
Palaeohistology helps reveal taxonomic variability in exceptionally large temnospondyl humeri from the Upper Triassic of Krasiejów, SW Poland
ELŻBIETA M. TESCHNER, LAURENT GARBAY, PIOTR JANECKI, and DOROTA KONIETZKO-MEIER
Teschner, E.M., Garbay, L., Janecki, P., and Konietzko-Meier, D. 2023. Palaeohistology helps reveal taxonomic variability in exceptionally large temnospondyl humeri from the Upper Triassic of Krasiejów, SW Poland. Acta Palaeontologica Polonica 68 (1): 63–74.
For more than twenty years, palaeontological excavations have been carried out at the Upper Triassic site of Krasiejów (south-west Poland), providing thousands of skeletal elements belonging to various tetrapod groups. However, almost all bones are preserved in a disarticulated state. This generates problems in taxonomic assignment among closely related groups, e.g., stereospondyl amphibians. As far as cranial elements, the pectoral girdle bones and the intercentra are very diagnostic, while all other remaining skeletal elements are difficult to unambiguously assign between either the capitosaurid Cyclotosaurus intermedius or the trematosaurid Metoposaurus krasiejowensis, both originating from Krasiejów. Because comparative postcranial material from other Triassic localities worldwide is very rare or even absent, assignment on a genus level is even more problematic. With the help of bone histology, we tested the taxonomic assignment of two exceptionally large humeri with only marginally differing morphology. Both humeri show lamellar-zonal bone and preserve between four to seven visible growth cycles, but with a markedly different amount of primary tissue, a varying degree of remodelling and distinct growth patterns. Considering the fact that both humeri are almost the same size, they show different histological ontogenetic stages, implying an assignment to two different genera. UOBS 02116 shows limited remodelling, with a large amount of primary bone preserved as alternating thick zones and thin annuli, it is considered to represent C. intermedius. UOBS 02116, showing limited remodelling, with a large amount of primary bone preserved as alternating thick zones and thin annuli, and considered to represent C. intermedius. UOBS 02431, showing advanced remodelling, residuals of primary bone, distinctly decreasing thickness of zones towards the outer cortex, and thick annuli, is assigned to M. krasiejowensis. This study shows that temnospondyl amphibians might show only minor differences in humeral morphology, making histology an even more valuable tool for taxonomic assignment at a genus level.
Key words: Amphibia, Temnospondyli, Capitosauria, Cyclotosaurus, palaeohistology, Late Triassic, Krasiejów, Poland.
Elżbieta M. Teschner [eteschner@uni.opole.pl, ORCID: https://orcid.org/0000-0001-5961-489X ], Opole University, Institute of Biology, Oleska 22, 45-052 Opole, Poland; University of Bonn, Institute of Geosciences, Nussallee 8, 53115 Bonn, Germany.
Laurent Garbay[garblaur@gmail.com; ORCID: https://orcid.org/0000-0001-6048-2468 ], Natural History Museum Luxembourg, Department of Paleontology, 25 rue Münster, 2160 Luxembourg City, Luxembourg.
Piotr Janecki [piotr.janecki@gmail.com; ORCID: https://orcid.org/0000-0002-3418-9089 ], Dinosaur Park, JuraPark Krasiejów, 1 Maja 10, 46-040 Krasiejów, Poland.
Dorota Konietzko-Meier [dmeier@uni-bonn.de; ORCID: https://orcid.org/0000-0003-4932-7402 ], University of Bonn, Institute of Geosciences, Nussallee 8, 53115 Bonn, Germany.
Received 30 September 2022, accepted 9 December 2022, available online 13 February 2023.
Copyright © 2023 E.M. Teschner et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Temnospondyli Zittel, 1888, is one of the largest non-amniote tetrapod orders, distributed globally with a wide stratigraphic occurrence ranging from the lower Carboniferous to the Lower Cretaceous (Holmes and Carroll 1977; Warren et al. 1997). The systematics of Temnospondyli are based almost exclusively on cranial characters (Schoch 2013). Articulated postcranial material of Mesozoic temnospondyls is very rare, e.g., Dutuitosaurus ouazzoui (Dutuit, 1976), Paracyclotosaurus davidi Watson, 1958, and therefore intraspecific variability at the generic level is not well understood. This makes the assignment of disarticulated bones to a specific genus or species even more difficult and problematic when at least two temnospondyl amphibian species co-occur together in a single locality. This scenario applies to the Norian, Upper Triassic, Krasiejów clay pit in Poland, preserving at least two temnospondyl amphibians, the capitosaurid Cyclotosaurus intermedius Sulej and Majer, 2005, and a trematosaurid Metoposaurus krasiejowensis (Sulej, 2002).
The family Cyclotosauridae Shishkin, 1964, includes eight valid species of Cyclotosaurus Fraas, 1889, with a narrow stratigraphic occurrence (Carnian–Rhaetian), mainly known from European localities in Germany, Poland, France, Luxembourg, Greenland (Meyer and Pleninger 1844; Fraas 1913; Kuhn 1932, 1942; Buffetaut and Wouters 1986; Milner et al. 1996; Sulej and Majer 2005; Witzmann et al. 2016; Marzola et al. 2017; Konietzko-Meier et al. 2019), but also from Thailand (Ingavat and Janvier 1981). The cyclotosaurid cranial material is highly diagnostic, e.g., closed otic notch and the position of the orbits (Damiani 2001; Schoch and Milner 2000), however, the postcranial material is very scarce, and thus not considered as taxonomically valuable. Cyclotosaurus intermedius from Krasiejów is the first species of Cyclotosaurus for which, in addition to the skulls and mandibles, a scapulocoracoid, an interclavicle, a fragmentary clavicle, a cleithrum and a humerus have been described (Sulej and Majer 2005). The only other published morphological description of a cyclotosaurid long bone is based on the German cf. Cyclotosaurus sp. humerus from Bonenburg (Konietzko-Meier et al. 2019).
The family Metoposauridae includes 10 valid species known from Africa, Asia, Europe and South America (Gee and Jasinski 2021). Although D. ouazzoui is the only articulated metoposaurid species, Metoposaurus krasiejowensis and Panthasaurus maleriensis (Roychowdhury, 1965) are known from numerous skeletal elements, including long bones (Roychowdhury 1965; Sengupta 2002; Sulej 2007), which were also studied at the histological level (Konietzko-Meier and Klein 2013; Konietzko-Meier and Sander 2013; Teschner et al. 2018, 2020).
Although the vertebral shape and the ornamentation pattern of the shoulder girdle elements of Cyclotosaurus are characteristic and differ from Metoposaurus (Sulej and Majer 2005; Sulej 2007), it is difficult to distinguish these two taxa based on the long bone morphology. The humerus of C. intermedius described by Sulej and Majer (2005) possesses a broad proximal and distal head, a weakly defined deltopectoral crest and supinator process, a thin entepicondyle and a pronounced ectepicondyle. Sulej (2007) described the humerus of M. krasiejowensis as a short bone with wide proximal and distal end with a clear torsion, and a pronounced ectepicondyle with a short and thin supinator process, whereas the entepicondyle is thicker than the supinator process. Barycka (2007) described a series of M. krasiejowensis humeri as well-ossified bones with well-developed processes, e.g., supinator process, wide and flattened ends, small degrees of torsion unrelated to the bone length, generally obliquely positioned radial condyles, and less prominent deltopectoral crests. Moreover, Barycka (2007) noted, that the smallest bones possess smooth surfaces and weakly developed processes, crests and tuberosities, but a well-developed supinator process, deltopectoral crest and medial epicondyle. In contrast, the lattisimi process, subcoracoscapular process and lateral epicondyle are well-defined in the larger forms. In general, both Barycka (2007) and Sulej (2007) pointed out a high morphological variation within the metoposaurid humeri, and therefore, a difficulty in clear defining diagnostic features (Barycka 2007; Sulej 2007). Therefore, the distinction between trematosaurids and cyclotosaurids is problematic at the morphological level, especially with disarticulated material, as the characters might change during ontogeny.
