
La Piquera in central Iberian Peninsula: A new key vertebrate locality for the Early Pliocene of western Europe
PEDRO PIÑERO, DAVID MANUEL MARTÍN-PEREA, PALOMA SEVILLA, JORDI AGUSTÍ, HUGUES-ALEXANDRE BLAIN, MARC FURIÓ, and CÉSAR LAPLANA
Piñero, P., Martín-Perea, D.M., Sevilla, P., Agustí, J., Blain, H.-A., Furió, M., and Laplana, C. 2023. La Piquera in central Iberian Peninsula: A new key vertebrate locality for the Early Pliocene of western Europe. Acta Palaeontologica Polonica 68 (1): 23–46.
We introduce the new vertebrate site of La Piquera (Duero Basin, central Iberian Peninsula), provide a detailed description of the lithostratigraphy and present a preliminary faunal list. The fossil vertebrate association includes amphibians (with representatives of the families Salamandridae, Alytidae, Pelodytidae, and Bufonidae), squamate reptiles (with members of the families Agamidae, Lacertidae, Anguidae, Scincidae, Blanidae, ?Erycidae, and Colubridae/Psammophiidae), and mammals (with representatives of the families Soricidae, Erinaceidae, Muridae, Cricetidae, Gerbillidae, Gliridae, Sciuridae, Vespertilionidae, Rhinolophidae, Miniopteridae, Megadermatidae, Leporidae, Prolagidae, and Bovidae). The association indicates an intermediate biostratigraphic position between the southern Iberian sites of Sifón-413 (at about 5.33–5.23 Ma) and Botardo-D (between 4.79 and 4.63 Ma), the new locality being correlated with the earliest Ruscinian (lower part of the MN14 unit). The ecological affinities of the identified small vertebrates suggest the presence of a landscape dominated by open herbaceous meadows in the surroundings of the fossil-site during the earliest Pliocene, with the occasional presence of woodland patches and stable water bodies under relatively dry and warm environmental conditions. La Piquera therefore enhances our knowledge on the vertebrate community recorded in central Iberian Peninsula during the earliest Pliocene. With more than 1200 remains, La Piquera becomes a key locality for the Early Pliocene of southwestern Europe, central Spain, where this time-span is poorly represented.
Key words: Amphibia, Reptilia, Mammalia, herpetofauna, micromammals, biochronology, palaeoecology, Duero Basin, Spain, Early Pliocene.
Pedro Piñero [ppinero@iphes.cat; ORCID: https://orcid.org/0000-0002-5626-2777 ] and Hugues-Alexandre Blain [hablain@iphes.cat; ORCID: https://orcid.org/0000-0002-9920-2707 ], IPHES-CERCA, Institut Català de Paleoecologia Humana i Evolució Social, Zona Educacional 4, Campus Sescelades URV (Edifici W3), 43007 Tarragona, Spain; Àrea de Prehistòria, Universitat Rovira i Virgili (URV), Avinguda de Catalunya 35, 43002 Tarragona, Spain.
David Manuel Martín-Perea [davidmam@ucm.es; ORCID: https://orcid.org/0000-0002-3238-0904 ], Departamento de Paleobiología, Museo Nacional de Ciencias Naturales (CSIC), José Gutiérrez Abascal 2, 28006 Madrid, Spain; Departamento de Geodinámica, Estratigrafía y Paleontología, Facultad de Ciencias Geológicas, Universidad Complutense de Madrid, Ciudad Universitaria, 28040 Madrid, Spain; Institute of Evolution in Africa, Covarrubias 36, 28010 Madrid, Spain.
Jordi Agustí [jordi.agusti@icrea.cat; ORCID: https://orcid.org/0000-0002-7240-1992 ], ICREA, Institució Catalana de Recerca i Estudis Avançats, Pg. Lluís Companys 23, 08010 Barcelona, Spain; IPHES-CERCA, Institut Català de Paleoecologia Humana i Evolució Social, Zona Educacional 4, Campus Sescelades URV (Edifici W3), 43007 Tarragona, Spain; Àrea de Prehistòria, Universitat Rovira i Virgili (URV), Avinguda de Catalunya 35, 43002 Tarragona, Spain.
Paloma Sevilla [psevilla@ucm.es; ORCID: https://orcid.org/0000-0001-7425-7173 ], Departamento de Geodinámica, Estratigrafía y Paleontología, Facultad de Ciencias Geológicas, Universidad Complutense de Madrid, Ciudad Universitaria, 28040 Madrid, Spain.
Marc Furió [marc.furio@uab.cat; ORCID: https://orcid.org/0000-0002-4582-3268 ], Serra Húnter fellow, Departament de Geologia, Universitat Autònoma de Barcelona, 08193, Bellaterra, Spain; Institut Català de Paleontologia Miquel Crusafont, Universitat Autònoma de Barcelona, Edifici ICTA-ICP, Carrer de les Columnes s/n, Campus de la UAB, 08193, Cerdanyola del Vallès, Barcelona, Spain.
César Laplana [cesar.laplana@gmail.com; ORCID: https://orcid.org/0000-0002-2067-4091 ], MAR, Museo Arqueológico y Paleontológico de la Comunidad de Madrid, Plaza de las Bernardas s/n, 28801 Alcalá de Henares, Spain.
Received 5 July 2022, accepted 28 November 2022, available online 6 February 2023.
Copyright © 2023 P. Piñero et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
La Piquera site (N 41°25’56”, W 3°40’37”, 1128 m above see level) is located less than a kilometer south from Carabias (Segovia, Spain; Fig. 1). The site was discovered in 2014 by one of the authors (CL) in an inactive aggregate quarry called La Piquera (Fig. 1), which revealed several karstic cavities carved into Late Cretaceous dolostone and limestone. A report notifying the discovery was sent to the Territorial Service of Culture of Segovia and to the Directorate General of Cultural Heritage of the Junta de Castilla y León. The karstic infill was initially inferred a Neogene age (Late Miocene–Pliocene) given the surrounding sedimentary deposits in the study area (García-Cortés et al. 2008).
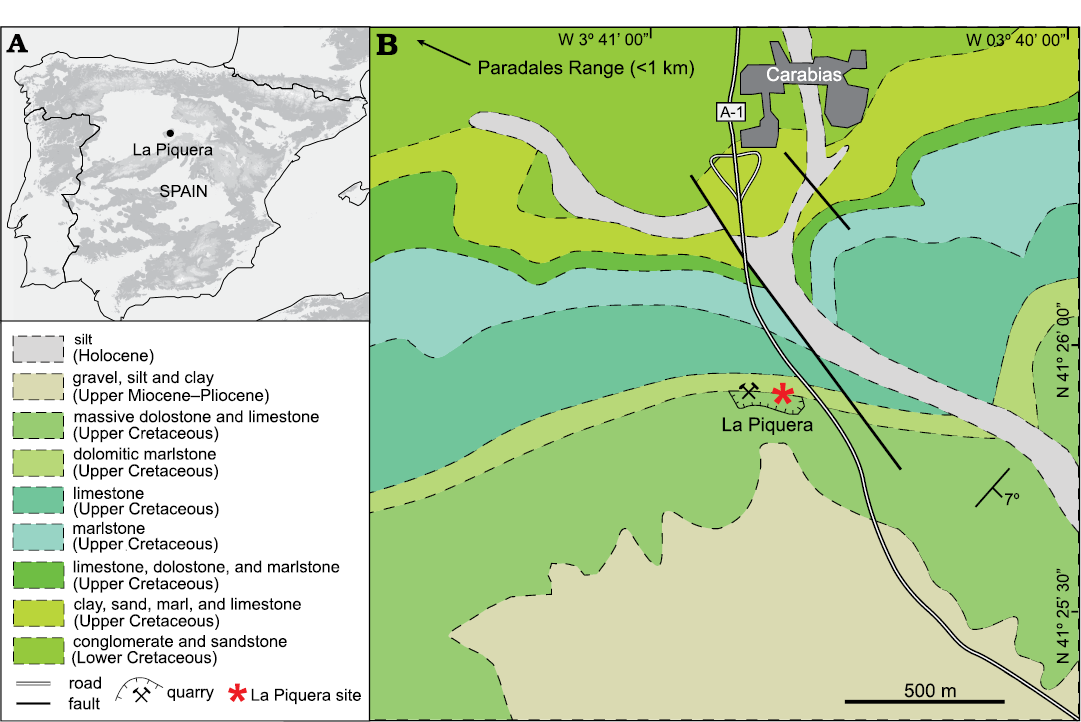
Fig. 1. Geographic and geological context of La Piquera site, Lower Pliocene, Duero Basin, Spain. A. Map of the Iberian Peninsula showing the location of La Piquera site. B. Geological map of the study area and location of La Piquera quarry and site (modified from García-Cortés et al. 2008).
The Pliocene (5.33–2.58 Ma) was the last significant warm epoch in Earth’s climate history before the onset of Pleistocene glacial-interglacial dynamics. The Pliocene marine record is well known from several sections and cores, but continental sections are scarce in western Europe. In particular, the earliest Pliocene is not so well represented due to the scarcity of sites from this time. In this regard, the new site of La Piquera represents one of the few localities in the Iberian Peninsula from the earliest Pliocene, together with Sifón-413 in the Fortuna Basin (Piñero and Agustí 2019), Puerto de la Cadena in the Murcia-Carrascoy Basin (Piñero et al. 2017b), Calicasas 3 and 4 and Purcal 4 and 7 in the Granada Basin (García-Alix et al. 2008a–c), Peralejos E in the Teruel Basin (Adrover et al. 1988; Mein et al. 1990), and Botardo C, D in the Guadix-Baza Basin (Martín-Suárez 1988; Piñero and Agustí 2020). Yet, there is a lack of information from continental Early Pliocene vertebrate faunas, especially from the Duero Basin.
The Duero Basin, located in central Iberian Peninsula, represents the largest continental Neogene basin of Spain. The age of the stratigraphic successions of this basin ranges from the Early Miocene (Agenian, MN2) to the Early Pliocene (early Ruscinian, MN14) (López et al. 1987; Calvo Sorando et al. 1993; Luengo et al. 2009, 2010). However, localities providing vertebrate faunas around the Mio-Pliocene transition are limited to Castillejo de Robledo, a site that has only yielded remains of rodents and lagomorphs (Luengo et al. 2009). This site was initially placed in the Early Pliocene (Luengo et al. 2009), although its chronology was later reconsidered and the site was placed at the end of the Late Miocene (Morales et al. 2013). Consequently, a great effort was placed in the recent sampling campaigns at La Piquera to recover a representative number of vertebrate remains. This effort has proved to be worthwhile, as evidenced by the unprecedented number of fossils collected. Indeed, La Piquera has yielded one of the most extensive collections of small vertebrates from the earliest Pliocene in western Europe. Thereby, the significance of La Piquera lies both in its chronology and in the outstanding fossil vertebrate sample size. This provides an opportunity to better understand changes in faunal communities in the Iberian Peninsula after the Messinian Salinity Crisis (Krijgsman et al. 1999).
In this paper, we introduce the site of La Piquera by providing a detailed description of the lithostratigraphy and presenting a preliminary faunal list. This enables us to estimate the age of the new site and to infer the prevalent palaeoenvironmental conditions during its formation.
Institutional abbreviations.—UCM, Complutense University of Madrid, Spain
Abbreviations.—dp, lower deciduous premolar; M, upper molar; m, lower molar; MN, Mammal Neogene; P, upper premolar; p, lower premolar; t, tubercle.
Material and methods
Lithostratigraphic levels were described, measured, and logged using scaled drawings and photographed. Sediment types have been classified based on particle size distributions according to Blott and Pye (2012).
The fossil material referred to here was collected from the La Piquera site during the 2021 sampling campaign, including remains from all palaeontological levels indistinctly. All the sediment retrieved during this campaign was water-screened using superimposed 4-, 1-, and 0.5-mm mesh sieves. The collection from La Piquera includes more than a thousand identified small mammal teeth, mandibles and maxillae corresponding to 25 different taxa, and more than 200 disarticulated cranial and postcranial bones of amphibians and squamate reptiles comprising at least 18 taxa (see Table 1). Three large mammal tooth fragments have also been recovered. Although these fossils are currently spread out, housed in the Complutense University of Madrid (UCM), the Spanish National Natural Sciences Museum (MNCN), the Archaeological and Palaeontological Museum of the Community of Madrid (APM), and the Catalan Institute of Human Paleoecology and Social Evolution (IPHES), the final repository is going to be the Geodynamics, Stratigraphy and Paleontology Department at UCM (Madrid, Spain). Small mammal teeth are illustrated by means of micrographs taken with Environmental Scanning Electron Microscopy (ESEM) at the Servei de Recursos Científics i Tècnics de la Universitat Rovira i Virgili (Tarragona) with the exception of the sciurid and lagomorph teeth, whose photographs were taken with an Olympus SC180 digital camera coupled to an Olympus SZX12 binocular at the APM.
Table 1. Vertebrate faunal list of the site of La Piquera.
|
Order |
Family |
Species |
|
Caudata |
Salamandridae |
Pleurodeles sp. |
|
Salamandridae indet. |
||
|
Anura |
|
Anura indet. |
|
Alytidae |
Discoglossus sp. |
|
|
Pelodytidae |
Pelodytes sp. |
|
|
Bufonidae |
Bufotes cf. B. viridis group |
|
|
Squamata |
|
Serpentes indet. |
| |
(non-snake) Squamata indet. |
|
|
Agamidae |
Agamidae indet. |
|
|
Lacertidae |
Lacertidae indet. 1 |
|
|
Lacertidae indet. 2 |
||
|
Anguidae |
Ophisaurus sp. |
|
|
Scincidae |
Scincidae indet. |
|
|
Blanidae |
Blanus sp. |
|
|
Erycidae |
Erycidae indet. |
|
|
Colubridae / |
morphotype 1 |
|
|
morphotype 2 |
||
|
Colubridae/Psammophiidae indet. |
||
|
Eulipotyphla |
Soricidae |
Myosorex meini |
|
Paenelimnoecus cf. pannonicus |
||
|
Neomyini indet. |
||
|
Erinaceidae |
Parasorex ibericus |
|
|
Chiroptera |
Rhinolophidae |
Rhinolophus sp. 1 Rhinolophus sp. 2 |
|
Vespertilionidae |
Myotis sp. 1 Myotis sp. 2 |
|
|
Megadermatidae |
Megadermatidae indet. |
|
|
Miniopteridae |
Miniopterus sp. |
|
|
Rodentia |
Muridae |
Stephanomys dubari |
|
Apodemus gorafensis |
||
|
Paraethomys meini |
||
|
Occitanomys alcalai |
||
|
Castillomys gracilis |
||
|
Cricetidae |
Apocricetus cf. barrierei |
|
|
Ruscinomys lasallei |
||
|
Blancomys aff. sanzi |
||
|
Gerbillidae |
Debruijnimys sp. |
|
|
Gliridae |
Eliomys truci |
|
|
Glis cf. minor |
||
|
Sciuridae |
Atlantoxerus sp. |
|
|
Lagomorpha |
Leporidae |
Trischizolagus sp. |
|
Prolagidae |
Prolagus sp. 1 Prolagus sp. 2 |
|
|
Artiodactyla |
Bovidae |
Bovidae indet. |
The systematic nomenclature generally follows Speybroeck et al. (2020) for extant herpetofaunal taxa, with the exception of higher taxonomical categories that follow Georgalis and Smith (2020) for reptiles (especially snakes), and Hedges (2014) for scincids. Osteological nomenclature follows Sanchiz (1998) and Bailon (1999) for amphibians, Augé (2005) and Barahona and Barbadillo (1997) for lizards, and Szyndlar (1984, 1991) for snakes. We employed the tooth terminology proposed by Van de Weerd (1976) for the descriptions of murids, Mein and Freudenthal (1971) for cricetids, Freudenthal (2004) for glirids, Wood and Wilson (1936) for gerbilids, Sinitsa and Pogodina (2019) for sciurids, Reumer (1984) for soricids, Mein and Martín Suárez (1993) for erinaceids, Angelone and Sesé (2009) for prolagids, López-Martínez et al. (2007) for leporids, and Menu and Popelard (1987) and Sevilla (1988) for bats.
Geological setting
La Piquera site is located in the southeastern sector of the Duero Basin, ~20 km north of the Central System (Somosierra Range) and less than a kilometer southeast from the Pradales Range (Fig. 1B). The Pradales Range, with peaks above 1300 m above see level (masl), towers above the rest of the study area (average of 1000 masl). This range has a WSW-ENE lineation, and is composed of Mesozoic rocks: Triassic claystone, sandstone, conglomerate, marlstone, and dolostone; Jurassic limestone; and Cretaceous claystone, sandstone, conglomerate, marlstone, limestone, and dolostone (García-Cortés et al. 2008). Of these, in our study area, only the Cretaceous materials are exposed, alongside Upper Miocene–Pliocene and Holocene deposits (García-Cortés et al. 2008; Fig. 1B).
The site of La Piquera consists of a small (around 1.4 m deep and 1 m width) vertically developed karstic cavity carved into Maastrichtian (Late Cretaceous) dolostone and limestone beds completely filled with sediments (Figs. 1B, 2, 3). Its sedimentary filling can be divided into 16 lithostratigraphic units, six of which bear small vertebrate remains (Fig. 2).

Fig. 2. Stratigraphic context of La Piquera site, Lower Pliocene, Duero Basin, Spain. A. Photograph of the La Piquera karstic infill and limit with dolostone and limestone bedrock, showing levels with paleontological remains. B. La Piquera stratigraphic section highlighting the six lithostratigraphic units with small vertebrate content.
