
First Jurassic brittlestar from Neuquén Basin, Argentina
DÉBORA M. CAMPETELLA, EVANGELINA E. PALÓPOLO, MAXIMILIANO N. RODRÍGUEZ, BEN THUY, JUAN J. PONCE, NOELIA B. CARMONA, and SILVIO CASADÍO
Campetella, D.M., Palópolo, E.E., Rodríguez, M.N., Thuy, B., Ponce, J.J., Carmona, N.B., and Casadío, S. 2023. First Jurassic brittlestar from Neuquén Basin, Argentina. Acta Palaeontologica Polonica 68 (1): 143–153.
Articulated fossil ophiuroids from South America were reported for the Devonian, Cretaceous, Eocene, and Miocene. Here we report the first Jurassic record of an articulated ophiuroid from the Sierra Chacaicó Formation (early Pliensbachian–Sinemurian) in Neuquén Basin, Argentina, and discuss the taphonomic processes that allowed its preservation. The Sierra Chacaicó Formation represents the onset of the Early Jurassic extensive marine transgression in the basin. The basal section comprises shoreface and offshore Gilbert-type delta system, which was affected by hyperpycnal discharges. The middle and upper sections are represented by offshore deposits, affected by storms and eroded by hyperpycnal channel-levee systems. The ophiuroid specimen was found in levels of massive, fine, tuffaceous sandstone beds and covered by coarse sandstone containing a large amount of plant debris and organic matter. It was preserved articulated, with a complete disc and almost complete arms. Based on the microstructure of the spine-bearing lateral arm plates, the ophiuroid is assigned to Sinosura, an extinct genus of the family Ophioleucidae, widespread in the Lower Jurassic deposits of Europe but previously unknown from other parts of the world. The posture of the ophiuroid, with one arm curved distally and extended in one direction and the other four arms symmetrically oriented in the opposite direction, suggests a walking or escape movement frozen in time. This implies that the ophiuroid was buried alive by sediment thick enough to prevent successful escape. The taphonomic and sedimentologic evidence indicates that the fossil material was found in hyperpycnal deposits accumulated in offshore positions, which carried a high concentration of sediment in suspension.
Key words: Echinodermata, Ophiuroidea, Sinosura, Pliensbachian, Sierra Chacaicó Formation, Neuquén Basin, Argentina.
Débora M. Campetella [dmcampetella@unrn.edu.ar; ORCID: https://orcid.org/0000-0002-0763-3420 ], Universidad Nacional de Río Negro, Estados Unidos 750, (R8332EXZ) General Roca, Río Negro, Argentina.
Evangelina E. Palópolo [eepalopolo@unrn.edu.ar; ORCID: https://orcid.org/0000-0002-1272-5637 ], Maximiliano N. Rodriguez [mnrodriguez@unrn.edu.ar; ORCID: https://orcid.org/0000-0003-0286-0111 ], Noelia B. Carmona [ncarmona@unrn.edu.ar; ORCID: https://orcid.org/0000-0003-0923-3719 ], and Silvio Casadío [scasadio@unrn.edu.ar; ORCID: https://orcid.org/0000-0002-8130-641X ], Universidad Nacional de Río Negro, General Roca, Río Negro, Argentina; Instituto de Investigación en Paleobiología y Geología, Río Negro, Argentina; Consejo Nacional de Investigaciones científicas y Tecnológicas (CONICET), Av. Roca 1242, (R8332EXZ) General Roca, Río Negro, Argentina.
Ben Thuy [ben.thuy@mnhn.lu; ORCID: https://orcid.org/0000-0001-8231-9565 ], Department of Paleontology, Research & Collections, Natural History Museum Luxembourg. 25, rue Münster, L-2160 Luxembourg.
Juan J. Ponce [jponce@unrn.edu.ar; ORCID: https://orcid.org/ 0000-0003-4148-5668 ], Servicio Geológico Minero Argentino (SEGEMAR), Centro General Roca, Río Negro, Argentina.
Received 12 October 2022, accepted 15 December 2022, available online 8 February 2023.
Copyright © 2022 D.M. Campetella et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Preservation potential of articulated echinoderms depends on their body plan, the nature of their multi-elemental skeleton, the coelomic cavity extension and the strength of ossicle articulations (Ausich et al. 2001; Brusca and Brusca 2003). Additionally, environmental parameters (such as salinity, oxygenation, sediment supply, presence of scavengers, etc.), and taphonomic processes (e.g., disarticulation, transport, fragmentation and bioerosion, among others) contribute to complete disintegration of specimens within hours to days (Brett et al. 1997; Nebelsick 2004).
Ophiuroids are characterized by a small, round disc and long, flexible arms. The ophiuroid skeleton consists of a multitude of millimeter-sized, calcitic ossicles. The arms are composed of a central ossicle called vertebrae, surrounded by dorsal, ventral, and lateral ossicles, the latter generally yielding spines. Due to the weak ossicle connections, ophiuroid skeletons generally disarticulate rapidly after death (Brett et al. 1997; Thuy et al. 2012; Reid et al. 2019).
As a result, ophiuroid remains are most commonly found as isolated skeletal ossicles in micropaleontological sieving residues (Thuy et al. 2020). Articulated fossil ophiuroids, in contrast, are exceptional fossils restricted to particular taphonomic windows such as “brittle star beds”, i.e., layers containing hundreds or thousands of specimens (Aronson 1989; Martínez et al. 2010). Occurrences of ophiuroids preserved intact are interpreted as a result of the combination of multiple factors: (i) little or no transport; (ii) rapid burial without subsequent reworking; (iii) anoxia or dysoxia; and (iv) occurrence in fine to very fine sediments (e.g., Seilacher 1976; Shroat-Lewis 2007; Zatoń et al. 2008; Reid et al. 2019).
Articulated fossil ophiuroids from South America were reported mainly from the Devonian (Ruedemann 1916; Haude 1995; Fraga and Vega 2020), Cretaceous (Fernández et al. 2019), Eocene (Furque and Camacho 1949; Malumián and Olivero 2005), and Miocene (Caviglia et al. 2007; Martínez and del Río 2008; Martínez et al. 2010). In the Neuquén Basin, ophiuroids are recorded in the Agua de la Mula Member of the Lower Cretaceous Agrio Formation, both as isolated ossicles and as articulated remains (Fernández et al. 2019). The present finding represents the oldest Mesozoic and the first Jurassic record of an ophiuroid from South America. The aim of the present publication is to describe a new record of an articulated ophiuroid from the Pliensbachian Sierra Chacaicó Formation, and to discuss its systematic position, and to interpret the taphonomic processes involved in its preservation.
Institutional abbreviations.—CONICET, Consejo Nacional de Investigaciones Científicas y Técnicas, Ciudad Autónoma de Buenos Aires, Argentina; IIPG, Instituto de Investigación en Paleobiología y Geología, General Roca, Río Negro, Argentina; MOZ-PI, Museo Oslacher de Zapala, Colección Paleoinvertebrados, Zapala, Neuquén Argentina; SEGEMAR, Servicio Geológico Minero Argentino, Centro General Roca, Río Negro, Argentina; UNRN, Universidad Nacional de Río Negro, General Roca, Río Negro, Argentina.
