
Unusual Miocene hydrocarbon-seep faunas from the Brisighella area in northern Italy: embedded in clastics and first records of the lucinid bivalves Megaxinus and Miltha
STEFFEN KIEL, MARCO SAMI, and MARCO TAVIANI
Ancient hydrocarbon-seep sites known as “Calcari a Lucina” are common in Miocene strata of northern Italy and typically consist of carbonate deposits dominated by large lucinid, bathymodiolin, and vesicomyid bivalves. Here we report two new sites found in Upper Miocene strata at Monte Mauro near Brisighella in the Emilia-Romagna province. One is unusual by being embedded in unconsolidated siltstone without any carbonate, but yet, consisting of the typical, seep-restricted bivalves Bathymodiolus moroniae and Archivesica aharoni vesicomyid clams and bathymodiolin mussels. The second deposit is dominated by the lucinid Megaxinus bellardianus, which has never been reported from a Miocene seep deposit in this region, despite being common in coeval siliciclastic sediments nearby. This species emphasizes biogeographic relationships between Upper Miocene seep faunas in the Mediterranean region and the tropic Indo-West Pacific Ocean.
Methane seeps along continental margins harbor lush faunal communities composed mainly of animals living in symbiosis with chemotrophic bacteria, from which they derive their nutrition (Paull et al. 1985, Dubilier et al. 2008). Due to this in situ food source, these faunas are believed to have a unique evolutionary history, closely linked to that of similar faunal communities living at hydrothermal vents (McArthur and Tunnicliffe 1998; Vrijenhoek 2013). The anaerobic oxidation of methane at these sites facilitates carbonate precipitation (Peckmann and Thiel 2004; Cochran et al. 2022) and hence fossil examples of these faunas are a common feature of uplifted deep-water strata, especially from the late Mesozoic onward (Kiel 2010; Hryniewicz 2022). This fossil record has provided unique insights into the evolution of these ecosystems, showing a major role for brachiopods in their earlier history (Campbell and Bottjer 1995; Kiel and Peckmann 2019), recurrent morphological adaptations among different clades at different times (Hryniewicz et al. 2017), resistance towards extinction events (Kiel and Little 2006), and marine sulfate concentrations as a potential major driver of ecologic and evolutionary change (Kiel 2015).
The Mediterranean Sea today has a rather depauperate seep fauna compared to that of the major oceanic continental margins (Olu et al. 2004; Taviani et al. 2013; Taviani 2014). The fossil record indicates that this is a geologically young feature, as typical “oceanic” faunal elements such as large bathymodiolin mussels and vesicomyid clams were common and widespread at Miocene methane-seep deposits throughout Italy (Moroni 1966; Taviani 1994; Kiel and Taviani 2017). These seep deposits, locally known as “Calcari a Lucina” (Lucina limestone), occur from the Turin hills along the Apennine Mountain chain all the way to Calabria in the south (Manzoni 1876; Taviani 1994; Conti et al. 2021; Hryniewicz 2022). Despite their biogeographic and evolutionary importance, the documentation and identification of the taxa composing these faunas is still incomplete. The first thorough monograph of the “Calcari a Lucina” mollusk fauna was published by Moroni (1966), though this was before the discovery of vent and seep communities in the modern oceans (Corliss et al. 1979; Paull et al. 1984) and the rich taxonomic work on their fauna (Desbruyères et al. 2006). The present contribution is part of our ongoing efforts to document the taxonomic and ecologic diversity of seep faunas in the Neogene of Italy (Taviani et al. 2011; Vinn et al. 2012; Sami and Taviani 2015; Kiel and Taviani 2017, 2018; Kiel et al. 2018), to better understand the biogeographic and evolutionary history of the Mediterranean seep fauna.
Institutional abbreviations.—MSF, Museo Civico di Scienze Naturali, Faenza, Italy; NRM, Swedish Museum of Natural History, Stockholm, Sweden.
