
Eocene phymaraphiniid demosponges from South Western Australia: filling the gap
ANDRZEJ PISERA, MARIA ALEKSANDRA BITNER, and JANE FROMONT
Pisera, A., Bitner, M.A., and Fromont, J. 2023. Eocene phymaraphiniid demosponges from South Western Australia: filling the gap. Acta Palaeontologica Polonica 68 (2): 261–272.
We describe two new genera of phymaraphiniid lithistid sponges Twertupia gen. nov. and Pickettispongia gen. nov. from the upper Eocene Pallinup Formation of South Western (SW) Australia based on new, rich and very well preserved material. Type material of these two genera, earlier described from poorly preserved material, were originally attributed to Thamnospongia subglabra and Stachyspongia neoclavatela (in case of species of Twertupia), and to Discoderma tabelliformis (case of species of Pickettispongia). This is the first record of bodily preserved phymaraphiniid sponges from Eocene rocks, as well as from the southern hemisphere. We discuss extant and fossil representatives of Phymaraphiniidae and their geographical distribution, concluding that the present day occurrences of these sponges are the result of a much larger Mesozoic Tethyan distribution.
Key words: Lithistid sponges, Demospongiae, Phymaraphiniidae, Eocene, extant, Australia.
Andrzej Pisera [apis@twarda.pan.pl; ORCID: https://orcid.org/0000-0001-7112-6063 ] and Maria Aleksandra Bitner [bitner@twarda.pan.pl; ORCID: https://orcid.org/0000-0002-2864-2967 ], Institute of Paleobiology, Polish Academy of Sciences, ul. Twarda 51/55, 00-818 Warszawa, Poland.
Jane Fromont [janefromont@iinet.net.au; ORCID: https://orcid.org/0000-0002-8887-4452 ], Western Australian Museum, Perth, Australia.
Received 3 January 2023, accepted 7 April 2023, available online 9 May 2023.
Copyright © 2023 A. Pisera et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The first report and descriptions of sponges from the Eocene rocks (Pallinup Formation, Bremer Basin; Fig. 1) of SW Australia were by Chapman and Crespin (1934) and Laubenfels (1953); a review of their collections was included in the study of fossil Australian sponges by Pickett (1983). More recently a representative of the extant Brachiaster Wilson, 1925, was described from these rocks by Pisera and Bitner (2007). Gammon et al. (2000a, b, 2003) and Gammon and James (2003) provided a review of the sponge fauna in a geological context and offered an environmental interpretation. Loose spicules were first reported by Hinde (1910), and more recently were studied in detail by Łukowiak (2015, 2016), while intact nonlithistid demosponges were described by Łukowiak and Pisera (2017). Despite these studies, the fossil content of the Pallinup Formation, in particular lithistid demosponges that comprise the majority of the fossil fauna, remains mostly undescribed.
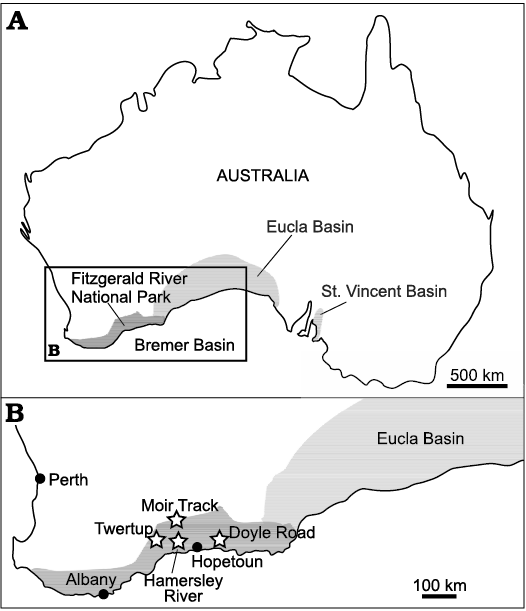
Fig. 1. Palaeogeography of the Eocene in SW Australia (A) and location of investigated sections (B, stars). Eocene sediment shaded. Based on Gammon et al. (2000b).
The aim of the present paper is the revision of the most common (numerically) lithistid sponges that belong to the family Phymaraphiniidae. The species described had been reported from the Eocene of SW Australia by Chapman and Crespin (1934) and Laubenfels (1953), but they were erroneously ascribed to the wrong genera and families. Gammon et al. (2000a) mentioned the presence of phymaraphiniid lithistids in these rocks but did not provide any additional details.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: http://zoobank.org/urn:lsid:zoobank.org:pub:20476566-0D3C-4AFE-B2D6-8B3566687FEE.
Institutional abbreviations.—GSWA, Geological Survey of Western Australia, Perth; MNHN, Musée national d’Histoire naturelle, Paris, France; WA, University of Western Australia, Perth; WAM, Western Australian Museum, Perth, Australia; ZPAL, Institute of Paleobiology, Polish Academy of Sciences, Warszawa, Poland.
Other abbreviations.—FRNP, Fitzgerald River National Park; st., SEM stub.
Geological setting
Eocene sponge rich facies from SW Australia, that are today attributed to the upper Eocene Pallinup Formation in Bremer Basin (Fig. 1), have been known for over a century (Hinde 1910; Chapman and Crespin 1934; Laubenfels 1953) and recently their stratigraphical, sedimentological, palaeoenvironmental and palaeobiogeographical aspects were described in detail by Gammon et al. ( 2000a, b, 2003) and Gammon and James (2001, 2003). All sponges described here were collected from the Fitzgerald Member, the topmost member of the Pallinup Formation represented by heterogeneous units of spiculite, spongiolite, terrigenous sandstone and mudstone. The Fitzgerald Member consists of spiculites and spongiolites (Gammon et al. 2000b) and is interpreted as being deposited in a shallow water inner shelf shore-face embayment-archipelago (Gammon et al. 2000a; Gammon and James 2001, 2003).
Material and methods
The first specimens of the sponges described here were collected by Paul Gammon during his PhD thesis fieldwork. The majority of the specimens that form the basis of the present study were collected during two field expeditions to the Fitzgerald River National Park (FRNP) area organised by AP and financed by the Polish granting agencies (grant numbers KBN 3 PO4D 03924 and NCN 2016/21/B/ST10/02332). The first expedition took place in 2004 (the field team included AP, Paul Gammon, and John Pickett), and the second one in 2017 (the field team included AP, MAB, JF, and David Sutton, Perth; Geraldine and Steve Janicke, Albany, Australia (both Friends of FRNP) helped us during part of the fieldwork).
The investigated sponges were first studied with a binocular microscope to recognise desma types from the choanosomal skeleton and to search for preserved ectosomal spicules. Small fragments of the upper surface, with both desmas and ectosomal spicules, were studied under SEM in the Institute of Paleobiology, Warszawa. The type material described by Chapman and Crespin (1934) and Laubenfels (1953) which was studied in 2017, is housed in the collections of the University of Western Australia and the Geological Survey of Western Australia, Perth. The new collected material described here is housed in the Western Australian Museum (WAM 2023.2, WAM 2023.3) and the Institute of Paleobiology (ZPAL Pf. 14).
Systematic palaeontology
Phylum Porifera Grant, 1836
Class Demospongiae Sollas, 1885
Subclass Heteroscleromorpha Cárdenas, Pérez, and Boury-Esnault, 2012
Order Tetractinellida Marshall, 1876
Family Phymaraphiniidae Schrammen, 1924
Genus Twertupia nov.
Type species: Thamnospongia subglabra Chapman and Crespin, 1934, by monotypy. Hamersley River area, SW Australia, upper Eocene.
Etymology: From Twertup field station, where many specimens of this species were found.
Diagnosis.—As for the type species, by monotypy.
Twertupia subglabra (Chapman and Crespin, 1934) comb. nov.
