
Chaetognath grasping spines from the Devonian of Poland: their structure and geochemistry
HUBERT WIERZBOWSKI and BŁAŻEJ BŁAŻEJOWSKI
Wierzbowski, H. and Błażejowski, B. 2023. Chaetognath grasping spines from the Devonian of Poland: their structure and geochemistry. Acta Palaeontologica Polonica 68 (1): 103–116.
Previously unidentified small (up to 1.3 mm in length), gently curved hollow spines composed of calcium phosphate and derived from the upper Famennian (Devonian) of the Holy Cross Mountains in central Poland are described. They are similar to the type species of Phakelodus, but show some distinct morphological differences, therefore, have been included into Phakeloides polonicus gen. et sp. nov. The studied specimens of that species show relatively massive mineral structure characterized by significant porosity with well-preserved major structural features. The outer layer of the spines is, in contrary, fragmentarily preserved, and exposes distinct mosaic-like ornamentation of the surface of the middle layer, which consists of obliquely arranged, shallow furrows. Geochemical analyses of the Devonian Phakeloides polonicus gen. et sp. nov. spines have revealed the presence of a weaker mineralized structure, compared to conodont apatite, composed of a diagenetic phosphate phase. It is characterized by moderate cathodoluminescence intensity, elevated concentrations of iron and sulphur as well as decreased concentrations of strontium, calcium, and phosphorus. The “Orsten” type, early diagenetic phosphatization of the outer layer of the spines is not observed in studied specimens of Phakeloides polonicus gen. et sp. nov., contrary to the previously investigated Furongian (Cambrian) material. This points to the low rate of diagenetic phosphatization, which was likely enabled by very slow sedimentation and long residence time of the spines close to sediment-water interface at varying redox conditions and significant flux of phosphate ions.
Key words: Conodonta, Protoconodonta, Chaetognatha, elemental ratios, microstructure, phosphatized spines, Devonian, Holy Cross Mountains, Poland.
Hubert Wierzbowski [hubert.wierzbowski@pgi.gov.pl; ORCID: https://orcid.org/0000-0003-2682-2945 ] Polish Geological Institute-National Research Institute, ul. Rakowiecka 4, 00-975 Warsaw, Poland.
Błażej Blażejowski [bblazej@twarda.pan.pl; ORCID: https://orcid.org/0000-0001-6320-9870 ] (corresponding author) Institute of Paleobiology, Polish Academy of Sciences, ul. Twarda 51/55, 00-818 Warsaw, Poland.
Received 4 July 2022, accepted 2 February 2023, available online 7 March 2023.
Copyright © 2023 H. Wierzbowski and B. Blażejowski. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The occurrence of phosphatic chaetognath grasping spines, previously included into protoconodonts, is generally restricted to the lower Palaeozoic (cf. Bengtson 1976, 1983; Szaniawski 1982, 1983, 2002, 2005, 2009, 2015; Andres 1988; McIlroy and Szaniawski 2000; Carlorosi et al. 2019). The spines, mostly assigned to the genera Phakelodus (earlier Prooneotodus) and Protoherzina, are considered a product of diagenetic phosphatization of originally organic chaetognath elements based on similarity in their architecture and microstructure, findings of the natural clusters of spines arranged in a form of chaetognath grasping apparatus (Szaniawski 1982, 2002, 2009, 2015; McIlroy and Szaniawski 2000; Vannier et al. 2007), their conserved plastic deformations (Doguzhaeva et al. 2002; Szaniawski 2002; Wierzbowski et al. 2021) and a growing number of discoveries of preserved fossil carbonaceous spines, which sometimes co-occur with remnants of soft tissues of chaetognaths (Szaniawski 2005; Vannier et al. 2007; Briggs and Caron 2017; Shu et al. 2017; Slater et al. 2018; Slater and Willman 2019). In addition, Wierzbowski et al. (2021) have recently documented the presence of a few generations of diagenetic phosphate phases in Furongian (Cambrian) chaetognath Phakelodus sp. spines from the Öland Island (Sweden). A thin, outermost layer of the spines is composed of early diagenetic apatite phase, which incrusted the original cuticle integument, contributing to the preservation of fine details of their surface. Its mineralization is assigned to the “Orsten”-type phosphatization process known from the excellently preserved three-dimensional small invertebrate fossils of the early Palaeozoic age (cf. Müller and Walossek 1985; Waloszek 2003; Maas et al. 2006; Wierzbowski 2021).
There are, however, other Cambrian–lowermost Ordovician problematic protoconodont-like fossils that show a strong resemblance to chaetognath grasping spines. Some of the protoconodont-like cones assigned to the genera Coelocerodontus and Virodus have recently been interpreted as chaetognaths grasping spines (Szaniawski 2015). Fossils assigned to Rhombocorniculum cancellatum despite structural affinity to chaetognath grasping spines are included into pseudoconodonts and regarded as sclerites of Hallucigenia-type lobopodians or other incertae sedis (Brasier 1989; Guoxiang et al. 2003; Caron et al. 2013; Kouchinsky et al. 2015).
A relatively large number of discoveries of Cambrian–Lower Ordovician chaetognath phosphatic grasping spines may be linked to phosphorus re-mobilization and high phosphate concentrations in ancient pore waters. It was probably linked to elevated phosphorus input to the oceans and diminished sediment mixing and ventilation before the Ordovician diversification of burrowing organisms, which may have contributed to the elevated position of oxic-anoxic interface within the sediment and fluctuating redox conditions in the upper part of sediment column (Allison and Briggs 1993; Brassier and Callow 2007; Dornbos 2011; Creveling et al. 2014). Rare findings of phosphatized chaetognath grasping spines are, nevertheless, reported from upper Palaeozoic deposits (Doguzhaeva et al. 2002).
The present paper is devoted to newly found upper Palaeozoic phosphatic chaetognath grasping spines from the upper Famennian (Devonian) of the Holy Cross Mountains in central Poland. The spine assemblage is composed of numerous, almost identical, isolated elements. The spines have been investigated morphologically and microstructurally. The present study highlights disparities in the microstructure and chemical composition among the new chaetognath material, consisting of secondary phosphate, co-occurring conodont elements built of primary phosphate, and previously described phosphatized chaetognath spines of the Furongian (Cambrian) age from Sweden (cf. Wierzbowski et al. 2021). The taxonomical affinity of studied Devonian fossils to previously described Cambrian forms is also discussed. The purpose of this study is to find apparent similarities and/or differences in the morphology, construction and structure of phosphatized spines of early and late Palaeozoic chaetognaths along with the specific pathways of their diagenetic mineralization processes. This study also demonstrates the potential of upper Palaeozoic deposits for further research on the evolution and physiological functions of chaetognaths and other problematic spines of invertebrates.
Institutional abbreviations.—ZPAL, Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland.
Other abbreviations.—CAI, conodont colour-alteration index.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in ZooBank: urn:lsid:zoobank.org:pub:26225CF8-3682-414D-A16B-4883AC67E04B.
Geological setting
Newly found upper Famennian (Devonian) chaetognath grasping spines come from the extensively exploited Ostrówka Quarry, which is situated on the Ostrówka Hill, near the village of Gałęzice, in the south-western part of the Holy Cross Mountains in central Poland (Fig. 1A, B). In the quarry are exposed upper Devonian limestone beds, which are covered by lower Carboniferous carbonate-mudstone strata (Fig. 1C). This lithological succession was formed as a result of drowning of a Frasnian (upper Devonian) shallow-water carbonate platform deposits, which are overlain by a discontinuity surface and condensed Famennian (upper Devonian) limestone beds. Tournaisian and Visean (lower Carboniferous) deep water limestone-clay beds, black siliceous shale and carbonate gravity flow deposits occur higher in the section (Szulczewski et al. 1996).
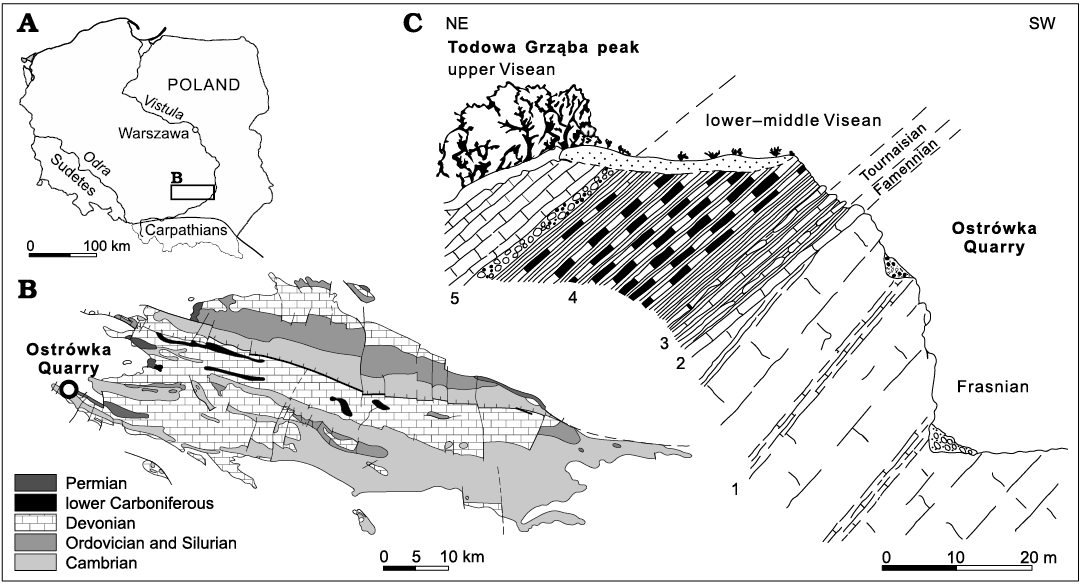
Fig. 1. Geographical location (A) and simplified geologic map (B) of the Holy Cross Mountains with location of the studied exposure. C. Cross-section through the ledge of the Ostrówka Quarry. 1, peritidal carbonates (Kowala Formation); 2, condensed crinoidal-cephalopod limestone beds; 3, clay with limestone intercalations; 4, black radiolarian shale (Zaręby Beds); 5, carbonate gravity flows (after Szulczewski et al. 1996).
