
The oldest putative prosbolid insect from Africa
ANDRÉ NEL and ROSE PREVEC
Nel, A. and Prevec, R. 2023. The oldest putative prosbolid insect from Africa. Acta Palaeontologica Polonica 68 (1): 95–102.
Karooprosbole magnifica gen. et sp. nov., oldest African cicadomorphan Prosbolidae, is described from middle Permian deposits (Abrahamskraal Formation, Karoo Supergoup) at the Onder Karoo locality in South Africa. It is compared to the genera currently included in Prosbolidae but also those in the Tettigarctidae: Cicadoprosbolinae. The limits and definitions of the two groups are discussed, because the new taxon has diagnostic characters of venation common to both groups. Also, some characters currently used to define these groups are clearly subject to homoplasy. This suggests that a phylogenetic analysis of the whole extant and fossil Cicadomorpha is needed to better define these families and verify their monophyly.
Key words: Hemiptera, Cicadomorpha, Prosbolidae, monophyly, Permian, South Africa.
André Nel [anel@mnhn.fr, ORCID: https://orcid.org/0000-0002-4241-7651 ], Institut de Systématique, Évolution, Biodiversité (ISYEB) Muséum national d’Histoire naturelle, CNRS, Sorbonne Université, EPHE, Université des Antilles, CP50, 57 rue Cuvier 75005 Paris, France.
Rose Prevec [r.prevec@am.org.za, ORCID: https://orcid.org/0000-0001-6674-1547 ], Department of Earth Sciences, Albany Museum, Makhanda, South Africa & Department of Botany, Rhodes University, Makhanda, South Africa.
Received 11 December 2022, accepted 27 January 2023, available online 6 March 2023.
Copyright © 2023 A. Nel and R. Prevec. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Prosbolidae Handlirsch, 1906, is a rather large family of Cicadomorpha that ranged from the late Carboniferous to the Cretaceous (Paleobiology database at https://paleobiodb.org/classic/basicTaxonInfo?taxon_no=191167). There is currently no phylogenetic analysis of the Cicadomorpha that encompasses both fossil and extant taxa, except the one proposed by Shcherbakov and Popov (2002: fig. 179) in which the Cicadina (Fulgoromorpha, Cicadomorpha, Prosboloidea) are polyphyletic and paraphyletic and are postulated to have given rise to the Heteroptera. Additionally, the superfamily Prosboloidea is paraphyletic in this phylogeny, and give rise to the Cicadomorpha and the Heteroptera. As a result, Shcherbakov and Popov (2002) did not propose apomorphic characters strictly applicable to the Prosbolidae. Hamilton (1992) included in the Prosboloidea the families Stenoviciidae Evans, 1956, and Serpentivenidae Shcherbakov, 1984 (included into Scytinopteroidea Handlirsch, 1906, by Szwedo, 2018), and Hylicellidae Evans, 1956 (in Hylicelloidea Evans, 1956, by Szwedo 2018). Szwedo (2018) only put the two families Prosbolidae and Maguviopseidae Shcherbakov, 2011, in the Prosboloidea. There is no recent revision of the family Prosbolidae. The positions of several prosbolid genera have changed, some more than once, and in some cases without justification (see below). Thus any new taxonomic work is not easy to achieve.
Here we describe from the Onder Karoo fossil locality in the south-western Karoo Basin, South Africa (Prevec et al. 2022), two new fossil forewings showing several characters of Prosbole Handlirsch, 1904 (type genus of the Prosbolidae), but also of the genus Cicadoprosbole Becker-Migdisova, 1947 (currently in Tettigarctidae: Cicadoprosbolinae, see Shcherbakov 2009). This means that the fossils are potentially of great interest for a future phylogenetic analysis of these insects.
The South African record of the Prosbolidae is rather modest when compared to that of Asia (Russian Federation, China), viz. a “Permocicada sp. indet.” (Pretorius et al. 2021), Permocicada thompsoni Van Dijk and Geertsema, 1999, Beaufortiscus dixi Riek, 1976, and Stenotegmocicada triclades van Dijk and Geertsema, 1999, all from the latest Permian age (Van Dijk and Geertsema 1999). Thus the new fossils are the oldest South African records for the family.
Institutional abbreviations.—AM, Albany Museum, Makhanda, South Africa.
Other abbreviations.—CuA cubitus anterior; CuP cubitus posterior; cua-cup intercubital crossvein; PCu postcubitus; RA radius anterior; RP radius posterior; r-m radial-median crossvein; ScP subcostal posterior.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:8448049A-5C93-4AB8-B19A-A87F6245E26C.
Material and methods
The fossils were collected under a South African Heritage Resources Agency (SAHRA) permit (ID# 2310 to RP), and the specimens have been accessioned to the palaeontological collections of the Department of Earth Sciences at the Albany Museum, Makhanda, Eastern Cape, South Africa. They were examined and photographed with a Zeiss Discovery V8 incident light microscope coupled with an Axiovision 208 camera. To improve photograph quality, we used polarizing filters and alcohol immersion. Drawings were made with Adobe Creative Cloud and Affinity graphics software. The fossils were excavated from a highly fossiliferous, olive-grey siltstone layer at the Onder Karoo locality (Sutherland District of the Northern Cape Province, South Africa; see Prevec et al. 2022). The host rocks are attributed to the lowermost Abrahamskraal Formation, Beaufort Group, Karoo Supergroup, close to the boundary with the underlying Waterford Formation of the Ecca Group. Regional vertebrate biostratigraphy, stratigraphic correlation and geochronology, as well as an age estimate obtained from U-Pb dating of zircons from a clay layer at the base of the fossiliferous deposit, have provided an age of 266–269 Ma for the fossil assemblage, dating it as Wordian (see Prevec et al. 2022).