Unit 1 is a red (2.5YR 4/6), 13-cm-thick, very well sorted fine and very fine sandstone. Overlying it, Unit 2 hosts the first fossiliferous level (Level 1). Unit 2 is a yellowish red (5YR 5/8) coarsening upwards sequence, from fine and medium (lower 4 cm) to coarse (uppermost 1 cm) sandstone. Abundant microfaunal remains are found in the lower 1 cm and uppermost 1 cm of the unit (Level 1). Unit 3 is a red (2.5YR 4/6) very fine sandy and silty deposit, very well sorted and 11 cm thick. Unit 4 is a 10-cm-thick coarsening upwards sequence, from fine to coarse red (2.5YR 4/6) sandstone. Unit 5 is composed of three coarsening upwards sequences, from fine and very fine sandstone to medium and coarse sandstone. Abundant microfaunal remains are found throughout the three sequences (Level 2). Unit 6 is a very well sorted, red (2.5YR 4/6) quartzofeldspathic fine sandstone. Unit 7 is a thin, 4-cm-thick, well sorted, light grey (5Y 7/1) fine sand, with abundant microfauna in the lowermost part (Level 3). Unit 8 is composed of several centimetric very fine to fine red (2.5YR 4/6) sandstone levels. Unit 9 is a thin, <3-cm-thick, light grey (7.5YR 7/1) coarsening upwards level, changing from fine to coarse sandstone, and contains copious amounts of microfaunal remains (Level 4). Unit 10 is a 15-cm-thick red (2.5YR 4/6) laminated siltstone. Unit 11 is a 5-cm-thick medium to coarse pale red (2.5YR 6/2) sandstone. Unit 12 is composed of a thin (4 cm) claystone. Both Unit 11 and Unit 12 contain abundant microfaunal remains and belong to palaeontological Level 5. Unit 13 is a 6-cm-thick pale red (2.5YR 6/2) siltstone, overlain by Unit 14, which is a 4-cm-thick sandstone. Unit 15, which contains Level 6, is a coarsening upwards bed, changing from siltstone to coarse sandstone with some pebbles and abundant palaeontological remains. The stratigraphic sequence ends with Unit 16, which is composed of 23 cm of pale red (2.5YR 6/2) siltstone.
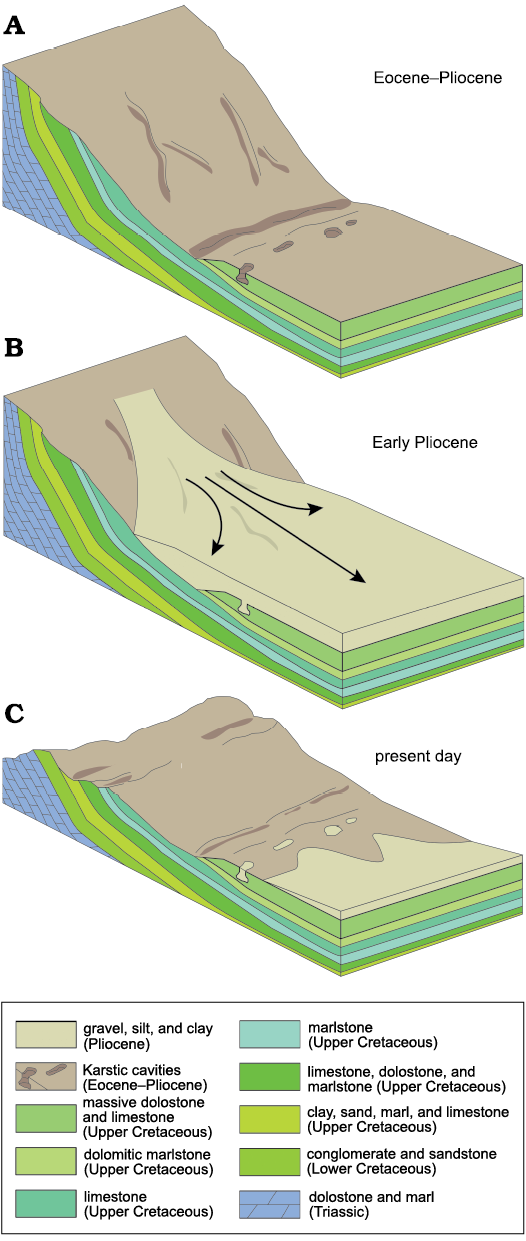
Fig. 3. La Piquera schematic block diagrams showing geological history. A. Eocene to Pliocene. Alpine orogeny causes Pradales Range to build. Local topographic variations lead to the carving of karstic cavities into Maastrichtian dolostone and limestone beds. B. Early Pliocene. Pradales Range dismantling and erosion create alluvial fans. Medial to distal deposits infill the karstic cavities. C. Present day. Weathering and erosion remove Pliocene alluvial fan deposits and expose karstic cavities.
Results
Faunal content.—The vertebrate association from the La Piquera site (Figs. 4–8) is highly diversified, comprising urodeles, anurans, lizards, snakes, artiodactyls, insectivores, bats, rodents, and lagomorphs (Table 1).
Herpetofauna.—Concerning amphibians and squamate reptiles, La Piquera documents a rather high diversity with one urodele (a salamandrid), three anurans (one alytid, one pelodytid, and one bufonid), seven lizards (one agamid, three lacertids, one scincid, one anguid, and one blanid), and three snakes (one potential erycid, and two colubrids or psammophiids). The faunal list is composed of: Pleurodeles sp., Salamandridae indet., Discoglossus sp., Pelodytes sp., Bufotes cf. B. viridis group, Anura indet., Agamidae indet., Lacertidae indet. 1 (small-sized with mainly bicuspid teeth), Lacertidae indet. 2 (middle-sized with mainly bicuspid teeth), indeterminate lizard 1 (small-sized with tricuspid teeth), Scincidae indet., Ophisaurus sp., Blanus sp., (non-snake) Squamata indet., ?Erycidae indet., Colubridae/Psammophiidae (morphotype 1), Colubridae/Psammophiidae (morphotype 2), Colubridae/Psammophiidae indet., and Serpentes indet. A complete osteological description of these fossils will be done in a future publication. Only the most important taxa are described here.
Amphibians: Pleurodeles is represented by a few trunk vertebrae at La Piquera. These vertebrae (Fig. 4A) preserve a small, concave-shaped bony lamina that connects the ventral side of the prezygapophyses to the ventral costal process. This character is only found in Pleurodeles (i.e., Haller-Probst and Schleich 1994).
Discoglossus is documented by a small ilium (Fig. 4B). This ilium bears a dorsally swollen tuber superior followed by a low dorsal crest on its anterior branch, a rather long pars ascendens, a small but clearly developed pars descendens, and a smooth posteromedial face that has an interiliac groove clearly visible in posterior view and an interiliac tubercle. Only Discoglossus possesses such set of characteristics (e.g., Roček 1994).
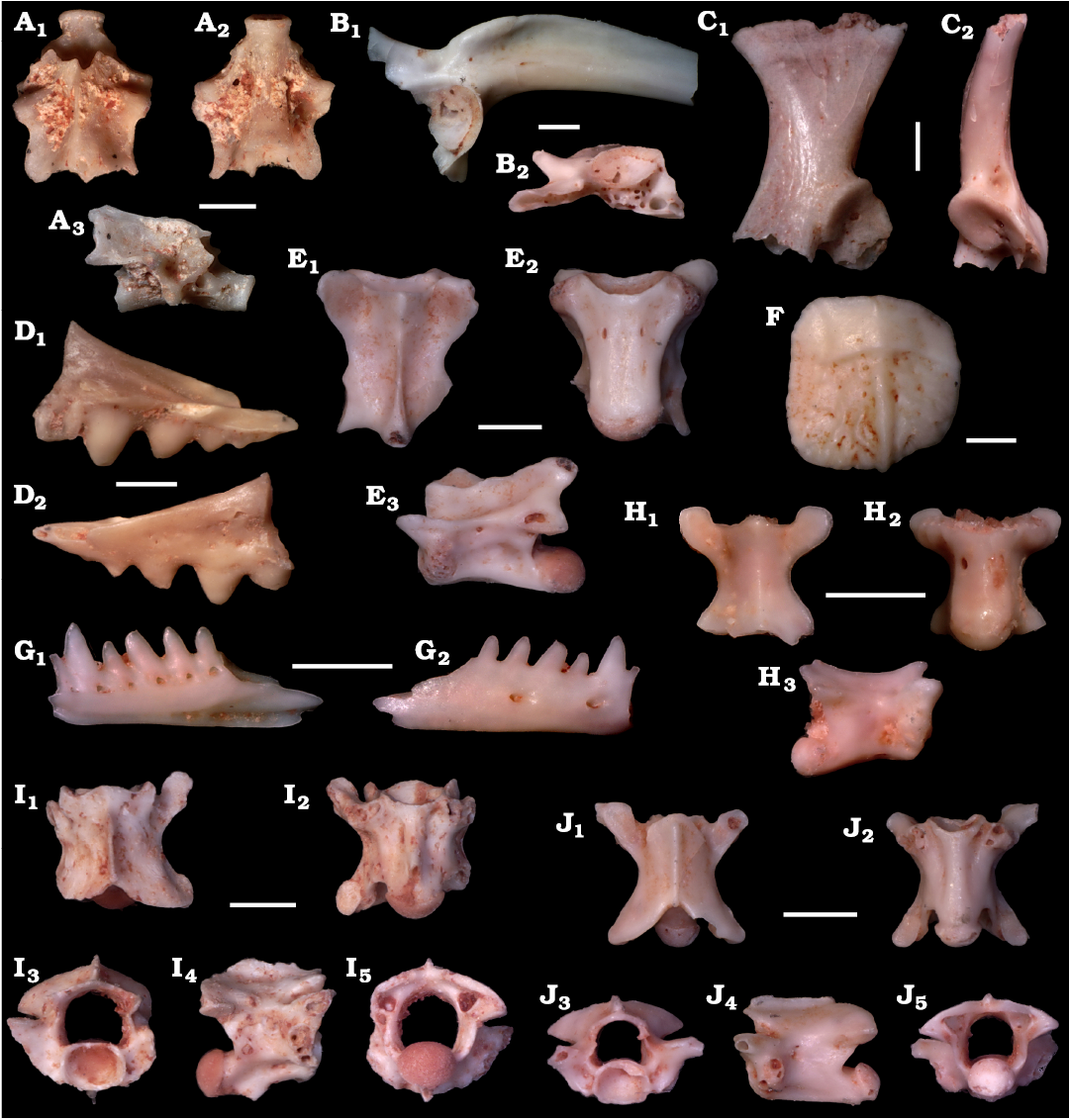
Fig. 4. Digital images of amphibians and reptiles from La Piquera site, Lower Pliocene, Duero Basin, Spain. A. Salamandrid Pleurodeles sp., trunk vertebra in dorsal (A1), ventral (A2), and right lateral (A3) views, UCM-LPQ-CAU-1. B. Alytid Discoglossus sp., right ilium in lateral (B1) and posterior (B2) views, UCM-LPQ-ANU-1. C. Bufonid Bufotes cf. B. viridis group, left scapula in dorsal (C1) and posterior (C2) views, UCM-LPQ-ANU-2. D. Agamidae indet., maxilla? in mesial (D1) and lateral (D2) views, UCM-LPQ-SQU-1. E, F. Anguid Ophisaurus sp. E. Trunk vertebra in dorsal (E1), ventral (E2), and left lateral (E3) views, UCM-LPQ-SQU-2. F. Osteoderm, UCM-LPQ-SQU-3. G, H. Amphisbaenian Blanus sp. G. Right dentary in mesial (G1) and lateral (G2) views, UCM-LPQ-SQU-4. H. Trunk vertebra in dorsal (H1), ventral (H2), and right lateral (H3) views, UCM-LPQ-SQU-5. I. ?Erycidae indet., anterior trunk vertebra in dorsal (I1), ventral (I2), anterior (I3), right lateral (I4), and posterior (I5) views, UCM-LPQ-SQU-6. J. Colubridae/Psammophiidae (morphotype 1), trunk vertebra in dorsal (J1), ventral (J2), anterior (J3), left lateral (J4), and posterior (J5) views, UCM-LPQ-SQU-7. Scale bars 1 mm.
Pelodytes is represented by a small ilium with no tuber superior and dorsal crest, and a typically slender humerus bearing a straight diaphysis and a condyle slightly moved laterally.
Bufotes cf. B. viridis group is represented by a scapula (Fig. 4C). The scapula is longer than wide, and bears a robust, detached glenoid process that is clearly visible in dorsal view. The attribution to the B. viridis group is based on the sinuous concave anterior margin of the pars acromialis and the processus glenoidalis, whereas in all other species but Bufo verrucosissimus it is convex, and the presence of a shallow supraglenoid fossa on the base of the glenoid process, well visible on the posterior view, whereas in Bufo gr. B. bufo this fossa is absent and in Epidalea calamita this fossa is generally deeper (Bailon 1999).
Lizards: Agamid lizards are represented by a sole fragment of maxilla or dentary, bearing the typical acrodont dentition (Fig. 4D).
Lacertid lizards are documented by a small number of dentaries. Their pertaining to the family Lacertidae is mainly based on the tooth morphology and their pleurodont insertion on the bone. The teeth are isodont, cylindrical and mono-, or bicuspid and on the dentaries the Meckelian canal is wide open. Some variation concerning the size of the fossil elements as well as differences observed between the types of cuspids represented on dentaries permit to cautiously propose several morphotypes, as listed above.
Scincid lizards are represented by some fragmentary elements, mainly dentaries, bearing pleurodont, isodont, cylindrical, and monocuspid teeth with a blunt apex. In lingual view, each tooth apex shows more or less visible ornamentation, with delicate vertical striations limited ventrally by a transverse groove.
Anguid lizards are represented by a few trunk vertebrae (Fig. 4E) and one osteoderm (Fig. 4F). The trunk vertebrae have a total length of less than 5 mm. They are procoelous, longer than wide, with a long, low neural spine, and prolonged posteriorly in an interzygapophyseal tip. In ventral view, the centrum is subtriangular, with concave lateral margins and a slightly convex ventral surface in transverse section. The osteoderm is slightly longer than wide and dorsally bears relatively discrete dermal ornamentation, formed of anastomosing grooves (vermiculate ornamentation). The presence of a well-marked longitudinal keel suggests a mediodorsal position for this osteoderm. Attribution to Ophisaurus is based on the subtriangular centrum with concave lateral margins in the trunk vertebrae (in contrast to the almost parallel lateral margins presents in Anguis or to the more robust subtriangular centrum with straight lateral margins in Pseudopus; e.g., Klembara and Rummel 2018), and the presence of a longitudinal keel on the osteoderm (present in Ophisaurus and Pseudopus but absent in Anguis; Hoffstetter 1962).
Amphisbaenian squamate Blanus sp. is represented by two incomplete dentaries (Fig. 4G) and three trunk vertebrae (Fig. 4H). The preserved parts of the dentaries suggest that they were short and robust, bearing sub-pleurodont and monocuspid teeth. The anterior teeth are cylindrical and inclined anteriorly whereas the central and posterior teeth are cone-shaped, with a wide base and a posteromedially directed apex. The Meckel’s groove is open along its whole length. The trunk vertebrae are procoelous, with a cotyle and a condyle dorsoventrally flattened. They show a dorsoventrally flattened neural arch, with a concave posterior end and a well-marked interzygapophyseal constriction. The neural spine is reduced, and in lateral view the synapophyses are globular and egg-shaped, which is characteristic of posterior trunk vertebrae. The ventral surface of the centrum is flat, with slightly convex margins. All these characters permit an attribution of the trunk vertebrae to Amphisbaenia. Currently, no clear diagnostic feature allowing identification at family level is known for isolated trunk vertebrae of this group of squamates (i.e., Georgalis et al. 2018). Here their attribution to Blanidae relies on their close association with the dentaries.
Snakes: A unique small cervical vertebra is reported to a potential erycid snake (Fig. 4I). In dorsal view it is wider than long with a well-marked interzygapophyseal constriction. The neural spine is relatively strong, and it is low and long. The zygosphene is wide and possesses a convex anterior margin. The articular surface of the preserved prezygapophysis is well developed and more or less oval-shaped. In ventral view, the centrum is anteriorly wide, short, and convex. In ventral view, the haemal keel is slender and well delimited laterally, and it ends in a short and relatively robust hypapophysis with a sigmoid ventral margin in lateral view. In anterior view, the cotyle is slightly flattened dorso-ventrally. There is no evidence of paracotylar foramina. The only preserved prezygapophysis lies upward and the neural canal is wide and roughly trapezoidal-shaped. Small sized trunk vertebrae, wider than long, in combination with low neural arches characterize the erycids (Hoffstetter and Rage 1972; Szyndlar 1991; Szyndlar and Schleich 1994; Georgalis 2019). The absence of caudal vertebrae precludes a definite attribution to erycid snakes.
Colubrid and/or psammophiid snakes are represented by five vertebrae. Two morphotypes can be recognized among the material. The morphotype 1 is represented by a unique trunk vertebra (Fig. 4J). The centrum has a length of 1.5 mm and is 1.57 times longer than wide. The neural spine is interpreted as having been originally extremely low. The prezygapophyseal processes, although broken, seem to have been long. The zygosphene is unfortunately incomplete but seems to have been narrow. In ventral view, the haemal keel is very large on its whole length, with well-defined lateral margins. Among colubrids, these characters are only observed in genus Telescopus (Szyndlar 1991, 2005), however the most diagnostic character is missing: parapophyses twice longer than the diapophyses (eroded on our fossil). Among psammophiids, this vertebra is somewhat reminiscent of Rhageris moilensis (Georgalis and Szyndlar 2022), but the margins of the neural arch are not straight in posterior view on our fossil.
The morphotype 2 corresponds to a rather elongated medium sized trunk vertebra (centrum length is around 4.0 mm). In ventral view, the centrum is narrow, with slightly divergent lateral margins towards the front and has a long and narrow haemal keel over its entire length with well-defined lateral margins. In dorsal view, the articular surfaces of the prezygapophyses are sub-oval and anteriorly directed. The prezygapophyseal processes, although broken, seem to have been long and pointed. The neural spine seems to be moderately high. In posterior view, the posterior edges of the neural arch are more or less straight and forming an angle close to 90°. When compared with extant Iberian colubrid snakes, its morphology fits with the one usually observed in species of Hemorrhois and Malpolon. When compared with species of Psammophis, our fossil is much less elongated.
