
New palaeoscolecid plates from the Cambrian Stage 3 of northern Mongolia
IBAN GOÑI, CHRISTIAN B. SKOVSTED, LUOYANG LI, GUOXIANG LI, MARISSA J. BETTS, DORJ DORJNAMJAA, GUNDSAMBUU ALTANSHAGAI, BATKHUYAG ENKHBAATAR, and TIMOTHY P. TOPPER
Goñi, I., Skovsted, C.B., Li, L., Li, G., Betts, M.J., Dorjnamjaa, D., Altanshagai, G., Enkhbaatar, B., and Topper, T.P. 2023. New palaeoscolecid plates from the Cambrian Stage 3 of northern Mongolia. Acta Palaeontologica Polonica 68 (1): 117–125.
New material of disarticulated paleoscolecid remains have been found in “Small Shelly/Skeletal Fossils” assemblages from Cambrian Stage 3 extracted from a section in the Khubsugul Lake region of northern Mongolia. The current material is composed of isolated phosphatic plates, rendering the whole-body reconstruction and comparisons difficult. However, the morphology of the plates is unique enough to warrant description of a new genus and species Floraconformis egiinensis. The new taxon is characterised by a stellate depression network spreading from the middle that separates numerous elevations. Floraconformis egiinensis gen. et sp. nov. represents one of the oldest records of isolated palaeoscolecid plates.
Key words: Small Shelly Fossils, biomineralization, palaeoscolecid, Cambrian Explosion, Mongolia.
Iban Goñi [iban.goni@ens-lyon.fr; ORCID: https://orcid.org/0000-0001-6672-2696 ], Université de Lyon, UCBL, ENSL, CNRS, LGL-TPE, 69622 Villeurbanne, France; Department of Palaeobiology, Swedish Museum of Natural History, Box 50007, SE-104 05 Stockholm, Sweden.
Christian B. Skovsted [Christian.skovsted@nrm.se; ORCID: https://orcid.org/0000-0001-7366-7680 ] and Timothy P. Topper [Timothy.Topper@nrm.se; ORCID: https://orcid.org/0000-0001-6720-7418 ], Department of Palaeobiology, Swedish Museum of Natural History, Box 50007, SE-104 05 Stockholm, Sweden; Shaanxi Key Laboratory of Early Life and Environments, State Key Laboratory of Continental Dynamics and Department of Geology, Northwest University, Xi’an 710069, China.
Luoyang Li [liluoyang@ouc.edu.cn; ORCID: https://orcid.org/0000-0002-8276-1000 ], Department of Palaeobiology, Swedish Museum of Natural History, Box 50007, SE-104 05 Stockholm, Sweden; Frontiers Science Center for Deep Ocean Multispheres and Earth System, Key Lab of Submarine Geosciences and Prospecting Techniques, MOE and College of Marine Geosciences, Ocean University of China, Qingdao 266100, China.
Guoxiang Li [gxli@nigpas.ac.cn; ORCID: https://orcid.org/0000-0002-1598-4765 ], State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing 210008, China.
Marissa J. Betts [marissa.betts@une.edu.au; ORCID: https://orcid.org/0000-0002-4884-825X ], Shaanxi Key Laboratory of Early Life and Environments, State Key Laboratory of Continental Dynamics and Department of Geology, Northwest University, Xi’an 710069, China; Palaeoscience Research Centre/LLUNE, School of Environmental and Rural Science, University of New England, Armidale, New South Wales 2351, Australia.
Dorj Dorjnamjaa [ddorj2001@yahoo.com; ORCID: https://orcid.org/0000-0002-6141-0519 ], Institute of Paleontology, Mongolian Academy of Sciences, Ulaanbaatar 15160, Mongolia.
Gundsambuu Altanshagai [shagaiceo@gmail.com; ORCID: https://orcid.org/0000-0003-2851-0697 ] and Batkhuyag Enkhbaatar [battar_paleo@yahoo.com; ORCID: https://orcid.org/0000-0002-2905-3312 ], Institute of Paleontology, Mongolian Academy of Sciences, Ulaanbaatar 15160, Mongolia; School of Arts and Sciences, National University of Mongolia, Ulaanbaatar 14200, Mongolia.
Received 10 October 2022, accepted 5 January 2023, available online 8 February 2023.
Copyright © 2023 I. Goñi et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Palaeoscolecids represent an extinct class of millimetric to decimetric worm-like organisms characterized by an annulated, cylindrical body with a cuticle consisting of numerous individual button-shaped plates or platelets. Recovered as body fossils and isolated phosphatic or phosphatized plates, palaeoscolecids occur widely from Cambrian Series 2/Stage 3 to the upper Silurian (Müller and Hinz-Schallreuter 1993; Ivantsov and Wrona 2004; Conway Morris and Peel 2010; Topper et al. 2010; Garcia-Bellido et al. 2013; Wang et al. 2014; Smith 2015; Kouraiss et al. 2018; Wendruff et al. 2020; Whitaker et al. 2020; Leibach et al. 2021; Shi et al. 2022). Most studies agree that palaeoscolecids sensu lato represent a grade within ecdysozoans, although their precise biological affinity within the Ecdysozoa remains unresolved (Smith 2015). However, palaeoscolecids sensu stricto (see Smith 2015; Smith and Dhungana 2022) are more likely to be the stem lineage of either Priapulida or Nematomorpha (Harvey et al. 2010; Wills et al. 2012). Recent studies proposed the creation of a new clade Palaeoscolecidomorpha (Shi et al. 2022), which is equivalent to Palaeoscolecida sensu lato in Harvey et al. (2010). Significantly, this clade is introduced as stem-group Priapulida, not as a potential stem lineage of the Nematomorpha.