Other abbreviations.—EDS, Energy Dispersive Spectroscopy.
Geological setting
The Sierra de Chacaicó locality is situated in the central-western area of the Neuquén Basin, 50 km South from the town of Zapala (Fig. 1A). It represents an anticlinal structure, whose axis is oriented to the NNE and the associated deposits verge towards the west (Franzese et al. 2007). The age of the rocks exposed at Sierra de Chacaicó ranges from the late Paleozoic to the Early Cretaceous (Pángaro et al. 2009). The basement is composed of metamorphic rocks from Piedra Santa Formation (Devonian–upper Carboniferous) (Franzese 1995), and it is intruded by the Chachil Plutonic Complex (upper Carboniferous–lower Permian) (Leanza 1990; Leanza and Hugo 1997; Leanza et al. 2005; Pángaro et al. 2009). Syn-rift deposits comprise volcanic, pyroclastic and sedimentary rocks from the Lapa Formation (Fig. 1B, C) of Late Triassic–Early Jurassic (early Sinemurian) age (Leanza and Hugo 1997; Leanza et al. 2005; Franzese et al. 2007; Pángaro et al. 2009). The beginning of the post-rift deposition period is represented by the Sierra Chacaicó Formation (Fig. 1B, C) (Lower Jurassic; Sinemurian–lower Pliensbachian) which is composed of volcaniclastic beds deposited in shallow-marine environments (Gulisano and Gutiérrez Pleimling 1995; Leanza and Hugo 1997; Leanza et al. 2005; Pángaro et al. 2009; Ponce et al. 2015).
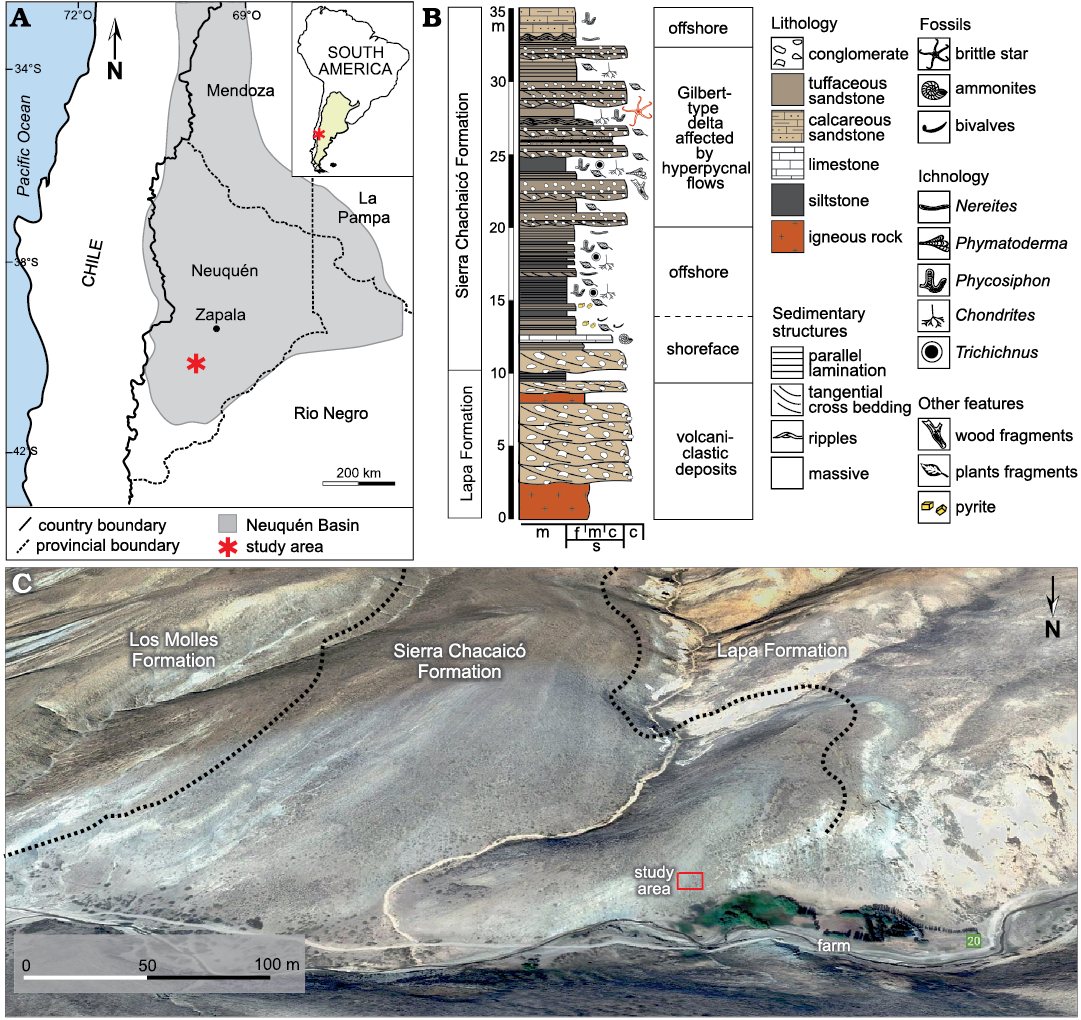
Fig. 1. Location and sedimentological section in the studied area. A. Location map of the Neuquén basin in west-central Argentina, showing the study area. B. Sedimentological section in the Arroyo Lapa locality, showing the massive levels where the ophiuroid specimen was collected. C. Satellite image of the Arroyo Lapa locality showing the boundaries of the Lapa, Sierra Chacaicó and Los Molles formations (Google Earth, access April 2022).
The Arroyo Lapa is a classic section (Fig. 1C) exposed at the margins of the homonymous stream, cutting out the South Margin of the Sierra de Chacaicó anticline structure. In this section, the oldest rocks of the Cuyo Group correspond to the Sierra Chacaicó Formation, representing the onset of the Early Jurassic extensive marine transgression in the Neuquén Basin (Volkheimer 1973; Leanza and Hugo 1997). The content of ammonites and radiolarians described in this area allowed assigning the Sierra Chacaicó Formation to the Pliensbachian (Hillebrandt 1981; Pujana 1996). In the basal section, this unit comprises a thick succession of calcareous and tuffaceous sandstone beds, which towards the top are intercalated with heterolites with abundant phytodetritus. These deposits are attributed to shoreface and offshore, Gilbert-type deltaic system affected by hyperpycnal flows (Carmona et al. 2016). The middle and upper sections are represented by offshore environments, reworked by storms and eroded by hyperpycnal flows, that produce channel-levee systems (Ponce et al. 2015; Carmona et al. 2016). The Sierra Chacaicó Formation is covered by a thick succession of tabular mudstone and sparse fine tuffaceous sandstone with tabular geometry, included in the Los Molles Formation (Weaver 1931) and interpreted as offshore-shelf deposits affected by storms and eroded by hyperpycnal flows (Volkheimer 1973; Volkheimer and Quattrocchio 1993; Gulisano and Gutiérrez Pleimling 1995; Ponce et al. 2015; Campetella 2022).
