
Early Katian, Late Ordovician, heliolitine corals from southern Kuruktag in northeastern Tarim Basin of China
YU-NONG CUI and GUANG-XU WANG
Cui, Y.-N and Wang G.-X. 2023. Early Katian, Late Ordovician, heliolitine corals from southern Kuruktag in northeastern Tarim Basin of China. Acta Palaeontologica Polonica 68 (2): 273–295.
Heliolitines are a major tabulate coral group, which experienced their early diversification in the Katian (Late Ordovician). Fossils of this group are well represented in the Kuruktag area of northeastern Tarim Basin, Northwest China, but detailed studies of corals from this area are still lacking. Here, we systematically describe early Katian heliolitines of the Tarim Block based on new material from the lower Katian Yuanbaoshan Formation of southern Kuruktag, which include the plasmoporellids Plasmoporella xinjiangensis and Plasmoporella grandis, the sibiriolitids Mongoliolites obliterans and Mongoliolites sp., the protoheliolitid Wormsipora sp., the proporid Acdalopora sokolovi, the pseudoplasmoporid Navoites irregularis, and the heliolitid Apekinella zeravshanica. A faunal comparison indicates that the biogeographic connections of Tarim Block are closest to Chu-Ili and South Tienshan, but relatively weaker with Qilian and North China.
Key words: Tabulata, Heliolitida, biogeography, Katian, Ordovician, Tarim, China.
Yu Nong Cui [yncui@nigpas.ac.cn; ORCID: https://orcid.org/0000-0002-7566-7411 ] and Guang Xu Wang [gxwang@nigpas.ac.cn; ORCID: https://orcid.org/0000-0001-7946-5537 ], State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, 39 East Beijing Road, Nanjing 210008, China; University of Chinese Academy of Sciences, Beijing 100049, China.
Received 15 September 2022, accepted 13 February 2023, available online 14 April 2023.
Copyright © 2023 Y.-N Cui and G.X. Wang This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
As a major clade of tabulate corals, heliolitines first appeared in the Sandbian (early Late Ordovician) and diversified significantly later in the Katian (Scrutton 1984; Webby et al. 2004; Cui et al. 2022). However, as noted by Cui and Wang (2021), Late Ordovician heliolitines have not been well characterized in a global scale, with those from only a few regions relatively well documented, and this hinders our understanding of their early evolution and biogeographic pattern during the Late Ordovician.
Abundant Katian heliolitine corals are known from the Tarim Basin, northwestern China, with main fossil localities from the Kuruktag area (Fig. 1). However, largely due to the remoteness and inaccessibility, comprehensive studies of these corals of the area are still lacking. Previously, they were documented by Yu (1960), Wang (1981), and Kong and Song (1990), with a total of six genera and 17 species recognized. These taxa, however, are not properly described and illustrated, and pending revision.
Cui and Wang (2021) recently revised the heliolitine fauna of late Katian age from northern Kuruktag. Here, a systematic study of early Katian heliolitines from the southern part of the area is presented based on new material, which is followed by a discussion concerning faunal affinities of the block.
Institutional abbreviations.—NIGP, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, China.
Other abbreviations.—TaD, tabularium diameter; Ta5, number of tabulae per 5 mm; WT, wall thickness.
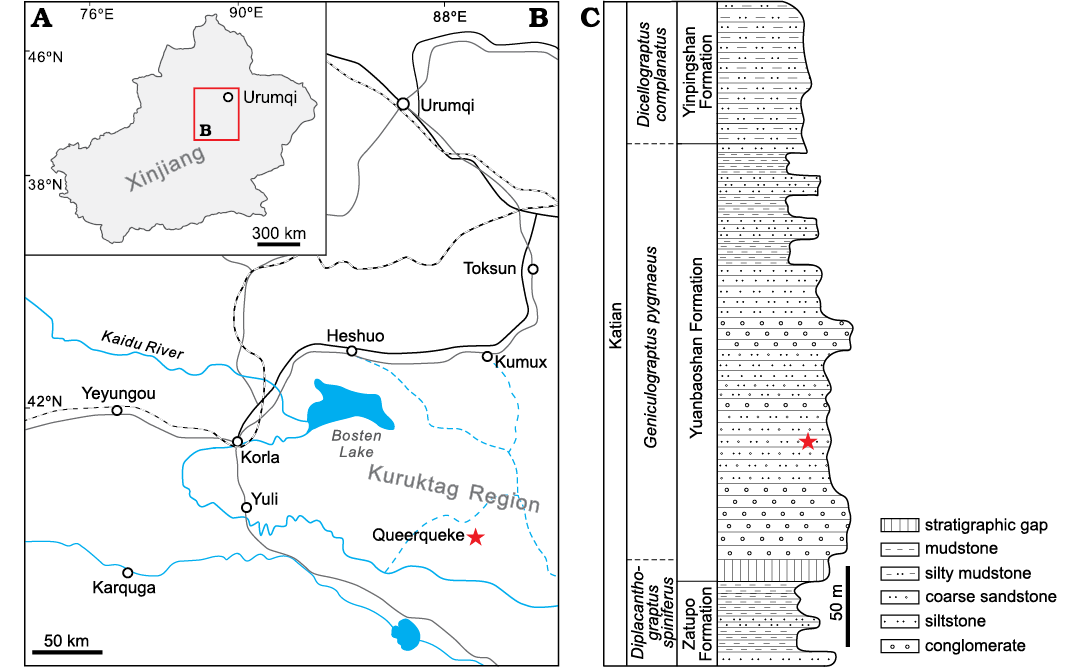
Fig. 1. Map of the Xinjiang Uygur Autonomous Region (A) and southern Kuruktag, northeastern Tarim Basin (B) in Northwest China showing location of the studied area (star). C. Stratigraphic column of the studied section of Queerqueke Montain, southern Kuruktag with position of the studied material (star)
Geological setting
Stratigraphy and material.—Kuruktag Mountains, located on the northeastern margin of Tarim Basin, is one of the major areas with extensive Ordovician outcrops in the Tarim Block, and these rocks are dominated by siliciclastics in the southern part of the area, but by carbonate rocks in the north (Norin 1937; Lai 1982; Zhong and Hao 1990, Zhou et al. 1990; Zhang and Munnecke 2016). Among the fossils abundant in these strata are graptolites (Chen et al. 1990, 2012), brachiopods (Zhan et al. 2014; Zhang et al. 2017; Sproat and Zhan 2018, 2019), and corals (Yu 1960; Kong and Song 1990; Cui and Wang 2021).
In southern Kuruktag, the Upper Ordovician succession includes the Zatupo, Yuanbaoshan, and Yinpingshan formations (Zhong and Hao 1990). The main lithology of the Yuanbaoshan Formation that yields the present coral material is thick-bedded coarse sandstone intercalated with thin-bedded siltstone and mudstone (Fig. 1). A complete Upper Ordovician graptolite succession recognized from this area comprises the Diplacanthograptus spiniferus, Genuculograptus pygmaeus, and Dicellograptus complanatus biozones from the Zatupo, Yuanbaoshan, and lower Yinpingshan formations, respectively (Zhong and Hao 1990; Chen et al. 2012). The Yuanbaoshan Formation was therefore considered to be equivalent to the upper Pagoda Formation (early Katian) of South China. In addition, Nankinolithus sp., a trilobite characteristic of the upper Katian Linhsiang Formation, was found in the overlying Yinpingshan Formation (Lin et al. 1990), supporting such an age assignment of the Yuanbaoshan Formation.
The present material includes a total of 312 specimens, all from the middle part of the Yuanbaoshan Formation of the Queerqueke Mountain. Of them, 73 well-preserved specimens with 162 thin sections are prepared for systematic studies.
Faunal composition and biogeography.—Heliolitine corals of early Katian age are known from both southern and northern parts of the Kuruktag area. In northern Kuruktag, Wang (1981) first documented Protoheliolites orientalis Yu, 1960, from the Uligezitag Formation (lower Katian), which is transferred herein to Wormsipora. Kong and Song (1990) subsequently described Plasmoporella xinjiangensis Lin and Wang in Wang, 1981, and Protoheliolites tenuis Kong and Song, 1990, from the same horizon.
In southern Kuruktag, Plasmoporella subtilis Yu, 1960, and Taeniolites compacta Kong and Song, 1990, were previously described by Kong and Song (1990) from the Yuanbaoshan Formation. The present systematic study further identifies eight species of six genera, including the plasmoporellids Plasmoporella xinjiangensis Lin and Wang in Wang, 1981, and P. grandis Bondarenko, 1958, the sibiriolitids Mongoliolites obliterans (Bondarenko 1958) and Mongoliolites sp., the protoheliolitid Wormsipora sp., the proporid Acdalopora sokolovi Bondarenko, 1958, the pseudoplasmoporid Navoites irregularis (Lin and Chow 1977), and the heliolitid Apekinella zeravshanica Ospanova, 2012a.
Previously, some authors considered that Tarim was biogeographically closely related to South China and Qilian based on evidence from Late Ordovician corals as a whole (e.g., Zhong and Hao 1990; Han et al. 2017). Herein, we make a comparison of the Kuruktag fauna with those of similar early Katian age from Chu-Ili (southern Kazakhstan), South Tienshan, Qilian, North China, South China, and Australia. The results indicate that Tarim has closest biogeographic links to Chu-Ili and South Tienshan (Fig. 2), which is consistent with the biogeographic analyses of the late Katian heliolitine corals from northern Kuruktag of the same block (Cui and Wang 2021).
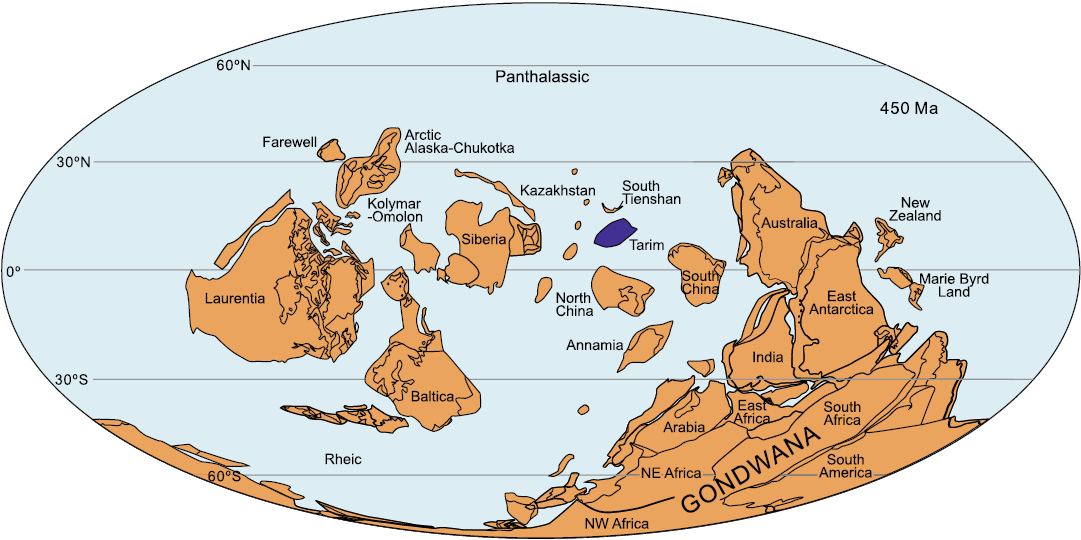
Fig. 2. Distribution of major tectonic units in the early Katian (Late Ordovician), with the Tarim Block (marked in purple). Modified from Torsvik and Cocks (2013).
Early Katian heliolitine corals from the Dulankara Formation of Chu-Ili is similarly typified by the abundance of species of Plasmoporella, accompanied by minor amount of Acdalopora (Sokolov 1955; Keller 1956; Bondarenko 1958, 1992). Among them, Plasmoporella obliterans (now revised as Mongoliolites obliterans, see discussion below) and Acdalopora sokolovi are only known in Tarim outside Chu-Ili, supporting the close biogeographic relationship between the two terranes.
Heliolitines in South Tienshan mainly occur in the Chashmankalon and Archalyk members of the Zeravshan-Gissar region, the uppermost Altyaul Formation of the Zirabulak Mountains, and the Oidynbulak Formation of the Kuldzhuktau Mountains, all of late Katian age (Kim et al. 2007; Ghobadi Pour et al. 2023), with major components including species of Plasmoporella, Navoites, Apekinella, Acdalina, and Kimilites, and Propora (Kim 1966; Dzyubo 1972; Leleshus 1975; Kim and Apekin 1978; Sladkovskaja 1987; Ospanova 1984, 1991, 2014, 2017). From the species of the above fauna, Apekinella zeravshanica is also identified from the current material, representing the only record outside South Tienshan, and Plasmoporella convexotabulata is comparable with Plamoporella maxima from the Hadabulaktag Formation (late Katian in age) of the northern Kuruktag, Tarim, indicating a close faunal link between South Tienshan and Tarim.
Our comparison also suggests a relatively close biogeographic connection of Tarim with Qilian and North China, as evidenced by some faunal similarity at species level. Heliolitines of the same age documented from the Koumenzi Formation of Qilian contain abundant Plasmoporella, which is accompanied by minor Wormsipora, Propora, and Khangailites (Yu 1960, 1962; Deng and Li 1979; Cui and Wang 2021), with Wormsipora sinkiangensis and Khangailites sinkiangensis in common with the heliolitine fauna from the slightly higher Hadabulaktag Formation (Dicellograptus complanatus Biozone) of northern Kuruktag in Tarim. The heliolitines from the lower Katian Beiguoshan and Taoqupo formations of North China include species of Plasmaporella, Wormsipora, Heliolites, and Khangailites (Li and Lin 1982; Deng 1984; Lin 1985; Jiang et al. 2013; Cui and Wang 2021). In this fauna, Khangailites sinkiangensis and Plasmoporella maxima are also known from the upper Katian Hadabulaktag Formation of northern Kuruktag (Cui and Wang 2021).
In contrast, the faunal connection of Tarim with Australia and South China seems weak. Early Katian heliolitines of Australia are characterized by the presence of species of Plasmoporella, Navoites, and Heliolites (Wang et al. 2021). Species of the former two genera are present in Tarim, but without species in common with Australia. Contemporary heliolitines from the upper part of the Yenwashan Formation in South China are solely represented by Protoheliolites heliolitiformis Lin and Chow, 1980; this species does not occur in Tarim Block, adding support for their weak faunal link. However, the limited heliolitine data of this age in South China render this conclusion less reliable.