Results
Here we comment on two deposits with mollusks characteristic of the “Calcari a Lucina” fauna. They were found only a few meters apart, on the northern corner of a vineyard along Via Monte Mauro, at 44°13’59.6”N 11°42’21.2”E, SWW of Faenza and just to the west of Brisighella (Fig. 1). Stratigraphically, the sediments enclosing these deposits belong to the upper Tortonian–?lower Messinian (Upper Miocene) pre-evaporitic “Euxinic Shales” (Sami and Taviani 2019). The vineyard bordering the outcrops belong to the Tenuta Cassano vinery, and the sites were hence reported as Ca’ Cassano 1 and 2 by Sami and Taviani (2019), who briefly discussed their stratigraphy, lithology and associated macrofauna.
Ca’ Cassano 1 is a lens, about 60 cm thick and at least 250 cm wide, of unconsolidated silty sandstone, gray with ochre weathering surfaces. Molluscan fossils are preserved mainly as internal molds with little or no shell material (Table 1, Fig. 2), though small shell fragments are dispersed throughout the lens. The most abundant taxa are the bathymodiolin mussel Bathymodiolus moroniae Kiel and Taviani, 2017, reaching 10 cm in length, and the vesicomyid clam Archivesica aharoni Kiel and Taviani, 2017, up to 10 cm in length. Less common are specimens of the lucinid Lucinoma perusina (Sacco, 1901) and of the mytilid Samiolus iohannesbaptistae Kiel and Taviani, 2017. Gastropods are rare but relatively diverse, including the neritid Thalassonerita megastoma Moroni, 1966, a large buccinid previously identified as “Chrysodomus hoernesi Bellardi, 1873” by Sami and Taviani (2019) up to 8 cm tall, the colloniid Homalopoma domeniconii Moroni, 1966, a large gastropod (possibly cf. “Galeodea sp.”), and specimens commonly identified as the pyramidellid “Phasianema taurocrassum Sacco, 1895” (i.e., Moroni 1966), though we think it more likely belongs to the vetigastropod family Cataegidae (McLean and Quinn 1987; Warén and Rouse 2016).
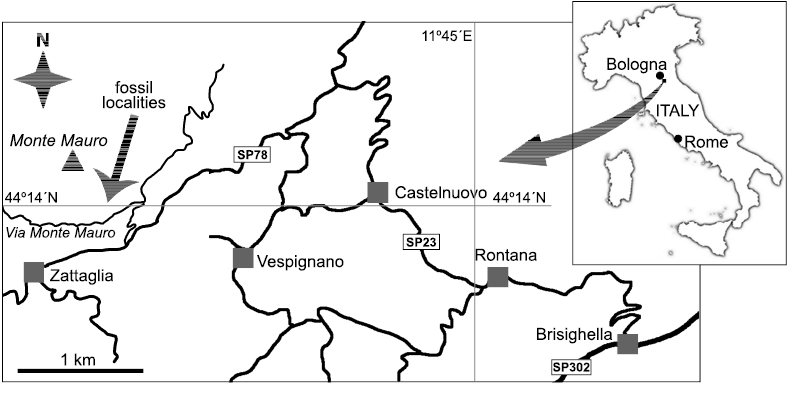
Fig. 1. The location of the Ca’ Cassano sites near Brisighella in northern Italy

Fig. 2. Mollusk fauna from the Upper Miocene of Italy, Ca’ Cassano 1 deposit, and the deposit itself. A. The lucinid bivalve Lucinoma perusina (Sacco, 1901), NRM Mo 197311, view on RV of internal mold showing elongate anterior adductor muscle scar. B. The mytilid bivalve Samiolus iohannesbaptistae Kiel and Taviani, 2017, MSF 2340, view on inner side of a LV. C. The mytilid bivalve Bathymodiolus moroniae Kiel and Taviani, 2017, NRM Mo 197312, view on LV of internal mold. D. The vesicomyid bivalve Archivesica aharoni Kiel and Taviani, 2017, NRM Mo 197313, view on LV of internal mold. E. Large gastropod, possibly “Galeodea sp.”, MSF 2341, internal mold. F. Large buccinid gastropod, MSF 2342, apertural view. G. Outcrop image of the deposit, hammerhead c. 18 cm wide.