Material and methods
The fossil material was collected at the Arroyo Lapa locality (GPS coordinates: 39°23’21.6”S; 70°25’21.4”W), where a conventional sedimentological analysis was conducted, comprising measurement of the stratigraphic section and a detailed facies description. Integration of these data allowed inferring the depositional environment and the main taphonomic features found in this setting.
In particular, taphonomic attributes such as transport, disarticulation, replacement and recrystallization were evaluated. Mineralogical, textural and compositional description of the surrounding sediment was made based on a thin section perpendicular to stratification in a binocular microscope Nikon SMZ800.
The fossil was observed with a ZEISS®-AXIO
Imager A2m optical microscope, and photographed with AxioCamEL®.
Furthermore, detailed SEM images were taken using Zeiss® Scanning
Electronic Microscope, model Evo MA 15, and EDS studies were performed
in order to determine cement and framboidal surrounding structures.
Fossil measurements were taken with ZEN 2 core v.2.4 software.
Illustrations and figures were made using CorelDraw and Adobe
Illustrator software.
The studied ophiuroid was temporarily transported to the Instituto de Investigación en Paleobiología y Geología (IIPG) for identification and imaging and subsequently deposited at Museo Olsacher de Zapala under MOZ-PI-5930 number. Fossil study was authorized by Dirección Provincial de Patrimonio Cultural, Gobierno de la Provincia de Neuquén.
Results
The Sierra Chacaicó Formation, exposed at the Arroyo Lapa section, is characterized by a progradational sequence (Fig. 2A). The studied section begins with a fossiliferous limestone layer yielding ammonites and continues with siliciclastic deposits composed by an intercalation of massive, very fine, tuffaceous sandstone and siltstone beds, with parallel lamination, tangential cross-bedding and symmetrical ripples at the top of the beds (Table 1). These layers intercalate with tabular, massive, coarse sandstone to fine conglomerate (Fig. 2A), showing parallel lamination and cross-stratification structures in some cases (Table 1). The aforementioned strata bear numerous plants and phytodetritus (Fig. 2B). The biogenic structures associated with massive and laminated siltstone are dominated by the ichnospecies Chondrites intricatus Brongniart, 1823 (Fig. 2C), followed in abundance by Trichichnus isp. (Fig. 2D), Phycosiphon incertum Fischer-Ooster, 1858 (Fig. 2E), and Phymatoderma isp. (Fig. 2F; Table 1). On the other hand, in the massive, laminated and rippled sandstone deposits, Chondrites intricatus and Phycosiphon incertum are present, albeit Nereites isp. (Fig. 2G) is the dominant ichnogenus (Table 1). These trace fossils are found isolated and present low ichnodiversity and abundance. Interestingly, in finer deposits, the occurrence of pyrite framboids was also recorded.

Fig. 2. Outcrop characteristics of the Sierra Chacaicó section. A. Panoramic view of the prograding clinoforms of Gilbert-type delta affected by hyperpycnal discharge in Sierra Chacaicó Formation at Arroyo Lapa locality. B. Levels of tuffaceous sandstone facies with phytodetritus towards the top (arrows). C. Chondrites intricatus Brongniart, 1823 (Ch) in massive siltstone. D. Trichichnus isp. (Tr) at the top of very fine massive tuffaceous sandstone. E. View in section of Phycosiphon incertum Fischer-Ooster, 1858 (Ph) in very fine massive tuffaceous sandstone. F. View in section of Phymatoderma isp. (Phm) in massive siltstone. G. Nereites isp. (Ne) at the top of medium tuffaceous sandstone.
Table 1. Facies table exposing the main sedimentological, ichnological and depositional features.
|
Facies |
Lithology |
Sedimentary structures |
Ichnogenera |
Paleoenvironmental interpretation |
|
F1 |
siltstone |
massive |
Chondrites, Phycosiphon, Phymatoderma, Trichichnus |
flocculation through hyperpycnal plume |
|
F2 |
siltstone |
parallel |
Chondrites, Phycosiphon, Trichichnus |
precipitation through marine water column or precipitation through hypopycnal plume |
|
F3 |
limestone |
massive |
– |
precipitation from saturated
sea water in absence |
|
F4 |
very fine to fine |
massive |
Chondrites, Phycosiphon |
rapid loss of flow competence that inhibits the development of sedimentary structures associated to hyperpycnal flows |
|
F5 |
very fine to fine |
parallel |
Nereites |
traction processes in low to
high flow regimen associated |
|
F6 |
very fine to fine |
tangential cross-bedding |
– |
deposited from a unidirectional
flow under conditions |
|
F7 |
very fine to medium tuffaceous sandstone |
symmetrical ripples |
Nereites |
reworking of moderate energy
oscillatory flows related |
|
F8 |
coarse sandstone to fine conglomerates |
massive |
– |
rapid loss of flow competence that inhibits the development of sedimentary structures associated to hyperpycnal flows |
|
F9 |
coarse sandstone to fine conglomerate |
parallel |
– |
traction processes in low to
high flow regimen associated |
|
F10 |
coarse sandstone to fine conglomerate |
tangential cross-bedding |
– |
deposited from a unidirectional
flow under conditions |
The fossil ophiuroid was found 20 m from the base of the Sierra Chacaicó Formation (Fig. 1B), in a massive fine tuffaceous sandstone, between two levels of fine tuffaceous sandstone with parallel lamination. Based on petrographic analysis of the fossil-bearing level, it was classified as a quartz wacke (Dott 1964 modified by Williams et al. 1982). The quartz grains are very fine, poorly sorted, angular to subangular (Fig. 3A, B), and are associated with a lower proportion of plagioclase, muscovite, biotite, pyrite, and glauconite grains (Fig. 3A–D). Abundant organic remains consisting of bryozoans (Fig. 3D), foraminiferal and radiolarian remains (Fig. 3E), sponge spicules, pyritized bivalves (Fig. 3F), and phytodetritus were also observed (Fig. 3).