Evidence from well-documented brachiopods and trilobites from Tarim Block also supports our view. The brachiopod fauna found from the Lianglitag Formation (lower Katian) of the Bachu area in northwestern Tarim Basin is characterized by common occurrence of triplesiides, camerelloids and atrypides, and the rarity of orthides and strophomenides, indicating a close biogeographic connection with the Chu-Ili, North China and Qilian (Rong et al. 2017). The trilobites from the slightly older Yingan Formation of the Kalpin area in northwestern Tarim Basin include species of Lisogorites, Taklamakania, and Ampyxinella (Zhou et al. 1995, 2014). These fossils are also known in Kazakhstan, South China and North China, showing consistency with our coral data.
Systematic palaeontology
Morphological terminology adopted in this work largely follows Hill (1981) and Dixon and Jell (2012), and biometric methods of Young and Elias (1995) are used.
Class Anthozoa Ehrenberg, 1834
Subclass Tabulata Milne-Edwards and Haime, 1850
Order Heliolitida Frech, 1897
Superfamily Proporicae Sokolov, 1949
Family Plasmoporellidae Kiaer, 1903
Genus Plasmoporella Kiaer, 1899
Type species: Plasmoporella convexotabulata forma typica Kiaer, 1899; Upper Ordovician, Norway.
Remarks.—We follow the diagnosis of Plasmoporella suggested by Hill (1981), but we emphasize the variable septal development in this genus. Its comparison with other similar genera was recently discussed by Cui and Wang (2022).
Plasmoporella xinjiangensis Lin and Wang in Wang, 1981
Figs. 3, 4, 5.
1981 Plasmoporella xinjiangensis sp. nov.; Lin and Wang in Wang 1981: 70, pl. 36: 2.
1986 Plasmoporella xinjiangensis Lin and Wang in Wang, 1981; Lin and Wang 1986: 52, pl. 8: 3a, 3b.
1990 Plasmoporella xinjiangensis Lin and Wang in Wang, 1981; Kong and Song 1990: 157, pl. 28: 2.
1990 Plasmoporella yuanbaoshanensis sp. nov.; Kong and Song 1990: 158, pl. 27: 3.
2015 Plasmoporella xinjiangensis Lin and Wang in Wang, 1981; Yang et al. 2015: 87, pl. 74: 3, 4.
Material.—Forty-five sectioned specimens, NIGP 200945–200989 from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, China.
Emended diagnosis.—Plasmoporella with TaDs 1.60–2.35 mm. Trabecular septa long some extending to corallite axis. Tabulae incomplete. Coenenchyme consisting of dissepiments, with vertical trabeculae present.
Description.—Eight almost complete coralla are measured, of low domical to high bulbous form, with the largest 80.24 mm wide and 33.96 mm high, and the smallest 44.12 mm wide and 16.72 mm high. Corallites rounded, 1.82–2.35 mm (average 2.14 mm) in diameter. Corallites outlined by 12 thick well-developed trabeculae, with walls of tabularia defined largely by downturned margins of coenenchymal dissepiments. Septa generally lachrymiform in transverse section, with a maximum width of 0.30–0.65 mm (average 0.42 mm). The trabeculae may extend into corallites and coenenchyme, some extending to the corallite axis. Septa spinose in axial margin and appearing as numerous black dots in transverse section. Tabulae incomplete, slightly convex, Ta5 23–26. Corallite centers 2.10–3.20 mm apart (average 2.58 mm). Intercorallite spaces filled with coenenchyme of convex dissepiments. Dissepiments generally 0.16–0.49 mm (average 0.31 mm), 24–28 in 5 mm. A large number of trabeculae scattered in the coenenchyme, often passing through a few dissepiments.
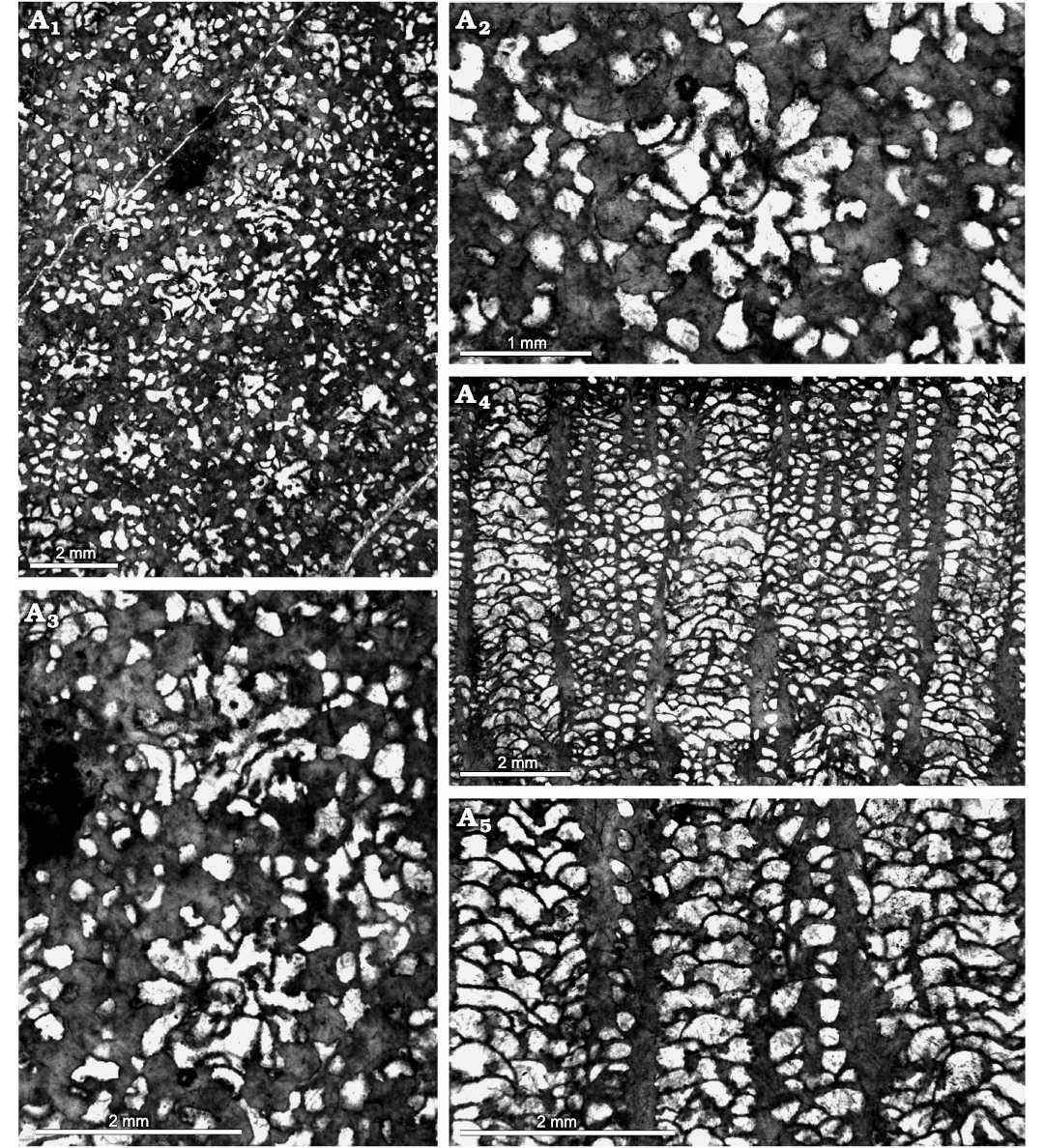
Fig. 3. Heliolitine tabulate coral Plasmoporella xinjiangensis Lin and Wang in Wang, 1981, from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, Northwest China. NIGP 200961. A1–A3, transverse sections. A4, A5, longitudinal sections.
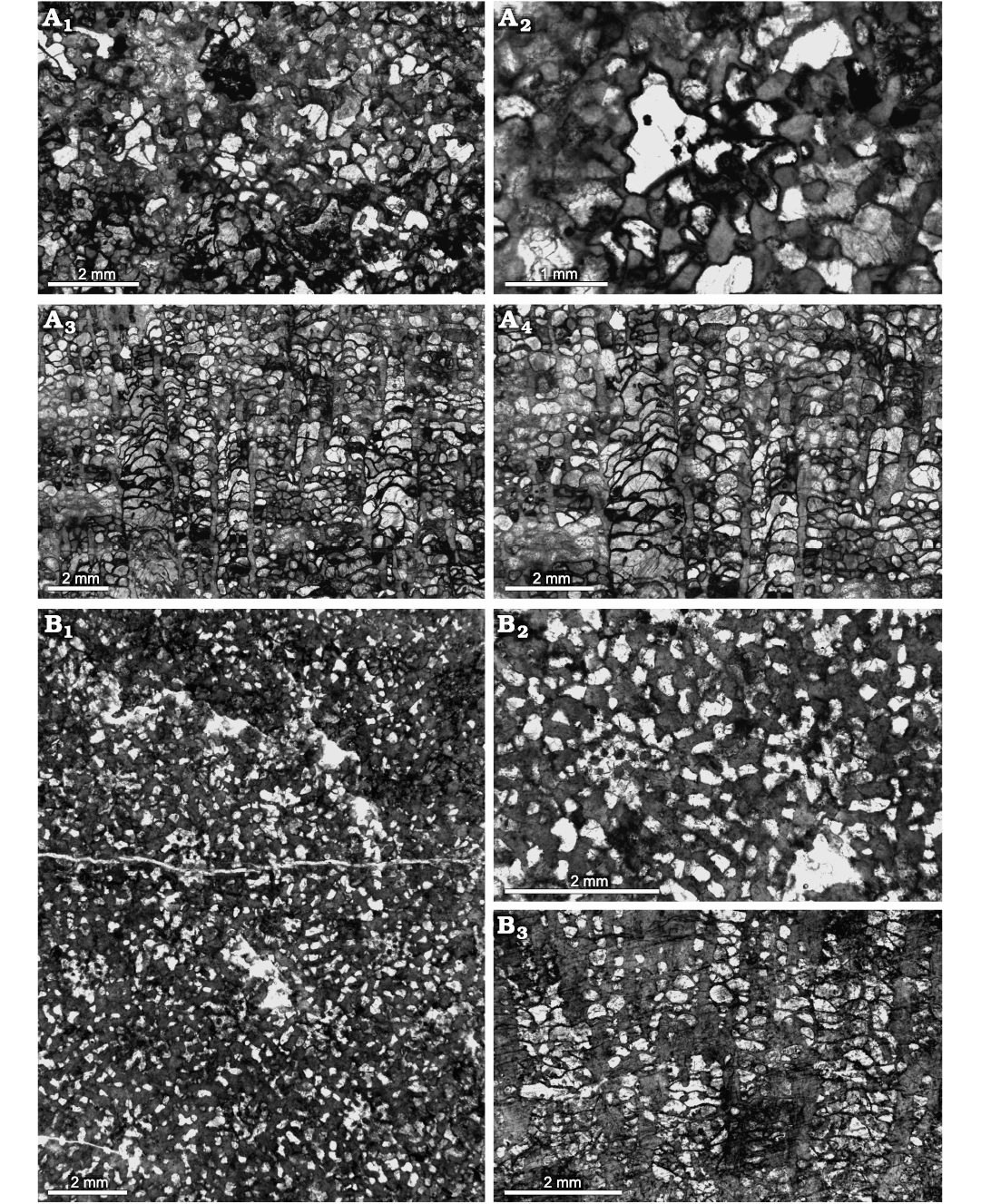
Fig. 4. Heliolitine tabulate coral Plasmoporella xinjiangensis Lin and Wang in Wang, 1981, from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, Northwest China. A. NIGP 200945. A1, A2, transverse sections. A3, A4, longitudinal sections. B. NIGP 200973. B1, B2, transverse sections. B3 longitudinal section.
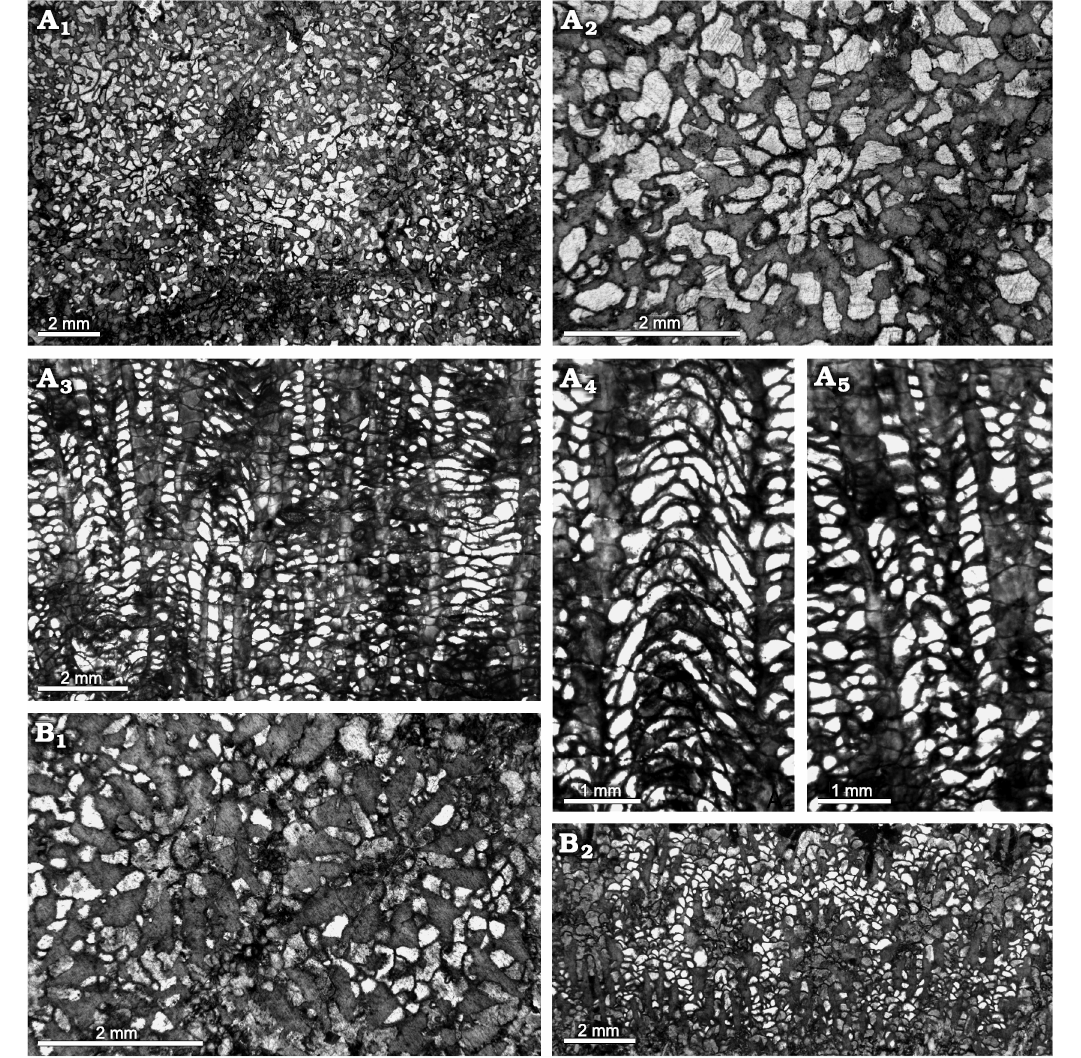
Fig. 5. Heliolitine tabulate coral Plasmoporella xinjiangensis Lin and Wang in Wang, 1981, from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, Northwest China. A. NIGP 200955. A1, A2, transverse sections. A3–A5, longitudinal sections. B. NIGP 200946. B1, transverse section. B2, longitudinal section.