Facies development and stratigraphic completeness of Famennian exposures from the northern wall of the Ostrówka Quarry is variable. The most complete Famennian section is located on the Todowa Grząba peak in the Gałęzice region (Fig. 1; Szulczewski 1978; Szulczewski et al. 1996). Currently, only a few relatively thin beds of extremely condensed crinoidal-cephalopod Famennian limestone can be found above an erosional discontinuity surface developed on the top of shallow-water Frasnian “Amphipora limestones” (Racki 1993; Szulczewski et al. 1996; Berkowski 2002). Stratigraphical studies of the condensed unit have revealed a standard succession of Famennian conodont zones, interrupted by several internal paraconformities (cf. Szulczewski 1971, 1978; Szulczewski et al. 1996). The oldest deposits covering the unconformity belong to the upper part of the middle Famennian Conditolepis marginifera Zone, while the youngest deposits represent the upper Famennian Dasbergina trigonica Zone (Szulczewski et al. 1996; Dzik 2006). The D. trigonica Zone roughly corresponds to Ziegler and Sandberg’s (1984) Siphonodella praesulcata Zone (cf. Szulczewski et al. 1996; Dzik 2006). The materials studied herein come from cephalopod limestone beds which were formed in neritic environment during the pelagic stage of deposition (Szulczewski 1978; Szulczewski et al. 1996).
Material and methods
The upper Famennian (Devonian) chaetognath spines and conodont elements are derived from archival rock samples collected by Michał Szulczewski (Faculty of Geology, University of Warsaw, Poland) from the uppermost part of dark grey cephalopod limestone beds (sample Ost-5) of the northern wall of the Ostrówka Quarry (latitude 50.843667, longitude 20.400528). Several chaetognaths grasping spines of Phakeloides polonicus gen. et sp. nov. and co-occurring conodont elements belonging to species of Dasbergina, Palmatolepis, Polygnathus, and Polynodosus were studied for the biostratigraphy, macro- and microstructure, cathodoluminescence and chemical composition (Table 1). All specimens were separated from carbonate rocks by dissolving in 10% acetic acid, washing, drying, and picking out of the residue under a binocular microscope by Hubert Szaniawski (ZPAL). Most of the recovered specimens exhibit excellent state of preservation and different ontogenetic growth stages.
Scanning electron microscope (SEM) and cathodoluminescence imaging were conducted at the Institute of Paleobiology, Polish Academy of Sciences in Warsaw, Poland. The architecture and microstructure of specimens were investigated using Philips XL-20 SEM and Thermo Fisher Scientific Quattro S ESEM devices after coating the whole or fragmented specimens with platinum. Thin sections made from chaetognath spines and conodont elements were studied using a petrographic microscope (Zeiss, Axio Scope A1) and, after coating with carbon, using a hot cathodoluminescence microscope (HC1-LM) with a beam of 14 keV and the current density of 0.1 μA/mm.
The electron microprobe studies were conducted at the Micro-area Analysis Laboratory of the Polish Geological Institute-National Research Institute in Warsaw, Poland. Concentrations of Ca, P, F, S, Sr, Fe, Mn, Ba, Si, Ti, Na, K, Mg, Cl, and Ce in thin sections made from different morphological parts of chaetognath grasping spines and conodont elements were measured by means of wavelength-dispersive spectrometers (WDS) of the Cameca SX100 electron microprobe analyser. The analyses were carried out using a 15 kV acceleration voltage, a 20 nA beam current, and a beam diameter of 5 μm. Natural minerals provided by Cameca were used as analytical standards. Average detection limits of the WDS analyses were as follows: Ca (299 ppm), P (202 ppm), F (1304 ppm), S (184 ppm), Sr (999 ppm), Fe (666 ppm), Mn (653 ppm), Ba (1466 ppm), Si (117 ppm), Ti (696 ppm), Na (194 ppm), K (181 ppm), Mg (118 ppm), Cl (170 ppm), Ce (1108 ppm). Mn, Ba, Ti, K, Mg and Ce elements were excluded from the interpretation as their detection limits were close to their measured concentrations. To show mineral density of different structural zones of the fossils backscattered electron (BSE) analysis was conducted. All figured specimens are deposited at the Institute of Paleobiology, Polish Academy of Sciences (ZPAL).
Systematic palaeontology
Phylum Chaetognatha Leuckart, 1854
Class, Order, and Family uncertain
Genus Phakeloides nov.
Zoobank LCID: urn:lsid:zoobank.org:act:A9F30FE6-A721-4FF8-99 CE-26BC533F154A
Type species: Phakeloides polonicus sp. nov.; see below.
Etymology: A combination of the generic name Phakelodus and the suffix -oides, implying likeness and referring to the similarity of this new genus to Cambrian chaetognath grasping spines of Phakelodus. Gender masculine.
Diagnosis.—As for species, by monotypy.
Phakeloides polonicus sp. nov.
Fig. 2A–C.
Zoobank LCID: urn:lsid:zoobank.org:act:1E251F23-2036-4AEF-8E 1F-16129CBD8C32
Etymology: From Latin Polonia, name of Poland, the country from which the specimen was collected.
Type material: Holotype, ZPAL Cg. 2/Ost-Ch.22 (Fig. 2A), well-preserved complete grasping spine. Paratypes: ZPAL Cg. 2/Ost-Ch.7, ZPAL Cg. 2/Ost-Ch.5 (Fig. 2B, C), almost complete grasping spines from the type locality and horizon.
Type locality: Ostrówka Quarry close to the Todowa Grząba peak, Gałęzice region, Holy Cross Mountains, central Poland.
Type horizon: Lagovignathus styriacus Zone, upper Famennian, Upper Devonian.
Diagnosis.—Small, gently curved hollow and round spines (approximate length 0.9–1.3 mm and width 0.1–0.2 mm); internal cavity long. The spine walls consisting of three layers: (i) the remainders of a thin outer layer which are fragmentarily preserved at the bottom of the spines, (ii) a thick middle layer composed of massive mineral structure with locally preserved fibril pattern and (iii) a thin, inner, microcrystalline layer. Furrows on the surface of the middle layer arranged in intersecting strips forming a mosaic-like or reticular ornamentation, most pronounced at the tip and in the central region of the spines. Ornamentation disappearing towards the spine’s base; individual furrows running obliquely around the spine. The internal cavity terminating, in some specimens, with open holes at the spine’s tips.
Remarks.—The upper Famennian (Devonian) Phakeloides polonicus gen. et sp. nov. is similar in architecture to the upper Cambrian and Early Ordovician species of the genus Phakelodus but is relatively round and devoid of longitudinal ornamentation (e.g., Müller 1959; Zhang in An et al. 1983; Szaniawski 2002; Wierzbowski et al. 2021). The characteristic feature of the Phakeloides polonicus gen. et sp. nov., readily distinguishing it from all previously named Palaeozoic chaetognath grasping spines, is reticular, surface ornamentation consisting of obliquely crossing furrows. In contrast, species of Phakelodus have longitudinal ridges and narrow oblique furrows that mark growth lines on the surface of grasping spines (see Szaniawski 2002: fig. 3A). The function of holes at the tips of some spines (Fig. 2B2) is not clear but we cannot rule out that cracking or chipping of the spine tip may have exposed its internal cavity.
The similarity in architecture of the grasping spines in species of Phakelodus and those of the Recent species of Sagitta chaetognaths concerns several major features of their morphology mentioned by Szaniawski (2002).
Stratigraphic and geographic range.—Type horizon and locality only.