We follow the wing venation terminology of Nel et al. (2012) and Bourgoin et al. (2015) for the Acercaria (including Hemiptera), as modified by Schubnel et al. (2020) concerning the presence of a PCu vein in the Pterygota. We prefer to use the term M for the median vein as there is no argument favoring the presence of a MA vs. MP in the Hemiptera.
Systematic palaeontology
Class Insecta Linnaeus, 1758
Order Hemiptera Linnaeus, 1758
Infra-order Cicadomorpha Latreille, 1802
Family Prosbolidae Handlirsch, 1906
Genus Karooprosbole nov.
Zoobank LSID: urn:lsid:zoobank.org:act:694C8DC3-4475-42B9-BEA 9-17369F1F01C7.
Type species: Karooprosbole magnifica sp. nov., by monotypy; see below.
Etymology: Named after the Onder Karoo locality and the genus name Prosbole. Gender feminine.
Diagnosis.—As for the monotypic type species.
Karooprosbole magnifica sp. nov.
Figs. 1, 2.
Zoobank LSID: urn:lsid:zoobank.org:act:F23591E1-6470-4ECA-948 C-86EED0FAB4A7.
Etymology: Named after the wonderful preservation of the material, with color patterns.
Type material: Holotype AM14858a, b, adpression of a tegmen, part and counterpart; paratype AM11270a, b, adpression of a tegmen, part and counterpart, from the type locality and horizon.
Type horizon: Wordian Stage, Guadelupian, Permian.
Type locality: Low road-cutting and associated abandoned quarry, Onder Karoo locality, Sutherland District of the Northern Cape Province, South Africa.
Diagnosis.—Tegmen characters only. Presence of irregular veinlets in distal part of costal area; nodal line crossing RP midway between base of RP and its first fork, and crossing M basad its first fork; basal part of CuA curved; posterior branch of RA with a series of weak anterior branches; base of RP distinctly closer to base of M than to fork of RA, but not very close to base of M.
Description.—Holotype AM14858a, b: A complete forewing (tegmen), 13.8 mm long, 5.0 mm wide; ScP forming a curved vein appressed to R+M+CuA ending into R just distad point of separation between R and M+CuA; a very broad area between radius and costal margin, 1.2 mm in widest part, with transverse, darkened, and curved veinlets; division of radius into RA and RP 5.7 mm from wing base; base of RP far basad distal fork of RA (2.3 mm) and rather far distad base of M (2.0 mm); a weak crossvein between RA and RP closing a triangular cell; RA(+ScP) straight, with a terminal fork into a rather long and oblique anterior branch (re-emergence of ScP?) along prominent nodal break and a longer posterior branch, defining an elongate cell along costal margin; posterior branch of RA with four very weak anterior branches; RP ending distally with two closely parallel terminal branches, both anteriorly directed; M basally connected to CuA and radius, M and CuA diverging from R 3.3 mm from wing base, stem of M 3.5 mm long; M ending with three main branches, anterior-most branch of M forked twice, second branch forked again; first and second branches defining two closed cells; an intra-median crossvein between second and third branch of M; basal part of CuA strongly curved, 3.4 mm long, with a distal fork; anterior branch of CuA fused for a short length with M and separated again distally; a crossvein between RP and M closing an elongate and narrower cell; a nodal line; a darkened spot between CuA and M; nodal line crossing CuA at its first fork, M slightly basad, 0.3 mm its first fork, and RP 1.3 mm distad base of RP and 2.1 mm basad crossvein r-m; cua-cup curved and long, 1.1 mm long; CuP straight, weaker than CuA; PCu straight; anal area with a long anal vein distally ending into PCu and two sigmoidal veinlets between it and PCu.
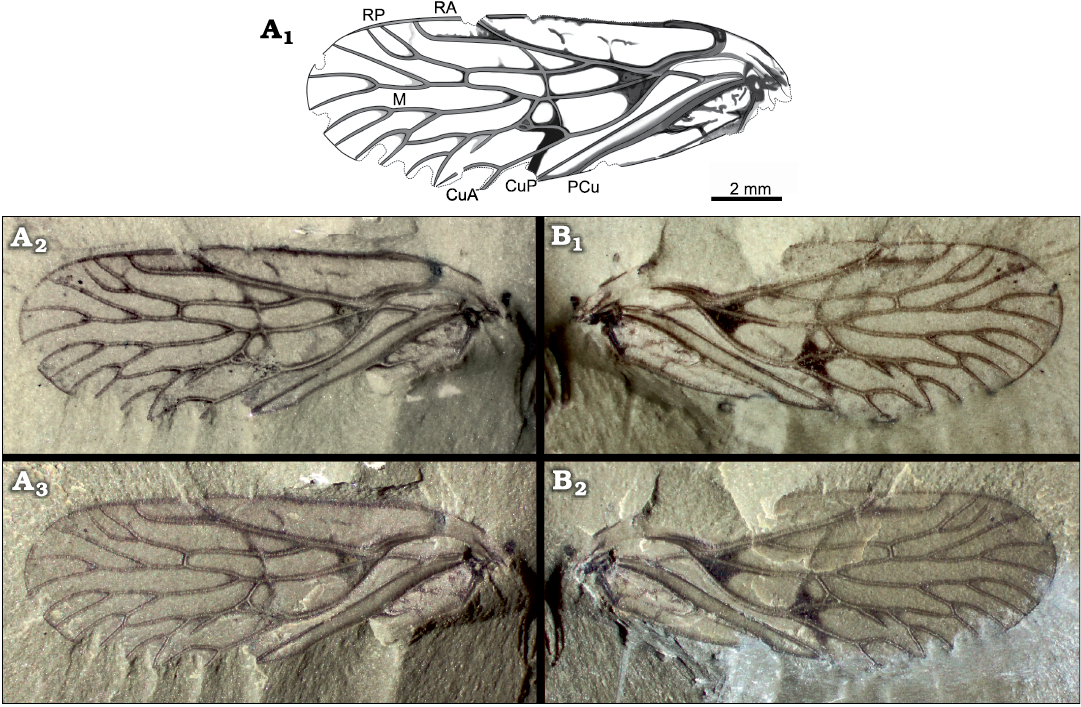
Fig. 1. Cicadomorph insect Karooprosbole magnifica gen. et sp. nov., holotype, Guadelupian, Onder Karoo, South Africa. A. AM14858a, composite drawing (A1), photograph under alcohol (A2), with incident light (A3). B. AM14858b, photograph under alcohol (B1), with incident light (B2).
