
Pereboriidae: a Permian clade of hemipteran insects with disjunctive distribution in the Northern and Southern parts of Pangea
ANDRÉ NEL, EKHONA ZOZO, ROMAIN GARROUSTE, and ROSE PREVEC
Nel, A., Zozo, E., Garrouste, R., and Prevec, R. 2023. Pereboriidae: a Permian clade of hemipteran insects with disjunctive distribution in the Northern and Southern parts of Pangea. Acta Palaeontologica Polonica 68 (1): 85–93.
Afropereboria magnifica gen. et sp. nov., is the oldest African representative of the Permian “cicada-like” family Pereboriidae. The family diagnosis is discussed and emended. Synapomorphic characters of the Pereboriidae are proposed. The two genera Jifara and Scytophara are excluded from this family, confirming that Pereboriidae are strictly Permian. The relationships between the two Permian families Pereboriidae and Ignotalidae and the Triassic Curvicubitidae are still not clear and will need a phylogenetic analysis. We propose new putatively apomorphic diagnostic characters for the Pereboriidae. We also discuss some characters supposed to discriminate the Pereboriidae from the Ignotalidae. The Permian Pereboriidae had a disjunctive distribution between the northern and southern parts of Pangea, under cool temperate climates, separated by the intertropical zones.
Key words: Insecta, Hemiptera, Pereborioidea, disjunctive distribution, Pangea, Africa.
André Nel [anel@mnhn.fr; ORCID: https://orcid.org/0000-0002-4241-7651 ] (corresponding author) and Romain Garrouste [garroust@mnhn.fr; ORCID: https://orcid.org/0000-0002-0880-5781 ], Institut de Systématique, Évolution, Biodiversité (ISYEB) Muséum national d’Histoire naturelle, CNRS, Sorbonne Université, EPHE, Université des Antilles, CP50, 57 rue Cuvier 75005 Paris, France.
Ekhona Zozo [ekhonazozo@gmail.com; ORCID: https://orcid.org/0000-0001-5351-6180 ], Department of Zoology and Entomology, Albany Museum, Makhanda, South Africa
Rose Prevec [r.prevec@am.org.za; ORCID: https://orcid.org/0000-0001-6674-1547 ] (corresponding author), Department of Earth Sciences, Albany Museum, Makhanda, South Africa & Department of Botany, Rhodes University, Makhanda, South Africa.
Received 13 September 2022, accepted 31 November 2022, available online 19 January 2023.
Copyright © 2023 A. Nel et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Pereboriidae is a small family of large Permian cicada-like hemipterans, mainly known from their forewings (tegmina). Despite the revision by Shcherbakov (1984), this family is still rather poorly defined. Some taxa, currently attributed to this group, probably belong to other lineages. Their taxa are currently recorded from Russia and Brazil, and an enigmatic genus and species described from Southern Africa (Riek 1976). Thus, the family seems to have a disjunctive distribution between the northern and southern parts of Pangea during the Permian.
Over the past four years, our team has discovered and searched for fossil plants and insects in the new fossil locality of Onder Karoo in the south-western Karoo Basin, near Sutherland in the Northern Cape Province of South Africa. It has yielded an extraordinary quantity of arthropods, some of which correspond to the first African records of some clades (Prevec et al. 2022).
Here we describe the oldest cicada-like insects of the family Pereboriidae from Africa. We complement the diagnosis of the family and discuss the phylogenetic positions of the included genera.
Institutional abbreviations.—AM, Albany Museum, Makhanda, South Africa.
Other abbreviations.—C, costa; CuA, cubitus anterior; CuP, cubitus posterior; M, median vein; RA, radius anterior; RP, radius posterior; ScP, subcosta posterior.
Nomenclatural acts.—This work and the nomenclatural acts it contains, have been registered in ZooBank: urn:lsid:zoobank.org:pub:DFE39AD5-8510-4BFB-AB94-CD419F103423
Material and methods
The fossils were collected under a South African Heritage Resources Agency (SAHRA) permit (ID # 2310 to RP), and the specimens have been accessioned to the palaeontology collections of the Department of Earth Sciences at the Albany Museum, Makhanda, Eastern Cape, South Africa. They were photographed with a Nikon D7200 SLR digital camera, and examined with a Zeiss Discovery V8 incident light microscope. To improve photo quality, we used polarizing filters and alcohol immersion. Drawings were made using Adobe Creative Cloud software. The fossils were excavated from a highly fossiliferous, olive-grey siltstone layer at the Onder Karoo locality (Sutherland District of the Northern Cape Province, South Africa; see Prevec et al. 2022). The host rocks are attributed to the lowermost Abrahamskraal Formation, Beaufort Group, Karoo Supergroup, close to the boundary with the underlying Waterford Formation of the Ecca Group. Regional biostratigraphy, stratigraphic correlation and geochronology, as well as an age estimate obtained from U-Pb dating of zircons from a clay layer at the base of the fossiliferous deposit, have provided an age of 266–269 Ma for the fossil assemblage, placing it within the Wordian Stage (Guadelupian) (see Prevec et al. 2022).