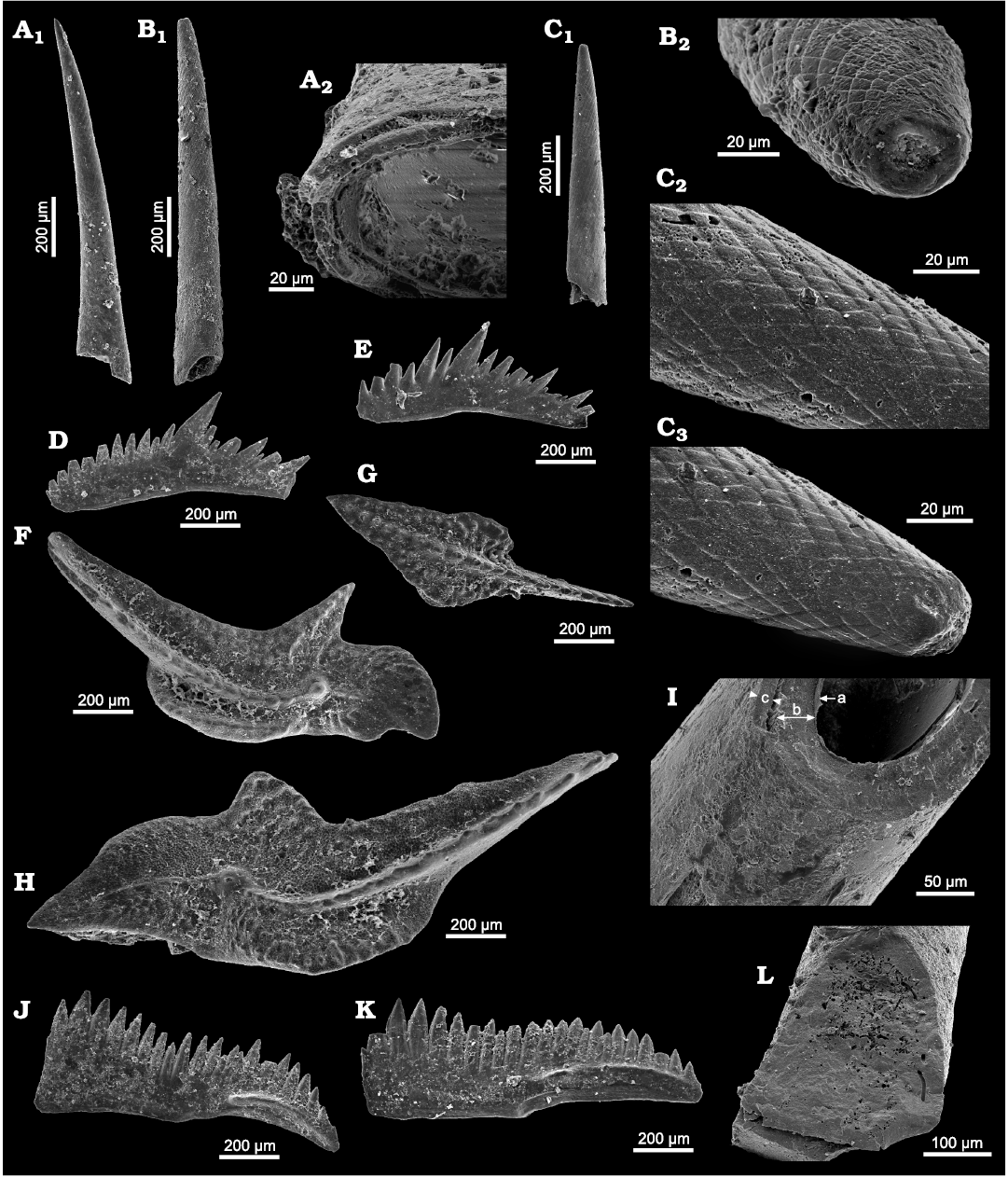
Fig. 2. Devonian chaetognath grasping spines Phakeloides polonicus gen. et sp. nov. (A–C, I, L) and conodonts (D–H, J, K) from the upper Famennian, Ostrówka Quarry, Holy Cross Mountains, Poland. A. ZPAL Cg. 2/Ost-Ch.22, A1, almost complete spines; A2, three layers in the wall observed at the base of the spines. B. ZPAL Cg. 2/Ost-Ch.5, B1, almost complete spines; B2, the hole at the tip of the spine. C. ZPAL Cg. 2/Ost-Ch.7, C1, almost complete spines; C2, C3, intersecting furrows create mosaic-like, reticular ornamentation of the surface of the middle layer close to the spine’s tip. D, E. Polygnathus sp., P2 element. D. ZPAL Cg. 2/Ost-C.14. E. ZPAL Cg. 2/Ost-C.26. F. Palmatolepis trachytera Ziegler, 1960, from the Lagovignathus styriacus Zone, ZPAL Cg. 2/Ost-C.25, P1 element. G. Idiognathodontid Dasbergina granulosa (Ziegler, 1962) from the Lagovignathus styriacus Zone, ZPAL Cg. 2/Ost-C.4, P1 element. H. Palmatolepis schindewolfi Müller, 1956, from the Lagovignathus styriacus Zone, ZPAL Cg. 2/Ost-C.15, P1 element. I. ZPAL Cg. 2/Ost-Ch.10, three layers in the wall: a thin, most internal, layer (a), the middle layer (b), and the outer layer (c). J, K. Platform-bearing elements of Polynodosus lauriformis (Dreesen and Dusar, 1974), from the Lagovignathus styriacus Zone, P1 element. J. ZPAL Cg. 2/Ost-C.13. K. ZPAL Cg. 2/Ost-C.21. L. ZPAL Cg. 2/Ost-Ch.9, irregular boring on the surface and inside the middle and inner layers.
Results
Petrographic microscope and scanning electron microscope studies.—The petrographic microscope studies have revealed the existence of a fragmentary, very thin, dark brown, opaque cortex of the Devonian Phakeloides polonicus gen. et. sp. nov. spines and mostly translucent, patchy light brown to dark brown inner area, which predominantly represents the bulk middle layer (Fig. 3A; cf. Wierzbowski et al. 2021). A thin, internal layer is often visible on the surface of internal cavity of the spines, which is similar to Cambrian chaetognath grasping spines (cf. Szaniawski 1983, 2002). The phosphate crystals of all zones of the studied spines are arranged in the same optical orientation and show undulose extinction pattern. The internal cavity of the spines is mostly empty, albeit clotted, dark brown, diagenetic phosphate infillings occur in some places.
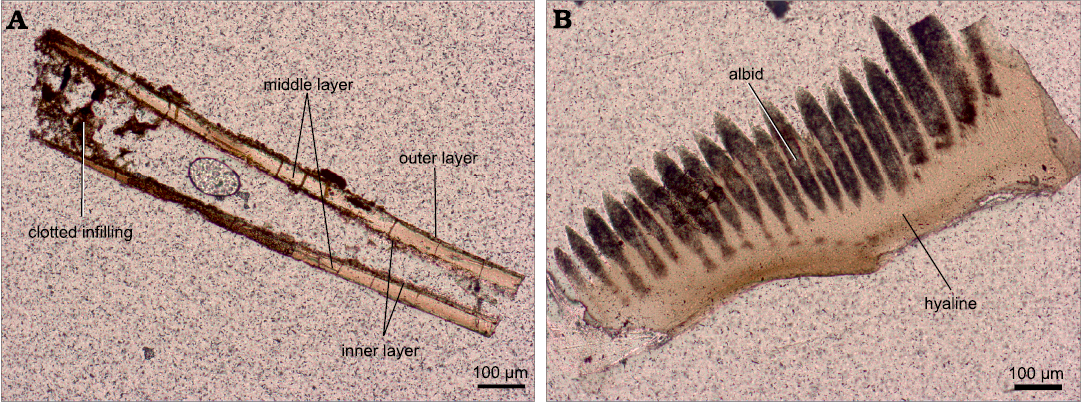
Fig. 3. Transmitted light images of thin sections of Devonian chaetognath spine and conodont element from the upper Famennian, Ostrówka Quarry, Holy Cross Mountains, Poland. A. Longitudinal section of Phakeloides polonicus gen. et sp. nov. grasping spine showing a very thin and partially preserved, dark brown, opaque cortex; a mostly translucent, light brown to dark brown, in patches, inner area representing the bulk middle layer and a thin, internal layer fragmentarily visible on the surface of internal cavity of the spine, ZPAL Cg. 2/Ost-Ch.30. B. Longitudinal section of Polynodosus sp. conodont element showing the top, brown and opaque albid (white matter) tissue and the underlying, translucent hyaline (lamellar) tissue, ZPAL Cg. 2/Ost-C.35.
The studied conodont elements belonging to species of Dasbergina, Palmatolepis, Polygnathus, and Polynodosus show pale yellow to light brown colours, which points to their low thermal alteration demonstrated by CAI (conodont colour-alteration index) of 1 (cf. Epstein et al. 1977; Rejebian et al. 1987). The conodont elements consist of tissues distinguishable in transmitted light zones: the top, brown and opaque albid (white matter) tissue, the translucent hyaline (lamellar) tissue (Fig. 3B) and, in some cases, remainders of dark brown, opaque and porous basal body of disordered mineral structure (cf. Wright 1989; Trotter et al. 2007; Zhuravlev and Gerasimova 2015; Zhuravlev 2017).
Scanning microscope observations do not point to the presence of a distinct outer layer in the majority of studied Devonian Phakeloides polonicus gen. et sp. nov. grasping spines, which contrasts to Furongian (Cambrian) grasping spines of Phakelodus sp. (cf. Szaniawski 1982, 1983, 2002; Wierzbowski et al. 2021). A thin, cylindrical, outer zone of microcrystalline fabric, which likely represents fragments of the outer layer, is, however, visible in some specimens, especially at the base of the spines (Fig. 4D2). The major portion of the spines, belonging structurally to the middle layer, consists of fused granular grains or a massive mineral structure characterized by the perpendicular, uneven or conchoidal fracture with minor breaks forming the second-order cleavage plane (Fig. 4A, B). The surface of the spines reveals a reticular or mosaic-like texture consisting of obliquely crossing furrows, which likely represents primary ornamentation of the middle layer (Fig. 4A, C, D2). Parallel fibrils of the middle layer, arranged obliquely to the spine axis, which were reported from other fossil and modern chaetognath spines, are visible locally on broken surfaces of studied specimens of Phakeloides polonicus gen. et sp. nov. (Fig. 4C; cf. Szaniawski 1982, 1983, 2002; Doguzhaeva et al. 2002; Vannier et al. 2007).

