
An Eocene sea turtle from the eastern North Pacific fills a Paleogene gap
ASHLEY W. POUST, PATRICIA A. HOLROYD, and THOMAS A. DEMÉRÉ
Sea turtles (Testudines: Chelonioidea) represent a monophyletic group with a global distribution in the fossil record, although most of our knowledge of their diversity and morphology comes from the Atlantic Ocean and Tethyan Sea (Evers and Benson 2019; Gentry et al. 2019). By contrast, the record of their evolution in the vast Pacific Basin prior to the Neogene is paltry. Late Cretaceous chelonioids are known from northern California (Parham et al. 2003), British Columbia (Nicholls 1992; Nicholls and Meckert 2002), Japan (Hirayama and Chitoku 1996), Australia (Kear et al. 2003; Kear 2006; Kear and Lee 2006), and New Zealand (Wiffen 1981), indicating they occupied at least portions of both the North and South Pacific Oceans at this time. Pacific Paleocene specimens are entirely unknown, and to date the Eocene record has been limited to chelonioids and dermochelyids (Köhler 1995a, b; Grant-Mackie et al. 2011) from New Zealand, and an informal report of a sea turtle from Oregon (Orr and Orr 2009). Here, we describe a new sea turtle fossil from the middle Eocene of California which demonstrates that members of this clade were indeed present in the North Pacific at this time.
Institutional abbreviations.—IRSNB, Royal Belgian Institute of Natural Sciences, Brussels, Belgium; SDNHM, San Diego Natural History Museum (run and operated by SDSNH, San Diego Society of Natural History), San Diego, USA.
Geologic setting
Stratigraphy.—SDSNH 103374 was recovered from middle Eocene strata during 2004 construction of the Bressi Ranch planned community in Carlsbad, San Diego County, California, USA (Fig. 1A). The upper estuarine/lagoonal stratigraphic unit of the Santiago Formation that produced SDSNH 103374 is an ~18 meter-thick complex sequence of cross-bedded sandstone, siltstone, and mudstone beds in a ~110 meter-thick Eocene section (Fig. 1B). Ripple cross-stratification, trough cross-stratification, graded bedding, and bioturbation were all observed in this unit. Several beds contained abundant carbonaceous horizons (e.g., SDSNH locality 5560) with leaf compressions of terrestrial plants, and others (e.g., SDSNH localities 5550, 5555, and 5561) had dense concentrations of estuarine mollusks. Typically, thicker cross-bedded strata were composed of yellow to yellowish-orange, lenticular, well-cemented