
New records of marsupials from the Miocene of Western Amazonia, Acre, Brazil
NARLA S. STUTZ, PATRICIA HADLER, FRANCISCO R. NEGRI, LAURENT MARIVAUX, PIERRE-OLIVIER ANTOINE, FRANÇOIS PUJOS, TIAGO R. JACÓ, EMMANUELLE M. FONTOURA, LEONARDO KERBER, ANNIE S. HSIOU, ROBERTO VENTURA SANTOS, ANDRÉ M.V. ALVIM, and ANA MARIA RIBEIRO
Stutz, N.S., Hadler, P., Negri, F.R., Marivaux, L., Antoine, P.-O., Pujos, F., Jacó, T.R., Fontoura, E.M., Kerber, L., Hsiou, A.S., Ventura Santos, R., Alvim, A.M.V., and Ribeiro, A.M. 2023. New records of marsupials from the Miocene of Western Amazonia, Acre, Brazil. Acta Palaeontologica Polonica 68 (3): 457–475.
The Amazonian region covers a significant part of the South American continent and harbors outstanding biodiversity. However, much of its history is still unknown. This situation has begun to change with paleontological field efforts over the last decades, which have been proving that fossils can be common in this region. Despite their great current species richness and abundance in the area today, marsupials have a sparse fossil record, restricted to a few specimens from handful Cenozoic Amazonian localities. Here we present new records of fossil marsupial teeth from the Solimões Formation (lower Eocene–Pliocene), on the Juruá and Envira riverbanks (Acre, Northwestern Brazil). The localities investigated yield at least four distinct didelphid didelphimorphians at PRE 06 (Ponto Rio Envira: Marmosini ?Marmosa sp., Didelphis cf. D. solimoensis, Thylamys? colombianus, plus unidentified didelphids), and two paucituberculatans from the Juruá River localities (Ponto Rio Juruá: the palaeothentid Palaeothentinae indet. at PRJ 25 and PRJ 33’, and Abderitidae indet. from PRJ 33). In agreement with the associated mammalian faunas, most of the didelphids, except for Thylamys? colombianus from PRE 06, indicate a (?early) Late Miocene age for this locality. Conversely, the abderitid specimens found in situ at PRJ 33 would match a Middle Miocene age. The palaeothentids found at PRJ 25 and PRJ 33’ localities cannot be considered for biostratigraphic inferences, since they were found outside a stratigraphic context. Nevertheless, these paucituberculatans considerably add to our knowledge, as they are the first ever recorded in Brazilian Amazonia.
Key words: Mammalia, Metatheria, Didelphimorphia, Paucituberculata, systematics, historical biogeography, Miocene, Brazil.
Narla S. Stutz [narlasstutz@gmail.com; ORCID: https://orcid.org/0000-0002-1937-3902 ], Programa de Pós-Graduação em Geociências, Universidade Federal do Rio Grande do Sul, Av. Bento Gonçalves 9500, 91501-970, Porto Alegre, Rio Grande do Sul, Brazil, and Laboratoire de Paléontologie, Institut des Sciences de l’Évolution de Montpellier (ISE-M, UMR 5554, CNRS/IRD), Université de Montpellier, Place Eugène Bataillon, 34095, Montpellier, France.
Patricia Hadler [patricia.hadler@ufsc.br; ORCID: https://orcid.org/0000-0003-4897-3111 ], Laboratório de Paleontologia, Universidade Federal de Santa Catarina, Campus Universitário s/n, 88040-900, Florianópolis, Santa Catarina, Brazil.
Francisco R. Negri [frnegriufac@gmail.com; ORCID: https://orcid.org/0000-0001-7396-0334 ], Laboratório de Paleontologia, Campus Floresta, Universidade Federal do Acre, Estrada do Canela Fina Km 12, 69980-000, Cruzeiro do Sul, Acre, Brazil.
Laurent Marivaux [laurent.marivaux@umontpellier.fr; ORCID: https://orcid.org/0000-0002-2882-0874 ] and Pierre-Olivier Antoine [pierre-olivier.antoine@umontpellier.fr; ORCID: https://orcid.org/0000-0001-9122-1818 ], Laboratoire de Paléontologie, Institut des Sciences de l’Évolution de Montpellier (ISE-M, UMR 5554, CNRS/IRD), Université de Montpellier, Place Eugène Bataillon, 34095, Montpellier, France.
François Pujos [fpujos@mendoza-conicet.gob.ar; ORCID: https://orcid.org/0000-0002-6267-3927 ], Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), and Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales (IANIGLA), CONICET-UNCUYO-Mendoza, Av. Ruiz Leal s/n, Parque Gral. San Martín, 5500, Mendoza, Argentina.