Remarks.—Our specimens show closest resemblance to the holotype of Plasmoporella xinjiangensis Lin and Wang in Wang, 1981, documented from the Upper Ordovician Bulongor Formation of Hobuksar in northwestern Xinjiang, differing only in having longer septa. For this reason, we assign these specimens to P. xinjiangensis.
The specimens described as P. xinjiangensis by Lin and Wang (1986) from the same level and locality as the holotype and by Kong and Song (1990) from the Uligezitag Formation (lower Katian) of northern Kuruktag in northeastern Xinjiang are almost identical to the type material of the species, with the Uligezitag material having slightly smaller corallites (1.6–2.0 mm in diameter). Yang et al. (2015) also recorded P. xinjiangensis from the Sandbian Lianglitag Formation of Well 70 of central Tarim, the tabulae of which are more complete. Plasmoporella yuanbaoshanensis Kong and Song, 1990, from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag is attributable to P. xinjiangensis based on its comparable corallite size, which only differs in having weakly developed septa.
The diagnosis of P. xinjiangensis is revised herein based on all the material now assigned to this species, with major revisions including an extension of the range of corallite size (1.60–2.35 mm) and the variation of septal length. Its comparison with similar species is shown in Table 1 and discussed below. Plasmoporella multitrabeculata Lin and Chow, 1977, and Plasmoporella cystosa Lin and Chow, 1977, both from the middle Sanqushan Formation (upper Katian) of Jiangxi, southeastern China, can be distinguished from P. xinjiangensis by their much smaller corallites (1.3–1.6 mm in diameter) and larger corallites (2.1–2.5 mm in diameter), respectively. Plasmoporella cystosa further differs in possessing much shorter septa.
Table 1. Comparison of Plasmoporella species discussed in the present paper. Abbreviations: CoD, corallite diameter; TaD, tabularium diameter; Ta5, number of tabulae per 5 mm. Species names in bold refer to those described in this paper.
| |
Corallite size (mm) |
Tabulae |
Septa |
|
Plasmoporella xinjiangensis Lin and Wang in Wang, 1981 |
TaD = 1.80–2.35 |
incomplete, Ta5 = 23–26 |
extending to the corallite axis |
|
Plasmoporella multitrabeculata Lin and Chow, 1977 |
CoD = 1.3–1.6 |
incomplete |
long |
|
Plasmoporella cystosa Lin and Chow, 1977 |
CoD = 2.1–2.5 |
incomplete and irregular |
long |
|
Plasmoporella grandis Bondarenko, 1958 |
TaD = 3.0–4.5 |
mostly complete, Ta5 = 13–17 |
moderately long and fork-shaped |
|
Plasmoporella bifida Bondarenko, 1958 |
CoD = 2.8–3.0 |
incomplete, Ta5 = 11–13 |
moderately long and fork-shaped |
|
Plasmoporella gigantea Lin and Wang, 1986 |
CoD = 4.2–5.0 |
mostly complete, Ta5 = 6–10 |
short |
|
Plasmoporella qiakertensis Lin and Wang, 1985 |
CoD = 3.0–3.5 |
complete, Ta5 = 10–12 |
short and fork-shaped |
Stratigraphic and geographic range.—The Yuanbaoshan Formation (lower Katian) of southern Kuruktag, northeastern Tarim, China; the Bulongor Formation (Katian) of Hobuksar, northwestern Xinjiang, China; the Uligezitag Formation (lower Katian) of northern Kuruktag, northeastern Tarim, China; the Lianglitag Formation (lower Katian) of Well 70 of central Tarim, Xinjiang, China.
Plasmoporella grandis Bondarenko, 1958
Figs. 6, 7.
1958 Plasmoporella grandis sp. nov.; Bondarenko 1958: 210, pl. 4: 3, 4.
Material.—Ten sectioned specimens, NIGP 200990–200999 from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, China.
Emended diagnosis.—Species of Plasmoporella with TaDs 3.00–4.50 mm. Septa long and thin, Tabulae complete to incomplete, strongly convex axially.
Description.—Ten complete coralla are measured, which are mostly low-domed, and sometimes bulbous or high-domed, with the largest 88.57 mm wide and 37.30 mm high, and the smallest 30.70 mm wide and 22.80 mm high. Corallites rounded and slightly stellated, with tabularium diameters varying from 3.02–4.11 mm (average 3.65 mm). Corallites walls outlined by edges of coenenchymal dissepiments. Septa of trabecular type, sometimes bifurcated, 12 in number, normally extending no more than one-eighth the TaD into tabularia, with maximum widths ranging from 0.53 to 0.87 mm (average 0.65 mm). Septa inclined upwardly and shown as dark dots in transverse section. Tabulae mostly complete, some strongly convex axially, closely spaced, with Ta5 13–17. Corallites separated by narrow cenenchyme, with their centers 4.26–4.35 mm apart (average 4.31 mm). Coenenchyme consisting of dissepiments, with vertical trabeculae observed. Dissepiments generally 0.68–1.16 mm wide (average 0.87 mm). Frequency of dissepiments similar throughout at 22–26 per 5 mm.
Remarks.—The present material is conspecific with Plasmoporella grandis Bondarenko, 1958, from the Upper Ordovician Dulankara Formation of the Chu-Ili Terrane in southern Kazakhstan. Their major difference lies in the more complete tabulae of the former. On this basis, we include this feature in the diagnosis of this species. This species was assigned to Granulina by Bondarenko (1992), which was recently regarded as a synonym of Plasmoporella by Cui et al. (2022).
A comparison of Plasmoporella grandis with similar species is shown in Table 1 and discussed below. In the trabecular development P. grandis strongly resembles Plasmoporella bifida Bondarenko, 1958, also from the Dulankara Formation of southern Kazakhstan. The major differences are smaller corallites (2.8–3.0 mm in diameter) and more incomplete tabulae of the latter. Plasmoporella gigantea Lin and Wang, 1986, from the Upper Ordovician Bulongor Formation of Hobuksar in northwestern Xinjiang differs from P. grandis in having much larger corallites (4.2–5.0 mm in diameter) and sparser tabulae (6–7 per 5 mm). Plasmoporella qiakertensis Lin and Wang, 1985, from the Upper Ordovician Gabosar Formation of Gabosar in northern East Junggar, Xinjiang, is distinguished by smaller corallite size (3.0–3.5 mm in diameter) and more complete tabulae.
Stratigraphic and geographic range.—The Yuanbaoshan Formation (lower Katian) of southern Kuruktag, northeastern Xinjiang, China; the Dulankara Formation (lower Katian) of the Chu-Ili Terrane, southern Kazakhstan.
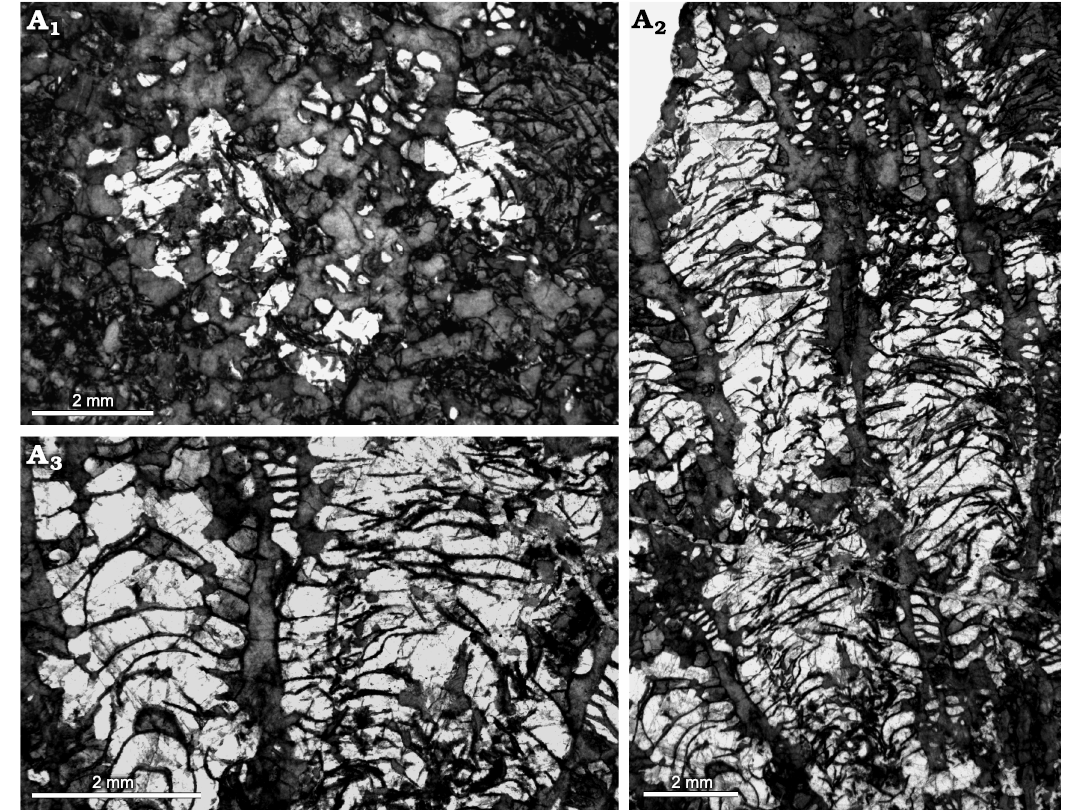
Fig. 6. Heliolitine tabulate coral Plasmoporella grandis Bondarenko, 1958, from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, Northwest China. NIGP 200990. A1, transverse section. A2, A3, longitudinal sections.
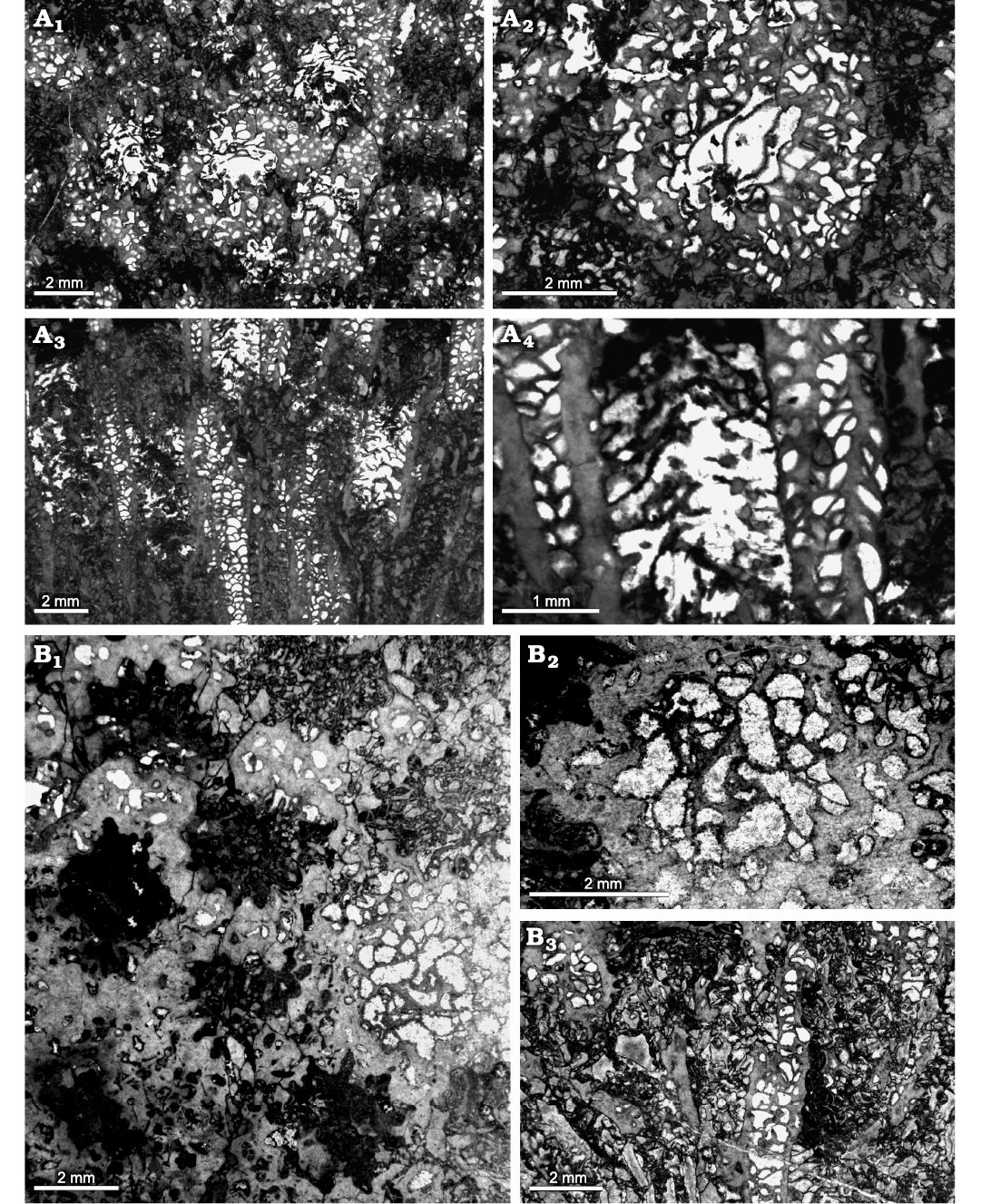
Fig. 7. Heliolitine tabulate coral Plasmoporella grandis Bondarenko, 1958, from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, Northwest China. A. NIGP 200991. A1, A2, transverse sections. A3, A4, longitudinal sections. B. NIGP 200993. B1, B2, transverse sections. B3, longitudinal section.
Family Sibiriolitidae Lin in Lin and Chow, 1977
Genus Mongoliolites Bondarenko and Minzhin, 1977
Type species: Mongoliolites paradoxides Bondarenko and Minzhin; 1977, Katian (Upper Ordovician), Khanay Range, central Mongolia.
Mongoliolites obliterans (Bondarenko, 1958)
Fig. 8.
1958 Plasmoporella obliterans sp. nov.; Bondarenko 1958: 214, 215, pl. 5: 5–7; pl. 6: 4, 5.
Material.—Three sectioned specimens, NIGP 201000–201002 from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, China.