The size was used as a proxy for genus determination, thus, based on the known skull length (small- to middle-sized Metoposaurus crania and large-sized Cyclotosaurus crania), the small-sized long bones were attributed to Metoposaurus, whereas larger-sized specimens were attributed to Cyclotosaurus (Sulej and Majer 2005; Sulej 2007). However, large metoposaurid skulls are known for D. ouazzoui (67 cm, Dutuit 1976), and a large (ca. 75 cm long) Metoposaurus skull was excavated from Krasiejów during the 2021 field campaign, so the taxonomic assignment of the postcrania is very doubtful based solely on their size. The lack of comparative Cyclotosaurus material and thus the unknown ontogenetic variation among cyclotosaurid humeri makes the final determination difficult.
A histological study conducted on a series of M. krasiejowensis humeri revealed the presence of two histotypes (Teschner et al. 2018). Histotype I showed a growth pattern of alternating growth cycles composed of a moderately vascularised zone and an avascular annulus together, both of a similar thickness and progressive remodelling with ontogeny, whereas histotype II represented humeri growing with a very thick, highly vascularized zone and a very thin annulus and only marginal remodelling. Teschner et al. (2018) primarily proposed that the observed difference in the humeral growth pattern could be a result of various populations separated in time and/or space, and therefore showing a different growth pattern due to variation in local environmental conditions. However, an affiliation to two different taxonomic groups was not ruled out (Teschner et al. 2018). Later, the histology of a cf. Cyclotosaurus sp. humerus from Bonenburg showed a clear difference in the growth pattern between M. krasiejowensis and cf. Cyclotosaurus sp., making the histology of the latter similar to histotype II observed in M. krasiejowensis humeri (Teschner et al. 2018; Konietzko-Meier et al. 2019). Hence, the high variation in morphological characters in M. krasiejowensis humeri and the occurrence of two histological growth patterns could be linked to an interspecific variation rather than to an intraspecific variation.
The aim of this study is to find out whether the differences in the amphibian humeral morphology and histology indeed reflect the intraspecific variability in the Krasiejów assemblage. Here we describe for the first time the histology and the growth pattern of two exceptionally large humeri, initially assigned to C. intermedius, from the Late Triassic of Krasiejów. We compared it with the known histological growth pattern of M. krasiejowensis, another temnospondyl amphibian co-occurring at this site. Moreover, we test whether the humeral microstructure provides information about environmental adaptation and allows us to distinguish the mode of life between Metoposaurus and Cyclotosaurus.
Institutional abbreviations.—IGPB, Institute of Geosciences, Section Palaeobiology, University of Bonn, Germany; LWL-MFN P, Landschaftsverband Westfalen-Lippe, Museum für Naturkunde, Abteilung Paläontologie, Münster, Germany. UOBS/UOPB, old/new collection catalogue, Biosystematics/Palaeobiology, University of Opole, Opole, Poland; ZPAL, Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland.
Other abbreviations.—An, annulus; EFS, External Fundamental System; LAG, Line of Arrested Growth; Zo, zone.
Material and methods
Krasiejów is a village in south-west Poland located approximatelly 30 km north-west from Opole. The locality exhibits Upper Triassic, most-probably Norian strat (Bodzioch and Kowal-Linka 2013; Jewuła et al. 2019). The multi-taxic bone-bearing layers preserve thousands of disarticulated bones, belonging to both amniote and non-amniote taxa (e.g., Sulej 2002; Sulej and Majer 2005; Dzik and Sulej 2007).
The sectioned material includes two large temnospondyl humeri, UOBS 02116 (146.4 mm; Fig. 1; Table 1) and UOBS 02431 (141.6 mm; Fig. 2; Table 1), belonging to two temnospondyl amphibians, initially assigned to Cyclotosaurus intermedius. Both bones originated from Krasiejów and are housed at the Institute of Biology, University of Opole. The material was the subject of a muscle reconstruction study; however, no morphological description was provided, and the taxonomic assignment was based solely on the large size of the humeri (Lech 2010). In addition, two Metoposaurus krasiejowensis humeri (UOPB 00109 representing histotype I and UOBS 00547 representing histotype II sensu Teschner et al. (2018) and cf. Cyclotosaurus sp. LWL-MNF P 64371 humerus (Konietzko-Meier et al. 2019) were used for bone compactness analyses (Table 1).
Table 1. Measurements (in mm) of humeri of Cyclotosaurus spp. and Metoposaurus krasiejowensis (Sulej, 2002). *Originally published as M. krasiejowensis (Lech 2010; Teschner et al. 2018), herein revised as Cyclotosaurus intermedius.
|
Specimen number |
Taxon |
Total length [mm] |
Midshaft width [mm] |
Distal width [mm] |
Proximal width [mm] |
Torsion [°] |
Compactness [%] |
Reference |
|
UOBS 02116 |
Cyclotosaurus intermedius* |
141.6 |
40 |
86 |
50 |
55 |
85 |
this study |
|
UOBS 02431 |
Metoposaurus krasiejowensis |
146.4 |
48 |
84 |
48 |
83 |
71 |
this study |
|
UOBS 00574 |
Cyclotosaurus intermedius* |
65.4 |
20.5 |
36.1 |
24 |
65 |
82 |
Teschner et al.
2018; |
|
UOPB 00109 |
Metoposaurus krasiejowensis |
69.2 |
22.1 |
43.4 |
26.5 |
75 |
66 |
|
|
LWL-MFN P 64371 |
cf. Cyclotosaurus sp. |
167 |
50 |
87 |
50 |
34 |
78 |
Konietzko-Meier et al. 2019; this study |
|
ZPAL Ab III 887/3 |
Cyclotosaurus intermedius |
130 |
32 |
62 |
48 |
70 |
– |
Sulej and Majer 2005; this study |
Both specimens were measured using a sliding calliper with precision of 0.05 mm. The histological samples were taken from the histological midshaft of both humeri (Figs. 1–3) yielding the most complete growth record (Konietzko-Meier and Klein 2013; Teschner et al. 2018). The taken samples were processed into standard petrographic thin-sections in the IGPB laboratory in Bonn (Germany) applying the technique described in Klein and Sander (2007). Wet silicon carbide (SiC) abrasive powders with grit sizes of 600 and 800 were used to grind and polish to a final thin-section thickness ranging between 25–40 μm. Osteohistological examination was performed with a Leica DM LP polarising light microscope and photographs were taken with a Leica DFC 420 camera attached to the microscope. The overview pictures in polarised light were merged using Photoshop 5. For the microanatomical analyses of UOBS 02116 and 02431, as well as of UOPB 00109, UOBS 00574, and LWL-MNF P 64371 the thin-sections were scanned (Fig. 3; Table 1) with an Epson Perfection 750V PRO scanner and transformed into black (bone) and white (cavities) images (Fig. 4; Table 1). In order to calculate the bone compactness, the bw-counter pixel counting software developed by ©Peter Göddertz (IGPB) was used to measure the ratio of compact bone vs. cavities (Table 1). The detailed morphology and histology of the additional specimens used for bone compactness analyses (UOPB 00109, UOBS 00574, and LWL-MNF P 64371) was described previously (Teschner et al. 2018; Konietzko-Meier et al. 2019). The morphological nomenclature follows Barycka (2007) and Sulej (2007) and the histological nomenclature follows Francillon-Vieillot et al. (1990) and de Buffrénil et al. (2021). It is important to mention that metaplastic bone forms in bones of dermal origin, however, the herein studied long bones are of a periosteal origin (de Buffrénil et al. 2021). The term “metaplastic-like bone” used in this study is not linked to the bone origin but is referred exclusively as a structural resemblance observed previously in Temnospondyli (Konietzko-Meier et al. 2019; Teschner et al. 2020) and therefore, does not represent a true metaplastic bone.