Figs. 2A–D, 3, 4.
1934 Thamnospongia subglabra sp. nov.; Chapman and Crespin 1934: 116, pl. 9: 17–18, pl. 10: 19.
non 1934 Thamnospongia neoclavatella sp. nov.; Chapman and Crespin 1934: 115, pl. 9: 16.
1953 Stachyspongia neoclavatella (Chapman and Crespin, 1934); Laubenfels 1953: 108, text-fig. 2.
?1983 “Thamnospongia” neoclavatella Chapman and Crespin, 1934; Pickett 1983: 101.
1983 “Thamnospongia” subglabra Chapman and Crespin, 1934; Pickett 1983: 101.
Diagnosis.—Subcylindrical to cylindrical ramose phymaraphiniid covered with irregularly distributed conical mamelons (they can be nearly smooth when young/smaller) that are covered with sinuous, radially arranged shallow canals on their surface. Ectosomal spicules are irregular, strongly tuberculate discotriaenes to phyllotriaenes.
Material.—Thamnospongia subglabra holotype (GSWA H1) and paratypes (GSWA H16, H19) from Hamersley River area, Australia, Eocene, and the material of Stachyspongia neoclavatella (WA 23732.2 specimens illustrated in Laubenfels 1953, here re-illustrated Fig. 2A–D), exact locality unknown, SW Australia, Eocene. Over 500 newly collected specimens (WAM 2023.2a, b, ZPAL Pf. 14/1–9, and more than 400 specimens unnumbered) from Fitzgerald River National Park (Hamersley River area, Twertup area, Moir Track), and Doyle Road, east of Hopetoun, SW Australia, Eocene (Fig. 1B).
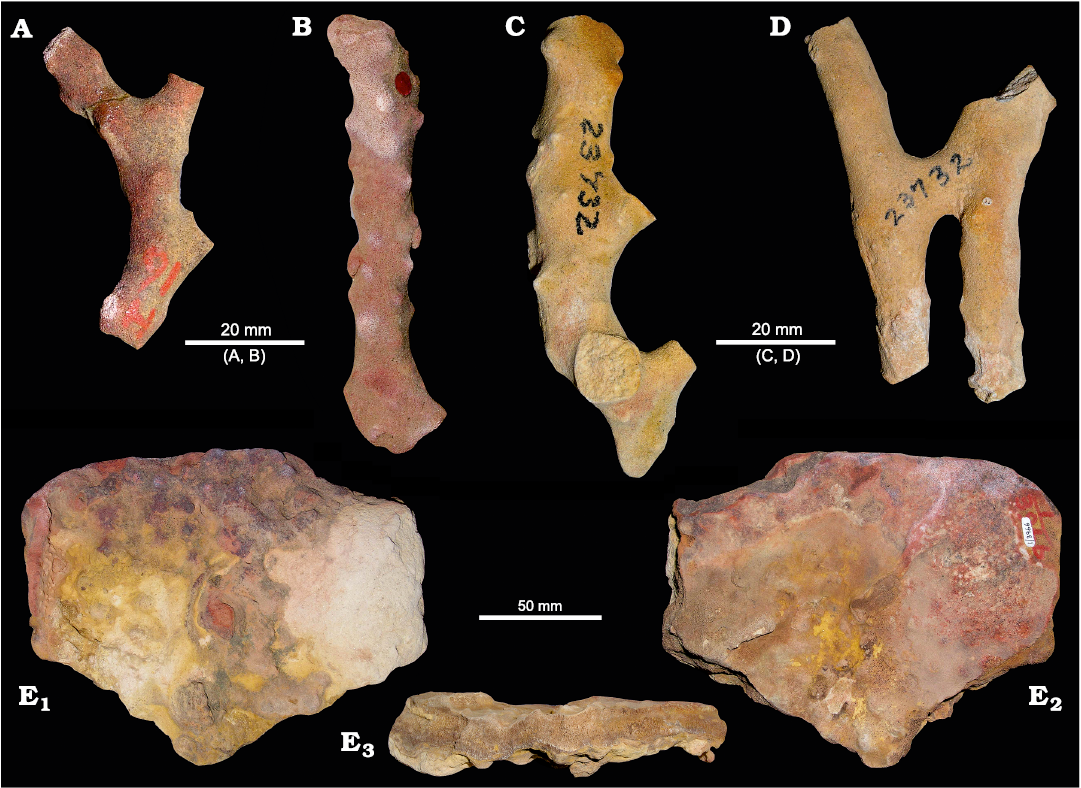
Fig. 2. Specimens of phymaraphiniid sponges illustrated by Chapman and Crespin (1934) and Laubenfels (1953). A, B. Type material of Twertupia subglabra (Chapman and Crespin, 1934) (originally Thamnospongia subglabra; Chapman and Crespin 1934: pl. 9: 17, 18), Hamersley River area, Australia, upper Eocene. A. Holotype, GSWA H1. B. Paratype GSWA H16. C, D. Twertupia subglabra (originally Stachyspongia neoclavatella; Laubenfels 1953: text-fig. 2A right and left), SW Australia, exact locality unknown, upper Eocene, WA23732 (two different specimens with the same collection number). E. Type material of Pickettispongia tabelliformis (Chapman and Crespin, 1934) (originally Discodermia tabelliformis; Chapman and Crespin 1934: pl. 7: 7), Hamersley River area, Australia, upper Eocene. GSWA 1/3966, E1, E2, opposite surface views, E3, section view.
Description.—Subcylindrical to cylindrical sponges (Fig. 3A–J), rarely slightly compressed (Fig. 3K), usually ramose, reaching 2 cm in diameter and up to 15 cm high. When smaller (young), they are nearly smooth, later bearing wide conical outgrowths (mamelons) that are distributed without any order on the surface. These outgrowths are up to 5 mm high and covered with sinuous, shallow, radially arranged furrows (not developed in small/young specimens) that spread over the surface of the sponge. Surface of choanosomal desma skeleton finely porous (Fig. 4B, C) showing strongly sculptured triders, with pore diameters between 0.05 and 0.2 mm, pores dense but irregularly distributed. In some parts of the surface, the skeleton may be dense, without visible openings, due to the development of a secondary skeleton composed of flattened modified desmas resembling rhizoclones (Fig. 4D) that cover canal openings.