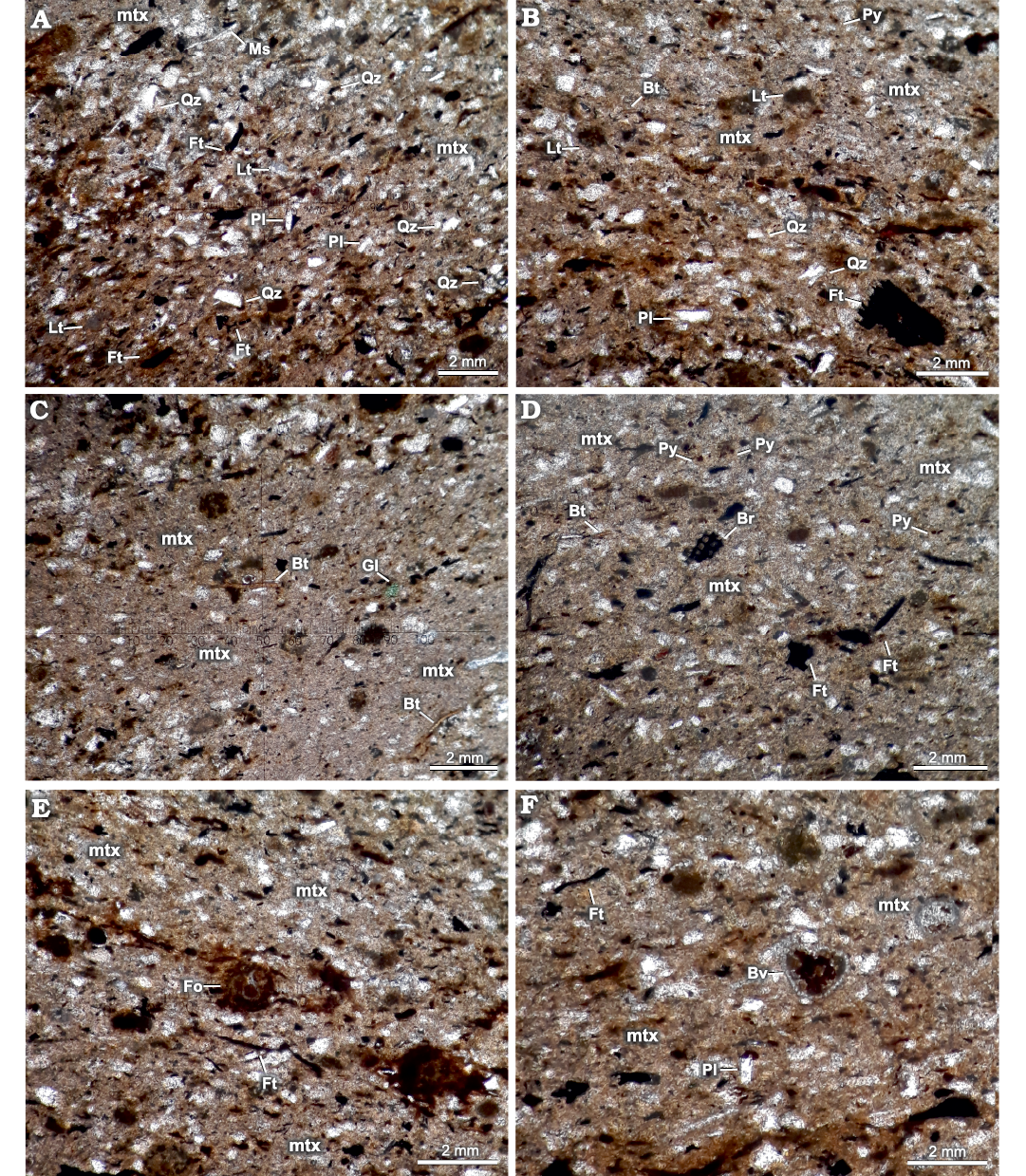
Fig. 3. Microscopic characteristics of the ophiuroid-bearing level. In all photomicrographs it is possible to identify the matrix (mtx). A. Photomicrograph of the level containing the ophiuroid fossil, showing phytodetritus (Ft), lithic fragments (Lt), muscovite (Ms), plagioclase (Pl), and angulose quartz (Qz). B. Photomicrograph of the level containing the ophiuroid fossil, showing biotite (Bt), phytodetritus (Ft), lithic fragments (Lt), plagioclase (Pl), framboidal pyrite (Py), and angulose quartz (Qz). C. Photomicrograph of the level containing the ophiuroid fossil showing biotite (Bt) and glauconite (Gl). D. Photomicrograph of the level containing the ophiuroid fossil showing fragments of bryozoans (Br), biotite (Bt), phytodetritus (Ft), and framboidal pyrite (Py). E. Photomicrograph of the level containing the ophiuroid fossil showing foraminifera (Fo), and phytodetritus (Ft). F. Photomicrograph of the level containing the ophiuroid fossil showing piritized bivalve (Bv), phytodetritus (Ft), and plagioclase (Pl).
Unfortunately, only one side of the ophiuroid specimen was available for study, whereas the counterpart was lost during recovery in the field. Skeletal material is moderately to poorly preserved (Fig. 4A). Some disc and arm ossicles are lost, probably due to dissolution, while others are preserved as low-magnesium calcite plates, showing structural differences with other ossicles (Fig. 4C). Most of the arm and disc ossicles are covered by limonite and alterated pyrite framboids, growing above the pores of the original stereom structure (Fig. 4D, G, H). In addition, pyrite alteration is present in the border of the specimen, demarcating the fossil boundaries (Fig. 4A, C, F).
The specimen is preserved with the disc in subhorizontal position, and an arm extended in one direction, partially curved distally. The other four arms are symmetrically oriented in the opposite direction and lack the distal segments (Fig. 4A, B).