Results
Morphological description.—UOBS 02116 is 141.6 mm long (Fig. 1; Table 1), has a short shaft (40 mm) and a slender proximal end (50 mm) compared to its wide distal end (86 mm). Anteriorly, the distal end has a pronounced 6.3 mm long ectepicondyle (epicondylus lateralis sensu Barycka 2007) and 10.9 mm long supinator process (Fig. 1A1, A2). Proximally, the ectepicondyle proceeds into the radial crest (adductor crest sensu Barycka 2007) before ending in the 28.5 mm long latissimi process (Fig. 1A1). Anterior to the ectepicondyle, the short supinator process is located dorsally, the supinator process is longer than the ectepicondyle and extends proximally into the deltopectoral crest (Fig. 1A1, A2). Posteriorly, the humeroradial tuberosity (Fig. 1) is located dorsally, with its rugose insertion of the grand pectoral muscle (Dutuit 1976) or biceps brachii (Ochev 1972). The epicondylar depression forms a groove between the supinator process and ectepicondyle, and is 14.5 mm wide (Fig. 1A1). On the opposite side of the distal end, the entepicondyle (medial epicondyle sensu Barycka 2007) proceeds proximally (16.4 mm), and is wider than the supinator process. The postero-dorsal depression forming a groove between the latissimi process and subcoracoscapular process is 12.8 mm wide. Despite four processes, the proximal end is compact and thick, with an oval-shaped scapulocoracoid articular facet (a major axis of 25.5 mm and a minor axis of 22 mm). In comparison, the distal end shows a 14 mm thin rectangular shape antero-posteriorly with only the ectepicondyle and supinator process disrupting the shape. The proximal and the distal articulation facets form an angle of 55° (Table 1).
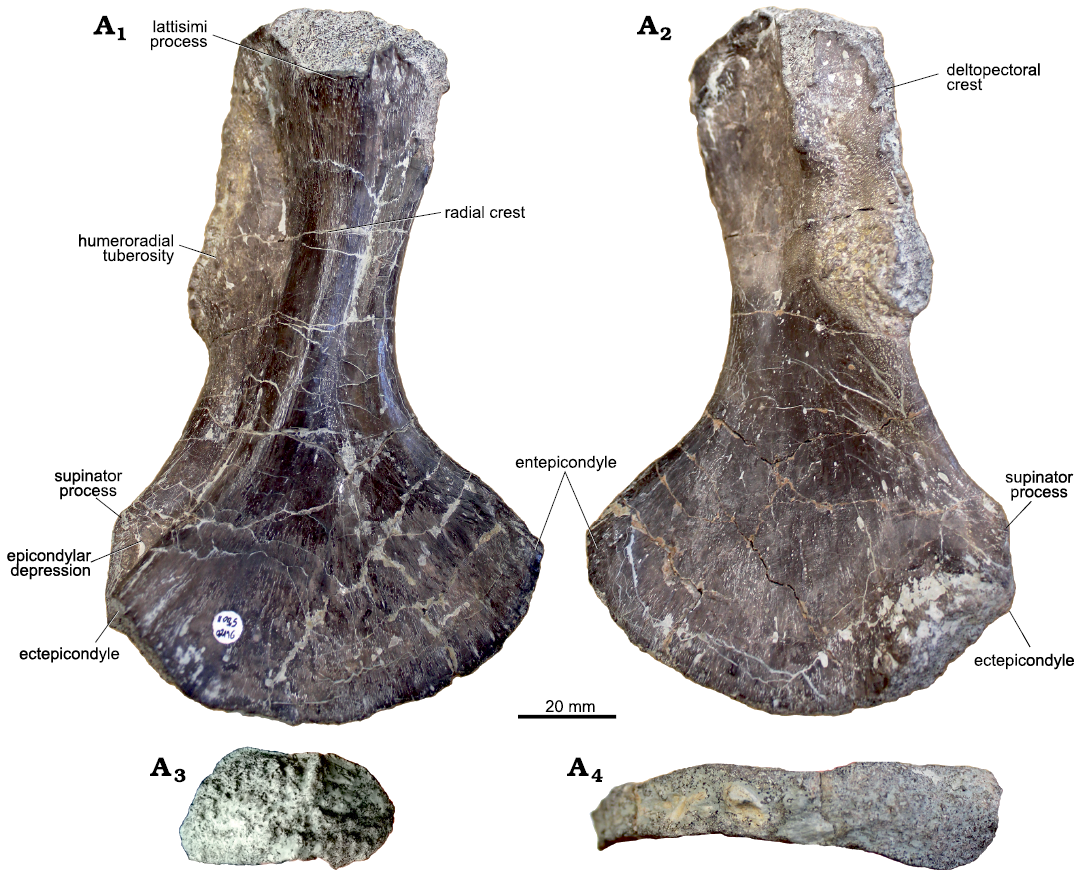
Fig. 1. Morphology of humerus of capitosaurid amphibian Cyclotosaurus intermedius Sulej and Majer, 2005, UOBS 02116, from Krasiejów (Poland), Upper Triassic, in dorsal (A1), ventral (A2), proximal (A3), and distal (A4) views.
UOBS 02431 is 146.4 mm long (Fig. 2; Table 1) and has a short and slender shaft of the same width as the proximal end (48 mm), whereas the distal end is wider (84 mm). Anteriorly, the distal end shows a pronounced 11.1 mm long ectepicondyle and a 15.8 mm long supinator process (Fig. 2A1, A2). Proximally, the ectepicondyle proceeds into the radial crest and ends in a 21 mm long latissimi process (Fig. 2A1). Anterior to the ectepicondyle, the short, dorsally situated supinator process is longer than the ectepicondyle and extends proximally into the deltopectoral crest (Fig. 2A1, A2). Posteriorly, the humeroradial tuberosity (Fig. 2A1) with a rugose insertion of the grand pectoral muscle (Dutuit 1976) or biceps brachii (Ochev 1972) is preserved. The epicondylar depression is 18 mm wide and forms a groove between the supinator process and ectepicondyle (Fig. 2A1). On the opposite side of the distal end, the entepicondyle proceeds proximally into the 20.7 mm wide subcoracoscapular process, being thicker than the supinator process. The postero-dorsal depression forming a groove between the latissimi process and subcoracoscapular process, is 14.6 mm wide. Despite four processes, the proximal end is compact and thick, with an oval-shaped scapulocoracoid articular facet (a major axis of 45 mm and a minor axis of 29 mm). In comparison, the distal end shows a 13.5 mm thin rectangular shape with only the ectepicondyle and supinator process disrupting the shape. The proximal and distal articulation facet form an angle of 83° (Table 1).