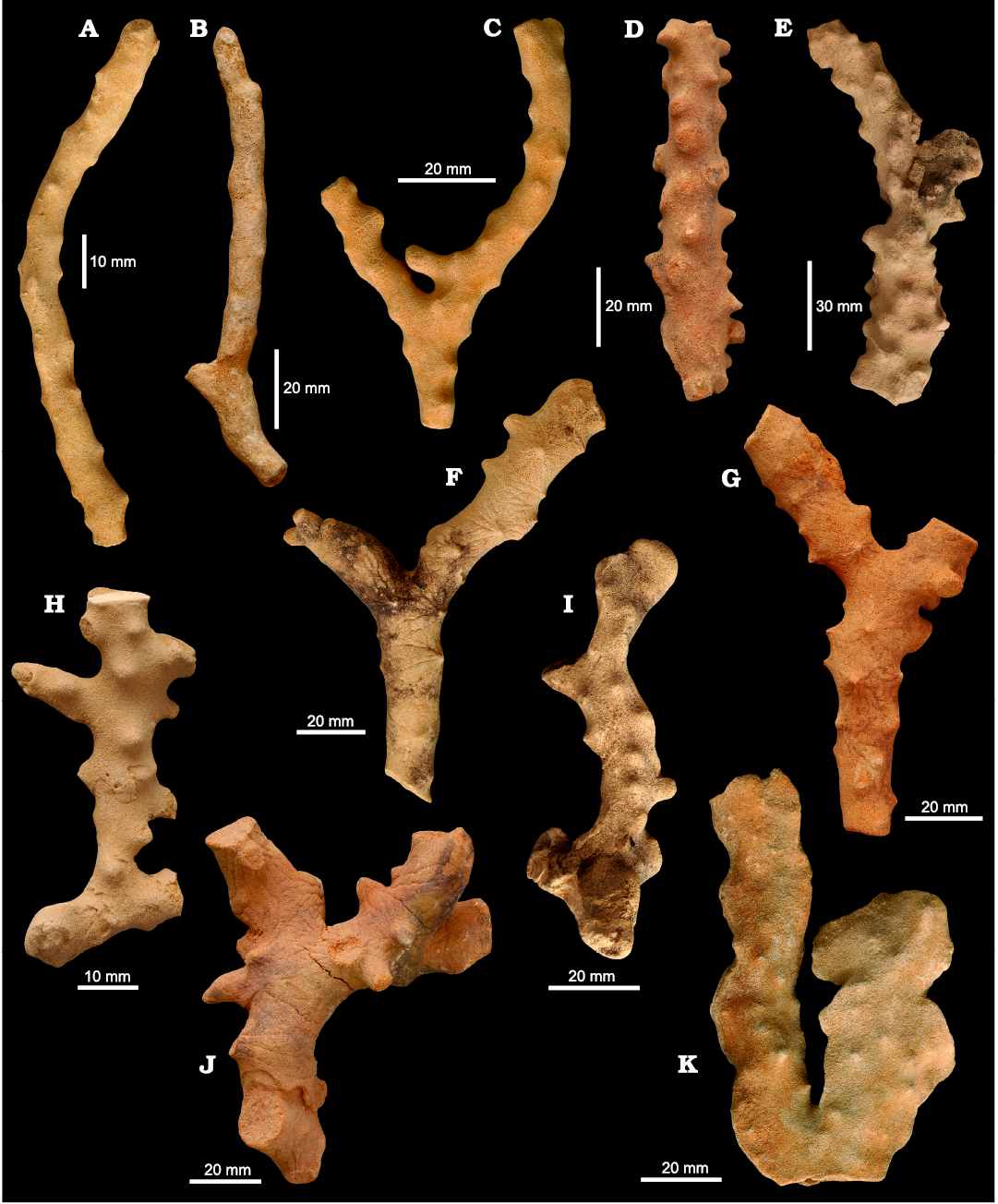
Fig. 3. Morphological variability of the phymaraphiniid sponge Twertupia subglabra (Chapmann and Crespin, 1934) from the Fitzgerald River National Park (A, Doyle Road; B–D, F, G, J, K, Twertup area; E, H, I, Hamersley River area), Australia, upper Eocene. A. ZPAL Pf. 14/1. B. ZPAL Pf. 14/2. C. WAM 2023.2a. D. ZPAL Pf. 14/3. E. ZPAL Pf. 14/4. F. ZPAL Pf. 14/5. G. ZPAL Pf. 14/6. H. ZPAL Pf. 14/7. I. ZPAL Pf. 14/8. J. WAM 2023.2b. K. ZPAL Pf. 14/9.
The choanosomal desma skeleton consists of regular triders (Fig. 4C) covered by complex large tubercles (button-like) in the central part (slightly mushroom-shaped), that are covered by smaller tubercles, and half-ring-like elevations (also granulated) on arms in the proximal part; in the distal part much smaller and smooth tubercles occur (Fig. 4F). Central large tubercle may be dome-like and granulated or with a flattened top and smooth tubercle or low conical elevation in the centre (Fig. 4G–I). When young, the principal tubercles may be smooth (Fig. 4E). Trider arms usually divided (dichotomously or trichotomously) in distal part, forming strongly branched zygomes (Fig. 4F). In some parts the skeleton may be very dense (no pores are visible) due to additional spicules resembling rhizoclones (Fig. 4D) (most probably modified, flattened triders). Ectosomal spicules are irregular and strongly incised discotriaenes to phyllotriaenes, with tuberculate surface, that are 296–378 μm in diameter, between them may occur rhizoclone-like modified desmas (Fig. 4A). No microscleres have been found.

Fig. 4. Skeletal features of the phymaraphiniid sponge Twertupia subglabra (Chapmann and Crespin, 1934), from the Fitzgerald River National Park (A, B, F, H, Doyle Road; C, D, E, G, I, Hamersley River area), Australia, upper Eocene. A. Upper surface showing ectosomal disco/phyllotriaenes; between them rhizoclone-like modified desmas are visible, ZPAL Pf. 14/st.A822. B. General view of the choanosomal skeleton with canal openings, ZPAL Pf. 14/st.A819. C. Detailed view of choanosomal skeleton surface showing trider desmas and their sculpture in oblique view, ZPAL Pf. 14/st.906. D. Surface of choanosomal skeleton showing development of the secondary skeleton composed of flattened modified triders covering canal openings, ZPAL Pf. 14/st.906. E. Young trider desma partly incorporated into the skeleton, ZPAL Pf. 14/st.A743. F. Surface of choanosomal skeleton showing variable sculpturing of the triders, ZPAL Pf. 14/st.A819. G. Details of trider sculpture and articulations, ZPAL Pf. 14/st.906. H, I. Trider sculpture variability. H. ZPAL Pf. 14/st.A748. I. ZPAL Pf. 14/st.A821.
Remarks.—This is the most common sponge in the Eocene rocks of the FRNP area.
Chapman and Crespin (1934) attributed these sponges to the Cretaceous Thamnospongia Hinde, 1883, based on their general shape which is comparable to the Cretaceous species T. glabra Hinde, 1883. However, Thamnospongia has dichotriaenes as ectosomal spicules, not disco- or phyllotriaenes, and tetraclones as choanosomal desmas, not triders, and thus belongs probably to the Phymatellidae Schrammen, 1910. Laubenfels (1953) who realized the error, took a different, although erroneous, approach and compared the sponges to the Cretaceous Stachyspongia Zittel, 1878, characterised by rhizoclones as choanosomal desmas. Stachyspongia also has a deep cylindrical spongocoel, entirely lacking in the studied specimens. The only comparable taxon is Pholidocladia Hinde, 1883, from the Late Cretaceous of Europe, composed of bushy cylindrical branches with triders as desmas (very similar to those in the studied specimens) and smooth irregular discotriaenes as ectosomal spicules. Considering the age difference, large geographical distance, and difference in ectosomal spicules, we consider the studied species belonging to a new genus.
Additionally, Laubenfels (1953) synonymized the two species of Chapman and Crespin (1934), i.e., Thamnospongia neoclavatella and Thamnospongia subglabra, erroneously basing it solely on external shape, and assigned them to the Stachyspongia, as Stachyspongia neoclavatella (Chapman and Crespin, 1934). Thamnospongia neoclavatella has tetraclones and phyllotriaenes as ectosomal spicules and thus belongs probably to the Theonellidae Lendenfeld, 1903, while Stachyspongia is a rhizomorine demosponge (desmas as rhizoclones).
Stratigraphic and geographic range.—Upper Eocene, Fitzgerald Member, Pallinup Formation; Fitzgerald River National Park (Hamersley River area, Twertup area, Moir Track) and Doyle Road, SW Australia.
Genus Pickettispongia nov.
Type species: Discodermia tabelliformis Chapman and Crespin, 1934, by monotypy. Hamersley River area, SW Australia, upper Eocene.
Etymology: In honour of John Pickett, Australian palaeontologist and student of fossil sponges.
Diagnosis.—As for the type species, by monotypy.
Pickettispongia tabelliformis (Chapman and Crespin, 1934) comb. nov.
Figs. 2E, 5, 6.