sandstone bodies, which were fine- to medium-grained and shelly at the base and graded upwards into siltstone or greenish-gray massive mudstone. One mudstone stratum (SDSNH locality 5567) produced well-preserved remains of stomatopod crustaceans (Haug et al. 2013). Another, (SDSNH locality 5570, dubbed the “fish bed”) contained partial and complete skeletons of bony fishes and produced the chelonioid fossil described below.
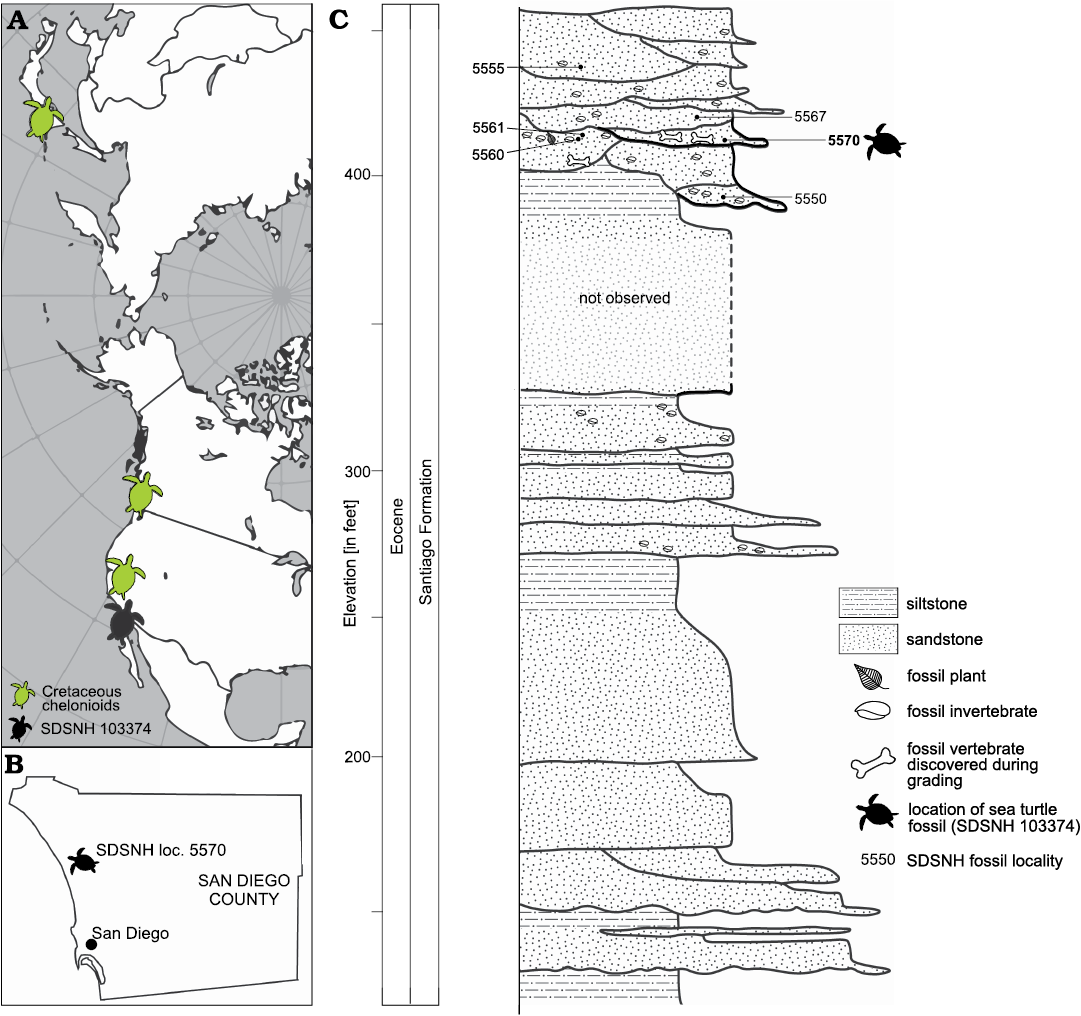
Fig. 1. Geographic and geologic context of sea turtle fragments (Chelonioidea gen. et sp. indet.). A. Map of northern Pacific showing sea turtle localities. B. Position of Bressi Ranch locality of the Santiago Formation (SDSNH loc. 5570) in Southern California, USA. C. Stratigraphic column of the Bressi Ranch.
The “fish bed” is a localized, thin stratum of bluish-gray, organic-rich, ripple-laminated and bioturbated siltstone. Roughly cylindrical burrows, about 1 cm in diameter, penetrate the siltstone both vertically and horizontally. The burrows are filled with medium- to coarse-grained sandstone with dispersed shells and shell hash derived from the immediately overlying stratum. The fish skeletons, as well as whole and fragmentary leaves occur concordant to bedding, closely packed, and often stacked. The fish assemblage includes haemulids (grunts), soleids (soles), clupeids (herrings and shads), ariids (catfish), and Genartina sp. (bony-tongued fish). Plant fossils are dominated by leaves of the cocoplum, Chrysobalanus sp. Also occurring in this horizon are fragments of carbonized wood, spotty occurrences of pyrite/marcasite, mollusk and barnacle shells, and shark and crocodile teeth.
Depositional environment.—Invertebrates from the “fish bed” include the gastropods Ampullella schencki, Nerita triangulata, Neverita globosa, Potamides carbonicola, Umpquaia oregonensis, and the bivalves Cuneocorbula torreyensis, Ostrea idrianensis, and Pelecyora aequilateralis (Givens and Kennedy 1978; Squires 1992, 1999). These faunal elements are typical of Eocene lagoonal intertidal marsh flat and tidal creek facies reported elsewhere in coastal San Diego County (Boyer and Warme 1975; Clifton 1979; Eisenberg and Abbott 1985; Warme 1991). In the Bressi Ranch area, these lagoonal facies directly overlie steep, irregular paleotopography suggesting a proximal depositional setting within a broader lagoonal habitat protected by adjacent headlands.