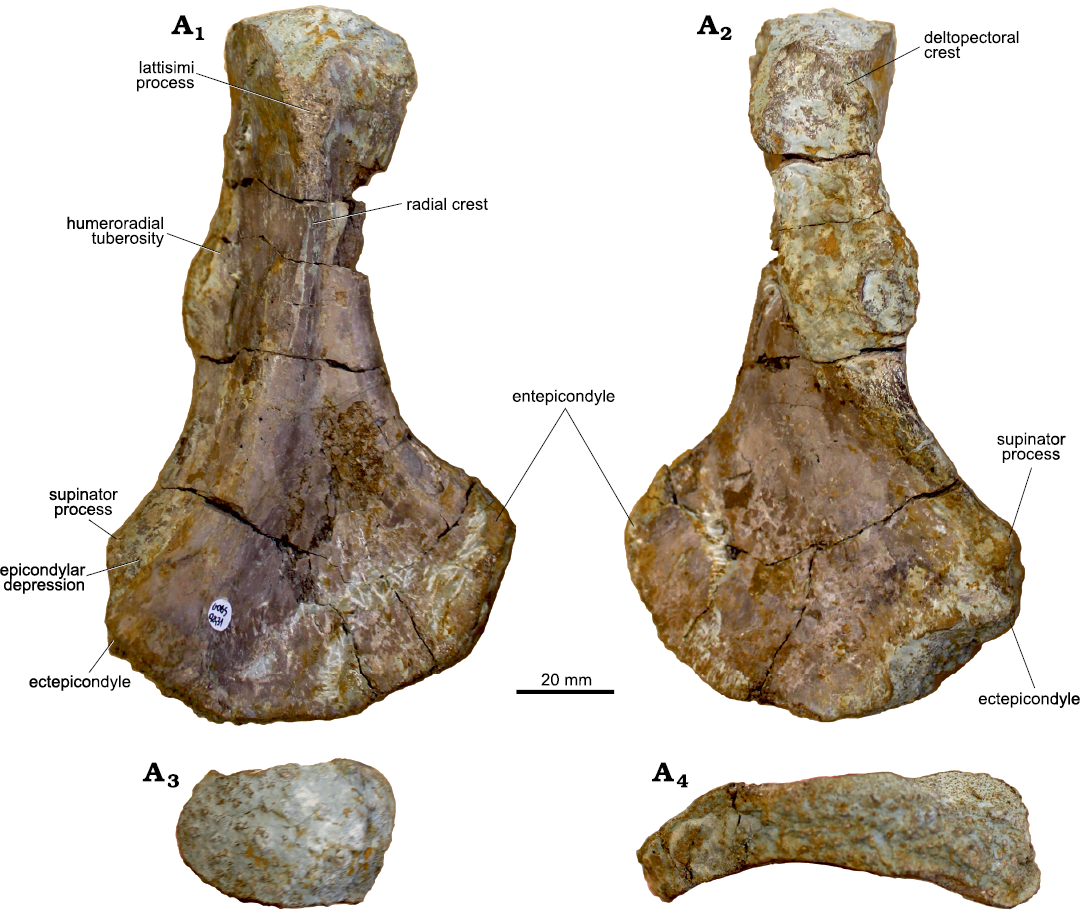
Fig. 2. Morphology of humerus of trematosaurid amphibian Metoposaurus krasiejowensis (Sulej, 2002), UOBS 02431, from Krasiejów (Poland), Upper Triassic in dorsal (A1), ventral (A2), proximal (A3), and distal (A4) views.
Microstructure.—UOBS 02116 (Fig. 3A; Table 1) represents a humerus sectioned at the histological midshaft with the nutrient canal preserved on the ventral side, whereas humerus UOBS 02431 (Fig. 3B; Table 1) has the nutrient canal positioned on the dorso-lateral side. Both humeri are triangular in cross-section and the apex of the humerus, which corresponds with the radial crest (Figs. 1, 2), is on the dorsal side of the bone. UOBS 02116 is prominently flattened (Fig. 3A). In UOBS 02116 the shorter flange borders the lateral side and the longer flange runs towards the medial side. In UOBS 02431 the flanges are of almost equal length (Fig. 3B). The bone compactness value equals 85% for UOBS 02116 (Table 1; Fig. 4A) and 71% for UOBS 02431 (Table 1; Fig. 4B). The bone compactness for the Metoposaurus krasiejowensis UOPB 00109 is 66% (Table 1; Fig. 4C) and 82% for UOBS 00574 (Table 1; Fig. 4D). For LWL-MFN P 64371 cf. Cyclotosaurus sp. the bone compactness is 78% (Table 1; Fig. 4E).
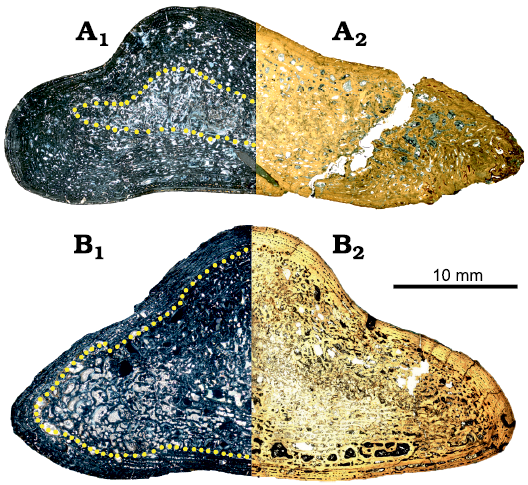
Fig. 3. Histology of humerus of capitosaurid amphibian Cyclotosaurus intermedius Sulej and Majer, 2005, UOBS 02116 (A) and trematosaurid amphibian Metoposaurus krasiejowensis (Sulej, 2002), UOBS 02431 (B), from Krasiejów (Poland), Upper Triassic. A1, B1, in polarized light; A2, B2, in normal transmitted light. The dotted line marks the border between the perimedullary region (internal) and the primary cortex (external).
General histology.—Neither of the two humeri, UOBS 02116 and 02431, shows a medullary cavity (Fig. 3), instead the perimedullary region is developed in both individuals. The boundary between the perimedullary region, composed of secondary trabeculae, and the primary cortex is gradual, without a sharp border (Fig. 3). The primary cortex preserved around the section in both specimens consists of parallel-fibred bone with varying degrees of collagen fibre organization (Fig. 5). On the medial side of the section UOBS 02116, the tissue becomes less unorganized and fibrous, resembling a “metaplastic-like tissue”, which reaches up to the outermost surface of the bone (Fig. 5A3, A4). The vascular density is moderate to high in both sections (Fig. 5). The vascular canals in the primary cortex are mostly longitudinal primary osteons (Fig. 5A1, A2, B1, B2). The simple, longitudinal vascular canals are scattered in the inner cortex, whereas towards the outermost cortex they are arranged in rows (Fig. 5B5, A6). Secondary osteons are preserved in both sections; however, they are more numerous in the UOBS 02431 than in UOBS 02116 (Fig. 5A6, B). Osteocyte lacunae are not common in both sections; however, they are more numerous in UOBS 02116 than in UOBS 02431 (Fig. 5A6, B5). Moreover, they are larger in UOBS 02116 and possess well-developed canaliculi (Fig. 5A5), whereas the osteocytes in UOBS 02413 lack canaliculi (Fig. 5B3). There are regions of well-vascularized but less organised parallel-fibered bone, referred as zones (zo; Fig. 5A6, B5), and regions of well-organized parallel-fibered bone without or only with few vascular canals, referred as annuli (an; Fig. 5A6, B5).
Growth pattern.—UOBS 02116: The innermost part of the section preserves crushed secondary trabeculae (Figs. 3A, 5A6). The primary matrix reaches very deep into the section (Figs. 2A1, A2, 5A6) and the primary tissue is distributed almost throughout the section. In the inner part of the cortex, the erosion cavities are numerous, unorganized, and vary in shape and size (Figs. 4A, B, 5B3). The erosion cavities are lined by lamellar bone, and are embedded in a less organised primary parallel-fibred bone matrix, with remains of the remodelled thick, first zone (Fig. 5A1, A2, A6). This is followed by a remodelled and thin, first annulus (Fig. 5A6). The second zone (Fig. 5A6) consists of a combination of elongated primary osteons, secondary osteons, and a few large, scattered erosion cavities; however, towards the outer cortex, they are more organized and primarily arranged in rows. Next, a thin, avascular layer is present, resembling the second annulus (Fig. 5A6). The third zone (Fig. 5A6) is similar in thickness to the second zone, but with a lower number of large erosion cavities, but with numerous primary osteons. Further, a thick, third annulus is preserved in the outer cortex (Fig. 5A6). In the outermost cortex a thin, fourth zone consisting of one row of primary osteons embedded in a higher organized parallel-fibered bone is preserved (Fig. 5A6). The primary osteons are not roundish but elongated (parallel to the surface), and only in the region of the radial crest do the canals become more radial. Prominent rest lines occur only in the outermost cortex, inside the thin, fourth annulus (Fig. 5A6). Clear LAGs or an EFS are absent in this section. Generally, the zones are distinctly thicker than the annuli and no thinning of the zones towards the outer cortex is observed in this sample.