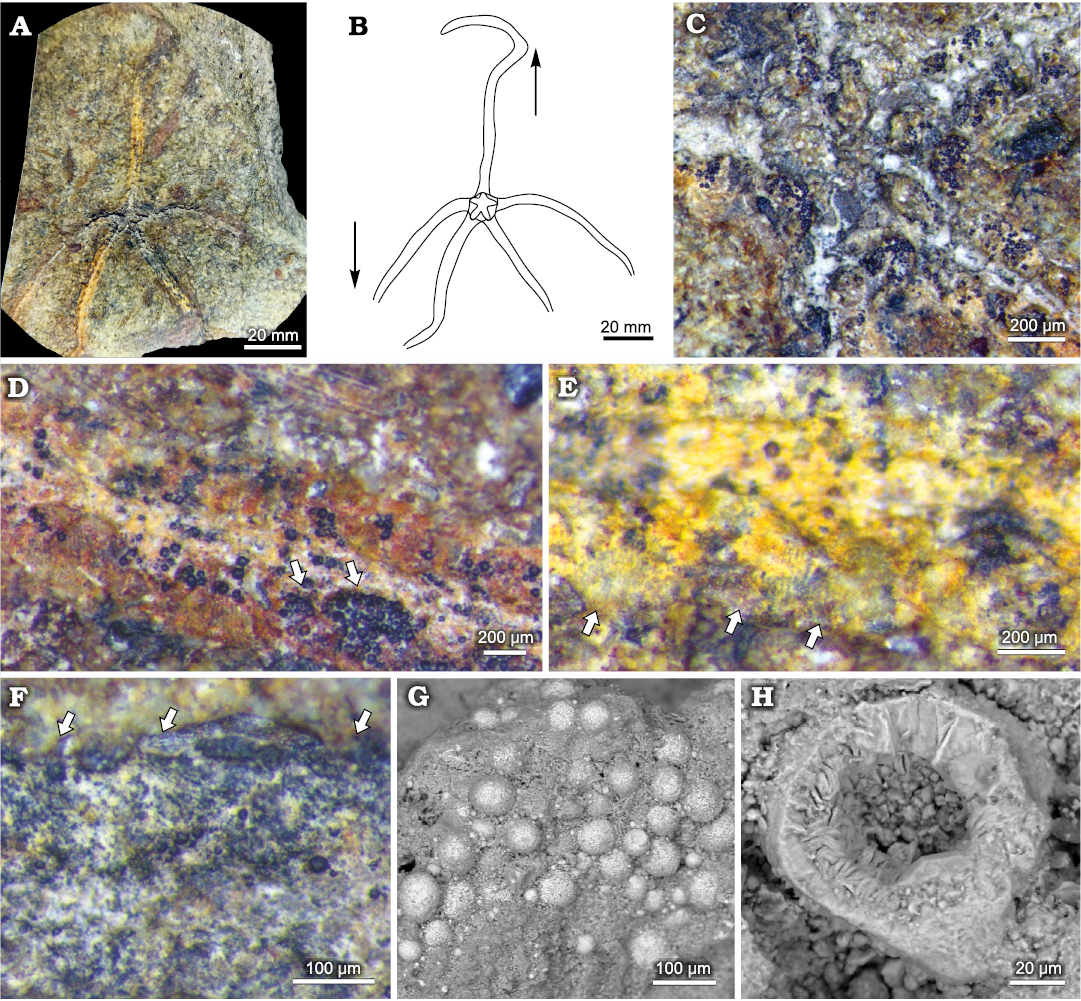
Fig. 4. Ophioleucid ophiuroid Sinosura sp. (MOZ-PI-5930) from the lower Pliensbachian of Sierra Chacaicó Formation, Arroyo Lapa locality, Neuquén, Argentina. A. General view of the studied specimen. B. Diagram of ophiuroid burial position, note that the unpaired arm is oriented in the opposite direction of the other four arms. C. Detail of the central disc. D. Close view of the unpaired arm, arrows showing pyrite framboids. E. Close view of a paired arm, arrows showing limonite covering striated lateral ossicles. F. Unpaired arm close view, arrows showing spines. G. SEM caption of arm lateral ossicle, stereom structure is obscured by the presence of pyrite framboid aggregation. H. Radiolarian test preserved near the ophiuroid arm.
Systematic palaeontology
Phylum Echinodermata Klein, 1778
Subphylum Asterozoa Zittel, 1895
Class Ophiuroidea Gray, 1840
Superorder Ophintegrida O’Hara, Hugall, Thuy, Stöhr, and Martynov, 2017
Order Ophioleucida O’Hara, Hugall, Thuy, Stöhr, and Martynov, 2017
Family Ophioleucidae Matsumoto, 1915
Genus Sinosura Hess, 1964
Type species: Acroura brodiei (Wright, 1866), by original designation; middle Lias (Lower Jurassic), Hewletts Hills near Cheltenham, Gloucestershire, UK.
Diagnosis (modified from Thuy and Numberger-Thuy 2021). —Ophioleucid genus with very large, rounded triangular radial shields surrounded by small disc scales covered by large, scattered granules; arms slender, cylindrical; large tentacle pores covered by large, oval scales; lateral arm plates thin, fragile, strongly arched, with conspicuous vertical striation on outer surface; ventral portion strongly protruding ventro-proximalwards; numerous small spine articulations directly adjacent to distalmost stripe of outer surface striation; row of spine articulations distally bordered by very wide, thin, smooth shelf forming a ventralward-protruding ventro-distal tip of the lateral arm plate; regular arm spines with a coarse longitudinal striation; in some species small, specialized, saw-like dorsalmost arm spines.
Sinosura sp.
Figs. 4, 5.
Material.—Single articulated specimen preserved as 3D ossicles and moulds (MOZ-PI-5930) from Arroyo Lapa, Neuquén, Argentina, Sierra Chacaicó Formation, Lower Jurassic.
Description.—Disc diameter is 2.06 mm, each arm segment is 0.53 mm long, and the maximum arm length is 16.79 mm (Fig. 4A, C). The specimen exposes the ventral side. The interradial disk skeleton is poorly preserved and shows no discernible structures such as disc scales or genital plates. The mouth skeleton is composed of five pairs of slender oral plates, with oval lateral oral papillae sensu lato and a single, slender, pointed ventralmost tooth (Fig. 5A). The exact number and arrangement of the lateral oral papillae sensu lato are not discernible. The adoral shields are slender and meet in front of a large, arrow-shaped oral shield. The vertebrae are preserved both in 3D and as molds (Fig. 4D, E, 5A) and mostly expose a large ventral furrow. Some of the lateral arm plates are sufficiently well preserved to show microstructural details. They are relatively long and very thin. Their outer surface shows a fine, very well developed vertical striation composed of imbricate lamellae (Fig. 5B, C). The spine articulations are small and directly adjacent to the distalmost lamella of the outer surface striation. They are composed of arched dorsal and ventral lobes (Fig. 5B) encompassing a comma-shaped muscle opening, and carry relatively thick, cylindrical arm spines whose length equals at least half of the segment length (Fig. 4F, 5C). Some arm spines are disarticulated and/or broken (Fig. 4E, F). The ventral portion of the lateral arm plates seems to be very large and strongly protruding ventralwards. It is deeply incised by a large, ventralwards pointing tentacle notch (Fig. 5C).

Fig. 5. Ophioleucid ophiuroid Sinosura sp. (MOZ-PI-5930) from the lower Pliensbachian of Sierra Chacaicó Formation, Arroyo Lapa locality, Neuquén, Argentina. A. Mouth skeleton of the studied specimen in ventral view, showing the vertebrae (V) of two arms, one oral shield (OS) proximally bordered by the adoral shields (AOS), a pair of oral plates (OP) and a ventralmost tooth (T). B. Detail of the lateral arm plates (LAP) showing the vertical striation and the spine articulations composed of dorsal (DL) and ventral lobes (VL), with an arrowing showing pointing to the distal (di) end of the arm. C. Detail of proximal arm segments in ventral view, showing the striated lateral arm plates (LAP), the arm spines (AS) and the tentacle notches (TN), with an arrowing showing pointing to the distal (di) end of the arm.
Remarks.—In spite of the poor preservation of the ophiuroid skeleton, some diagnostic characters are visible and allow for a taxonomic interpretation of the specimen. The features pertaining to the lateral arm plates are of greatest value. Their overall shape and fragility, the conspicuous outer surface striation, the shape and position of the spine articulations and the size and orientation of the tentacle pores all suggest an assignment to the ophioleucid genus Sinosura (Hess 1964; Thuy and Numberger-Thuy 2021). The shape of the oral shield and the oral papillae and teeth are in agreement with an assignment to Sinosura (Hess 1964). The very large ventral portion of the lateral arm plates and the large size and conical shape of the arm spines suggest closest affinities with Sinosura dieschbourgae Thuy and Numberger-Thuy, 2021, from the lowermost Toarcian of Luxembourg and Germany. The poor preservation of the specimen from the Neuquén Basin, however, precludes a conclusive comparison with the currently known species of the genus.