The Ca’ Cassano 2 deposit, stratigraphically about 7 meters above Ca’ Cassano 1, is an isolated limestone deposit, consisting of gray, carbonate-cemented, detrital siltstone with occasional micritic patches, calcite-filled voids, finely dispersed, small shell fragments, and a diverse mollusk fauna (Table 1). The most common bivalve is the lucinid Megaxinus bellardianus (Mayer, 1864), reaching 43 mm in length, 40 mm in height, and 23 mm in thickness (Fig. 3D–F). It shows some variation in shell shape, especially regarding the position of the umbo, but the specimens match those reported as Megaxinus bellardianus by Sacco (1901: 75–77, pl. 17: 29–37). This taxon is recorded by Sami and Taviani (2019) as “Lucinoma sp.”. An extremely flat lucinid bivalve reaching 40 × 40 mm at a width of 13 mm is here identified as Miltha sp. (Fig. 3G–J). It was reported previously as “Myrtea sp.” by Sami and Taviani (2019), the ten available specimens show a style of preservation similar to that of Megaxinus bellardianus, indicating that the extremely flat shell is original and not the result of a taphonomic process. In contrast to Ca’ Cassano 1, the Ca’ Cassano 2 site also included a few large specimens of the lucinid bivalve Meganodontia hoernea (Des Moulins, 1868), reaching 15 cm in length. The gastropod fauna is similar to that of Ca’ Cassano 1.

Fig. 3. Mollusks of the Upper Miocene of Italy, Ca’ Cassano 2 deposit. A. The mytilid bivalve Bathymodiolus moroniae Kiel and Taviani, 2017, MSF 2343, view on RV of articulated specimen. B. The mytilid bivalve Samiolus iohannesbaptistae Kiel and Taviani, 2017, MSF 2344, isolated right valve. C. The lucinid bivalve Lucinoma perusina (Sacco, 1901), NRM Mo 197314, view on left valve of internal mold. D–F. The lucinid bivalve Megaxinus bellardianus (Mayer, 1864). D. NRM Mo 197315, articulated specimen showing sculpture and healed shell injuries. E. NRM Mo 197316, dorsal view showing lunule and narrow escutcheon. E. NRM Mo 197316, dorsal view showing lunule and narrow escutcheon. F. NRM Mo 197317, view on LV of internal mold showing pallial line, anterior adductor muscle scar, and pallial blood vessel scar. G–J. The lucinid bivalve Miltha sp. G. MSF 2345, internal mold, view on LV showing anterior adductor muscle scar. H. MSF 2346, outside of LV with preserved shell material. I. MSF 2347, outside of LV with preserved shell material. J. MSF 2348, dorsal view on articulated specimen. K. The vesicomyid bivalve Archivesica aharoni Kiel and Taviani, 2017, NRM Mo 197319, internal mold of RV. L. The colloniid gastropod Homalopoma domeniconii Moroni, 1966, MSF 2349, apertural view. M. The gastropod “Phasianema taurocrassum Sacco, 1895”, NRM Mo 197320, backside of specimen embedded in rock matrix. N. The neritid gastropod Thalassonerita megastoma (Moroni, 1966), NRM Mo 197321, backside of specimen embedded in rock matrix. O. The buccinid gastropod Neptunea hoernesi subdilatata Moroni, 1966, MSF 2350, apertural view. All specimens coated with ammonium chloride.
Discussion
Carbonate formation is a characteristic feature of hydrocarbon seeps because the main geochemical process, the anaerobic oxidation of methane, results in a local increase in alkalinity, triggering carbonate precipitation (Peckmann and Thiel 2004; Cochran et al. 2022). Hence, the mass occurrence in the unconsolidated siltstone lens of Ca’ Cassano 1 of taxa that are typically considered obligate to cold seeps, such as Bathymodiolus moroniae and Archivesica aharoni, is unusual; elsewhere in Miocene strata of the Apennines, these species are typically found in massive carbonate mounds (Taviani 1994, 2011; Conti et al. 2021). One exception are the vesicomyid-rich horizons (reported as ?Calyptogena sp., but most likely belonging to Archivesica) from carbonate-free sandy silty mudstone associated with submarine slides in the turbiditic Miocene Marnoso-Arenacea Formation in the Romagna Apennines (Lucente and Taviani 2005). Occurrences of seep-inhabiting bivalves, especially vesicomyids, in bedded siltstone or mudstone, are common in the Oligocene of Alaska (Kiel and Amano 2010) and the Neogene of Japan (Majima et al. 2005) and are considered as allochthonous. Re-deposition of the shells by a turbidity current is certainly a possible explanation for Ca’ Cassano 1.