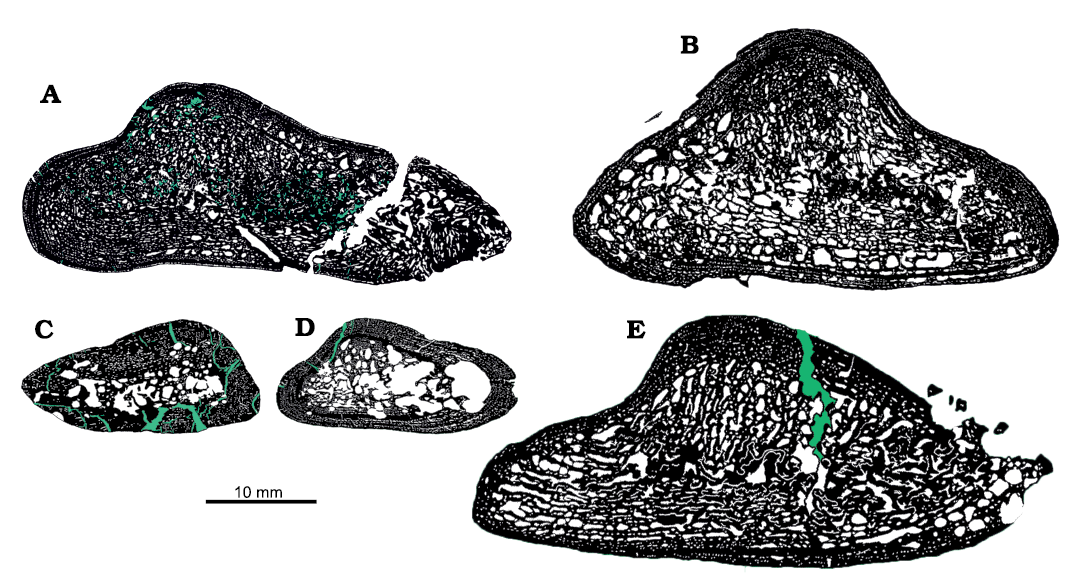
Fig. 4. Microanatomy of the humeri of temnospondylian amphibians from Triassic of Krasiejów (Poland), Norian (A–D) and Bonenburg (Germany), Rhaetian (E). A, C. Capitosaurid Cyclotosaurus intermedius Sulej and Majer, 2005. A. UOBS 02116. C. UOBS 00574 (histotype II sensu Teschner et al. 2018). B, D. Trematosaurid Metoposaurus krasiejowensis (Sulej, 2002). B. UOBS 02431. D. UOPB 00109 (histotype I sensu Teschner et al. 2018). E. cf. Cyclotosaurus, LWL-MFN P 64371. Black, bone; white, pores; green, diagenetic cracks.

Fig. 5. Detailed histology of the humeri of capitosaurid amphibian Cyclotosaurus intermedius Sulej and Majer, 2005, UOBS 0216 (A) and trematosaurid amphibian Metoposaurus krasiejowensis (Sulej, 2002), UOBS 02431 (B); from Krasiejów (Poland), Upper Triassic. A1, A2, close-up of the primary cortex consisting of less-organised parallel-fibered bone; A3, fibrous, “metaplastic-like tissue” on the medial bone side; A4, close-up from A3 of the unorganized, fibrous “metaplastic-like tissue”; B1, B2, close-up of the primary cortex consisting of less-organised and well-organised parallel-fibered bone; A5, osteocyte lacunae with well-preserved canaliculi; B3, osteocyte lacunae lacking canaliculi; B4, B5, growth pattern with preserved residuals of the thick annuli and zones (zo I–III) and thin annuli and zones (zo IV–VII); A6, growth pattern with preserved thin annuli and thick zones (zo I–IV), the dotted line marks the border between the perimedullary region and the cortex. Arrows in A5 and B3 indicate osteocyte lacunae; in B4, B5, and A6 indicate the annuli. Growth pattern in B4 figured on the lateral section side, in B5 and A5 on the ventral side; note the cortex thickness variation between B4 and B5. A1, A3, A4, A6, B1, B4, B5 in polarized light and A2, A5, B2, B3 in normal transmitted light. Abbreviations: an, annulus; ec, erosion cavity; pmr, perimedullary region; pos, primary osteon; sos, secondary osteon; zo, zone.
UOBS 02431: The central most part of the section consists of diagenetically crushed secondary trabeculae (Figs. 3B, 4B), and together with large erosion cavities they represent the perimedullary region. In the inner part of the cortex, on the dorso-medial side, a highly remodelled layer with large erosion cavities is present, resembling the first zone (Fig. 5B4). Between the erosion cavities, remains of more organized parallel-fibred bone are visible, resembling a thick, first annulus (Fig. 5B4). Next, a thick, extensively remodelled second zone with multiple, scattered erosion cavities is present (Fig. 5B4). Following that, the residual of the second, thick annulus is preserved (Fig. 5B4), and further a thick, third zone is deposited (Fig. 5B4, B5). The third annulus is thin and avascular (Fig. 5B4, B5). Zones four to seven are very thin and consist of a single row of simple vascular canals and primary and secondary osteons, which become more remodelled towards the section centre (Fig. 5B4, B5). They are best to observe on the ventral side (Fig. 5B5). In the outermost cortex, the thin and avascular annuli (fourth to seventh) are present (Fig. 5B5); moreover, each of the annuli contains a prominent rest line. Generally, the zones become distinctly thinner towards the outermost cortex (Fig. 5B4, B5).
Discussion
Morphological and ontogenetic variation.—The humeri sectioned herein are exceptionally large (Figs. 1, 2; Table 1) compared to the other amphibian humeri originating from the Krasiejów site (Barycka 2007; Sulej 2007; Teschner et al. 2018); however, they are morphologically very similar to each other. The most striking differences are the degree of torsion between the distal and proximal head in UOBS 02116 (55°; Fig. 1; Table 1) and UOBS 02431 (83°; Fig. 2; Table 1) and the thickness of the ectepicondyle, which is almost twice larger in UOBS 02431 (Fig. 1) than in UOBS 02116 (Fig. 2; Table 1). These two characters make UOBS 02116 thinner and flatter in the lateral view than UOBS 02431.
However, it is not clear how important these characters are in the context of taxonomic assignment. Sulej and Majer (2005) provided the degree of torsion between the proximal and distal head of 70° for C. intermedius (130 mm; Table 1), while the only other known cyclotosaurid humerus of cf. Cyclotosaurus sp. (167 mm, Table 1) shows a torsion of 34° (Konietzko-Meier et al. 2019). Barycka (2007) noted a degree of torsion ranging 22–40° for M. krasiejowensis (59.8–107.5 mm). Sulej (2007) did not provide measurements, but pointed out that torsion vary greatly, with the angle even exceeding 90°. Teschner (2015) measured additional M. krasiejowensis humeri (65.4–82.6 mm), which also showed strong torsional variability (55–85°) that was not related to ontogenetic development. Based on the data for metoposaurids, it is not possible to state how important torsion is for a taxonomic diagnosis. Sulej (2007) observed that the ectepicondyle is very pronounced in M. krasiejowensis humeri.
Other morphological features that are helpful in distinguishing Cyclotosaurus from Metoposaurus are the shape of the proximal head and the anterior edge (Sulej and Majer 2005; Barycka 2007; Sulej 2007). Sulej (2007) notes that the proximal head in C. intermedius is slender and has a narrower ectepicondyle and much shorter anterior edge of the shaft than that of M. krasiejowensis (Sulej 2007). However, Sulej (2007) emphasises that the length of the anterior margin is variable in Metoposaurus, making the latter feature doubtful and non-diagnostic.
After comparing the above-mentioned features with the bones described herein, the humerus UOBS 02116 is significantly less torted and posesses a thinner ectepicondyle than UOBS 02431, but the proximal flange is wide in both specimens. Both humeri have the same shape of the anterior margin of the shaft and both lack a pronounced supinator process (Table 1). Nevertheless, it is important to point out that the strong torsion of humerus UOBS 02431 makes the estimation of the shape of the anterior margin more difficult. Therefore, taxonomic assignment of the humeri to a certain genus based solely on morphology is doubtful, as the range of ontogenetic variability in species of Cyclotosaurus is unknown due to the lack of comparative material. Furthermore, no large M. krasiejowensis humeri are known, neither from Krasiejów nor from other locality worldwide, hence no ontogenetic series are known and no further studies on morphological development were conducted to date.