Paratype AM11270a, b: A nearly complete forewing (tegmen), with anal area and CuP lacking in main part, wing ca. 13.8 mm long, 5.1 mm wide; venation nearly identical to that of holotype except for the presence of a crossvein between anterior and median branches of M, absence of fusion of distal branches of median and posterior branches of M, and presence of a distal crossvein between M and CuA; area between radius and costal margin 1.3 mm in widest part; base of RP far basad distal fork of RA (2.1 mm) and rather far distad base of M (1.5 mm); stem of M 3.6 mm long; basal part of CuA 3.4 mm long, a weak nodal line visible through dilatations of RP, M, and CuA and a darkened spot between CuA and M; nodal line crossing CuA at its first fork, M slightly basad, 0.4 mm its first fork, and RP 1.4 mm distad base of RP and 1.8 mm basad crossvein r-m; cua-cup curved and long, 1.1 mm long, weaker than CuA; a small part of CuP present.
Stratigraphic and geographic range.—Type locality and horizon only.
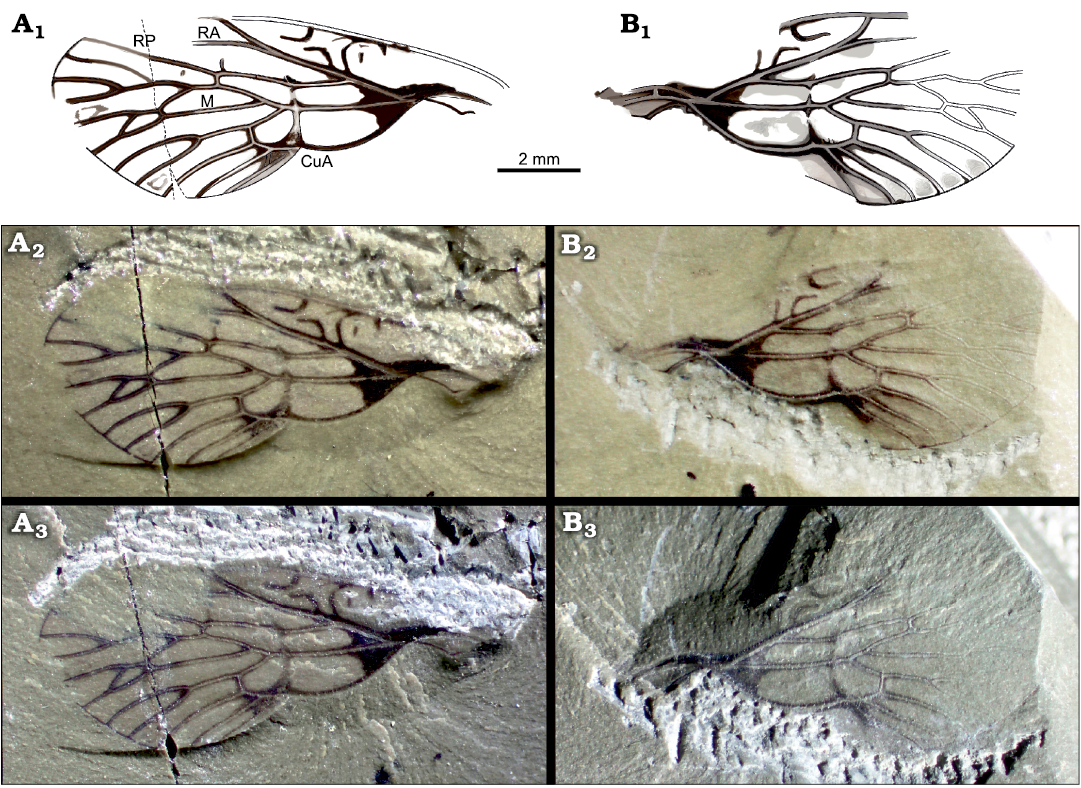
Fig. 2. Cicadomorph insect Karooprosbole magnifica gen. et sp. nov., paratype, Guadelupian, Onder Karoo, South Africa. A. AM11270b, drawing (A1), photographed under alcohol (A2), with incident light (A3). B. AM1270a, drawing (B1), photographed under alcohol (B2), with incident light (B3).