Emended diagnosis.—Mongoliolites species with TaDs 0.75–1.01 mm. Corallite walls formed by the fusion of peripheral parts of septa. Septa wedge-shaped, short. Tabulae mostly complete, sparsely spaced.
Description.—Only one single complete corallum measured, of low domical form, 33.79 mm wide and 22.72 mm high. Corallites rounded with clear outlines, TaDs 0.75–1.01 mm (average 0.92 mm). Corallite walls consisting of thickened trabeculae. Septa triangular to lachrymiform in shape, 12 in number, 0.13–0.19 mm long (average 0.15 mm), with maximum widths ranging from 0.10 to 0.19 mm, extending one-fifth of the TaD. Corallite walls weakly to moderately developed. Tabulae mostly complete and sparse, generally convex, some strongly concave or curved, Ta5 12–16. Corallites commonly separated by narrow coenenchyme, with centers 1.04–1.66 mm apart (average 1.33 mm). Dissepiments flat or slightly convex, generally 0.21–0.44 mm wide (average 0.35 mm). The number of dissepiments in 5 mm 20–22.
Remarks.—Our material is clearly conspecific with the holotype of Plasmoporella obliterans Bondarenko, 1958, from the Upper Ordovician Dulankara Formation of the Chu-Ili Terrane in southern Kazakhstan, except that the former has slightly smaller (0.75–1.01 mm) corallite size. We therefore extend the range of corallite size in the revised diagnosis of this species. This species was originally assigned by Bondarenko (1958) to Plasmoporella on the basis of the development of 12 trabeculae, but later given by her (Bondarenko 1992) an uncertain taxonomic position. In this paper, we attribute this species to Mongoliolites based on the presence of corallite wall.
A comparison of Mongoliolites obliterans with similar species is shown in Table 2 and discussed below. Mongoliolites paradoxides Bondarenko and Minzhin, 1977, from central Mongolia differs from M. obliterans in having larger corallites (1.0–1.5 mm in diameter), more complete tabulae, and less developed septa. Mongoliolites sp. described below is separated by larger corallite size (1.51–1.67 mm in diameter) and shorter septa.
Stratigraphic and geographic range.—The Yuanbaoshan Formation (lower Katian) of southern Kuruktag, northeastern Tarim, China; the Dulankara Formation (lower Katian) of the Chu-Ili Terrane, southern Kazakhstan.

Fig. 8. Heliolitine tabulate coral Mongoliolites obliterans (Bondarenko, 1958), from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, Northwest China. A. NIGP 201001. A1, A2, transverse sections. A3, A4, longitudinal sections. B. NIGP 201002. B1, B2, transverse sections. B3, B4, longitudinal sections.
Table 2. Comparison of Mongoliolites species discussed in the present paper. Abbreviations: CoD, corallite diameter; TaD, tabularium diameter; Ta5, number of tabulae per 5 mm. Species names in bold refer to those described in this paper.
| |
Corallite size (mm) |
Tabulae |
Septa |
Coenenchyme |
|
Mongoliolites obliterans (Bondarenko, 1958) |
TaD = 0.75–1.01 |
complete and sparse, Ta5 = 12–16 |
0.13–0.19 mm |
moderate developed |
|
Mongoliolites paradoxides Bondarenko and Minzhin, 1977 |
CoD = 1.0–1.5 |
complete |
short |
weakly developed |
|
Mongoliolites sp. |
TaD = 1.51–1.67 |
mostly complete, Ta5 = 8–10 |
short |
moderate developed |
|
Mongoliolites sp. |
TaD = 1.51–1.67 |
mostly complete, Ta5 = 8–10 |
0.11–0.15 mm |
moderate developed |
|
Mongoliolites paradoxides Bondarenko and Minzhin, 1977 |
CoD = 1.0–1.5 |
complete |
short |
weakly developed |
|
Mongoliolites giganteus White and Yang, 2004 |
CoD = 3.0–3.8 |
complete and sparse, Ta5 = 2–6 |
short |
weakly developed |
|
Mongoliolites contiguus (Hall, 1975) |
TaD = 1.3–1.7 |
complete , Ta5 = 6–14 |
well developed |
weakly developed |
Mongoliolites sp.
Fig. 9.
Material.—One sectioned specimen, NIGP 201003, from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, China.
Description.—One single complete corallum measured, of low domical form, 21.16 mm wide and 7.83 mm high. Corallites rounded or sub-rounded, TaDs 1.51–1.67 mm (average 1.56 mm). Corallite walls thick, straight to weakly wavy. Septa short, 0.11–0.15 mm in length, arranged in 12 rows in later growth stages. Tabulae complete, flat, slightly sagging or convex, with some incomplete, Ta5 8–10. Corallites mostly in contact, or separated by dissepiments, with centers 1.75–2.13 mm apart (average 1.88 mm). Coenenchyme less developed. Dissepiments generally flat or slightly convex, average 14–16 per 5 mm.
Remarks.—The present material has better developed coenenchyme compared with other described Mongoliolites species, and probably represents a new species. However, it is left in open nomenclature due to limited material.
A comparison of Mongoliolites sp. with similar species is shown in Table 2 and discussed below. Mongoliolites paradoxides Bondarenko and Minzhin, 1977, from the Upper Ordovician of central Mongolia is generally similar to Mongoliolites sp. discussed herein, but differs in having smaller corallite size (1.0–1.5 mm in diameter) and less developed coenenchyme. Mongoliolites? giganteus recorded by White and Yang (2004) from the Stile End Formation (upper Katian) of the Stockdale Farm, Longsleddale, Lake District, England differs in larger corallite size (3.0–3.8 mm in diameter), sparser tabulae (2–6 per 5 mm) and less developed coenenchyme. Wang et al. (2021) described M. contiguus (Hall 1975) from the “Trelawney Beds” of the New England Region in northeastern New South Wales, which differs in possessing much less developed coenenchyme and more septa.
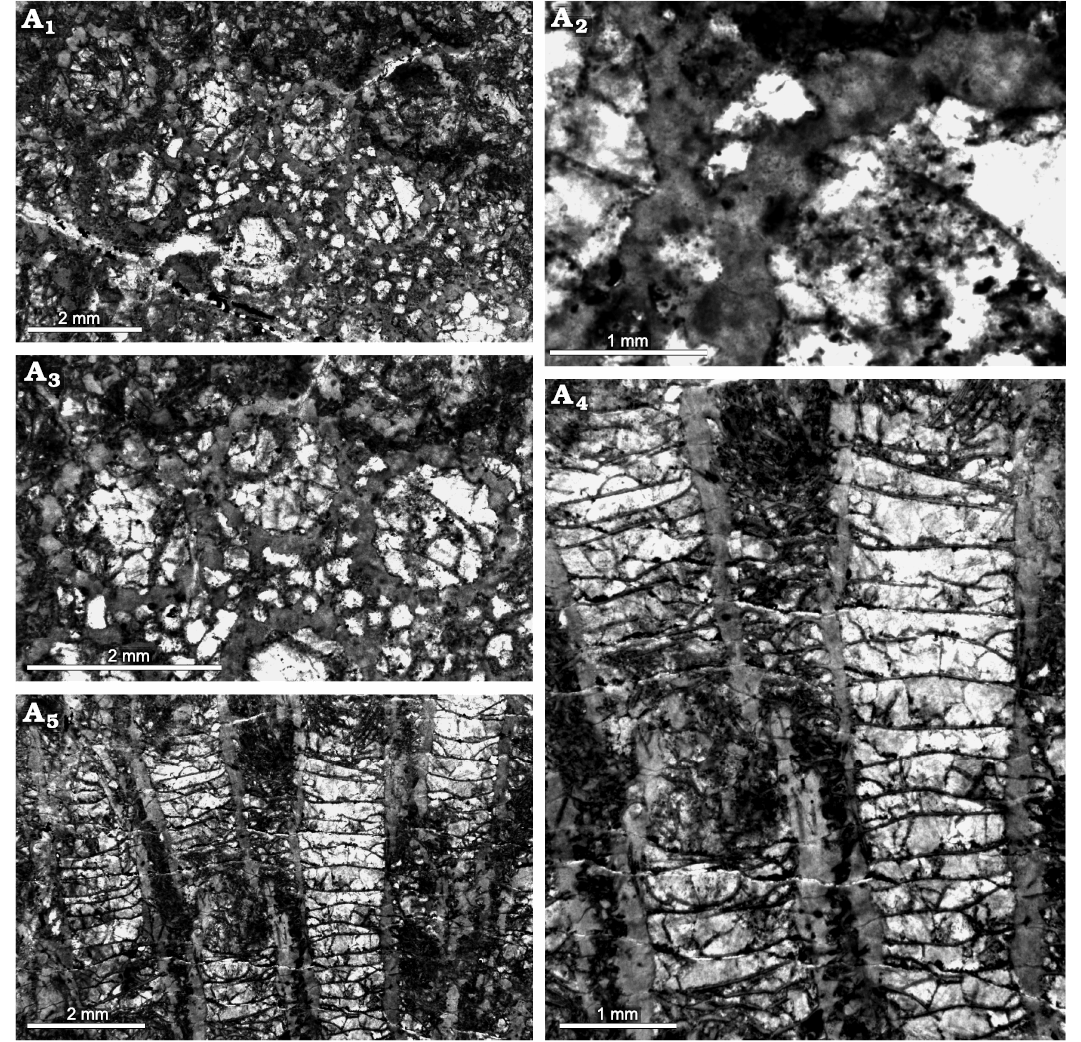
Fig. 9. Heliolitine tabulate coral Mongoliolites sp. from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, Northwest China. NIGP 201003. A1–A3, transverse sections. A4, A5, longitudinal sections.
Family Proheliolitidae Kiaer, 1899
Genus Wormsipora Sokolov, 1955
Type species: Heliolites hirsutus Lindström, 1899, Upper Ordovician, Sweden.
Remarks.—Our concept of Wormsipora largely follows Hill (1981), with an inclusion of the presence of discontinuous tubular walls in the diagnosis. For a comparison with other similar genera, see Cui and Wang (2021).
Wormsipora sp.
Fig. 10.
Material.—One sectioned specimen, NIGP 201004, from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, China.
Description.—The corallum of low domical form, 37.07 mm wide and 14.06 mm high. Corallites slightly stellate, with TaDs 0.76–0.83 mm (average 0.81 mm). Corallite walls longitudinally plicated, 0.07–0.12 mm thick (average 0.08 mm). Septa 12 in number, each consisting of a series of discrete spines. Septal spines 0.15–0.22 mm long (average 0.18 mm), commonly extending no more than one-third the TaD. Tabulae complete, horizontal or slightly sagging or convex, Ta5 10–14. Corallites often in contact or separated by narrow coenenchyme, with their centers 0.84–1.08 mm apart (average 0.93 mm). Coenenchymal tubules weakly developed, commonly 0.17–0.22 mm in diameter (average 0.21 mm). Tubular walls discontinuous, 0.04–0.05 mm thick. Diaphragms complete, flat, slightly convex or oblique, mostly 24–26 per 5 mm.
Remarks.—The present specimen is characterized by small corallites and short septa, which distinguish it from other known Wormsipora forms, and may represent a new species. However, an open nomenclature is used due to limited material. It shows closest similarities to the type material of Wormsipora orientalis (Yu, 1960) documented from the Hadabulaktag Formation (upper Katian) of northern Kurutag, Xinjiang, but is distinguished by much smaller corallites (0.76–0.83 mm in diameter), as well as more complete tabulae and tubular walls.
A comparison of Wormsipora sp. with similar species is shown in Table 3 and discussed below. Wormsipora hirsutus (Lindström, 1899) from the Upper Ordovician of Sweden differs from Wormsipora sp. in having better developed coenenchyme and septal spines. Wormsipora hiruta minor White and Yang, 2004, from the Upper Ordovician Glyn Limestone of North Wales, is separable by larger corallite size (1.3–1.5 mm in diameter) and more concave tabulae.
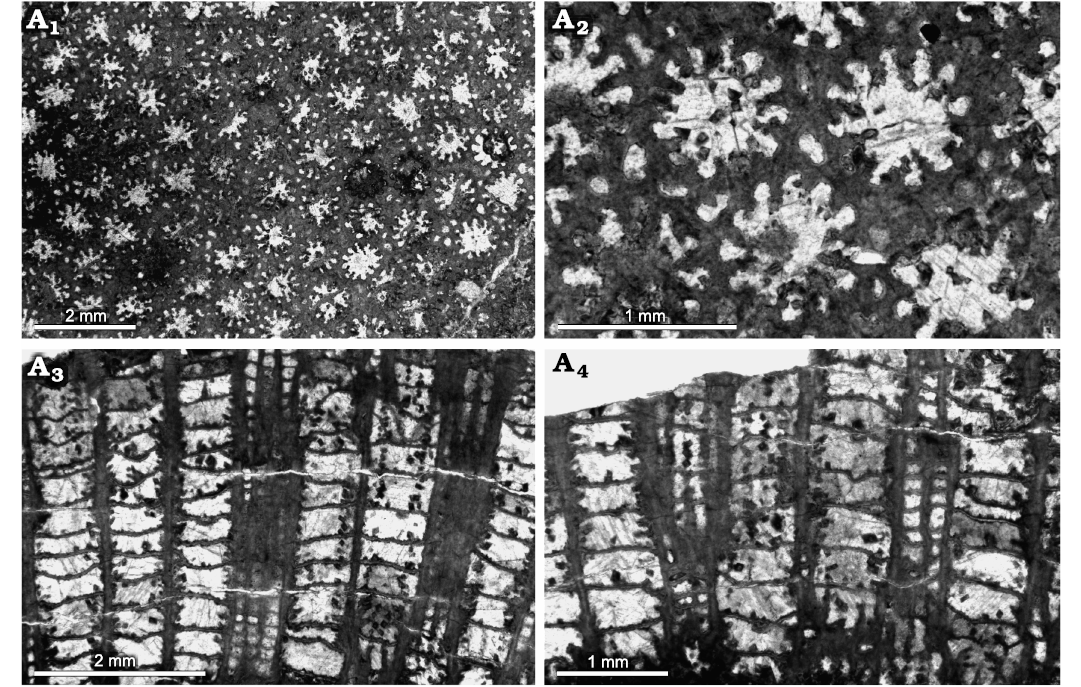
Fig. 10. Heliolitine tabulate coral Wormsipora sp. from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, Northwest China. NIGP 201004. A1, A2, transverse sections. A3, A4, longitudinal sections.