Correlation and geologic age.—The age of the Santiago Formation is based on biochronological evidence from mammalian fossils, collected from its component members B and C and correlated with portions of the Uintan and Duchesnean North American Land Mammal Ages (Golz 1976; Golz and Lillegraven 1977; Walsh 1996), and limited paleomagnetic work indicating correlation with Chron C20R through Chron C18R (Prothero 2001).
Member B contains index fossils for Uintan biochron Ui1 and Ui2 (~45–43 Ma; Robinson et al. 2004). Mammals from the lower part of member C are correlated to Uintan biochron Ui3 (~42–41 Ma), whereas those from the upper part of member C are correlated with Uintan biochron Ui3 and the Duchesnean biochron (~39–40 Ma; Robinson et al. 2004). No terrestrial mammals were recovered from the Bressi Ranch stratigraphic sequence. However, the topographic and stratigraphic position of the Bressi Ranch “fish bed” relative to nearby Santiago Formation outcrops strongly suggest a correlation with member B. This position for the deposit allows us to assign the sea turtle fossil a likely age of middle Eocene (Lutetian, 45–43 Ma).
Systematic palaeontology
Testudines Batsch, 1788
Cryptodira Cope, 1868
Chelonioidea Baur, 1893
Gen. et sp. indet.
Fig. 2.
Material.—SDSNH 103374, a partial nuchal and right and left first peripherals (collected by Gino Calvano, March, 2004) from SDSNH locality 5570 (“fish bed”), middle Eocene (Lutetian, 43–45 Ma), San Diego County, California, USA. This locality occurred within informal member B of the Santiago Formation. Additional locality information is available on request from SDNHM.
Measurements (in mm).—Width 120.6; ant/post height, lateral 21.2; ant/post height, midline 33.7; maximum thickness 6.7; width of “keel” 14.6.
Description.—SDSNH 103374 is a thin, “boomerang”-shaped fossil 6.7 mm at its thickest point, tapering to 2 mm at the acute anterior edge. The nuchal and partial right and left first peripherals are preserved, together forming what would have been a slightly dorsoventrally arched embayment extending concavely into the anterior portion of the carapace. The sutures are poorly expressed, but a complex suture is visible 22.1 mm from the midline on the visceral surface (equivalent to ventral view) of the left side of the nuchal embayment. The concave anterior opening measures 91.9 mm in width. The dorsal surface is subtly sculptured, and no sulci indicating the position of the scales are visible. The external cortical bone is thin and spongy; cancellous internal bone (diploe) with large pore spaces was exposed by erosion. This bone structure is typical in cryptodiran marine turtles, in contrast to marine or freshwater pleurodires and more pelagic cryptodires which generally have a thicker external cortical wall and thinner internal diploe (Scheyer 2009; Scheyer et al. 2014). On the visceral surface a thickened and slightly raised midline ridge is present for articulation with the 8th cervical vertebra. Due to erosion, it is not possible to determine whether the articular surface was formed as a blunt facet or raised pedestal (see e.g., character 120 of Cadena and Parham 2015; or characters 62 and 126 of Anquetin 2012). The posterior edge of the preserved nuchal is worn, so it is not possible to determine whether it was sutured to adjacent bones or separated by fontanelles. Two minute foramina are located bilaterally close to the midline of the nuchal. These structures are rimmed with intact cortical bone and are not taphonomic artefacts. The transverse torus is well expressed, and there is no costiform process. The first peripherals are facultatively fused to the nuchal, although a small indentation in the surface of the nuchal opening and faint impressions in the dorsal surface indicate where the bones meet. The first peripherals are incomplete, but the exterior outline would have been angled or “elbow-shaped” as seen in some other chelonioids (Lapparent de Broin et al. 2018).