Table 1. List of species found at the Ca’ Cassano sites in the Late Miocene-aged pre-evaporitic “Euxinic Shales” in the Emilia-Romagna province in northern Italy.
|
Species |
Ca’ Cassano 1 |
Ca’ Cassano 2 |
|
Bathymodiolus moroniae Kiel and Taviani, 2017 |
abundant |
uncommon |
|
Samiolus
iohannesbaptistae |
rare |
rare |
|
Lucinoma
perusina |
uncommon |
uncommon |
|
Meganodontia
hoernea |
|
uncommon |
|
Megaxinus
bellardianus |
|
abundant |
|
Miltha sp. |
|
common (40 mm) |
|
Archivesica
aharoni |
common |
uncommon |
|
Homalopoma
domeniconii |
rare |
rare |
|
“Phasianema
taurocrassum |
rare |
rare |
|
Thalassonerita
megastoma |
rare |
rare |
|
Large gastropod |
rare |
|
|
Neptunea
hoernesi subdilatata |
|
rare |
|
Large buccinid gastropod |
rare (80 mm) |
|
Alternatively, a few accumulations of “seep taxa” without associated carbonate precipitation, but on organic-rich sediments, are known from the modern ocean. Turbidity current deposits resulting from the 1929 Grand Banks earthquake on the Laurentian Fan in the deep NW Atlantic Ocean were colonized by vesicomyid and thyasirid clams, and siboglinid tube worms (Mayer et al. 1988). Likewise, vesicomyid bivalves are common in organic- and sulfide-rich sediments of the Congo deep-sea fan in the tropical eastern Atlantic Ocean, and are believed to sustain themselves on hydrogen sulfide generated by the degradation of the locally abundant organic matter (Olu et al. 2017). Yet another example are solemyid and nucinellid bivalves inhabiting sediments in fjords in New Zealand, where sulfide is generated by decomposing forest litter and marine algae (McLeod et al. 2010). Such settings with organic-rich sediments could be an alternative explanation for the mass occurrence of seep taxa in the unconsolidated siltstone lens of Ca’ Cassano 1. However, the faunal communities in the Laurentian and Congo fans and the New Zealand fjords host mainly infaunal and semi-infaunal bivalves but no bathymodiolins, unlike the Ca’ Cassano 1 deposit. Thus, re-deposition might be the more likely scenario for the formation of the Ca’ Cassano 1 deposit.
The lucinid Megaxinus is common in Neogene strata of the Mediterranean region (Sacco 1901), including at Pliocene seep deposits (Cau et al. 2015; Kiel and Taviani 2018) and whale falls (Danise et al. 2010). The genus occurs mostly in Neogene deposits of the Mediterranean region, and today in the Mediterranean Sea and western Indian Ocean (Glover and Taylor 1997). Megaxinus bellardianus found at Ca’ Cassano 2 is the first record of this genus and species at a Miocene Mediterranean seep deposit. A species of Megaxinus was recently reported from Upper Miocene seep deposits in the Philippines (Kiel et al. 2022), thus Megaxinus bellardianus from the Ca’ Cassano 2 deposit emphasizes faunal relationships between the Mediterranean region and the tropical Indo-West Pacific Ocean during the Miocene. Together with the flat Miltha sp., it also emphasizes the heterogeneity among faunal assemblages in the “Calcari a Lucina” seep deposits, as recently indicated by an Upper Miocene faunule including the lucinid bivalve Anodontia mioinflata, the bathymodiolin Idas aff. tauroparva, and abundant serpulid worms (Kiel et al. 2018).