Discussion
The two forewings under description can be attributed to the same genus and species because they share the most crucial forewing characters in dimensions, venation, and pattern of coloration. They only differ in the number and disposition of the distal-most branches of M and CuA, characters that can vary greatly even in the extant Cicadoidea. They have a pattern of venation similar to those of the Prosbolidae: large wings, broad costal area, presence of a nodal line, and can be attributed with confidence to the Cicadomorpha. If we use Shcherbakov and Popov (2002: fig. 179) strategy to discriminate the cicadomorphan “groups”, affinities with the Pereborioidea would be excluded because their forewings have a “prolific” venation and apical portion of ScP with longitudinal branches. The Dysmorphoptilidae are excluded because the tegmen in the new fossils is not punctate. The Cercopoidea are also excluded for the same reason as Dysmorphoptilidae and because the ScP is fused to R. The weakly defined nodal flexion line and the broad costal area exclude the Cicadoidea. The Membracoidea and Hylicelloidea are excluded on the basis of the broad tegmen seen in the new specimens, even if some Mesojabloniidae (Mesojablonia kukalovae Storozhenko, 1992, Triassic of Kyrgyzstan) have broad tegmina resembling that of the new fossils, but without any nodal line (Shcherbakov 2011).
The Palaeontinoidea would be excluded because their forewings have a hypertrophied nodal flexion line (Shcherbakov and Popov 2002), but the Dunstaniidae have a nodal line even less pronounced than in Palaeontinidae. Shcherbakov (1984) proposed the “transverse m-cu (the only one or the first) long passing in the basal part along the nodal line and/or in the middle part, directly longitudinally” as a crucial character for the Palaeontinoidea. In the new fossils, the m-cu does not run along the nodal line and is not directed longitudinally in its mid part, excluding affinities with this group.
Evans (1956: 223) indicated that Cicadoprosbole (Jurassic of Kyrgyzstan) differs other prosbolid genera in the position of the nodal line, crossing M well distad its first fork and RP well distad its base in the former vs. crossing M just basad or at first fork and RP at its base in the latter. The new forewings have the nodal line crossing M basad its first fork as in Prosbolidae, but they differ from the Prosbolidae in the nodal line crossing RP distinctly distad its base, in a position similar to that in Cicadoprosbole, viz. ca. midway between base of RP and r-m crossvein, much more distal than in the Prosbolidae, for instance Prosbole affinis (Permian of the Russian Federation) has the nodal line crossing RP slightly distad its base.
Among the other Cicadoprosbolinae, the nodal lines in Burmaprosbole Qiao, Zhang, Zhang, Ren, and Yao, 2021 (mid-Cretaceous Burmese amber), Shuraboprosbole Becker-Migdisova, 1947 (Jurassic of China, Kyrgyzstan, UK, junior synonym Paraprosbole Whalley, 1985, after Chen et al. 2016), and Sanmai Chen, Zhang, Wang, Zheng, and Zheng, 2016 (Jurassic of China) are in the same situation as in Cicadoprosbole, with the nodal line crossing M at its fork or distal to it (Hamilton 1990; Wang and Zhang 2009; Qiao et al. 2021). Architettix Hamilton, 1990 (Cretaceous of Brazil) has the nodal line crossing RP at its base, as in the Prosbolidae, but crossing M well distad its fork (Hamilton 1990). The genus Diphtheropsis Martynov, 1937 (Jurassic of Kyrgyzstan) is based on a poorly preserved fossil tegmen, but with M forked at the nodal line, unlike the new fossils (Martynov 1937; Shcherbakov 2009). Elkinda Shcherbakov, 1988 (Cretaceous of Russian Federation) has the nodal line crossing RP much closer to r-m than to the base of RP (Shcherbakov 1988). Hylaeoneura Lameere and Severin, 1897 (Cretaceous of Belgium), is based on the distal half of a tegmen, with the nodal line not preserved but with a pattern of the crossveins between main veins strongly different from that of the new fossils (Lameere and Severin 1897; Jarzembowski 2002: fig. A1). Turutanovia Becker-Migdisova, 1949 (Cretaceous of Mongolia, Jurassic of Kazakhstan), is based on an incomplete tegmen showing only the part of the nodal line between the posterior branch of M and CuA. Thus it is clearly crossing M distad its basal fork (Becker-Migdisova 1949: fig. 19).
The presence of anterior branches of the posterior branch of RA would correspond to the situation in the Cicadoprosbolini sensu Shcherbakov (2009), but Qiao et al. (2021) indicated that the situation is not stable in this tribe as Burmaprosbole has a simple posterior branch of RA. Also the prosbolid Dictyoprosbole Martynov, 1935, has some anterior branches on this vein. It also has the nodal line crossing RP in a quite distal position, close to midway between the base of RP and r-m (Martynov 1935).