The faunal list presented here for La Piquera fills a gap between the Miocene and the Late Pliocene faunas, and represents the first mentions for the Spanish Early Pliocene for Pleurodeles (previously known from the Upper Pliocene of Las Higueruelas and Camp dels Ninots; Mazo 1996; Gómez de Soler et al. 2012), agamid lizards (only recovered in the Upper Pliocene of Sarrión 1; HAB personal observations), and an erycid snake (represented in the Upper Pliocene by Eryx primitivus in Gorafe 5 and Eryx aff. E. primitivus in Moreda A; Szyndlar and Schleich 1994; Bailon 1991). Interestingly, besides an isolated mention of agamid lizards (similar to “Tinosaurus”) in the Eocene of Spain (Bolet 2017), La Piquera would probably represent the earliest mention of an extant genus in the Iberian Peninsula, whereas the family is well known from the Miocene and Pliocene of Italy and Greece (Delfino et al. 2008; Georgalis et al. 2019). The presence of a tricuspid toothed lizard in La Piquera is also of interest. Iguanidae, Teiidae, and some lacertid lizards are known to possess pleurodont teeth, which are flared and tricuspid (Augé 2005). Iguanidae (only known in Europe from the Upper Cretaceous, Eocene, and Oligocene; Augé 2005; Smith 2009; Augé and Pouit 2012; Bolet 2017) and teiid lizards (currently restricted to the Americas, and known as fossils from the Eocene of France; Augé 2005; Augé and Brizuela 2020) can be reasonably discarded. Further comparisons with extant and extinct tricuspid lacertids may probably extend their occurrence to the Early Pliocene.
Mammals.—La Piquera shows a highly diversified mammal association, with six bats (two rhinolophids, two vespertilionids, one megadermatid, and one miniopterid), four insectivores (three soricids, and one erinaceid), 12 rodents (five murids, three cricetids, one gerbillid, two glirids, and one sciurid), three lagomorphs (one leporid, and two prolagids), and one artiodactyl (a bovid).
Chiropterans: Bat remains in the new site of La Piquera are distinctly less abundant than those of other small mammal groups. However, a preliminary overview of the material recovered so far has evidenced a relatively high species richness, among which the families Vespertilionidae, Rhinolophidae, Miniopteridae, and Megadermatidae are represented. The material consists predominantly of isolated teeth, although mandibular and maxillary fragments carrying teeth have been recovered, as well as a couple of baculi and the distal part of a humerus. Fragments of phalanges are also relatively abundant, but unfortunately these only provide evidence of the presence of chiropterans and have no further taxonomic value.
Rhinolophid remains are the most numerous and they come from at least two different species discernible from size differences observed in homologous teeth (Fig. 5B, C). Rhinolophid isolated teeth are easily recognizable in fossil assemblages in which they are present thanks to their delicate appearance, with slender cusps, nyctalodont lower molars carrying a thin labial cingulum, first two upper molars with a distolingual heel, and flattened upper canines with a robust labial cingulum (Sevilla 1988).
Vespertilionids have yielded a lower number of remains, but seem to be represented as well by several species belonging to the genus Myotis (Fig. 5D, E). In this genus, the teeth have a more robust appearance, due to thicker cusps. The upper molars have no talon, and the lower molars are typically myotodont with relatively shorter trigonids and thick cingula on the labial side (Sevilla 1988).
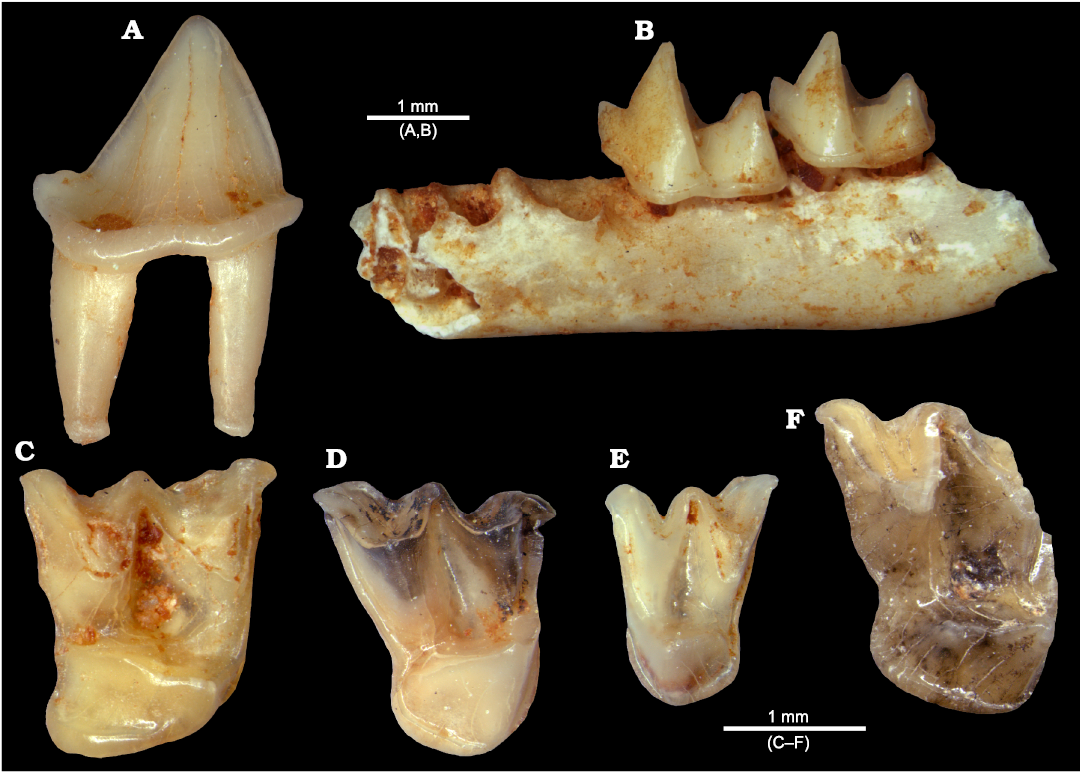
Fig. 5. Digital images of chiropterans from La Piquera site, Lower Pliocene, Duero Basin, Spain. A. Megadermatidae indet., UCM-LPQ-CHI-1, right p4 in lateral view. B. Rhinolophid Rhinolophus sp. 1, UCM-LPQ-CHI-2, left mandibular fragment with m2 and m3 in lateral view. C. Rhinolophid Rhinolophus sp. 2, UCM-LPQ-CHI-3, right M1 in occlusal view. D. Vespertilionid Myotis sp. 1, UCM-LPQ-CHI-4, right M1 in occlusal view. E. Vespertilionid Myotis sp. 2, UCM-LPQ-CHI-5, left M2 in occlusal view. F. Miniopterid Miniopterus sp., UCM-LPQ-CHI-6, right M2 in occlusal view.
Besides, the families Miniopteridae and Megadermatidae are also represented, although so far by a single specimen each. The former family comprises a single genus, Miniopterus, with several extant species and a few fossil species in the European Neogene. The second upper molar found in La Piquera is partially broken (Fig. 5F), its anterolabial part missing so that the characteristic hook-like parastyle of all Miniopterus species is not observable. However, the overall outline, the shape of the metacone and protocone, and the small hypocone shelf are features that enable to assign this tooth to this genus (Sevilla and López-Martínez 1986).
Most interesting is the fourth lower premolar belonging to a megadermatid (Fig. 5A). Although this family is well represented in European Miocene assemblages (Sigé and Legendre 1983; Storch 1999), it is rather rare in the Pliocene, with no records of this epoch in the Iberian Peninsula (Sevilla 1991). This tooth is laterally compressed instead of squarish as in vespertilionids or rhinolophids, with a carnivore-like appearance; a well-developed cingulum is found both on its labial and lingual sides, extending on the distal side to form a short shelf. The current distribution of megadermatids is restricted to tropical and subtropical regions in Eastern Asia and central Africa. Fossils belonging to this family of bats are recorded in Oligocene and Miocene European sites, but are no longer found since sometime in the Early to Late Pliocene transition. Their disappearance from Europe was probably caused by climate change from subtropical to temperate conditions. Thus, the record of this megadermatid fossil in La Piquera supports the persistence of warm, subtropical climatic conditions in Iberia at least during the Early Pliocene.
Insectivores: The insectivore assemblage includes four different species, namely three soricids (Myosorex meini, Paenelimnoecus cf. pannonicus, Neomyini indet.) and one galericine erinaceid (Parasorex ibericus).
Myosorex meini (Fig. 6A) is the main insectivore species represented in La Piquera. Its most evident differential traits are the non-pigmented teeth, the transversely elongated upper molars (mainly M1 and M2), the position of the mental foramen under the p4, the short talonids of m1 and m2, and the presence of a tiny a2 hanged between the a1 and the p4 in excellently preserved specimens. The occurrence of Myosorex meini reinforces the post-Messinian age of the site because Myosorex is a genus with an African origin, which reached the Iberian Peninsula during the Mediterranean desiccation (Furió et al. 2007). The predominance of this insectivore (about two thirds of the total insectivore assemblage) compared with the low frequencies of its counterparts is indicative of rather dry environmental conditions.
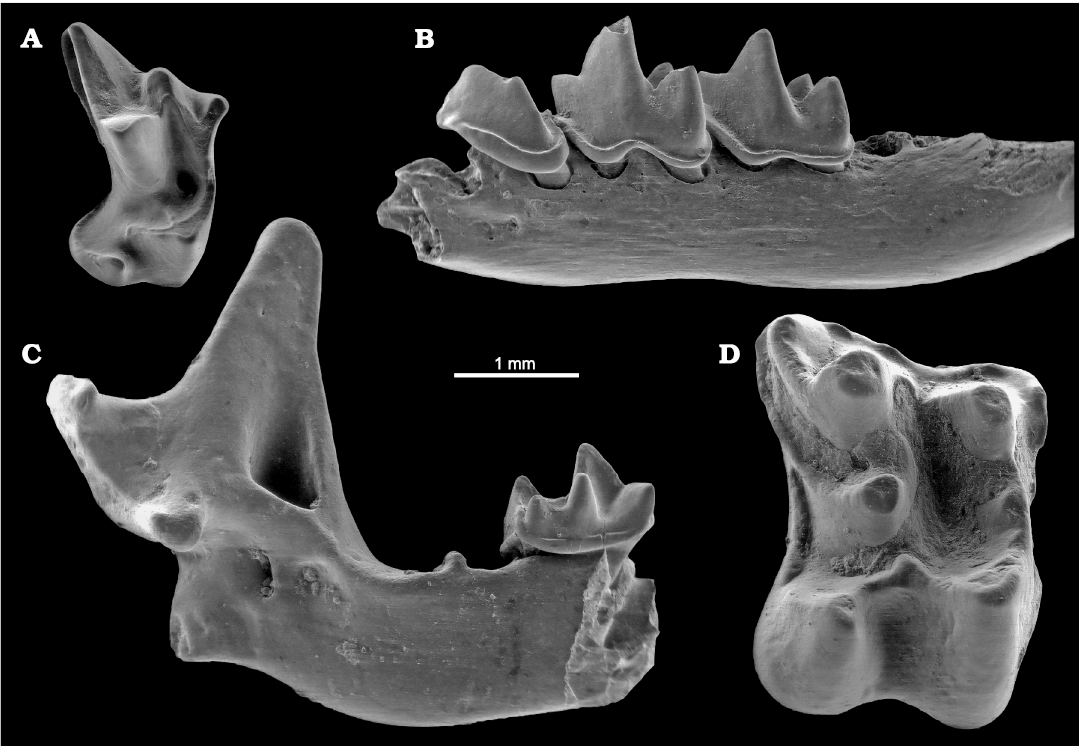
Fig. 6. ESEM images of insectivores from La Piquera site, Lower Pliocene, Duero Basin, Spain. A. Soricid Myosorex meini Jammot, 1977, UCM-LPQ-EUL-1, right M1 in occlusal view. B. Soricid Neomyini indet., UCM-LPQ-EUL-2, left mandibular fragment with p4–m2 in lateral view. C. Neomyini indet., UCM-LPQ-EUL-3, left mandibular fragment with m2 in lateral view. D. Erinaceid Parasorex ibericus (Mein and Martín-Suárez, 1993), UCM-LPQ-EUL-4, right M1 in occlusal view.
Paenelimnoecus is a frequent form in the Eurasian small mammal assemblages during most of the Miocene and Pliocene. Its small dimensions are only comparable with the Etruscan shrew (Suncus etruscus), the smallest present-day soricid. Other than its extremely reduced size, this extinct genus is characterized by the lack of entoconid cristids and the rather straight labial cingulids in m1 and m2. As a minor component of the insectivore assemblage, not many elements have been recovered hitherto, so the specific ascription is still to be confirmed. The ecological preferences of this extinct genus of tiny shrews are yet unknown, but according to its widespread distribution, the genus Paenelimnoecus rather thrived under all kinds of environmental conditions. In such sense it is to be noticed that this genus was likely one of the few soricids not strictly dependent on high environmental wetness as deduced by its frequent occurrence in Mio-Pliocene localities from the southeast of Spain influenced by dry external conditions (e.g., Minwer-Barakat et al. 2010; Van Dam et al. 2014; Furió and Agustí 2017; Piñero et al. 2018b).
The occurrence of an undetermined form of Neomyini (Fig. 6B, C) is clearly evidenced by several mandibular elements preserving the condyle, in which a small upper articular facet is separated from an elongated lower articular facet by a long and rather narrow interarticular area. Other typical traits within the tribe are the weak to intermediate degree of pigmentation of the teeth, the undulated labial cingulids, and the clear presence of entoconid cristids in m1 and m2. The first Neomyini occurrences in Spain correspond to the latest Miocene faunas (Furió and Agustí 2017). Hitherto, the only reported genus of this tribe in the Early Pliocene faunas of Spain is Asoriculus (Rofes and Cuenca-Bescós 2006), which was one of the most successful insectivores by the end of the Pliocene and the earliest Pleistocene (Furió et al. 2018). However, the available material from the studied locality does not preserve traits that are diagnostic at the genus or species level. In fact, other members of this tribe have been reported from different localities close to the transition from the Miocene to the Pliocene around Europe (e.g., Kowalski 1956; Rzebik-Kowalska 1998; Rzebik-Kowalska and Nesin 2010). Thus, even though the most likely genus of Neomyini present in La Piquera should be Asoriculus, other options cannot be discarded. In any case, the presence of neomyine forms is usually considered an indication of continental water bodies in the nearby area.
Parasorex (Fig. 6D) is a very frequent element within Middle Miocene to Pliocene small mammal assemblages from Spain. Parasorex ibericus is typical in Late Miocene and Early Pliocene faunas (Mein and Martín-Suárez 1993; Minwer-Barakat et al. 2012; Furió and Agustí 2017; Furió et al. 2018). Its occurrence in La Piquera is clearly evidenced by the rather large dental elements found and the transversely elongated upper molars with the posterior cingula interrupted by the posterior arm of the metaconule. As in the case of the soricid Paenelimnoecus, Parasorex is ecologically a eurytopic form, with a wide range of tolerance to external conditions. This galericine erinaceid is present in almost all the fossil sites close to the Mio-Pliocene transition from southwestern Europe, being better represented in fossil assemblages with low deduced environmental moistures.
Rodents: The rodent assemblage from La Piquera (Fig. 7) is highly diversified, comprising the cricetids Ruscinomys lasallei, Apocricetus cf. barrierei, and Blancomys aff. sanzi, the murids Stephanomys dubari, Apodemus gorafensis, Paraethomys meini, Occitanomys alcalai, and Castillomys gracilis, the gerbil Debruijnimys sp., the glirids Eliomys truci and Glis cf. minor, and the sciurid Atlantoxerus sp.
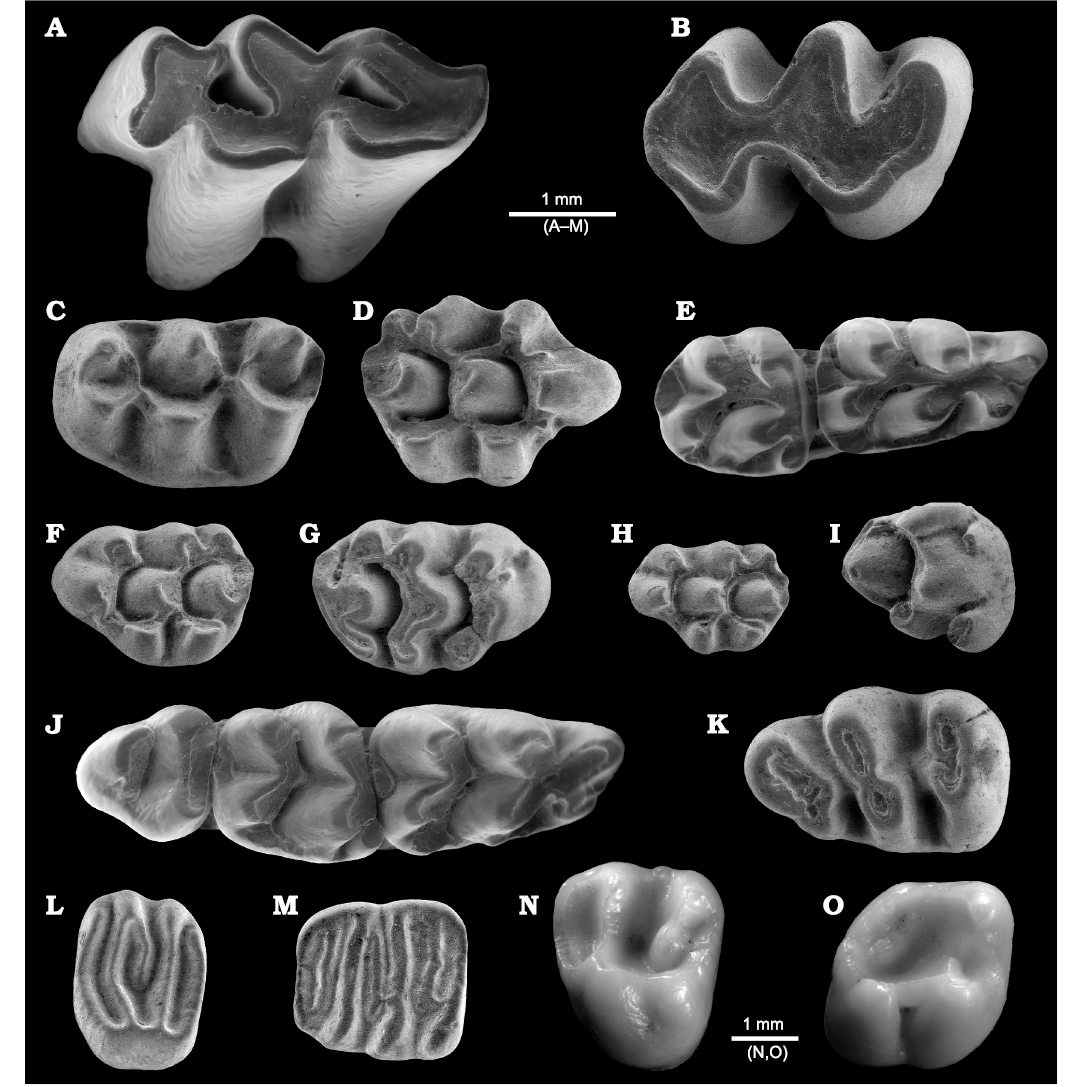
Fig. 7. ESEM images of rodents from La Piquera site, Lower Pliocene, Duero Basin, Spain, all in occlusal view. A. Cricetid Ruscinomys lasallei Adrover, 1969, UCM-LPQ-ROD-1, left M1. B. Cricetid Blancomys aff. sanzi Adrover, Mein, and Moissenet, 1993, UCM-LPQ-ROD-2, right M1. C. Cricetid Apocricetus cf. barrierei (Mein and Michaux, 1970), UCM-LPQ-ROD-3, left M1. D, E. Murid Stephanomys dubari Aguilar, Michaux, Bachelet, Calvet, and Faillat, 1991. D. UCM-LPQ-ROD-4, right M1. E. UCM-LPQ-ROD-5, right m1 and m2. F. Murid Occitanomys alcalai Adrover, Mein, and Moissenet, 1988, UCM-LPQ-ROD-6, left M1. G. Murid Apodemus gorafensis Ruiz Bustos, Sesé, Dabrio, Peña, and Padial, 1984, UCM-LPQ-ROD-7, right M1. H. Murid Castillomys gracilis van de Weerd, 1976, UCM-LPQ-ROD-8, left M1. I, J. Murid Paraethomys meini (Michaux, 1969). I. UCM-LPQ-ROD-9, right M2. J. UCM-LPQ-ROD-10, right m1, m2, and m3. K. Gerbillid Debruijnimys sp., UCM-LPQ-ROD-11, left m1. L. Glirid Eliomys truci Mein and Michaux, 1970, UCM-LPQ-ROD-12, left M1/2. M. Glirid Glis cf. minor, UCM-LPQ-ROD-13, left m1. N, O. Sciurid Atlantoxerus sp. N. UCM-LPQ-ROD-14, left M1/2. O. UCM-LPQ-ROD-15, right m1/2.
