.—Four specimens from block B (NRM Mo 192619–192622), one specimen from block I (NRM Mo 192626). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Remarks.—These small (up to 2.0 mm diameter), poorly preserved, low-spired shells most closely resemble species of the heterobranch Xylodiscula, but the imperfectly preserved protoconch prevents a more certain assignment. Xylodiscula occurs on sunken wood, both recent (Marshall 1988) and fossil (Kiel and Goedert 2007), but is also known from hydrothermal vents (Warén and Bouchet 1993) and other reducing environments (Høisæter and Johannessen 2001; Hawe et al. 2014).
Family Rhizoridae Dell, 1956
Genus Volvulella Newton, 1891
Type species: Bulla acuminata Bruguière, 1792, by typification of replaced name; Recent, European waters.
Volvulella aff. minoensis Itoigawa, 1958
Fig. 20N.
Material.—One specimen from block I (NRM Mo 192641: H = 12.8 mm, W = 7.4 mm). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Remarks.—The pointed apex characteristic for Volvulella is eroded away in the only available specimen, but its former presence is indicated by the blunt apical peg and the apically broken aperture. The type specimens of V. minoensis illustrated by Itoigawa (1958: 178, pl. 26: 8, 9) from the Middle Miocene Mizunami Group in south-central Honshu, are reported to have fine incisions at both ends; these, too, may have been eroded away in NRM Mo 192641. The other Miocene Japanese Volvulella species reported by Itoigawa (1958) are more slender than V. minoensis.
Similar in shape, size and having a smooth shell is the extant Volvulella ovulina (Adams in Sowerby, 1850) which occurs in 10–477 m depth, from Japan to southeastern Indonesia (Valdés 2008). Slightly more globular and with fine incisions near the base is Volvulella volvulaeformis (Seguenza, 1879) from the Italian Pliocene (Brunetti 2020), whereas the Early Miocene V. staphylemorpha Harzhauser, 2014, from shallow-marine deposits in Kerala, SW India, has a more slender shell (Harzhauser 2014).
Family Ringiculidae Philippi, 1853
Ringiculidae gen. et sp. indet.
Fig. 20O.
Material.—One specimen from block I (NRM Mo 192642: H = 6.2 mm, W = 5.2 mm). Upper Miocene of the “shale quarry” within the Northern Cement Corporation quarry in Pangasinan province, Luzon, Philippines.
Remarks.—This small globular shell with about two whorls preserved shows 17 fine spiral incisions on the last whorl and three folds on the parietal and columellar lip, suggesting affinities to Ringicula. Helwerda (2015) reported numerous species of Ringicula from Plio-Pleistocene strata in the Philippines, but the poor preservation of the present specimen does not allow a more detailed comparison.
Acknowledgements
We would like to thank Kazutaka Amano (Joetsu University of Education, Japan), Alan G. Beu (GNS Science, Lower Hutt, New Zealand), David Jablonski (University of Chicago, USA), P. Graham Oliver (National Museum of Wales, Cardiff, UK), and John D. Taylor (British Museum of Natural History, London, UK) for insights and discussions of the species; Takuma Haga (National Museum of Nature and Science, Tsukuba, Japan), Wenceslao Mago (Mines and Geosciences Bureau of the Philippines, Quezon City, Philippines), Ryuichi Majima (Yokohama National University, Japan), Kerve M. Supnet, David C. Policarpio, and Renato Bautista (all University of the Philippines, Quezon City, Philippines) for help with collecting, preparing, and identifying fossils; the Northern Cement Corporation, especially Alfredo D. Ballesteros, Jr., Myra Vanessa A. Frijellana, and Oliver R. Gorrospe, who provided access to the quarry, and all the staff who served as guides during the fieldwork. We also thank the two reviewers Kazutaka Amano and Marco Taviani (ISMAR-CNR, Bologna, Italy) for their critical comments on the manuscript. Financial support was provided by the Swedish Research Council (Vetenskapsrådet) through grant 2016-03920 to SK.
References
Abbott, R.T. 1974. American Seashells, the Marine Mollusca of the Atlantic and Pacific Coast of North America. Second edition. 663 pp. Van Nostrand Reinhold, New York.
Adams, H. and Adams, A. 1853–1858. The Genera of Recent Mollusca Arranged According to Their Organization. Vol. 1: vi–xl + 484 pp.; Vol. 2: 661 pp. John Van Voorst, London. Crossref
Amano, K. 2003. Predatory gastropod drill holes in Upper Miocene cold seep bivalves, Hokkaido, Japan. The Veliger 46: 90–96.
Amano, K. and Ando, H. 2011. Giant fossil Acharax (Bivalvia: Solemyidae) from the Miocene of Japan. The Nautilus 125: 207–211.
Amano, K. and Jenkins, R.G. 2007. Eocene drill holes in cold-seep bivalves of Hokkaido, northern Japan. Marine Ecology 28: 108–114. Crossref
Amano, K. and Jenkins, R.G. 2011. New fossil Bathymodiolus (s. l.) (Mytilidae, Bivalvia) from Oligocene seep-carbonates in eastern Hokkaido, Japan, with remarks on the evolution of Bathymodiolus (s. l.). The Nautilus 125: 29–35.
Amano, K. and Kiel, S. 2010. Taxonomy and distribution of fossil Archivesica (Vesicomyidae, Bivalvia) in Japan. The Nautilus 124: 155–165.
Amano, K. and Kiel, S. 2012. Two Neogene vesicomyid species (Bivalvia) from Japan and their biogeographic implications. The Nautilus 126: 79–85.
Amano, K., Hamuro, T., and Hamuro, M. 2004. Latest early-earliest middle Miocene deep-sea molluscs in the Japan Sea borderland—the warm-water Higashibessho fauna in Toyama Prefecture, central Japan. Paleontological Research 8: 29–42. Crossref
Amano, K., Jenkins, R.G., Aikawa, M., and Nobuhara, T. 2010. A Miocene chemosynthetic community from the Ogaya Formation in Joetsu: evidence for depth-related ecologic control among fossil seep communities in the Japan Sea back-arc basin. Palaeogeography, Palaeoclimatology, Palaeoecology 286: 164–170. Crossref
Amano, K., Little, C.T.S., and Campbell, K.A. 2018. Lucinid bivalves from Miocene hydrocarbon seep sites of eastern North Island, New Zealand, with comments on Miocene New Zealand seep faunas. Acta Palaeontologica Polonica 63: 371–382. Crossref
Amano, K., Miyajima, Y., Jenkins, R.G., and Kiel, S. 2019a. The Miocene to Recent biogeographic history of vesicomyid bivalves in Japan, with two new records of the family. The Nautilus 133: 48–56.
Amano, K., Miyajima, Y., Nakagawa, K., Hamuro, M., and Hamuro, T. 2019b. Chemosymbiotic bivalves from the lower Miocene Kurosedani Formation in Toyama Prefecture, central Honshu, Japan. Paleontological Research 23: 208–219. Crossref
Aoki, S. 1954. Mollusca from the Miocene Kabeya Formation, Joban coal-field, Fukushima Prefecture, Japan. Science Reports of the Tokyo Kyoiku Daigaku, Section C 17: 23–40.