We follow the wing venation terminology of Nel et al. (2012) and Bourgoin et al. (2015) for the Acercaria (including Hemiptera).
Systematic palaeontology
Insecta Linnaeus, 1758
Hemiptera Linnaeus, 1758
Prosbolopsemorpha Szwedo, 2018
Pereborioidea Zalessky, 1930
Pereboriidae Zalessky, 1930
Type genus: Pereboria Zalessky, 1930; Baitugan Formation, Roadian–Wordian (Guadelupian), Perebore, Russian Federation.
Included genera: Gondwanaptera Pinto and Pinto de Ornellas, 1981 (Kungurian, Cisuralian, Permian, Irati Formation, Paraná Basin, Brazil), Neuropibrocha Becker-Migdisova, 1961 (Roadian–Wordian, Guadelupian, Permian, Kuznetsk Basin, Russian Federation), Afropereboria gen. nov. (see below), possibly also Borisrohdendorfia Becker-Migdisova, 1959 (Roadian–Wordian, Guadelupian, Permian, Kuznetsk Basin, Russian Federation) and Perissovena Riek, 1976 (Lopingian, Permian, Normandien Formation, KwaZulu-Natal, South Africa). We support the exclusion of Jiphara Ren in Ren et al., 1995 (Lower Cretaceous, China) as proposed by Shcherbakov (2020), and exclude Scytophara Martynov, 1937 (upper Permian, Kargala, Russian Federation) (see below).
Genus Afropereboria nov.
Zoobank LSID: urn:lsid:zoobank.org:act:968E0FE8-94F5-4BF4-921 B-004DD547CB28
Type species: Afropereboria magnifica sp. nov.; by monotypy, see below.
Etymology: Named after Africa and Pereboria.
Diagnosis.—Forewing characters: base of RP much more basal to level of branches of M; all branches of M well-defined; area of RP broad with numerous branches; numerous small rectangular cells in apical part of forewing.
Stratigraphic and geographic range.—Wordian Stage (Guadelupian), Onder Karoo locality, Sutherland District of the Northern Cape Province, South Africa.
Afropereboria magnifica sp. nov.
Figs. 1–5.
Zoobank LSID: urn:lsid:zoobank.org:act:458DB3B7-F3B7-4C07-AD 11-F7D9EBDB6687
Etymology: Named after the wonderful state of preservation of the wings, showing traces of coloration and detailed venation.
Type material: Holotype: AM11157a, b, adpression of a forewing, part and counterpart. Paratypes: AM11447a, b, adpression of a hind wing, part and counterpart; AM11141a, b, adpression of a forewing, part and counterpart. All form type locality and horizon.
Type horizon: Wordian Stage (Guadelupian).
Type locality: Onder Karoo locality, Sutherland District of the Northern
Cape Province, South Africa.
Diagnosis.—As for the genus. Presence of several broad dark zones in apical part of forewing and of hind wing.
Description.—Holotype (AM11157a, b; Figs. 1, 2). Part and counterpart of a forewing, with anal area (clavus) and base of wing missing. Wing with dark bands in apical part and a large dark spot between RP and M(P); basal half punctuate, probably sclerotized; wing large, preserved part 40 mm long, wing ca. 15.7 mm wide; costal area between C and R+ScP broad, 3.7 mm wide, with a series of transverse irregular veinlets, more or less curved; base of ScP, if present, not preserved; ScP apparently fused or appressed to R distally; concave RP separating from RA 31.7 mm from wing apex; RA with several anterior branches with a net of cells between RA and C near wing apex, one long posterior branch of RA between main RA and RP; RP divided into two main branches 4.3 mm distal of its base, anterior branch of RP subdivided into two branches, each of them being bifurcate; posterior branch of RP posteriorly pectinate with four branches; a very broad and large area between RP and M, 5.7 mm wide and ca. 20.6 mm long, with a series of irregular more or less transverse veinlets defining large cells between them; M divided into four main branches, each of them being forked several times; no nodal line at all, but one distal crossvein rp-m; a very broad and long area between M and CuA, 4.2 mm wide, with a series of irregular more or less transverse veinlets defining large cells between them; CuA long and posteriorly curved, CuA posteriorly pectinate with at least four branches, 12.8 mm from its apex; even if the basal parts of M and CuA are not preserved, their general courses are clearly converging, so that these veins are certainly basally fused; a net of crossveins dividing apical part of wing into small rectangular cells; concave CuP simple and straight.