So far, Sinosura has been recorded from the Jurassic and, albeit with some doubt, from the uppermost Cretaceous of Europe (Stöhr et al. 2021). Species of this genus were especially common components of Early Jurassic soft-bottom communities and also occurred in poorly oxygenated settings (Thuy et al. 2011; Thuy and Numberger-Thuy 2021). The specimen described herein adds the first South American record of Sinosura and thereby significantly expands the paleogeographic range of the genus.
Discussion
Rapid burial, low oxygen concentration and low decay rates are commonly claimed as the main factors allowing for the preservation of articulated brittle star skeletons in a marine depositional setting (Schäfer 1972; Brett et al. 1997; Martínez et al. 2010).
In the Sierra Chacaicó Formation, the ophiuroid specimen was found in levels of massive fine tuffaceous sandstone accumulated by mechanical processes, covered by coarse sandstone with a large amount of plant debris and organic matter. This type of deposit can be produced by the continuous aggradation of a sustained flow with high concentration of suspended sediments, or in areas of slope change where hydraulic jumps and collapse of the suspended load occur (Bursik and Woods 1996; Branney and Kokelaar 2002; Ponce and Carmona 2011; Ponce et al. 2018). Both processes are produced by hyperpycnal flows. Hyperpycnal discharges provide a large volume of organic matter (phytodetritus and wood fragments) and fresh water, as interstitial fluid, resulting in brackish conditions of the systems. In addition, the presence of laminated mudstone with framboidal pyrite and limonite, and the occurrence of the ichnospecies Chondrites intricatus and Trichichnus isp., generated by infaunal deposit feeders or chemosymbiotic organisms (Bromley and Ekdale 1984; Savrda and Bottjer 1991; Kędzierski et al. 2015), suggests dysoxic conditions (Wignall and Newton 1998; Ponce et al. 2015; Carmona et al. 2016; Rickard 2019; Campetella et al. 2020).
Ophiuroids can escape rapid burial when covering sediment is 1–3 cm thick, moving towards the sediment surface, but are entombed alive when the sediment cover is thicker than 5 cm (Reid 2017). In some cases, ophiuroids can move within the sediment, trying to escape from the sediment deposition, but eventually they die and are preserved in walking or escape positions (Ishida and Fujita 2001; Reid et al. 2019). The disc and arm arrangement of the studied specimen is similar to the “walking posture” of fossil and Recent Ophiura sarsii sarsii illustrated by Ishida and Fujita (2001). Usually, this posture is adopted by ophiuroids while moving towards the direction of the unpaired arm (Fig. 4A, B; Ishida and Fujita 2001; Reid 2017), suggesting that the studied brittle star was buried alive by sediment thick enough to prevent it from escaping (Reid et al. 2019). It cannot be entirely ruled out that the ophiuroid was transported alive before being buried. The absence of broken arms as a result of mechanical stress during transport, however, suggests burial in situ. This implies that the ophiuroid lived in dysoxic conditions. In this respect, our finding parallels previous reports of Sinosura from dysoxic settings (Hess 1991; Thuy and Numberger-Thuy 2021).
Specimen preservation suggests that, after burial, the taphonomic history was complex, involving several stages. The organism was entombed below the oxic-anoxic redox interface by deposition of a thick layer of sediment carrying a high amount of particulate organic matter (POM). During early diagenesis, skeletal material was replaced by more stable low-magnesium calcium carbonate. At a later time, an iron enriched fluid circulated through the sediment, generating the precipitation of pyrite framboids above the ossicles, obliterating the stereom structures and partially dissolving the skeletal calcite. Finally, the specimen was exposed and experienced oxidation processes, resulting in the weathering of pyrite into limonite and other ferric oxides, as commonly happens with fossils preserved in dysoxic and euxinic layers (Glass et al. 2003; Hernick et al. 2008).
Conclusions
The studied material represents the earliest Mesozoic record of brittle stars in South America.
The specimen belongs to the extinct ophioleucid genus Sinosura and represents the first record of this genus outside Europe.
The brittle star specimen is preserved almost completely, in situ and in a “walking posture”, suggesting that the organism was buried alive. Low oxygenation and rapid, definite burial favored the preservation of the articulated ossicles.
The taphonomic and sedimentologic evidence indicates that these levels were accumulated in a prodelta/offshore setting, associated with episodic hyperpycnal flows.
Acknowledgements
This research was supported by three PhD scholarships given by CONICET to Débora Campetella, Evangelina Palópolo, and Maximiliano Rodríguez, PI UNRN 40-A-616 given to Noelia Carmona, PI UNRN and PUE 0031CO from the Instituto de Investigación en Paleobiología y Geología, General Roca, Río Negro, Argentina. All these contributions are gratefully acknowledged. We also whish to thank the referees Yoshiaki Ishida (Tokyo, Japan) and Diana E. Fernández (Universidad de Buenos Aires, Argentina) and the editors who assisted in substantially improving the manuscript.
References
Aronson, R.B. 1989. Brittlestar beds: Low-predation anachronisms in the British isles. Ecology 70: 856–865. Crossref
Ausich, W.I., Jangoux, M., and Lawrence, J.M. 2001. Echinoderm taphonomy. Echinoderm Studies 6: 171–227.
Branney, M.J. and Kokelaar, B.P. 2002. Pyroclastic Density Currents and the Sedimentation of Ignimbrites. Geological Society of London, Memoirs 27: 1–137.
Brett, C.E., Moffat, H.A., and Taylor, W.L. 1997. Echinoderm taphonomy, taphofacies, and Lagerstätten. The Paleontological Society Papers 3: 147–190. Crossref
Bromley, R.G. and Ekdale, A.A. 1984. Chondrites: a trace fossil indicator of anoxia in sediments. Science 224: 872–874. Crossref
Brongniart, A.T. 1823. Observations sur les fucoids. Société d’Histoire Naturelle de Paris, Mémoires 1: 301–320.
Brusca, R.C. and Brusca, G.J. 2003. Invertebrates. 2nd Edition. 936 pp. Sinauer Associates Inc., Sunderland.
Bursik, M.I. and Woods, A.W. 1996. The dynamics and thermodynamics of large ash flows. Bulletin of Volcanology 58: 175–193. Crossref
Campetella, D.M. 2022. Sedimentología e Icnología de la Formación Los Molles en las Localidades de Picún Leufú, Sierra de Chacaico y Cordillera del Viento, Cuenca Neuquina, Argentina. 214 pp. Unpublished Ph.D. Thesis, Universidad Nacional de Río Negro, General Roca.
Campetella, D.M., Carmona, N.B., Ponce, J.J., Wetzel, A., Rodríguez M.N., and Parada, M.N. 2020. Trace fossils as tools to unravel oxygen conditions: A case study from Los Molles Formation (Toarcian–lower Callovian). Revista de la Asociación Geológica Argentina 77: 463–477.