Audzijonyte, A., Krylova, E.M., Sahling, H., and Vrijenhoek, R.C. 2012. Molecular taxonomy reveals broad trans-oceanic distributions and high species diversity of deep-sea clams (Bivalvia: Vesicomyidae: Pliocardiinae) in chemosynthetic environments. Systematics and Biodiversity 10: 403–415. Crossref
Aurelio, M.A., Peña, R.E., and Taguibao, K.J.L. 2013. Sculpting the Philippine archipelago since the Cretaceous through rifting, oceanic spreading, subduction, obduction, collision and strike-slip faulting: Contribution to IGMA5000. Journal of Asian Earth Sciences 72: 102–107. Crossref
Beets, C. 1942. Beiträge zur Kenntnis der angeblich oberoligocänen Mollusken-Fauna der Insel Buton, Niederländisch-Ostindien. Leidsche Geologische Mededeelingen 13: 255–328.
Beets, C. 1953. Reconsideration of the so-called Oligocene fauna in the asphaltic deposits of Buton (Malay Archipelago). Leidse Geologische Mededelingen 17: 237–258.
Bellardi, L. 1875. Monografia delle nuculidi trovate finora nei terreni terziari del Piemonte e della Liguria. 32 pp. Il R. Liceo Gioberi nell’Anno Scolastico 1874–1875, Torino. Crossref
Berry, S.S. 1958. Notices of new eastern Pacific Mollusca. II. Leaflets in Malacology 1: 83–90.
Beu, A.G. and Maxwell, P.A. 1990. Cenozoic Mollusca of New Zealand. New Zealand Geological Survey Paleontological Bulletin 58: 1–518.
Blow, W.H. 1969. Late Middle Eocene to Recent planktonic foraminiferal biostratigraphy. In: P. Bronnimann, and H.H. Renz (eds.), Proceedings of the 1st International Conference on Planktonic Microfossils, Geneva, Vol. 1, 199–422. Geneva.
Bonfitto, A. and Morassi, M. 2006. A new genus of Indo-West Pacific Turridae (Gastropoda: Prosobranchia). The Veliger 48: 136–142.
Borson, S. 1825. Saggio di orittografia piemontese. Memorie della Reale Accademia delle Scienze di Torino 29: 251–318.
Bouchet, P. and Cosel, R. von 2004. The world’s largest lucinid is an undescribed species from Taiwan (Mollusca: Bivalvia). Zoological Studies 43: 704–711.
Bronn, H.G. 1831. Italiens Tertiär-Gebilde und deren organische Einschlüsse. 176 pp. Groos, Heidelberg. Crossref
Brugnone, G. 1880. Le conchiglie plioceniche selle vicinaze di Caltanisetta. Bolletino Societá Malacologia Italiana 6: 85–158.
Bruguière, J.G. 1789–1816. Encyclopédic méthodique ou par ordre de matières. Histoire naturelle de Vers, des Mollusques. 758 pp. Panckouke, Paris. Crossref
Brunetti, M.M. 2020. Rediscovery of Volvulella volvulaeformis (G. Seguenza, 1879) in the Italian Pliocene (Gastropoda Rhizoridae). Biodiversity Journal 11: 435–437. Crossref
Burmeister, H. 1837. Handbuch der Naturgeschichte, Vol. 2, Zoologie. i–xii + 369–858. Enslin, Berlin.
Callomon, P. and Snyder, M.A. 2007. On the genus Fusinus in Japan III: Nine further species, with type selections. Venus 66: 19–47.
Callomon, P. and Snyder, M.A. 2019. The genus Fusinus in the northwestern Pacific. Special Publication of the Malacological Society of Japan 4: 1–122.
Campbell, K.A. 2006. Hydrocarbon seep and hydrothermal vent paleoenvironments and paleontology: Past developments and future research directions. Palaeogeography, Palaeoclimatology, Palaeoecology 232: 362–407. Crossref
Campbell, K.A. and Bottjer, D.J. 1995. Brachiopods and chemosymbiotic bivalves in Phanerozoic hydrothermal vent and cold seep environments. Geology 23: 321–324. Crossref
Carpenter, P.P. 1864. Supplementary report on the present state of our knowledge with regard to the Mollusca of the West Coast of North America. Reports of the British Association to the Advancement of Science 33: 517–686.
Cernohorsky, W.O. 1975. The taxonomy of some Indo-Pacific Mollusca. Part 3. With descriptions of new taxa and remarks on an Ecuadorian fossil species of Turridae. Records of the Auckland Institute and Museum 12: 213–234.
Chen, C., Watanabe, H.K., Nagai, Y., Toyofuku, T., Xu, T., Sun, J., Qiu, J.-W., and Sasaki, T. 2019. Complex factors shape phenotypic variation in deep-sea limpets. Biology Letters 15: 20190504. Crossref
Chen, C., Xu, T., Fraussen, K., and Qiu, J.-W. 2021. Integrative taxonomy of enigmatic deep-sea true whelks in the sister-genera Enigmaticolus and Thermosipho (Gastropoda: Buccinidae). Zoological Journal of the Linnean Society 193: 230–240. Crossref
Clarke, A.H. 1989. New mollusks from undersea oil seep sites off Louisiana. Malacology Data Net 2: 122–134.
Coan, E.V., Scott, P.V., and Bernard, F.R. 2000. Bivalve Seashells of Western North America. viii + 764 pp. Santa Barbara Museum of Natural History, Santa Barbara.
Conrad, R.A. 1849. Fossils from the northwestern America. In: J.D. Dana (ed.), U.S. Exploration Expedition, 1838–1842, under Charles Wilkes. Geology, Volume 10, 723–728 (appendix). C. Sherman, Philadelphia.
Cosel, R. von and Bouchet, P. 2008. Tropical deep-water lucinids (Mollusca: Bivalvia) from the Indo-Pacific: essentially unknown, but diverse and occasionally gigantic. Mémoires du Muséum National d’Histoire Naturelle 196: 115–213.
Cosel, R. von and Marshall, B.A. 2003. Two new species of large mussels (Bivalvia: Mytilidae) from active submarine volcanoes and a cold seep off the eastern North Island of New Zealand, with description of a new genus. The Nautilus 117: 31–46.
Cosel, R. von and Olu, K. 2009. Large Vesicomyidae (Mollusca: Bivalvia) from cold seeps in the Gulf of Guinea off the coasts of Gabon, Congo and northern Angola. Deep-Sea Research II 56: 2350–2379. Crossref
Cossmann, M. and Peyrot, A. 1917. Conchologie néogénique de l’Aquitaine. Tome 3, Gastropodes, Scaphopodes et Amphineures. Actes de la Société linnéenne de Bordeaux 69: 285–365.
Costa, O.G. 1830. Catalogo sistematico e ragionato de’ testacei delle Due Sicilie. 1–8 + i–cxxxii pp. Tipografia della Minerva, Napoli. Crossref
Couto, D.R., Bouchet, P., Kantor, Y.I., Simone, L.R.L., and Giribet, G. 2016. A multilocus molecular phylogeny of Fasciolariidae (Neogastropoda: Buccinoidea). Molecular Phylogenetics and Evolution 99: 309–322. Crossref
Cox, L.R. 1960. Gastropoda. General characteristics of Gastropoda. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology. Part I. Mollusca, 1, I85–I169. Geological Society of America and University of Kansas Press, Lawrence.