Fig. 1. Hemipteran insect Afropereboria magnifica gen. et sp. nov. (holotype AM11157a) from Onder Karoo locality, South Africa, lowermost Abrahamskraal Formation, Wordian Stage (Permian). Forewing, photograph (A1), explanatory drawing (A2).

Fig. 2. Hemipteran insect Afropereboria magnifica gen. et sp. nov. (holotype AM11157b) from Onder Karoo locality, South Africa, lowermost Abrahamskraal Formation, Wordian Stage (Permian). Forewing, photograph dry specimen (A1), under alcohol (A2).
Paratype (AM11447a, b; Figs. 3). Part and counterpart of distal part of a hind wing. Wing with a dark band along margin and a large dark spot between M and CuA; wing large, preserved part 29.2 mm long and 24.2 mm wide; Costal area and ScP not preserved; RA with several anterior branches with a net of cells between RA and C near wing apex, RP with three main branches, each subdivided several times; a broad and large area between RP and M, at least 3.5 mm wide in its distal part, with a series of irregular more or less transverse veinlets defining large cells between them; distal crossvein rp-m indistinct; M forked twice, and covering a rather narrow area; CuA with an anterior branch posteriorly pectinate and a trifurcation on posterior branch; no nodal line at all; a very broad and long area between CuA and CuP, at least 4.5 mm wide, with a series of irregular more or less transverse veinlets defining large cells between them; concave CuP long, curved and simple anal vein posteriorly pectinate; a broad and long area between CuP and anal vein.
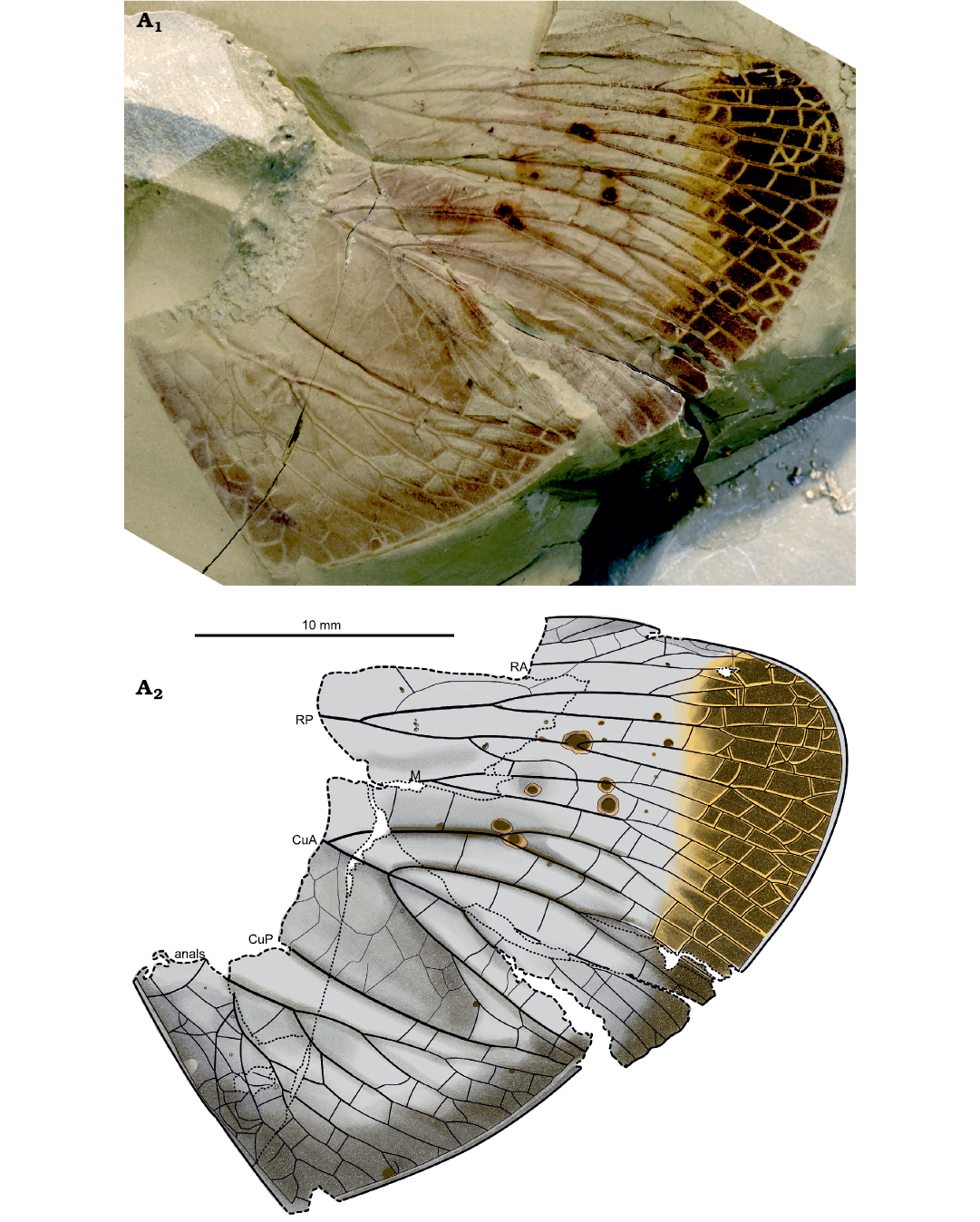
Fig. 3. Hemipteran insect Afropereboria magnifica gen. et sp. nov. (paratype AM11447a) from Onder Karoo locality, South Africa, lowermost Abrahamskraal Formation, Wordian Stage (Permian). Hind wing, photograph under alcohol (A1), explanatory drawing (A2).
Paratype (AM11141a, b; Figs. 4, 5). Adpression of distal half of a forewing, ca. 34.0 mm long; pattern of coloration the same as in holotype; branching of veins RA, RP and M also nearly the same as in holotype, especially for trifurcation on posterior branch of M; a very broad and large area between RP and M, 5.5 mm wide; distal half of the area between ScP and Costa preserved, very broad, 4.8 mm wide, apparently more sclerotized than rest of wing; ScP appressed to R/RA; series of long oblique veinlets emerging from ScP weakly marked and narrower than other veins.
Stratigraphic and geographic range.—Onder Karoo locality, lowermost Abrahamskraal Formation, Wordian Stage.