Ruscinomys lasallei (Fig. 7A) is the second most numerous rodent, with 15% of the total rodent abundance. This species is characterized by highly hypsodont teeth. In the M1 a deep groove separates the anterocone into two lobes. However, this groove does no reach the base of the crown, the two lobes of the anterocone becoming widely confluent as wear increases. The lingual lobe of the anterocone is rounded and smaller than the labial lobe. The anterior ectoloph reaches the paracone at lower height than the posterior ectoloph. In the m1, the anteroconid is rounded and does not present an anterolophid. A labially oriented mesolophid is present, connecting the paraconid at mid height of the crown. The genus Ruscinomys was erected by Depéret (1890) to define a large-sized and high-crowned cricetid from the Pliocene of Perpignan, Ruscinomys europaeus. Ruscinomys is common in the Late Miocene and Early Pliocene faunas of western Europe, showing increase in tooth size and crown height over time (Minwer-Barakat et al. 2009a). Ruscinomys lasallei was defined as a species intermediate between the Turolian R. schaubi and the Ruscinian R. europaeus. The continuous record of this genus from the Turolian to the lower Ruscinian in the Granada Basin supports the hypothesis of a lineage Ruscinomys schaubi–Ruscinomys lasallei–Ruscinomys europaeus (García-Alix et al. 2008b).
Apocricetus is scarcely represented in the rich sample from La Piquera, with only one M1 (Fig. 7C) and a fragment of m1 preserving the anterconoid, protoconid, and metaconid. The M1 lacks any evidence of cingulum ridge at the anterior wall of the tooth. The anterocone is double, the two lobes of the anterocone connecting posteriorly to a wide prelobe. The protolophule is posterior, and the mesoloph is absent. The tooth presents an anterior metalophule while the posterior metalophule is absent, the posteroloph connecting directly to the posterior wall of the metacone. Regarding the fragmentary m1, it presents a wide anteroconid slightly subdivided into three cusps. There is a very short prelophid, the two anterolophulids and the anterior arms of the protoconid and metaconid being almost confluent and drawing an “X” shape. The mesolophid is apparently absent. The shape of the M1 and the slight tripartition of the anteroconid in the m1 strongly point to Apocricetus barrierei, which is present in several Early Pliocene faunas known from the Betic and the Teruel basins (Freudenthal et al. 1998; García-Alix et al. 2008b; Mansino et al. 2017b; Piñero et al. 2017a; among others). However, more material will be necessary in order to confirm this assignment. This cricetid was originally identified in the Early Pliocene faunas of southern France, being first described as Cricetus barrierei (see Mein and Michaux 1970). Later, this species was included in Apocricetus by Freudenthal et al. (1998). Apocricetus barrierei arose from the smaller Apocricetus alberti close to the Miocene/Pliocene boundary (Freudenthal et al. 1998). The lowermost Pliocene site of Purcal-4 (between 5.2 and 5.3 Ma; Martín-Suárez et al. 1998) yielded the oldest population clearly assigned to Apocricetus barrieri (García-Alix et al. 2008b; Piñero and Agustí 2019). The species of Apocricetus constitute the anagenetic lineage Apocricetus aff. plinii–Apocricetus plinii–Apocricetus alberti–Apocricetus barrierei–Apocricetus angustidens. This lineage underwent gradual morphological and biometrical changes throughout the Late Miocene and Pliocene (MN11–MN16) (Freudental et al. 1998; Mansino et al. 2014).
The cricetid Blancomys aff. sanzi is surprisingly abundant among rodents from La Piquera, with 42 specimens (Fig. 7B). The genus is poorly known, its material being scarce and its record very discontinuous. The oldest record of Blancomys is found at the site of Almenara-Casablanca M, associated with a Late Miocene fauna including Apodemus gudrunae, Stephanomys ramblensis, Paraethomys meini, Apocricetus alberti, Debruijnimys almenarensis, Myocricetodon jaegeri, Pseudomeriones abbreviatus, and Calomyscus sp. (Agustí et al. 2011; Minwer-Barakat et al. 2018). As in the case of allochtonous elements such as Debruijnimys almenarensis, Pseudomeriones abbreviatus, Myocricetodon jaegeri, and Calomyscus sp., its entry in the Iberian Peninsula is most probably related with the Messinian Salinity Crisis, which opened new dispersal routes with Africa and Asia (Agustí 1989; Agustí et al. 2006; García-Alix et al. 2016; Booth-Rea et al. 2018). Blancomys disappeared in the Late Pliocene. Other than Almenara-Casablanca M, the species of Blancomys are present in several sites from eastern and, although more rarely, southern Spain, including the Teruel, Alcoy, Granada, and Guadix-Baza basins (Mazo et al. 1985; Adrover 1986; Sesé 1989; Castillo 1990; Mein et al. 1990, Adrover et al. 1993b; García-Alix et al. 2008b; Maldonado-Garrido et al. 2017; among others). In contrast, Blancomys appears to be absent from the rich late Neogene sequences of the Fortuna Basin (Agustí et al. 2006; Piñero and Agustí 2019). Blancomys sanzi (type locality La Gloria 5, Teruel Basin) is so far the most archaic species of the genus, being smaller and less hypsodont than the species B. neglectus (type locality Layna) and B. meini (type locality Sarrión) (Van de Weerd et al. 1977; Adrover 1986). The abundant material coming from La Piquera coincides in shape with the sample from the type locality of La Gloria 5, although it presents smaller dimensions, suggesting that we are dealing with a more archaic population.
Stephanomys dubari (Fig. 7D, E) represents the most abundant rodent at La Piquera, making 65% for the whole rodent association, and 85% of the total murid sample. The teeth show the main features of Stephanomys, including high crown, development of longitudinal crests in the lower teeth, and presence of posterior crests and spurs on the tubercles 1 and 3 (t1 and t3) in the upper teeth. The material from La Piquera is intermediate between Stephanomys ramblensis and Stephanomys cordii both in size and in stephanodonty and hypsodonty, pointing towards Stephanomys dubari, a taxon defined by Aguilar et al. (1991) as an intermediate species between S. ramblensis and S. cordii. The morphology of the molars from La Piquera is also consistent with S. dubari from its type locality, Castelnou 3 (Aguilar et al. 1991), and other Mio-Pliocene localities (Mansino et al. 2017a; García-Alix et al. 2008c; Piñero and Agustí 2019). The other Stephanomys species from the Pliocene and Pleistocene are larger, have higher crowns and more pronounced stephanodonty than the studied material. The sample of La Piquera is undoubtedly the most extensive collection of S. dubari recorded to date. The temporal distribution of this frequent murid extends from the Late Miocene to the Early Pliocene, being first recorded at Venta del Moro (6.2 Ma; Gibert et al. 2013; Mansino et al. 2017a). The transitional replacement of Stephanomys dubari by Stephanomys cordii took place sometime between 4.9 and 4.6 Ma (Piñero and Agustí 2019). García-Alix et al. (2008c) proposed the anagenetic evolutionary line Occitanomys adroveri–Stephanomys ramblensis–Stephanomys dubari–Stephanomys cordii. This lineage underwent a continuous increase in size, crown height, and stephanodonty over time, which is supported by the very complete and continuous record of Stephanomys from the Fortuna Basin. The oldest evidence of Stephanomys has been documented at the late Turolian site of Sifón-1, at about 6.9–6.7 Ma (Piñero and Agustí 2019).
The murid Occitanomys alcalai is well represented at La Piquera (Fig. 7F), being the second murid in abundance (48 specimens). The sample from La Piquera differs from Occitanomys hispanicus in the better development of the t6–t9 connection. The teeth are similar in size to Occitanomys sondaari but, unlike that species, they have well-developed labial cingula in the lower molars, and t1bis and t6–t9 connection in the upper molars (Van de Weerd 1976). Occitanomys brailloni can be distinguished from Occitanomys species from La Piquera by its larger size and more frequent and better developed longitudinal connections (Michaux 1969). Occitanomys adroveri is larger and has more frequency and better development of longitudinal connections in the lower molars. In addition, the t1 in the M1 is in a more anterior position (Van de Weerd 1976). The specimens from La Piquera are consistent with some of the diagnostic features of O. alcalai, including the relatively high crown, well-developed t1bis, absence of isolated tubercles, well-developed spurs in the upper molars, and symmetrical anteroconid (Adrover et al. 1988). The size is also close to that of O. alcalai from Peralejos E (type locality; Adrover et al. 1988). This small-sized Occitanomys species is very common in Late Miocene and Early Pliocene Iberian faunas, having a wide stratigraphic range (Van de Weerd 1976; Adrover et al. 1988, 1993b; García-Alix et al. 2008c; Minwer-Barakat et al. 2009b; Mansino et al. 2015a, 2016b, 2017a; among others). Alike Stephanomys ramblensis, its oldest record has been reported at Sifón-1 (MN13; Piñero and Agustí 2019), whereas its last occurrence is recorded at the Lower Pliocene site of Gorafe-4 (upper MN14), between 4.30 and 4.19 Ma (Martín-Suárez 1988; Piñero et al. 2018a). Occitanomys is known also from the Upper Pliocene (MN16), with the species Occitanomys brailloni.
Apodemus gorafensis is scarcely represented at La Piquera (20 specimens) (Fig. 7G). The molars lack a longitudinal crest in the lower teeth and have a well-developed medial anteroconid (tma) in the m1, two diagnostic traits of Apodemus (Martín-Suárez and Mein 1998). These molars are larger or slightly larger than in the Miocene species Apodemus lugdunensis, Apodemus barbarae, and Apodemus meini, the Mio-Pliocene Apodemus gudrunae, and the Plio-Pleistocene Apodemus atavus (Van de Weerd 1976; Martín-Suárez 1988; Martín-Suárez and Freudenthal 1993; Piñero and Agustí 2020; Agustí et al. 2022; among others). Apodemus agustii differs from the Apodemus sample of La Piquera by the presence of less pronounced stephanodonty in the upper molars, and a funnel delimited by the anteroconid, protoconid, and metaconid in m1 (Martín-Suárez 1988). The specimens from La Piquera fit the size of A. gorafensis from its type locality, Gorafe-A (Ruiz-Bustos et al. 1984). In addition, they show typical traits of A. gorafensis: presence of a well-developed labial cingulum and a large tma and c1 in the m1, presence of elongated t7 and posterior spur in the t3 of the M1, and weak t6–t9 connection in the M2. This species was first described by Ruiz-Bustos et al. (1984) from the Lower Pliocene level of Gorafe-A (Guadix-Baza Basin), having been subsequently identified in several Late Miocene and Pliocene faunas from the Iberian Peninsula (Adrover et al. 1993b; García-Alix et al. 2008c; Mansino et al. 2015b; Piñero et al. 2017a; among others). The oldest record of A. gorafensis was reported at the Upper Miocene level of Sifón-3 (MN13, upper Turolian), at about 6.7 Ma (Piñero and Agustí 2019). Martín-Suárez and Mein (1998) suggested that this murid arose from A. gudrunae. However, Piñero and Agustí (2019) proposed that A. gorafensis was either an immigrant to the Iberian Peninsula or the result of a quick cladogenetic process, based on evidence from the Fortuna Basin.
Castillomys gracilis (Fig. 7H) is relatively well represented in La Piquera. The material consists of 28 isolated teeth, with low crown, small size, absence of t7, development of posterior spur in the t1 of the M2, and presence of a longitudinal spur in the m1. This morphology is consistent with that of Castillomys. The specimens from La Piquera differ from Castillomys crusafonti, Castillomys rivas, and Castillomys africanus by their smaller size and lesser development of the t1–t5 and t3–t5 connections (Michaux 1969; Martín-Suárez and Mein 1991; Coiffait 1991; Minwer-Barakat et al. 2005). The presence of an isolated t3 in the M2, and the incomplete longitudinal connection in some m1 are in accordance with Castillomys gracilis. Moreover, the size is similar to that of C. gracilis from its type locality, Caravaca (Van de Weerd 1976). Castillomys gracilis is considered a typical species of the Lower Pliocene in the Iberian Peninsula (Van de Weerd 1976; Mein et al. 1990). Apparently, the first occurrence of the genus Castillomys coincides with the beginning of the Pliocene (Piñero and Agustí 2019). Martín-Suárez and Mein (1991) proposed the evolutionary lineage Castillomys gracilis–Castillomys crusafonti–Castillomys rivas. Along this phyletic lineage, an increase in size and a better development of the longitudinal connections in all molars occur. Castillomys from La Piquera would apparently represent the earliest record of this genus, which became extinct at the Early/Middle Pleistocene boundary (Agustí et al. 1999).
Paraethomys meini represents the least abundant murid at La Piquera, with only five identified specimens (Fig. 7I, J). The material is consistent with the typical features of Paraethomys: t9 absent in the M2, tma absent in the m1, weak or moderate labial cingulum in the lower molars, and presence of a small funnel of enamel in the m1. The specimens from La Piquera are smaller and have the distal spurs on the t1 and t3 less developed than in Paraethomys baeticus, Paraethomys abaigari, and Paraethomys jaegeri (Montenat and de Bruijn 1976; Adrover et al. 1988; Piñero and Verzi 2020). Paraethomys from La Piquera differs from Paraethomys balearicus in having slightly smaller size and lower-crowned molars (Torres-Roig et al. 2019). Paraethomys belmezensis can be distinguised from the studied molars by the presence of a well-developed tma connected to the lingual lobe of the anteroconid (Castillo Ruiz 1992). The molars from La Piquera are close in size and morphology to Paraethomys meini from its type locality, Sète (France; Michaux 1969). The genus Paraethomys was a widely distributed murid in both southwestern Europe and northern Africa since the Late Miocene. The populations on both shores of the Mediterranean underwent divergent evolutionary processes after their split at the end of the Messinian Salinity Crisis (De Bruijn 1973; Jaeger et al. 1975; Coiffait and Coiffait 1981; Renaud et al. 1999; among others). The North African representatives of the genus reached the late Middle Pleistocene, whereas the European lineage became extinct during the Pliocene. Paraethomys meini dispersed into Europe probably from northern Africa, prior to the Messinian Salinity Crisis, at about 6.2 Ma (Garcés et al. 1998, 2001; Agustí et al. 2006; Gibert et al. 2013). It is the oldest representative of the genus in Europe, whose stratigraphic range extends from the upper Turolian (MN13, Upper Miocene) to the upper Ruscinian (MN15, Pliocene). Paraethomys became more diversified during the Pliocene, when larger species such as Paraethomys baeticus, Paraethomys jaegeri, Paraethomys abaigari, Paraethomys belmezensis, and Paraethomys balearicus occurred (Montenat and De Bruijn 1976; Castillo-Ruiz 1992; Adrover et al. 1988; Torres-Roig et al. 2019; Piñero and Verzi 2020). The large-sized Paraethomys baeticus would have diverged from Paraethomys meini through asymmetric speciation at about 4.6 Ma, representing the first form of the phyletic lineage Paraethomys baeticus–Paraethomys abaigari–Paraethomys jaegeri, which is characterized by a gradual increase of size and a progressive development of longitudinal connections in the molars along the Pliocene. The genus must have vanished from Europe at some moment slightly before the Ruscinian/Villanyian boundary (Piñero and Verzi 2020).
Debruijnimys was established by Castillo and Agustí (1996) on the basis of the species Debruijnimys julii, from the Upper Pliocene site of Asta Regia (Jerez Basin). Besides the type-species, the formerly described Protatera almenarensis was also included in this genus by Agustí and Casanovas-Vilar (2003). In the same paper, these authors also recognized a third unnamed species, Debruijnimys sp., from the site of Alcoy 4B. This species was smaller and had less developed longitudinal spurs than D. julii. Moreover, it also differs from D. almenarensis because of its more reduced size and smaller, rounded anteroconid. Isolated teeth of this small Debruijnimys species are also present at the Early Pliocene faunas of Sifón-413 (Piñero and Agustí 2019), Botardo D (Piñero and Agustí 2020), and Puerto de la Cadena (Piñero et al. 2017b). The specimens from La Piquera coincide in size and shape with the scarce material coming from Alcoy 4B and the other above-mentioned sites. Debruijnimys sp. (Fig. 7K) from La Piquera is the largest sample of this unnamed species recovered to date (ten isolated teeth). The genus Debruijnimys is of African origin and probably dispersed into the Iberian Peninsula during the Messinian Salinity Crisis (Agustí et al. 2006; Minwer-Barakat et al. 2009a). The oldest species of the genus is Debruijnimys almenarensis (Agustí 1990), which is restricted to a short interval close to the Miocene/Pliocene boundary. However, this genus persisted over the Pliocene with Debruijnimys sp. and Debruijnimys julii (Castillo and Agustí 1996; Piñero et al. 2017a, Mansino et al. 2015b). Agustí and Casanovas-Vilar (2003) proposed two different dispersal waves, so that in the first immigration wave Debruijnimys almenarensis reached the Iberian Peninsula, whereas Debruijnimys sp. arrived during the second one. Debruijnimys sp. probably evolved into Debruijnimys julii sometime between 4.8 and 4.4 Ma (Piñero and Agustí 2020), the latter persisting until the late Ruscinian (MN15) (Castillo and Agustí 1996).