Here we report fossils found in the form of isolated plates in a SSFs assemblage from the Khubsugul Lake region of northern Mongolia. All names applied to palaeoscolecid plates in SSFs assemblages (for example the common taxon Hadimopanella Gedik, 1977) are regarded as form taxa, due to their isolated nature (Topper et al. 2010). Indeed, making the connection between isolated plates and a whole-body taxon is difficult. Different types of plates may co-occur on the same animal (e.g., Palaeoscolex spinosus Ivantsov and Wrona, 2004 or Hadimopanella oezgueli Gedik, 1977; Barragán et al. 2014b) and morphologically identical plates may occur in different biological species (e.g., Rhomboscolex chaoticus Müller and Hinz-Schallreuter, 1993, and Corallioscolex gravius Müller and Hinz-Schallreuter, 1993). Nonetheless, the vast majority of plates described herein are sufficiently different from any previously described palaeoscolecid fossils to merit taxonomic description as a new genus and species, Floraconformis egiinensis gen. et sp. nov.
Nomenclatoral acts.—This published work and the nomenclatural acts it contains, have been registered in ZooBank: urn:lsid:zoobank.org:pub:D9B96732-5853-49C5-9AA8-FFF4A283F85E
Institutional abbreviations.—NRM, Swedish Museum of Natural History, Stockholm, Sweden.
Other abbreviations.—SSFs, Small Shelly Fossils.
Material and methods
The samples were collected during the 2019 field campaign to northern Mongolia. All fossils documented here are from a single measured stratigraphic section, named Khuhtolgoi (Fig. 1; 50°08’31.6” N, 100°04’47.7” E). The section intersects an interval within the Erkhelnuur Formation of the Khubsugul Group. The Erkhelnuur Formation is about 2000 m thick in this region and is composed of alternating limestone and dolostone beds (Anttila et al. 2021). The base of the 131.9 m thick (true thickness) Khuhtolgoi section is characterised by dark-grey limestone and dolostone beds that transitions to dark-grey micritic limestone. Trilobite fragments appear at 87.74 m above the base of the section followed by archaeocyathids at 128.04 m. The presence of trilobites and archaeocyathids suggest correlation with the recently defined Middle Member of the Erkhelnuur Formation (Antilla et al. 2021). However, as the Khuhtolgoi section is fault bounded at both bottom and top, its exact stratigraphic position within the Erkhelnuur Formation cannot be precisely determined.
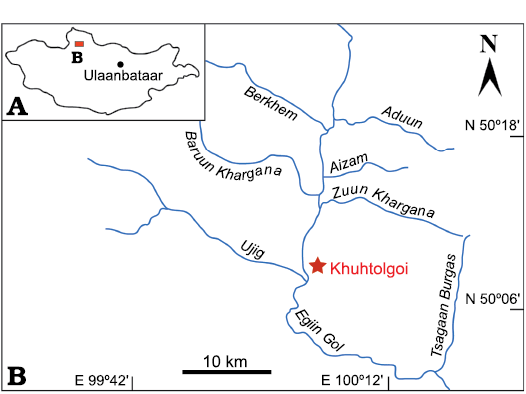
Fig. 1. A. Map of Mongolia showing location of the studied section. B. Precise location of the section Khuhtolgoi.
The isolated palaeoscolecid plates have been recovered from six samples situated 121.84–130 m above the base of the Khuhtolgoi section. The age of the section is likely to represent the unnamed Cambrian Series 2, Stage 3 and can be correlated with the Atdabanian stage of Siberia (Dorjnamjaa and Altanshagai 2015; Korovnikov and Lazarev 2021). The presence of Rhombocorniculum cancellatum (Cobbold, 1921) suggests correlation with upper part of Cambrian Stage 3 (Steiner et al. 2007; Topper et al. 2007, 2009; Wotte and Sundberg 2017; Korovnikov et al. 2018; Wallet et al. 2022).
To extract fossils from the rock, limestone samples were washed and placed in a solution of 5% acetic acid for several days. Acetic residues were sieved, dried, and the faunal elements were hand-picked from the residue under a binocular microscope. Selected specimens were set on a stub and gold coated, to be analysed with a Quanta 650 FEG SEM housed at the NRM, using an accelerating voltage of 15 kV. In this study, a total of 260 plates were found and 80 were selected for morphological analysis and measurements. Measurements were obtained using the tools available in the SEM during the imaging process. All measurements (see SOM, Supplementary Online Material available at http://app.pan.pl/SOM/app68-Goni_etal_SOM.pdf) represent the mean diameter. The material is stored at the NRM, in the Department of Palaeobiology.
The morphological terminology used herein predominantly follows that of Topper et al. (2010) and Whitaker et al. (2020). Palaeoscolecid plates are generally characterized by an ornamented outer side and a smooth inner side. The outer side of the plates are typically ornamented by a series of protuberances, referred to as nodes that can vary widely in morphology and display a range of configurations. The plates of Floraconformis egiinensis gen. et sp. nov. however, do not possess nodes, but are instead characterized by a depression network, that covers the outer side. The depression network defines a maximum of three categories of elevations: a large central elevation, smaller second-generation elevations, and third-generation elevations (Fig. 2). A brim represents the peripheral region of the plate but is only present when the network does not reach the edge of the plate.
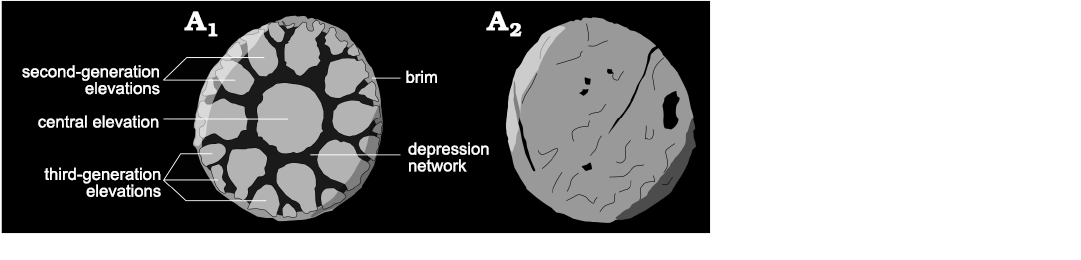
Fig. 2. Schematic drawing of palaeoscolecid Floraconformis egiinensis gen. et sp. nov. plate showing the terminology used, based on specimen NRM X11497; outer (A1) and inner (A2) side.