Table 3. Comparison of Wormsipora species discussed in the present paper. Abbreviations: CoD, corallite diameter; TaD, tabularium diameter; Ta5, number of tabulae per 5 mm. Species names in bold refer to those described in this paper.
| |
Corallite size (mm) |
Tabulae |
Septa |
Coenenchyme |
|
Wormsipora sp. |
TaD = 0.76–0.83 |
complete, Ta5 = 10–14 |
0.15–0.22 mm |
weakly developed |
|
Wormsipora orientalis (Yu, 1960) |
TaD = 1.24–1.56 |
complete, Ta5 = 10–18 |
moderately long |
weakly developed |
|
Wormsipora hirsutus (Lindström, 1899) |
CoD = 1.2 |
complete |
long |
developed |
|
Wormsipora hiruta minor White and Yang, 2004 |
CoD = 1.3–1.5 |
complete and concave, Ta5 = 10–12 |
long |
developed |
Family Proporidae Sokolov, 1949
Genus Acdalopora Bondarenko, 1958
Type species: Acdalopora sokolovi Bondarenko, 1958, Upper Ordovician, Chu-Ili Mountain region, southern Kazakhstan.
Remarks.—We follow the diagnosis of Acdalopora by Hill (1981). This genus was considered by Young and Noble (1990) to be synonymous with Propora Milne-Edwards and Haime, 1849, because he believed that well-developed septal plates typical of Acdalopora were within the normal variation of Propora. Given the taxonomic importance of septal plates, we consider Acdalopora as a valid genus, as suggested by some authors (e.g., Hill 1981; Bondarenko 1992).
Forms closely similar to Acdalopora include Wormsipora Sokolov, 1955, and Kimilites Sladkovskaja, 1987. Wormsipora is distinguished from Acdalopora by possessing entirely tubular coenenchyme, incomplete tubular walls, and septal spines. Kimilites differs in having better developed coenenchyme and septal spines.
Acdalopora sokolovi Bondarenko, 1958
Figs. 11, 12.
1958 Acdalopora sokolovi sp nov.; Bondarenko 1958: 216, pl. 5: 1–3.
1964 Acdalopora sokolovi Bondareko, 1958; Kovalevsky 1964: 41, pl. 2: 2.
non 1975 Acdalopora sokolovi Bondareko, 1958; Leleshus 1975: 11, 12, pl. 4: 1, 2.
non 1978 Acdalopora sokolovi Bondareko, 1958; Kim and Apekin 1978: 62, 63, pl. 6: 1.
1986 Acdalopora sokolovi Bondarenko, 1958; Lin and Wang 1986: 49, pl. 7: 4a, 4b.
?1986 Acdalopora junggarensis sp nov.; Lin and Wang 1986: 48, pl. 6: 1a, 1b.
2012b Acdalopora sokolovi Bondarenko, 1958; Ospanova 2012b: 33, pl. 7.
Material.—Eight sectioned specimens, NIGP 201005–201012 from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, China.
Emended diagnosis.—Species of Acdalopora with TaDs 0.62–1.59 mm. Corallite walls wavy. Septa long; mostly laminate, with spines developed near the axial ends. Tabulae complete, horizontal.
Description.—Five complete coralla are measured, of low domical or columnar form, with the largest 123.61 mm wide and 35.42 mm high, and the smallest 44.88 mm wide and 13.08 mm. Corallites slightly stellate, TaDs 1.18–1.59 mm (average 1.34 mm). Corallite walls complete, moderately crenulated, 0.06–0.12 mm thick (average 0.08 mm). Each corallite having 12 long septa. Septa 0.15–0.36 mm long (average 0.27 mm), tapering toward the axis; mostly laminate, with spines developed near the axis. Tabulae complete, sub-horizontal or slightly convex, Ta5 14–16. Spacing of corallites variable, often in contract or slightly separated, with their centers 1.57–2.33 mm apart (average 1.91 mm). Coenenchyme consisting of overlapping small domed dissepiments. The width of the smaller and larger dissepiments 0.10–0.21 mm (average 0.13 mm), and 0.23–0.36 mm (average 0.27 mm), respectively. Dissepiments 20–22 per 5 mm.
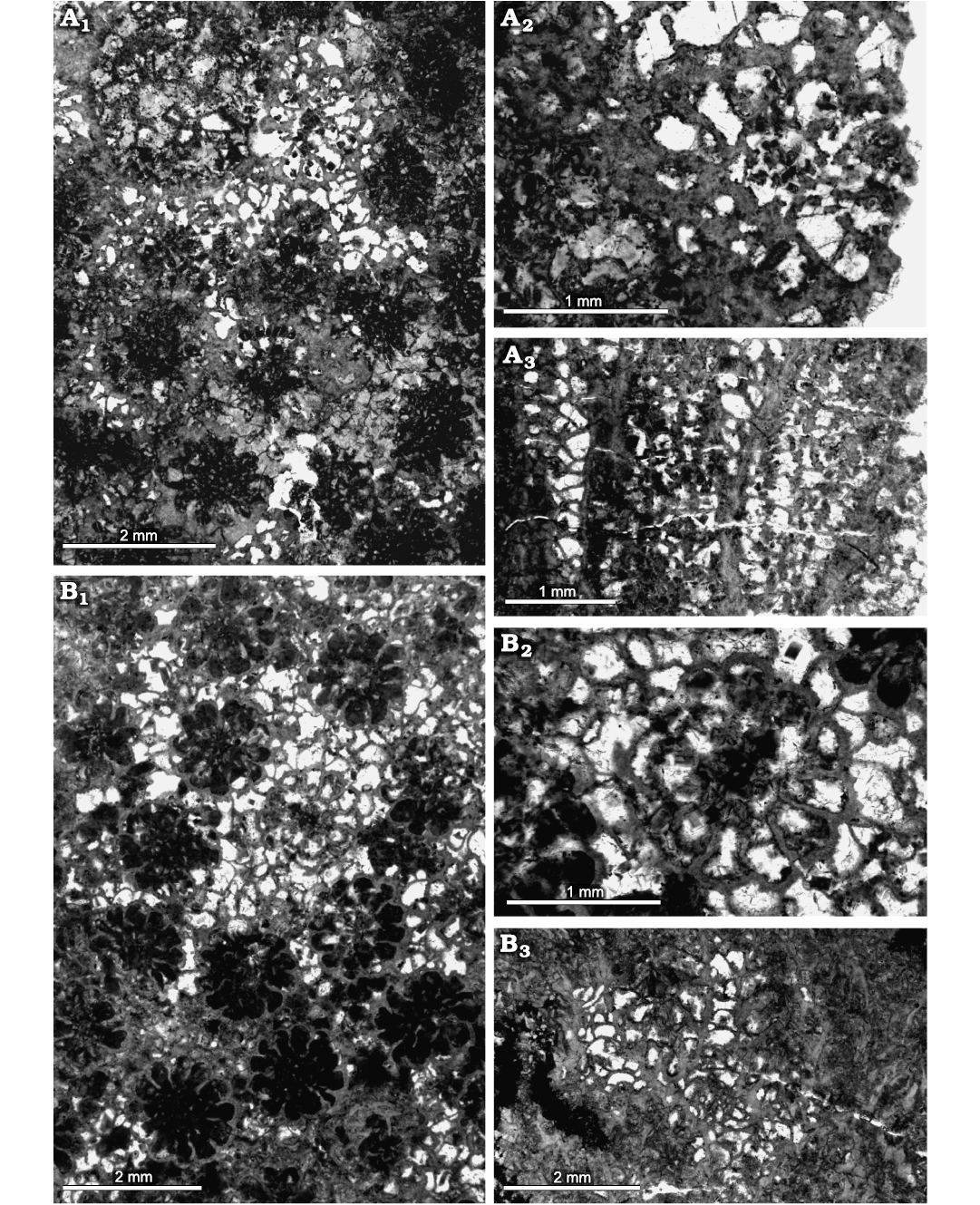
Fig. 11. Heliolitine tabulate coral Acdalopora sokolovi Bondarenko, 1958, from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, Northwest China. A. NIGP 201009. A1, A2, transverse sections. A3, longitudinal section. B. NIGP 201008. B1, B2, transverse sections. B3, longitudinal section.
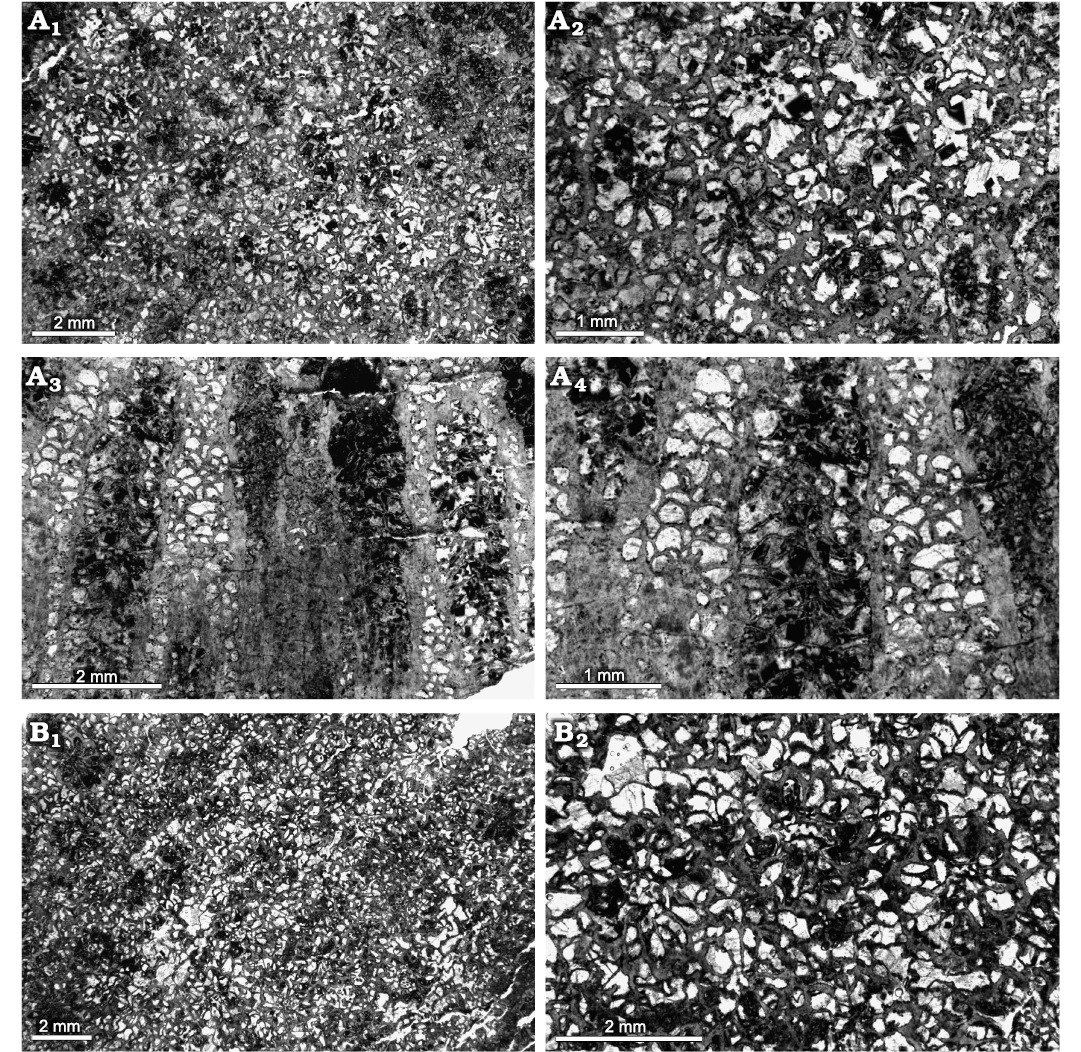
Fig. 12. Heliolitine tabulate coral Acdalopora sokolovi Bondarenko, 1958, from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, Northwest China. A. NIGP 201012. A1, A2, transverse sections. A3, A4 longitudinal sections. B. NIGP 201007. B1, B2, transverse sections.
Remarks.—These specimens are most likely conspecific with the type material of Acdalopora sokolovi Bondarenko, 1958, from the Upper Ordovician Dulankara Formation of the Chu-Ili Terrane in southern Kazakhstan, differing only in having slightly smaller corallite size (0.62–1.35 mm) of the latter. This type material was refigured by Ospanova (2012a).
The material described by Lin and Wang (1986) as Acdalopora sokolovi is from the Upper Ordovician Bulongor Formation of Hobuksar in northwestern Xinjiang. It is essentially identical to the holotype of A. sokolovi, except only in its denser tabulae (20–24 per 5 mm). Acdalopora junggarensis Lin and Wang, 1986, recorded from the same locality and horizon is a probable synonym of A. sokolovi, which differs only in having a slightly larger corallite size (1.2–1.7 mm in diameter), and more convex tabulae.
Specimens identified as A. sokolovi are also known from the Upper Ordovician of Zeravshan Ridge, Tajikistan (Leleshus 1975), and the Archalyk Member (upper Katian) of the Shakhriomon area in Uzbekistan (Kim and Apekin 1978). They have smooth and incomplete corallite walls and inconsistent septal number, and were referred to Acdalina asiatica by Ospanova (2012b), which is followed in this study.
The diagnosis of A. sokolovi is revised herein based on all the material now assigned to this species, with major revisions including an extension of the range of corallite size (0.62–1.59 mm), and major revisions including an addition of the development of septal spines. Its comparison with similar species is shown in Table 4 and discussed below. A. mira Chekhovich, 1977, from the Upper Ordovician Hondell Bed of western Tuva strongly resembles A. sokolovi in corallite size and septal development. The main differences lie in less developed septal plates and better developed coenenchyme of the former. Acdalopora qiakuertensis Lin and Wang, 1985, from the Upper Ordovician Gabosar Formation of Gabosar in northern East Junggar, Xinjiang, also show similarities to A. sokolovi in many aspects, but is distinguished by having shorter septal plates, as well as complete and slightly concave tabulae. Acdalopora breviana Yu, 1960, from the Upper Ordovician (probably Sanqushan Formation of late Katian age) of Jiangxi, southeastern China, has similarly well-developed septal spines. It differs from the present material in having slightly larger corallite size (1.5–1.75 mm in diameter), shorter septa and more convex tabulae.