Eliomys truci (Fig. 7L) is the dominant glirid in the association of La Piquera, but its prevalence among the total sample of rodents is only around 1.5% (six isolated teeth). The M1/2 presents a very simple dental pattern, composed of anteroloph, protoloph, precentroloph, metaloph, and posteroloph. All these lophs (except the precentroloph) are lingually connected by a continuous endoloph. The precentroloph is labially fused to the protoloph. There are no clearly differentiated cusps. The M3 also presents a very simple dental pattern, composed of anteroloph, protoloph, postcentroloph, metaloph, and posteroloph. The postcentroloph is labillay fused to the metaloph. As in the case of the M1/2, no clearly differentiated cusp can be recognized. Eliomys truci was originally described in the Early Pliocene fauna of Hautimagne (southern France, Mein and Michaux 1970). Later, this species has been recognized in the Upper Miocene and Lower Pliocene of a number of Neogene basins in Spain: Teruel (Van de Weerd 1976; Adrover 1986; Adrover et al. 1993b), Fortuna (Piñero et al. 2018b; Piñero and Agustí 2019), Guadix-Baza (García-Alix et al. 2008b; Piñero and Agustí 2020), and Granada (García-Alix et al. 2008b). It differs from the younger species of the genus (Eliomys yevesi, Eliomys intermedius, and Eliomys quercinus) by its smaller size and more complex dental pattern. The stratigraphic range of Eliomys truci is very wide. The oldest doubtless record of this dormouse comes from the early Vallesian fauna of Pedregueras 2A (MN9; Daams and Freudenthal 1988), whereas its youngest occurrence is recorded at Tollo de Chiclana-13 (MN16, lower Villanyian; García-Alix et al. 2008b).
Glis is represented at La Piquera by a single m1 (Fig. 7M). The dental pattern is composed of seven ridges: anterolophid, anterotropid, metalophid, centrolophid, mesolophid, posterotropid, and posterolophid. The anterolophid and metalophid are lingually fused by an endolophid; this is also the case for the metalophid and the posterolophid. The dimensions of the tooth (1.77 × 1.60 mm) fit the values of the species Glis minor from the type locality of Podlesice (Poland), although more material will be necessary in order to confirm this assignment. Glis minor was originally defined at the Pliocene site of Podlesice as a subspecies Glis sackdillingensis minor (see Kowalski 1956). However, in his revision of the Pliocene and Pleistocene dormice from Poland, Kowalski (1963) decided to elevate the rank of this taxon to the species level based on its smaller dimensions. Glis minor is a widely distributed species, having been identified in the Plio-Pleistocene of France (Les Valerots; Chaline 1972), Italy (Moncuco Torinese; Angelone et al. 2011), Germany (Gundersheim 4 and Sondershausen; Fejfar and Storch 1990; Hellmund and Ziegler 2012), Austria (Richardhof-Golfplatz, Richardhof-Wald, Kohfidisch; Daxner-Höck and Höck 2009), Poland (Podlesice, Węże, Rębielice; Kowalski 1963), and Greece (Tourkobounia-1, Notio 1 and Komanos 1; De Bruijn and van der Meulen 1975; Hordijk and De Bruijn 2009). The extant Glis glis lives in forests and open woodland, and is mainly arboreal.
Sciurids are represented by scarce (one dp4, one m1/2 and two M1/2) but well-preserved material (Fig. 7N, O). Its biometric homogeneity suggests that it corresponds to a single taxon. The lower molars show an entolophid, which, according to Cuenca-Bescós (1988), is a distinguishing feature of the tribe Xerini. Within faunas from the final part of the Late Miocene and the beginning of the Early Pliocene, this tribe is represented in the Iberian Peninsula by Atlantoxerus and Heteroxerus (Sesé 2006), which share a very similar dental pattern (Peláez-Campomanes 2001). Typically, the molars of Atlantoxerus species have larger dimensions than those of Heteroxerus, although some Atlantoxerus species share with Heteroxerus their reduced sizes. The sciurid molars from La Piquera have larger dimensions than those of the Heteroxerus species, excluding their attribution to that genus, so they are ascribed to Atlantoxerus. During the Late Miocene and Early Pliocene, two species of Atlantoxerus occurred in the study area: Atlantoxerus margaritae and Atlantoxerus adroveri (De Bruijn and Mein 1968; Van de Weerd 1976; Adrover et al. 1993a; García-Alix et al. 2007; Minwer Barakat et al. 2009a; Mansino et al. 2015b, 2016a; among others). A third species, Atlantoxerus cuencae, has been reported from the Late Miocene faunas of southern France (Aguilar et al. 1995). The size of the La Piquera lower (L = 2.75 mm; W = 2.92 mm; n = 1) and upper molars (L = 2.53 mm; W = 3.1 mm; n = 2; mean values) is similar to some populations of the species A. adroveri (Concud 3; Van de Weerd 1976) and A. cf. margaritae (Villalba Alta; Adrover et al. 1993a), but the small size of the sample prevents further identification of this material. Nowadays, the ground squirrel Atlantoxerus is represented by the single species Atlantoxerus getulus from North Africa. It lives under arid or semiarid conditions, preferring sparsely vegetated areas of the Atlas Mountains (De Bruijn 1999; Krystufek et al. 2016).
Lagomorphs: The lagomorph assemblage from La Piquera includes three distinct forms that differ in size. The largest corresponds to a leporid because, in its lower dentition, the talonids and trigonids of its molariform teeth are joined by a small lingual isthmus, and the third lower molar is bilobate (López-Martínez 1989; Laplana et al. 2015). Within the faunas from the final part of the Late Miocene and the beginning of the Early Pliocene, leporids are represented in the Iberian Peninsula by two genera, Alilepus and Trischizolagus (López-Martínez 1989, 2008; Sesé 2006). The presence of an anteroflexid and a small paraflexid on p3 (Fig. 8A) rule out the inclusion of the La Piquera leporid in Alilepus, whose representatives lack these structures (López-Martínez 1989; Čermak et al. 2015) and confirm its attribution to Trischizolagus. The scarcity of material (only one p3, the most diagnostic element, is available to date, Fig. 8A) prevents us from specifying the attribution of this form beyond its generic ascription.
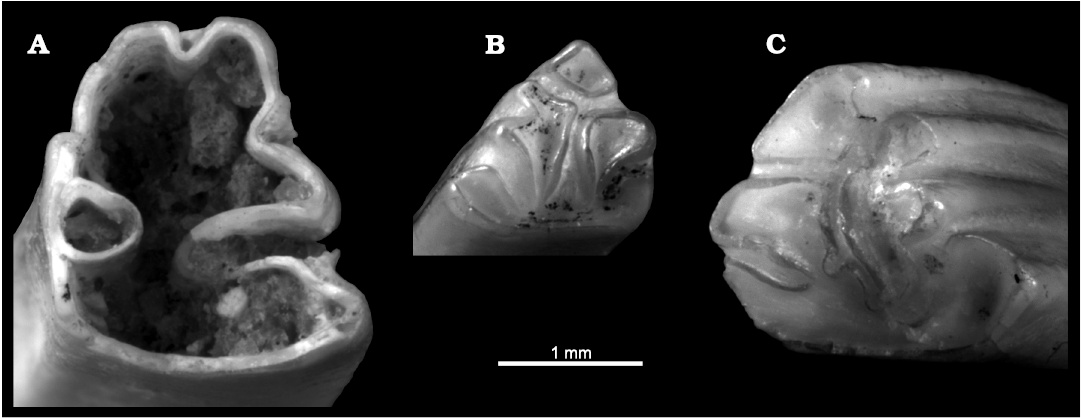
Fig. 8. Lagomorphs from La Piquera site, Lower Pliocene, Duero Basin, Spain. A. Leporid Trischizolagus sp., UCM-LPQ-LAG-1, basal view of an unworn left p3. B. Prolagid Prolagus sp. 1, UCM-LPQ-LAG-2, left p3 in occlusal view. C. Prolagid Prolagus sp. 2, UCM-LPQ-LAG-3, right p3 in occlusal view.
Trischizolagus is a widespread, extinct leporid. The geographic distribution of the genus includes much of the Palearctic region, having been recorded from western Europe to China, and probably northwestern Africa (López-Martínez 1989; Geraads 1994; Averianov and Tesakov 1997; Qiu and Storch 2000; Čermák and Wagner 2013; Čermák et al. 2019; among others). The Iberian Trischizolagus record includes the most archaic species of the genus, i.e., Late Miocene Trischizolagus crusafonti and Early Pliocene Trischizolagus maritsae (Janvier and Montenat 1971; López-Martínez 1989; Piñero et al. 2017b). These two species are smaller than the type species Trischizolagus dumitrescuae from the Late Pliocene of eastern Europe (Radulesco and Samson 1967). Although no transitional populations have yet been found, Trischizolagus has been proposed as a probable ancestor of Oryctolagus and Lepus, as well as for other leporines (López-Martínez 2008). The main habitats of these extant lagomorphs are open lands such as Mediterranean shrubs.
Concerning the other two lagomorph species from La Piquera, their smaller size, the absence of any isthmus connecting the two lobes of the lower molariform teeth, the presence of three lobes in the m2, and the absence of m3 allow them to be included in the family Prolagidae (López-Martínez 1989; Laplana et al. 2015), while the presence of three labial and two lingual flexids allows them to be attributed to Prolagus (López-Martínez 1989). The most conspicuous difference between the two forms of Prolagus from La Piquera (Prolagus sp. 1 and 2) is their size (Fig. 8B, C). Ongoing studies on the dental morphology of both forms will make it possible to determine their specific ascription.
Prolagus is one of the most common Neogene and Quaternary European mammals (López-Martínez 2001). It was a long-lived, widely distributed and abundant prolagid genus that inhabited Europe, western Asia and North Africa (Angelone 2008). Like most pikas, the majority of Prolagus species are small sized. The Prolagus lineage includes about 22 different species, being present in Europe since the Early Miocene to the Holocene (López-Martínez 1989, 2001; Angelone 2008; among others). According to López-Martínez (2001), Prolagus species were usually thermophile wetland dwellers.
Artiodactyls: The large mammal remains from La Piquera are very scarce, comprising only three specimens, corresponding to three bovid teeth fragments: a left DP3 (length = 7.2 mm: width = 6.4 mm), which can be attributed to an antilopini close to Gazella spp. or Hispanodorcas spp. according to its morphology and size (Alcalá 1994); a P3 fragment (length = 14.4 mm) belonging to a larger sized bovid with a moderately brachydont dentition; and an enamel fragment of a selenodont tooth, which could be decidual, very polished due to erosion, and that can be identified as a bunodont bovid upper molar.
Discussion
Stratigraphy of La Piquera site.—The cosmopolitanism, high evolutionary rates, and rapid diversification of small mammals, and particularly rodents, make them an excellent tool for correlating European faunas. Thereby, the stratigraphy of La Piquera can be estimated based on its micromammal content.
The rodents Ruscinomys lasallei, Eliomys truci, Stephanomys dubari, Occitanomys alcalai, Apodemus gorafensis, and Paraethomys meini are considered to be common taxa of the Miocene–Pliocene transition in the Iberian Peninsula. These rodents can be found in both Upper Miocene (upper Turolian, MN13) and Lower Pliocene (lower Ruscinian, MN14) localities (Martín-Suárez 1988; Mein et al. 1990; Adrover et al. 1993b; Agustí et al. 2006; García-Alix et al. 2008b, c; Minwer-Barakat et al. 2009a, b, 2012; Mansino et al. 2015a, b, 2016b, 2017a, b; Piñero and Agustí 2019, 2020; Piñero et al. 2017b, 2018a; among others). However, considering biostratigraphic criteria, the presence of Castillomys gracilis implies an Early Pliocene age for La Piquera fauna. Although there are up to three samples of Castillomys gracilis presumably identified in the Late Miocene fauna (Salobreña, Almenara Casablanca-M, and La Dehesa-16; Aguilar et al. 1984; García-Alix 2006; Agustí et al. 2011), the assignment of this material to the species is doubtful, most probably belonging to Occitanomys alcalai (see Piñero and Agustí 2019). Thereby, the first occurrence of Castillomys gracilis would correspond to the Lower Pliocene, as was originally proposed by Van de Weerd (1976) and Mein et al. (1990). The presence of Debruijnimys sp. is a further evidence supporting the Early Pliocene age of La Piquera fauna, since so far this gerbil has only been identified in sites assigned to the lower part of the MN14 unit (lower Ruscinian) (Agustí and Casanovas-Vilar 2003; Piñero and Agustí 2019, 2020; Piñero et al. 2017b). In addition, Apocricetus barrierei has been used to characterize the Lower Pliocene in the Iberian Peninsula, being indicative of an early Ruscinian age (see García-Alix et al. 2008b and references therein).
The insectivore assemblage also supports this chronology. The simultaneous occurrence of Parasorex, Myosorex, Paenelimnoecus, and a Neomyini species (usually Asoriculus gibberodon) is typical of the Lower Pliocene in Spain (Furió et al. 2018). Whereas Myosorex and Asoriculus (Neomyini, in general) find their first occurrences at MN13, contemporary to the Messinian Salinity Crisis, Parasorex and Paenelimnoecus have their last occurrences at MN15 and MN16, respectively. Thus, the whole insectivore assemblage is consistent with an early Ruscinian age (MN14), when the four forms are rather frequent and abundant in the Iberian Peninsula.
At La Piquera there is no clear evidence of any reptile taxa with African affinities (waiting for a better attribution of snake material), that could be linked with a Messinian dispersal event, maybe with the exception of the potential erycid, as proposed by Szyndlar and Schleich (1994), and if the two snake taxa that were described above as Colubridae/Psammophiidae are indeed psammophiids, then they are the product of a Messinian dispersal from Africa to Iberia, as was recently demonstrated for the psammophiid Psammophis odysseus from Salobreña (southern Spain; Georgalis and Szyndlar 2022).
In the Fortuna Basin, the lowermost Pliocene level of Sifón-413 (5.33–5.23 Ma; Garcés et al. 2001; Piñero and Agustí 2019) shares with La Piquera the presence of Occitanomys alcalai, Paraethomys meini, Stephanomys dubari, Debruijnimys sp., Apocricetus cf. barrierei, Ruscinomys cf. lasallei, and Eliomys truci. Sifón-413 provided a large sample of rodent teeth belonging to a diversified assemblage in which the murid Castillomys gracilis is still absent, thus indicating a somewhat older age than La Piquera material.
The rodent assemblage from Puerto de la Cadena (southeastern Spain), dated to 5.0–4.9 Ma (Piñero et al. 2017b), is very similar to that of La Piquera. Similar taxa have been recognized in this level such as Apocricetus cf. barrierei, Debruijnimys sp., Paraethomys cf. meini, Stephanomys cf. dubari, and Ruscinomys lasallei, suggesting that they have a very close age. Similarly, in the Teruel Basin, the early Ruscinian locality of Peralejos-E shares with La Piquera some taxa such as Apocricetus barrierei, Apodemus gorafensis, Paraethomys meini, Stephanomys dubari (according to García-Alix et al. 2008c), and Occitanomys alcalai (Adrover et al. 1988; Mein et al. 1990). Therefore, both sites are to be placed in a close biostratigraphic position.
The localities of Purcal-4 and 7 and Calicasas-3 and 4 (García-Alix et al. 2008a–c), in the Granada Basin, have some taxa in common with La Piquera such as Occitanomys alcalai, Stephanomys dubari, Paraethomys meini, Apodemus gorafensis (Purcal-4, Calicasas-3), Castillomys gracilis (Calicasas-3 and 4), Ruscinomys lasallei (Purcal-4 and Calicasas-3), and Apocricetus barrierei (Purcal-4 and 7). Thus, these sites must be very close in age. Purcal-4, a level correlated to the base of the Pliocene, has been dated to between 5.2 and 5.3 Ma (Martín-Suárez et al. 1998). In the Guadix-Baza Basin, Botardo-D is dated between 4.79 and 4.63 Ma (Piñero et al. 2018b; Piñero and Agustí 2020) and has some elements in common with La Piquera such as Apocricetus barrierei, Paraethomys meini, Apodemus gorafensis, Castillomys gracilis, Debruijnimys sp., and Eliomys truci. However, Botardo-D includes Stephanomys cordii, descendant of Stephanomys dubari, suggesting a slightly younger age than La Piquera.
The nearby locality of Castillejo de Robledo (Luengo et al. 2009), in the Duero Basin, records some species also found in La Piquera. However, the Stephanomys species from Castillejo de Robledo was assigned to S. medius, which, without biometric data nor images, does not allow us to ascertain whether it corresponds to Stephanomys dubari or its descendant Stephanomys cordii. A more accurate study of the rodent fauna from this site would be required to be able to correlate precisely Castillejo de Robledo with La Piquera. Nevertheless, the record of the microtoid cricetid Celadensia at Castillejo de Robledo confirms its earliest Pliocene age (Mein et al. 1990), so these two sites would be stratigraphically close.