Tiago R. Jacó [jaco.tiago@gmail.com; ORCID: https://orcid.org/0000-0002-5903-4673 ], Programa de Pós-Graduação em Sistemática e Conservação da Diversidade Biológica (PPGSCBIO), Secretaria do Meio Ambiente e Infraestrutura do Rio Grande do Sul, Universidade Estadual do Rio Grande do Sul, R. Dr. Salvador França 1427, 90690-000, Porto Alegre, Rio Grande do Sul, Brazil.
Emmanuelle M. Fontoura [emmanuelle.fontoura@acad.ufsm.br; ORCID: https://orcid.org/0000-0002-3907-618X ], Programa de Pós-Graduação em Biodiversidade Animal (PPGBA), Universidade Federal de Santa Maria, Av. Roraima 1000, 97105-900, Santa Maria, Rio Grande do Sul, and Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia (CAPPA), Universidade Federal de Santa Maria, R. Maximiliano Vizzotto 598, 97230-000, São João do Polêsine, Rio Grande do Sul, Brazil.
Leonardo Kerber [leonardokerber@gmail.com; ORCID: https://orcid.org/0000-0001-8139-1493 ], Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia (CAPPA), Universidade Federal de Santa Maria, R. Maximiliano Vizzotto 598, 97230-000, São João do Polêsine, Rio Grande do Sul, Brazil.
Annie S. Hsiou [anniehsiou@ffclrp.usp.br; ORCID: https://orcid.org/0000-0003-2392-6191 ], Laboratório de Paleontologia, Universidade de São Paulo, Av. Bandeirantes 3900, 14040-901, Ribeirão Preto, São Paulo, Brazil.
Roberto Ventura Santos [robertoventurasantos@gmail.com; ORCID: https://orcid.org/0000-0001-6071-8100 ] and André M. V. Alvim [andre.mavaal@gmail.com; ORCID: https://orcid.org/0000-0001-5050-4977 ], Laboratório de Geocronologia, Instituto de Geociências, Universidade de Brasília (UnB), Campus Universitário Darcy Ribeiro ICC - Ala Central, 70910-900, Brasília, Distrito Federal, Brazil.
Ana Maria Ribeiro [ana-ribeiro@sema.rs.gov.br; ORCID: https://orcid.org/0000-0003-4167-8558 ], Seção de Paleontologia, Museu de Ciências Naturais, Secretaria do Meio Ambiente e Infraestrutura, R. Dr. Salvador França 1427, 90690-000, Porto Alegre, Rio Grande do Sul, and Programa de Pós-Graduação em Geociências, Universidade Federal do Rio Grande do Sul, Av. Bento Gonçalves 9500, 91501-970, Porto Alegre, Rio Grande do Sul, Brazil.
Received 22 January 2023, accepted 6 July 2023, available online 29 September 2023.
Copyright © 2023 N.S. Stutz et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Amazon plains and lowlands cover approximately 40% of South America, being the largest forested subregion of the Neotropics. It harbors an outstanding biodiversity, with a high degree of endemism, encompassing one of the richest mammal faunas on the planet, especially in its western part (Patterson and Costa 2012; Meseguer et al. 2022). Nevertheless, the configuration of Amazonia substantially changed throughout the Cenozoic. For example, during the Miocene, but before the Late Miocene, a much larger region, the pan-Amazonia, comprised the drainage basins of the Amazon, Orinoco, and Magdalena rivers, which were all connected, sometimes extending even to the northern part of the Paraná River basin (Hoorn et al. 2010b). The Middle Miocene, particularly, was marked by the existence of the Pebas Mega-Wetland System (PMWS), formed by wetlands, shallow lakes, floodplains, and swamps, which covered a broad area of Western Amazonia from 18–10 Ma (Hoorn et al. 2010b; Boonstra et al. 2015). Around 9 Ma ago, the PMWS gave way to a fluvial/fluviotidal system in Western Amazonia. At about the same time, Andean sediments reached the Atlantic coast for the first time via the drainage system, originating the Amazon River fluvial system as it exists today. These events were influenced by important tectonic activity related to the intensification of the uplifting process in the Northern Andes (Hoorn et al. 2010a, b, 2017).