Fig. 4. Hemipteran insect Afropereboria magnifica gen. et sp. nov. (paratype AM11141a), from Onder Karoo locality, South Africa, lowermost Abrahamskraal Formation, Wordian Stage (Permian). Forewing, photograph under alcohol.

Fig. 5. Hemipteran insect Afropereboria magnifica gen. et sp. nov. (paratype AM11141b), from Onder Karoo locality, South Africa, lowermost Abrahamskraal Formation, Wordian Stage (Permian). Forewing, photograph dry specimen.
Discussion
All three specimens clearly belong to the same species characterized by the multiplication of the cells and veinlets in distal part of wing, similar patterns of coloration and similar venations. AM11157 and AM11141 are forewings because of the presence of a straight and concave CuP, a broad area between C and R(+ScP), and the shape of CuA and M. AM11141 has a broad costal area, typical of forewings of Cicadomorpha, while the hind wings have much narrower ones. AM11447 corresponds to a hind wing with a broad anal area. AM11141 differs from AM11157 in the presence of a trifurcation of posterior branch of M, a character not sufficient for a species separation.
The numerous branches of RA, RP, M, and CuA, plus the presence of numerous cells in the apical part of wing are characters of the Permian cicada-like Pereboriidae. These tegmina are very similar to those of Gondwanaptera (Permian of Brazil), Neuropibrocha (Permian of Russia), and Pereboria (Permian of Russia). Thus Afropereboria gen. nov. can be accurately attributed to this family.
The previously described taxa in this family are based on forewings, thus the present discovery of a hind wing gives new information on this structure. The hind wing differs from the forewing in the narrower area between M and CuA, the vein M covering a narrower area than in forewing, the broader area between CuA and CuP. Another remarkable character is the presence of a broad anal area with numerous branches of anal vein.
Shcherbakov (1984) considered that the Pereboriidae is the type family of the Pereborioidea, a group that would also comprise the Ignotaliidae Riek, 1973. Shcherbakov (2000: fig. 2) and Shcherbakov and Popov (2002) considered the Pereboriidae as a “grade”, a paraphyletic group that has given birth to the Permian Ignotalidae Riek, 1973, and the Triassic Curvicubitidae Hong, 1984. Shcherbakov (1996) supposed that the Pereboriidae should be “the more primitive of the three”, which is incongruous from a phylogenetic perspective.