Systematic palaeontology
Superphylum Ecdysozoa Aguinaldo, Turbeville, Linford, Rivera, Garey, Raff, and Lake, 1997
Phylum uncertain
Clade Palaeoscolecidomorpha Shi, Howard, Edgecombe, Hou, and Ma, 2022
Class Palaeoscolecida Conway Morris and Robison, 1986
Order uncertain
Family ?Palaeoscolecidae Whittard, 1953
Genus Floraconformis nov.
ZooBank LSID: urn:lsid:zoobank.org:act:C99D6FB6-64A2-49A4-8281-5002BAD9A925
Etymology: From Latin flora, flower and conformis, like; referring to the flower-like shape of plates.
Type species: Floraconformis egiinensis gen. et sp. nov. by monotypy; see below.
Diagnosis.—As for the type species.
Remarks.—The isolated plates described here are referred to the Palaeoscolecida based on the similarities to dermal plates of known palaeoscolecid worms (Müller and Hinz-Schallreuter 1993; Topper et al. 2010; Botting et al. 2012). Typical features of palaeoscolecid plates are present, like a phosphatic composition, circular to elliptical general shape, possessing one ornamented side, a porous core and a hyaline outer layer. As such it is reasonable to include these Mongolian fossils within the Palaeoscolecida.
Because Floraconformis egiinensis gen. et sp. nov. plates are not tuberculate it cannot readily be accommodated within the diagnosis of the family Palaeoscolecidae as proposed by Botting et al. (2012). Elevations on Floraconformis egiinensis gen. et sp. nov. plates described here are formed from the extension of the depression network. In contrast, plates belonging to the species of Hadimopanella Gedik, 1977, Milaculum Müller, 1973, Kaimenella Märss, 1988, and Utahphospha Müller and Miller, 1976, have elevations that are extensions of an otherwise solid surface. However, in other aspects, the plates fit well within the range of plate morphologies documented from the family and Floraconformis egiinensis gen. et sp. nov. is here regarded as a likely member of the Palaeoscolecidae. This assignment is supported by the presence of individual plates lacking ornamentation across a number of formally described palaeoscolecid genera, such as Scathascolex Smith, 2015, and Utahscolex Whitaker, Jamison, Schiffbauer, and Kimmig, 2020, that have been assigned to the Palaeoscolecidae based on the presence of features such as an annulated cuticle, that can only be observed on body fossils. The wide variety of ornamentation (or lack thereof) on the surface of plates seen across members of the family indicates that the definition of the family may need to be revised to better incorporate the morphological variability observed in isolated phosphatic plates.
The plates described herein are morphologically distinct from previously described palaeoscolecid plates, including those belonging to species of Hadimopanella, Milaculum, Kaimenella, or Utahphospha. All of these form genera bear nodes on their convex outer surface, whereas the elevations on Floraconformis egiinensis gen. et sp. nov. plates are created by the stellate depression network. The top surfaces of elevations are concave. Moreover, plates belonging to species of Milaculum and Kaimenella are elongated ovals and not circular as in Floraconformis egiinensis gen. et sp. nov. The only known palaeoscolecid plates that bear some resemblance to those of Floraconformis egiinensis gen. et sp. nov. are that of a three-dimensionally preserved specimen from Australia (1 km north of Mt Murray, Duchess, Queensland; upper Templetonian stage in the Australian scheme; correlated with upper Wuliuan) described by Müller and Hinz-Schallreuter (1993) as Palaeoscolecida gen. et sp. indet. The single published image of this specimen shows a section through the cuticle, parallel to the surface and small plates are visible (Müller and Hinz-Schallreuter 1993: text-fig. 17D). The plates are circular, with a central delineated elevation and smaller elevations arranged around the central one, apparently separated by furrows. Unfortunately, the plates are lacking a detailed description, preventing precise comparison to Floraconformis egiinensis gen. et sp. nov. It is difficult to evaluate if this Australian specimen can be accommodated into the newly introduced genus Floraconformis. Alternatively, the specimen may represent a related taxon, but the lack of information regarding the Australian material prevents a definitive conclusion.
Floraconformis gen. nov. should be considered as a form genus (like Hadimopanella, Milaculum, Kaimenella, and Utahphospha) pending further analyses, as neither articulated material nor microplates have been found in the Mongolian material.
Stratigraphic and geographic range.—Cambrian Series 2, Stage 3; Khuhtolgoi section, northern Mongolia.
Floraconformis egiinensis gen. et sp. nov.
Figs. 3–7.
ZooBank LSID: urn:lsid:zoobank.org:act:7B85B938-9948-490A-B1F5-78649C6076AC
Etymology: For the river Egiin Gol that flows near the section and the type locality.
Holotype: NRM X11462; well preserved plate (Fig. 3).
Type locality: Khuhtolgoi, along the Egiin Gol river, south of Khubsugul Lake, northern Mongolia.
Type horizon: Samples Khu1-141.8 to Khu1-167.2, upper Erkhelnuur Formation, Khubsugul Group, Cambrian Series 2, Stage 3.
Material.— Holotype: NRM X11462; paratypes: NRM 11463, 11465, 11467, 11489–11535, 11554, 11555. Type material only.
Diagnosis.—Circular to slightly elliptical, biconvex, phosphatic plates. The outer side displays a stellate depression network that creates a series of elevations. The outer face of each elevation is slightly concave. The central elevation is the largest elevation and can occupy up to 30% of the entire width of the plate. A second large circular elevation is delimited from the brim of the plate and is typically divided into one or two concentric rings of smaller elevations by the depression network. Small tubercles are present on the brim when the outer hyaline layer is preserved.
Description.—All plates are circular to slightly elliptical in outline (Figs. 3, 4), with a diameter ranging from 98 to 233 µm (mean 147 µm, n = 80; see SOM). Both inner and outer sides are convex. The surface of the inner side is usually smooth, although a fingerprint-like structure can be observed in a few specimens (Fig. 5A). The outer side displays various ornamentations based on the formation of the depression network; a deeply incised network of furrows separates a series of elevations (Figs. 3, 4). This network demarcates a central circle with radiating and bifurcating rays, which result in a stellate appearance. Morphological variation of the stellate pattern depends predominantly on the development of the depression network. In some samples, a wide marginal brim can be observed when the depression network is focused in the middle of the plate (Fig. 4O), but in others, the brim is narrow or absent when the depression network reaches the margin of the plate (Fig. 4A). The depression network defines a variety of elevations that range from circular to oval in shape, with a slightly concave top surface (Figs. 3, 6). The top of the elevations is usually wider than in the middle, giving the impression that the margins of the elevations slightly overhang the depressions (Figs. 3, 6).