Among the families that have been put in the Prosboloidea in the past, the new fossils strongly differ from the Eoscarterellidae Evans, 1956 (currently in Dysmorphoptiloidea, see Szwedo 2018), the Maguviopseidae Shcherbakov, 2011, in having a nodal line (Evans 1956; Shcherbakov 2011). The Serpentivenidae Shcherbakov, 1984 have the base of RP opposite the fork of RA (Becker-Migdisova 1962; Shcherbakov 1984). The Stenoviciidae Evans, 1956, also lack a nodal line and the base of RP is opposite the fork of RA (Evans 1956). The Hylicellidae (especially the type genus Hylicella Evans, 1956, Triassic of Australia) share with the new fossils what could be a branch of M that merges into the anterior branch of CuA (appearing as a long oblique vein between M and CuA) (Evans 1956). Lambkin (2020: 525) indicated that a “suggestion” of nodal line exists in these bugs, thus less pronounced than in the new fossil. He also added that the pre-nodal area is finely to somewhat coarsely punctate, and that RP is simple in the Hylicellidae, unlike in the new fossil. Also, the basal cell situated between R+M+CuA, CuP, and cua-cup, is much narrower in the Hylicellidae than in the new fossil. The Hylicellidae are all Mesozoic except for Tychtoscytina kusnezkiensis Becker-Migdisova, 1952 (Permian of the Russian Federation), based on a rather poorly preserved tegmen but that shows a simple RP too (Becker-Migdisova 1952). Their relationships with the Prosboloidea remain to be clarified.
Among the Prosbolidae, Evanscicada Becker-Migdisova, 1962 (Permian of the Russian Federation), Prosbolecicada Pinto, 1987 (Permian of Brazil), and Sojanoneura Martynov, 1928 (Permian of the Russian Federation) have the nodal line crossing M well distad its first fork and RP crossing it just distad its base, unlike the new fossils (Becker-Migdisova 1962; Pinto 1987). Sojanoneura also has the base of RP close to the fork of RA (Becker-Migdisova 1940). Prosbole has the nodal line crossing M at its fork but crossing RP near its base (Becker-Migdisova 1940; Carpenter 1992; Prokop et al. 2015). Kaltanopsis Becker-Migdisova, 1961 (Permian of the Russian Federation), Kaltanetta Becker-Migdisova, 1961 (Permian of the Russian Federation), Orthoscytina Tillyard, 1926 (Permian of Australia, Russian Federation, and South Africa), Elliptoscarta Tillyard, 1926 (Permian of Australia), Homaloscytina Tillyard, 1926 (Permian of Australia), Permobrachus Evans, 1943 (Permian of Australia), Permojassus Tillyard, 1926 (Permian of Australia), and Cicadopsylla Martynov, 1931 (Permian of the Russian Federation, based on an incomplete tegmen) have no clear nodal line, but they differ from the new fossils in the fork of M distad that of CuA and a reduced number of branches of RP and M (Tillyard 1926; Martynov 1931; Evans 1943; Becker-Migdisova 1961; Carpenter 1992). Permojassus also differs from the new fossils in the base of RP close to the fork of RA. Permobrachus has a very different shape of CuA with very distal branches. Kaltanopsis has no fork of RA, a character suggesting it does not belong to the Prosbolidae. Prosbolidinella Martins-Neto and Rohn, 1996, is based on a poorly preserved tegmen, with no indication of a nodal line. It differs from the new fossils, in having the base of RP very close to that of M and the CuA being nearly straight in the part basad its fork (Martins-Neto and Rohn 1996). Stenoscytina Tillyard, 1926, and Stenotegmocicada Van Dijk and Geertsema, 1999, also have no indication of a nodal line, and differ from the new fossils in the base of RP being very close to that of M (Van Dijk and Geertsema 1999).
Dictyoprosbole (Permian of the Russian Federation) shares with the new fossils the presence of veinlets in the costal area, but strongly differs from them with the presence of a net of cells all over the tegmen, and with a nodal line that crosses M at its fork.
Austroprosbole Evans, 1943 (Permian of Australia), and Austroprosboloides Riek, 1973 (Permian of South Africa), were originally included in the Prosbolidae (Evans 1943; Riek 1973), were then moved to the Dunstaniidae by Shcherbakov (1984), and finally returned into the Prosbolidae by Carpenter (1992), each time without argument. The venation of the tegmina in these taxa clearly bears a closer resemblance to that of Prosbole than that of Dunstania, especially in the shape of the nodal line and of RP. They differ from the new fossils in the nodal line crossing M at its fork. Neurobole Riek, 1976 (Permian of South Africa) is based on a fragment of the costo-distal part of a wing, showing no nodal line and no costal area. Its RA and RP have numerous branches, unlike the other Prosbolidae (Riek 1976). It probably does not belong to this family, as indicated by Carpenter (1992).
Orthoprosbole Martynov, 1935 (Permian of the Russian Federation), Mitchelloneura Tillyard, 1921, and Anomaloscytina Davis, 1942 (both Permian of Australia) are based on hind wings that cannot be accurately compared to the new fossils except that they have many more branches of RP, M, and CuA, and far fewer crossveins between them (Tillyard 1921; Martynov 1935; Davis 1942; Carpenter 1992). Pervestigia Becker-Migdisova, 1961 (Permian of France and of Russian Federation) is only known from hind wings. It differs from the new fossils in having the base of RP close to the fork of RA (Becker-Migdisova 1961). Mitchelloneura Tillyard, 1921 (Permian of Australia) is also based on a hind wing. It differs from the new fossils in the presence of more branches of RP (Tillyard 1921).
Archeglyphis Martynov, 1930 (Carboniferous of Russian Federation), is based on a very incomplete distal part of a tegmen, without any indication of a nodal line. Its attribution to the Prosbolidae is poorly grounded. It differs from the new fossils in the absence of crossveins between main veins and a very different shape of the costal area, which is not greatly expanded (Martynov 1930: fig. 10).
Beaufortiscus Riek, 1976 (upper Permian of Natal) differs from the new fossils in the absence of a posterior branch of RA (a character quite curious for a Prosbolidae, suggesting it does not belong to this family) and the presence of far fewer branches of M. Otherwise, the course of the nodal line is similar (Riek 1976; Van Dijk and Geertsema 1999).