1934 Discodermia tabelliformis sp. nov.; Chapman and Crespin 1934: 113, pl. 7: 7–8.
1983 “Discodermia” tabelliformis; Pickett 1983: 100.
Diagnosis.—Large flattened phymaraphiniid composed of two closely adhering plates; both sides identical, covered with numerous wide and low conical outgrowths/mamelons with radially arranged sinuous bands of denser skeleton. Desmas are triders with strong sculpture, ectosomal spicules are irregularly tuberculated disco- to phyllotriaenes.
Material.—Holotype (GSWA 1/3966, Fig. 2E) from Hamersley River area, SW Australia, Eocene. Newly collected material (WAM 2023.3a, b, ZPAL Pf. 14/10–11, and 19 specimens unnumbered): 12 specimens from the Hamersley River area, six specimens from the Twertup area, five specimens from Doyle Road (Fig. 1B); in each case from large to small fragments.
Description.—Plate-likes sponges that are up to 18×15 cm large (they are not complete) and up to 1 cm thick (Fig. 5A–C). Both sides of the plates have the same morphology; they are covered with low, widely conical outgrowths (mamelons) that vary strongly in size and distribution between various specimens. The mamelons display sinuous, radially arranged bands of denser skeleton that are up to 1 mm wide (Fig. 5B, C1).
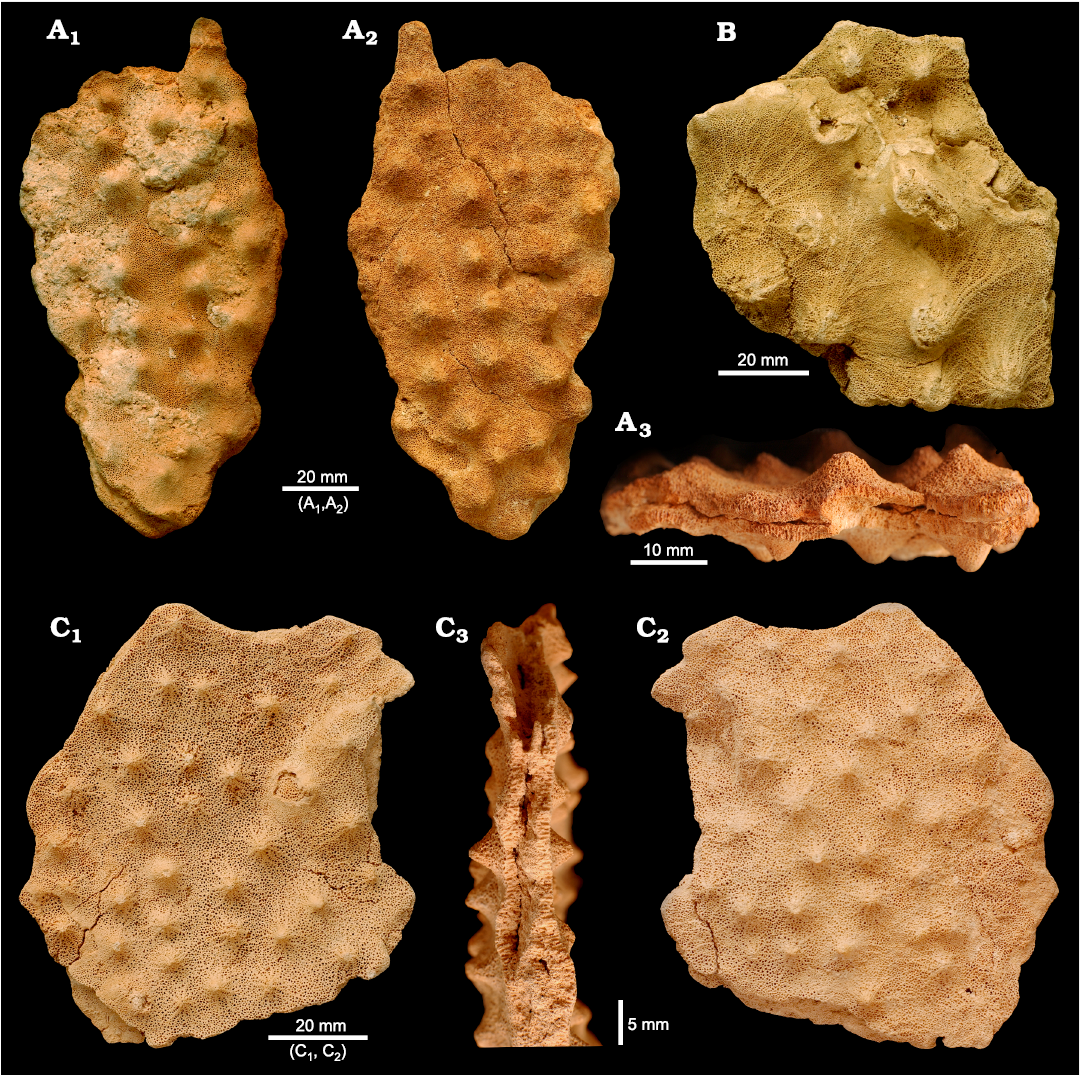
Fig. 5. Morphological variability of the phymaraphiniid sponge Pickettispongia tabelliformis (Chapmann and Crespin, 1934) from the Fitzgerald River National Park (B, Doyle Road; A, C Hamersley River area), Australia, upper Eocene. A. Opposite side views (A1, A2) and transverse section (A3) showing narrow slit in the middle, WAM 2023.3/a. B. ZPAL PF. 14/10. C. Opposite side views (C1, C2) and transverse section (C3) showing narrow slit in the middle, ZPAL Pf. 14/11.
The surface of the choanosomal skeleton is covered with openings that are round to elongate, 0.1–0.4 (exceptionally 0.6) mm wide, and densely distributed (Fig. 6B, E). They are separated only by a narrow skeletal bridge in some parts or more wide skeletal bands in others. Desmas are typical triders (Fig. 6C, D, F–I) with a central tubercle (slightly mushroom-like shaped) that is hemispherical and tuberculate (Fig. 6C, D, G) or nearly flat with a large central tubercle (Fig. 6F–I); both types are observed in the same specimen or even side by side. Trider arms bear proximally half-ring-like bands (one per arm) that are covered with tiny tubercles, and smaller smooth tubercles distally (Fig. 6C, D, H); the arms are usually divided (dichotomously or trichotomously) in distal part, and forming strongly branched zygomes (Fig. 6C, G). Ectosomal spicules are spinose discotriaenes with irregular margins measuring 100–164 μm in diameter; between them rhizoclone-like modified desmas may occur (Fig. 6A). No microscleres have been found.