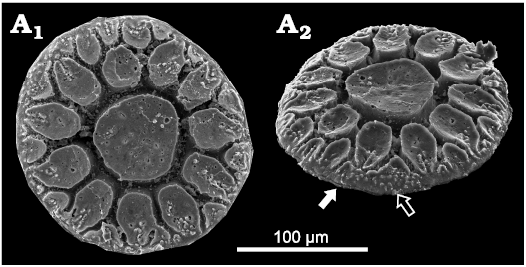
Fig. 3. Palaeoscolecid Floraconformis egiinensis gen. et sp. nov., plate from Khuhtolgoi section, northern Mongolia, Cambrian Stage 3. Holotype NRM X11462 in dorsal (A1) and dorso-lateral (A2) views. The white arrow shows the small gap present on few plates, at the bound between inner and outer sides. The black arrow shows small tubercles that are present on the brim, when the outer hyaline layer is preserved.
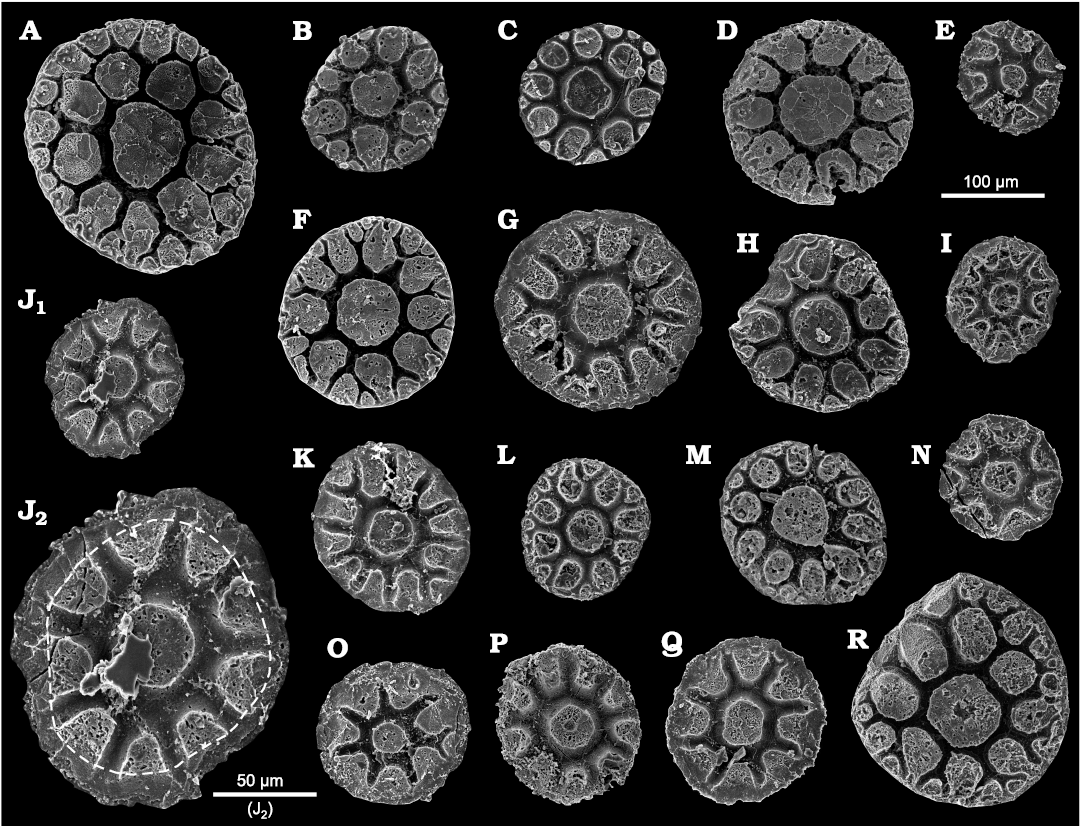
Fig. 4. Palaeoscolecid Floraconformis egiinensis gen. et sp. nov., plates (in dorsal view) from Khuhtolgoi section, northern Mongolia, Cambrian Stage 3. A. NRM X11465. B. NRM X11493. C. NRM X11502. D. NRM X11492. E. NRM X11489. F. NRM X11497. G. NRM X11495. H. NRM X11521. I. NRM X11508. J. NRM X11505 in general (J1) and close-up (J2) views; the dashed line in J2 represents the second large elevation delimited from the brim mentioned in the diagnosis. K. NRM X11500. L. NRM X11513. M. NRM X11499. N. NRM X11490. O. NRM X11496. P. MNRM X11491. Q. NRM X11523. R. NRM X11463.
There is almost always a single central circular to elliptical elevation that is larger than the surrounding elevations. This central elevation occupies approximately 30% of the entire plate width (see SOM). The central elevation is surrounded by a ring of between 6 and 11 second-generation elevations that are delimited from the brim by a step (Fig. 4J2; see also Fig. 4O, R). The second-generation elevations are smaller than the central elevation and have two main characteristics: (i) they are disposed radially around and adjacent to the central elevation, forming a ring and (ii) they are usually elongated toward the margin of the plate. Sometimes a third generation of elevations (formed between the second-generation elevations as the depression network bifurcates towards the edge) are visible closer to the edge of the plate (Figs. 3, 4B). These have a similar shape to the other elevations but are smaller. Total number of elevations varies up to 33, depending on the development of the depression network (which may be variable across the plate). The space between the top of the elevations (or the width of the depression network) is also highly variable and ranges 7.6–21.6 µm (measurements in plan view).