Szwedo et al. (2004: 16) attributed the Permian family Prosbolopseidae Becker-Migdisova, 1946, to the Pereborioidea, but the same authors also indicated that the Prosbolopseidae belong to “the paraphyletic group Prosboloidea Handlirsch, 1906, after Shcherbakov (1984)”. Szwedo (2018) included the Pereborioidea (for Curvicubitidae, Ignotalidae, and Pereboriidae) in a new clade Prosbolopsemorpha.
Shcherbakov (1984) proposed the following diagnosis for the Pereborioidea: “costal break absent; costal ‘complex of veins’ margin/‘hypocosta’ at base not separated, if weakly split, then with notch near the middle; CP not subdivided at base, if it weakly expressed then the costal field has a median projection. Sc without arc, fusing with Ra or R, or free over its whole extent, distally convex. Ra bears post-nodal branches. Nodal line expressed weakly or absent. Transverse m-cu (first) short, not passing along nodal line, without longitudinal segment. CuA without displacement along nodal line. Anal field wide, weakly inclined (the structure of the field in the Ignotaliidae is unknown). Macrosculpture, if expressed, is concentrated in the prenodal part of the wing, and in the post-nodal part is only developed in the form of patches, covered by granules”. He added the following diagnosis for the Pereboriidae: “Wing anterior margin at base moderately convex, edge pronounced, CP well developed, costal margin without median notch. ScP falls into RA or into R. First segment of CuA developed (basal cell distinct). Macrosculpture developed.”
The polarity “absence of a notch in median portion of costal margin (median projection)” is uncertain because of the lack of phylogenetic analysis. Also, a notch is present in Gondwanaptera and Neuropibrocha, and this part of the forewing is unknown in Pereboria (Zalessky 1930; Pinto and Pinto de Ornellas 1981: pl. 1). Also, the “presence of a notch” is a doubtful character for the Ignotaliidae, as no such notch is figured in Riek (1973: fig. 10) for Ignotala mirifica Riek, 1973, and the costal margin is not preserved in Megoniella multinerva Riek, 1973, second taxon listed in Ignotaliidae by Shcherbakov (1984). It would seem likely that the Pereboriidae have a notch and the Ignotaliidae have none.
Shcherbakov (1996: 37) indicated that the Ignotalidae are characterized by the vein ScP being “secondarily released” and “the basal cell of the forewing opened at apex”. The first character is not present in Afropereboria, precluding any affinities with this family.
Shcherbakov (2022: 230) proposed the following emended diagnosis for the Ignotaliidae: “moderately to very large, extremely polyneurous cicadas. Tegmen: Costal margin strongly convex at least proximally, area anterior to Sc wide. Sc free or distally connected with RA, with weak prenodal branches. Basal cell reduced. Membrane without punctures or granules. Hind wing: Costal margin strongly convex at base, nearly straight distally, with series of coupling hooks. R stem short and oblique; R with several, M with few, and CuA with many branches in bundle.” The ScP appressed to R/RA and the membrane with punctures exclude Afropereboria gen. nov. from this family.
The Curvicubitidae currently comprises the two genera Beaconiella Evans, 1963 (Middle Triassic of Australia) and Curvicubitus Hong, 1984 (considered a junior synonym of Beaconiella by Shcherbakov 2021) (Triassic of China), after Shcherbakov (1996) (see also Evans 1963; Szwedo et al. 2004). Note that Hamilton (1992: 428) suggested to erect a new family for Beaconiella but did not formally make it. Both have numerous branches of RA, RP, M, and CuA, but both differ from the Pereboriidae and Afropereboria in the area between RP and RA being distinctly broader than that between RP and M, and with the presence of a distinct nodal line (Evans 1963; Hong 1984; Shcherbakov 1996, 2021). Beaconiella lacks the numerous posterior branches of RA.
It is not easy at all to define the states of characters that would be putative apomorphies for the Pereboriidae. Nevertheless, Afropereboria gen. nov. has the following characters of the Pereboriidae: ScP basally fused or closely appressed to Radius; no nodal line. The characters of the forewing base are not preserved.