All the above-mentioned sites are older than those reporting two coexisting Paraethomys species (Paraethomys meini and Paraethomys baeticus; see Piñero and Verzi 2020 and references therein). In La Piquera, two Paraethomys species are not yet distinguished. Despite this locality having yielded a huge number of rodent teeth, no specimen could be assigned to Paraethomys baeticus. This murid is a biostratigraphic marker of the upper part of the MN14 unit widely distributed in the Iberian Peninsula, whose chronological span ranges from ca. 4.6 to 4.3 Ma (Piñero and Verzi 2020). In this regard, La Piquera fauna is older than the Early Pliocene faunas recording Paraethomys baeticus such as Sifón Pista in the Fortuna Basin (Piñero and Agustí 2019), La Bullana 3 and 2B in the Cabriel Basin (Mansino et al. 2015b), La Gloria-4 and Celadas-9 in the Teruel Basin (Adrover et al. 1993b), Gorafe A and Baza-1 in the Guadix-Baza Basin (Ruiz Bustos et al. 1984; Piñero et al. 2017a), Calicasas-5A and Purcal-13 in the Granada Basin (García-Alix et al. 2008a), and Alcoi Barranc Sud-2, 3, and 3A in the Alcoy Basin (Mansino et al. 2015a), among others.
The rodent association from La Piquera therefore indicates an intermediate biostratigraphic position between the Lower Pliocene levels of Sifón-413 (5.33–5.23 Ma) and Botardo-D (4.79–4.63 Ma), the new locality being correlated with the lower part of the MN14 unit (lower Ruscinian) (see Fig. 9).
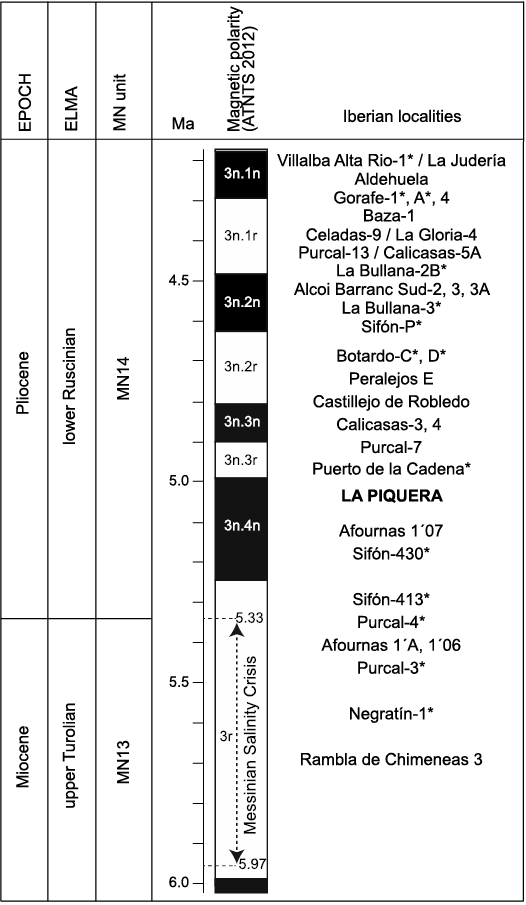
Fig. 9. Biostratigraphic position of the La Piquera locality, placing other Spanish sites for comparison. The localities marked with an asterisk are calibrated with paleomagnetic data (Opdyke et al. 1997; Martín-Suárez et al. 1998; Garcés et al. 2001; Mansino et al. 2015b; Piñero et al. 2017b, 2018a). Abbreviations: ATNTS, Astronomically Tuned Neogene Time Scale; ELMA, European Land Mammal Ages; MN, Mammal Neogene.
Palaeoenvironment.—Small vertebrates have been widely used as palaeoclimatic and palaeoecological indicators (e.g., López-García et al. 2008). Amphibian and reptile species are particularly useful for palaeoenvironmental reconstructions (Blain et al. 2009, 2016; Sánchez-Bandera et al. 2020; among others). Small mammal assemblages also provide interesting information on the past environmental and climatic conditions (e.g., Martín-Suárez et al. 2001; Minwer-Barakat et al. 2005; Montuire et al. 2006; Furió et al. 2011; Piñero et al. 2016). Here, we provide a preliminary palaeoenviromental inference based on the ecological requirements of the small vertebrate taxa.
As a whole, the herpetofaunal assemblage from La Piquera suggests a relatively dry and warm, open environment (Pelodytes, Bufotes viridis, possibly also agamids, scincids and lacertids, Blanus, and ?Erycidae) evolving laterally to more humid scrubby and open-woodland conditions (Ophisaurus). Amphibians (Pleurodeles and Discoglossus) document the nearby presence of permanent water. Scincids and Blanus also indicate the presence of loose (sandy and stony) soils that they nowadays use for their thermoregulation and as shelter (Pleguezuelos and Martínez-Rica 1997; Salvador 1997; Carrascal and Salvador 2002–2016; García-París et al. 2004; Pleguezuelos et al. 2004). Mean annual precipitation, using the Paleoprecipitation tool method (Böhme et al. 2006), can be estimated around 374 mm, a rather low value especially when compared to the 700–750 mm observed today in the region of study.
As for insectivores, the soricid Paenelimnoecus and the erinaceid Parasorex are ecologically eurytopic forms, with wide ranges of tolerance to external conditions. However, the dominance of the soricid Myosorex meini (2/3 of insectivores) is indicative of rather dry environmental conditions. This would be supported by the complete absence of desmanines, talpids that are linked to permanent water streams and that are abundant in the lowermost Pliocene in other areas of the Iberian Peninsula, such as the Teruel and Granada basins (Rümke 1985; Minwer-Barakat et al. 2020). The presence of continental water bodies in the nearby area is attested by the occurrence of Neomyini indet. It is supported by the occurrence of Prolagus, a lagomorph linked to wetlands.
The presence of a megadermatid among the chiropterans supports the presence of warm subtropical conditions when the deposit was formed. All Recent megadermatids have tropical and subtropical distributions, some of which linked to savanna environments thus agreeing with the palaeoenvironmental conditions indicated by the accompanying taxa.
The rodent assemblage is mostly dominated by Ruscinomys and Stephanomys, both taxa considered to be inhabitants of open grassy areas (Van Dam and Weltje 1999; Piñero et al. 2017a). The two taxa account for a prevalence of about 80% among rodents. Although less abundant, Castillomys gracilis, Occitanomys alcalai, Blancomys sanzi, and Apocricetus barrierei are also interpreted as dwellers of open herbaceous meadows (García-Alix et al. 2008d and references therein; Piñero et al. 2017a). The open, bare habitat is represented by Debruijnimys sp. and Atlantoxerus sp. (<1%). Likewise, the woodland elements (according to García-Alix et al. 2008d) are poorly represented (1.5% of rodents), with only Eliomys truci and Glis cf. minor. The ecological preferences of Apodemus gorafensis and Paraethomys meini are unknown, but they account for a very small part of the total rodent sample (3.5%).
Overall, the ecological affinities assigned to the identified small vertebrates suggest the presence of a landscape dominated by open herbaceous areas in the surroundings of La Piquera during the earliest Pliocene, with the occasional development of forest patches and stable water bodies under relatively dry and warm environmental conditions.
Conclusions
The diversity and abundance of small vertebrate remains in La Piquera, a new locality from the Iberian Peninsula, is extraordinary. With more than 1200 remains, representing 25 families, La Piquera is in the way of becoming a key locality for the Early Pliocene faunas of southwestern Europe, central Spain, where this time-span is poorly represented. Among the material, there are representatives of urodeles (Salamandridae), anurans (Alytidae, Pelodytidae, and Bufonidae), lizards (Agamidae, Lacertidae, Anguidae, Scincidae, and Blanidae), snakes (?Erycidae and Colubridae/Psammophiidae), artiodactyls (Bovidae), insectivores (Soricidae and Erinaceidae), bats (Rhinolophidae, Vespertilionidae, Megadermatidae, and Miniopteridae), rodents (Cricetidae, Gerbillidae, Muridae, Gliridae, and Sciuridae), and lagomorphs (Leporidae and Prolagidae). The faunal assemblage suggests an earliest Pliocene age (at about 5.3–4.6 Ma), lower part of the MN14 unit (early Ruscinian), with the presence of Debruijnimys sp., Castillomys gracilis, Stephanomys dubari, Apodemus gorafensis, Paraethomys meini, Occitanomys alcalai, Apocricetus cf. barrierei, and Ruscinomys lasallei, among others. The ecological requirements of the small vertebrates point to the dominance of open grassy areas with a scarce presence of forested environments under relatively dry and warm conditions during the earliest Pliocene in the Duero Basin.
The La Piquera collection represents the richest sample of small vertebrates from the beginning of the Pliocene documented to date in the Iberian Peninsula, contributing to an improved knowledge of the local biostratigraphic succession during the Neogene. Some taxonomic groups would deserve further investigation, and some of them probably include new species. Therefore, it is expected that La Piquera will be the subject of several taxonomic works. Aside from providing new information on the local Early Pliocene palaeoenvironment, the detailed study of the vertebrates from this site will shed light on the fauna that lived in central Iberian Peninsula after the Messinian Salinity Crisis.
Author’s contributions
The study is based on fieldwork by CL, PS, and DMM, with the contribution of all co-authors. Study of the facies was carried out by DMM, the systematic study of herpetofauna by HAB, and the study of micromammals by PP, PS, JA, MF, and CL. Biostratigrapic interpretation was developed from discussions by PP, whereas palaeoecological interpretations are based on discussions between PP and HAB. The manuscript was prepared by PP and DMM with contributions from all co-authors.
Acknowledgements
The authors would here like to express their gratitude to Jorge Morales (Museo Nacional de Ciencias Naturales, CSIC, Spain) for kindly provinding description and identification of the artiodactyl specimens. We are especially grateful to the referees Raef Minwer-Barakat Requena (University of Granada, Spain) and Georgios Georgalis (Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Kraków, Poland), and the editor Olivier Lambert (Institut royal des Sciences naturelles de Belgique, Brussels, Belgium) for their very constructive comments and suggestions on the manuscript. IPHES is part of the CERCA Program (Generalitat de Catalunya) and has received financial support from the Spanish Ministry of Science and Innovation through the “María de Maeztu” program for Units of Excellence (CEX2019-000945-M). This research is also supported by projects 2017SGR-859 (Generalitat de Catalunya), PRP-PID2021-123092NB-C21 and PRP-PID2021-122533NB-I00 (Spanish Ministry of Science and Innovation). PP is supported by a “Juan de la Cierva-Incorporación” contract (grant IJC2020-044108-I) funded by MCIN/AEI/10.13039/501100011033 and “European Union NextGenerationEU/PRTR”.
References
Adrover, R. 1986. Nuevas faunas de roedores en el Mio-Plioceno continental de la región de Teruel (España). Interés Bioestratigráfico y Paleoecológico. 423 pp. Instituto de Estudios Turolenses, Teruel.
Adrover, R., Mein, P., and Moissenet, E. 1988. Contribución al conocimiento de la fauna de roedores del Plioceno de la región de Teruel. Revista del Instituto de Estudios Turolenses 79: 89–151.
Adrover, R., Mein, P. and Moissenet, E. 1993a. Los Sciuridae del Turoliense superior, Rusciniense y Villanyiense de la región de Teruel. Paleontologia i Evolució 26–27: 85–106.
Adrover, R., Mein, P., and Moissenet, E. 1993b. Roedores de la transición Mio-Plioceno de la región de Teruel. Paleontologia i Evolució 26–27: 47–84.
Alcalá, L. 1994. Macromamíferos neógenos de la fosa de Alfambra-Teruel. 554 pp. Instituto de Estudios Turolenses, Teruel.
Aguilar, J.P., Brandy, L.D., and Thaler, L. 1984. Les rongeurs de Salobreña (sud de l’Espagne) et le problème de la migration messinienne. Paléobiologie continentale 14: 3–17.
Aguilar, J.P., Calvet, M., and Michaux, J. 1995. Les rongeurs du gisement karstique miocène supérieur de Caltelnou 1 (Pyrénées-Orientales, France). Geobios 28: 501–510. Crossref
Aguilar, J.P., Michaux, J., Bachelet, B., Calvet, M., and Faillat, J.P. 1991. Les nouvelles faunes de rongeurs proches de la limit mio-pliocene en Roussillon. Implications biostratigraphiques et biogéographiques. Palaeovertebrata 20: 147–174.
Agustí, J. 1989. On the peculiar distribution of some muroid taxa in the Western Mediterranean. Bolletino della Società Paleontologia Italiana 28: 147–154.
Agustí, J. 1990. The Miocene Rodent Succession in Eastern Spain: a zoogeographical appraisal. In: E. Lindsay, V. Fahlbusch, and P. Mein (eds.), European Neogene Mammal Chronology, 375–404. Springer Science & Business Media, New York. Crossref
Agustí, J. and Casanovas-Vilar, I. 2003. Neogene gerbils from Europe. Deinsea 10: 13–22.
Agustí, J., Chochishvili, G., Lozano-Fernández, I., Furió, M., Piņero, P., and de Marfà, R. 2022. Small mammals (Insectivora, Rodentia, Lagomorpha) from the Early Pleistocene hominin-bearing site of Dmanisi (Georgia). Journal of Human Evolution 170: 103238. Crossref
Agustí, J., Garcés, M., and Krijgsman, W. 2006. Evidence for African-Iberian exchanges during the Messinian in the Spanish mammalian record. Palaeogeography, Palaeoclimatology, Palaeoecology 238: 5–14. Crossref
Agustí, J., Oms, O., and Parés, J.M. 1999. Calibration of the Early–Middle Pleistocene transition in the continental beds of the Guadix-Baza Basin (SE Spain). Quaternary Science Reviews 18: 1409–1417. Crossref
Agustí, J., Santos-Cubedo, A., Furió, M., De Marfá, R., Blain, H.A., Oms, O., and Sevilla, P. 2011. The late Neogene–early Quaternary small vertebrate succession from the Almenara-Casablanca karst complex (Castellón, Eastern Spain): chronologic and paleoclimatic context. Quaternary International 243: 183–191. Crossref
Angelone, C. 2008. Contribution of complex discrete Fourier transform (CDFT) analysis to the systematics of the genus Prolagus (Ochotonidae, Lagomorpha, Mammalia). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 249: 129–138.
Angelone, C. and Sesé, C. 2009. New characters for species discrimination within the genus Prolagus (Ochotonidae, Lagomorpha, Mammalia). Journal of Paleontology 83: 80–88. Crossref
Angelone, C., Colombero, S., Esu, D., Giuntelli, P., Marcolini, F., Pavia, M., Trenkwalder, S., van den Hoek Ostende, L.W., Zunino, M., and Pavia, G. 2011.Moncucco Torinese, a new post-evaporitic Messinian fossiliferous site from Piedmont (NW Italy). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 259: 89–104. Crossref
Augé, M.L. 2005. Evolution des lézards du Paléogène en Europe. Mémoires du Muséum national d’Histoire naturelle 192: 1–369.
Augé, M.L. and Brizuela, S. 2020. Transient presence of a teiid lizard in the European Eocene suggests transatlantic dispersal and rapid extinction. Palaeobiodiversity and Palaeoenvironments 100: 793–817. Crossref
Augé, M.L. and Pouit, D. 2012. Presence of iguanid lizards in the European Oligocene Lazarus taxa and fossil abundance. Bulletin de la Société géologique de France 183: 653–660. Crossref
Averianov, A. and Tesakov, A.S. 1997. Evolutionary trends in Mio-Pliocene Leporinae, based on Trischizolagus (Mammalia, Lagomorpha). Paläontologische Zeitschrift 71: 145–153. Crossref
Bailon, S. 1991. Amphibiens et Reptiles du Pliocène et du Quaternaire de France et d’Espagne: Mise en Place et Evolution des Faunes. 499 pp. Unpublished Ph.D. Thesis, Université de Paris VII, Paris.
Bailon, S. 1999. Différenciation ostéologique des anoures (Amphibia, Anura) de France. In: J. Desse and N. Desse-Berset (eds.), Fiches d’ostéologie animale pour l’archéologie, Serie C: varia, 3–41. Centre de Recherches archéologiques-CNRS, Valbonne.
Barahona, F. and Barbadillo, L.J. 1997. Identification of some Iberian lacertids using skull characters. Revista Española de Herpetología 11: 47–62.
Blain, H.A., Bailon, S., Cuenca-Bescos, G., Arsuaga, J.L., Bermúdez de Castro, J.M., and Carbonell, E. 2009. Long-term climate record inferred from early–middle Pleistocene amphibian and squamate reptile assemblages at the Gran Dolina Cave, Atapuerca, Spain. Journal of Human Evolution 56: 55–65. Crossref
Blain, H.A., Lozano-Fernández, I., Agustí, J., Bailon, S., Menéndez Granda, L, Espigares Ortiz, M.P., Ros-Montoya, S., Jiménez Arenas, J.M., Toro-Moyano, I., Martínez-Navarro, B., and Sala, R. 2016. Refining upon the climatic background of the early Pleistocene hominid settlement in Western Europe: Barranco León and Fuente Nueva-3 (Guadix-Baza Basin, se Spain). Quaternary Science Reviews 144: 132–144. Crossref
Blott, S.J. and Pye, K. 2012. Particle size scales and classification of sediment types based on particle size distributions: Review and recommended procedures. Sedimentology 59: 2071–2096. Crossref
Böhme, M., Ilg, A., Ossig, A., and Kuchenhoff, H. 2006. New method to estimate paleoprecipitation using fossil amphibians and reptiles and the middle and late Miocene precipitation gradients in Europe. Geology 34: 425–428. Crossref
Bolet, A. 2017. First early Eocene lizards from Spain and a study of the compositional changes between late Mesozoic and early Cenozoic Iberian lizard assemblages. Palaeontologia Electronica 20.2.20A: 1–22. Crossref
Booth-Rea, G., R., Ranero, C. and Grevemeyer, I. 2018. The Alboran volcanic-arc modulated the Messinian faunal exchange and salinity crisis. Scientific Reports 8: 13015. Crossref
Calvo Sorando, J.P., Daams, R., Morales, J., López Martínez, N., Agustí, J., Anadón, P., Armenteros, I., Cabrera, L., Civis Llovera, J., Corrochano, A., Díaz Molina, M., Elizaga, E., Hoyos, M., Martín-Suárez, E., Martínez, J., Moissenet, E., Muñoz, A., Pérez-García, A., Pérez-González, A., Portero, J.M., Robles, F., Santisteban, C., Torres, T., van der Meulen, A.J., Vera, J.A., and Mein, P. 1993. Up-to-date Spanish continental Neogene synthesis and paleoclimatic interpretation. Revista de la Sociedad Geologica de España 6: 29–40.
Carrascal, L.M. and Salvador, A. (eds.) 2002–2016. Enciclopedia Virtual de los Vertebrados Españoles. Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/.