Cuvier, G.L.C.F.D. 1795. Second Mémoire sur l’organisation et les rapports des animaux à sang blanc, dans lequel on traite de la structure des Mollusques et de leur division en ordre, lu à la société d’Histoire Naturelle de Paris, le 11 prairial an troisième. Magazin Encyclopédique, ou Journal des Sciences, des Lettres et des Arts 2: 433–449.
Dall, W.H. 1891. On some new or interesting west American shells obtained from the dredgings of the U.S. Fish Commission steamer Albatross in 1888, and from other sources. U.S. National Museum Proceedings 14: 173–191. Crossref
Dall, W.H. 1896. Diagnoses of new species of mollusks from the west coast of America. Proceedings of the U.S. National Museum of Natural History 18: 7–20. Crossref
Dall, W.H. 1901. Synopsis of the Lucinacea and of the American species. Proceedings of the U.S. National Museum of Natural History 23: 779–833. Crossref
Dall, W.H. 1903. Contributions to the Tertiary fauna of Florida, with especial reference to the silex beds of Tampa and the Pliocene beds of the Caloosahatchie River, including in many cases a complete revision of the generic groups treated of and their American species. Part VI. Concluding the work. Wagner Free Institute of Science of Philadelphia, Transactions 3: xiv + 1219–1654. Crossref
Dall, W.H. 1908a. A gigantic Solemya and a new Vesicomya. The Nautilus 22: 61–63.
Dall, W.H. 1908b. A revision of the Solenomyacidae. The Nautilus 22: 1–2.
Dall, W.H. and Simpson, C.T. 1901. The Mollusca of Porto Rico. United States Fishery Commission, Bulletin 20: 351–524.
David, S.J., Stephan, J.F., Delteil, J., Muller, C., Butterlin, J., Bellon, H., and Billedo, E. 1997. Geology and tectonic history of southeastern Luzon, Philippines. Journal of Asian Earth Sciences 15: 435–452. Crossref
Dell, R.K. 1956. The archibenthal Mollusca of New Zealand. Dominion Museum Bulletin 18: 1–235.
Des Moulins, C. 1868. Descriptions et figures de quelques coquilles fossiles du terrain tertiaire et de la craie (Gironde, Dordogne, Royan). Actes de la Société linnéenne de Bordeaux 26: 357–379.
Dickerson, R.E. 1916. Stratigraphy and fauna of the Tejon Eocene of California. University of California Publications in Geological Sciences 9: 363–524.
Distel, D.L., Baco, A.R., Chuang, E., Morrill, W., Cavanaugh, C.M., and Smith, C.R. 2000. Do mussels take wooden steps to deep-sea vents? Nature 403: 725–726. Crossref
Doumet, E. 1840. Pleurotoma in Mollusques. Magasin de Zoologie Series 2 year II: pl. 10 + 2 pp.; pl. 11 + 2 pp.
Fisher, C.R. 1995. Toward an appreciation of hydrothermal-vent animals: their environment, physiological ecology, and tissue stable isotope values. In: S.E. Humphris, R.A. Zierenberg, L.S. Mullineaux, and R.E. Thomson (eds.), Seafloor Hydrothermal Systems: Physical, Chemical, Biological, and Geochemical Interactions (Geophysical Monograph Series), 297–316. Blackwell, Washington, DC. Crossref
Fleming, J. 1828. A History of British Animals, Exhibiting the Descriptive Characters and Systematical Arrangement of the Genera and Species of Quadrupeds, Birds, Reptiles, Fishes, Mollusca and Radiata of the United Kingdom; Including the Indigenous, Extirpated, and Extinct Kinds; Together with Periodical and Occasional Visitants. xxiii + 554 pp. Bell & Bradfute, Edinburgh. Crossref
Fraussen, K. 2008. Enigmaticolus, a new genus of deep water buccinids (Gastropoda: Buccinidae), with description of a new species from Madagascar. Gloria Maris 46: 74–82.
Fraussen, K. and Stahlschmidt, P. 2016. The extensive Indo-Pacific deep-water radiation of Manaria E.A. Smith, 1906 (Gastropoda: Buccinidae) and related genera, with descriptions of 21 new species. Mémoires du Muséum national d’Histoire naturelle 208: 363–456.
Gabb, W.M. 1866–1869. Cretaceous and Tertiary fossils. California Geological Survey, Paleontology 2: 1–299.
Galgana, G., Hamburger, M., McCaffrey, R., Corpuz, E., and Chen, Q. 2007. Analysis of crustal deformation in Luzon, Philippines using geodetic observations and earthquake focal mechanisms. Tectonophysics 432: 63–87. Crossref
Garcia, E.F. 2001. A new species of Latirus (Gastropoda: Fasciolariidae) from the southern Philippines. Novapex 2: 149–151.
Gill, F.L., Harding, I.C., Little, C.T.S., and Todd, J.A. 2005. Palaeogene and Neogene cold seep communities in Barbados, Trinidad and Venezuela: An overview. Palaeogeography, Palaeoclimatology, Palaeoecology 227: 191–209. Crossref
Glover, E.A. and Taylor, J.D. 1997. New species and records of Rastafaria and Megaxinus (Bivalvia: Lucinidae) from the western Indian Ocean and Red Sea, with a reappraisal of Megaxinus. Journal of Conchology 36: 1–18.
Glover, E.A. and Taylor, J.D. 2016. Lucinidae of the Philippines: highest known diversity and ubiquity of chemosymbiotic bivalves from intertidal to bathyal depths (Mollusca: Bivalvia: Lucinidae). Mémoires du Muséum National d’Histoire Naturelle 208: 65–234.
Goedert, J.L. and Squires, R.L. 1990. Eocene deep-sea communities in localized limestones formed by subduction-related methane seeps, southwestern Washington. Geology 18: 1182–1185. Crossref
Golikov, A.N. and Starobogatov, Y.I. 1975. Systematics of prosobranch gastropods. Malacologia 15: 185–232.
Grant, U.S.I. and Gale, H.R. 1931. Catalogue of the marine Pliocene and Pleistocene Mollusca of California and adjacent regions. San Diego Society of Natural History, Memoir 1: 1–1036.
Gray, J.E. 1824. Shells. A Supplement to the Appendix of Captain Parry’s Voyage for the Discovery of a North-West Passage, in the Years 1819–20: Containing an Account of the Subjects of Natural History. ccxl–ccxlvi. Murray, London.
Gray, J.E. 1840. Synopsis of the Contents of the British Museum. 370 pp. Trustees of the British Museum, London.