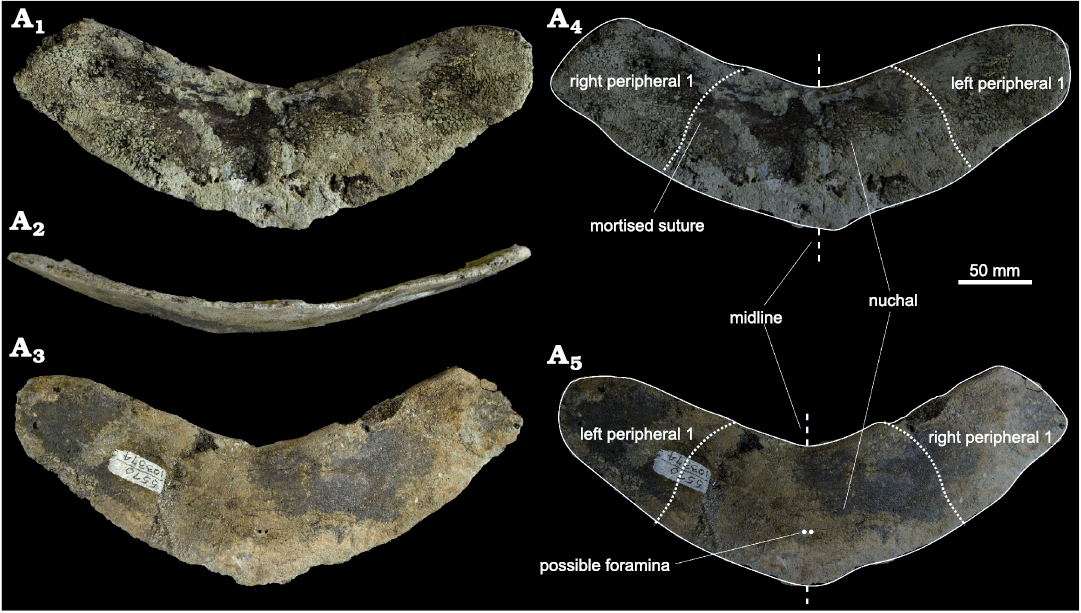
Fig. 2. Photographs of sea turtle nuchal and peripherals of Chelonioidea gen. et sp. indet. (SDSNH 103374) from the Eocene Santiago Formation of California, USA (SDSNH loc. 5570). Ventral (A1, A4), anterior (A2), and dorsal (A3, A5) views.
The wider-than-long nuchal is dissimilar in shape and dimensions to that of dermochelyids, but consistent with that seen in other chelonioids with marked nuchal embayments. The curvature of the nuchal embayment is less than in many Cretaceous chelonioids (e.g., ctenochelyids; Zangerl 1953; Gentry 2018) and greater than in Eocene stem-cheloniids (e.g., European Puppigerus Cope, 1970 [Moody 1974], Ashleychelys Weems and Sander, 2014, Procolpochelys Hay, 1908, and Carolinochelys Hay, 1923 [Weems and Sanders 2014]). In thinness and degree of the nuchal embayment, SDSNH 103374 is most similar to Eochelone brabantica Dollo, 1903, and to a lesser extent Eochelone monstigris Grant-Mackie, Hill, and Gill, 2011, and/or Argillochelys sp. (Grant-Mackie et al. 2011; Lapparent de Broin et al. 2018). It differs from E. monstigris in lacking an acute corner at the edge of the nuchal embayment. SDSNH 103374 is more strongly curved and emarginate than similarly thin-shelled forms such as Erquelinessia gosseleti (IRSNB 1628; Lapparent de Broin et al. 2018).
The phylogenetic placement of incomplete carapacial specimens is challenging, due to both a paucity of characters and a lack of Paleogene taxa in most recent phylogenetic analyses of chelonioids. When included, Paleogene taxa are basal to extant taxa (e.g., Evers and Benson 2019). Weems and Brown (2017) provide the most comprehensive recent phylogenetic analysis (including multiple Paleogene taxa) based largely on cranial characters, and consistently recovered several weakly-supported clades among Paleogene and Neogene chelonioids. Notably, they recognized a clade comprising Eocene Erquelinessia gosseleti and the Miocene Californian and Peruvian taxon Pacifichelys. This phylogenetic hypothesis suggests a long ghost lineage for the origin of Pacifichelys.
Lapparent de Broin et al. (2018) informally grouped several thin-shelled Eocene–Oligocene species, including European Eochelone spp., Puppigerus camperi, and Glarichelys spp., united in part by the shape of the nuchal region, i.e., having a slight protrusion of the anterior carapacial border delimited by an “elbowed” peripheral 1 and an “anterior transversal” or slightly notched nuchal border. These authors did not consider Eochelone monstigris from New Zealand, but both it and SDSNH 103374 would appear to fall within this informal group. However, it is not clear if this is a monophyletic group, or whether it corresponds to one or more of the clades recognized by Weems and Brown (2017) as the closest Paleogene sister group to crown Cheloniidae.