Nevertheless, some of the humeri published earlier as M. krasiejowensis show similar characters to C. intermedius, e.g., weakly developed supinator process and an overall slim shape; however, all were significantly smaller than the C. intermedius humeri studied herein (Sulej 2007; Barycka 2007; Teschner et al. 2018). Due to the very small number of known humeri of C. intermedius, it cannot be ruled out that some of the bones examined so far and assigned to M. krasiejowensis based on their small size (Sulej 2007; Barycka 2007; Teschner et al. 2018) indeed represent juvenile C. intermedius humeri (Table 1), especially, since M. krasiejowensis humeri are exceptionally variable morphologically (Sulej 2007; Barycka 2007). The high variability among M. krasiejowensis (Barycka 2007; Sulej 2007) might thus be related to interspecific, rather than intraspecific variation. Therefore, a revision of the morphological characters of the stereospondyl postcranial material is necessary.
Since there are only two large humeri assigned to Cyclotosaurus (C. intermedius and cf. Cyclotosaurus sp.) and no other comparative material is available worldwide, bone histology might shed new light on the biology of the animals and help with taxonomic assignment.
Histology and growth pattern.—The thin-sections of both humeri are triangular in cross-section, however UOBS 02116 is much flatter (Fig. 3). The nutrient canal enters from the the ventral side in UOBS 02116 (Fig. 3A), whereas in UOBS 02431 it enters from dorso-lateral side (Fig. 3B). Further, UOBS 02116 possesses a distinct “metaplastic-like structure” on the medial side, corresponding with the deltopectoral crest (Figs. 3B, 4A, 5A3).
At the histological level, both humeri show a lamellar-zonal bone tissue composed of parallel-fibred bone with similar vascular density and preservation of alternating growth marks (zones and annuli; Figs. 3, 5A1, A4, A5, B2, B3. However, the zones in UOBS 02431 are mostly preserved only in the outermost portion of the cortex and consist only of one row of vascular canals, separated by thin avascular annuli (Fig. 5B4, B5). Moreover, only residuals of the primary tissue are still preserved locally, next to the lateral margin in the deeper part of the cortex (Fig. 3B). In this region, the presence of thick annuli alternating with thinner zones is preserved. In UOBS 02116, on the other hand, the alternating growth marks are visible throughout the entire section and can be followed into the deeper parts of the cortex (Figs. 3A, 5A6). The zones are of an equal thickness and are separated by thin annuli. Interestingly, LAGs are not preserved in either sample, which might be related to climatic and/or environmental influence and confirms a rather mild climate during the Late Triassic in Krasiejów, as previously suggested for other taxa from this locality (Teschner et al. 2020, 2022). Only in UOBS 02431, in the external most cortex, few rest lines occur, indicating slowed growth (Francillon-Vieillot et al. 1990; de Buffrénil et al. 2021). Moreover, UOBS 02116 does not show a clear trend of decreasing thickness of growth marks towards the outer cortex; instead the bone was still actively growing as expressed by the presence of thick zones (Fig. 5A6).
Ontogenetic age is related to bone remodelling, thus, the older the specimen the more remodelled the tissue (Klein and Sander 2008; Konietzko-Meier et al. 2012; Mitchell et al. 2017). The most striking difference between these two bones is the degree of remodelling (Fig. 3). Considering that both bones are of similar size, the amount of the primary tissue in UOBS 02116 is high and is still preserved in the deeper portion of the cortex (Fig. 3A1), whereas in UOBS 02431 it is limited only to the outer cortex (Fig. 3B1). The distinctly extensive remodelling and a clear decrease of growth rate observed in the humerus UOBS 02431, indicates an older ontogenetic stage of that bone when compared to the second humerus UOBS 02116, which was still actively growing. The growth pattern of the histologically older bone (UOBS 02431) resembles the pattern known for M. krasiejowensis long bones, growing with unusually thick annuli (Konietzko-Meier and Klein 2013; Konietzko-Meier and Sander 2013). The humerus (120 mm) of the metoposaurid Panthasaurus maleriensis shows extensive remodelling and a similar thinning of the zones towards the outer cortex (Teschner et al. 2020). Unfortunately, no humeri of the metoposaurid Dutuitosaurus ouazzoui from Morocco were examined, however, the femora (79–157 mm) show a similar growth pattern to M. krasiejowensis, except for the deposition of distinct LAGs in the Moroccan taxon, which are absent in the Polish taxon (Steyer et al. 2004). Additionally, a similar position of the nutrient canal entering from the dorso-lateral side was observed in UOPB 00109 of M. krasiejowensis (Teschner et al. 2018). Further, neither UOBS 02431 nor the M. krasiejowensis humeri studied in Teschner et al. (2018) show a similar “metaplastic-like structure”.
The only other histologically examined cyclotosaurid humerus was described by Konietzko-Meier et al. (2019). Unfortunately, the growth pattern of cf. Cyclotosaurus sp. (LWL-MFN P 64371) was not described, nevertheless, the observation made on the available material shows a high amount of primary tissue also preserved in the inner portion of the bone, whereas the growth marks contain a thin annulus and a thick zone (Konietzko-Meier et al. 2019), similar to the growth pattern present in the humerus UOBS 02116 (Figs. 3A, 5B3). Furthermore, the latter specimen shows a distinct “metaplastic-like structure” in the region corresponding with the deltopectoral crest on the medial side (Fig. 5A3, A4), similar to the tissue observed in the cf. Cyclotosaurus sp. humerus from Bonenburg (Konietzko-Meier et al. 2019). Unfortunately, the cf. Cyclotosaurus sp. specimen from Bonenburg was not sectioned at the exact midshaft plane, and therefore, the nutrient canal is not visible in the thin-section (Konietzko-Meier et al. 2019).
The difference in the ontogenetic stage in the sampled bones is significant (Fig. 3) as it would not be expected for bones of similar size. The age, together with the different histological characters, strongly suggests that the observed variability is related to the presence of two different taxa and does not represent intraspecific variation. Thus, the younger bone represents a medium-sized humerus of C. intermedius (UOBS 02116; Figs. 1, 3A, 4A, 5B3), while the older bone belongs to a very large and old Metoposaurus (UOBS 02431; Figs. 2, 3B, 4B, 5A5, B2).
If growth pattern is a reliable indicator for taxonomic differences, the question arises whether the previously observed histotypes in M. krasiejowensis humeri actually represent an intraspecific variation or rather express an interspecific variation (Teschner et al. 2018). Therefore, the assumption previously applied to the long bones from Krasiejów that small-sized animals belong to Metoposaurus and large-sized humeri belong to Cyclotosaurus is thus invalid. After the re-evaluation of the humeri described in Teschner et al. (2018) we can certainly state that humeri described as histotype I, e.g., UOPB 00109 (Fig. 4D), growing with alternating zones and annuli represent small- to medium-sized M. krasiejowensis humeri. In contrast, humeri described as histotype II, e.g., UOBS 00574 (Fig. 4C), outlying by its growth with a very thick zone and only one, extremely thin annulus and a high degree of vascularisation (Teschner et al. 2018), represents a juvenile Cyclotosaurus humerus. Moreover, UOBS 00574 proposed herein to be a juvenile Cyclotosaurus, does not show a distinct torsion between the distal and proximal head, and despite its high vascularity, the bone compactness value is 82% (Table 1; Fig. 4C). In contrast, the compactness of the M. krasiejowensis humeri sampled by Teschner et al. (2018) ranges between 55–76% and for UOPB 00109 it is 66% (Table. 1; Fig. 4D). The cyclotosaurid, cf. Cyclotosaurus sp. from Bonenburg (Konietzko-Meier at al. 2019) shows a similar growth pattern as UOBS 02116, which is proposed herein to represent a species of Cyclotosaurus. Moreover, the bone compactness of the cyclotosaurid LWL-MFN P 64371 is also relatively high with 78% (Table. 1; Fig. 4E).