Gray, J.E. 1853. On the division of ctenobranchous gasteropodous Mollusca into larger groups and families. Annals and Magazine of Natural History 2: 124–132. Crossref
Guilding, L. 1834. Observations on Naticina and Dentalium, two genera of molluscous animals. Transactions of the Linnean Society of London 17: 29–35. Crossref
Habe, T. 1953. Fissurellidae in Japan (2). Publications of the Seto Marine Biological Laboratory 3: 33–50. Crossref
Hadorn, R. and Fraussen, K. 2005. Revision of the genus Granulifusus Kuroda & Habe 1954, with description of some new species (Gastropoda: Prosobranchia: Fasciolariidae). Archiv für Molluskenkunde 134: 129–171. Crossref
Hall, R. 2002. Cenozoic geological and plate tectonic evolution of SE Asia and the SW Pacific: computer-based reconstructions, models, and animations. Journal of Asian Earth Sciences 20: 353–431. Crossref
Hanna, G.D. 1924. Rectifications of nomenclature. Proceedings of the California Academy of Sciences, Fourth Series 13: 151–186.
Harzhauser, M. 2014. A seagrass-associated Early Miocene Indo-Pacific gastropod fauna from South West India (Kerala). Palaeontographica A 302: 73–178. Crossref
Hasegawa, K., Okutani, T., and Tsuchida, E. 2000. Family Turridae. In: T. Okutani (ed.), Marine Mollusks in Japan, 618–667. Tokai University Press, Tokyo.
Haszprunar, G. 1986. Die klado-evolutionäre Klassifikation-Versuch einer Synthese. Zeitschrift für Phylogenetische Systematik und Evolutionsforschung 24: 98–109. Crossref
Hawe, A., Paroll, C., and Haszprunar, G. 2014. Interactive 3D-anatomical reconstruction and affinities of the hot-vent gastropod Xylodiscula analoga Warén & Bouchet, 2001 (Ectobranchia). Journal of Molluscan Studies 80: 315–325. Crossref
Helbling, G.S. 1779. Beyträge zur Kenntniß neuer und seltener Konchylien. Aus einigen Wienerischen Sammlungen. Abhandlungen einer Privatgesellschaft in Böhmen, zur Aufnahme der Mathematik, der vaterländischen Geschichte, und der Naturgeschichte 4: 102–131.
Helwerda, R.A. 2015. Acteonidae, Bullinidae and Ringiculidae (Gastropoda: Heterobranchia) from the Plio-Pleistocene of the Philippines. Zootaxa 3990: 197–220. Crossref
Helwerda, R.A., Wesselingh, F.P., and Williams, S.T. 2014. On some Vetigastropoda (Mollusca, Gastropoda) from the Plio-Pleistocene of the Philippines with descriptions of three new species. Zootaxa 3755: 101–135.
Hickman, C.S. 1972. Review of the bathyal gastropod genus Phanerolepida (Homalopomatinae) and description of a new species from the Oregon Oligocene. The Veliger 15: 107–112.
Hikida, Y., Suzuki, S., Togo, Y., and Ijiri, A. 2003. An exceptionally well-preserved seep community from the Cretaceous Yezo forearc basin in Hokkaido, northern Japan. Paleontological Research 7: 329–342. Crossref
Hinds, R.B. 1843. Descriptions of new species of Nucula from the collections of Sir Edward Belcher, C.B., and Hugh Cuming, Esq. Proceedings of the Zoological Society of London 1843: 97–101.
Hirayama, K. 1954. On some Miocene species of Lucinoma from Japan, with description of two new species. Japanese Journal of Geology and Geography 25: 101–115.
Høisæter, T. and Johannessen, P.J. 2001. Xylodiscula planata sp. nov., a “lower” heterobranch gastropod from Norwegian waters. Sarsia 86: 325–332. Crossref
Hörnes, M. 1856. Die fossilen Mollusken des Tertiär-Beckens von Wien, I, Univalven. Abhandlungen der kaiserlich-königlichen geologischen Reichsanstalt 3: 1–736.
Hryniewicz, K., Amano, K., Jenkins, R.G., and Kiel, S. 2017. Thyasirid bivalves from Cretaceous and Paleogene cold seeps. Acta Palaeontologica Polonica 62: 705–728. Crossref
Hryniewicz, K., Bakayeva, S., Heneralova, L., Hnylko, O., Jenkins, R.G., and Kaim, A. 2020. Taphonomy and palaeoecology of deep-water chemosymbiotic bivalves from the Eocene of Outer Eastern Carpathians, Ukraine. Palaeogeography, Palaeoclimatology, Palaeoecology 553. Crossref
Hryniewicz, K., Miyajima, Y., Amano, K., Georgieva, M.N., Jakubowicz, M., Jenkins, R.G., and Kaim, A. 2021. Formation, diagenesis and fauna of cold seep carbonates from the Miocene Taishu Group of Tsushima (Japan). Geological Magazine 158: 964–984. Crossref
Hu, C.-H. 1995. Molluscan fossils from various localities in Taiwan Island (continued) [in Chinese]. Taiwan Molluscan Fossils [National Museum of Natural Science, Taichung] 4: 1927–2166.
Hutton, F.W. 1885. Descriptions of some Tertiary shells. Transactions of the New Zealand Institute 17: 313–332.
Hybertsen, F. and Kiel, S. 2018. A middle Eocene seep deposit with silicified fauna from the Humptulips Formation in western Washington State, USA. Acta Palaeontologica Polonica 63: 751–768. Crossref
Itoigawa, J. 1958. On some cephalaspid Opisthobranchia from the Japanese Miocene. Transactions and Proceedings of the Palaeontological Society of Japan, New Series 29: 175–184.
Jenkins, R.G., Kaim, A., and Hikida, Y. 2007a. Antiquity of the substrate choice among acmaeid limpets from the Late Cretaceous chemosynthesis-based communities. Acta Palaeontologica Polonica 52: 369–373. Crossref
Jenkins, R.G., Kaim, A., Hikida, Y., and Tanabe, K. 2007b. Methane-flux-dependent lateral faunal changes in a Late Cretaceous chemosymbiotic assemblage from the Nakagawa area of Hokkaido, Japan. Geobiology 5: 127–139.
Johnson, S.B., Krylova, E.M., Audzijonyte, A., Sahling, H., and Vrijenhoek, R.C. 2017. Phylogeny and origins of chemosynthetic vesicomyid clams. Systematics and Biodiversity 15: 346–360. Crossref
Kaim, A., Jenkins, R.G., and Hikida, Y. 2009. Gastropods from Late Cretaceous hydrocarbon seep deposits in Omagari and Yasukawa, Nakagawa area, Hokkaido, Japan. Acta Palaeontologica Polonica 54: 463–690. Crossref
Kamada, Y. 1962. Tertiary marine Mollusca from the Joban coal-field, Japan. Special Papers, Palaeontological Society of Japan 8: 1–187.
Kamenev, G.M. 2009. North Pacific species of the genus Solemya Lamarck, 1818 (Bivalvia: Solemyidae), with notes on Acharax johnsoni (Dall, 1891). Malacologia 51: 233–261. Crossref
Kanie, Y. and Kuramochi, T. 1995. Acharax yokosukaensis n. sp. (giant bivalve) from the Miocene Hayama Formation of the Miura Peninsula, south-central Japan. Science Report of the Yokosuka City Museum 43: 51–57.
Kanie, Y. and Kuramochi, T. 2002. Review of the family Solemyidae (Mollusca: Bivalvia) from Japan, and description of the new species. Science Report of the Yokosuka City Museum 49: 49–60.