Tropical regions have long been considered devoid of fossils. However, in Amazonia, for example, fossil remains have proven to be relatively common, being found mainly on riverbanks during the dry seasons (Stutz et al. 2022 and references therein). Indeed, increased field efforts and intensive wet-screening of potentially fossil-bearing sediments, mainly in Western Amazonia, have demonstrated that this region harbored a great mammal diversity over middle Eocene–Holocene times, with approximately 23 orders, 89 families, and 320 species, including Allotheria, Metatheria, and Eutheria (Antoine et al. 2017). Nonetheless, there may be some biases in our current knowledge about the fossil record from the low and middle latitudes of South America, due to taphonomic factors and surface collecting approaches, with, for instance, a greater representation of large mammals in historical findings (Antoine et al. 2017). Metatheria, despite currently possessing great richness and abundance in the area, with didelphid didelphimorphians (Gardner 2007), are known as fossils only by a few samples from a handful of localities, such as Santa Rosa, Río Acre, Madre de Dios, the Fitzcarrald Arch localities, Contamana, and Tarapoto/Juan Guerra vicinity in Peru (Czaplewski 1996; Goin and Candela 2004; Goin and de los Reyes 2011; Antoine et al. 2013, 2016, 2021; Tejada-Lara et al. 2015; Marivaux et al. 2020; Stutz et al. 2022); Cerdas (Croft et al. 2016) and Quebrada Honda (Engelman et al. 2017, 2020) in Bolivia; Urumaco, in Venezuela (Linares 2004); Castilletes Formation (Suárez et al. 2016) and the La Venta local fauna in Colombia (Goin 1997; Suárez Gómez 2019); and Acre, in Brazil (Czaplewski 1996; Cozzuol et al. 2006). Thus, this study aims to partly address this knowledge gap by describing new records of fossil marsupials from the upper Juruá and Envira riverbanks in the Western Amazonian Acre Basin (State of Acre, Brazil). Once described, compared, and taxonomically assigned, these remains provide valuable information on South American marsupials’ biostratigraphy and historical biogeography.
Institutional abbreviations.—UFAC, Universidade Federal do Acre, Acre, Brazil; UFSC, Universidade Federal de Santa Catarina, Florianópolis, Brazil.
Other abbreviations.—D/d, upper/lower deciduous; L, length; M/m, upper/lower molar; P/p, upper/lower premolar; PMWS, Pebas Mega-Wetland System; PRE, Ponto Rio Envira; PRJ, Ponto Rio Juruá; SALMA, South American Land Mammal Age; StA–D, stylar cusps A–D (from anterior to posterior); tal, talonid; tri, trigonid; W, width.
Geological setting
Several field campaigns along the upper Juruá and Envira rivers occurred between 2008 and 2022. The Juruá and Envira river areas are located in the Acre Basin within the State of Acre, Brazil (Fig. 1). They comprise outcrops of the Solimões Formation, part of the eastern extension of the Amazonian Foreland Basin. The Solimões Formation corresponds to the last depositional cycle of the Acre Basin, with thin fluvial-lacustrine sediments spanning a lower Eocene–Pliocene time interval (Cunha 2007), with its upper levels (Neogene) cropping out mostly on riverbanks and road cuts (Lacerda et al. 2021). U-Pb dating on detrital zircons collected at two fossil-yielding sites of the Solimões Formation indicate a Tortonian, Late Miocene age (Bissaro Júnior et al. 2019): one on the banks of the Purus River (8.9±0.13 Ma) and the other on the Acre River (8.5±0.5 Ma). Those ages agree with previous estimations using biochronological data from fossil faunas recovered in Southeastern Acre, which mostly placed it in the Huayquerian South American Land Mammal Age (SALMA; e.g., Negri et al. 2010; Ribeiro et al. 2013; Kerber et al. 2017; Hsiou et al. 2022). Palynostratigraphic analyses on samples collected near the city of Cruzeiro do Sul (Northwestern Acre) indicate a Miocene age for the Solimões Formation and point to a correlation with Pebas and Ipururo formations exposed in Peru (Silva-Caminha et al. 2020). The same authors also suggested a late Middle Miocene marine incursion in the region, thereby extending by 400 km to the south the extend of this previously reported incursion event in Western Amazonia (Silva-Caminha et al. 2020).
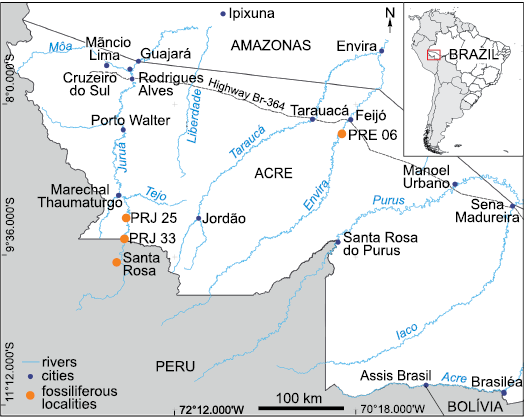
Fig. 1. Location map of the studied localities PRJ 25 and PRJ 33, on the banks of the upper Juruá River, and PRE 06, on the banks of the Envira River, State of Acre, Brazil, as well as the location of Santa Rosa, in Peru, where fossil marsupials have also been found (Goin and Candela 2004). Geographic Coordinate System WGS 84.