Mode of life as deduced from bone compactness.—Only a few humeri from Mesozoic Temnospondyli have been examined histologically. Sanchez et al. (2010) sampled the capitosaurid Mastodonsaurus giganteus, however, these authors did not provide a detailed description of the histology nor figured the section, therefore, neither comparison of the growth pattern nor of the bone compactness is possible. The Indian capitosaurids Paracyclotosaurus crookshanki and Cherninia denwai (Mukherjee et al. 2020) show a varied growth pattern within the sampled bones, and a preservation of various bone tissues (woven fibred bone in juveniles and an incipient fibro-lamellar bone with the tendency to parallel-fibred and lamellar bone deposition later in ontogeny). Mukherjee at al. (2020) proposed a different niche occupation for the two examined taxa, a passive benthic predation for Ch. denwai, and a shallow water predation with a possible terrestrialization for P. crookshanki. Moreover, P. crookshanki shows a clear torsion in the bones whereas in Ch. denwai this character is variable (Mukherjee et al. 2020).
In general, UOBS 02431, referred here as Metoposaurus krasiejowensis, shows a higher degree of vascularisation and therefore has a less compact cortex (bone compactness value of 71%; Table 1; Fig. 4B). UOBS 02116 has a more compact cortex (bone compactness value of 85%; Table 1; Fig. 4A) which could indicate occupation of different ecological niches in the water column. Therefore, the more compact bone observed for Cyclotosaurus intermedius could be an adaptation to a lifestyle close to the water surface and a possible degree of terrestrialisation, as in the modern giant salamanders Andrias spp. or as proposed for P. crookshanki (Mukherjee et al. 2020), while the lower bone porosity value of UOBS 02431 indicates a more active swimming lifestyle as proposed previously for metoposaurids (Kalita et al. 2022). However, even if the animals have similar bone lengths, they do not represent an ontogenetically corresponding stage, but rather represent two different species. Therefore, the reflected change in bone density (85% for UOBS 02116 and 71% for UOBS 02431; Fig. 4A, B; Table 1) does not represent an environmental signal or a mode of life. Instead, the tendency of increasing porosity in the older specimen, as observed herein in UOBS 02431, was described in M. krasiejowensis dermal bones of the pectoral girdle (Kalita et al. 2022). Thus, no information about the mode of life and or niche occupation, based on the bone compactness study, can be provided.
Conclusions
The fossil record of the stereospondyl postcranial skeleton is poor. Herein two sectioned humeri, despite their similar bone length and a uniform morphology, show a different growth pattern at the histological level. Therefore, the described specimens are assigned to two different temnospondyl genera. The humerus UOBS 02116 shows a low degree of torsion between the proximal and distal end, and is attributed to Cyclotosaurus intermedius. On the other hand, the specimen UOBS 02431, with a prominent degree of torsion between the proximal and distal head of 83°, is attributed to Metoposaurus krasiejowensis. Histologically, the specimen UOBS 02116 shows more compact bone than the specimen UOBS 02431. Both humeri show a slightly varied growth pattern, with the Cyclotosaurus humerus showing four visible cycles and the Metoposaurus humerus preserving seven visible growth cycles. Despite a similar bone length, UOBS 02116 represents a histologically younger individual than UOBS 02431. The older age of the latter is reflected by the distinctive thinning of zones and thickening of annuli towards the outer cortex, as well as advanced remodelling. The absence of clear LAGs in both specimens suggests that the climate in Krasiejów was rather mild during the Late Triassic, as previously proposed. Furthermore, this study indicates that capitosaurids and trematosaurids share a similar humeral morphology and therefore, assignment at the genus level of disarticulated material is problematic, but bone histology is a valuable tool in the taxonomic assignment of Temnospondyli.
Acknowledgements
Olaf Dülfer (IGPB), Krzysztof Książkiewicz (University of Wrocław, Poland) are thanked for the preparation of the specimens. Tomasz Szczygielski and Tomasz Sulej (ZPAL) are thanked for proving access to the collection with comparative material. Sudipta Kalita (IGPB) is acknowledged for the preparation of figures for the bone compactness analysis. We are also grateful to the Editor Daniel E. Barta (Oklahoma State University, Tahlequah, USA), and the reviewers Damien Germain (National Musuem of Natural History, Paris, France) and Justyna Słowiak-Morkovina (ZPAL) for their helpful comments and suggestions. This research has been funded by the Polish National Science Centre (NCN) grant number: UMO-2016/23/N/ST10/02179.
References
Barycka, E. 2007. Morphology and ontogeny of the humerus of the Triassic temnospondyl amphibian Metoposaurus diagnosticus. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 243: 351–361. Crossref
Bodzioch, A. and Kowal-Linka, M. 2012. Unraveling the origin of the Late Triassic multitaxic bone accumulation at Krasiejów (S Poland) by diagenetic analysis. Palaeogeography, Palaeoclimatology, Palaeoecology 346: 25–36. Crossref
Buffetaut, E. and Wouters, G. 1986. Amphibian and reptile remains from the Upper Triassic of Saint-Nicolas-de-Port (eastern France) and their biostratigraphic significance. Modern Geology 10: 133–145.
Damiani, R.J. 2001. A systematic revision and phylogenetic analysis of Triassic mastodonsauroids (Temnospondyli: Stereospondyli). Zoological Journal of the Linnean Society 133: 379–482. Crossref
de Buffrénil, V., de Ricqlès, A.J., Zylberg, L., and Padian, K. (eds.). 2021. Vertebrate Skeletal Histology and Paleohistology. 837 pp. CRC Press, Paris. Crossref
Dutuit, J.M. 1976. Introduction à l’étude paléontologique du Trias continental marocain. Description des premiers stegocephales recueillis dans le couloir d’Argana (Atlas occidental). Mémoires du Museum national d‘histoire naturelle Série C 36: 1–253.
Dzik, J. and Sulej, T. 2007. A review of the early Late Triassic Krasiejów biota from Silesia, Poland. Phytopatologia Polonica 64: 3–27.
Fraas, E. 1889. Die Labyrinthodonten der schwäbischen Trias. Palaeontographica 36: 1–158.
Fraas, E. 1913. Neue Labyrinthodonten aus der schwäbischen Trias. Palaeontographica 60: 275–294.
Francillon-Vieillot, H., de Buffrénil, V., Castanet, J., Géraudie, J., Meunier, F.J., Sire, J.Y., Zylberg, L., and de Ricqlès, A. 1990. Microstructure and mineralization of vertebrate skeletal tissues. Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends 1: 471–530. Crossref
Gee, B.M. and Jasinski, S.E. 2021. Description of the metoposaurid Anaschisma browni from the New Oxford Formation of Pennsylvania. Journal of Paleontology 95: 1061–1078. Crossref
Holmes, R. and Carroll, R. 1977. A temnospondyl amphibian from the Mississippian of Scotland. Bulletin of the Museum of Comparative Zoology 147: 489–511.
Ingavat, R. and Janvier, P. 1981. Cyclotosaurus cf. posthumus Fraas (Capitosauridae, Stereospondyli) from the Huai Hin Lat Formation (Upper Triassic), Northeastern Thailand: with a note on capitosaurid biogeography. Geobios 14: 711–725. Crossref
Jewuła, K., Matysik, M., Paszkowski, M., and Szulc, J. 2019. The late Triassic development of playa, gilgai floodplain, and fluvial environments from Upper Silesia, southern Poland. Sedimentary Geology 379: 25–45. Crossref
Kalita, S., Teschner, E.M., Sander, P.M., and Konietzko-Meier, D. 2022. To be or not to be heavier: the role of dermal bones in the buoyancy of the Late Triassic temnospondyl amphibian Metoposaurus krasiejowensis. Journal of Anatomy 241: 1459–1476. Crossref
Klein, N. and Sander, P.M. 2007. Bone histology and growth of the prosauropod dinosaur Plateosaurus engelhardti von Meyer, 1837 from the Norian bonebeds of Trossingen (Germany) and Frick (Switzerland). Special Papers in Palaeontology 77: 169–206.