Table 4. Comparison of Acdalopora species discussed in the present paper. Abbreviations: CoD, corallite diameter; TaD, tabularium diameter; Ta5, number of tabulae per 5 mm. Species name in bold refers to those described in this paper.
| |
Corallite size (mm) |
Tabulae |
Septa |
|
Acdalopora sokolovi Bondarenko, 1958 |
TaD = 0.62–1.59 |
complete, Ta5 = 14–16 |
0.15–0.36 mm |
|
Acdalopora irregularis Lin and Wang, 1986 |
CoD = 1.5–2.0 |
complete, Ta5 = 13–15 |
long |
|
Acdalopora mira Chekhovich, 1977 |
CoD = 1.0–1.5 |
complete, Ta5 = 10–16 |
long |
|
Acdalopora qiakuertensis Lin and Wang, 1985 |
CoD = 1.1–1.7 |
complete and concave, Ta5 = 9–10 |
short |
|
Acdalopora breviana Yu, 1960 |
CoD = 1.5–1.75 |
complete and convex, Ta5 = 11–13 |
short |
Stratigraphic and geographic range.—The Yuanbaoshan Formation (lower Katian) of southern Kuruktag, northeastern Tarim, China; the Dulankara Formation (lower Katian) of the Chu-Ili Terrane, southern Kazakhstan; the Bulongor Formation (Katian) of Hobuksar, northwestern Xinjiang, China; the Upper Ordovician of Chingiz Range, Kazakhstan.
Family Pseudoplasmoporidae Bondarenko, 1963
Genus Navoites Leleshus and Ospanova, 1979
Type species: Navoites simmetricus Leleshus and Ospanova, 1979, Upper Ordovician, Zeravshan-Gissarskiy mountain, Tajikistan.
Remarks.—The revised concept of Navoites proposed by Wang et al. (2020) is accepted herein.
Navoites irregularis (Lin and Chow, 1977)
Figs. 13, 14.
1977 Heliolites sinensis irregularis sp nov.; Lin and Chow 1977: 181, 182, pl. 52: 7a, 7b, not pl. 52: 5a, 5b.
2015 Heliolites sinensis irregularis Lin and Chow, 1977; Dai et al. 2015: 52, pls. 47, 48: 7E, 8E.
?2017 Heliolites sinensis irregularis Lin and Chow, 1977; Han et al. 2017: pl. 15: 6.
Material.—Three sectioned specimens, NIGP 201013–201015 from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, China.
Emended diagnosis.—Navoites with TaDs 0.73–1.15 mm. Corallite walls thin. Septa short. Tabulae complete, sub-horizonal or slightly convex, Ta5 14–16. Corallites commonly separated by 2–4 rows of tubules. Tubular walls incomplete. Diaphragms complete, flat, slightly sagging or convex, 22–24 per 5 mm.
Description.—Three almost complete coralla are measured, of low domical form, with the largest 50.46 mm wide and 21.92 mm high, and the smallest 27.21 mm wide and 21.92 mm. Corallites rounded or sub-rounded, TaDs 0.82–0.96 mm (average 0.92 mm). Walls complete, slightly faceted, WT 0.03–0.09 mm (average 0.05 mm). Septa numbering 12, short, extending one-ninth the TaD. The width of septa 0.11–0.16 mm (average 0.14 mm). Tabulae complete, slightly convex, Ta5 15–16. Corallites commonly separated by 2–4 rows of tubules, with centers 1.13–1.86 mm apart (average 1.54 mm). Tubules having 5 or 6 sides, 0.32 –0.37 mm in diameter, with 12 surrounding each corallite. Tubular walls incomplete, commonly 0.04–0.05 mm thick. Diaphragms generally complete, flat, slightly sagging or convex, mostly 24 per 5 mm.
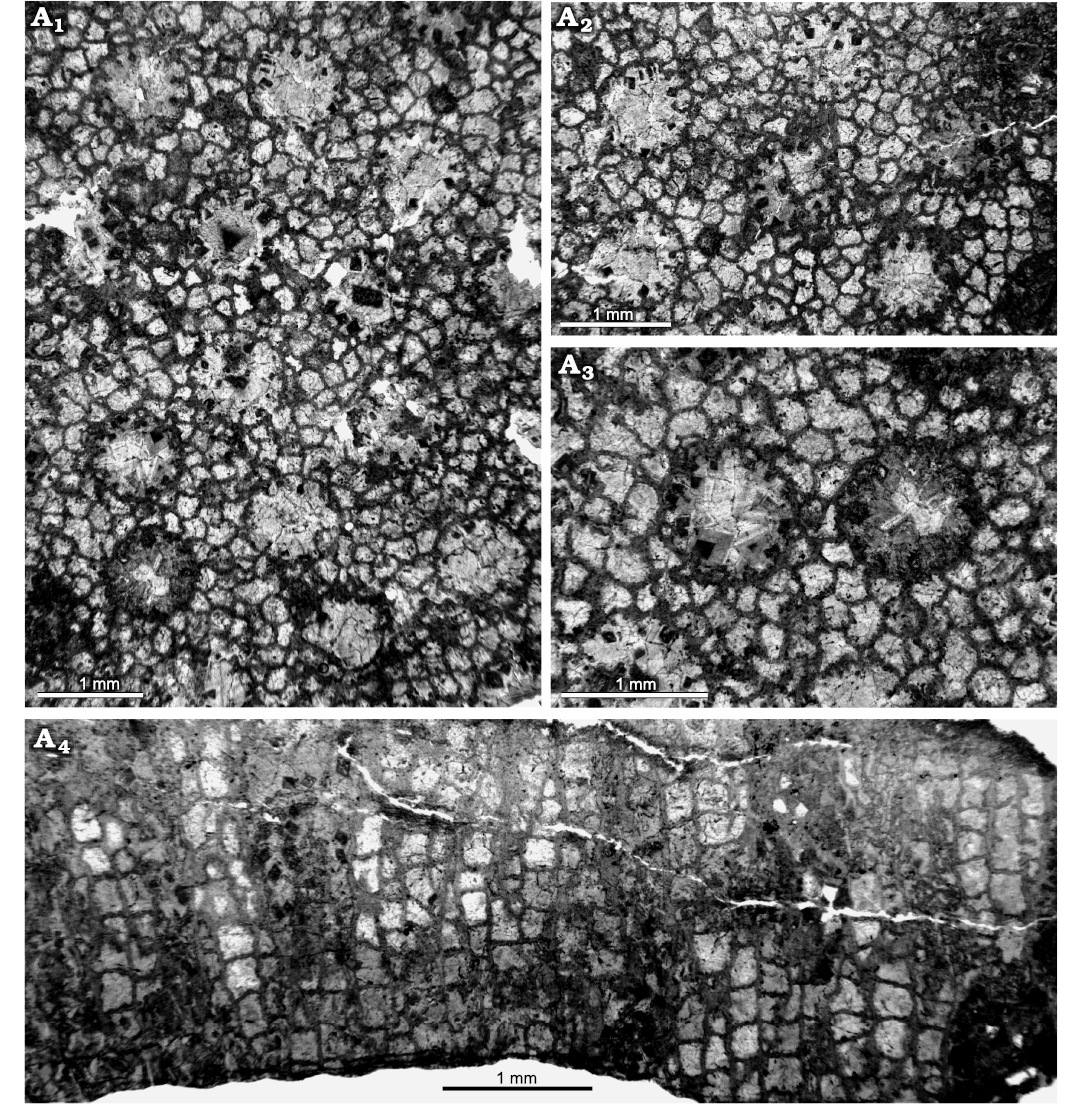
Fig. 13. Heliolitine tabulate coral Navoites irregularis (Lin and Chow, 1977), from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, Northwest China. NIGP 201013. A1–A3, transverse sections. A4, longitudinal section.
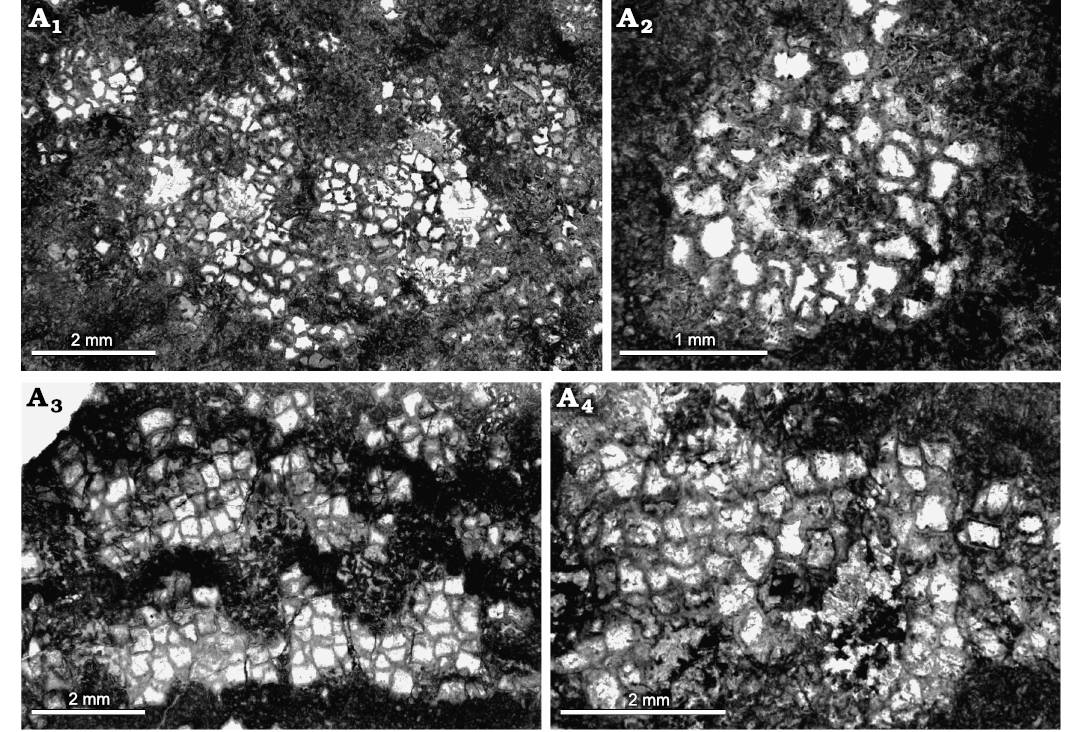
Fig. 14. Heliolitine tabulate coral Navoites irregularis (Lin and Chow, 1977), from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, Northwest China. NIGP 201014. A1, A2, transverse sections. A3, A4, longitudinal sections.
Remarks.—Specimens clearly conspecific with the present material were identified by Lin and Chow (1977) as Heliolites sinensis irregularis, from the middle Sanqushan Formation (upper Katian) of Jiangxi, southeastern China. Their major difference lies in the slightly larger corallites (1.00–1.15 mm in diameter) of the former, and we therefore assign our material to this species. The original generic assignment of H. sinensis irregularis was based on its tubular coenenchyme. We assign this species to Navoites because of the consistent presence of 12 coenenchymal tubules (Dixon and Jell 2012; Zhen et al. 2017; Wang et al. 2020).
The material described by Dai et al. (2015) as H. sinensis irregularis is from the upper Xiazhen Formation (upper Katian) of Jiangxi, Southeast China. It is essentially identical to the holotype of Navoites irregularis, except in its smaller corallite size (0.73–0.81 mm in diameter), and we therefore treat them as conspecific. Han et al. (2017) identified H. sinensis irregularis from the lower Katian “Yingan Formation” of the Kalpin area, northwestern Xinjiang. Their material was poorly illustrated, and its attribution to N. irregularis needs confirmation.
The present revised diagnosis of N. irregularis is based on all the material now assigned to this species, and the major revision is an extension of the range of corallite size (0.73–1.15 mm). Its comparison with similar species is shown in Table 5 and discussed below. Navoites simmetricus Leleshus and Ospanova, 1979, documented from the Upper Ordovician Archalyk Member of the Shakhriomon area, western Zeravshan Ridge, Tajikistan, differs from N. irregularis in having smaller corallite size (around 0.6 mm in diamter) and more complete tubular walls. The material described by Wang et al. (2020) as Navoites cargoensis (Hill, 1957) from the upper Katian Malachis Hill Formation of New South Wales differs in having smaller corallite size (0.67–0.76 mm in diameter), less developed septal plates, and more complete tubular walls.
Table 5. Comparison of Navoites species discussed in the present paper. Abbreviations: CoD, corallite diameter; TaD, tabularium diameter; Ta5, number of tabulae per 5 mm. Species name in bold refers to those described in this paper.
| |
Corallite size (mm) |
Tabulae |
Septa |
Coenenchyme |
|
Navoites irregularis (Lin and Chow, 1977) |
TaD = 0.82–1.15 |
complete and convex, Ta5 = 14–16 |
short |
developed |
|
Navoites simmetricus Leleshus and Ospanova, 1979 |
CoD = 0.6 |
complete, Ta5 = 10–24 |
short |
well developed |
|
Navoites cargoensis (Hill, 1957) |
TaD = 0.67–0.76 |
complete, Ta5 = 5–15 |
absent or weakly developed |
developed |
|
Navoites breviseptata (Lin and Chow, 1977) |
CoD = 0.7–0.85 |
complete, Ta5 = 17–20 |
short |
developed |
|
Navoites crassimuralis (Lin and Huang, 1986) |
CoD = 0.8–0.9 |
complete and concave, Ta5 = 10–12 |
weakly developed |
developed |
Two further species with 12 coenenchymal tubules are transferred herein to Navoites, also exhibiting similarities to N. irregularis. They include Stelliporella breviseptata Lin and Chow, 1977, from the middle Sanqushan Formation (upper Katian) of Jiangxi, southeastern China, and Heliolites crassimuralis Lin and Huang, 1986, from the lower Katian Beiguoshan Formation of Ningxia, northwestern China. Navoites breviseptata differs from N. irregularis in the development of smaller corallites (0.7–0.85 mm in diameter), denser tabulae and shorter septa, and Navoites crassimuralis is distinguished by having less developed septa, more concave tabulae, and more complete diaphragms.
Stratigraphic and geographic range.—The Yuanbaoshan Formation (lower Katian) of southern Kuruktag, northeastern Tarim, China; the middle Sanqushan Formation (upper Katian) of Jiangxi, Southeast China; the upper Xiazhen Formation (upper Katian) of Jiangxi, Southeast China; the “Yingan Formation” (lower Katian) of Kalpin, northwestern Tarim, China.
Family Heliolitidae Lindström, 1876
Genus Apekinella Ospanova, 2012a
Type species: Apekinella zeravshanica Ospanova, 2012a, Upper Ordovician, Shakhriomon-2 section, Shakhriomon district, west of Zeravshan Ridge, Tajikistan.
Remarks.—The diagnostic features of Apekinella were discussed by Ospanova (2012a). The genus was originally assigned to Stelliporellidae Bondarenko, 1971. However, this family is characterized by long septal plates forming a polygonal structure at the axis, a feature being absent in Apekinella. Therefore, Apekinella is placed herein in Heliolitidae Lindström, 1876.
Stelliporella Wentzel, 1895, and Derivatolites Bondarenko, 1971, have similarly longer septal plates, both of which can be distinguished from Apekinella by the presence of axial structure. Derivatolites further differs in having more widely spaced corallites and the development of an axial structure formed by connection of opposite septa.