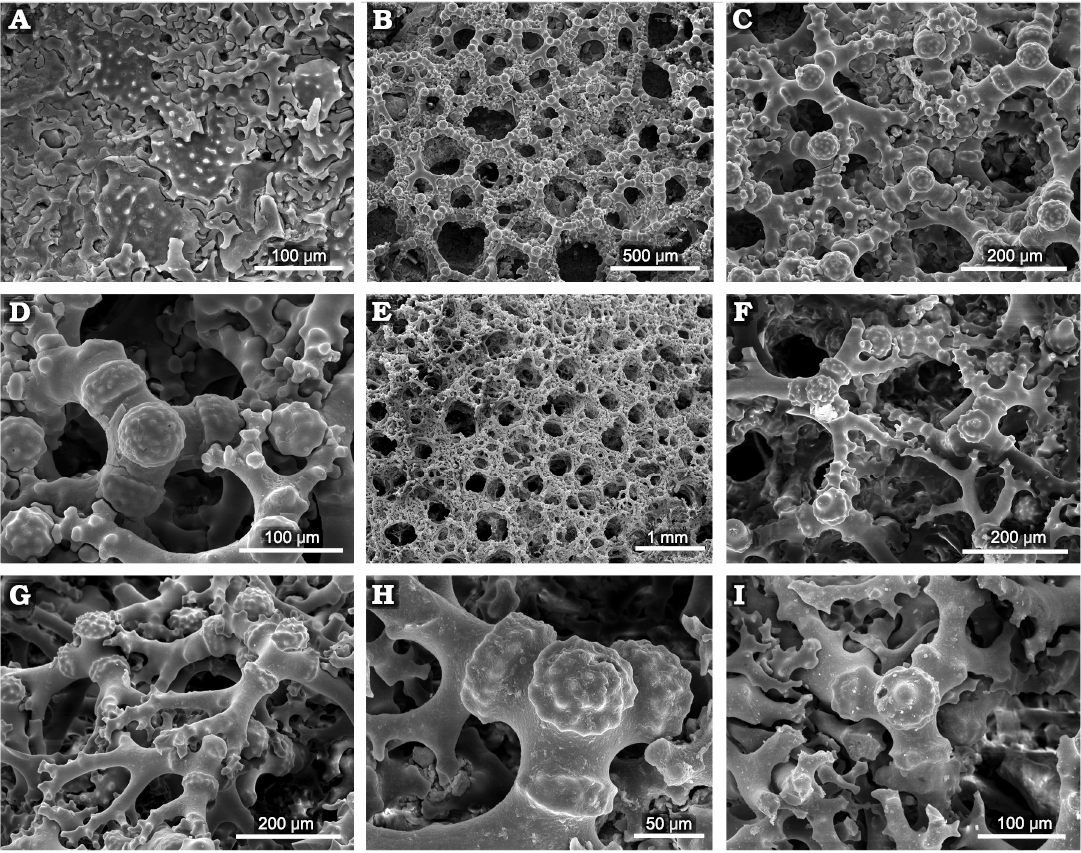
Fig. 6. Skeletal features of morphological variability of the phymaraphiniid sponge Pickettispongia tabelliformis (Chapmann and Crespin, 1934), from the Fitzgerald River National Park (A–D, Doyle Road; E–I, Hamersley River area), Australia, upper Eocene. A. Upper surface showing ectosomal discotriaenes; between them rhizoclone-like modified desmas are visible, ZPAL Pf. 14/st.A839. B. General view of the choanosomal skeleton with canal openings, ZPAL Pf. 14/st.A823. C. Detailed view of choanosomal skeleton surface showing trider desmas and their articulations, ZPAL Pf. 14/st.A823. D. Surface of choanosomal skeleton showing trider with hemispherical central tubercle, ZPAL Pf. 14/st.A823. E. Slightly eroded surface of the choanosomal skeleton showing densely distributed canal openings, ZPAL Pf. 14/st.914. F. Details of triders showing variability in sculpture of desmas, ZPAL Pf. 14/st.825. G. Oblique view of triders showing articulations, ZPAL Pf. 14/st.914. H, I. Details of trider sculpture. H. ZPAL Pf. 14/st.914. I. ZPAL Pf. 14/st.913.
Remarks.—These sponges were originally described as Discodermia tabelliformis (family Theonellidae) by Chapman and Crespin (1934). However, neither desmas nor ectosomal discotriaenes from the type material were illustrated nor clearly described. The newly collected specimens have triders as desmas that are typical of Phymaraphiniidae, thus the species is transferred to this family. The habitus of this sponge is unique, not only among Phymaraphiniidae but generally among lithistids, hence the designation of a new genus.
Initially this species appears to be an encrusting form. However, both sides of the specimens are the same (Fig. 5A1, A2, C1, C2), clearly indicating that it was an erect, flat sponge. This is also supported by the observation that in the middle of the flat sponge body is a narrow slit about 1 mm wide (Fig. 5A3, C3), and the appearance of the skeletal structures indicate that the growth of the sponge in thickness was in two opposite directions.
Stratigraphic and geographic range.—Upper Eocene, Fitzgerald Member, Pallinup Formation, Fitzgerald River National Park (Hamersley River area, Twertup area), and Doyle Road, east of Hopetoun, SW Australia (Fig. 1B).
Discussion
The complex sculpture of the trider desmas in the studied Eocene species resembles Cretaceous species from Europe (Schrammen 1910; Świerczewska-Gładysz and Jurkowska 2022: figs. 5A, 7A, B, 8A), and to a smaller degree extant Kaliapsis Bowerbank, 1869, from Madagascar (Figs. 7, 8), but not most of the Atlantic-Mediterranean phymaraphiniid species which are characterised by desmas covered by simple smooth tubercles (see Carvalho and Pisera 2019; Carvalho et al. 2020). In addition, the Caribbean species Exsuperantia clava (Schmidt, 1879) has mushroom-like tubercles covered with smaller tubercles. All these species are characterised by smooth phyllotriaenes. The ornamentation of desmas of the Cretaceous species from Poland, as illustrated by Świerczewska-Gładysz and Jurkowska (2022), shows large variability in the same specimen, and the same sculpture variability is noted in the Eocene material from Australia.
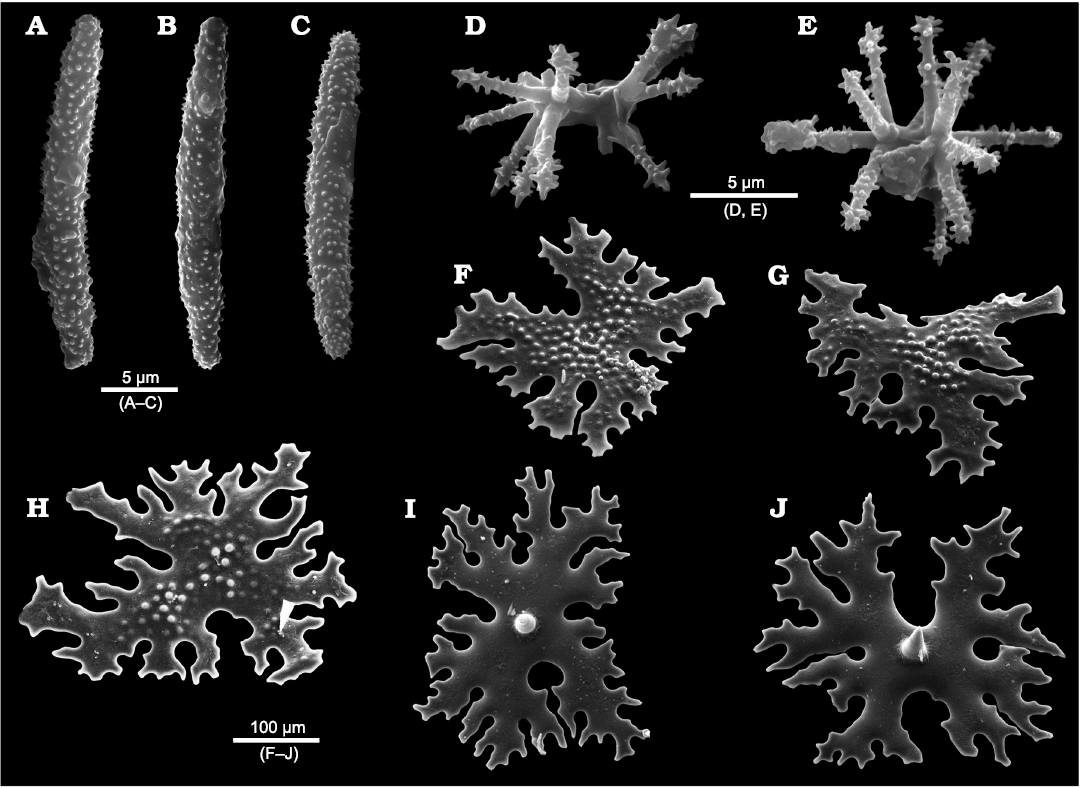
Fig. 7. Loose spicules of the extant phymaraphiniid sponge Kaliapsis incrustans (Vacelet and Vasseur, 1971) from Madagascar, MNHN E531, Tu63. A–C. Microrhabds. D, E. Amphiasters. F–J. Ectosomal phyllotriaenes in top (F–H, note tuberculation) and lower surface views (I, J, note short rhabd).