Kanno, S. 1960. Part II, Palaeontology. In: J. Arai, and S. Kanno (eds.), The Tertiary System of the Chichibu Basin, Saitama Prefecture, central Japan, 123–396. Japan Society for the Promotion of Science, Ueno, Tokyo.
Karig, D.E. 1983. Accreted terrains in the northern part of the Philippines archipelago. Tectonics 2: 211–236. Crossref
Kase, T., Isaji, S., Aguilar, Y.M., and Kiel, S. 2019. A large new Wareniconcha (Bivalvia: Vesicomyidae) from a Pliocene methane seep deposit in Leyte, Philippines. The Nautilus 133: 26–30.
Kenk, V.C. and Wilson, B.R. 1985. A new mussel (Bivalvia: Mytilidae) from hydrothermal vents in the Galapagos Rift Zone. Malacologia 26: 253–271.
Kiel, S. 2008. Fossil evidence for micro- and macrofaunal utilization of large nekton-falls: examples from early Cenozoic deep-water sediments in Washington State, USA. Palaeogeography, Palaeoclimatology, Palaeoecology 267: 161–174. Crossref
Kiel, S. 2010a. An Eldorado for paleontologists: the Cenozoic seep deposits of western Washington State, USA. In: S. Kiel (ed.), The Vent and Seep Biota, 433–448. Springer, Heidelberg. Crossref
Kiel, S. 2010b. The fossil record of vent and seep mollusks. In: S. Kiel (ed.), The Vent and Seep Biota, 255–278. Springer, Heidelberg. Crossref
Kiel, S. 2013. Lucinid bivalves from ancient methane seeps. Journal of Molluscan Studies 79: 346–363. Crossref
Kiel, S. 2015. Did shifting seawater sulfate concentrations drive the evolution of deep-sea vent and seep ecosystems? Proceedings of the Royal Society B 282: 20142908. Crossref
Kiel, S., Aguilar, Y.M., and Kase, T. 2020a. Mollusks from Pliocene and Pleistocene seep deposits in Leyte, Philippines. Acta Palaeontologica Polonica 65: 589–627. Crossref
Kiel, S., Amano, K., and Jenkins, R.G. 2008. Bivalves from Cretaceous cold-seep deposits on Hokkaido, Japan. Acta Palaeontologica Polonica 53: 525–537. Crossref
Kiel, S. and Goedert, J.L. 2007. Six new mollusk species associated with biogenic substrates in Cenozoic deep-water sediments in Washington State, USA. Acta Palaeontologica Polonica 52: 41–52.
Kiel, S. and Hansen, B.T. 2015. Cenozoic methane-seep faunas of the Caribbean region. PLoS ONE 10: e0140788. Crossref
Kiel, S. and Little, C.T.S. 2006. Cold seep mollusks are older than the general marine mollusk fauna. Science 313: 1429–1431. Crossref
Kiel, S. and Peckmann, J. 2008. Paleoecology and evolutionary significance of an Early Cretaceous Peregrinella-dominated hydrocarbon-seep deposit on the Crimean Peninsula. Palaios 23: 751–759. Crossref
Kiel, S. and Taviani, M. 2017. Chemosymbiotic bivalves from Miocene methane-seep carbonates in Italy. Journal of Paleontology 91: 444–466. Crossref
Kiel, S. and Taviani, M. 2018. Chemosymbiotic bivalves from the late Pliocene Stirone River hydrocarbon seep complex in northern Italy. Acta Palaeontologica Polonica 63: 557–568. Crossref
Kiel, S., Campbell, K.A., and Gaillard, C. 2010. New and little known mollusks from ancient chemosynthetic environments. Zootaxa 2390: 26–48. Crossref
Kiel, S., Hybertsen, F., Hyžný, M., and Klompmaker, A.A. 2020b. Mollusks and a crustacean from early Oligocene methane-seep deposits in the Talara Basin, northern Peru. Acta Palaeontologica Polonica 65: 109–138. Crossref
Kojima, S. and Ohta, S. 1997. Calyptogena okutanii n. sp., a sibling species of Calyptogena soyae Okutani, 1957 (Bivalvia: Vesicomyidae). Venus 56: 189–195.
Krishtofovich, L.V. 1936. Shells of the group Thyasira bisecta (Conrad) from the Tertiary deposits of the west coast of Kamchatka [in Russian]. Trudy Neftânogo-Razvedočnogo Instituta, Seriâ A 88: 3–67.
Krishtofovich, L.V. 1964. Molluscs from the Tertiary deposits of Sakhalin [in Russian]. Transactions of the All-Union Petroleum Scientific-Research Geological-Exploring Institute 232: 1–344.
Krylova, E.M. and Cosel, R. von 2011. A new genus of large Vesicomyidae (Mollusca, Bivalvia, Vesicomyidae, Pliocardiinae) from the Congo margin with the first record of the subfamily Pliocardiinae in the Bay of Biscay (northeastern Atlantic). Zoosystema 33: 83–99. Crossref
Krylova, E.M. and Sahling, H. 2006. Recent bivalve molluscs of the genus Calyptogena (Vesicomyidae). Journal of Molluscan Studies 72: 359–395. Crossref
Kuroda, T. 1931. Fossil Mollusca. In: F. Homma (ed.), Geology of the Central Part of Shinano, Part 4, 1–90 [in Japanese]. Kokon Shoin, Tokyo.
Kuroda, T., Habe, T., and Oyama, K. 1971. The Sea Shells of Sagami Bay. xix + 741 pp. (Japanese text), 489 pp. (English text), 51 pp. (Index). Maruzen Co., Tokyo.
Laws, C.R. 1947. Tertiary Mollusca from Hokianga District, North Auckland. Transactions of the Royal Society of New Zealand 76: 537–541.
Linnaeus, C. 1758. Systema Naturae (tenth edition). 824 pp. Laurrentii Salvii, Holmiae.
Little, C.T.S., Herrington, R.J., Maslennikov, V.V., Morris, N.J., and Zaykov, V.V. 1997. Silurian hydrothermal vent community from the southern Urals, Russia. Nature 385: 146–148. Crossref
Lorentz, R.A. 1984. Stratigraphy and sedimentation of late Neogene sediments on the southwest flank of Luzon Central Cordillera, Philippines. Journal of Philippine Geology 38: 1–24.
MacNeil, F.S. 1960. Tertiary and Quaternary Gastropoda from Okinawa. U.S. Geological Survey Professional Paper 339: 1–148. Crossref
Magtoto, C.Y. and Fernando, A.G. 2017. Calcareous nannofossil biostratigraphy of the mudstone-limestone interbeds exposure in Northern Cement Corporation (NCC) Quarry, Sison, Pangasinan (Philippines). Bulletin of the National Science Museum C 43: 49–63.