Two kinds of textures can be observed on the plate surface: a thin hyaline outer layer, which covers a porous core. The hyaline external layer is predominantly smooth, but infrequently it bears some small tubercles on the brim (Fig. 3) or on the most external elevations (Fig. 4A). Generally, the hyaline layer is not preserved, revealing the porous core’s fibrous structure (Fig. 6). Around the circumference of the plate at the junction between the inner and the outer sides, a furrow of a few micrometres can be observed (Fig. 3: white arrow). This furrow is observed on plates, whether the hyaline layer is preserved or not.
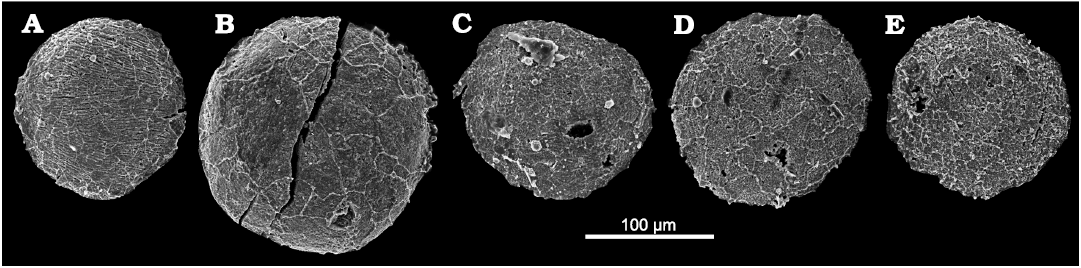
Fig. 5. Palaeoscolecid Floraconformis egiinensis gen. et sp. nov., plates (in ventral view) from Khuhtolgoi section, northern Mongolia, Cambrian Stage 3. A. NRM X11516; note the fingerprint-like ornamentation on this plate. B. NRM X11522. C. NRM X11498. D. NRM X11518. E. NRM X11501.
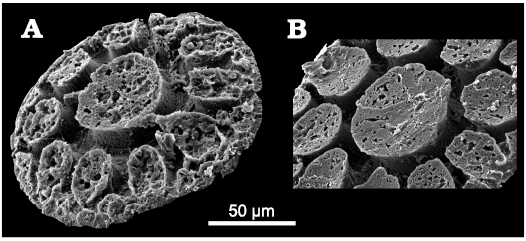
Fig. 6. Fibrous structures observed on elevations on the dorsal sides of palaeoscolecid Floraconformis egiinensis gen. et sp. nov., plate from Khuhtolgoi section, northern Mongolia, Cambrian Stage 3. these structures are only visible when the outer hyaline layer is absent. A. NRM X11499. B. NRM X11497.
Although most plates conform to the general pattern described above, a few plates in the collection differ slightly. In particular, one specimen consists of two merged plates (Fig. 7A). In this specimen, one plate is slightly larger in diameter (134 vs. 108 µm), but the size of the elevations and the width of the depression network are approximately the same across both plates. The junction between the plates is a straight furrow, narrower but slightly deeper than the usual depression network. Usually, plates are isolated and circular (Fig. 4), without any deformation along the edge, suggesting that they were not arranged immediately next to each other in the cuticle. Even though this specimen is an isolated case among 260 specimens, it indicates that plates could be formed in direct association with each other within the cuticle. In another unusual specimen (Fig. 7B), second-generation elevations are not elongated radially toward the edge of the plate, but are elongated parallel to the edge around the central elevation. A third unusual type of plate has no well-defined large central elevation (Fig. 7C). In this specimen, there are two approximately central elevations that are slightly larger than the other elevations.
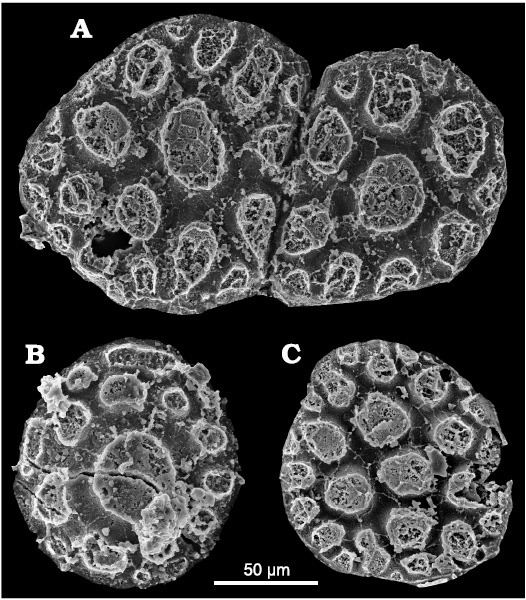
Fig. 7. Unusual plates of palaeoscolecid Floraconformis egiinensis gen. et sp. nov. from Khuhtolgoi section, northern Mongolia, Cambrian Stage 3. A. NRM X11467; two merged plates. B. NRM X11524; second and third-generation elevations are not elongated toward the margin but parallel to it. C. NRM X11510; absence of the usual larger central elevation.
Remarks.—Articulated-body preservation of Cambrian palaeoscolecids reveals that plates across a range of morphologies can occur in the scleritome of a single animal (Müller and Hinz-Schallreuter 1993; Ivantsov and Wrona 2004; Topper et al. 2010; Smith 2015; Streng et al. 2017). The development of different plate morphologies in palaeoscolecid scleritomes may be related to particular locations on the body, environmental influences, or different developmental stages (Brock and Cooper 1993; Müller and Hinz-Schallreuter 1993; Harvey et al. 2010; Barragán et al. 2014a). Floraconformis egiinensis gen. et sp. nov. from Mongolia is known exclusively from isolated plates but these show a distinctive type of plate morphology and ornamentation (Fig. 4). The characteristic depression network that defines the concave elevations differentiates Floraconformis egiinensis gen. et sp. nov. from all other palaeoscolecid form taxa, and from plate morphologies of articulated taxa, with the possible exception of the plates of Palaeoscolecida gen. et sp. indet. from the Templetonian stage of Australia (correlated with Wuliuan; drilling Duchess 18.1 km north of Mt Murray, Queensland; Triplagnostus gibbus Zone; Müller and Hinz-Schallreuter 1993: text-fig. 17D). Although the morphology of the plates from Australia bears some similarities to that of Floraconformis egiinensis gen. et sp. nov., there are key differences. In particular, the plates from Australia are smaller (maximum diameter 70 µm) than the material from Mongolia described herein (minimum diameter 98 µm), and the elevations do not appear concave as in the Mongolian material.