Today phymaraphiniids are a relict group of lithistid demosponges. Recent representatives (Fig. 9) are known from the Atlantic (from Azores and Caribbean) and Indo-Pacific (Pisera and Lévi 2002; Carvalho and Pisera 2019; Carvalho et al. 2020). Recently a new occurrence (new species and probably new genus) was reported from the submarine caves in the Mediterranean (Pisera et al. 2022).
There are three genera of extant Phymaraphiniidae, i.e., Exsuperantia Özdikmen, 2009 (=Rimella Schmidt, 1879), Kaliapsis Bowerbank, 1869, and Lepidothenea Laubenfels, 1936. The position of Lepidothenea (monospecific) among phymaraphiniids is doubtful in our opinion (see also comments by Carvalho et al. 2020), due to the fact that its desmas are unknown, while the ectosomal spicules and microscleres fit with the Theonellidae.
The genus Exsuperantia contains three species, all known from the Atlantic (Cuba, Azores, and a seamount south of Azores ), and their taxonomic details were recently elaborated by Carvalho and Pisera (2019) and Carvalho et al. (2020). They are small clavate to columnar, globular to ficiform shaped sponges which have ectosomal spicules that are smooth phyllotriaenes to discotriaenes. In this respect they resemble Cretaceous representatives of the family that are much larger sponges and are characterised by various morphological types (club, ear-shaped, etc.).
The genus Kaliapsis with three species is reported from the Indian Ocean and probably the Pacific (Vacelet et al. 1976). They are small thinly encrusting sponges possessing strongly tuberculate phyllotriaenes to discotriaenes. The species are K. permolis Topsent, 1890, from the Island of Reunion, K. incrustans (Vacelet and Vasseur, 1971) from Madagascar, and K. cidaris Bowerbank, 1869, from the South Seas (probably Pacific). All of them are very poorly known and illustrated. It was found that the first species, K. permolis is a theonellid (Siliquarispongia Hoshino, 1981) not a phymaraphiniid sponge, due to the character of its desmas (Pisera and Lévi 2002). This was also found for K. aspera (Carter, 1880) (see Pisera and Lévi 2002). K. cidaris is known only from the holotype material (in slides), while spicules of K. incrustans were illustrated in drawings without figures of the choanosomal desmas that are the defining character. Its spicular component is illustrated here for the first time using SEM (Figs. 7, 8). These two Indo-Pacific species are very small, thin encrusting sponges characterised by strongly tuberculate disco- to phyllotriaenes, similar to the Eocene phymaraphiniids reported here. There are two other thinly encrusting lithistid species, reported by Carter (1880: 143, 144, pl. 7: 45, 46) from Ceylon, attributed to the genus Corallistes Schmidt, 1870, having trider-like tuberculated desmas resembling triders of phymaraphiniid sponges. The similarity of desmas of Corallistes aculeata (Carter 1880: pl. 7: 34) and C. verrucosus (Carter 1880: pl. 7: 46) to those in Kaliapsis was already observed by Sollas (1888). Unfortunately there is no information about ectosomal spicules or microscleres of these species in Carter (1880), thus these sponges may equally belong to Desmanthus Topsent, 1893. This problem needs further study if new material becomes available. In summary, the genus Kaliapsis contains only two species, i.e., K. incrustans and K. cidaris with three possible others to be confirmed.

Fig. 8. Choanosomal desma skeleton of the extant phymaraphiniid sponge Kaliapis incrustans (Vacelet and Vasseur, 1971) from Madagascar, MNHN E531, Tu63, composed of trider desmas.
Bodily preserved Cretaceous phymaraphiniids are represented by seven genera and numerous species that have been reported from the Aptian to Campanian in Europe (Fig. 9; Reid 2004). They are known from Great Britain (Hinde 1883), Germany (Schrammen 1910; Wagner 1963; Schneider et al. 2013), France (Moret 1926), Poland (Hurcewicz 1966), and Spain (Lagenau-Hérenger 1962; Rosales et al. 1995). Recently Świerczewska-Gładysz and Jurkowska (2022) in their monograph on phymaraphiniids and theonellids from the Cretaceous of Poland discussed in detail their stratigraphical and geographical distribution in the Upper Cretaceous rocks of Europe. Intact fossil phymaraphiniid lithistids have never been reported outside Europe, or from post Mesozoic rocks.
Apart from those bodily preserved phymaraphiniids, there is a geological record of loose phymaraphiniid desmas (that are sufficiently characteristic to be certain of their taxonomic position) that covers a much longer time span (Wiedenmayer 1994). The oldest record of phymaraphiniid triders is from the upper Norian (Late Triassic) (Mostler 1986; Wiedenmayer 1994) of the Alps and from the Triassic/Jurassic boundary beds (Mostler 1976; Wiedenmayer 1994) of the same area. The next occurrence of such triders is from the Upper Jurassic of Alps (Mostler 1986; Wiedenmayer 1994) and Schwäbische Alb (Reif 1967; Wiedenmayer 1994). Loose phymaraphiniid spicules were also reported from the Eocene rocks of Western Australia (Hinde 1910) and Ukraine (Łukowiak et al. 2022).
The present-day and fossil distribution of phymaraphiniids (Fig. 9) is geographically wide but highly punctuated and incomplete (especially bodily preserved sponges) and not overlapping. Such a pattern can be explained by a much wider past Tethyan distribution of phymaraphiniid lithistids, and the occurrences today considered to be a relict of this Tethyan fauna. This pattern is also known among other sponges, i.e, Vetulina Schmidt, 1879 (see Pisera et al. 2018; Schuster et al. 2018) and some brachiopods (e.g., Bitner and Cahuzac 2013; Bitner and Motchurova-Dekova 2016), and it may be expected to occur among other groups. These observations support Reid’s (1967) opinion that the Tethys was a factor in shaping the distribution of modern sponge fauna in warm regions.
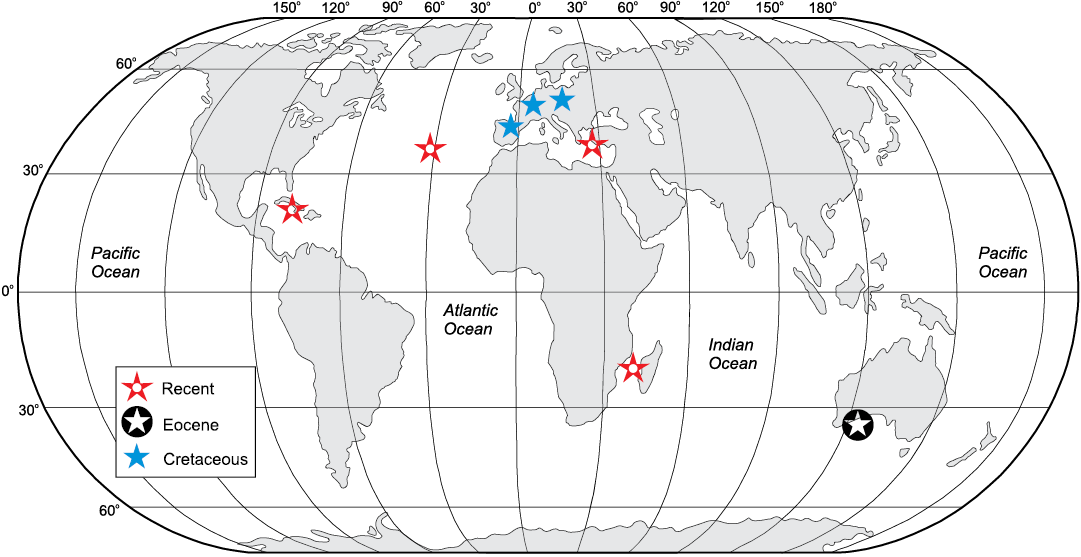
Fig. 9. Geographic distribution of extant and fossil representatives of the demosponge family Phymaraphiniidae.