Apekinella zeravshanica Ospanova, 2012a
Fig. 15.
2012a Apekinella zeravshanica sp nov.; Ospanova 2012a: 45, pl. 5: 1a, 1b.
Material.—Two sectioned specimens, NIGP 201016 and NIGP 201017, both from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, China.
Emended diagnosis.—Species of Apekinella with TaDs 1.00–1.80 mm. Septa long, strongly curved, almost extending to the axis. Tabulae complete, horizontal or slightly concave. Corallites commonly separated by 1–5 rows of discontinuous tubules.
Description.—One complete corallum is measured, of low domical form, 27.58 mm wide and 10.88 mm high. Corallites rounded or sub-rounded, with TaDs varying from 1.36 to 1.70 mm (average 1.51 mm). Corallite walls complete, slightly wavy, 0.04–0.08 mm thick (average 0.07 mm). Septa laminate, 12 in number, with a maximum width of 0.15–0.42 mm (average 0.33 mm). Septa long, strongly curved, almost extending to the axis. Tabulae complete, mostly horizontal or slightly convex, Ta5 15–16. Corallites contiguous or separated by narrow coenenchyme. Coenenchyme consisting of polygonal tubules formed by discontinuous vertical elements, with up to 15 tubules surrounding each corallite. Corallites separated by 1–5 rows of tubules, with centers 1.13–1.86 mm apart (average 1.67 mm). Tubules commonly complete, convex, concave or oblique. Diaphragms 0.22–0.32 mm in diameter, 19–20 per 5 mm.
Remarks.—These specimens differ from the type material of Apekinella zeravshanica Ospanova, 2012a, from the Upper Ordovician Archalyk Member of the Shakhriomon area, western Zeravshan Ridge, Tajikistan, only in having lager crorallites, the latter being about 1.0–1.2 mm in diameter. For this reason, we assign these specimens to Apekinella zeravshanica and revise the range of corallite size as 1.00–1.80 mm in the diagnosis of this species.
Stratigraphic and geographic range.—The Yuanbaoshan Formation (lower Katian) of southern Kuruktag, northeastern Tarim, China; the Archalyk Member (upper Katian) of the Shakhriomon area, western Zeravshan Ridge, Tajikistan.
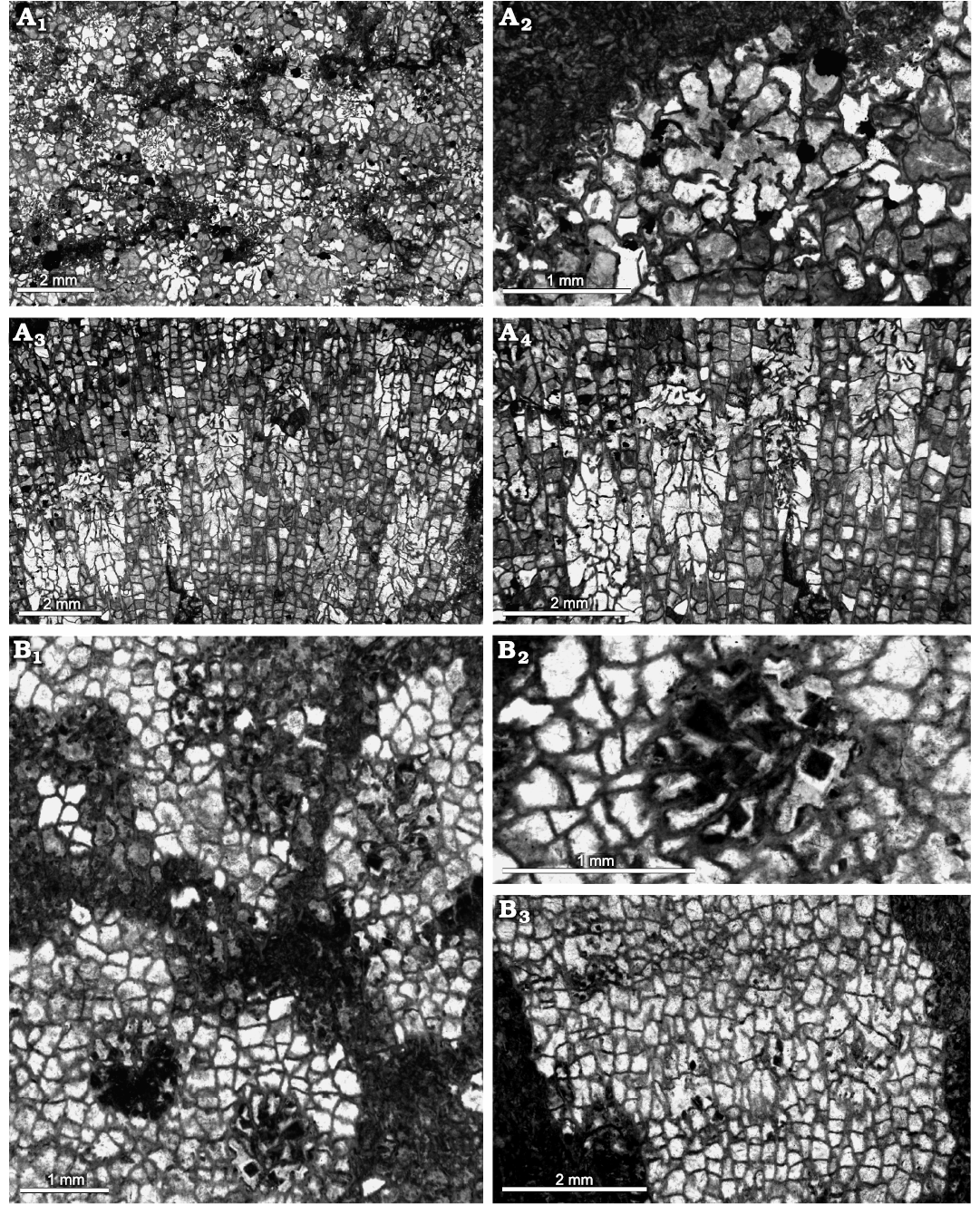
Fig. 15. Heliolitine tabulate coral Apekinella zeravshanica Ospanova, 2012a, from the Yuanbaoshan Formation (lower Katian) of southern Kuruktag, Tarim Basin, Northwest China. A. NIGP 201016. A1, A2, transverse sections. A3, A4, longitudinal sections. B. NIGP 201017. B1, B2, transverse sections. B3, longitudinal section.
Conclusions
In this paper, we revise the early Katian heliolitine coral fauna consisting of eight species of six genera from the Kuruktag area of northeastern Tarim Basin, Northwest China. This fauna is characterized by the dominance of Plasmoporella, accompanied by minor Mongoliolites, Wormsipora, Acdalopora, Navoites, and Apekinella. A comparison with contemporaneous faunas from adjacent blocks suggests that the biogeographic connections of Tarim Block are closest to Chu-Ili and South Tienshan, but relatively weaker with Qilian and North China.
Acknowledgements
We are very grateful to Renbin Zhan, Yi Wang, Peng Tang, Xiaocong Luan, Xiaole Zhang, Hongyong Zhang, Pu Huang, and Yuchen Zhang (all from NIGP), and Jianbo Liu (Peking University, Beijing, China) for their kind provision of the precious specimens examined in this work. We also thank two anonymous referees for their comments on our manuscript. Financial support for this study came from National Natural Science Foundation of China (42072007), Strategic Priority Research Program of Chinese Academy of Sciences (XDB26000000), Youth Innovation Promotion Association of CAS (2020311), and the State Key Laboratory of Palaeobiology and Stratigraphy (LPS).
References
Bondarenko, O.B. 1958. Heliolitida and Tabulata from the Ordovician of Chu-Iliysk Mountains [in Russian]. Trudy Geologičeskogo Instituta, Akademiâ Nauk SSSR 9: 197–228.
Bondarenko, O.B. 1963. Revision of the genus Plasmopora [in Russian]. Paleontologičeskij žurnal 1: 42–52.
Bondarenko, O.B. 1971. Members of the new family Stelliporellidae (heliolitids) [in Russian]. In: V.N. Dubatolov (ed.), Tabulâty i Geliolitoidei Paleozoâ SSSR, Tr. II Vsesoûznogo Simpoziuma po Izučeniû Iskopaemyh Korallov SSSR, 166–178. Nauka, Moskva.
Bondarenko, O.B. 1992. Sistema Geliolitoidej. 206 pp. Moskovskoe Obŝestvo Ispytatelej Prirody, Moskva.
Bondarenko, O.B. and Minzhin, C. [Minžin, Č] 1977. Morphology and astogeny of some Late Ordovician corals of Bayan Kongor (central Mongolia) [in Russian]. In: L.P. Tamarinov (ed.), Bespozvonočnye Paleozoâ Mongolii, 20–31. Nauka, Moskva.
Chekhovich, V.D. [Čehovič, V.D.] 1977. New species of Late Ordovician and Late Silurian Heliolitoidea of Tuva [in Russian]. In: G.A. Stukalina (ed.), Novye Vidy Drevnih Rastenij i Bespozvonočnyh SSSR, Vypusk 4, 19–24. Nauka, Moskva.
Chen, X., Ni, Y.N., and Jiang, N.R. 1990. Graptolites [in Chinese]. In: South Xinjiang Petroleum Prospecting Corporation, Xinjiang Petroleum Administration Bureau, and Institute of Petroleum and Geosciences, Dian-Qian-Gui Petroleum Prospecting Bureau (eds.), Sinian to Permian Stratigraphy and Palaeontology of the Tarim Basin, Xinjiang Volume 1, Kuruktag Area, 122–131. Nanjing University Press, Nanjing.
Chen, X., Zhang, Y.D., Li, Y., Fan, J.X., Tang, P., Chen, Q., and Zhang, Y.Y. 2012. Biostratigraphic correlation of the Ordovician black shales in Tarim Basin and its peripheral regions. Science China Earth Sciences 55: 1230–1237. Crossref
Cui, Y.N. and Wang, G.X. 2021. Revision of late Katian (Late Ordovician) heliolitine corals from northern Kuruktag in northeastern Tarim Basin of China. Alcheringa 45: 178–189. Crossref
Cui, Y.N., Wang, G.X., and Percival, I.G. 2022. Early heliolitine tabulate corals from the Sandbian (Upper Ordovician) in the Yunnan-Sichuan border area, Southwest China. Palaeoworld [published online, https://doi.org/10.1016/j.palwor.2022.01.004]. Crossref
Dai, M.J., Liu, L., Lee, D.J., Peng, Y.B., and Miao, A.S. 2015. Morphometrics of Heliolites (Tabulata) from the Late Ordovician, Yushan, Jiangxi, South China. Acta Geologica Sinica 89: 38–54. Crossref
Deng, Z.Q. 1984. Middle and Upper Ordovician corals from marginal areas of Ordos Platform [in Chinese with English summary]. Bulletin of Nanjing Institute of Geology and Palaeontology, Academia Sinica 8: 306–312.
Deng, Z.Q. and Li, Z.R. 1979. Tabulata [in Chinese]. In: Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (eds.), Palaeontological Atlas of Northwest China, Qinghai Volume, Part 2, 6–16. Geological Publishing House, Beijing.
Dixon, O.A. and Jell, J.S. 2012. Heliolitine tabulate corals from Late Ordovician and possibly early Silurian allochthonous limestones in the Broken River Province, Queensland, Australia. Alcheringa 36: 69–98. Crossref
Dzyubo, P.S. [Dzûbo, P.S.] 1972. The Upper Ordovician and Silurian tabulates and heliolitides of Kuldzhuktau [in Russian]. In: A.S. Masumov and R.A. Abdullaev (eds.), Novye Dannye po Faune Paleozoâ i Mezozoâ Uzbekistana, 18–40. FAN Uzbek SSR, Taškent.
Ehrenberg, C.G. 1834. Beiträge zur physiologischen Kenntniss der Corallenthiere im allgemeinen, und besonders des Rothen Meeres, nebst einem Versuche zur physiologischen Systematik derselben. Königlichen Akademie der Wissenschaften, Physikalisch–Mathematische Klasse 1832: 225–380.
Frech, F. 1897. Referat, J. Wentzel: Kenntniss der Zoantharia tabulate. Neues Jahrbuch für Mineralogie, Geologie und Palaöntologie, Beilage-Bände 1897: 212–214.
Ghobadi Pour, M., Popov, L.E., Kim, A.I., Abduazimova, Z.M., Mikolaichuk, A.V., Kim, I.A., Ospanova, N., Erina, M.V., Salimova, F.A., Klishevich, I., Saidov, M.S., Davlatov, N.H., Abdiyev, N.H., Koldybekov, O.Y., Gordeev, D., Rustamov, S.T., and Yangiboev, S.B. 2023. The Ordovician of Central Asia (Kyrgyzstan, Uzbekistan and Tajikistan). Geological Society, London, Special Publications 533 [published online, https://doi.org/10.1144/SP533-2022-52]. Crossref
Hall, R.L. 1975. Late Ordovician coral faunas from north-eastern New South Wales. Journal and Proceedings of the Royal Society of New South Wales 108: 75–93. Crossref
Han, X.M., Zhang, X.Y., Yang, B.Z., Sun, J.G., Zhou, F.Q., Zhao, P.F., and Wang, Q.T. 2017. Discovery and geological significance of the coral fossils of Yingan Formation in Keping, Xinjiang [in Chinese]. Geological Science and Technology Information 36: 13–19.
Hill, D. 1957. Ordovician corals from New South Wales. Journal and Proceedings of the Royal Society of New South Wales 91: 97–107. Crossref
Hill, D. 1981. Rugosa and Tabulata. In: C. Teichert (ed.), Treatise on Invertebrate Paleontology. Coelenterata, Part F, Supplement 1, 1–762. The Geological Society of America and The University of Kansas, Boulder and Lawrence.
Jiang, H.X., Bao, H.P., Sun, L.Y., Wu, Y.S., and Diao, J.B. 2013. Tabulate and rugose corals from the Ordovician reefs in the southern edge of the Ordos Basin and their paleoecology significance [in Chinese with English abstract]. Acta Palaeontologica Sinica 52: 243–255.
Keller, B.M. 1956. A general review of the Ordovician stratigraphy of the Chu-Ili range [in Russian]. Trudy Geologičeskogo Instituta, Akademii Nauk SSSR 1: 5–49.
Kiaer, J. 1899. Die Korallenfaunen der Etage 5 des norwegischen Silursystems. Palaeontographica 46: 1–60.
Kiaer, J. 1903. Revision der mittelsilurischen Heliolitiden und neue Beitrage zur Stammesgeschichte derselben. Videnskabs-Selskabet i Kristiania, Skrifter 1, Math-Naturvidensk Klasse 10: 1–58.