According to Barragán et al. (2014a), the size of a palaeoscolecid plate is defined and fixed at its inception whilst the external ornamentation is secondary and variable. Articulated palaeoscolecid taxa typically bear plates that are between 20 and 60 µm in diameter (Müller and Hinz-Schallreuter 1993; Topper et al. 2010; Botting et al. 2012; Wang et al. 2014; Streng et al. 2014). However, larger plates comparable to those reported herein have been recovered from acid isolated plate assemblages. Two form species, Hadimopanella knappologica (Bengtson 1977) and Hadimopanella oezguli Gedik, 1977 have comparable or larger plate diameters of 460 and 400 µm maximum, respectively (Ivantsov and Wrona 2004; Kouchinsky et al. 2015; Barragán et al. 2014a). The size of Floraconformis egiinensis gen. et sp. nov. falls within the overall size range of Cambrian palaeoscolecid isolated plates. Further distinguishing characteristics include the presence of third-generation elevations and a brim in Floraconformis egiinensis gen. et sp. nov., not seen on the Australian plates. The palaeoscolecid specimen from Australia is also significantly younger (late Templetonian in the Australian scheme; correlated with late Wuliuan) than Floraconformis egiinensis gen. et sp. nov. from Mongolia (Atdabanian Stage in the Siberian scheme; correlated with Cambrian Stage 3). Pending additional material from Australia for comparison, we conclude that while Palaeoscolecida gen. et sp. indet. of Müller and Hinz-Schallreuter (1993) may be a closely related taxon, it is unlikely to be conspecific with Floraconformis egiinensis gen. et sp. nov. from Mongolia.
Stratigraphic and geographic range.—Type horizon and locality only.
Discussion
In addition to Floraconformis egiinensis gen. et sp. nov., two fragmentary fossil specimens (Fig. 8), morphologically similar to species of the genus Stoibostrombus Conway Morris and Bengtson in Bengtson et al., 1990, are present 167.2 m above the base of the section. Although the biological affinity of Stoibostrombus remains unclear, the taxon has tentatively been affiliated with palaeoscolecid worms (Skovsted et al. 2011). The morphology of the Mongolian fragments fits with the diagnosis of the family Stoibostrombidae Conway Morris and Bengtson, 1990, however, the basal ornamentation is distinctive. The basal ornament consists of separate, small, button-shaped elements with concave top surfaces similar to the elevations in Floraconformis egiinensis gen. et sp. nov. A thin hyaline layer covers these elements (Fig. 8A3), under which the support is porous (Fig. 8B3). The differences in basal ornament indicates that these fragmentary cones from Mongolia do not conform to the type and only species, Stoibostrombus crenulatus Conway Morris and Bengtson in Bengtson et al., 1990 (Bengtson et al. 1990: figs. 93–98). We instead suggest that they may be sclerites associated with Floraconformis egiinensis gen. et sp. nov. or a similar taxon, possibly representing the aboral spines that are preserved on some palaeoscolecid specimens (Müller and Hinz-Schallreuter 1993: text-fig. 11; Duan et al. 2012: fig. 2).
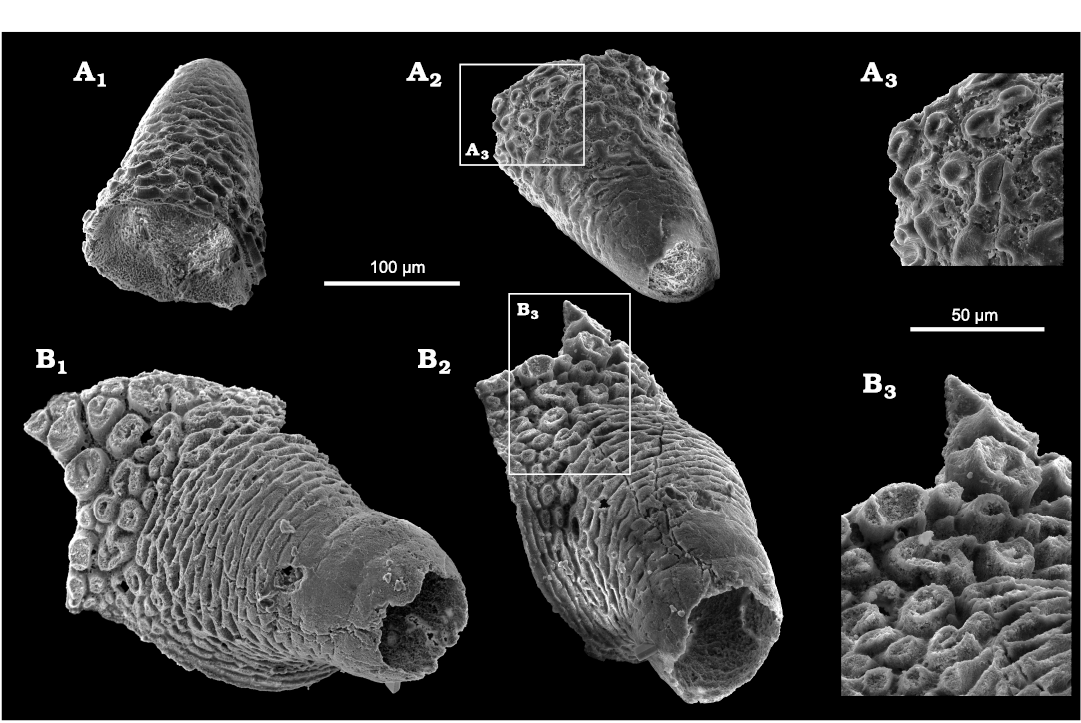
Fig. 8. Two conical fossils regarded as palaeoscolecid fragments from Khuhtolgoi section, northern Mongolia, Cambrian Stage 3. A. NRM X11556 in latero-abapical (A1) and latero-apical (A2) views, with a close-up on button-shaped elements (A3). B. NRM X11556 in latero-apical (B1, B2) views, with a close-up on button-shaped elements (B3).