Nevertheless, after re-examining the original figures of Gondwanaptera, Neuropibrocha, and Pereboria, we propose a new set of characters appropriate for these taxa, meaning, very broad and large areas of same widths, between RP and M and between M and CuA, traversed by irregular veinlets and with their anterior and posterior margins (R, M and CuA) distinctly curved (putative apomorphies to separate them from the Ignotaliidae); a broad area between C and R(+ScP), with a series of long veinlets emerging obliquely from ScP, narrower than the other veins; RA with several distal anterior and posterior branches; RP and M with numerous branches, CuA with several distal branches separating at the same point; a net of small cells in apical part of wing. These characters (especially the presence of several anterior branches of RA, and of broad and large areas between RP, M and CuA, and the numerous branches of these main veins) can be considered as putative synapomorphies of these taxa. They complete the diagnosis proposed by Shcherbakov (1984).
All these characters are present in Afropereboria gen. nov. It has no nodal line, a character that discriminates it from the Prosboliidae. Some taxa in this latter family have also numerous branches of main veins, e.g., Dictyoprosbole membranosa Martynov, 1935 (Martynov 1935; Becker-Migdisova 1961).
Afropereboria differs from Gondwanaptera as follows: base of RP much more basad level of branches of M vs. just distad them in Gondwanaptera; branches of M all well-defined vs. only two main well-defined branches in Gondwanaptera; area of RP broader with more numerous branches in Afropereboria; the numerous small cells in apical part of forewing are very irregular in Gondwanaptera vs. rectangular in Afropereboria; CuA has a long and simple anterior branch in Afropereboria, absent in Gondwanaptera (Pinto and Pinto de Ornellas 1981).
Afropereboria differs from Pereboria and Neuropibrocha in the larger area of RP, of the same width as that of M, vs. distinctly narrower in Pereboria and Neuropibrocha (Zalessky 1930; Becker-Migdisova 1961). Afropereboria shares with Neuropibrocha the rather rectangular small cells of apical part of forewing, while these are irregular in Pereboria.
Borisrohdendorfia Becker-Migdisova, 1959, a genus based on the extreme apical part of a forewing, is currently attributed to the Pereboriidae. This taxon shares with this family the numerous branches of main veins plus the presence of a series of small cells between them. These cells are very irregular, while they are rectangular in Afropereboria (Becker-Migdisova 1959; Rohdendorf 1962). The attribution of Borisrohdendorfia to the Pereboriidae remains rather poorly supported due to the lack of crucial structures of the basal two-third of the wing.
Perissovena Riek, 1976, is an unusual fossil from the upper Permian of KwaZulu-Natal Province, South Africa. It is based on a wing, probably a forewing, with the basal and apical parts missing. Riek (1976) only tentatively attributed it to the Pereboriidae. It shares with the Pereboriidae the numerous branches of main veins and large areas between RP, M, and CuA. However, it lacks the net of small cells in the distal part of wing that is typical of the group. It strongly differs from Afropereboria in the very large area of CuA while that of M is strongly reduced, and in the branched posterior branch of RA (Riek 1976).
Several other genera have been placed in the Pereboriidae, but these attributions are equivocal. In Jiphara (Mesozoic of China), RA, RP, M, and CuA branch many times, as is typical of the Pereboriidae, but it strongly differs from this family in lacking the numerous small cells in the apical part of the forewing, and in the absence of large and broad areas between RP, M, and CuA (Ren et al. 1995). As many extant Hemiptera similarly have numerous branches of R, M, and CuA, these characters are not sufficient for an attribution to the Pereboriidae. We support the exclusion of Jiphara from this family as proposed by Shcherbakov (2020). The removal of this Mesozoic taxon from consideration supports our assertion that the Pereboriidae are an exclusively Permian family.
Scytophara Martynov, 1937, is based on a complete forewing from the Permian of Russia. Its RA has numerous branches, but RP and CuA have very few; also the areas between RP, M, and CuA are not especially broad and large (Martynov 1937). Its attribution to the Pereboriidae is unlikely.
Conclusions
The present discoveries demonstrate that the Pereboriidae were clearly present in the Permian of Africa. The Pereboriidae seem to have had a disjunctive palaeo-distribution, in sub-Angara (Siberia) and Gondwana (South America and Africa). These different areas of Pangea were all under cool temperate climates, but were separated by the much warmer and drier intertropical zones (Shi and Waterhouse 2010). Similar Permian distributions are known for some “mixed floral elements” of northern and southern origins. Srivastava and Agnihotri (2010) suggested that the antiquity of these plant clades could explain this phenomenon, viz. “their ancestry or affiliation with Carboniferous floras” that shared a much broader global distribution prior to the development of a strong intertropical barrier. This hypothesis seems unlikely for the Pereboriidae that have no representatives at all in the Carboniferous. Thus the distribution of the Pereboriidae suggests that some faunal exchanges of insects between the southern and northern Permian land masses were possible during the Permian, despite the climatic obstacles, maybe along the coastal margins of Pangea. Similar results have been observed among the vertebrate Therapsida (Smith et al. 2006), although these need to be taken with caution in some cases (Angielczyk and Kurkin 2003).