Fig. 4. Scanning electron microscope images of Devonian chaetognath grasping spines Phakeloides polonicus gen. et sp. nov. (A–D) and conodont elements (E) from the upper Famennian, Ostrówka Quarry, Holy Cross Mountains, Poland. A. ZPAL Cg. 2/Ost-Ch.31, cross-section throughout the spine showing massive mineral structure of the middle layer characterized by the conchoidal fracture, a reticular ornamentation of the surface of the middle layer is partially visible. B. ZPAL Cg. 2/Ost-Ch.32, longitudinal section throughout the spine showing a massive mineral structure characterized by uneven fracture, which is superimposed on minor breaks forming the second-order cleavage plane. A thin, cylindrical internal layer composed of fine-grained calcium phosphate crystals is visible in the innermost portion of the spine wall, around the internal cavity. C. ZPAL Cg. 2/Ost-Ch.33, a surface of the middle layer of the spine showing oblique to the spine axis, primary fibrils, visible at its slightly, broken part, and less noticeable surface striations of the reticular ornamentation. D. ZPAL Cg. 2/Ost-Ch.34, D1, cross-section throughout the spine showing porous internal zones of the middle layer, which are divided by solid cylindrical zones, a thin, cylindrical internal layer is visible in the innermost portion of the spine wall; D2, three layers in the wall of the basal portion of the spine: a thin, most internal, layer (a), the middle layer (b), and the outer layer (c). E. ZPAL Cg. 2/Ost-C.38, cross-section throughout Polynodosus sp. conodont element showing innermost pillar-like structure of densely packed, weakly lamellar, albid (white matter) tissue, and outer hyaline tissue composed of oblique apatite lammelae. The outermost portion of the conodont element consists of paralamellar variety of the hyaline tissue of more chaotic structure.
The innermost portion of the spines, around the internal cavity, usually shows the presence of a thin, cylindrical internal layer of microcrystalline fabric composed of granular calcium phosphate crystals (Fig. 4B, D1). Pitting or significant internal porosity is observed within many parts of the studied Phakeloides polonicus gen. et sp. nov. spines. In some places, in the cross-section of the spine walls are visible narrow porous zones dividing solid well-mineralized zones (Fig. 4D1). Networks of small borings are often observed in cross sections of the spines (Fig. 4A).
In contrast, studied conodont elements consist of densely packed, weakly lamellar, albid (white matter) tissue, which is typical of the crown tips and innermost parts of conodont elements composed of the strongly lamellar, hyaline tissue (Fig. 4E). The oblique lamellae of the hyaline tissue of lower parts of conodont elements are very characteristic. They pass into more chaotic structure of lateral parts of the conodont platform, which may be assigned to the paralamellar variety of the lamellar tissue (Fig. 4E; cf. Zhuravlev 2017).
Cathodoluminescence imaging.—Phakeloides polonicus gen. et sp. nov. spines show weak to moderate red cathodoluminescence, excluding narrow fractures and fragments of outermost rim areas which show bright luminescence colours (Fig. 5A). In conodont elements, very weak yellow-red luminescence is observed in albid tissue, whereas the bulk of hyaline tissue and remnants of basal body are non-luminescent (Fig. 5B).
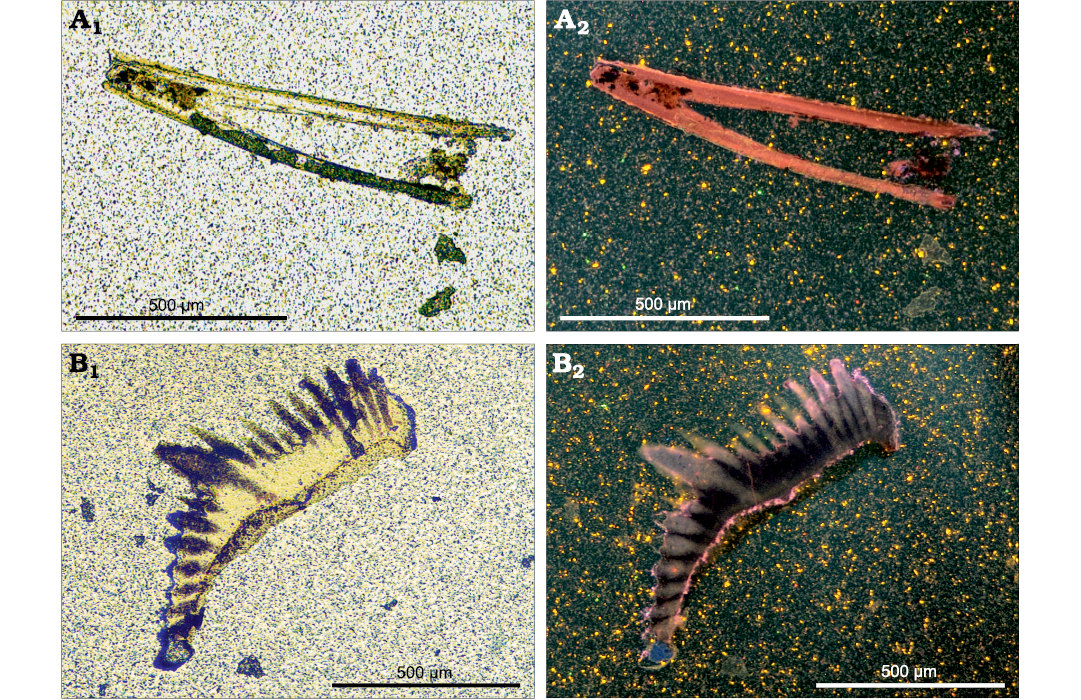
Fig. 5. Transmitted light (A1, B1) and hot cathodoluminescence images (A2, B2) of Devonian Phakeloides polonicus gen. et sp. nov. grasping spine (A, ZPAL Cg. 2/Ost-Ch.41) and Polygnathus sp. conodont element (B, ZPAL Cg. 2/Ost-C.45) from the upper Famennian, Ostrówka Quarry, Holy Cross Mountains, Poland. A2, weak to moderate red cathodoluminescence of the grasping spine and bright luminescence of some parts of its outermost rim area. B2, very weak yellow-red luminescence of conodont albid tissue and non-luminescent hyaline tissue.
Electron microprobe and backscattered electron studies.—Electron microprobe studies have revealed low calcium and phosphorus concentrations in the middle layer of the studied Devonian Phakeloides polonicus gen. et. sp. nov. spines (Ca ~31.8 wt.%, P ~13.4 wt.%), which are much lower than concentrations of the same elements in co-occurring conodont elements (albid crown: Ca ~37.8 wt.%, P ~18.3 wt.%; hyaline crown: Ca ~38.6 wt.%, P ~18.0), except for their basal body (Ca ~33.9 wt. %, P ~14,4 wt.%; cf. Fig. 6A, B). It should be noted that relatively low calcium and phosphorus concentrations were previously measured from the middle layer and internal cavity infilling of Cambrian, Phakelodus sp. spines from Sweden, unlike to higher concentrations of the same elements in their outer layer (Wierzbowski et al. 2021).
Fluoride concentrations are relatively high in all studied Devonian fossils (middle layer of Phakeloides polonicus gen. et. sp. nov. spines: ~3.6 wt.%; conodont albid crown: ~4.2 wt.%; conodont hyaline crown: ~4.2 wt.%; conodont basal body: ~4.1 wt.%; Fig. 6C). Comparable fluoride concentrations were previously reported from the middle layer and cavity infilling of Cambrian, Phakelodus sp. spines from Sweden (Wierzbowski et al. 2021). Much lower fluoride content (~2.4 wt.%) was measured from the outer layer of the latter fossils (Wierzbowski et al. 2021).
High sulphur and iron concentrations are observed in the middle layer of Devonian Phakeloides polonicus gen. et. sp. nov. spines (S ~0.59 wt.%, Fe ~0.24 wt.%) and basal body of coeval conodont elements (S ~0.49 wt.%; Fe ~0.16 wt.%; Fig. 6D, E). Albid and hyaline tissues of conodont elements show much lower concentrations of these elements (albid crown: S ~0.05 wt.%; Fe <0.07 wt.%; hyaline crown: S ~0.06 wt.%; Fe <0.07 wt.%). It is worth noting that elevated sulphur and iron concentrations were previously found in all parts of Cambrian, Phakelodus sp. spines from Sweden (Wierzbowski et al. 2021).