Falsia Becker-Migdisova, 1946, seems to be a nomen nudum. Lariojaprosbole Martins-Neto and Gallego, 2001 (Triassic of Argentina), is based on the basal third of a tegmen, with the costal area and the nodal line not preserved. If the interpretation of Martins-Neto and Gallego (2001) is accurate, it has a fork of M in a very basal position, unlike all the other Prosbolidae. This taxon probably does not belong to this family.
Permocicada Martynov, 1928 (Jurassic of China, Permian of Russian Federation and South Africa) differs from the new fossils in the very distal position of the base of RP, close to the fork of RA, the absence of veinlets in the costal area and the absence of intra-median crossveins (Martynov 1928; Becker-Migdisova 1940; Van Dijk and Geertsema 1999). The exact position of the nodal line is somewhat uncertain (Martynov 1928): Becker-Migdisova (1940) did not figured it in any representative of the genus except in Permocicada pusilla Becker-Migdisova, 1940, where it is in a very distal position. Permocicadopsis Becker-Migdisova, 1940 (Permian of Russian Federation) differs from the new fossils in the same characters as for Permocicada. Permodiphtheroides Martynov, 1928 (Permian of Russian Federation) is based on a poorly preserved fossil, but the base of RP is close to the fork of RA, unlike in the new fossils (Martynov 1928). Permodiphthera Tillyard, 1926 (Permian of Australia) differs from the new fossils in the base of RP being distinctly closer to the fork of RA than to that of M, and in the nodal line crossing RP close to its base and M at its fork (Tillyard 1926). Permoglyphis Tillyard, 1926 (Permian of Australia) has also the base of RP close to the fork of RA and the nodal line crossing RP close to its base, but as in the new fossils, the nodal line crosses M quite basad its fork.
Sinisbole Lin, 1986 (Triassic of China) is based on an incomplete distal third of a tegmen, apparently not showing a nodal line. If it shares with the new fossils the base of RP closer to that of M than to the fork of RA, but not extremely close to it, it differs from the latter in the very long and straight CuA basad its fork (Lin 1986).
The new fossils differ from the Permian South African genus Austroprosboloides Riek, 1973 (Permian of South Africa, a taxon originally attributed to the “Cicadoprosbolidae”, transferred into the palaeontinoid Dunstaniidae by Shcherbakov (1984, 2009), but put in the Prosbolidae by Carpenter (1992) without any revision) in the longer and distinctly oblique anterior branch of RA, and the branched posterior branch of RA, the nodal line basal to first fork of M and basal to crossvein m-cua, and m-cua quite short (Riek 1973).
Conclusions
The new fossil taxon strongly differs from all the genera currently placed in Prosbolidae and Cicadoprosbolinae, and therefore can be attributed to a new genus and species. Nevertheless the lack of a robust phylogenetic analysis of the whole extant and fossil Cicadomorpha, plus the weakness of the characters used to define and separate the Prosbolidae from the other families in this clade, render uncertain the attributions of all the genera currently in this family (except for Prosbole itself). Thus the attribution of the new genus and species to the Prosbolidae rather than to the Cicadoprosbolinae is only tentative. The position of the nodal line of Karooprosbole magnifica gen. et sp. nov. is “intermediate” between the two groups, calling into question their current delimitation. A phylogenetic analysis of the Prosboloidea and other Cicadomorpha is necessary. The verification of the monophyly of the Prosbolidae is particularly important to our understanding of insect evolution because this family is supposed to have survived the Permian/Triassic crisis and the Carnian pluvial episode, disappearing during the Cretaceous.
Acknowledgements
We sincerely thank Jacek Szwedo (University of Gdańsk, Poland) and an anonymous referee for their very useful comments on the first version of the paper. Specimen AM11270 was prepared by Romain Garrouste (Muséum national d’Histoire naturelle, Paris, France). We acknowledge the following funding bodies for their contributions: Genus (Department of Science and Innovation-National Research Foundation, Centre of Excellence in Palaeosciences) grants CoE2016-600, CoE2016-496, CoE2017-059 (RP); National Research Foundation, African Origins Platform grants 98822 and 117685 (RP).
References
Becker-Migdisova, E.E. 1940. Fossil Permian cicadas of the family Prosbolidae from the Soyana River [in Russian]. Trudy Paleontologičeskogo Instituta Akademii Nauk SSSR 11: 1–79.
Becker-Migdisova, E.E. 1946. Contributions to the knowledge of comparative morphology of the Recent and Permian Homoptera [in Russian]. Izvestiâ Akademii nauk SSSR 6: 741–766.
Becker-Migdisova, E.E. 1947. Cicadoprosbole sogutensis g. n., sp. n. A transitional form between the Permian Prosbolidae and the Recent Cicadidae [in Russian]. Doklady Akademii nauk SSSR 55 (5): 441–444.
Becker-Migdisova, E.F. 1949. Mesozoic Homoptera from Central Asia [in Russian]. Trudy Paleontologičeskogo Instituta Akademii Nauk SSSR 40: 1–68.
Becker-Migdisova, E.E. 1952. New Homoptera from the Permian of Kuzbass and notes on the Ipsviciidae] [in Russian]. Trudy Paleontologičeskogo Instituta Akademii Nauk SSSR 40: 177–186.