Carmona, N., Ponce, J.J., and Wetzel, A. 2016. Sedimentología e Icnología de la Formación Sierra Chacaicó en la localidad de Arroyo Lapa, Cuenca Neuquina, Argentina. In: E. Bedatou and A.E. Mehl (eds.), Actas VII Congreso Latinoamericano de Sedimentología, XV Reunión Argentina de Sedimentología. 53 pp. Santa Rosa, La Pampa.
Caviglia, S.E., Martínez, S., and del Río, C.J. 2007. A new Early Miocene species of Ophiocrossota (Ophiuroidea) from Southern Patagonia, Argentina. Neues Jahrbuch für Geologie und Palaontologie Abhandlungen 245: 147–152. Crossref
Dott, R.H., Jr. 1964. Wacke, graywacke, and matrix: what approach to immature sandstone classification. Journal of Sedimentary Petrology 34: 625–632. Crossref
Fernández, D.E., Giachetti, L., Stöhr, S., Thuy, B., Pérez, D.E., Comerio, M., and Pazos, P.J. 2019. Ofiuroideos del Cretácico Inferior de Patagonia: primer registro fósil articulado para el Mesozoico de América del Sur. Andean Geology 46: 421–432. Crossref
Fischer-Ooster, C. von 1858. Die fossilen Fucoiden der Schweizer Alpen, nebst Erörterung über deren geologisches Alter. 74 pp. Huber und Companie, Bern.
Fraga, M.C. and Vega, C.S. 2020. Asterozoans from the Devonian of the Paraná basin, south Brazil. Journal of South American Earth Sciences 97: 102398. Crossref
Franzese, J.R. 1995. El Complejo Piedra Santa (Neuquén, Argentina): parte de un cinturón metamórfico neopaleozoico del Gondwana suroccidental. Andean Geology 22: 193–202.
Franzese, J.R., Veiga, G.D., Muravchik, M., Ancheta, M.D., and D’Elía, L. 2007. Estratigrafía de “sin-rift” (Triásico Superior-Jurásico Inferior) de la Cuenca Neuquina en la sierra de Chacaico, Neuquén, Argentina. Revista Geológica de Chile 34: 49–62. Crossref
Furque, G. and Camacho, H.H. 1949. El Cretácico Superior de la costa atlántica de Tierra del Fuego. Revista de la Asociación Geológica Argentina 4: 263–297.
Glass, A., Ausich, W.I., and Copper, P. 2003. New cyclocystoid (Phylum Echinodermata) from Anticosti Island, Quebec, and its bearing on cyclocystoid life modes. Journal of Paleontology 77: 949–957. Crossref
Gray, J.E. 1840. A synopsis of the genera and species of the Class Hypostoma (Asterias, Linnaeus). Annals and Magazine of Natural History 6: 175–184. Crossref
Gulisano, C.A. and Gutiérrez Pleimbing, A. 1995. Field guide: The Jurassic of the Neuquén Basin. Neuquén province. Asociación Geológica Argentina, Serie E 2: 1–111.
Haude, R. 1995. Echinodermen aus dem Unter-Devon der argentinischen Praekordillere. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 197: 37–86. Crossref
Hernick, L.V.A., Landing, E., and Bartowski, K.E. 2008. Earth’s oldest liverworts—Metzgeriothallus sharonae sp. nov. from the Middle Devonian (Givetian) of eastern New York, USA. Review of Palaeobotany and Palynology 148: 154–162. Crossref
Hess, H. 1964. Die Ophiuren des englischen Jura. Eclogae geologicae Helvetiae 57: 756–801.
Hess, H. 1991. Neue Schlangensterne aus dem Toarcium und Aalenium des Schwäbischen Jura (Baden-Württemberg). Stuttgarter Beiträge zur Naturkunde, Serie B 180: 1–11.
Hillebrandt, A. 1981. Faunas de amonites del Liásico inferior y medio (Hettangiano hasta Pliensbachiano) de América del Sur (excluyendo Argentina). In: W. Volkheimer and E. Musacchio (eds.), Cuencas Sedimentarias del Jurásico y Cretácico América del Sur 2: 499–538.
Ishida, Y. and Fujita, T. 2001. Escape behavior of epibenthic ophiuroids buried in the sediment: Observations of extant and fossil Ophiura sarsii sarsii. In: A.N. Barker (ed.), Echinoderms 2000, 285–292. CRC Press, Boca Ratón, Florida.
Kędzierski, M., Uchman, A., Sawlowicz, Z., and Briguglio, A. 2015. Fossilized bioelectric wire—the trace fossil Trichichnus. Biogeosciences 12: 2301–2309. Crossref
Klein, J.T. 1778. Naturalis Dispositio Echinodermatum. Accessit Lucubratiuncula de Aculeis Echinorum Marinorum et Specilegium de Belemnitis. 278 pp. Edita et Descriptionibus Novisque Inventis et Synonymis Auctorum Auca a Nathanaele Godofredo Leske. Officina Gleditdchiana, Lipsiae.
Leanza, H. 1990. Estratigrafía del Paleozoico y Mesozoico anterior a los movimientos intermálmicos en la comarca del Cerro Chachil. Revista de la Asociación Geológica Argentina 45: 272–299.
Leanza, H.A. and Hugo, C.A. 1997. Hoja Geológica 3969-III Picún Leufú, provincias del Neuquén y Río Negro. Programa Nacional de Cartas Geológicas de la República Argentina a escala 1: 250.000. Instituto de Geología y Recursos Minerales. SEGEMAR, Boletín 218: 1–135.
Leanza, H.A., Llambías, E., and Carbone, O. 2005. Unidades estratigráficas delimitadas por discordancias en los depocentros de la Cordillera del Viento y la Sierra de Chacaico durante los inicios de la Cuenca Neuquina. In: 6 Congreso de Exploración y Desarrollo de Hidrocarburos, 13. Instituto Argentino del Petróleo y del Gas, Mar del Plata.
Malumián, N. and Olivero, E.B. 2005. Shallow-water late middle Eocene crinoids from Tierra del Fuego: a new southern record of a retrograde community structure. Scientia Marina 69: 349–353. Crossref
Martínez, S.A. and Río, D. 2008. A new, first fossil species of Ophioderma Müller and Troschel, 1842 (Echinodermata: Ophiuroidea) (Late Miocene, Argentina). Zootaxa 1841: 43–52. Crossref
Martínez, S., del Río, C.J., and Pérez, D.E. 2010. A brittle star bed from the Miocene of Patagonia, Argentina. Lethaia 43: 1–9. Crossref
Matsumoto, H. 1915. A new classification of the Ophiuroidea: with description of new genera and species. Proceedings of the Academy of Natural Sciences of Philadelphia 68: 43–92.