Numerous taxa of fossil vertebrates have already been reported for the Solimões Formation, such as diverse extinct forms of crocodylomorphs, chelonians, squamates, birds, and fishes (Ribeiro et al. 2013; Hsiou et al. 2022). Regarding mammals, among the most frequent taxa are rodents (e.g., Kerber et al. 2016, 2017, 2018, 2019a, b, 2022; Fontoura et al. 2022), xenarthrans (e.g., Negri 2004) and notoungulates (e.g., Bergqvist et al. 2002). Remains of other mammalian taxa such as litopterns, astrapotheres, sirenians, cetaceans, primates, and chiropterans have also been reported (see Negri et al. 2010; Ribeiro et al. 2013; Hsiou et al. 2022 for a comprehensive review). As for marsupials, very few studies have been published, limited to unidentified didelphids reported for the Acre River by Czaplewski (1996) and the oldest species of the extant genus Didelphis Linnaeus, 1758, Didelphis solimoensis Cozzuol, Goin, de los Reyes, and Ranzi, 2006, also from a locality on the Acre River (Cozzuol et al. 2006).
The fieldwork along the Juruá River was carried out in a segment bracketed between the city of Marechal Thaumaturgo and the community of Foz do Breu, at the border with Peru (upper Juruá River). Approximately 30 localities were surveyed and designated as PRJ (Ponto Rio Juruá). Two localities on the upper Juruá riverbanks provided marsupial fossils, namely PRJ 25 (Figs. 1–3; 9°11’06.8”S 72°42’37.2”W) and PRJ 33 (Figs. 1, 4; 9°24’24.6”S 72°43”27.6”W).
The locality of PRJ 25 is located near the Belford property, at the left margin of the Juruá River. The outcrop is composed of two parts: a horizontal surface identified as “H”, and a vertical outcrop identified as “V” (Fig. 2A, B). The horizontal surface corresponds to the lithologies between 0 and 0.45 m in the stratigraphic profile (Fig. 2A). It consists of the intercalation of fractured red siltstone and fine-grained yellowish sandstone dipping 15–18º S. The siltstone and sandstone were cemented by carbonate. The vertical outcrop corresponds to the interval lithologies between 0.5 and 1.6 m in the profile and consisted of the same sedimentary sequence as the horizontal surface. In this part occurs four levels of continuous carbonate concretions within the siltstone. The strata were less inclined than the horizontal part of the outcrop (10–15º). The horizontal part of the outcrop is covered by fossil-rich Miocene–Holocene fluvial sand and gravel deposits as shown in Fig. 2. These allochthonous deposits are related to the erosion of upstream strata transported by the river stream and deposited recently in this outcrop (Fig. 2C). The corresponding fossil fragments include silicified wood and vertebrate remains. The latter specimens provide markers of different epochs (e.g., Astrapotheria and Purussaurus remains). All the fossil specimens from PRJ 25 originate from these out-of-sequence sands and gravels, including the only dental fragment of marsupial retrieved (UFAC-CS 418). From this ensemble, Kerber et al. (2017) also reported fossil caviomorphs, likely of Miocene affinities (unidentified potamarchine and erethizontid).

Fig. 2. Stratigraphic section (A) and photos (B, C) of the locality PRJ 25, with the Solimões Formation as a substratum, located on the left margin of the Juruá River, State of Acre, Brazil. CTS, Holocene conglomerate, with a mixture of Miocene–Holocene pebbles and fossils (mostly silicified wood and vertebrates), which may cover indifferently any Miocene layers. Vertebrate fossils originate from the sand and gravel emerging on C (star). Abbreviations: H, horizontal; V, vertical.

Fig. 3. Detailed horizontal profile of the locality PRJ 25, on the left margin of the Juruá River, State of Acre, Brazil. Fossils were found in the gravel alluvial deposit and lateral sandstone/siltstone.