Acknowledgements.—We thank the reviewers Kazutaka Amano (Joetsu University of Education, Japan) and Tom DeVries (Burke Museum, Seattle, USA) for their constructive reviews of the manuscript. Financial support was provided by the Swedish Research Council (Vetenskapsrådet) through grant 2016-03920 to SK. This is ISMAR-CNR Bologna scientific contribution n. 2070.
References
Bellardi, L. 1873. I Molluschi dei terreni Terziari del Piemonte e della Liguria, Parte I. Cephalopooda, Pteropoda, Heteropoda, Gastropoda (Muricidae et Tritonidae). 264 pp. Stamperia Reale, Torino. Crossref
Campbell, K.A. and Bottjer, D.J. 1995. Brachiopods and chemosymbiotic bivalves in Phanerozoic hydrothermal vent and cold seep environments. Geology 23: 321–324. Crossref
Cau, S., Franchi, F., Roveri, M., and Taviani, M. 2015. The Pliocene-age Stirone River hydrocarbon chemoherm complex (Northern Apennines, Italy). Marine and Petroleum Geology 66: 582–595. Crossref
Cochran, J.K., Landman, N.H., Jakubowicz, M., Brezina, J., Naujokaityte, J., Rashkova, A., Garb, M.P., and Larson, N.L. 2022. Geochemistry of cold hydrocarbon seeps: an overview. In: A. Kaim, N.H. Landman, and J.K. Cochran (eds.), Ancient Hydrocarbon Seeps. Topics in Geobiology 53: 267–321. Crossref
Conti, S., Argentino, C., Fioroni, C., Salocchi, A.C., and Fontana, D. 2021. Miocene seep-carbonates of the northern Apennines (Emilia to Umbria, Italy): an overview. Geosciences 11: 53. Crossref
Corliss, J.B., Dymond, J., Gordon, L.I., Edmond, J.M., Von Herzen, R.P., Ballard, R.D., Green, K., Williams, D., Bainbridge, A., Crane, K., and Van Andel, T.H. 1979. Submarine thermal springs on the Galápagos Rift. Science 203: 1073–1083. Crossref
Danise, S., Dominici, S., and Betocchi, U. 2010. Mollusk species at a Pliocene shelf whale fall (Orciano Pisano, Tuscany). Palaios 25: 449–456. Crossref
Desbruyères, D., Segonzac, M., and Bright, M. 2006. Handbook of deep-sea hydrothermal vent fauna. Second completely revised version. Denisia 18: 1–544.
Dubilier, N., Bergin, C., and Lott, C. 2008. Symbiotic diversity in marine animals: the art of harnessing chemosynthesis. Nature Reviews Microbiology 6: 725–740. Crossref
Glover, E.A. and Taylor, J.D. 1997. New species and records of Rastafaria and Megaxinus (Bivalvia: Lucinidae) from the western Indian Ocean and Red Sea, with a reappraisal of Megaxinus. Journal of Conchology 36: 1–18.
Hryniewicz, K. 2022. Ancient hydrocarbon seeps of the world. In: A. Kaim, N.H. Landman, and J.K. Cochran (eds.), Ancient Hydrocarbon Seeps. Topics in Geobiology 53: 571–647. Crossref
Hryniewicz, K., Jakubowicz, M., Belka, Z., Dopieralska, J., and Kaim, A. 2017. New bivalves from a Middle Devonian methane seep in Morocco: the oldest record of repetitive shell morphologies among some seep bivalve molluscs. Journal of Systematic Palaeontology 15: 19–41. Crossref
Kiel, S. 2010. The fossil record of vent and seep mollusks. In: S. Kiel (ed.), The Vent and Seep Biota. Topics in Geobiology 33: 255–278. Crossref
Kiel, S. 2015. Did shifting seawater sulfate concentrations drive the evolution of deep-sea vent and seep ecosystems? Proceedings of the Royal Society B 282: 20142908. Crossref
Kiel, S. and Amano, K. 2010. Oligocene and Miocene vesicomyid bivalves from the Katalla district in southern Alaska, USA. The Veliger 51: 76–84.