Kim, A.I. 1966.Tabulâtomorfnye Korally Paleozoâ Zeravšano-Gissarskoj Gornoj Oblasti. 71 pp. Ministerstvo Geologii Uzbekskoj SSR, Taškent.
Kim, A.I. and Apekin, Y.N. [Apekin, Û.N.] 1978. Tabulata and Heliolitoidea [in Russian]. In: B.S. Sokolov and E.A. Jolkin (eds.), Pograničnye sloi Ordovika i Silura Altae-Saânskoj Oblasti i Tyan’-Ŝanâ, 58–64. Trudy Institut Geologii i Geofiziki, Sibirskoe Otdelenie, Akademiâ Nauk SSSR, Moskva.
Kim, A.I., Popov, L.E., and Ivanova, O.B. 2007. Ordovician system–outline of stratigraphy. In: A.I. Kim, F.A. Salimova, and N.A. Meshchankina (eds.), Palaeontological Atlas of Phanerozoic Faunas and Floras of Uzbekistan, Volume 1, Palaeozoic, 25–71. Republic of Uzbekistan State Committee on Geology and Mineral Resources, Tashkent.
Kong, L. and Song, X.L. 1990. Corals [in Chinese with English summary]. In: South Xinjiang Petroleum Prospecting Corporation, Xinjiang Petroleum Administration Bureau, and Institute of Petroleum and Geosciences, Dian-Qian-Gui Petroleum Prospecting Bureau (eds.), Sinian to Permian Stratigraphy and Palaeontology of the Tarim Basin, Xinjiang Volume 1, Kuruktag Area, 154–160. Nanjing University Press, Nanjing.
Kovalevskiy, O.P. [Kovalevskij, O.P.] 1964. Some Upper Ordovician Heliolitoidea from the Chingiz range [in Russian]. Trudy Vsesoûznyj Naučno-Issledovatel’skij Geologičeskij Institut (VSEGEI), Novaâ seriâ 93: 35–47.
Lai, C.G. 1982. Stratigraphy of China (No. 5), The Ordovician System of China [in Chinese]. 297 pp. Geological Publishing House, Beijing.
Leleshus, V.L. [Lelešus, V.V.] 1975. Upper Ordovician Proporidae (Heliolitoidea) from the Zeravshan-Hissar mountainous region [in Russian]. In: M.R. Džalilov (ed.), Voprosy Paleontologii Tadžikistana, 7–24. Doniš, Dušanbe.
Leleshus, V.L. and Ospanova, N.K. 1979. New Late Ordovician Heliolitoidea from central Asia. Münstersche Forschungen zur Geologie und Paläontologie 47: 147–155.
Li, Y.X. and Lin, B.Y. 1982. Subclass Tabulata [in Chinese]. In: Xi’An Institute of Geology and Mineral Resources (ed.), Palaeontological Atlas of Northwest China, Shaanxi-Gansu-Ningxia Volume, Part 1, Precambrian and Early Palaeozoic, 50–93. Geological Publishing House, Beijing.
Lin, B.Y. 1985. Upper Ordovician tabulate corals from the Ejin Banner, Nei mongol (Inner Mongolia) and their stratigraphical significance [in Chinese with English abstract]. Geological Review 31: 118–124.
Lin, B.Y. and Chow, X.H. 1977. Late Ordovician Tabulata and Heliolitida from Zhejiang and Jiangxi provinces, and their stratigraphic significance [in Chinese]. Professional Papers of Stratigraphy and Palaeontology 3: 108–208.
Lin, B.Y. and Chow, X.H. 1980. Some Middle Ordovician corals from Jiangshan County, Zhejiang [in Chinese with English abstract]. Bulletin of the Chinese Academy of Geological Sciences 1: 28–41.
Lin, B.Y. and Huang, H.P. 1986. Late Ordovician tabulate corals from Xinjiang and Ningxia [in Chinese with English summary]. Bulletin of the Chinese Academy of Geological Sciences 12: 127–149.
Lin, B.Y. and Wang, B.Y. 1985. Some Late Ordovician heliolitoid corals from Jiabosar Formation of Jiabosar District, Xinjiang [in Chinese with English abstract]. Acta Palaeontologica Sinica 24: 351–357.
Lin, B.Y. and Wang, B.Y. 1986. Upper Ordovician tabulate corals from western Junggar, Xinjiang, China [in Chinese with English abstract]. Professional Papers in Stratigraphy and Palaeontology 16: 37–72.
Lin, H.L., Zhou, Z.Y., and Luo, H.L. 1990. Trilobites [in Chinese with English summary]. In: South Xinjiang Petroleum Prospecting Corporation, Xinjiang Petroleum Administration Bureau, and Institute of Petroleum and Geosciences, Dian-Qian-Gui Petroleum Prospecting Bureau (eds.), Sinian to Permian Stratigraphy and Palaeontology of the Tarim Basin, Xinjiang Volume 1, Kuruktag Area, 109–122. Nanjing University Press, Nanjing.
Lindström, G. 1876. On the affinities of the Anthozoa Tabulata. Annals and Magazine of Natural History, Series 4 18: 1–17. Crossref
Lindström, G. 1899. Remarks on the Heliolitidae. Kongliga Svenska Vetenskaps-Akademiens Handlingar 32: 1–140.
Milne-Edwards, H. and Haime, J. 1849. Mémoire sur les polypiers appartenant aux groupes naturels des Zoanthaires perforés et des Zoanthaires tabulés. Academie des Sciences de Paris, Comptes Rendus 29: 257–263.
Milne-Edwards, H. and Haime, J. 1850. A Monograph of the British Fossil Corals. Part I. Introduction. Corals from the Tertiary and Cretaceous Formations. ixxxv + 72 pp. Palaeontographical Society, London. Crossref
Norin, E. 1937. Geology of Western Quruqtagh, Eastern Tien-Shan. Reports from the Scientific Expedition to the North-Western Provinces of China under Leadership of Dr. Sven Hedin (III), Geology I. 194 pp. Bokförlags Aktiebolaget Thule, Stockholm.
Ospanova, N.K. 1984. The new data about the age of the Archalyk suite (the mountainous region of Zerafhano-Gissar) [in Russian]. Doklady Akademii Nauk Tadžikskoj SSR 27: 399–401.
Ospanova, N.K. 1991. Heliolitoidea (Ordovician) [in Russian]. In: M.R. Džalilov (ed.), Atlas Iskopaemoj Fauny i Flory Tadžikistana. Ordovik. Silur. Devon, 13–17. Institut Geologii, Akademiâ Nauk Tadžikskoj SSR, Dušanbe.
Ospanova, N.K. 2012a. Leleshusites gen. nov., Apekinella gen. nov. and Senexites gen. nov. are new genera of the Late Ordovician Heliolitida of the southern Tien Shan [in Russian]. In: Sovremennye Voprosy Regional’noj Geodinamiki i Mineragenii Pamiro-Tân’-Šanâ, 44–57. Doniš, Dušanbe.
Ospanova, N.K. 2012b. New Ordovician heliolitine corals family Acdalinidae fam. nov. [in Russian]. In: Sovremennye Voprosy Regional’noj Geodinamiki i Mineragenii Pamiro-Tân’-Šanâ, 27–43. Doniš, Dušanbe.
Ospanova, N.K. 2014. Atlas Verhneordovikskih Geliolitid Ûžnogo Tân’-Šanâ. 194 pp. Akademiâ Nauk Respubliki Tadžikistana, Institut Geologii, Sejsmostojskogo Stroitel’stva i Sejsmologii, Dušanbe.
Ospanova, N.K. 2017. New Data about the Upper Ordovician Heliolitida of South Tien Shan. 97 pp. Lambert Academic Publishing House, Saarbrucken.
Rong, J.Y., Zhan, R.B., Huang, B., Xu, H.K., Fu, L.P., and Li, R.Y. 2017. Ordovician brachiopod genera on type species of China. In: J.Y. Rong, Y.G. Jin, S.Z. Shen, and R.B. Zhan (eds.), Phanerozoic Brachiopod Genera of China, 87–243. Science Press, Beijing.
Scrutton, C.T. 1984. Origin and early evolution of tabulate corals. Palaeontographica Americana 54: 110–118.
Sladkovskaja, M.G. [Sladkovskaâ, M.G.] 1987. Tabulatomorphous corals from the Upper Ordovician of south-eastern Uzbekistan [in Russian]. Paleontologičeskij žurnal 1987: 22–29.
Sokolov, B.S. 1949. Tabulata and Heliolitida [in Russian]. In: M.Z. Âniševskij (ed.), Atlas Rukovodâŝih form Iskopaemyh Faun SSSR, II: Silurijskaâ Sistema, 75–98. Gosgeoltekhizdat, Moskva.
Sokolov, B.S. 1955. Tabulates from Paleozoic of European part of USSR. Introduction (General aspects of systematics and evolution of tabulates) [in Russian]. Trudy VNIGRI 85: 1–527.
Sproat, C.D. and Zhan, R.B. 2018. Altaethyrella (Brachiopoda) from the Late Ordovician of the Tarim Basin, Northwest China, and its significance. Journal of Paleontology 92: 1005–1017. Crossref
Sproat, C.D. and Zhan, R.B. 2019. Paleogeographic and paleoecological significance of Schachriomonia (Brachiopoda) from the Upper Ordovician of the Tarim Basin, Northwest China. Journal of Paleontology 93: 1075–1087. Crossref
Torsvik, T.H. and Cocks, L.R.M. 2013. New global palaeogeographical reconstructions for the Early Palaeozoic and their generation. Geological Society, London, Memoirs 38: 5–24. Crossref
Wang, B.Y. 1981. Tabulata [in Chinese]. In: Regional Work Team of Geological Survey, Xinjiang Geological Bureau (ed.), Palaeontological Atlas of Northwest China. Xinjiang-Uygur Autonomous Region Volume, Part 1, Late Proterozoic and Early Palaeozoic, 39–73. Geological Publishing House, Beijing.
Wang, G.X., Percival, I.G., and Zhen, Y.Y. 2020. The youngest Ordovician (latest Katian) coral fauna from eastern Australia, in the uppermost Malachis Hill Formation of central New South Wales. Alcheringa 44: 356–378. Crossref
Wang, G.X., Percival, I.G., Zhen, Y.Y., and Webby, B.D. 2021. Late Ordovician corals from allochthonous clasts in the Devonian Drik-Drik Formation of northeastern New South Wales, Australia. Proceedings of the Linnean Society of New South Wales 143: 51–86.
Webby, B.D., Elias, R.J., Young, G.A., Neuman, B.E.E., and Kaljo, D. 2004. Corals. In: B.D. Webby, F. Paris, M.L. Droser, and I.G. Percival (eds.), The Great Ordovician Biodiversification Event, 124–146. Columbia University Press, New York. Crossref
Wentzel, J. 1895. Zur Kenntnis der Zoantharia Tabulata. Denkschriften der kaiserlich-königlichen Akademie der Wissenschaften, mathematisch-naturwissenschaftliche Klasse 62: 476–516.
White, D.E. and Yang, S.W. 2004. British Ordovician tabulate corals. Monograph of the Palaeontographical Society 157: 1–92. Crossref
Yang, H.J., Wu, Y.S., Xiao, Z.Y., Huang, Z.B., Jiang, H.X., Pan, W.Q., Liu, L.J., Yang, Z.L., and Chang, X.L. 2015. Cyanobacteria, Algae, Bryozoan, Coral, and Stromatoporoid Fossils from Ordovician of Tarim Basin [in Chinese]. 201 pp. Geological Publishing House, Beijing.
Young, G.A. and Elias, R.J. 1995. Latest Ordovician to earliest Silurian colonial corals of the east-central United States. Bulletins of American Paleontology 108: 1–153.
Young, G.A. and Noble, J.P.A. 1990. Silurian Proporidae and Plasmoporidae (Anthozoa, Tabulata) from the Chaleurs Bay Region, Canada. Journal of Paleontology 64: 184–199. Crossref
Yu, C.M. 1960. Late Ordovician corals of China [in Chinese with Russian abstract]. Acta Palaeontologica Sinica 82: 65–102.
Yu, C.M. 1962. Late Ordovician corals from Menyuan, Qinghai Province [in Chinese]. In: Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (eds.), Geology of Qilian Mountain 4, 1–11. Science Press, Beijing.
Zhan, R.B., Rong, J.Y., Jin, J.S., Liang, Y., Yuan, W.W., Zhang Y.D., and Wang, Y. 2014. Discovery of a Late Ordovician Foliomena fauna in the Tarim desert, Northwest China. Palaeoworld 23: 125–142. Crossref
Zhang, Y.C., Zhan, R.B., Wang, Y., and Tang, P. 2017. Foliomena Fauna from the Zatupo Formation (Katian, Upper Ordovician) of Kuruktag, southeastern Xinjiang, NW China [in Chinese with English abstract]. Acta Palaeontologica Sinica 56: 322–330.
Zhang, Y.D. and Munnecke, A. 2016. Ordovician stable carbon isotope stratigraphy in the Tarim Basin, NW China. Palaeogeography, Palaeoclimatology, Palaeoecology 458: 154–175. Crossref
Zhen, Y.Y., Wang, G.X. and Percival, I.G. 2017. Conodonts and tabulate corals from the Upper Ordovician Angullong Formation of central New South Wales, Australia. Alcheringa 41: 141–168. Crossref
Zhong, D. and Hao, Y.X. 1990. Ordovician System [in Chinese with English summary]. In: South Xinjiang Petroleum Prospecting Corporation, Xinjiang Petroleum Administration Bureau, and Institute of Petroleum and Geosciences, Dian-Qian-Gui Petroleum Prospecting Bureau (eds.), Sinian to Permian Stratigraphy and Palaeontology of the Tarim Basin, Xinjiang, Volume 1, Kuruktag Area, 40–104. Nanjing University Press, Nanjing.
Zhou, Z.Y., Chen, X., Wang, Z.H., Wang, Z.Z., Li, J., Geng, L.Y., Fang, Z.J., Qiao, X.D., and Zhang, T.R. 1990. Ordovician System [in Chinese]. In: Z.Y. Zhou and P.J. Chen (eds.), Biostratigraphy and Geological Evolution of Tarim, 56–130. Science Press, Beijing.
Zhou, Z.Y., Webby, B.D., and Yuan, W.W. 1995. Ordovician trilobites from the Yingan Formation of northwestern Tarim, Xinjiang, northwestern China. Alcheringa 19: 47–72. Crossref
Zhou, Z.Y., Yin, G.Z., and Zhou, Z.Q. 2014. Ordovician (Darriwilian–early Katian) trilobite faunas of northwestern Tarim, Xinjiang, China. Memoirs of the Association of Australasian Palaeontologists 46: 1–142.
Acta Palaeontol. Pol. 68 (2): 273–295, 2023
https://doi.org/10.4202/app.01023.2022