Majima, R., Jenkins, R.G., Kase, T., Aguilar, Y.M., Nanjo, T., Wani, R., Wada, H., Fernando, A.G.S., and Hayashi, H. 2010. In situ Calyptogena colonies from Pliocene back-arc basin fills in Leyte Island, Philippines. Journal of the Geological Society of Japan 116: xv–xvi. Crossref
Majima, R., Kase, T., Kawagata, S., Aguilar, Y.M., Hagino, K., and Maeda, M. 2007. Fossil cold-seep assemblages from Leyte Island, Philippines. Journal of Geography 116: 643–652. Crossref
Majima, R., Nobuhara, T., and Kitazaki, T. 2005. Review of fossil chemosynthetic assemblages in Japan. Palaeogeography, Palaeoclimatology, Palaeoecology 227: 86–123. Crossref
Makiyama, J. 1941. Evolution of the gastropod genus Siphonalia with accounts on the Pliocene species of Totomi and other examples. The Ketienzian fauna series, no. 1. Memoirs of the College of Science, Kyoto Imperial University, Series B 16: 75–93.
Marshall, B.A. 1988. Skeneidae, Vitrinellidae and Orbitestellidae (Mollusca: Gastropoda) associated with biogenic substrata from bathyal depths off New Zealand and New South Wales. Journal of Natural History 22: 949–1004. Crossref
Martin, K. 1883. Palaeontologische Ergebnisse von Tiefbohrungen auf Java, nebst allgemeineren Studien ueber das Tertiaer von Java, Timor und einiger anderer Inseln. Sammlungen des Geologischen Reichs-Museums in Leiden. Serie 1, Beiträge zur Geologie Ost-Asiens und Australiens 3: 1–380.
Martin, K. 1891–1906. Die Fossilien von Java auf Grund einer Sammlung von Dr. R.D.M. Verbeek. Sammlung des Geologischen Reichsmuseum in Leiden, Neue Serie 1: 1–331. Crossref
Martin, K. 1933. Eine neue tertiäre Molluskenfauna aus dem indischen Archipel. Leidsche Geologische Mededeelingen 6: 7–32.
Marwick, J. 1931. The Tertiary Mollusca of the Gisborne District. New Zealand Geological Survey Bulletin 113: 1–177.
Matsubara, T. 2011. Miocene shallow marine molluscs from the Hokutan Group in the Tajima area, Hyogo Prefecture, southwest Japan. Bulletin of the Mizunami Fossil Museum 37: 51–113.
McLean, J.H. 1981. The Galapagos Rift limpet Neomphalus: Relevance to understanding the evolution of a major Paleozoic–Mesozoic radiation. Malacologia 21: 291–336.
McLean, J.H. and Quinn, J.F.J. 1987. Cataegis, new genus of three new species from the continental slope (Trochidae: Cataeginae new subfamily). The Nautilus 101: 111–116. Crossref
Meek, F.B. 1864. Check list of the invertebrate fossils of North America—Miocene. Smithsonian Miscellaneous Collection 7: 1–32. Crossref
MGB 2010. Geology of the Philippines. Second Edition. 532 pp. Mines and Geosciences Bureau, Quezon City, Philippines.
Miyajima, Y., Nobuhara, T., and Koike, H. 2017. Taxonomic reexamination of three vesicomyid species (Bivalvia) from the middle Miocene Bessho Formation in Nagao Prefecture, central Japan, with notes on vesicomyid diversity. The Nautilus 131: 51–66.
Miyajima, Y., Watanabe, Y., Jenkins, R.G., Goto, A.S., and Hasegawa, T. 2018. Diffusive methane seepage in ancient deposits: examples from the Neogene Shin’etsu sedimentary basin, central Japan. Journal of Sedimentary Research 88: 449–466. Crossref
d’Montfort, D.P. 1810. Conchyliologie systématique et classification méthodique des coquilles: Offrant leur figures, leur arrengement générique, leur description charactéristiques, leur noms; ainsi que leur synonymie en plusieurs langues. 676 pp. F. Schoell, Paris.
Moore, E.J. 1963. Miocene marine mollusks from the Astoria Formation in Oregon. U.S. Geological Survey Professional Paper 419: 1–109. Crossref
Morassi, M. and Bonfitto, A. 2010. New raphitomine gastropods (Gastropoda: Conidae: Raphitominae) from the South-West Pacific. Zootaxa 2526: 54–68. Crossref
Moroni, M.A. 1966. Malacofauna del “Calcare a Lucine” di S. Sofia, Forlì. Palaeontographica Italica 60: 69–87.
Morton, B. and Scott, P.H. 1989. The Hong Kong Galeommatacea (Mollusca: Bivalvia) and their hosts, with descriptions of new species. Asian Marine Biology 6: 129–160.
Mörch, A.O.L. 1852. Catalogus conchyliorum quae reliquit D. Alphonso d’Aguirra et Gadea Comes de Yoldi. 170 pp. Hafnie, Copenhagen. Crossref
Nagao, T. and Huzioka, K. 1941. Fossil Acila from Hokkaido and Karafuto (Saghalien). Journal of the Faculty of Science, Hokkaido Imperial University, Series IV 6: 113–141.
Nesbitt, E.A., Martin, R.A., and Campbell, K.A. 2013. New records of Oligocene diffuse hydrocarbon seeps, northern Cascadia margin. Palaeogeography, Palaeoclimatology, Palaeoecology 390: 116–129. Crossref
Newton, R.B. 1891. Systematic List of the F.E. Edwards Collection of British Oligocene and Eocene Mollusca in the British Museum (Natural History), With References to the Type-specimens from Similar Horizons Contained in Other Collections Belonging to the Geological Department of the Museum. xxviii + 365 pp. British Museum, London.
Noda, H. 1988. Molluscan fossils from the Ryukyu Islands, southwest Japan Part 2. Gastropoda and Pelecypoda from the Shinzato Formation in the middle part of Okinawa-jima. Science Reports of the Institute of Geoscience University of Tsukuba Section B Geological Sciences 9: 29–85.
Okutani, T. 1957. Two new species of bivalves from the deep water in Sagami Bay collected by the R.V. Soyo-Maru. Bulletin of Tokai Regional Fisheries Research Laboratory 17: 27–31.
Okutani, T. 1962. Report on the archibenthal and abyssal Lamellibranchiate Mollusca mainly collected from Sagami Bay and adjacent waters by the RV Soyo-Maru during the years 1955–1960. Contributions from the Tokai Regional Fisheries Research Laboratory 160: 1–40.
Okutani, T. and Tsuchiya K. 2000. Family Fasciolariidae. In: T. Okutani (ed.), Marine Mollusks in Japan, 505–519. Tokai University Press, Tokyo.
Okutani, T. 2001. Six new bathyal and shelf trochoidean species in Japan. Venus 60: 121–127.
Okutani, T. 2002. A new thyasirid Conchocele novaeguinensis n. sp. from a thanatocoenosis associated with a possible cold seep activity off New Guinea. Venus 61: 141–145.
Okutani, T. 2011. Bizarre lucinid bivalves from southwestern Japan, including a new species, and relatives in adjacent waters. Venus 69: 115–122.
Okutani, T. and Fujiwara, Y. 2000. Gastropod fauna of a thermal vent site on the North Knoll of Iheya Ridge, Okinawa Trough. Venus 59: 123–128.
Okutani, T., Fujikura, K., and Kojima, S. 2000. New taxa and review of vesicomyid bivalves collected from the Northwest Pacific by deep sea research systems of Japan Marine Science & Technology Center. Venus 59: 83–101.