Conclusions
Floraconformis egiinensis gen. et sp. nov. is the first palaeoscolecid recorded from northern Mongolia. Despite recovery as isolated plates, specimens of Floraconformis egiinensis gen. et sp. nov. are morphologically unique and can therefore be used to designate a new species. This contributes to the diversity of Cambrian palaeoscolecid form taxa, of which only members of four genera have been previously described: Hadimopanella Gedik, 1977, Milaculum Müller, 1973, Kaimenella Märss, 1988, and Utahphospha Müller and Miller, 1976. Despite significant faulting in the region, Floraconformis egiinensis gen. et sp. nov. can be confidently correlated with Cambrian Stage 3 and therefore represents one of the oldest reports of isolated palaeoscolecid plates in the fossil record.
Acknowledgements
Thanks are given to our field chef Usukhbayar and our Mongolian drivers Bayarsaikhan and Batkhuyag for assistance in the field. We also thank Julien Kimmig (Department of Geosciences, University Park, USA), Thomas Wotte (Department of Palaeontology, Freiberg, Germany), and the handling editors, for their constructive comments regarding the paper. The authors acknowledge financial support from the Swedish Research Council to T.P.T (VR2017-05183) and C.B.S. (VR2016-04610, VR2021- 04295), a Young Thousand Talents Plan of China and National Natural Science Foundation of China to TPT (42072003) and GL (41890844). MJB is supported by research funds from Northwest University, Xi’an and the University of New England, Australia.
References
Aguinaldo, A.M., Turbeville, J.M., Linford, L.S., Rivera, M.C., Garey, J.R., Raff, R.A., and Lake, J.A. 1997. Evidence for a clade of nematodes, arthropods and other moulting animals. Nature 387: 489–493. Crossref
Anttila, E., Macdonald, F., and Bold, U. 2021. Stratigraphy of the Khuvsgul Group, Mongolia. Mongolian Geoscientist 26: 2–15. Crossref
Barragán, T., Esteve, J., García-Bellido, D.C., Zamora, S., and Álvaro, J.J. 2014a. Hadimopanella oezgueli Gedik, 1977: a palaeoscolecidan plate useless for taxonomic purposes. Palaeontologia Electronica 17: 1–20. Crossref
Barragán, T., Esteve, J., García-Bellido, D.C., Zamora, S., and Álvaro, J.J. 2014b. New Middle Cambrian palaeoscolecid plates of Hadimopanella oezgueli from the Cantabrian Mountains, northern Spain. GFF 136: 22–25. Crossref
Bengtson, S. 1977. Early Cambrian button-shaped phosphatic microfossils from the Siberian Platform. Palaeontology 20: 751–762.
Bengtson, S., Conway Morris, S., Cooper, B.J., Jell, P.A., and Runnegar, B.N. 1990. Early Cambrian fossils from South Australia. Memoirs of the Association of Australasian Palaeontologists 9: 1–364.
Botting, J.P., Muir, L.A., Van Roy, P., Bates, D., and Upton, C. 2012. Diverse middle Ordovician palaeoscolecidan worms from the Builth‐Llandrindod Inlier of central Wales. Palaeontology 55: 501–528. Crossref
Brock, G.A., and Cooper, B.J. 1993. Shelly fossils from the Early Cambrian (Toyonian) Wirrealpa, Aroona Creek, and Ramsay Limestones of South Australia. Journal of Paleontology 67: 758–787. Crossref
Cobbold, E.S. 1921. The Cambrian horizons of Comley (Shropshire) and their Brachiopoda, Pteropoda, Gasteropoda and etc. The Quarterly Journal of the Geological Society of London 76: 325–386. Crossref
Conway Morris, S. and Peel, J.S. 2010. New palaeoscolecidan worms from the Lower Cambrian: Sirius Passet, Latham Shale and Kinzers Shale. Acta Palaeontologica Polonica 55: 141–156. Crossref
Conway Morris, S. and Robison, R.A. 1986. Middle Cambrian priapulids and other soft-bodied fossils from Utah and Spain. The University of Kansas Paleontological Contributions 117: 1–22.
Dorjnamjaa, D. and Altanshagai, G. 2015. Concerning the original viewpoint of biogeologic accumulation of the old bedded phosphorites in the Khubsugul and Zavkhan basins of Mongolia. Open Journal of Geology 5: 666. Crossref
Duan, B., Dong, X.P., and Donoghue, P.C. 2012. New palaeoscolecid worms from the Furongian (upper Cambrian) of Hunan, South China: Is Markuelia an embryonic palaeoscolecid? Palaeontology 55: 613–622. Crossref
García-Bellido, D.C., Paterson, J.R., and Edgecombe, G.D. 2013. Cambrian palaeoscolecids (Cycloneuralia) from Gondwana and reappraisal of species assigned to Palaeoscolex. Gondwana Research 24: 780–795. Crossref
Gedik, I. 1977. Conodont stratigraphy in the Middle Taurus [in Turkish with English abstract]. Bulletin of the Geological Society of Turkey 20: 35–48.
Harvey, T.H.P., Dong, X.P., and Donoghue, P.C.J. 2010. Are palaeoscolecids ancestral ecdysozoans? Evolution and Development 12: 177–200. Crossref
Ivantsov, A.Y. and Wrona, R. 2004. Articulated palaeoscolecid plate arrays from the Lower Cambrian of eastern Siberia. Acta Geologica Polonica 54: 1–22.
Korovnikov, I.V. and Lazarev, F.D. 2021. New Findings of Lower Cambrian Trilobites in the Southern Hövsgöl Area (Mongolia) and Their Significance for Stratigraphy and Interregional Correlation. Russian Geology and Geophysics 62: 1350–1358. Crossref
Korovnikov, I.V., Novozhilova, N.V., and Tokarev, D.A. 2018. Distribution of Atdabanian (Early Cambrian) Trilobites, Archaeocyaths, and Small Shelly Fossils in the Altai-Sayan Folded Area. Paleontological Journal 52: 1481–1493. Crossref
Kouchinsky, A., Bengtson, S., Clausen, S., and Vendrasco, M.J. 2015. An early Cambrian fauna of skeletal fossils from the Emyaksin Formation, northern Siberia. Acta Palaeontologica Polonica 60: 421–512.