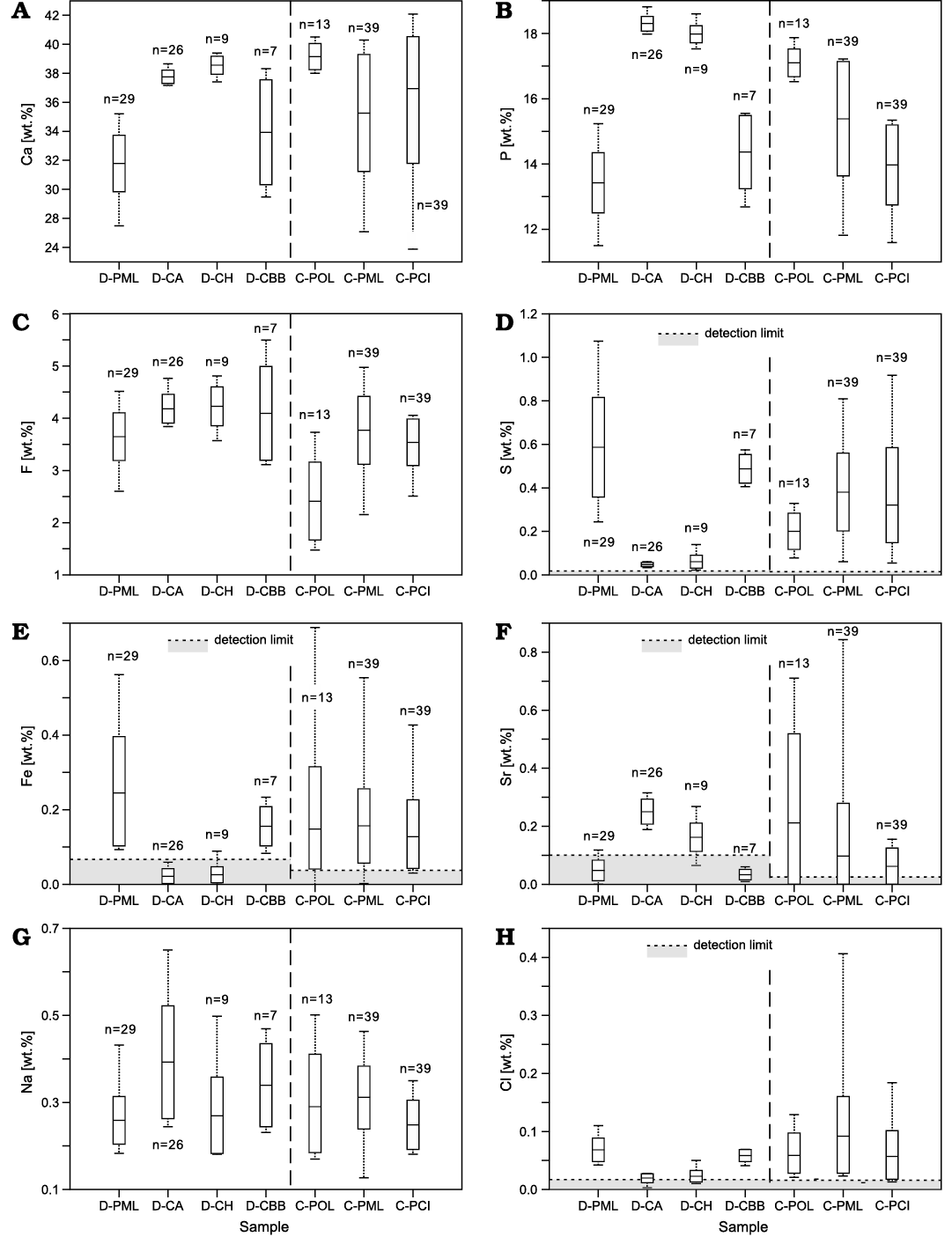
Fig. 6. Concentrations of Ca, P, F, S, Fe, Sr, Na and Cl (A–H) in different structural parts of studied Devonian (D-) conodont elements from the upper Famennian, Ostrówka Quarry, Holy Cross Mountains, Poland (new data) as well as previously studied Cambrian (C-) Phakelodus sp. spines from the Furongian of Sweden (from Wierzbowski et al. 2021). D-PML, Phakeloides polonicus gen. et sp. nov. middle layer; D-CA, Polygnathus sp. conodont albid; D-CH, Polygnathus sp. conodont hyaline; D-CBB, Polygnathus sp. conodont basal body; C-POL, Phakelodus sp. outer layer; C-PML, Phakelodus sp. middle layer; C-PCI, Phakelodus sp. cavity infilling. Dotted lines, a range of values; rectangles, 1 standard deviation limits; black horizontal bars, average elemental content.
Studied Devonian, Phakeloides polonicus gen. et. sp. nov. spines and basal body of conodont elements are characterized by much lower strontium content (<0.1 wt.%) than conodont albid (~0.25 wt.%) and hyaline tissues (0.16 wt.%; Fig. 6F). Relatively low strontium concentrations were also found in middle layer and internal cavity infilling of Cambrian, Phakelodus sp. spines from Sweden (Wierzbowski et al. 2021). Distinct differences in sodium and chlorine concentrations of various parts of chaetognath spines and conodont elements are not observed, except for slight chlorine depletion in conodont albid and hyaline tissues (Fig. 6).
BSE images have shown that the studied Devonian Phakeloides polonicus gen. et sp. nov. spines consist of apatite phases of similar density, which form the bulk middle and inner layers (Fig. 7). A thin cortex of slightly higher density, which may represent remainders of outer layer, is fragmentarily preserved and occurs in some specimens only (Fig. 7B). It was too thin to be studied for chemical composition using the electron microprobe. The same applies to minute grains of higher density visible in inner parts of the spine walls, which probably represent pyrite microcrystals (Fig. 7).
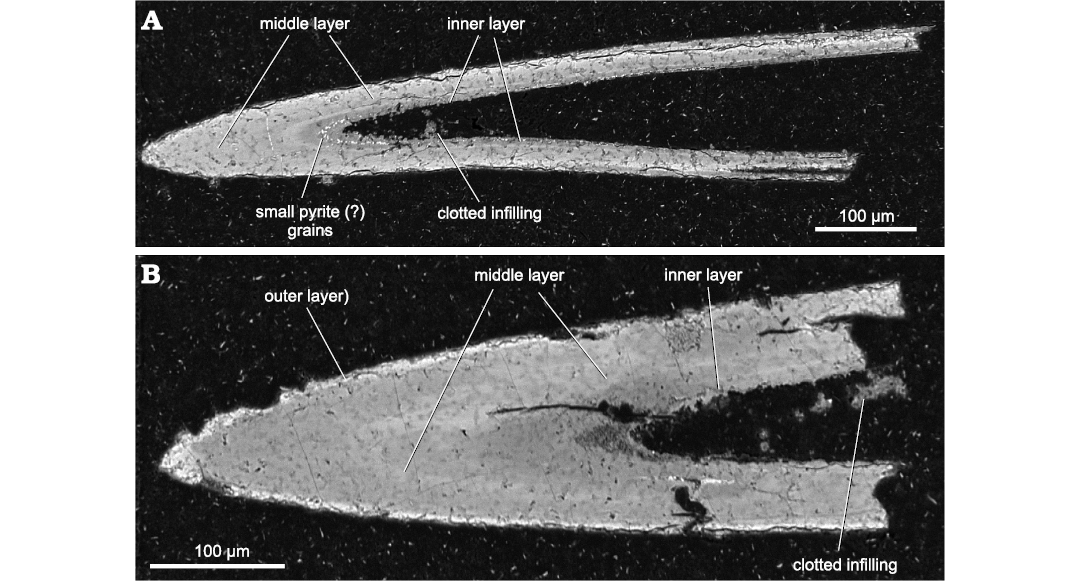
Fig. 7. Back scattered electron images of thin section made of Devonian Phakeloides polonicus gen. et sp. nov. spines from the upper Famennian, Ostrówka Quarry, Holy Cross Mountains, Poland. A. Longitudinal section of the spine showing the apatite phases of similar density of the middle and inner layers, ZPAL Cg. 2/Ost-Ch.48. B. Longitudinal section of the spine showing the apatite phases of similar density of the middle and inner layers as well as a thin cortex of slightly higher density, which may represent fragmentarily preserved outer layer, ZPAL Cg. 2/Ost-Ch.49.
Comparison of the structure of the spines of Devonian Phakeloides polonicus gen. et sp. nov., Cambrian Phakelodus sp. and Rhombocorniculum cancellatum.—The investigation of Devonian Phakeloides polonicus gen. et sp. nov. and Cambrian spines of Phakelodus sp. has shown significant similarity of these fossils in terms of their shape and morphology. They include a similar hollow and approximately round shape, layered internal structure, and the same manner of ontogenetic growth of individual elements into basal direction. Both Devonian Phakeloides polonicus gen. et sp. nov and Cambrian Phakelodus sp. chaetognath grasping spines are gently curved. Their wall structure likely consisted of a thin, outer, and an apparently homogenous, comparatively thick and fibrous middle layer as wells as a thin inner, laminated layer. The outer layer of Devonian Phakeloides polonicus gen. et sp. nov. is, however, fragmentarily preserved, mostly in basal areas of the spines (Fig. 2A), while relatively intact middle and inner layers of the same fossils extend around a large pulp cavity from the base to the tip of the spines. Both groups of spines show, in many places, a fibrous structure of the middle layer (Fig. 4C and Szaniawski 2002: fig. 4B).
Grasping spines of Cambrian Phakelodus sp. have oval or trapezoidal cross-section (Szaniawski 1983, 2002), whereas Devonian Phakeloides polonicus gen. et sp. nov. spines are more circular. The surfaces of both Cambrian Phakelodus sp. and the Devonian Phakeloides polonicus gen. et sp. nov. spines are ornamented but their details are different. The surface of Phakelodus sp. grasping spines is ornamented by longitudinal ridges and narrow oblique furrows (Szaniawski 1983, 2002: fig. 1). In contrast, the Phakeloides polonicus gen. et sp. nov. spines show a mosaic-like, rhomboidal ornamentation consisting of obliquely crossing shallow furrows (Fig. 2A–C).
It is worth noting that ornamentation of the Devonian Phakeloides polonicus gen. et sp. nov. spines greatly resembles the reticular, rhomboidal sculpture developed on the surface of Cambrian (Series 2 and 3) spines of Rhombocorniculum cancellatum interpreted presently as sclerites of a Hallucigenia-like lobopodian worm (Caron et al. 2013). The Rhombocorniculum spines despite mostly elongated, gently curved shape, the occurrence of dextral and sinistral forms, internal cavity and three structural layers differ from the phosphatized chaetognath grasping spines due to the presence of thicker spine walls and mostly coaxial, internal hollow tubules in the internal structure of the middle layer (Guaxiong et al. 2003), although their fibrous wall structure was also reported (Landing et al. 1980). In addition, the convex shape of rhomboidal sculpture of Rhombocorniculum spines (cf. Brasier, 1986: fig. 5, 1989: pl. 7.2; Landing 1995: fig. 5; Guaxiong et al. 2003: fig. 1; Caron et al. 2013: figs. 1, 2; Kouchinsky et al. 2015: figs. 49–52), which actually consists of inclined triangular scales raised above the smooth surface of the spines (Caron et al. 2013), differs from concave rhomboidal surface furrows of the studied Upper Devonian Phakeloides polonicus gen. et sp. nov. spines.