Klein, N. and Sander, P.M. 2008. Ontogenetic stages in the long bone histology of sauropod dinosaurs. Paleobiology 34: 247–263. Crossref
Konietzko-Meier, D. and Klein, N. 2013. Unique growth pattern of Metoposaurus diagnosticus krasiejowensis (Amphibia, Temnospondyli) from the Upper Triassic of Krasiejów, Poland. Palaeogeography, Palaeoclimatology, Palaeoecology 370: 145–157. Crossref
Konietzko-Meier, D. and Sander, P.M. 2013. Histology of long bones of Metoposaurus diagnosticus krasiejowensis (Temnospondyli) from the Late Triassic of Krasiejów (Opole, Silesia Region). Journal of Vertebrate Paleontology 33: 1003–1018. Crossref
Konietzko-Meier, D., Bodzioch, A., and Sander, P.M. 2012. Histological characteristics of the vertebral intercentra of Metoposaurus diagnosticus (Temnospondyli) from the Upper Triassic of Krasiejów (Upper Silesia, Poland). Earth and Environmental Science Transactions of the Royal Society of Edinburgh 103: 237–250. Crossref
Konietzko-Meier, D., Werner, J.D., Wintrich, T., and Sander, P.M. 2019. A large temnospondyl humerus from the Rhaetian (Late Triassic) of Bonenburg (Westphalia, Germany) and its implications for temnospondyl extinction. Journal of Iberian Geology 45: 287–300. Crossref
Kuhn, O. 1932. Labyrinthodonten und Parasuchier aus dem mittleren Keuper von Ebrach in Oberfranken. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie Abhandlungen 69: 94–144.
Kuhn, O. 1942. Über Cyclotosaurus hemprichi Kuhn und einige weitere Tetrapodenreste aus dem Keuper von Halberstadt. Beiträge zur Geologie von Thüringen 6: 181–197.
Lech, K.A. 2010. Reconstruction of forelimb musculature of temnospondyl amphibian Cyclotosaurus intermedius Sulej & Majer, 2005 from the Late Triassic of Poland. In: D. Nowakowski (ed.), Morphology and Systematics of Fossil Vertebrates, 52–66. DN Publisher, Wrocław.
Marzola, M., Mateus, O., Shubin, N.H., and Clemmensen, L.B. 2017. Cyclotosaurus naraserluki, sp. nov., a new Late Triassic cyclotosaurid (Amphibia, Temnospondyli) from the Fleming Fjord Formation of the Jameson Land Basin (East Greenland). Journal of Vertebrate Paleontology 37: 1303501. Crossref
Meyer, H.V. and Plieninger, T. 1844. Beiträge zur Paläontologie Württembergs, enthaltend die fossilen Wirbeltierreste aus den Triasgebilden mit besonderer Rücksicht auf die Labyrinthodonten des Keupers. 132 pp. Schweizerbart, Stuttgart.
Milner, A.R., Duffin, C., and Delsate, D. 1996. Plagiosaurid and capitosaurid amphibian material from the Late Triassic of Medernach, Grand-Duchy of Luxembourg: preliminary note. Bulletin de la Sociètè belge de Gèologie 104: 42–53.
Mitchell, J., Sander, P.M., and Stein, K.H. 2017. Can secondary osteons be used as ontogenetic indicators in sauropods? Extending the histological ontogenetic stages into senescence. Paleobiology 43: 321–342. Crossref
Mukherjee, D., Sengupta, D.P., and Rakshit, N. 2020. New biological insights into the Middle Triassic capitosaurs from India as deduced from limb bone anatomy and histology. Papers in Palaeontology 6: 93–142. Crossref
Ochev, V.G. [Očev, V.G.] 1972. Kapitozavroidnye labirintodonty ûgo-vostoka evropejskoj časti SSSR. 269 pp. Saratov State Univeristy, Saratov.
Roychowdhury, T. 1965. A new metoposaurid amphibian from the Upper Triassic Maleri Formation of Central India. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 250: 1–52. Crossref
Sanchez, S., Germain, D., de Ricqlès, A., Abourachid, A., Goussard, F., and Tafforeau, P. 2010. Limb-bone histology of temnospondyls: implications for understanding the diversification of palaeoecologies and patterns of locomotion of Permo-Triassic tetrapods. Journal of Evolutionary Biology 23: 2076–2090. Crossref
Schoch, R.R. 2013. The evolution of major temnospondyl clades: An inclusive phylogenetic analysis. Journal of Systematic Palaeontology 11: 673–705. Crossref
Schoch, R.R. and Milner, A.R. 2000. Stereospondyli. Encyclopedia of Palaeoherpetology. Part 3B. 203 pp. Verlag Dr. Friedrich Pfeil, Munich.
Sengupta, D.P. 2002. Indian metoposaurid amphibians revised. Paleontological Research 6: 41–65.
Shishkin, M.A. [Šiškin, M.A.] 1964. Suborder Stereospondyli [in Russian]. In: Y.A. Orlov (ed.), Osnovy Paleontologii. Zemnovodnye, presmykaûŝesâ, pticy, 83–122. Nauka, Moskva.
Steyer, J.S., Laurin, M., Castanet, J., and de Ricqlès, A. 2004. First histological and skeletochronological data on temnospondyl growth: palaeoecological and palaeoclimatological implications. Palaeogeography, Palaeoclimatology, Palaeoecology 206: 193–201. Crossref
Sulej, T. 2002. Species discrimination in the Late Triassic labyrinthodont Metoposaurus. Acta Palaeontologica Polonica 47: 535–546.
Sulej, T. 2007. Osteology, variability, and evolution of Metoposaurus, a temnospondyl from the Late Triassic of Poland. Palaeontologia Polonica 64: 29–139.
Sulej, T. and Majer, D. 2005. The temnospondyl amphibian Cyclotosaurus from the Late Triassic of Poland. Palaeontology 48: 157–170. Crossref
Teschner, E.M., Chakravorti, S., Sengupta, D.P., and Konietzko-Meier, D. 2020. Climatic influence on the growth pattern of Panthasaurus maleriensis from the Late Triassic of India deduced from paleohistology. PeerJ 8: e9868. Crossref
Teschner, E.M., Konietzko-Meier, D., and Klein, N. 2022. Growth and limb bone histology of aetosaurs and phytosaurs from the Late Triassic Krasiejów locality (SW Poland) reveals strong environmental influence on growth pattern. Contributions to Zoology 91 (3): 199– 232. Crossref
Teschner, E.M., Sander, P.M., and Konietzko-Meier, D. 2018. Variability of growth pattern observed in Metoposaurus krasiejowensis humeri and its biological meaning. Journal of Iberian Geology 44: 99–111. Crossref
Warren, A., Rich, T.H., and Vickers-Rich, P. 1997. The last labyrinthodonts. Palaeontographica A 247: 1–24. Crossref
Watson, D.M.S. 1958. A new labyrinthodont (Paracyclotosaurus) from the Upper Trias of New South Wales. Bulletin of the British Museum of Natural History, London (Geology) 3: 233–263.
Witzmann, F., Sachs, S., and Nyhuis, C. 2016. A new species of Cyclotosaurus (Stereospondyli, Capitosauria) from the Late Triassic of Bielefeld, NW Germany, and the intrarelationships of the genus. Fossil Record 19: 83–100. Crossref
Zittel, K.A. 1888. Vertebrata (Pisces, Amphibia, Reptilia, Aves). In: Handbuch der Palaeontologie: Palaeozoologie. 598 pp. Oldenbourg, Munich.
Acta Palaeontol. Pol. 68 (1): 63–74, 2023
https://doi.org/10.4202/app.01027.2022