Castillo, C. 1990. Paleocomunidades de micromamíferos de los yacimientos kársticos del Neógeno Superior de Andalucía Oriental. 255 pp. Unpublished Ph.D. Thesis, Universidad de Granada, Granada.
Castillo Ruiz, C. 1992. Paraethomys belmezensis nov. sp. (Rodentia, Mammalia) du pliocène de Córdoba (Espagne). Geobios 25: 775–780. Crossref
Castillo, C. and Agustí, J. 1996. Early Pliocene rodents (Mammalia) from Asta Regia (Jerez Basin, Southwestern Spain). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen 99: 25–43.
Čermák, S. and Wagner, J. 2013. The Pliocene record of Trischizolagus and Pliopentalagus (Leporidae, Lagomorpha, Mammalia) in Central Europe with comments on taxonomy and evolutionary history of Leporinae. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 268: 97–111. Crossref
Čermák, S., Angelone, C., and Sinitsa, M.V. 2015. New Late Miocene Alilepus (Lagomorpha, Mammalia) from Eastern Europe—a new light on the evolution of the earliest Old World Leporinae. Bulletin of Geosciences 90: 431–451. Crossref
Čermák, S., Joniak, P., and Rojay, F. 2019. A new early Pliocene locality Tepe Alagoz (Turkey) reveals a distinctive tooth phenotype of Trischizolagus (Lagomorpha, Leporidae) in Asia Minor. Palaeontologia Electronica 22.1.14A: 1–21. Crossref
Chaline, J. 1972. Les rongeurs du Pléistocène moyen et supérieur de France. Cahiers de Paléontologie. Ed du CNRS: 1–410.
Coiffait, B. 1991. Contribution des rongeurs du Néogène d’Afrique à la biostratigraphie d’Afrique du Nord occidentale. 389 pp. Unpublished Ph.D. Thesis, Université Nancy-1, France.
Coiffait, B. and Coiffait, P.E. 1981. Découverte d’un gisement de micromammifères d’âge Pliocène dans le Bassin de Constantine (Algérie): présence d’un Muridé nouveau, Paraethomys athmeniae n. sp. Palaeovertebrata 11: 1–15.
Cuenca-Bescós, G. 1988. Revisión de los Sciuridae del Aragoniense y del Rambliense en la fosa de Calatayud-Montalbán. Scripta Geologica 87: 1–116.
Daams, R. and Freudenthal, M. 1988. Cricetidae (Rodentia) from the type-Aragonian; the genus Megacricetodon. Scripta Geologica Special Issue 1: 39–132.
Daxner-Höck, G. and Höck, E. 2009. New data on Eomyidae and Gliridae (Rodentia, Mammalia) from the late Miocene of Austria. Annalen des Naturhistorischen Museums in Wien. Serie A 111: 375–444.
De Bruijn, H. 1973. Analysis of the data bearing upon the correlation of the Messinian with the Succession of Land Mammals. In: C.W. Drooger (ed.), Messinian Events in the Mediterranean, 260–262. Koninklijke Nederlandse Akademie van Wetenschappen, Amsterdam.
De Bruijn, H. 1999. Superfamily Sciuroidea. 1999. In: G.E. Rössner and K. Heissig (eds.), The Miocene Land Mammals of Europe, 271–280. Verlag Dr. Friedrich Pfeil, Munich.
De Bruijn, H. and Mein, P. 1968. On the mammalian fauna of the Hipparion-beds in the Calatayud-Teruel Basin (Prov. Zaragoza, Spain). Part V. The Sciurinae. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen Series B 71: 73–90.
De Bruijn, H. and Van der Meulen, A. 1975. The Early Pleistocene rodents from Tourkobounia-1 (Athens, Geece). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen B 78: 314–338.
Delfino, M., Kotsakis, T., Arca, M., Tuveri, C., Pitruzzella, G., and Rook, L. 2008. Agamid lizards from the Plio-Pleistocene of Sardinia (Italy) and an overview of the European fossil record of the family. Geodiversitas 30: 641–656.
Deperet, C. 1890. Les animaux pliocenes du Roussillon. Memoires de la Societe geologique de France, Paleontologie 3: 1–194.
Fejfar, O. and Storch, G. 1990. Eine Pliozäne (ober-Ruscinische) Kleinsäigerfauna aus Gunsdersheim, Rhein-hessen. 1. Nagetiere: Mammalia, Rodentia. Senkenbergiana Lethaea 71: 139–184.
Freudenthal, M. 2004. Gliridae (Rodentia, Mammalia) from the Eocene and Oligocene of the Sierra Palomera (Teruel, Spain). Treballs del Museu de Geologia de Barcelona 12: 97–173.
Freudenthal, M., Mein, P., and Martín-Suárez, E. 1998. Revision of Late Miocene and Pliocene Cricetinae (Rodentia, Mammalia) from Spain and France. Treballs del Museu de Geologia de Barcelona 7: 11–93.
Furió, M. and Agustí, J. 2017. Latest Miocene insectivores from Eastern Spain: Evidence for enhanced latitudinal differences during the Messinian. Geobios 50: 123–140. Crossref
Furió, M., Casanovas-Vilar, I., and Van den Hoek Ostende, L.W. 2011. Predictable structure of Miocene insectivore (Lipotyphla) faunas in Western Europe along a latitudinal gradient. Palaeogeography, Palaeoclimatology, Palaeoecology 304: 219–229. Crossref
Furió, M., Santos-Cubedo, A., Minwer-Barakat, R., Agustí, J. 2007. Evolutionary history of the African Soricid Myosorex (Insectivora, Mammalia) out of Africa. Journal of Vertebrate Paleontology 27: 1018–1032. Crossref
Furió, M., Van den Hoek Ostende L.W., Agustí J., and Minwer-Barakat, R. 2018. Evolution of the insectivore assemblages (Eulipotyphla, Mammalia) in Spain and their relation with Neogene and Quaternary climatic changes. Ecosistemas 27: 38–51. Crossref
Garcés, M., Krijgsman, W., and Agustı́, J. 1998. Chronology of the late Turolian deposits of the Fortuna basin (SE Spain): implications for the Messinian evolution of the eastern Betics. Earth and Planetary Science Letters 163: 69–81. Crossref
Garcés, M., Krijgsman, W., and Agustí, J. 2001. Chronostratigraphic framework and evolution of the Fortuna basin (Eastern Betics) since the Late Miocene. Basin Research 13: 199–217. Crossref
García-Alix, A. 2006. Bioestratigrafía de los depósitos continentales de la transición Mio-Plioceno de la cuenca de Granada. 414 pp. Unpublished Ph.D. Thesis, Universidad de Granada, Granada.
García-Alix, A., Minwer-Barakat, R., Martín-Suárez, E., and Freudenthal, M. 2007. New data on Mio-Pliocene Sciuridae (Rodentia, Mammalia) from southern Spain. Comptes Rendus Palevol 6: 269–279. Crossref
García-Alix, A., Minwer-Barakat, R., Martín-Suárez, E., and Freudenthal, M. 2008a. Biostratigraphy and sedimentary evolution of Late Miocene and Pliocene continental deposits of the Granada Basin (southern Spain). Lethaia 41: 431–446. Crossref
García-Alix, A., Minwer-Barakat, R., Martın-Suárez, E., and Freudenthal M. 2008b. Cricetidae and Gliridae (Rodentia, Mammalia) from the Miocene and Pliocene of southern Spain. Scripta Geologica 136: 1–37.
García-Alix, A., Minwer-Barakat, R., Martín-Suarez, E., and Freudenthal, M. 2008c. Muridae (Rodentia, Mammalia) from the Mio-Pliocene boundary in the Granada Basin (southern Spain). Biostratigraphic and phylogenetic implications. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 248: 183–215. Crossref
García-Alix, A., Minwer-Barakat, R., Martín-Suárez, E., Freudenthal, M., Aguirre, J., and Kaya, F. 2016. Updating the Europe-Africa small mammal exchange during the late Messinian. Journal of Biogeography 43: 1336–1348. Crossref
García-Alix, A., Minwer-Barakat, R., Martín-Suárez, E., Freudenthal, M., and Martín, J.M. 2008d. Late Miocene–Early Pliocene climatic evolution of the Granada Basin (southern Spain) deduced from the paleoecology of the micromammal associations. Palaeogeography, Palaeoclimatology, Palaeoecology 265: 214–225. Crossref
García-Cortés, A., Mansilla, H., Quintero, I., Torres, T., Rubio Pascual, F., and Gabaldón, V. 2008. Mapa geológico y Memoria de la Hoja nº403 (Maderuelo). Mapa Geológico de España E. 1:50.000 (MAGNA). Segunda Serie, Primera Edición. IGME, Madrid.
García-París, M., Montori, A., and Herrero, P. 2004. Amphibia. Lissamphibia. In: M.A. Ramos et al. (eds.), Fauna Ibérica, Vol. 24, 1–640. Museo Nacional de Ciencias Naturales, Madrid.
Georgalis, G.L. 2019. Poor but classic: The squamate fauna from the late Miocene of Pikermi, near Athens, Greece. Comptes Rendus Palevol 18: 801–815. Crossref
Georgalis, G.L. and Smith, K.T. 2020. Constrictores Oppel, 1811—the available name for the taxonomic group uniting boas and pythons. Vertebrate Zoology 70: 291–304.
Georgalis, G.L. and Szyndlar, Z. 2022. First occurrence of Psammophis (Serpentes) from Europe witnesses another Messinian herpetofaunal dispersal from Africa—biogeographic implications and a discussion of the vertebral morphology of psammophiid snakes. The Anatomical Record 305: 3263–3282. Crossref
Georgalis, G.L., Villa, A., and Delfino, M. 2018. The last amphisbaenian (Squamata) from continental Eastern Europe. Annales de Paléontologie 104: 155–159. Crossref
Georgalis, G.L., Villa, A., Ivanov, M., Vasilyan, D., and Delfino, M. 2019. Fossil amphibians and reptiles from the Neogene locality of Maramena (Greece), the most diverse European herpetofauna at the Miocene/Pliocene transition boundary. Palaeontologia Electronica 22.3.68: 1–99. Crossref
Geraads, D. 1994. Rongeurs et Lagomorphes du Pléistocène moyen de la “Grotte des Rhinocéros”, carrière Oulad Hamida 1, à Casablanca, Maroc. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 191: 147–172.
Gibert, L., Scott, G.R., Montoya, P., Ruiz-Sánchez, F.J., Morales, J., Luque, L., Abella, J., and Lería, M. 2013. Evidence for an African-Iberian mammal dispersal during the pre-evaporitic Messinian. Geology 41: 691–694. Crossref
Gómez de Soler, B., Campeny Vall-llosera, G., van der Made, J., Oms, O., Agustí, J., Sala, R., Blain. H.A., Burjachs, F., Claude, J., García Catalan, S., Riba, D., and Rosillo, R. 2012. A new key locality for the Pliocene vertebrate record of Europe: the Camp dels Ninots maar (NE Spain). Geologica Acta 10: 1–17.
Haller-Probst, M. and Schleich, H.H. 1994. Vergleichende osteologische Untersuchungen an einigen Urodelen Eurasiens (Amphibia: Urodela, Salamandridae, Proteidae). Courier Forschungsinstitut Senckenberg 173: 23–77.
Hedges, S.B. 2014. The high-level classification of skinks (Reptilia, Squamata, Scincomorpha). Zootaxa 3765: 317–338. Crossref
Hellmund, M. and Ziegler, R. 2012. A Ruscinian mammalian microfauna from a fissure filling near Sondershausen (Thuringia, Central Germany). Swiss Journal of Palaeontology 131: 77–94. Crossref
Hoffstetter, R. 1962. Observations sur les ostéodermes et la classification des Anguidés actuels et fossiles (Reptiles, Sauriens). Bulletin du Muséum national d’Histoire naturelle 34: 149–157.
Hoffstetter, R. and Rage, J.-C. 1972. Les Erycinae fossiles de France (Serpentes, Boidae). Compréhension et histoire de la sous-famille. Annales de Paléontologie (Vertébrés) 63: 161–190.
Hordijk, K. and De Bruijn, H. 2009. The succession of rodent faunas from the Mio-Pliocene lacustrine deposits of the Florina-Ptolemais-Servia Basin (Greece). Hellenic Journal of Geosiences 44: 21–103.
Jaeger, J.J., Michaux, J., and Thaler, L. 1975. Présence d’un rongeur muridé nouveau, Paraethomys miocaenicus n. sp., dans le Turolien supérieur du Maroc et d’Espagne. Implications paléogéographiques. Comptes Rendus de l’Académie des Sciences Paris 280: 1673–1676.
Janvier, P. and Montenat, C., 1971. Le plus ancien Léporité d’Europe Occidentale, Hispanolagus crusafonti n. gen. n. sp. du Miocène supérieur de Murcia (Espagne). Bulletin du Muséum national d’Histoire naturelle 42: 780–788.
Klembara, J. and Rummel, M. 2018. New material of Ophisaurus, Anguis and Pseudopus (Squamata, Anguidae, Anguinae) from the Miocene of the Czech Republic and Germany and systematic revision and palaeobiogeography of the Cenozoic Anguinae. Geological Magazine 155: 20–44. Crossref
Kowalski, K. 1956. Insectivores, bats and rodents from the Early Pleistocene bone breccia of Podlesice near Kroczyce (Poland). Acta Palaeontologica Polonica 1: 331–394.
Kowalski, K. 1963. The Pliocene and Pleistocene Gliridae (Mammalia, Rodentia) from Poland. Acta Zoologica Cracoviensa 14: 533–567.
Krijgsman, W., Hilgen, F.J., Raffi, I., Sierro, F.J., and Wilson, D.S. 1999. Chronology, causes and progression of the Messinian salinity crisis. Nature 400: 652–655. Crossref
Krystufek, B., Mahmoudi, A., Tesakov, A.S., Mateju, J., and Hutterer R. 2016. A review of bristly ground squirrels Xerini and a generic revision in the African genus Xerus. Mammalia 80: 521–540. Crossref
Laplana, C., Sevilla, P., Arsuaga, J.L., Arriaza, M.C., Baquedano, E., Pérez-González, A., and López-Martínez, N. 2015. How far into Europe did pikas (Lagomorpha: Ochotonidae) go during the Pleistocene? New evidence from Central Iberia. Plos One 10 (11): e0140513. Crossref
López, N., Agustí, J., Cabrera, L., Calvo Sorando, J.P., Civis, J., Corrochano, A., Daams, R., Díaz, M., Elizaga, E., Hoyos, M., Martínez, J., Morales, J., Portero, J.M., Robles, F., Santisteban, C., Torres, T., Alberdi, M.T., Álvarez Sierra, M., Belinchón, M., Carballeira, J., Cuenca, G., Freudenthal, M., García, E., Gibert, J., González, A., Junco, F., Lacomba, J.I., Mazo, A.V., van der Meulen, A., Moyà, S., Olivé, A., Ordóñez, S., Renzi, M., Sacristán, M.A., Sesé, C., Soria, D., Usera, J., and Zapata, J.J. 1987. Approach to the Spanish continental Neogene synthesis and paleoclimatic interpretation. Annals of the Hungarian Geological Institute 70: 383–391.
López-García, J.M., Blain, H.A., Cuenca Bescós, G., and Arsuaga, J.L. 2008. Chronological, environmental, and climatic precisions on the Neanderthal site of the Cova del Gegant (Sitges, Barcelona, Spain). Journal of Human Evolution 55: 1151–1155. Crossref
López-Martínez, N. 1989. Revisión sistemática de los Lagomorpha (Mammalia) del Terciario y Cuaternario de España. Memorias del Museo Paleontologico de la Universidad de Zaragoza 3: 1–344.
López-Martínez, N. 2001. Paleobiogeographical history of Prolagus, an European ochotonid (Lagomorpha). Lynx 32: 215–231.
López-Martínez, N. 2008. The lagomorph fossil record and the origin of the European rabbit. In: P.C. Alves, N. Ferrand, and K. Hackländer (eds.), Lagomorph Biology: Evolution, Ecology and Conservation, 27–46. Springer, Berlin. Crossref
López-Martínez, N., Likius, A., Mackaye, H.T., Vignaud, P., and Brunet, M. 2007. A new lagomorph from the Late Miocene of Chad (Central Africa). Spanish Journal of Palaeontology 22: 1–20. Crossref
Luengo, J., Garcés, M., Montes, M., Beamud, E., Nozal, F., López-Olmedo, F., and Armenteros, I. 2010. Magnetoestratigrafía del Neógeno de la Cuenca del Duero. MAGIVER 6: 22–25.
Luengo, J., Nozal, F., Peláez Campomanes, P., Montes, M., Armenteros, I., López Olmedo, F., and García Paredes, I. 2009. Leading factors of fluvial/lacustrine Castillejo de Robledo intrabasinal infill (southeastern border of the Duero Basin, Spain). In: A. Amorosi (ed.), Proceeding of the 27th IAS Meeting of Sedimentology, 582. Medimond, Bologna.
Maldonado-Garrido, E., Piñero, P., and Agustí, J. 2017. A catalogue of the vertebrate fossil record from the Guadix-Baza Basin (SE Spain). Spanish Journal of Palaeontology 32: 207–235. Crossref
Mansino, S., Crespo, V., Montoya, P., and Ruiz-Sánchez, F.J. 2017a. Muridae from the late Miocene site of Venta del Moro (Eastern Spain). Historical Biology 29: 677–691. Crossref
Mansino, S., Fierro, I., Montoya, P., and Ruiz-Sánchez, F.J. 2015a. Micromammal faunas from the Mio-Pliocene boundary in the Alcoy Basin (SE Spain): biostratigraphical and palaeoecological inferences. Bulletin of Geosciences 90: 555–576. Crossref
Mansino, S., Fierro, I., Tosal, A., Montoya, P., and Ruiz-Sánchez, F.J. 2017b. Micromammal biostratigraphy of the Alcoi Basin (Eastern Spain): remarks on the Pliocene record of the Iberian Peninsula. Geologica Acta 15: 121–134.