Irregular boring networks composed of minute meandering and branching tunnels, which are visible on the surface and inside of the middle and inner layers of studied chaetognath spines (Figs. 2I, 4A) can be regarded as secondary phenomena. These burrows in the phosphatized spines might have been formed by some food-seeking benthic organisms during short periods of their excavation.
Discussion
Biostratigraphy.—The stratigraphy and sedimentology of the Famennian limestone beds from the Ostrówka Quarry is documented by Szulczewski (1971), Szulczewski et al. (1996), and Dzik (2006). The studied samples have yielded a conodont assemblage with dominant Dasbergina granulose, Palmatolepis schindewolfi, Palmatolepis trachytera, Polynodosus lauriformis, and Polygnathus sp., which suggests that all studied chaetognath spines and conodonts are derived from the Lagovignathus styriacus Zone of the upper Famennian of the Upper Devonian (cf. Dzik 2006: fig. 3). All the previously unidentified small chaetognath grasping spines have been assigned to a new species, Phakeloides polonicus gen. et sp. nov.
Microstructure, cathodoluminescence, and chemical composition of Devonian Phakeloides polonicus gen. et sp. nov. grasping spines.—Weak to moderate red cathodoluminescence of the bulk interior of the studied Devonian Phakeloides polonicus gen. et sp. nov. spines may be related to remarkable incorporation of manganese and rare earth element (REE) into apatite crystal lattice during diagenetic processes (Fig. 5; Ségalen et al. 2008; Fischer et al. 2012, 2013; Wierzbowski 2021). In addition, low calcium and phosphorus contents, as well as elevated sulphur, iron, and fluoride concentrations of the middle layer of the Phakeloides polonicus gen. et sp. nov. spines (Fig. 6A–E) should be regarded, respectively, as a result of (i) its poorly mineralized, porous structure and/or the presence of significant amount of organic matter (cf. Szaniawski 1982, 1983, 2002) and (ii) the effect of supplying of certain chemical elements by pore waters in post-depositional processes (cf. Bocherens et al. 1994; Kohn et al. 1999; Wierzbowski 2021; Wierzbowski et al. 2021). This points to the re-crystallization or the secondary origin of phosphate mineral phases of the studied chaetognath spines. Their phosphatization, associated with pyrite incrustation, likely occurred in reducing pore waters, characterized by the abundance of dissolved manganese and iron and elevated REE concentrations (cf. Marshall 1992; Haley et al. 2004).
The diagenetic origin of chaetognath apatite may be substantiated by the frequent lack of the distinct incremental microstructure of the spines. The spines predominantly consist of randomly oriented, fused apatite microcrystals with pseudolamination emphasised by zones of higher porosity (Fig. 4B, D1). Such lamination pattern may arise because of the secondary phosphatization of different, morphological parts of previously organic (chitinous) spines, rather than their original biomineralization. Arranged obliquely to the spine axis and closely packed fibrils of the middle layer, which are distinctive of modern and fossil spines (cf. Szaniawski 1982, 1983, 2002; Doguzhaeva et al. 2002; Vannier et al. 2007), are observed in certain fragments of the studied specimens only (Fig. 4C). It indicates that the phosphatization process has only partially conserved the original microstructure of the spines. Minor internal microbreaks (cleavage) of the spines (Fig. 4A, B) shows the fragility of diagenetic apatite crystals.
The diagenetic phosphate mineralization was likely a selective process as co-occurring albid and hyaline tissues of conodont elements, which have common taphonomic history with the studied chaetognath spines, do not show any signs of re-crystallization. Conodont apatite is non-luminescent or shows very weak cathodoluminescence intensity and low concentrations of secondary sulphur and iron (Figs. 5, 6). In addition, high strontium concentrations in conodont albid and hyaline may be linked to their chemical equilibration with seawater. It is also worth noting that low value of CAI contradicts significant thermal alteration of the upper Famennian (Devonian) deposits of the Holy Cross Mountains in central Poland. Therefore, such process could not have been responsible for the pervasive re-crystallization of phosphate components of the studied rocks.
A thin, outer layer of Cambrian Phakelodus sp. spines from Sweden characterized by a dense mineral structure, low fluoride and elevated strontium concentration, which has been assigned to the early marine “Orsten” type phosphatization (cf. Wierzbowski et al. 2021), is mostly lacking in the studied Devonian Phakeloides polonicus gen. et sp. nov. The formation of the bulk middle layer of the Cambrian Phakelodus spines is, however, related to later stages of diagenesis similarly to the middle layer of the Devonian Phakeloides polonicus gen. sp. nov. spines (cf. Wierzbowski et al. 2021). This points to a difference in the pathways of phosphatization processes between Cambrian and Devonian chaetognath grasping spines, the latter of which did not undergo very early “Orsten”-like apatite impregnation of their cuticle layer.
The “Orsten” type phosphatization of small invertebrate organisms leading to preservation of their three-dimensional fossils with minor structural details and, at least, some phosphatized chaetognath spines is mostly known from the Cambrian–Lower Ordovician (cf. Müller 1979; Andres 1989; Müller and Walossek 1985; Maas and Waloszek 2001; Waloszek 2003; Maas et al. 2006; Eriksson et al. 2012, 2016; Shen et al. 2013; Wierzbowski et al. 2021; Olempska et al. 2023). It is assigned to the widespread phosphate mineralization in the earliest Palaeozoic sediments due to high load and concentrations of phosphorus in pore waters, varying redox conditions and low ventilation of deposits, before the Ordovician diversification of burrowing animals (Shen et al. 2000; Dornbos 2011; Creveling et al. 2014; Pruss et al. 2018). Less common findings of phosphatized small shelly fossils, including one discovery of chaetognath spines, are reported from the upper Palaeozoic, Mesozoic or Cenozoic deposits of various parts of the world (Dzik 1994; Duncan et al. 1998; Walossek 1999; Doguzhaeva et al. 2002; Dattilo et al. 2016; Pruss et al. 2018). It is, unfortunately, unclear if the reported, younger phosphatized fossils show any differences in timing and the pathway of diagenetic mineralization processes, which might be comparable to differences noted between the previously studied Cambrian Phakelodus sp. spines and the newly investigated Devonian Phakeloides polonicus gen. et sp. nov. material.
Specific structural features of Devonian Phakeloides polonicus gen. et sp. nov. grasping spines.—A removal of the cuticle layer of the studied chaetognath spines may have unveiled the concave reticular ornamentation of the surface of the middle layer (Fig. 4D2), which is not visible in other fossil or modern chaetognath spines with preserved outer layer. It is also possible that the mosaic-like ornamentation pattern may be a characteristic feature of some late Palaeozoic chaetognath grasping spines being not present in their fossil and modern counterparts.
The rhomboidal ornamentation of the Devonian, Phakeloides polonicus gen. et sp. nov. chaetognath grasping spines may also point to their affinity to problematic spines of the Cambrian Rhombocorniculum cancellatum. In fact, one of the major structural difference of Rhombocorniculum cancellatum spines, compared to those of Phakeloides sp., i.e., their porous internal structure and the presence of internal hollow tubules, parallel to the longitudal axis of the spines (cf. Guoxiang et al. 2003; Kouchinsky et al. 2015), can be a diagenetic feature related to incomplete apatite mineralization of the middle layer of originally organic spines. Although the middle layer of modern chaetognath spines is described as consisting of obliquely arranged fibrils or fiber bundles (cf. Szaniawski 1983; Bone et al. 1983), those structures are often arranged nearly parallel to the spine axis (see Szaniawski 2002: fig. 5; Doguzhaeva et al. 2002: fig. 6C). It is therefore possible that partial phosphatization of original, thicker and less tightly packed fiber structure of the middle layer of some Cambrian chaetognath spines might have produced hollow tubules and spongy mineral network similar to that of Rhombocorniculum cancellatum. On the other hand, the convex shape of rhomboidal sculpture of Rhombocorniculum cancellatum clearly differs from concave rhomboidal ornamentation of the studied Devonian, Phakeloides polonicus gen. et sp. nov. spines. Therefore, the resemblance of internal structures of both fossil groups may be related to convergent evolution. It is also possible that some Cambrian chaetognath spines of rhomboidal ornamentation are wrongly classified as belonging to Rhombocorniculum.
Another feature of the Devonian Phakeloides polonicus gen. et sp. nov. spines from the Holy Cross Mountains of central Poland is their approximately round shape, devoid of any signs of flattening and the lack of longitudinal lamination. They are different from the majority of more oval or trapezoidal fossil or modern spines showing external ridges or lamination (cf. Bone et al. 1983; cf. Doguzhaeva et al. 2002; Moreno and Kapp 2003; Szaniawski 2002, 2015). The lack of such structures can hardly be linked to the removal of the outer layer of the studied spines. In addition, the lack of flattening of the studied Devonian chaetognath grasping spines contradicts their deep burial before phosphatization (contrary to flattening of the Cambrian carbonaceous spines; Slater et al. 2018; Slater and Willman 2019). This may point to relatively long residence time of these fossils in the upper part of sediment column, in the oxygen-depleted environment, under fluctuating redox conditions and significant flux of phosphorus, which might have led to their phosphatization. The condensed Famennian limestone beds exposed in the Ostrówka Quarry show mixed lithology and represent a transition between the crinoidal and cephalopod limestone beds. The crinoidal limestone beds are distinctive of an earlier phase of drowning of the shallow-water carbonate platform, whereas the cephalopod limestone, from which the studied material is derived, represents a later, pelagic stage of deposition (e.g., Szulczewski et al. 1996). The condensed nature of these beds may have allowed accumulation of organic skeletal elements close to the sediment-water interface and their subsequent phosphatization.