Okutani, T., Hashimoto, J., and Fujikura, K. 1992a. A new species of vesicomyid bivalve associated with hydrothermal vents near Amami-Oshima Island, Japan. Venus 51: 225–233.
Okutani, T., Kojima, S., and Ashi, J. 1997. Further discovery of a new taxon of vesicomyid clam from the Nankai Trough, off Honshu, Japan. Venus 56: 185–188.
Okutani, T., Tsuchida, S., and Fujikura, K. 1992b. Five bathyal gastropods living within or near the Calyptogena-community of the Hatsushima Islet, Sagami Bay. Venus 51: 137–148.
Oliver, P.G. 2013. Description of Atopomya dolobrata gen. et sp. nov.: first record of bacterial symbiosis in the Saxicavellinae (Bivalvia). Journal of Conchology 41: 359–367.
Olu, K., Sibuet, M., Harmegnies, F., Foucher, J.-P., and Fiala-Médoni, A. 1996. Spatial distribution of diverse cold seep communities living on various diapiric structures of the southern Barbados prism. Progress in Oceanography 38: 347–376. Crossref
Pallary, P. 1904. Addition à la faune Malacologique de la Golfe de Gabès. Journal de Conchyliologie 52: 212–248.
Paull, C.K., Hecker, B., Commeau, R., Freeman-Lynde, R.P., Neumann, C., Corso, W.P., Golubic, S., Hook, J.E., Sikes, E., and Curray, J. 1984. Biological communities at the Florida Escarpment resemble hydrothermal vent taxa. Science 226: 965–967. Crossref
Pelseneer, P. 1889. Sur la classification phylogenetique des pélécypodes. Bulletin scientifique de la France et de la Belgique 20: 27–52.
Peña, R.E. 2008. Lexicon of Philippine Stratigraphy. 364 pp. The Geological Society of the Philippines, Inc., Manila.
Pérez-Barría, L. and Nielsen, S.N. 2020. Solemyidae (Bivalvia, Protobranchia) from the lower Miocene of south-central Chile, with description of a new species. Journal of Paleontology 94: 56–63. Crossref
Philippi, R.A. 1853. Handbuch der Conchyliologie und Malakozoologie. i–xx + 547 pp. Hermann Eduard Anton, Halle. Crossref
Pilsbry, H.A. 1895. Catalogue of the Marine Mollusks of Japan. 196 pp. Frederick Stearns, Detroit.
Pinet, N. and Stephan, J.F. 1990a. The Ilocos foothills and western Central Cordillera: a key region for the understanding of the geodynamic evolution of the Eurasian margin in Luzon area (Philippines). In: J. Aubouin, and J. Bourgois (eds.), Tectonics and Circum-Pacific Continental Margins, 165–180. VSP, Utrecht.
Pinet, N. and Stephan, J.F. 1990b. The Philippine wrench fault system in the Ilocos Foothills, northwestern Luzon, Philippines. Tectonophysics 183: 207–224. Crossref
Rafinesque, C.S. 1815. Analyse de la nature, ou Tableau de l’univers et des corps organisées. 224 pp. Barraveccia, Palermo. Crossref
Robba, E., Geronimo, I.D., Chaimanee, N., Negri, M.P., and Sanfilippo, R. 2004. Holocene and Recent shallow soft-bottom mollusks from the northern Gulf of Thailand area: Scaphopoda, Gastropoda, additions to Bivalvia. La Conchiglia 35: 1–288.
Ryan, W.B.F., S.M. Carbotte, J.C., O’Hara, S., Melkonian, A., Arko, R., Weissel, R.A., Ferrini, V., Goodwillie, A., Nitsche, F., Bonczkowski, J., and Zemsky, R. 2009. Global Multi-Resolution Topography (GMRT) synthesis data set. Geochemistry, Geophysics, Geosystems 10: Q03014. Crossref
Sacco, F. 1901. I molluschi dei terreni terziarii del Piemonte e della Liguria. Parte XXIX. 217 pp. Carlo Clausen, Torino.
Saether, K.P., Jingeng, S., Little, C.T.S., and Campbell, K.A. 2016. New records and a new species of bivalve (Mollusca: Bivalvia) from Miocene hydrocarbon seep deposits, North Island, New Zealand. Zootaxa 4154: 1–26. Crossref
Saether, K.P., Little, C.T.S., and Campbell, K.A. 2010a. A new fossil provannid gastropod from Miocene hydrocarbon seep deposits, East Coast Basin, North Island, New Zealand. Acta Palaeontologica Polonica 55: 507–517. Crossref
Saether, K.P., Little, C.T.S., Campbell, K.A., Marshall, B.A., Collins, M., and Alfaro, A.C. 2010b. New fossil mussels (Bivalvia: Mytilidae) from Miocene hydrocarbon seep deposits, North Island, New Zealand, with general remarks on vent and seep mussels. Zootaxa 2577: 1–45. Crossref
Saldivar-Sali, A. 1978. Reef exploration in the Philippines. In: Second Circum-Pacific Energy and Mineral Resources Conference, Honolulu, Hawaii, 30 July–4 August 1978. 54 pp.
Samadi, S., Puillandre, N., Pante, E., Boisselier, M.-C., Chen, W.-J., Corbari, L., Maestrati, P., Mana, R., Thubaut, J., Zuccon, D., and Hourdez, S. 2015. Patchiness of deep-sea communities in Papua New Guinea and potential susceptibility to anthropogenic disturbances illustrated by seep organisms. Marine Ecology 36 (Supplement 1): 109–132. Crossref
Sasaki, T., Okutani, T., and Fujikura, K. 2005. Molluscs from hydrothermal vents and cold seeps in Japan: A review of taxa recorded in twenty recent years. Venus 64: 87–133.
Schenck, H.G. 1936. Nuculid bivalves of the genus Acila. Geological Society of America Special Paper 4: 1–149.
Schepman, M.M. (ed.) 1913. The Prosobranchia of the Siboga Expedition, Part 5. 365–452. E.J. Brill, Leiden.
Seguenza, G. 1879. Le formazioni terziarie della provincia di Reggio (Calabria). Memorie della Reale Accademia dei Lincei, Classe di Scienze Fisiche, Matematiche e Naturalistiche 3: 1–446. Crossref
Shuto, T. 1958. Granulifusus from the Miyazaki Group. Transactions and Proceedings of the Palaeontological Society of Japan, New Series 31: 253–264.
Snyder, M.A. and Bouchet, P. 2006. New species and new records of deep-water Fusolatirus (Neogastropoda: Fasciolariidae) from the West Pacific. Journal of Conchology 39: 1–12.
Sowerby, G.B. 1850. Thesaurus conchyliorum, or Monographs of Genera of Shells. Vol. 2, 553–899 pp. Sowerby, London.
Squires, R.L. and Goedert, J.L. 1996. A new species of Thalassonerita? (Gastropoda: Neritidae?) from a Middle Eocene cold-seep carbonate in the Humptulips Formation, Western Washington. The Veliger 39: 270–272.