Acknowledgements
We thanks an anonymous referee and Jacek Szwedo (University of Gdańsk, Poland) for their very useful remarks on the first version of the paper. We thank Sydney Moyo (Albany Museum, Grahamstown, Republic of South Africa), Martin Villet (Rhodes University, Grahamstown, Republic of South Africa) for discussions, and the teams of students and preparators involved in excavating and sorting the many fossil specimens collected from the Onder Karoo locality in recent years. This research was funded in part by grants to RP from Genus (Department of Science and Innovation—National Research Foundation, Centre of Excellence in Palaeosciences): CoE2016-600, CoE2016-496, CoE2017-059; and the National Research Foundation, African Origins Platform: 98822, 117685.
References
Angielczyk, K.D., and Kurkin, A.A. 2003. Has the utility of Dicynodon for Late Permian terrestrial biostratigraphy been overstated? Geology 31: 363–366. Crossref
Bekker-Migdisova, E.E. 1946. Contributions to the knowledge of comparative morphology of the Recent and Permian Homoptera [in Russian]. Izvestiâ Akademii nauk SSSR 6: 741–766.
Becker-Migdisova, E.E. 1959. New genus of Borisrohdendorfia (Homoptera) from Kuznetska svita of Kuznetsk Basin. Paleontologičeskij žurnal 1959 (3): 138–140.
Becker-Migdisova, E.E. 1961. Order Homoptera [in Russian]. In: B.B. Rohdendorf, E.E. Becker-Migdisova, O.M. Martynova, and A.G. Šarov (eds.), Paleozojskie nasekomyje Kuznetska. Trudy Paleontologičeskogo Instituta Akademii Nauk SSSR 85: 286–393.
Bourgoin, T., Wang, R.-R., Asche, M., Hoch, H., Soulier-Perkins, A., Stroinski, A., Yap, S., and Szwedo, J. 2015. From micropterism to hyperpterism: recognition strategy and standardized homology-driven terminology of the forewing venation patterns in planthoppers (Hemiptera: Fulgoromorpha). Zoomorphology 134: 63–77. Crossref
Evans, J.W. 1963. The systematic position of the Ipsviciidae (Upper Triassic Hemiptera) and some new Upper Permian and Middle Triassic Hemiptera from Australia. Journal of the Entomological Society of Queensland 2: 17–23. Crossref
Hamilton, K.G.A. 1992. Lower Cretaceous Homoptera from the Koonwarra fossil bed in Australia with a new superfamily and synopsis of the Mesozoic Homoptera. Annals of the Entomological Society of America 85: 423–430. Crossref
Handlirsch, A. 1906–1908. Die fossilen Insekten und die Phylogenie der rezenten Formen. Ein Handbuch für Paläontologen und Zoologen. 1430 pp. W. Engelman, Leipzig. Crossref
Hong, Y.-C. 1984. Curvicubitidae fam. nov. (Lepidoptera? Insecta), from the Middle Triassic of Shaanxi [in Chinese]. Acta Paleontologica Sinica 23: 782–785.
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. 824 pp. Laurentius Salvius, Stockholm. Crossref
Martynov, A.V. 1935. About several insects from the Kolchuginsky region of the Kuznetsk Basin. Izvestiâ Akademii Nauk SSSR, Seriâ Biologičeskaâ 1935: 441–448.
Martynov, A.V. 1937. Permian fossil insects from Kargala and their relationships [in Russian]. Trudy Paleontologičeskogo Instituta Akademii Nauk SSSR 7: 8–92.
Nel, A., Prokop, J., Nel, P., Grandcolas, P., Huang, Di-ying, Roques, P., Guilbert, E., Dostál, O., and Szwedo, J. 2012. Traits and evolution of wing venation pattern in paraneopteran insects. Journal of Morphology 273: 480–506. Crossref
Pinto, I.D. and Pinto de Ornellas, L. 1981. Permian insects from Parana basin, South Brazil. 3. Hemiptera, 1. Pereboridae. Anais do Congresso Latino-Americano de Paleontologia 2: 209–219.