Conclusions
Devonian chaetognath grasping spines from the upper Famennian of the Holy Cross Mountains in central Poland have been assigned to the newly established Phakeloides polonicus gen. et sp. nov. species. It is known from an isolated occurrence in upper Palaeozoic rocks of the Holy Cross Mountains in central Poland and shows characteristic mosaic-like ornamentation of the surface.
The studied Devonian Phakeloides polonicus gen. et sp. nov. chaetognath grasping spines are built of diagenetic phosphate phases and possess poorly mineralized, porous mineral structure. It is demonstrated by moderate cathodoluminescence intensity, high concentrations of diagenetic sulphur, iron and fluoride, low concentrations of strontium as well as decreased contents of major elements including calcium and phosphorus. Phakeloides polonicus gen. et sp. nov. spines mostly consist of secondary, fairly massive, mineral structure of the middle layer, which has preserved major structural features of the spines including primary shape and layered-architecture. Minor structural details of the spines like fibrils of the middle layer are visible only locally. In addition, the outer layer of the spines is fragmentarily preserved, which has revealed characteristic, mosaic-like ornamentation of the surface of their middle layer. The remainders of the outer layer are mostly found at the base of the spines.
The observed features of Phakeloides polonicus gen. et sp. nov. spines likely arose because of their slow, post-depositional phosphatization, close to the sediment-water interface, in condensed, transgressional Upper Devonian sequence. The Devonian, Phakeloides polonicus gen. et sp. nov. spines possess a round shape, without longitudal lamination, contrary to many lower Palaeozoic spines and are devoid of early diagenetic “Orsten” type mineralization of the outer, cuticle layer (cf. Wierzbowski et al. 2021).
Acknowledgements
Hubert Szaniawski (ZPAL) and Michał Szulczewski (Faculty of Geology, University of Warsaw, Poland) are thanked for supplying separated phosphatic chaetognath spines and conodont elements from the upper Famennian (Devonian) condensed limestone of the Ostrówka Quarry in central Poland. We warmly acknowledge Piotr Maciak (President of the Management Board in Nordkalk Company, Kraków, Poland) for the invaluable support and enable research in Ostrówka Quarry. Paweł Bącal (ZPAL) provided invaluable assistance in the SEM investigations. The study was supported by the Polish Geological Institute-National Research Institute statutory funds (Project No. 62.9012.2203.00.0). Reviewers, Stefan Bengtson (Department of Palaeobiology, Museum of Natural History, Sweden) and Jean Vannier (Laboratoire de Géologie de Lyon, Université de Lyon, France) are thanked for valuable reviews of earlier versions of the manuscript and suggested improvements.
References
Allison, P.A. and Briggs, D.E.G. 1993. Exceptional fossil record: Distribution of soft-tissue preservation through the Phanerozoic. Geology 21: 527–530. Crossref
An, T.−X., Zhang, F., Xiang, W., Zhang, Y., Xu, W., Zhang, H., Jiang, D., Yang, C., Lin, L., Cui, Z., and Yang, X. 1983. The Conodonts of North China and the Adjacent Regions. 223 pp. Science Press, Beijing.
Andres, D. 1988. Strukturen, Apparate und Phylogenie primitive Conodonten. Palaeontographica A 200: 105–152.
Andres, D. 1989. Phosphatisierte Fossilien aus dem unteren Ordoviz von Südschweden. Berliner Geowissenschaftliche Abhandlungen 106: 9–19.
Bengtson, S. 1976. The structure of some Middle Cambrain conodonts, and the early evolution of conodont structure and function. Lethaia 9: 185–206. Crossref
Bengtson, S. 1983. A functional model for the conodont apparatus. Lethaia 16: 38. Crossref
Berkowski, B. 2002. Famennian Rugosa and Heterocorallia from southern Poland. Palaeontologia Polonica 61: 3–88.
Bocherens, H., Brinkman, D.B., Dauphin, Y., and Mariotti, A. 1994. Microstructural and geochemical investigations on Late Cretaceous archosaur teeth from Alberta, Canada. Canadian Journal of Earth Sciences 31: 783–792. Crossref
Bone, Q., Ryan, K.P., and Pulsford, A.L. 1983. The structure and composition of the teeth and grasping spines of chaetognaths. Journal of the Marine Biological Association UK 63: 929–939. Crossref
Brasier, M.D. 1986. The succession of small shelly fossils (especially conoidal microfossils) from English Precambrian–Cambrian boundary beds. Geological Magazine 123: 237–256. Crossref
Brasier, M.D. 1989. Towards a biostratigraphy of the earliest skeletal biotas. In: J.W. Cowie and M.D. Brasier (eds.), The Precambrian–Cambrian Boundary, 117–165. Clarendon Press, Oxford.
Brasier, M.D. and Callow, R.H.T. 2007. Changes in the pattern of phosphatic preservation across the Proterozoic–Cambrian transition. Memoirs of the Association of Australasian Palaeontologists 34: 377–389.
Briggs, D.E.G. and Caron, J.-B. 2017. A large Cambrian chaetognath with supernumerary grasping spines. Current Biology 27: 2536–2543. Crossref
Caron, J.-B., Smith, M.R., and Harvey, T.H.P. 2013. Beyond the Burgess Shale: Cambrian microfossils track the rise and fall of hallucigeniid lobopodians. Proceedings of the Royal Society B: Biological Sciences 280: 20131613. Crossref
Carlorosi, J., Esteban, S.B., and Tortello, M.F. 2019. Early Ordovician conodonts from the Santa Rosita Formation at Pantipampa, Iruya area, northestern Argentina. Andean Geology 46: 168–182. Crossref
Creveling, J.R., Knoll, A.H., and Johnston, D.T. 2014. Taphonomy of Cambrian phosphatic small shelly fossils. Palaios 29: 295–308. Crossref
Dattilo, B.F., Freeman, R.L., Peters, W.S., Heimbrock, W.P., Deline, B., Martin, A.J., Kallmayer, J.W., Reeder, J., and Argast, A. 2016. Giants among micromorphs: were Cincinnatian (Ordovician, Katian) small shelly phosphatic faunas dwarfed? Palaios 31: 55–70. Crossref
Doguzhaeva, L.A., Mutvei, H., and Mapes, R.H. 2002: Chaetognath grasping spines from the Upper Mississipian of Arkansas (USA). Acta Palaeontologica Polonica 47: 421–430.
Dornbos, S.Q. 2011. Phosphatization through the Phanerozoic. In: P.A. Allison and P.A. Bottjer (eds.), Taphonomy Process and Bias through Time. Topics in Geobiology Volume 32, 435–456. Springer, Dordrecht. Crossref
Dreesen, R. and Dusar, M. 1974. Refinement of conodont-biozonation in the Famenne area. In: J.P. Bouckaert and M. Street (eds.), International Symposium on Belgian Micropalaeontological Limits, Namur 1974. Geological Survey of Belgium Publication 13: 1–38
Duncan, I.J., Briggs, D.E.G., and Archer, M. 1998. Three-dimensionally mineralized insects and millipedes from the Tertiary of Riversleigh, Queensland, Australia. Palaeontology 41: 835–851.
Dzik, J. 1994. Evolution of “small shelly fossils” assemblages of the Early Paleozoic. Acta Palaeontologica Polonica 39: 247–313.
Dzik, J. 2006. The Famennian “Golden Age” of conodonts and ammonoids in the Polish part of the Variscan sea. Palaeontologia Polonica 63: 1–359.
Epstein, A.G., Epstein, J.B., and Harris, L.D. 1977. Conodont Color Alteration—an Index to Organic Metamorphism. Geological Survey Professional Paper 995: 1–27. Crossref
Eriksson, M.E., Terflet, F., Elofsson, R., and Marone, F. 2012. Internal soft-tissue anatomy of Cambrian “Orsten” arthropods as revealed by synchrotron X-ray tomographic microscopy. Plos One 7: e42582. Crossref
Eriksson, M.E., Terflet, F., Elofsson, R., Maas, A., Marone, F., Lindskog, A., Waloszek, D., Schmitz, B., and Stampanoni, M. 2016. Baring it all: undressing Cambrian “Orsten” phosphatocopine crustaceans using synchrotron X-ray tomographic microscopy. Lethaia 49: 312–326. Crossref
Fischer, J., Schneider, J.W., Voigt, S., Joachimski, M.M., Tichomirowa, M., Tütken, T., Götze, J., and Berner, U. 2013. Oxygen and strontium isotopes from fossil shark teeth: environmental and ecological implications for late Palaeozoic European basins. Chemical Geology 342: 44–62. Crossref
Fischer, J., Voigt, S., Franz, M., Schneider, J.W., Joachimski, M.M., Tichomirowa, M., Götze, J., and Furrer, H. 2012. Palaeoenvironments of the late Triassic Rhaetian Sea: implications from oxygen and strontium isotopes of hybodont shark teeth. Palaeogeography, Palaeoclimatology, Palaeoecology 353–355: 60–72. Crossref
Guoxiang, L., Zhu, M., and Steiner, M. 2003. Microstructure and functional morphology of the Early Cambrian problematic fossil Rhombocorniculum. Progress in Natural Science 13: 831–835. Crossref
Haley, B.A., Klinkhammer, G.P., and McManus, J. 2004. Rare earth elements in pore waters of marine sediments. Geochimica et Cosmochimica Acta 68: 1265–1279. Crossref
Kohn, M.J., Schoeninger, M.J., and Barker, W.W. 1999. Altered states: effects of diagenesis on fossil tooth chemistry. Geochemica et Cosmochimica Acta 63: 2737–2747. Crossref
Kouchinsky, A., Bengston, S., Clausen, S., and Vendrasco, M.J. 2015. An early Cambrian fauna of skeletal fossils from the Emyaksin Formation, northern Siberia. Acta Palaeontologica Polonica 60: 421–512.