Nebelsick, J.H. 2004. Taphonomy of Echinoderms: introduction and outlook. In: T. Heinzeller and J.H. Nebelsick (eds.), Echinoderms-München, 471–477. Taylor and Francis Group, London. Crossref
O’Hara, T.D., Hugall, A.F., Thuy, B., Stöhr, S., and Martynov, A.V. 2017. Restructuring higher taxonomy using broad-scale phylogenomics: The living Ophiuroidea. Molecular Phylogenetics and Evolution 107: 415–430. Crossref
Pángaro, F., Pereira, D., and Minucci, E. 2009. El sinrift de la dorsal de Huincul, Cuenca Neuquina: evolución y control sobre la estratigrafía y estructura del área. Revista de la Asociación Geológica Argentina 65: 265–277.
Ponce, J.J. and Carmona, N.B. 2011. Miocene deep-marine hyperpycnal channel levee complexes, Tierra del Fuego, Argentina: Facies associations and architectural elements. Sediment transfer from shelf to deep water-Revisiting the delivery system. AAPG Studies in Geology 61: 75–93. Crossref
Ponce, J.J., Carmona, N., and Montagna, A.O. 2018. Atlas de estructuras sedimentarias inorgánicas y biogénicas. Descripción, análisis e interpretación a partir de afloramientos, testigo corona y registros de imágenes de pozo. 166 pp. Fundación YPF, General Roca.
Ponce, J.J., Carmona, N.B., Montagna A.O., and Canale, N. 2015. Sedimentología e Icnología de los Sistemas Petroleros no Convencionales de la Cuenca Neuquina. (Los Molles-Lajas y Vaca Muerta-Quintuco), Guía de Campo. 112 pp. Fundación YPF y Universidad Nacional de Río Negro, General Roca.
Pujana, I. 1996. A New Lower Jurassic Radiolarian fauna from the Neuquén Basin, Central west Argentina. In: XIII Congreso Argentino de Geología, y II Congreso de Exploración de Hidrocarburos, Actas V, 133–142. Asociación Geológica Argentina, Buenos Aires.
Reid, M. 2017. Taphonomy, Palaeoecology and Taxonomy of an Ophiuroid-stylophoran Obrution Deposit from the Lower Devonian Bokkeveld Group, South Africa. 149 pp. Unpublished M.Sc. Thesis, University of Cape Town, Cape Town. Crossref
Reid, M., Taylor, W.L., Brett, C.E., Hunter, A.W., and Bordy, E.M. 2019. Taphonomy and paleoecology of an ophiuroid-stylophoran obrution deposit from the lower Devonian Bokkeveld Group, South Africa. Palaios 34: 212–228. Crossref
Rickard, D. 2019. Sedimentary pyrite framboid size-frequency distributions: A meta-analysis. Palaeogeography, Palaeoclimatology, Palaeoecology 522: 62–75. Crossref
Ruedemann, R. 1916. Account of some new or little known species of fossils mostly from Palaeozoic rocks of New York. Paleontological Contributions from the New York State Museum 189: 7–97.
Savrda, C.E. and Bottjer, D.J. 1991. Oxygen-related biofacies in marine strata: an overview and update. In: R.V. Tyson and T.H. Pearson (eds.), Modern and Ancient Continental Shelf Anoxia. Geological Society London Special Publication 58: 201–219. Crossref
Schäfer, W. 1972. Ecology and Palaeoecology of Marine Environments. 586 pp. Oliver & Boyd, Edinburgh.
Seilacher, A. 1976. Palökologie, Konstruktionen, Sedimentologie, Diagenese und Vergesellschaftung von Fossilien. Zentralblatt für Geologie und Paläontologie 2: 227–233.
Shroat-Lewis, R.A. 2007. Taphonomy of a Pliocene ophiuroid mass mortality lagerstätte in the Tirabuzón Formation, Baja California sur. 70 pp. Unpublished M.Sc. Thesis, University of North Carolina, Wilmington.
Stöhr, S., O’Hara, T., and Thuy, B. 2021. Sinosura Hess, 1964†. In: S. Stöhr, T. O’Hara, and B. Thuy (eds.), World Ophiuroidea Database. Accessed at: http://www.marinespecies.org/Ophiuroidea/aphia.php?p=taxdetails &id=745133 on 2022-09-26
Thuy, B. and Numberger-Thuy, L.D. 2021. Brittlestar diversity at the dawn of the Jenkyns Event (early Toarcian Oceanic Anoxic Event): new microfossils from the Dudelange drill core, Luxembourg. Geological Society, London, Special Publications 514: 83–119. Crossref
Thuy, B., Gale, A.S., and Reich, M. 2011. A new echinoderm Lagerstätte from the Pliensbachian (Early Jurassic) of the French Ardennes. Swiss Journal of Palaeontology 130: 173–185. Crossref
Thuy, B., Klompmaker, A.A., and Jagt, J.W. 2012. Late Triassic (Rhaetian) ophiuroids from Winterswijk, the Netherlands; with comments on the systematic position of Aplocoma (Echinodermata, Ophiolepididae). Zoosymposia 7: 163–172. Crossref
Thuy, B., Numberger-Thuy, L., and Jagt, J.W. 2020. A new ophiacanthid brittle star (Echinodermata, Ophiuroidea) from sublittoral crinoid and seagrass communities of late Maastrichtian age in the southeast Netherlands. PeerJ 8: e9671. Crossref
Volkheimer, W. 1973. Palinología estratigráfica del Jurásico de la Sierra de Chachico y adyacencias (Cuenca Neuquina, República Argentina). I: Estratigrafía de las formaciones Sierra Chacai Co, Los Molles, Cura Niyeu y Lajas. Ameghiniana 10: 105–131.
Volkheimer, W. and Quattrocchio, M. 1993. Formación Sierra Chacaicó. In: A.C. Riccardi and S. Damborenea (eds.), Léxico estratigráfico de la Argentina. Volumen 9: Jurásico. Asociación Geológica Argentina, Serie B 21: 393–394.
Weaver, C. 1931. Paleontology of the Jurassic and Cretaceous of West Central Argentina. Universty of Washington Memoirs 1: 1–469.
Wignall, P.B. and Newton, R. 1998. Pyrite framboid diameter as a measure of oxygen deficiency in ancient mudrocks. American Journal of Science 298: 537–552. Crossref
Williams, D.F., Turner, F.J., and Gilbert, C.M. 1982. Petrography (2nd Edition). 626 pp. W.H. Freeman, San Francisco.
Wright, T. 1866. A Monograph on the British fossil Echinodermata from the Oolitic Formations. Volume 2. Part 2. On the Ophiuroidea. The Palaeontographical Society: 132–154.
Zatoń, M., Salamon, M.A., Boczarowski, A., and Sitek, S. 2008. Taphonomy of dense ophiurdiedoid accumulations from the Middle Triassic of Poland. Lethaia 41: 47–58. Crossref
Zavala, C. and Arcuri, M. 2016. Intrabasinal and extrabasinal turbidites: Origin and distinctive characteristics. Sedimentary Geology 337: 36–54. Crossref
Zittel, K.A. 1895. Grundzüge der Paläontologie (Paläzoologie). 971 pp. Von R. Oldenbourg, München,
Acta Palaeontol. Pol. 68 (1): 143–153, 2023
https://doi.org/10.4202/app.01032.2022