Conclusions
We described two new genera of the phymaraphiniid lithistid sponges from the Eocene of SW Australia that were earlier erroneously attributed to other genera and different families. This occurrence is filling the stratigraphical gap between well-known Cretaceous phymaraphiniid sponges from Europe and Recent phymaraphiniids, as well as geographical gap (Cretaceous of Europe and Eocene of Australia). We have discussed the scope of extant phymaraphiniids and their present day and fossil occurrences concluding that the observed pattern results, most probably, from much wider Mesozoic Tethyan distribution.
Acknowledgements
Paul Gammon (Geological Survey of Canada, Ontario, Canada) and John Pickett (Geological Survey of New South Wales, Londonderry, Australia) are thanked for introducing AP to the studied outcrops in 2004; David Sutton (Perth, Australia) for his help with organisation of the fieldwork in 2017; Friends of FRNP are thanked for allowing us to use the Twertup station during both stays in Twertup in 2004 and 2017, and Geraldine and Steve Janicke (Albany, Friends of FRNP) for their help and company during part of the 2017 fieldwork. Jean Vacelet (Station Marine d’Endoume, Marseille, France) and Ewa Świeczewska-Gładysz (University of Łódź, Poland) are thanked for their careful reviews. We are indebted to the Department of Biodiversity and Attractions, Western Australia for collecting permits, FRNP park rangers for support and assistance and the traditional owners for allowing us to work in the area. Western Australian Museum, Perth, is thanked for logistic support and help with some equipment. The study was supported by KBN grant no. 3 PO4D 03924 and NCN grant no. 2016/21/B/ST10/02332 to AP.
References
Bitner, M.A. and Cahuzac, B. 2013. New record of Discradisca (Brachiopoda: Discinidae) from the Early Miocene of the Aquitaine Basin, south-western France. Comptes Rendus Palevol 12: 23–29. Crossref
Bitner M.A. and Motchurova-Dekova, N. 2016. Middle Miocene (Badenian) brachiopods from Yasen, northwestern Bulgaria: taxonomic composition and biogeographical significance. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 279: 7–22. Crossref
Bowerbank, J.S. 1869. A Monograph of the siliceo-fibrous sponges. Part I. Proceedings of the Zoological Society of London 1869 (1): 66–100. Crossref
Carter, H.J. 1880. Report on specimens dredged up from the Gulf of Manaar and presented to the Liverpool Free Museum by Capt. W. H. Cawne Warren. Annals and Magazine of Natural History, Series 5 6: 35–61. Crossref
Cárdenas, P., Pérez, T., and Boury-Esnault, N. 2012. Sponge systematics facing new challenges. In: M.A. Becerro, M.J. Uriz, M. Maldonado, and X. Turon (eds.), Advances in Sponge Science: Phylogeny, Systematics, Ecology. Advances in Marine Biology 61: 79–209. Crossref
Carvalho, F.C. and Pisera, A. 2019. Revision of the genus Exsuperantia Özdikmen, 2009 (Tetractinellida: Phymaraphiniidae) with description of a new species from the Atlantic Ocean. Zootaxa 4613: 135–151. Crossref
Carvalho, F.C., Cárdenas, P., Ríos, P., Cristobo, J., Rapp, H.T., and Xavier, J.R. 2020. Rock sponges (lithistid Demospongiae) of the Northeast Atlantic seamounts, with description of ten new species. PeerJ 8: e8703. Crossref
Chapman, F. and Crespin, I. 1934. The palaeontology of the Plantagenet Beds of Western Australia. Journal and Proceedings of the Royal Society of Western Australia 20: 103–136.
Gammon, P.R. and James, N.P. 2001. Palaeogeographical influence on late Eocene biosiliceous sponge-rich sedimentation, southern Western Australia. Sedimentology 48: 559–584. Crossref
Gammon, P.R. and James, N.P. 2003. Paleoenvironmental controls on Upper Eocene biosiliceous neritic sediments, Southern Australia. Journal of Sedimentary Research 73: 957–972. Crossref
Gammon, P.R., Bone, Y., Clarke, J.D.A., and James, N.P. 2003. Upper Eocene greenhouse deposits in southern Australia: paleoclimatology and paleoceanography. In: D.R. Prothero, L.C. Ivany, and E.A. Nesbitt (eds.), From Greenhouse to Icehouse: the Marine Eocene–Oligocene Transition, 107–118. Columbia University Press, New York.
Gammon, P.R., James, N.P., and Pisera, A. 2000a. Eocene spiculites and spongolites in south-western Australia: Not deep, not polar, but shallow and warm. Geology 28: 855–858. Crossref
Gammon, P.R., James, N.P., Bone, Y., and Clarke, J.D.A. 2000b. Sedimentology and lithostratigraphy of Upper Eocene sponge rich sediments, southern Western Australia. Australian Journal of Earth Sciences 47: 1087–1103. Crossref
Grant, R.E. 1836. Animal kingdom. In: R.B. Todd (ed.), The Cyclopaedia of Anatomy and Physiology. Volume 1, 107–118. Sherwood, Gilbert, and Piper, London.
Hinde, G.J. 1883. Catalogue of the Fossil Sponges in the Geological Department of the British Museum. 248 pp. British Museum (Natural History), London.
Hinde, G.J. 1910. On the fossil sponge spicules in a rock from deep Lead at Princess Royal Township, Norseman District, Western Australia. Bulletin of Geological Survey of Western Australia 36: 7–24.
Hoshino, T. 1981. Shallow-water demosponges of Western Japan, 1. Journal of Science of the Hiroshima University (B) 29 (1): 47–205.
Hurcewicz, H. 1966. Siliceous sponges from the Upper Cretaceous of Poland, Part I, Tetraxonia. Acta Palaeontologica Polonica 11: 15–129.
Lagenau-Héreneger, L. 1962. Contribution à l’étude des spongiaires siliceux du Crétacé inférieur. Mémoires de la Société Géologique de France, Nouvelle Série 41: 1–252.
Laubenfels, M.W. de 1936. A discussion of the sponge fauna of the dry Tortugas in particular and the West Indies in General, with material for a revision of the families and orders of the Porifera. Carnegie Institute of Washington Publication 467 (Tortugas Laboratory Paper 30): 1–225.
Laubenfels, M.W. de 1953. Fossil sponges of Western Australia. Journal and Proceedings of Royal Society of Western Australia 37: 105–117.
Lendenfeld, R. von. 1903. Porifera. Tetraxonia. In: F.E. Schulze (ed.), Das Tierreich 19, vi–xv + 1–168. Friedländer, Berlin.