Kouraiss, K., El Hariri, K., El Albani, A., Azizi, A., Mazurier, A., and Vannier, J. 2018. X-ray microtomography applied to fossils preserved in compression: Palaeoscolescid worms from the Lower Ordovician Fezouata Shale. Palaeogeography, Palaeoclimatology, Palaeoecology 508: 48–58. Crossref
Leibach, W.W., Lerosey-Aubril, R., Whitaker, A.F., Schiffbauer, J.D., and Kimmig, J. 2021. First palaeoscolecid from the Cambrian (Drumian, Miaolingian) Marjum Formation of western Utah, USA. Acta Palaeontologica Polonica 66: 663–678. Crossref
Märss, T. 1988. Early Palaeozoic hadimopanellids of Estonia and Kirgizia (USSR). Proceedings of Sciences of the Estonian SSR. Geology 37: 10–17. Crossref
Müller, K.J. 1973. Milaculum n. gen., ein phosphatisches Mikrofossil aus dem Altpalaeozoikum. Paläontologische Zeitschrift 47: 277–228. Crossref
Müller, K.J. and Hinz-Schallreuter, I. 1993. Palaeoscolecid worms from the Middle Cambrian of Australia. Palaeontology 36: 549–592.
Müller, K.J. and Miller, J.F. 1976. The problematic microfossil Utahphospha from the Upper Cambrian of the western United States. Lethaia 9: 391–395. Crossref
Shi, X., Howard, R.J., Edgecombe, G.D., Hou, X., and Ma, X. 2022. Tabelliscolex (Cricocosmiidae: Palaeoscolecidomorpha) from the early Cambrian Chengjiang Biota, and the evolution of seriation in Ecdysozoa. Journal of the Geological Society 179: jgs2021-060. Crossref
Skovsted, C.B., Brock, G.A., and Topper, T.P. 2011. Plate fusion in the problematic early Cambrian spine-like fossil Stoibostrombus from South Australia. Bulletin of Geosciences 86: 651–658. Crossref
Smith, M.R. 2015. A palaeoscolecid worm from the Burgess Shale. Palaeontology 58: 973–979. Crossref
Smith, M.R. and Dhungana, A. 2022. Discussion on “Tabelliscolex (Cricocosmiidae: Palaeoscolecidomorpha) from the early Cambrian Chengjiang Biota and the evolution of seriation in Ecdysozoa” by Shi et al. 2021. Journal of the Geological Society 179: jgs2021-111. Crossref
Steiner, M., Li, G., Qian, Y., Zhu, M., and Erdtmann, B.D. 2007. Neoproterozoic to early Cambrian small shelly fossil assemblages and a revised biostratigraphic correlation of the Yangtze Platform (China). Palaeogeography, Palaeoclimatology, Palaeoecology 254: 67–99. Crossref
Streng, M., Ebbestad, J.O.R., and Berg‐Madsen, V. 2017. Cambrian palaeoscolecids (Cycloneuralia) of southern Scandinavia. Papers in Palaeontology 3: 21–48. Crossref
Topper, T.P., Brock, G.A., Skovsted, C.B., and Paterson, J.R. 2009. Shelly Fossils from the Lower Cambrian Pararaia bunyerooensis Zone, Flinders Ranges, South Australia. Memoirs of the Association of Australasian Palaeontologists 37: 199–246.
Topper, T.P., Brock, G.A., Skovsted, C.B., and Paterson, J.R. 2010. Palaeoscolecid scleritome fragments with Hadimopanella plates from the early Cambrian of South Australia. Geological Magazine 147: 86–97. Crossref
Topper, T.P., Skovsted, C.B., Brock, G.A., and Paterson, J.R. 2007. New bradoriids from the lower Cambrian Mernmerna Formation, South Australia: systematics, biostratigraphy and biogeography. Memoirs of the Association of Australasian Palaeontologists 33: 67–100. Crossref
Wallet, E., Padel, M., Devaere, L.,
Clausen, S., Álvaro, J.J., and Laumonier, B. 2022. Cambrian Age 3
small shelly fossils from the Terrades inlier, southern Pyrenees,
Spain: Biostratigraphic and paleobiogeographic implications. Journal
of Paleontology 96: 552-582. Crossref
Wang, W., Muir, L.A., Botting, J.P., Feng, H., Servais, T., and Li, L. 2014. A Tremadocian (Early Ordovician) palaeoscolecidan worm from graptolitic shales in Hunan Province, South China. Palaeontology 57: 657–671. Crossref
Wendruff, A.J., Babcock, L.E., Kluessendorf, J., and Mikulic, D.G. 2020. Paleobiology and taphonomy of exceptionally preserved organisms from the Waukesha Biota (Silurian), Wisconsin, USA. Palaeogeography, Palaeoclimatology, Palaeoecology 546: 109631. Crossref
Whitaker, A.F., Jamison, P.G., Schiffbauer, J.D., and Kimmig, J. 2020. Re-description of the Spence Shale palaeoscolecids in light of new morphological features with comments on palaeoscolecid taxonomy and taphonomy. PalZ 94: 661–674. Crossref
Whittard, W.F. 1953. Palaeoscolex piscatorum gen. et sp. nov., a worm from the Tremadocian of Shropshire. Quarterly Journal of the Geological Society 109: 125–135. Crossref
Wills, M.A., Gerber, S., Ruta, M., and Hughes, M. 2012. The disparity of priapulid, archaeopriapulid and palaeoscolecid worms in the light of new data. Journal of Evolutionary Biology 25: 2056–2076. Crossref
Wotte, T. and Sundberg, F. A. 2017. Small shelly fossils from the Montezuman-Delamaran of the Great Basin in Nevada and California. Journal of Paleontology 91: 883–901. Crossref
Acta Palaeontol. Pol. 68 (1): 117–125, 2023
https://doi.org/10.4202/app.01030.2022