Concluding remarks
Middle Eocene marine vertebrates of the northeastern Pacific Ocean are poorly known compared with the rich record of the North American Gulf Coast or Atlantic Coastal Plain. Marine fish scales were reported from California’s Central Valley (David 1946). Isolated chondrichthyan teeth were noted from the Scripps Formation of San Diego County (Walsh 1991), the Llajas Formation of Ventura County, California (Squires 1984, 2001), the Coaledo, Nestucca, and Spencer formations of west-central Oregon (Welton 1972), and Tukwila Formation of northwestern Washington (Nesbitt 1998). The only substantive marine microvertebrate assemblage is the Lake Miramar fauna from the Mission Valley Formation in northern San Diego County, comprising 15 families of bony fish and 9 families of chondrichthyans (Roeder 1991). Bressi Ranch is unique in preserving macro-vertebrate remains and in the preservation of a marine reptile, providing a rare glimpse of the middle Eocene Pacific Coast vertebrate fauna. The chelonioid appears to belong to a geographically widespread group of thin-shelled forms that reached both the southwestern and northern Pacific during the middle Eocene. It has not escaped our notice that the macrotaphonomic biases hitherto limiting our knowledge of turtles in the eastern North Pacific may similarly be obscuring the true geographic extent of other successful marine vertebrates during the Paleogene.
Acknowledgements.—For access to specimens, we thank Thierry Smith (IRSNB), Amanda Millhouse (National Museum of Natural History, Smithsonian Institution, Washington, D.C., USA). Reviews from Edwin Cadena (Universidad del Rosario, Bogotá, Colombia) and Andrew Gentry (Alabama School of Mathematics and Science, USA) improved our manuscript. We thank Gino Calvano (SDNHM) for careful fieldwork, Katie McComas (SDNMH) for photographic help, and Kesler Randall (SDNHM) for comments and specimen access.
References
Anquetin, J. 2012. Reassessment of the phylogenetic interrelationships of basal turtles (Testudinata). Journal of Systematic Palaeontology 10: 3–45. Crossref
Boyer, J.E. and Warme, J.E. 1975. Sedimentology and trace fossils in the Eocene Delmar Formation and Torrey Sandstone, California. In: D.W. Weaver, G.R. Hornaday, and A. Tipton (eds.), Future Energy Horizons of the Pacific Coast, 65–98. Pacific Sections AAPG-SEPM-SEG, Los Angeles.
Cadena, E.A. and Parham, J.F. 2015. Oldest known marine turtle? A new protostegid from the Lower Cretaceous of Colombia. PaleoBios 32: 1–42. Crossref
Clifton, H.E. 1979. Tidal channel deposits of middle Eocene age, Torrey Pines State Reserve, California. In: P. Abbott (ed.), Eocene Depositional Systems, 35–42. Pacific Section SEPM, Los Angeles.
David, L.R. 1946. Some typical Upper Eocene fish scales from California. Contributions to Paleontology, Carnegie Institute of Washington Publication 551: 47–112
Eisenberg, L.I. and Abbott, P.L. 1985. Eocene lithofacies and geologic history, northern San Diego County. In: P.L. Abbott (ed.), On the Manner of Deposition of the Eocene Strata in Northern San Diego County, 19–36. San Diego Association of Geologists, San Diego.
Evers, S.W. and Benson, R.B. 2019. A new phylogenetic hypothesis of turtles with implications for the timing and number of evolutionary transitions to marine lifestyles in the group. Palaeontology 62: 93–134. Crossref
Gentry, A.D. 2018. Prionochelys matutina Zangerl, 1953 (Testudines: Pan-Cheloniidae) from the Late Cretaceous of the United States and the evolution of epithecal ossifications in marine turtles. PeerJ 6: e5876. Crossref
Gentry, A.D., Ebersole, J.A., and Kiernan, C.R. 2019. Asmodochelys parhami, a new fossil marine turtle from the Campanian Demopolis Chalk and the stratigraphic congruence of competing marine turtle phylogenies. Royal Society Open Science 6: 1–12. Crossref
Givens, C.R. and Kennedy, M.P. 1976. Middle Eocene mollusks of northern San Diego County, California. Journal of Paleontology 50: 954–975.
Golz, D.J. 1976. Eocene Artiodactyla of southern California. Los Angeles County Museum of Natural History, Bulletin 26: 1–85.
Golz, D.J. and Lillegraven, J.A. 1977. Summary of known occurrences of terrestrial vertebrates from Eocene strata of southern California. Contributions to Geology, University of Wyoming 15: 43–65.