Becker-Migdisova, E.E. 1961. Order Homoptera [in Russian]. In: B.B. Rohdendorf, E.E. Becker-Migdisova, O.M. Martynova, and A.G. Šarov (eds.), Paleozojskie nasekomye Kuzbassa. Trudy Paleontologičeskogo Instituta Akademii Nauk SSSR 85: 286–393.
Becker-Migdisova, E.E. 1962. Orders Homoptera and Heteroptera. In: B.B. Rohdendorf (ed.), Osnovy paleontologii. Traheinye i helicerovye, 162–194, 199–224. Izdatel’stvo Akademii Nauk SSSR, Nauka, Moscow. [English translation: B.B. Rohdendorf (ed.) 1991. Fundamentals of Palaeontology 9. Arthropoda, Tracheata, Chelicerata, 216–317. Amerind Publ. Co., Moscow]
Bourgoin, T., Wang, R.-R., Asche, M., Hoch, H., Soulier-Perkins, A., Stroinski, A., Yap, S., and Szwedo, J. 2015. From micropterism to hyperpterism: recognition strategy and standardized homology-driven terminology of the forewing venation patterns in planthoppers (Hemiptera: Fulgoromorpha). Zoomorphology 134: 63–77. Crossref
Carpenter, F.M. 1992. Superclass Hexapoda. In: R.C. Moore and R.L. Kaesler (eds.), Treatise on Invertebrate Paleontology. Part R. Arthropoda 4, Volume 4, i–ii + 279–655. The Geological Society of America and the University of Kansas, Boulder.
Chen, J., Zhang, H-C., Wang, B., Zheng, Y., and Zheng, X.-T. 2016. New Jurassic tettigarctid cicadas from China with a novel example of disruptive coloration. Acta Palaeontologica Polonica 61: 853–862. Crossref
Davis, C. 1942. Hemiptera and Copeognatha from the Upper Permian of New South Wales. Proceedings of the Linnean Society of New South Wales 67 (3–4): 111–122.
Evans, J.W. 1943. Upper Permian Homoptera from New South Wales. Records of the Australian Museum 21: 180–198. Crossref
Evans, J.W. 1956. Palaeozoic and Mesozoic Hemiptera. Australian Journal of Zoology 4: 165–258. Crossref
Hamilton, K.G.A. 1990. Homoptera. In: D.A. Grimaldi (ed.), Insects From the Santana Formation, Lower Cretaceous of Brazil. Bulletin of the American Museum of Natural History 195: 82–122.
Hamilton, K.G.A. 1992. Lower Cretaceous Homoptera from the Koonwarra fossil bed in Australia with a new superfamily and synopsis of the Mesozoic Homoptera. Annals of the Entomological Society of America 85: 423–430. Crossref
Handlirsch, A. 1904. Über einige Insektenreste aus der Permformation Russlands. Mémoires de l’Académie impériale des sciences de Saint-Petersbourg 16 (5): 1–7.
Handlirsch, A. 1906–1908. Die fossilen Insekten und die Phylogenie der rezenten Formen. Ein Handbuch für Paläontologen und Zoologen. Lieferung 8. 1430 pp. Wilhelm Engelman, Leipzig. Crossref
Jarzembowski, E.A. 2002. Visit to the Natural History Museum of Belgium. Proceedings of the Geologist’s Association 113: 175–179. Crossref
Lambkin, K.J. 2020. Revision of the Hylicellidae of the Late Triassic of Queensland (Hemiptera: Cicadomorpha: Hylicelloidea). Zootaxa 4790: 525–539. Crossref
Lameere, A. and Severin, G. 1897. Les insectes de Bernissart. Annales de la Société Entomologique de Belgique 41: 35–38.
Latreille, P.A. 1802. Histoire naturelle, générale et particulière des Crustacés et des insectes. Ouvrage faisant suite aux œuvres de Leclerc de Buffon et partie du cours complet d’Histoire naturelle rédigé par C.S. Sonnini. Tome 3. Familles naturelles des genres. xii + 467 pp. F. Dufart, Paris. Crossref
Lin, Q.-B. 1986. Early Mesozoic fossil insects from South China [in Chinese, with an English summary]. Palaeontologia Sinica B 170 (21): i–iii + 1–112.
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. 824 pp. Laurentius Salvius, Stockholm. Crossref
Martins-Neto, R.G. and Gallego, O.F. 2001. The Triassic insect fauna from Argentina. 4. Glosselytrodea and complements on Auchenorrhyncha. 1 Simposio Brasileiro de Paleoarthropodologia and 1rst International Meeting on Paleoarthropodology, Universidade de Vale do Rio dos Sinos, Acta Geologica Leopoldensia, Estudos Tecnologicos 24: 105–114.
Martins-Neto, R.G. and Rohn, 1996. Primeiro registro de inseto na Formação Rio do Rasto, Bacia do Paraná, com descrição de novo táxon. Revista Geociências 15: 243–251.
Martynov, A.V. 1928. Permian fossil insects of North-East Europe [in Russian]. Trudy Geologičeskogo Muzeâ Akademii Nauk SSSR 4: 1–118.
Martynov, A.V. 1930. Permian fossil insects from Tikhije Gory. Order Miomoptera. 2. The families Lemmatophoridae and Palaeomantidae [in Russian]. Izvestiâ Akademii Nauk SSSR 7: 1115–1134.
Martynov, A.V. 1931. New fossil insects from Tikhie Gory. 2. Neoptera (excluding Miomoptera) [in Russian]. Trudy Geologičeskogo Muzeâ, Akademii Nauk SSSR 8: 149–212.