Mansino, S., Ruiz-Sánchez, F.J., de Luque, L., Montoya, P., Gibert, L., Morales, J., Abellá, J., Crespo, V. D., and Scott, J.R. 2015b. First Early Pliocene micromammal faunas from the Venta del Moro area (Cabriel Basin, Spain): new data on the Messinian dispersal of Debruijnimys. Journal of Iberian Geology 41: 273–293. Crossref
Mansino, S., Ruiz-Sanchez, F.J., and Montoya, P. 2016a. Sciuridae (Rodentia, Mammalia) from the late Miocene of Venta del Moro (Cabriel Basin, Spain): biostratigraphical, phylogenetic and palaeoecological inferences. Historical Biology 30: 297–304. Crossref
Mansino, S., Ruiz-Sánchez F.J., Fierro, I., and Montoya, P. 2016b. Mio-Pliocene rodent assemblages from Alcoi Forn (Alcoy Basin, Eastern Spain). Biostratigraphical and palaeoclimatical inferences. Historical Biology 28: 1050–1065. Crossref
Mansino, S., Ruiz-Sánchez, F.J., Freudenthal, M., and Montoya, P. 2014. A new approach to the Late Miocene–Early Pliocene forms of the genus Apocricetus. Apocricetus alberti (Rodentia, Mammalia) from Venta del Moro (Cabriel Basin, Spain). Proceedings of the Geologists’ Association 125: 392–405. Crossref
Martín-Suárez, E. 1988. Sucesiones de micromamíferos en la Depresión de Guadix-Baza (Granada, España). 241 pp. Unpublished Ph.D. Thesis, Universidad de Granada, Granada.
Martín-Suárez, E. and Freudenthal, M. 1993. Muridae (Rodentia) from the lower Turolian of Crevillente (Alicante, Spain). Scripta Geologica 103: 65–118.
Martín-Suárez, E. and Mein, P. 1991. Revision of the genus Castillomys (Muridae, Rodentia). Scripta Geologica 96: 47–81.
Martín-Suárez, E. and Mein, P. 1998. Revision of the genera Parapodemus, Apodemus, Rhagamys and Rhagapodemus (Rodentia, Mammalia). Geobios 31: 87–97. Crossref
Martín-Suárez, E., Freudenthal, M., and Civis, J. 2001. Rodent palaeoecology of the Continental Upper Miocene of Crevillente (Alicante, SE Spain). Palaeogeography, Palaeoclimatology, Palaeoecology 165: 349–356. Crossref
Martín-Suárez, E., Oms, O., Freudenthal, M., Agustí, J., and Parés, J.M. 1998. Continental Mio-Pliocene transition in the Granada Basin. Lethaia 31: 161–166. Crossref
Mazo, A.V. 1996. Nuevos datos sobre el yacimiento de vertebrados pliocenos de Las Higueruelas (Ciudad Real, España). Bollettino del Museo Regionale di Scienze Naturali di Torino 14: 1–8.
Mazo, A.V., Sesé, C., Ruiz Bustos, A., and Peña, J.A. 1985. Geología y paleontología de los yacimientos Plio-Pleistocenos de Huéscar (Depresión de Guadix Baza, Granada). Estudios Geológicos 41: 467–493. Crossref
Mein, P. and Freudenthal, M. 1971. Une nouvelle classification des Cricetidae (Mammalia, Rodentia) du Tertiaire de l’Europe. Scripta Geologica 2: 1–37.
Mein, P. and Martín-Suárez, E. 1993. Galerix iberica sp. nov. (Erinaceidae, Insectivora, Mammalia) from the Late Miocene and Early Pliocene of the Iberian Peninsula. Geobios 26: 723–730. Crossref
Mein, P. and Michaux, J. 1970. Un nouveau stade dans l’évolution des rongeurs pliocènes de l’Europe sud-occidentale. Comptes Rendus des Séances de l’Académie des Sciences de Paris D 270: 2780–2783.
Mein, P., Moissenet, E., and Adrover, R. 1990. Biostratigraphie du Néogène Supérieur du bassin de Teruel. Paleontologia i Evolució 23: 121–139.
Menu, H. and Popelard, J.B. 1987. Utilisation des caractères dentaires pour la déterminatin des Vespertilioninés de l’ouest Européen. Le Rhinolophe 4: 1–88.
Michaux, J. 1969. Muridae (Rodentia) du Pliocène supérieur d’Espagne et du midi de la France. Paleovertebrata 3: 1–25. Crossref
Minwer-Barakat, R., Agustí, J., García-Alix, A., and Martín-Suárez, E. 2018. The European record of the gerbil Myocricetodon (Rodentia, Mammalia) and its bearing on the Messinian salinity crisis. Palaeogeography, Palaeoclimatology, Palaeoecology 506: 168–182. Crossref
Minwer-Barakat, R., García-Alix, A., Agustí, J., Martín-Suárez, E., and Freudenthal, M. 2009a. The micromammal fauna from Negratín-1 (Guadix Basin, southern Spain): new evidence of African-Iberian mammal exchanges during the Late Miocene. Journal of Paleontology 83: 854–879. Crossref
Minwer-Barakat, R., García-Alix, A., Martín-Suárez, E., and Freudenthal, M. 2005. Muridae (Rodentia) from the Pliocene of Tollo de Chiclana (Granada, South-Eastern Spain). Journal of Vertebrate Paleontology 25: 426–441. Crossref
Minwer-Barakat, R., García-Alix, A., Martín-Suárez, E., and Freudenthal, M. 2009b. Late Turolian micromammals from Rambla de Chimeneas-3: considerations on the oldest continental faunas from the Guadix Basin (southern Spain). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 251: 95–108. Crossref
Minwer-Barakat, R., García-Alix, A., Martín Suárez, E., and Freudenthal, M. 2010. Soricidae (Soricomorpha, Mammalia) from the Pliocene of Tollo de Chiclana (Guadix Basin, Southern Spain). Journal of Vertebrate Paleontology 30: 535–546. Crossref
Minwer-Barakat, R., García-Alix, A., Martín-Suárez, E., and Freudenthal, M. 2020. Early Pliocene Desmaninae (Mammalia, Talpidae) from southern Spain and the origin of the genus Desmana. Journal of Vertebrate Paleontology 40: e1835936. Crossref
Minwer-Barakat, R., García-Alix, A., Martín-Suárez, E., Freudenthal, M., and Viseras, C. 2012. Micromammal biostratigraphy of the Upper Miocene to lowest Pleistocene continental deposits of the Guadix Basin, southern Spain. Lethaia 45: 594–614. Crossref
Montenat, C. and De Bruijn, H. 1976. The Ruscinian rodent faunule from La Juliana (Murcia); its implication for correlation of continental and marine biozones. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen B 79: 245–255.
Montuire, S., Maridet, O., and Legendre, S. 2006. Late Miocene–Early Pliocene temperature estimates in Europe using rodents. Palaeogeography, Palaeoclimatology, Palaeoecology 238: 247–262. Crossref
Morales, J., Peláez-Campomanes, P., Abella, J., Montoya, P., Ruiz, F.J., Gibert, L., Scott, G., Cantalapiedra, J.L., and Sanisidro, O. 2013. The Ventian mammal age (Latest Miocene): present state. Spanish Journal of Palaeontology 28: 149–159. Crossref
Opdyke, N., Mein, P., Lindsay, E., Perez-Gonzales, A., Moissenet, E., and Norton, V.L. 1997. Continental deposits, magnetostratigraphy and vertebrate paleontology, late Neogene of Eastern Spain. Palaeogeography, Palaeoclimatology, Palaeoecology 133: 129–148. Crossref
Peláez-Campomanes, P. 2001. Revision of the Aragonian (Miocene) Atlantoxerus (Sciuridae). Journal of Paleontology 75: 418–426. Crossref
Piñero, P. and Agustí, J. 2019. The rodent succession in the Sifón de Librilla section (Fortuna Basin, SE Spain): implications for the Mio-Pliocene boundary in the Mediterranean terrestrial record. Historical Biology 31: 279–321. Crossref
Piñero, P. and Agustí, J. 2020. Rodents from Botardo-D and the Miocene–Pliocene transition in the Guadix-Baza Basin (Granada, Spain). Palaeobiodiversity and Palaeoenvironments 100: 903–920. Crossref
Piñero, P. and Verzi, D.H. 2020. A new early Pliocene murine rodent from the Iberian Peninsula and its biostratigraphic implications. Acta Palaeontologica Polonica 65: 719–731. Crossref
Piñero, P., Agustí, J., and Oms, O. 2018a. The late Neogene rodent succession of the Guadix-Baza Basin (south-eastern Spain) and its magnetostratigraphic correlation. Palaeontology 61: 253–272. Crossref
Piñero, P., Agustí, J., Blain, H.A., and Laplana, C. 2016. Paleoenvironmental reconstruction of the Early Pleistocene site of Quibas (SE Spain) using a rodent assemblage. Comptes Rendus Palevol 15: 659–668. Crossref
Piñero, P., Agustí, J., Furió, M., and Laplana, C. 2018b. Rodents and insectivores from the late Miocene of Romerales (Fortuna Basin, Southern Spain). Historical Biology 30: 336–359. Crossref
Piñero, P., Agustí, J., Oms, O., Blain, H.A., Laplana, C., Ros-Montoya, S., and Martínez-Navarro, B. 2017a. Rodents from Baza-1 (Guadix-Baza Basin, southeast Spain): filling the gap of the early Pliocene succession in the Betics. Journal of Vertebrate Paleontology 37: e1338294. Crossref
Piñero, P., Agustí, J., Oms, O., Fierro, I., Montoya, P., Mansino, S., Ruiz-Sánchez, F.R., Alba, D.M., Alberdi, M.T., Blain, H.A., Laplana, C., Van der Made, J., Mazo, A.V., Morales, J., Murelaga, X., Pérez-García, A., Pérez-Valera, F., Pérez-Valera, J.A., Sevilla, P., Soria, J.M., and Romero, G. 2017b. Early Pliocene continental vertebrate fauna at Puerto de la Cadena (SE Spain) and its bearing on the marine-continental correlation of the late Neogene of Eastern Betics. Palaeogeography, Palaeoclimatology, Palaeoecology 479: 102–114. Crossref
Pleguezuelos, J.M. and Martínez-Rica, J.P. 1997. Distribución y Biogeografia de los anfibios y reptiles de España. In: J.M. Pleguezuelos and J.P. Martínez-Rica (eds.), Monografías de la Revista Española de Herpetología 3, Distribución y Biogeografía de los anfibios y reptiles de España y Portugal, 1–542. Universidad de Granada, Granada.
Pleguezuelos, J.M., Márquez, R., and Lizana, M. (eds.). 2004. Atlas y Libro Rojo de los Anfibios y Reptiles de España. 587 pp. Dirección General de Conservación de la Naturaleza-Asociación Herpetologica Española, Madrid.
Qiu, Z.D. and Storch, G. 2000. The early Pliocene micromammalian fauna of Bilike, Inner Mongolia, China (Mammalia: Lipotyphla, Chiroptera, Rodentia, Lagomorpha). Senckenbergiana lethaea 80: 173–229. Crossref
Radulesco, C. and Samson, P. 1967. Contribution à la connaissance du complexe faunistique de Măluşteni-Bereşti (Pléistocène inférieur), Roumanie I. Ord. Lagomorpha, Fam. Leporidae. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 9: 544–563.
Renaud, S., Benammi, M., and Jaeger, J.J. 1999. Morphological evolution of the murine rodent Paraethomys in response to climatic variations (Mio-Pleistocene of North Africa). Paleobiology 25: 369–382. Crossref
Reumer, J.W.F. 1984. Ruscinian and early Pleistocene Soricidae (Insectivora, Mammalia) from Tegelen (The Netherlands) and Hungary. Scripta Geologica: 73: 1–173.
Roček, Z. 1994. Taxonomy and distribution of Tertiary discoglossids (Anura) of the genus Latonia v. Meyer, 1843. Geobios 27: 717–751. Crossref
Rofes, J. and Cuenca-Bescós, G. 2006. First evidence of the Soricidae (Mammalia) Asoriculus gibberodon (Petényi, 1864) in the Pleistocene of North Iberia. Rivista Italiana di Paleontologia e Stratigrafia 112: 301–315.
Ruiz Bustos, A., Sesé, C., Dabrio, C.J., Peña Ruano, J.A., and Padial, J.M. 1984. Geología y fauna de micromamíferos del nuevo yacimiento del Plioceno inferior de Gorafe-A (Depresión de Guadix-Baza, Granada). Estudios Geológicos 40: 231–241. Crossref
Rümke, C.G. 1985. A review of fossil and recent Desmaninae (Talpidae, Insectivora). Utrecht Micropaleontological Bulletins, Special Publication 4: 1–241.
Rzebik-Kowalska, B. 1998. Fossil history of shrews in Europe. In: J.M. Wojik and M. Wolsan (eds.), Evolution of Shrews, 23–92. Mammal Research Institute, Polish Academy of Science, Białowieża.
Rzebik-Kowalska, B. and Nesin, V.A. 2010. Erinaceomorpha and Soricomorpha (Insectivora, Mammalia) from the Late Miocene of Ukraine. 61 pp. Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Kraków.
Sánchez-Bandera, C., Oms, O., Blain, H.A., Lozano-Fernández, I., Bisbal-Chinesta, J.F., Agustí, J., Saarinen, J., Fortelius, M., Titton, S., Serrano-Ramos, A., Luzón, C., Solano-Garcia, J., Barsky, D., and Jiménez-Arenas, J.M. 2020. New stratigraphically constrained palaeoenvironmental reconstructions for the first human settlement in Western Europe: The Early Pleistocene herpetofaunal assemblages from Barranco León and Fuente Nueva 3 (Granada, SE Spain). Quaternary Science Reviews 243: 106466. Crossref
Sanchiz, B. 1998. Salientia. Handbuch der Paläoherpetologie, Part 4. 275 pp. Verlag Dr Friedrich Pfeil, Stuttgart.
Salvador, A. 1997. Reptiles. In: M.A. Ramos et al. (eds.), Fauna Ibérica, Vol. 10, 1–705. Museo Nacional de Ciencias Naturales, Madrid.
Sesé, C. 1989. Micromamíferos del Mioceno, Plioceno y Pleistoceno de la cuenca de Guadix-Baza (Granada). Trabajos sobre el Neógeno-Cuaternario 11: 185–214.
Sesé, C. 2006. Los roedores y lagomorfos del Neógeno de España. Estudios geológicos 62: 429–480. Crossref
Sevilla, P. 1988. Estudio paleontológico de los quirópteros del Cuaternario español. Paleontologia i Evolució 22: 113–233.
Sevilla, P. 1991. Murciélagos fósiles de España. In: J. Benzal and O. de Paz (eds.), Los Murciélagos de España y Portugal. Monografías del ICONA, 21–36. Colección Técnica, Madrid.
Sevilla, P. and López-Martínez, N. 1986. Comparative systematic value between Dental and External/Skeletal features in Western European Chiroptera. Mémoires du Muséum national d’Histoire naturelle de Paris, série C 53: 255–266.
Sigé, B. and Legendre, S. 1983. L’Histoire des peuplements de Chiroptères du bassin méditerranéenne: L’apport comparé des remplissages karstiques et des dépôts fluvio-lacustres. Mémoires de biospéologie 10: 209–225.
Sinitsa, M.V. and Pogodina, N.V. 2019. The evolution of early Spermophilus in eastern Europe and the antiquity of the Old World ground squirrels. Acta Palaeontologica Polonica 64: 643–667. Crossref
Smith, K.T. 2009. Eocene Lizards of the Clade Geiseltaliellus from Messel and Geiseltal, Germany, and the Early Radiation of Iguanidae (Reptilia: Squamata). Bulletin of the Peabody Museum of Natural History 50: 219–306. Crossref
Speybroeck, J., Beukema, W., Dufresnes, C., Fritz, U., Jablonski, D., Lymberakis, P., Martínez-Solano, I., Razzetti, E., Vamberger, M., Vences, M., Vörös, J., and Crochet, P.-A. 2020. Species list of the European herpetofauna e 2020 update by the taxonomic committee of the societas europaea herpetologica. Amphibia-Reptilia 41: 139–189. Crossref
Storch, G. 1999. Order Chiroptera. In: G.E. Rossner and K. Heissig (eds.), The Miocene Land Mammals of Europe, 81–90. Friedich Pfeil, München.
Szyndlar, Z. 1984. Fossil snakes from Poland. Acta Zoologica Cracoviensia 28: 1–156.
Szyndlar, Z. 1991. A review of Neogene and Quaternary snakes of Central and Eastern Europe. Part I: Scolecophidia, Boidae, Colubrinae. Estudios Geologicos 47: 103–126.
Szyndlar, Z. 2005. Snake fauna from the Late Miocene of Rudabánya. Palaeontographia Italica 90: 31–52.
Szyndlar, Z. and Schleich, H. 1994. Two species of the genus Eryx (Serpentes; Boidae; Erycinae) from the Spanish Neogene with comments on the past distribution of the genus in Europe. Amphibia-Reptilia 15: 233–248. Crossref
Torres-Roig, E., Piñero, P., Agustí, J., Bover, P., and Alcover, J.A. 2019. First evidence of endemic Murinae (Rodentia, Mammalia) in the early Pliocene of the Balearic Islands (western Mediterranean). Geological Magazine 156: 1742–1750. Crossref
Van Dam, J.A. and Weltje, G.J. 1999. Reconstruction of the Late Miocene climate of Spain using rodent paleocommunity successions: an application of end-member modelling. Palaeogeography, Palaeoclimatology, Palaeoecology 151: 267–305. Crossref
Van Dam, J.A., Furió, M., and Van Balen, R.T. 2014. Re-interpreting the biochronology of the La Celia and Los Gargantones mammal sites (Late Miocene, Murcia, Spain). Geobios 47: 155–164. Crossref
Van de Weerd, A. 1976. Rodent faunas of the Mio-Pliocene continental sediments of the Teruel-Alfambra region, Spain. Utrecht Micropaleontological Bulletin, Special Publication 2: 1–217.
Van de Weerd, A., Adrover, R., Mein, P., and Soria, D. 1977. A new genus and species of the Cricetidae (Mammalia, Rodentia) from the Pliocene of south-western Europe. Proceedings Koninklij ke Nederlandse Akademie van Wetenschappen B 80: 429–439.
Wood, A.E. and Wilson, R.W. 1936. A suggested nomenclature for the cusps of the cheek teeth of rodents. Journal of Paleontology 10: 388–391.
Acta Palaeontol. Pol. 68 (1): 23–46, 2023
https://doi.org/10.4202/app.01013.2022