Grant-Mackie, J.A., Hill, J., and Gill, B.J. 2011. Two Eocene chelonioid turtles from Northland, New Zealand. New Zealand Journal of Geology and Geophysics 54: 181–194. Crossref
Haug, C., Nyborg, T. and Vega, F.J. 2013. An exceptionally preserved upogebiid (Decapoda: Reptantia) from the Eocene of California. Boletin de La Sociedad Geologica Mexicana 65: 235–248. Crossref
Hirayama, R. and Chitoku, T. 1996. Family Dermochelyidae (Superfamily Chelonioidea) from the Upper Cretaceous of North Japan. Transactions and Proceedings of the Palaeontological Society of Japan, New Series 184: 597–622.
Kear, B.P. 2006. Reassessment of Cratochelone berneyi Longman, 1915, a giant sea turtle from the Early Cretaceous of Australia. Journal of Vertebrate Paleontology 26: 779–783. Crossref
Kear, B.P. and Lee, M.S. 2006. A primitive protostegid from Australia and early sea turtle evolution. Biology Letters 2: 116–119. Crossref
Kear, B.P., Boles, W.E. and Smith, E.T. 2003. Unusual gut contents in a Cretaceous ichthyosaur. Proceedings of the Royal Society of London B 270: 206–208. Crossref
Köhler, R. 1995a. An Eocene turtle humerus (Dermochelyidae, Psephophorus) from New Zealand. Studia Geologica Salmanticensia 30: 101–106.
Köhler, R. 1995b. A new species of the fossil turtle Psephophorus (Order Testudines) from the Eocene of the South Island, New Zealand. Journal of the Royal Society of New Zealand 25: 371–384. Crossref
Lapparent de Broin, F. de, Murelaga, X., Pérez-García, A., Farrés, F., and Altimiras, J. 2018. The turtles from the upper Eocene, Osona County (Ebro Basin, Catalonia, Spain): new material and its faunistic and environmental context. Fossil Record 21: 237–284. Crossref
Moody, R.T.J. 1974. The taxonomy and morphology of Puppigerus camperi (Gray), an Eocene sea-turtle from Northern Europe. Bulletin of the British Museum (Natural History), Geology 25: 153–186.
Nesbitt, E.A. 1998. Marine fauna of the middle Eocene Tukwila Formation, King County. Washington Geology 26 (1): 13–19.
Nicholls, E.L. 1992. Note on the occurrence of the marine turtle Desmatochelys (Reptilia: Chelonioidea) from the Upper Cretaceous of Vancouver Island. Canadian Journal of Earth Science 29: 377–380. Crossref
Nicholls, E.L. and Meckert, D. 2002. Marine reptiles from the Nanaimo Group (Upper Cretaceous) of Vancouver Island. Canadian Journal of Earth Sciences 39: 1591–1603. Crossref
Orr, E.L. and Orr, W.N. 2009. Oregon Fossils. 2nd Edition. 300 pp. Oregon State University Press, Corvallis.
Parham, J.F., Hilton, R.P., and Göhre, E.S. 2003. The oldest turtle from California and other new records of Late Cretaceous sea turtles from the Chico Formation. PaleoBios 23: 24–28.
Prothero, D.R. 2001. Magnetic stratigraphy of the middle–upper Eocene Santiago Formation, Orange and San Diego Counties, California. In: D.R. Prothero (ed.), Magnetic Stratigraphy of the Pacific Coast Cenozoic, 107–118. Pacific Section SEPM, Los Angeles.
Robinson, P., Gunnell, G.F., Walsh, S.L., Clyde, W.C., Storer, J.E., Stucky, R.K., Froehlich, D.J., Ferrusquia-Villafranca, I., and McKenna, M.C. 2004. Wasatchian through Duchesnean biochronology. In: M.O. Woodburne (ed.), Late Cretaceous and Cenozoic Mammals of North America, 106–155. Columbia University Press, New York. Crossref
Roeder, M.A. 1991. A fossil fish fauna from the Upper Eocene Mission Valley Formation at the Lake Miramar Filtration Plant, San Diego, San Diego County, California. In: P.L. Abbot and J.A. May (eds.), Eocene Geologic History, San Diego Region, 179–180. Pacific Section SEPM, Los Angeles.