Squires, R.L. and Gring, M.P. 1996. Late Eocene chemosynthetic? bivalves from suspect cold seeps, Wagonwheel Mountain, central California. Journal of Paleontology 70: 63–73. Crossref
Stimpson, W. 1851. A Revision of the Synonymy of the Testaceous Mollusks of New England, With Notes on Their Structure, And Their Geographical and Bathymetrical Distribution. 58 pp. Phillips, Sampson, and Company, Boston. Crossref
Sturany, R. 1896. Mollusken I (Prosobranchier und Opisthobranchier; Scaphopoden; Lamellibranchier) gesammelt von S.M. Schiff “Pola” 1890–94. Denkschriften der mathematisch-naturwissenschaftlichen Klasse der Kaiserlichen Akademie der Wissenschaften 63: 1–36.
Swainson, W. 1829–1833. Zoological Illustrations, or Original Figures and Descriptions of New, Rare, or Interesting Animals, Selected Chiefly from the Classes of Ornithology, Entomology, and Conchology, and Arranged According to Their Apparent Affinities. Second series. pls. 1–30 (1829), pls. 31–45 (1830), pls. 46–85 (1831), pls. 86–96 (1832), pls. 97–136 (1833). Baldwin & Cradock, London.
Sykes, E.R. 1930. On a new species of Turris from Japan. Proceedings of the Malacological Society of London 19: 82. Crossref
Taviani, M. 1994. The “calcari a Lucina” macrofauna reconsidered: Deep-sea faunal oases from Miocene-age cold vents in the Romagna Apennine, Italy. Geo-Marine Letters 14: 185–191. Crossref
Taviani, M., Angeletti, L., and Ceregato, A. 2011. Chemosynthetic bivalves of the family Solemyidae (Bivalvia, Protobranchia) in the Neogene of the Mediterranean Basin. Journal of Paleontology 85: 1067–1076. Crossref
Taylor, J.D. and Glover, E.A. 1997. A chemosymbiotic lucinid bivalve (Bivalvia: Lucinoidea) with periostracal pipes: functional morphology and description of a new genus and species. In: F.E. Wells (ed.), The Marine Flora and Fauna of the Houtman Abrolhos Islands, Western Australia, 335–361. Western Australian Museum, Perth.
Taylor, J.D. and Glover, E.A. 2000. Nomenclatural rectifications for Indo-Pacific Lucinidae. Journal of Conchology 37: 82.
Taylor, J.D., Glover, E.A., and Williams, S.T. 2008. Ancient shallow water chemosynthetic bivalves: systematics of Solemyidae from eastern and southern Australia (Mollusca, Bivalvia). Memoires of the Queensland Museum Nature 54: 75–104.
Taylor, J.D., Glover, E.A., Zuschin, M., Dworschak, P.C., and Waitzbauer, W. 2005. Another bivalve with dreadlocks: living Rasta lamyi from Aqaba, Red Sea (Bivalvia: Lucinidae). Journal of Conchology 38: 489–497.
Thiele, J. and Jaeckel, S. 1931. Muscheln der Deutschen Tiefsee-Expedition. II Teil. Deutsche Tiefsee-Expedition 1898–1899 21: 159–268.
Tsuda, K. 1959. New Miocene molluscs from the Kurosedani Formation in Toyama Prefecture, Japan. Journal of the Faculty of Science, Niigata University, Series II 3: 67–110.
Tunnicliffe, V. 1991. The biology of hydrothermal vents: Ecology and evolution. Oceanographic and Marine Biology Annual Review 29: 319–407.
Valdés, Á. 2008. Deep-sea “cephalaspidean” heterobranchs (Gastropoda) from the tropical southwest Pacific. Mémoires de Muséum National de Histoire Naturelle 196: 587–792.
Valdés, F., Sellanes, J., and D’Elía, G. 2013. Phylogenetic position of vesicomyid clams from a methane seep off central Chile (~36°S) with a molecular timescale for the diversification of the Vesicomyidae. Zoological Studies 51: 1154–1164.
Van Winkle, K. 1919. Remarks on some new species from Trinidad. Bulletins of American Paleontology 8: 19–27.
Vermeij, G.J. and Snyder, M.A. 2018. Proposed genus-level classification of large species of Fusininae (Gastropoda, Fasciolariidae). Basteria 82: 57–82.
Vilvens, C. 2016. New records and new species of Cataegis (Gastropoda: Seguenzioidea) from Solomon Islands. Novapex 17: 67–76.
Vrijenhoek, R.C. 2013. On the instability and evolutionary age of deep-sea chemosynthetic communities. Deep-Sea Research II 92: 189–200. Crossref
Warén, A. 1992. New and little known “Skeneimorph” gastropods from the Mediterranean Sea and the adjacent Atlantic Ocean. Bollettino Malacologico 27: 149–248.
Warén, A. and Bouchet, P. 1993. New records, species, genera, and a new family of gastropods from hydrothermal vents and hydrocarbon seeps. Zoologica Scripta 22: 1–90. Crossref
Warén, A. and Bouchet, P. 2001. Gastropoda and Monoplacophora from hydrothermal vents and seeps; new taxa and records. The Veliger 44: 116–231.
Warén, A. and Rouse, G.W. 2016. A new genus and species of Cataegidae (Gastropoda: Seguenzioidea) from eastern Pacific Ocean methane seeps. Novapex 17: 59–66.
Weinkauff, H.C. 1875. Ueber eine kritische Gruppe des Genus Pleurotoma Lam. sensu stricto. Jahrbücher der Deutschen Malakozoologischen Gesellschaft 2: 283–292.
Woodring, W.P. 1925. Miocene Mollusca from Bowden Jamaica, pelecypods and scaphopods. Carnegie Institution of Washington publications 336: 1–564. Crossref
Yabe, H. and Nomura, S. 1925. Notes on the Recent and Tertiary species of Thyasira from Japan. Science Reports of the Tohoku Imperial University, 2nd Series (Geology) 7: 83–95.
Yokoyama, M. 1928. Mollusca from the oil-field of the island of Taiwan. Report of the Imperial Geological Survey of Japan 101: 1–112.
Yumul, G.P.J., Dimalanta, C.B., Maglambayan, V.B., and Marquez, E.J. 2008. Tectonic setting of a composite terrane: A review of the Philippine island arc system. Geosciences Journal 12: 7–17. Crossref
Zhang, S., Zhang, J., and Zhang, S. 2020a. Integrative taxonomy reveals new taxa of Trochidae (Gastropoda: Vetigastropoda) from seamounts in the tropical western Pacific. Deep-Sea Research II 159: 103234. Crossref
Zhang, S. and Zhang, S. 2018. Three species of Siphonalia Adams, 1863 (Gastropoda, Buccinidae) from China seas, with descriptions of two new species. Journal of Oceanology and Limnology 36: 2333–2336. Crossref
Zhang, S., Zhang, S., and Chen, H. 2020b. Enigmaticolus inflatus sp. nov., a new buccinid species from a methane seep area in the South China Sea (Gastropoda: Neogastropoda). Zootaxa 4728: 385–389. Crossref
Acta Palaeontol. Pol. 67 (4): 917–947, 2022
https://doi.org/10.4202/app.00977.2022