Kiel, S. and Little, C.T.S. 2006. Cold seep mollusks are older than the general marine mollusk fauna. Science 313: 1429–1431. Crossref
Kiel, S. and Peckmann, J. 2019. Resource partitioning among brachiopods and bivalves at ancient hydrocarbon seeps: a hypothesis. PLoS ONE 14: e0221887.
Kiel, S. and Taviani, M. 2017. Chemosymbiotic bivalves from Miocene methane-seep carbonates in Italy. Journal of Paleontology 91: 444–466. Crossref
Kiel, S. and Taviani, M. 2018. Chemosymbiotic bivalves from the late Pliocene Stirone River hydrocarbon seep complex in northern Italy. Acta Palaeontologica Polonica 63: 557–568. Crossref
Kiel, S., Fernando, A.G., Magtoto, C.Y., and Kase, T. 2022. Mollusks from Miocene hydrocarbon-seep deposits in the Ilocos-Central Luzon Basin, Luzon Island, Philippines. Acta Palaeontologica Polonica 67: 917–947. Crossref
Kiel, S., Sami, M., and Taviani, M. 2018. A serpulid-Anodontia-dominated methane-seep deposit from the Miocene of northern Italy. Acta Palaeontologica Polonica 63: 569–577. Crossref
Lucente, C.C. and Taviani, M. 2005. Chemosynthetic communities as fingerprints of submarine sliding-linked hydrocarbon seepage, Miocene deep-sea strata of the Turscan-Romagna Apennines, Italy. Palaeogeography, Palaeoclimatology, Palaeoecology 227: 176–190. Crossref
Majima, R., Nobuhara, T., and Kitazaki, T. 2005. Review of fossil chemosynthetic assemblages in Japan. Palaeogeography, Palaeoclimatology, Palaeoecology 227: 86–123. Crossref
Manzoni, A. 1876. Della posizione geografica del calcare a Lucina pomum, Mayer. Bollettino del Regio Comitato Geologico Italiano 7: 209–216.
Mayer, L.A., Shor, A.N., Huges Clarke, J., and Piper, D.J.W. 1988. Dense biological communities at 3850 m on the Laurentian Fan and their relationship to the deposits of the 1929 Grand Banks earthquake. Deep-Sea Research 35: 1235–1246. Crossref
McArthur, A.G. and Tunnicliffe, V. 1998. Relics and antiquity revisited in the modern vent fauna. Geological Society of London, Special Publications 148: 271–291. Crossref
McLean, J.H. and Quinn, J.F.J. 1987. Cataegis, new genus of three new species from the continental slope (Trochidae: Cataeginae new subfamily). The Nautilus 101: 111–116. Crossref
McLeod, R.J., Wing, S.R., and Skilton, J.E. 2010. High incidence of invertebrate-chemoautotroph symbioses in benthic communities of the New Zealand fjords. Limnology and Oceanography 55: 2097–2106. Crossref
Moroni, M.A. 1966. Malacofauna del “Calcare a Lucine” di S. Sofia, Forlì. Palaeontographica Italica 60: 69–87.
Olu, K., Decker, C., Pastor, L., Caprais, J.C., Khripounoff, A., Morineaux, M., Ain Baziz, M., Menot, L., and Rabouille, C. 2017. Cold-seep-like macrofaunal communities in organic- and sulfide-rich sediments of the Congo deep-sea fan. Deep-Sea Research II 142: 180–196. Crossref
Olu, K., Sibuet, M., Fiala-Médoni, A., Gofas, S., Salas, C., Mariotti, A., Foucher, J.-P., and Woodside, J. 2004. Cold seep communities in the deep eastern Mediterranean Sea: composition, symbiosis and spatial distribution on mud volcanoes. Deep-Sea Research I 51: 1915–1936. Crossref
Paull, C.K., Hecker, B., Commeau, R., Freeman-Lynde, R.P., Neumann, C., Corso, W.P., Golubic, S., Hook, J.E., Sikes, E., and Curray, J. 1984. Biological communities at the Florida Escarpment resemble hydrothermal vent taxa. Science 226: 965–967. Crossref
Paull, C.K., Jull, A.J.T., Toolin, L.J., and Linick, T. 1985. Stable isotope evidence for chemosynthesis in an abyssal seep community. Nature 317: 709–711. Crossref
Peckmann, J. and Thiel, V. 2004. Carbon cycling at ancient methane-seeps. Chemical Geology 205: 443–467. Crossref
Sacco, F. 1901. I molluschi dei terreni terziarii del Piemonte e della Liguria. Parte XXIX. 217 pp. Carlo Clausen, Torino.