Scheyer, T.M. 2009. Conserved bone microstructure in the shells of long‐necked and short‐necked chelid turtles (Testudinata, Pleurodira). Fossil Record 12: 47–57. Crossref
Scheyer T.M., Danilov, L.G., Sukhanov, V.B., and Syromyatnikova, E.V. 2014. The shell bone histology of fossil and extant marine turtles revisited. Biological Journal of the Linnean Society 112: 701–718. Crossref
Squires, R.L. 1984. Megapaleontology of the Eocene Llajas Formation, Simi Valley, California. Contributions in Science, Natural History Museum of Los Angeles County 350: 1–76. Crossref
Squires, R.L. 1992. New morphologic and geographic data on the neritid gastropod Nerita (Theliostyla) triangulata Gabb, 1869, from the Eocene of the Pacific Coast of North America. The Veliger 35: 323–329.
Squires, R.L. 1999. Middle Eocene brackish-marine mollusks from the Matilija Sandstone at Matilija Hot Springs, Ventura County, southern California. Natural History Museum of Los Angeles County Contributions in Science 480: 1–30. Crossref
Squires, R.L. 2001. Additions to the Eocene megafossil fauna of the Llajas Formation, Simi Valley, southern California. Contributions in Science (Natural History Museum of Los Angeles County) 489: 1–40. Crossref
Walsh, S.L. 1991. Eocene mammal faunas of San Diego County. In: P.L. Abbot and J.A. May (eds.), Eocene Geologic History, San Diego Region, 161–193. Pacific Section SEPM, Los Angeles.
Walsh, S.L. 1996. Middle Eocene Mammalian faunas of San Diego County, California. In: D.R. Prothero and R.J. Emry (eds.), The Terrestrial Eocene–Oligocene Transition in North America, 75–119. Cambridge University Press, New York. Crossref
Warme, J.E. 1991. Delmar Formation and Torrey Sandstone as exposed along beach cliffs, Solana Beach, northern San Diego County. In: P.L. Abbott and J.A. May (eds.), Eocene Geologic History San Diego Region, 39–54. Pacific Section SEPM, Los Angeles.
Welton, B.J. 1972. Fossil sharks in Oregon. The Ore Bin 34 (10):161–170.Crossref
Weems, R.E. and Brown, K.M. 2017. More-complete remains of Procolpochelys charlestonensis (Oligocene, South Carolina), an occurrence of Euclastes (upper Eocene, South Carolina), and their bearing on Cenozoic pancheloniid sea turtle distribution and phylogeny. Journal of Paleontology 91: 1228–1243. Crossref
Weems, R.E. and Sanders, A.E. 2014. Oligocene pancheloniid sea turtles from the vicinity of Charleston, South Carolina, U.S.A. Journal of Vertebrate Paleontology 34 80–99. Crossref
Wiffen, J. 1981. The first Late Cretaceous turtles from New Zealand. New Zealand Journal of Geology and Geophysics 24: 293–299. Crossref
Wood, R.C., Johnson-Gove, J., Gaffney, E.S., and Maley, K.F. 1996. Evolution and phylogeny of leatherback turtles (Dermochelyidae) with descriptions of new fossil taxa. Chelonian Conservation and Biology 2: 266–286.
Zangerl R. 1953. The vertebrate fauna of the Selma Formation of Alabama. Part IV. The turtles of the family Toxochelyidae. Fieldiana, Geology Memoirs 3: 137–277. Crossref
Ashley W. Poust [apoust@sdnhm.org; ORCID: https://orcid.org/0000-0001-7955-613X ], San Diego Natural History Museum, Department of Paleontology, San Diego, California, 92101, USA; University of California Museum of Paleontology, University of California, Berkeley, California, 94720, USA.
Patricia A. Holroyd [pholroyd@berkeley.edu; ORCID: https://orcid.org/ 0000-0003-1292-6356 ], University of California Museum of Paleontology, University of California, Berkeley, California, 94720, USA.
Thomas A Deméré [tdemere@sdnhm.org; ORCID: https://orcid.org/0000-0001-5464-3528 ], San Diego Natural History Museum, Department of Paleontology, San Diego, California, 92101, USA.
Received 19 October 2022, accepted 26 January 2023, available online 6 March 2023.
Copyright © 2022 A.W. Poust et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Acta Palaeontol. Pol. 68 (1): 47–51, 2023
https://doi.org/10.4202/app.01034.2022