Martynov, A.V. 1935. Permian fossil insects from Arkhangelsk district. Part 5. Homoptera [in Russian]. Trudy Paleozoologičeskogo Instituta Akademii Nauk SSSR 4: 1–35.
Martynov, A.V. 1937. Liassic insects from Shurab and Kisyl-Kiya [in Russian with English summary]. Trudy Paleontologičeskogo Instituta Akademii nauk SSSR 7: 1–179.
Nel, A., Prokop, J., Nel, P., Grandcolas, P., Huang, Di-ying, Roques, P., Guilbert, E., Dostál, O., and Szwedo, J. 2012. Traits and evolution of wing venation pattern in paraneopteran insects. Journal of Morphology 273: 480–506. Crossref
Pinto, I.D. 1987. Permian insects from Parana basin, South Brazil. 5. Homoptera–Cicadoidea. Pesquisas (Zoologia) 19: 13–22. Crossref
Pretorius, A.I., Labandeira, C.C., Nel, A., and Prevec, R. 2021. Latest Permian insects from Wapadsberg Pass, southern Karoo Basin, South Africa. Austral Entomology 60: 560–570. Crossref
Prevec, R., Nel, A., Day, M.O., Muir, R.A., Matiwane, A., Kirkaldy, A.P., Moyo, S., Staniczek, A., Cariglino, B., Maseko, Z., Kom, N., Rubidge, B.S., Garrouste, R., Holland, A., and Barber-James, H.M. 2022. New South African Lagerstätte reveals middle Permian Gondwanan lakeshore ecosystem in exquisite detail. Communication Biology 5: article 1154. Crossref
Prokop, J., Szwedo, J., Lapeyrie, J., Garrouste, R., and Nel, A. 2015. New Middle Permian insects from Salagou Formation of the Lodève Basin in southern France (Insecta: Pterygota). Annales de la Société Entomologique de France (N.S.) 51: 14–51. Crossref
Qiao, D., Zhang, W.-W., Zhang, X., Ren, D., and Yao, Y.-Z. 2021. New genus and species of Tettigarctidae (Hemiptera: Cicadomorpha: Cicadoidea) from the mid-Cretaceous amber of northern Myanmar. Cretaceous Research 126: 104900. Crossref
Riek, E.F. 1973. Fossil insects from the Upper Permian of Natal. Annals of the Natal Museum 21: 513–532.
Riek, E.F. 1976. New Upper Permian insects from Natal, South Africa. Annals of the Natal Museum 22: 755–789.
Schubnel, T., Desutter-Grandcolas, L., Legendre, F., Prokop, J., Mazurier, A., Garrouste, R., Grandcolas, P., and Nel, A. 2020. To be or not to be: postcubital vein in insects revealed by microtomography. Systematic Entomology 45: 327–336. Crossref
Shcherbakov, D.Y. 1984. Systematics and phylogeny of Permian Cicadomorpha (Cimicida and Cicadina). Paleontological Journal 18: 87–97.
Shcherbakov, D.Y. 1988. New cicadas (Cicadina) from the Late Mesozoic of Transbaikalia. Paleontological Journal 22: 52–63.
Shcherbakov, D.Y. 2009. Review of the fossil and extant genera of the cicada family Tettigarctidae (Hemiptera: Cicadoidea). Russian Entomological Journal 17: 343–348.
Shcherbakov, D.Y. 2011. New and little-known families of Hemiptera Cicadomorpha from the Triassic of Central Asia—early analogs of treehoppers and planthoppers. Zootaxa 2836: 1–26. Crossref
Shcherbakov, D.Y. and Popov, Y.A. 2002. Superorder Cimicidea Laicharting, 1781 order Hemiptera Linné, 1758. The bugs, cicadas, plantlice, scale insects, etc. (= Cimicida Laicharting, 1781, = Homoptera Leach, 1815 + Heteroptera Latreille, 1810). In: A.P. Rasnitsyn, and D.L.J. Quicke (eds.), History of Insects, 143–157. Kluwer Academic Publishers, Dordrecht.
Storozhenko, S.Yu. 1992. New Mesozoic grylloblattid insects (Grylloblattida) from Central Asia. Paleontological Journal 26: 67–75.
Szwedo, J. 2018. The unity, diversity and conformity of bugs (Hemiptera) through time. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 107: 109–128. Crossref
Tillyard, R.J. 1921. Two fossil insect wings in the collection of Mr John Mitchell, from the Upper Carboniferous of New-Castle belonging to the order Homoptera. Proceedings of the Linnean Society of New South Wales 46: 413–422. Crossref
Tillyard, R.J. 1926. Upper Permian insects of New South Wales. 1. Introduction and the order Hemiptera. Proceedings of the Linnean Society of New South Wales 51: 1–30.
Van Dijk, D.E. and Geertsema, H. 1999. Permian insects from the Beaufort Group of Natal, South Africa. Annals of the Natal Museum 40: 137–171.
Wang, B. and Zhang, H.-C. 2009. Tettigarctidae (Insecta: Hemiptera: Cicadoidea) from the Middle Jurassic of Inner Mongolia, China. Geobios 42: 243–253. Crossref
Whalley, P.E.S. 1985. The systematics and palaeogeography of the Lower Jurassic insects of Dorset, England. Bulletin of the British Museum (Natural History), Geology 39 (3): 107–187.
Acta Palaeontol. Pol. 68 (1): 95–102, 2023
https://doi.org/10.4202/app.01046.2022