Sami, M. and Taviani, M. 2015. I Calcari a Lucina e i Gessi di Rontana. I Gessi di Brisighella e Rontana. Memorie dell’Istituto Italiano di Speleologia, Ser. II 28: 39–56.
Sami, M. and Taviani, M. 2019. La vita nei mari prima dei Gesso: I “Calcari a Lucina” e i carbonati pre-evaporitici di Monte Mauro. Memorie dell’Istituto Italiano di Speleologia, Ser. II 34: 49–73.
Taviani, M. 1994. The “calcari a Lucina” macrofauna reconsidered: Deep-sea faunal oases from Miocene-age cold vents in the Romagna Apennine, Italy. Geo-Marine Letters 14: 185–191. Crossref
Taviani, M. 2011. The deep-sea chemoautotroph microbial world as experienced by the Mediterranean metazoans through time. In: J. Reitner, N.-V. Quéric, and G. Arp (eds.), Advances in Stromatolite Geobiology. Lecture Notes in Earth Sciences 131: 277–295. Crossref
Taviani, M. 2014. Marine chemosynthesis in the Mediterranean Sea. In: S. Goffredo and Z. Dubinsky (eds.), The Mediterranean Sea: Its History and Present Challenges, 69–83. Springer, Dordrecht. Crossref
Taviani, M., Angeletti, L., and Ceregato, A. 2011. Chemosynthetic bivalves of the family Solemyidae (Bivalvia, Protobranchia) in the Neogene of the Mediterranean Basin. Journal of Paleontology 85: 1067–1076. Crossref
Taviani, M., Angeletti, L., Ceregato, A., Foglini, F., Froglia, C., and Trincardi, F. 2013. The Gela Basin pockmark field in the strait of Sicily (Mediterranean Sea): chemosymbiotic faunal and carbonate signatures of postglacial to modern cold seepage. Biogeosciences 10: 4653–4671. Crossref
Vinn, O., Kupriyanova, E.K., and Kiel, S. 2012. Systematics of serpulid tubeworms (Annelida, Polychaeta) from Cretaceous and Cenozoic hydrocarbon-seep deposits in North America and Europe. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 265: 315–325. Crossref
Vrijenhoek, R.C. 2013. On the instability and evolutionary age of deep-sea chemosynthetic communities. Deep-Sea Research II 92: 189–200. Crossref
Warén, A. and Rouse, G.W. 2016. A new genus and species of Cataegidae (Gastropoda: Seguenzioidea) from eastern Pacific Ocean methane seeps. Novapex 17: 59–66.
Steffen Kiel [steffen.kiel@nrm.se; ORCID: https://orcid.org/0000-0001-6281-100X ], Swedish Museum of Natural History, Department of Palaeobiology, Box 50007, 10405 Stockholm, Sweden.
Marco Sami [marco.sami@cheapnet.it; ORCID: https://orcid.org/0000-0003-2936-1603 ], Museo Civico di Scienze Naturali, 51, Via Medaglie D’Oro 51, 48018 Faenza, Italy.
Marco Taviani [marco.taviani@bo.ismar.cnr.it; ORCID: https://orcid.org/0000-0003-0414-4274 ], Institute of Marine Sciences, Italian National Research Council, Via Gobetti 101, 40129 Bologna, Italy; and Stazione Zoologica Anton Dohrn, Villa Comunale, 80121 Napoli, Italy.
Received 6 September 2022, accepted 2 December 2022, available online 19 January 2023.
Copyright © 2023 S. Kiel et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Acta Palaeontol. Pol. 68 (1): 127–132, 2023
https://doi.org/10.4202/app.01021.2022