Landing, E. 1995. Upper Placentian–Branchian series of mainland Nova Scotia (Middle–Upper Lower Cambrian): faunas, paleoenvironments, and stratigraphic revision. Journal of Paleontology 69: 475–495. Crossref
Landing, E., Nowlan, G.S., and Fletcher, T.P. 1980. A microfauna associated with Early Cambrian trilobites of the Callavia Zone, northern Antigonish Highlands, Nova Scotia. Canadian Journal of Earth Sciences 17: 400–418. Crossref
Maas, A. and Waloszek, D. 2001. Cambrain derivatives of the early arthropod stem lineage, pentastomids, tardigrades and lobopodians—an “Orsten” perspective. Zoologischer Anzeiger 240: 451–459. Crossref
Maas, A., Braun, A, Dong, X.-P., Donoghue, P.C.J., Müller, K.J., Olempska, E., Repetski, J.E., Siveter, D.J., Stein, M., and Waloszek, D. 2006. The “Orsten”—More than a Cambrian Konservat-Lagerstätte yielding exceptional preservation. Palaeoworld 15: 266–282. Crossref
Marshall, J.D. 1992. Climatic and oceanographic isotopic signals from the carbonate rock record and their preservation. Geological Magazine 129: 143–160. Crossref
McIlroy, D. and Szaniawski, H. 2000. A lower Cambrian protoconodont apparatus from the Placentian of southeastern Newfundland. Lethaia 33: 95–102. Crossref
Moreno, I. and Kapp, H. 2003. Structures of grasping spines and teeth in three species of chaetognaths from Antarctic waters. Polar Biology 26: 143–150. Crossref
Müller, K.J. 1956. Zur Kenntnis der Conodonten-Fauna des europaischen Devons, 1; Die Gattung Palmatolepis. Abhandlunghen and Senckenbergischen NaturforschendenGesellschaft 494: 1–70.
Müller, K.J. 1959. Kambrische Conodonten. Zeitschrift der Deutschen Geologischen Gesellschaft 111: 434–485. Crossref
Müller, K.J. 1979. Phosphatocopine ostracods with preserved appendages from the Upper Cambrian of Sweden. Lethaia 12: 1–27. Crossref
Müller, K.J. and Walossek, D. 1985. A remarkable arthropod fauna from the Upper Cambrain “Orsten” of Sweden. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 76: 161–172. Crossref
Olempska, E., Błażejowski, B., Waloszek, D., and Maas, A. 2023. Phosphatic bromalites and microfossils from the Furongian (Cambrian) of northern Poland (Baltica) and palaeobiological implications. Palaeogeography, Palaeoclimatology, Palaeoecology 610: 111350. Crossref
Pruss, S.B., Tosca, N.J., and Stark, C. 2018. Small shelly fossil preservation and the role of early diagenetic redox in the Early Triassic. Palaios 33: 441–450. Crossref
Racki, G. 1993. Evolution of the bank to reef complex in the Devonian of the Holy Cross Mountains. Acta Palaeontologica Polonica 37: 87–182.
Rejebian, V.A., Harris, A.G., and Huebner, J.S. 1987. Conodont color and textural alteration: An index to regional metamorphism, contact metamorphism, and hydrothermal alteration. Geological Society of America Bulletin 99: 471–479. Crossref
Ségalen, L., De Rafélis, M., Lee-Thorp, J.A., Maurer, A.-F., and Renard, M. 2008. Cathodoluminescence tools provide clues to depositional history in Miocene and Pliocene mammalian teeth. Palaeogeography, Palaeoclimatology, Palaeoecology 266: 246–253. Crossref
Shen, C., Pratt, B.R., Lan, T., Hou, J.-B., Chen, L., Hao, B.-Q., and Zhang, X.-G. 2013. The search for Orsten-type fossils in southern China. Paleoword 22: 1–9. Crossref
Shen, Y., Schidlowski, M., and Chu, X. 2000. Biogeochemical approach to understanding phosphogenic events of the terminal Proterozoic to Cambrian. Palaeogeography, Palaeoclimatology, Palaeoecology 158: 99–108. Crossref
Shu, D., Conway Morris, S., Han, J., Hoyal Cuthill, J.F., Zhang, Z., Cheng, M., and Huang, H. 2017. Multi-jawed chaetognaths from the Chengjiang-Lagerstätte (Cambrian, Series 2, Stage 3) of Yunnan, China. Palaeontology 60: 763–772. Crossref
Slater, B.J. and Willman, S. 2019. Early Cambrian small carbonaceous fossils (SCFs) from an impact crater in western Finland. Lethaia 52: 570–582. Crossref
Slater, B.J., Harvey, T.H.P., and Butterfield, N.J., 2018. Small carbonaceous fossils (SCFs) from the Terreneuvian (Lower Cambrian) of Baltica. Palaeontology 61: 417–439. Crossref
Szaniawski, H. 1982. Chaetognath grasping spines recognized among Cambrian protoconodonts. Journal of Paleontology 56: 806–810.
Szaniawski, H. 1983. Structure of protoconodont elements. Fossils and Strata 15: 21–27.
Szaniawski, H. 2002. New evidence for the protoconodont origin of chaetognaths. Acta Palaeontologica Polonica 47: 405–419.
Szaniawski, H. 2005. Cambrian chaetognaths recognized in Burgess Shale fossils. Acta Palaeontologica Polonica 50: 1–6.
Szaniawski, H. 2009. Fossil chaetognaths from the Burgess Shale: a reply to Conway Morris (2009). Acta Palaeontologica Polonica 54: 361–364. Crossref
Szaniawski, H. 2015. New group of the Early Palaeozoic conodont-like fossils. Estonian Journal of Earth Sciences 64: 91–94. Crossref
Szulczewski, M. 1971. Upper Devonian conodonts, stratigraphy and facial development in the Holy Cross Mts. Acta Geologica Polonica 21: 1–129.
Szulczewski, M. 1978. The nature of unconformities in the Upper Devonian–Lower Carboniferous condensed sequence in the Holy Cross Mts. Acta Geologica Polonica 28: 283–298.
Szulczewski, M., Belka, Z., and Skompski, S. 1996. The drowning of a carbonate platform: An example from the Devonian–Carboniferous of the southwestern Holy Cross Mountains, Poland. Sedimentary Geology 106: 21–49. Crossref
Trotter, J.A., Gerald, J.D.F., Kokkonen, H., and Barnes, C.R. 2007. New insights into the ultrastructure, permeability, and integrity of conodont apatite determined by transmission electron microscopy. Lethaia 40: 97–110. Crossref
Vannier, J., Steiner, M., Renvoisé, E., Hu, S.-X., and Casanova, J.-P. 2007. Early Cambrian origin of modern food webs: evidence from predator arrow worms. Proceedings of the Royal Society B 274: 627–633. Crossref
Walossek, D. 1999. On the Cambrian diversity of Crustacea. In: F.R. Schram and J.C. von Vaupel Klein (eds.), Crustaceans and the Biodiversity Crisis. Proceedings of the Fourth International Crustacean Congress, Amsterdam, The Netherlands, July 20–24, 1998, Vol. 1, 3–27. Brill Academic Publishers, Leiden.
Waloszek, D. 2003. The “Orsten” window—a three-dimensionally preserved Upper Cambrain meiofauna and its contribution to our understanding of the evolution of Arthropoda. Paleontological Research 7: 71–88. Crossref
Wierzbowski, H. 2021. Advances and challenges in palaeoenvironmental studies based on oxygen isotope composition of skeletal carbonates and phosphates. Geosciences 11: 419. Crossref
Wierzbowski, H., Szaniawski, H., and Błażejowski, B. 2021. Structural, chemical and isotope evidence for secondary phosphate mineralization of grasping spines of Early Paleozoic chaetognaths. Lethaia 54: 245–259. Crossref
Wright, J. 1989. Conodont apatite: structure and geochemistry. In: J.G. Carter (ed.), Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends. Short Course in Geology: Volume 5, Part II, 149– 163. American Geophysical Union, New York Crossref
Zhuravlev, A.V. 2017. Comparative histology of conodonts and early vertebrates. Vestnik IG Komi SC UB RAS 2017/4: 37–42. Crossref
Zhuravlev, A.V. [Žuravlev, A.V.] and Gerasimova, A.I. 2015. Albid tissue of the conodont elements: composition and forming models [in Russian with English summary]. Vestnik IG Komi SC UB RAS 2015/10: 21–27. Crossref
Ziegler, W. 1960. Conodonten aus dem rheinischen Unterdevon (Gedinnium) des Remscheider Sattels (Rheinisches Schiefergebirge). Palaeontologische Zeitschrift 34: 169–200. Crossref
Ziegler, W. 1962. Taxionomie und Phylogenie Oberdevonischer Conodonten und ihre stratigraphische Bedeutung. Abhandlunghen des Hessisches Landesamt für Bodenforschung 38: 1–166.
Ziegler, W. and Sandberg, C.A. 1984. Palmatolepis-based revision of upper part of standard Late Devonian conodont zonation. In: D.L. Clark (ed.), Conodont Biofacies and Provincionalism. GSA Special Papers 196: 179–194. Crossref
Acta Palaeontol. Pol. 68 (1): 103–116, 2023
https://doi.org/10.4202/app.01012.2022