Prevec, R., Nel, A., Day, M.O., Muir, R.A., Matiwane, A., Kirkaldy, A.P., Moyo, S., Staniczek, A., Cariglino, B., Maseko, Z., Kom, N., Rubidge, B.S., Garrouste, R., Holland, A., and Barber-James, H.M. 2022. New South African Lagerstätte reveals middle Permian Gondwanan lakeshore ecosystem in exquisite detail. Communication Biology 5: article 1154. Crossref
Ren, D., Lu, L.-W., Guo, Z.-G., and Ji, S.-A. 1995. Faunae and Stratigraphy of Jurassic–Cretaceous in Beijing and the Adjacent Areas [in Chinese, with English summary]. i–vii + 1–222 pp. Seismic Publishing House, Beijing.
Riek, E.F. 1973. Fossil insects from the Upper Permian of Natal. Annals of the Natal Museum 21: 513–532.
Riek, E.F. 1976. New Upper Permian insects from Natal, South Africa. Annals of the Natal Museum 22: 755–789.
Rohdendorf, B.B. (ed.) 1962. Osnovy paleontologii. Členistonogie. Traheinye i Helicerovye. 560 pp. Izdatel’stvo Akademii Nauk SSSR, Moskva. [English translation in 1991. Smithsonian Institution Libraries and the National Science Foundation, Washington, D.C.]
Shcherbakov, D.Y. 1984. Systematics and phylogeny of Permian Cicadomorpha (Cimicida and Cicadina). Paleontological Journal 18: 87–97.
Shcherbakov, D.Y. 1996. Origin and evolution of the Auchenorrhyncha as shown by the fossil record. pp. 31–45. In: C.W. Schaefer (ed.). Studies in the Hemipteran Phylogeny. Proceedings of the Thomas Say Publications in Entomology. 244 pp. Entomological Society of America, Lanham.
Shcherbakov, D.E. 2000. Permian faunas of Homoptera (Hemiptera) in relation to phytogeography and the Permo-Triassic crisis. Paleontological Journal 34: S.251–S.267.
Shcherbakov, D.E. 2020. New Homoptera from the Early Cretaceous of Buryatia with notes on the insect fauna of Khasurty. Russian Entomological Journal 29: 127–138. Crossref
Shcherbakov, D.E. 2021. New Curvicubitidae and Paraknightiidae (Homoptera: Cicadomorpha) from the Triassic of Central Asia. Russian Entomological Journal 30: 129–134. Crossref
Shcherbakov, D.E. 2022. New genus of Ignotalidae (Cicadomorpha) with notes on other Homoptera from the Permian and Triassic of the Tunguska Basin. Russian Entomological Journal 31: 230–233. Crossref
Shcherbakov, D.E. and Popov, Y.A. 2002. Superorder Cimicidea Laicharting, 1781 order Hemiptera Linné, 1758. The bugs, cicadas, plantlice, scale insects, etc. (= Cimicida Laicharting, 1781, = Homoptera Leach, 1815 + Heteroptera Latreille, 1810). In: A.P. Rasnitsyn and D.L.J. Quicke (eds.), History of Insects, 143–157. Kluwer Academic Publishers, Dordrecht.
Shi, G.R. and Waterhouse, J.B. 2010. Late Palaeozoic global changes affecting high-latitude environments and biotas: an introduction. Palaeogeography, Palaeoclimatology, Palaeoecology 298: 1–16. Crossref
Smith, R.M.H., Rubidge, B.S., and Sidor, C.A. 2006. A new burnetiid (Therapsida: Biarmosuchia) from the Upper Permian of South Africa and its biogeographic implications, Journal of Vertebrate Paleontology 26: 331–343. Crossref
Srivastava, A.K. and Agnihotri, D. 2010. Dilemma of late Palaeozoic mixed floras in Gondwana. Palaeogeography, Palaeoclimatology, Palaeoecology 298: 54–69. Crossref
Szwedo, J. 2018. The unity, diversity and conformity of bugs (Hemiptera) through time. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 107: 109–128. Crossref
Szwedo, J., Bourgoin, T., and Lefebvre, F. 2004. Fossil Fulgoromorpha (Hemiptera: Fulgoromorpha) of the World. An Annotated Catalogue with Notes on Hemiptera Classification. 199 pp. Studio 1, Warsaw.
Zalessky, M.D. 1930. Sur deux représentants nouveaux des Paléohémipères du Permien de la Kama et du Perebore dans le bassin de la Petchora. Bulletin de l’Académie des Sciences de l’URSS 7: 1017–1027.
Acta Palaeontol. Pol. 68 (1): 85–93, 2023
https://doi.org/10.4202/app.01022.2022