Łukowiak, M. 2015. Late Eocene siliceous sponge fauna of southern Australia: reconstruction based on loose spicules record. Zootaxa 3917: 1–65. Crossref
Łukowiak, M. 2016. Fossil and modern sponge fauna of southern Australia and adjacent regions compared: interpretation, evolutionary and biogeographic significance of the late Eocene “soft” sponges. Contributions to Zoology 85 (1): 13–35. Crossref
Łukowiak, M. and Pisera, A. 2017. Bodily preserved Eocene non-lithistid demosponge fauna from southern Australia: taxonomy and affinities Journal of Systematic Palaeontology 15: 473–497. Crossref
Łukowiak, M., Pisera, A., Stefanska, T., and Stefanskyi, V. 2022. High diversity of siliceous sponges in western Tethyan areas during the Eocene: palaeobiogeographical, ecological and taxonomic significance. Papers in Palaeontology 8 (1): e1416. Crossref
Marshall, W. 1876. Ideen über die Verwandtschaftsverhältnisse der Hexactinelliden. Zeitschrift für wissenschaftliche Zoologie 27 (1): 113–136.
Moret, L. 1926. Contribution l’étude des Spongiaires siliceux du Crétacé supérieur français. Mémoires de la Société Géologique de France, Nouvelle Série 5: 1–314.
Mostler, H. 1976. Poriferenspiculae der alpinen Trias. Geologisch-Paläontologische Mitteilungen Innsbruck 6: 1–42.
Mostler, H. 1986. Ein Beitrag zur Entwicklung phyllotriaener Megaskleren (Demospongea) aus oberjurassischen Beckensedimenten (Oberalmer Schichten, Nördliche Kalkalpen). Geologische-Paläontologische Mitteilungen Innsbruck 13: 297–329.
Özdikmen, H. 2009. Substitute names for eight sponge genus group names (Porifera). Munis Entomology & Zoology 4: 212–218.
Pickett, J. 1983. An annotated bibliography and review of Australian fossil sponges. Memoirs of the Australasian Palaeontologists 1: 93–120.
Pisera, A. and Bitner, M.A. 2007. The sponge genus Brachiaster (Pachastrellidae, Demospongiae) and its first known fossil representative from late Eocene of southwestern Australia. Alcheringa 31: 365–373. Crossref
Pisera, A. and Lévi, C. 2002. Family Phymaraphiniidae Schrammen, 1924. In: J.N.A. Hooper and R.W.M. Van Soest (eds.), Systema Porifera: Guide to the Classification of Sponges. Kluwer Academic/Plenum Publishers, New York. Crossref
Pisera, A., Gerovasileiou, V., and Digenis, M. 2022. Lithistid sponges from submarine caves of Crete Island: hidden diversity and false endemicity. In: C. Bouafif and A. Ouerghi (eds.), Proceedings of the 3rd Mediterranean Symposium on the Conservation of the Dark Habitats (Genoa, Italy, 21–22 September 2022): 71–74.
Pisera, A., Łukowiak, M., Fromont, J., and Schuster, A. 2018. First record of the genus Vetulina Schmidt, 1879 (Porifera: Demospongiae: Sphaerocladina) from the Indian Ocean with the description of two new species: biogeographic and evolutionary significance. Marine Biodiversity 48: 1529–1539. Crossref
Reid, R.E.H. 1967. Tethys and the zoogeography of some modern and Mesozoic Porifera. Systematics Association Publication 7: 171–181.
Reid, R.E.H. 2004. Mesozoic and Cenozoic lithistid demosponges: Tetracladina. In: R.L. Kaesler (ed.), Treatise on Invertebrate Paleontology. Part E (revised), Porifera, 199–317. The Geological Society of America and The University of Kansas, Boulder and Lawrence.
Reif, W.-E. 1967. Schwammspicula aus dem Weissen Jura Zeta von Natheim (Schwabische Alb). Palaeontographica 127: 85–102.
Rosales, I., Mehl, D., Fernández-Mendiola, P.A., and Garcia-Mondéjar, G. 1995. An unusual poriferan community in Albian of Islares (north Spain): Palaeoenvironmental and tectonic implications. Palaeogeography, Palaeoclimatology, Palaeoecology 119: 47–61. Crossref
Schmidt, O. 1870. Grundzüge einer Spongien-Fauna des atlantischen Gebietes. iii–iv + 88 pp. Wilhelm Engelmann, Leipzig.
Schmidt, O. 1879. Die Spongien des Meerbusen von Mexico (Und des caraibischen Meeres). In: Reports on the dredging under the supervision of Alexander Agassiz, in the Gulf of Mexico, by the USCSS “Blake”, Abtheilung I, Heft I, 1–32. Gustav Fischer, Jena.
Schneider, S., Jäger, M., Kroh, A., Mitterer, A., Niebuhr, B., Vodrážka, R., Wilmsen, M., Wood, C.J., and Zágoršek, K. 2013. Silicified sea life—Macrofauna and palaeoecology of the Neuburg Kieselerde Member (Cenomanian to Lower Turonian Wellheim Formation, Bavaria, southern Germany). Acta Geologica Polonica 63: 555–610. Crossref
Schrammen, A. 1910. Die Kieselspongien der oberen Kreide von Nordwestdeutschland. I. Tetraxonia, Monaxonia und Silicea incertae sedis. Palaeontographica, Supplement 5: 1–385.
Schrammen, A. 1924. Die Kieselspongien der oberen Kreide von Nordwestdeutschland III. Monographien Geologie und Palaeontologie, Serie 1 2: 1–159.
Schuster, A., Pisera, A., Kelly, M., Bell, L.J, Pomponi, S.A., Woerheide, G.E., and Erpenbeck, D. 2018. New species and a molecular dating analysis of Vetulina Schmidt, 1879 (Porifera: Demospongiae: Sphaerocladina) reveal an ancient relict fauna with Tethys origin. Zoological Journal of the Linnean Society 184: 585–604. Crossref
Sollas, W.J. 1885. A classification of the sponges. Annals and Magazine of Natural History (Series 5) 16 (95): 395. Crossref
Sollas, W.J. 1888. Report on the Tetractinellida collected by H.M.S. Challenger, during the years 1873–1876. In: Report on the Scientific Results of the Voyage of H.M.S. Challenger during the years 1873–1876. Zoology 25: 1–458.
Świerczewska-Gładysz, E. and Jurkowska, A. 2022. Campanian (Late Cretaceous) Theonellidae and Phymaraphiniidae (lithistid Demospongiae) from the Miechów and Mogilno-Łódź synclinoria (southern and central Poland): new data and taxonomic revision. Papers in Palaeontology 8: e1426. Crossref
Topsent, E. 1890. Études de Spongiaires. Revue biologique du Nord de la France 2: 289–298.
Topsent, E. 1893. Nouvelle série de diagnoses d’éponges de Roscoff et de Banyuls. Archives de Zoologie expérimentale et générale 3, Notes et Revue 10: xxxiii–xliii.
Vacelet, J. and Vasseur, P. 1971. Éponges des récifs coralliens de Tuléar (Madagascar). Téthys (Supplément 1): 51–126.
Vacelet, J., Vasseur, P., and Lévi, C. 1976. Spongiaires de la pente externe des récifs coralliens de Tuléar (Sud-Ouest de Madagascar). Mémoires du Muséum national d’Histoire naturelle (Zoologie) 49: 1–116.
Wagner, W. 1963. Die Schwammfauna der Oberkreide von Neuburg (Donau). Palaeontographica A 122: 166–248.
Wiedenmayer, F. 1994. Contribution of the knowledge of post-Paleozoic neritic and archibenthal sponges (Porifera). Schweizerische Paläontologische Mitteilungen 116: 1–147.
Wilson, H.V. 1925. Siliceous and horny sponges collected by the U.S. Fisheries steamer “Albatros” during the Philippine Expedition, 1907–1910. Bulletin of the United States National Museum 100: 273–506.
Zittel, K.A. 1878. Studien über fossile Spongien. II. Lithistidae. Abhandlungen der Mathematisch-Physikalischen Classe der Königlich-Bayerischen Akademie der Wissenschaften 13: 65–154.
Acta Palaeontol. Pol. 68 (2): 261–272, 2023
https://doi.org/10.4202/app.01052.2023