The locality PRJ 33 is situated on the left bank of the Juruá River and has about 2–3 m height (Fig. 4A). In this outcrop, occurs a layer of massive reddish siltstone with horizontal fractures and local faults filled by carbonate concretions. These fractures tend to be laterally discontinuous along the outcrop. The siltstone is in erosional contact with the 10–15 cm-thick level of fossiliferous conglomerate composed by clasts of carbonate concretions and rare quartz supported by a reddish sandy-siltstone matrix. The conglomerate level is in direct contact with a layer of massive fine grayish sandstone at the top of the section. At PRJ 33, three specimens were collected: (i) in situ within the conglomerate layer (UFAC-CS 101 and 109); (ii) in the Holocene mixed sediments that comprise reworked bones (PRJ 33’, UFAC-CS 65). These last sediments are placed at the bottom of the outcrop, close to the river, and have fossil specimens outside of their stratigraphic context. Ribeiro et al. (2013) and Kerber et al. (2017) have reported dental remains of the Paleogene caviomorph rodent Eoincamys and Marivaux et al. (2023) have recently described a Paleogene eosimiid primate extracted from the mixed sediments.
In the Envira River area, the outcrops were designated as PRE (Ponto Rio Envira). Six marsupial teeth were retrieved at the locality PRE 06, also called locality “Coqueiros” (Figs. 1, 5; 08°18’12.6”S 70°26’39.9”W). Four laterally equivalent sampling points (A–D) were surveyed (Fig. 5A), but only one (C) yielded fossil marsupials after wet-screening of the sediments. Briefly, the sedimentary sequence at sampling points A–C (Fig. 5B, C) comprises layers of grayish-green and red argillite, which are in erosive contact with the conglomeratic level very rich in fossil content. The conglomerate is composed of carbonate concretions and numerous fossil fragments (such as remains of fishes, reptiles, and mammals, currently under study) supported by a silty-sandy matrix. This conglomerate is in direct contact with intercalated layers of brown clay and very fine to fine yellowish sand (rhythmite). Sampling points A–C in section belong to a same stratigraphic horizon, while sampling point D consists of an accumulation of fossil fragments found on the Envira River floodplain and, hence, out of stratigraphical context. Regarding the fossil content from PRE 06, there is fossil wood, with a diverse flora reported by Kloster et al. (2017) that resembles the one currently present in the region. Furthermore, recent studies revealed a diverse record of caviomorph rodents from PRE 06A–C (Cardiatherium sp., Caviidae, Hydrochoerinae; Potamarchus sp. and cf. Pseudopotamarchus, Dinomyidae, Potamarchinae; and Neoepiblema sp., Neoepiblemidae; Fontoura et al. 2022), which suggest a Late Miocene age.

Fig. 4. Stratigraphic section (A) and photo (B) of the Miocene locality PRJ 33, Solimões Formation, located on the left bank of the Juruá River, State of Acre, Brazil.

Fig. 5. Photo (A) and stratigraphic section (B and C) of the Upper Miocene locality PRE 06, Solimões Formation, located on the right bank of the Envira River, State of Acre, Brazil.
Material and methods
The fossil specimens were collected through wet-screening of the sediments from previously recognized fossil-bearing layers (sands and conglomerates), using sieves of 0.6, 1, and 2 mm, and then sorted out using a stereomicroscope. The specimens are permanently housed in the collection of the Paleontology Laboratory of the Universidade Federal do Acre, Floresta campus, Cruzeiro do Sul, Acre (UFAC-CS).
The fossils were photographed with Scanning Electron Microscope (SEM) facilities of the Centro de Microscopia e Microanálise (CMM, UFRGS, JEOL JSM 6060) and at the Laboratório Central de Microscopia e Microanálise (LabCEMM, PUCRS, FEI Inspect F50). The measurements of maximum anteroposterior length and maximum labiolingual width were made using a manual Mitutoyo caliper, with a precision of 0.02 mm. The nomenclature of the molar structures and the Marsupialia systematics follow Goin et al. (2016). Taxonomic identifications were made according to Abello (2007), Voss and Jansa (2009), and Abello and Rubilar-Rogers (2012). They were directly compared with fossil metatherians from the locality TAR-31 (Juan Guerra, San Martín, Peru) deposited at the Vertebrate Paleontology Department of the Museo de Historia Natural of the Universidad Nacional Mayor San Marcos (MUSM) in Lima, Peru, and with specimens of modern didelphids deposited in the Coleção de Fragmentos Ósseos do Laboratório de Paleontologia and Coleção de Mamíferos do Departamento de Ecologia e Zoologia, Universidade Federal de Santa Catarina (UFSC), Florianópolis, Brazil, and at the Mammal Collection of the MUSM.
Systematic palaeontology
Class Mammalia Linnaeus, 1758
Infraclass Metatheria Huxley, 1880
Supercohort Marsupialia Gill, 1872
Order Didelphimorphia Gill, 1872
Family Didelphidae Gray, 1821
Subfamily Didelphinae Gray, 1821
Tribe Marmosini Hershkovitz, 1992
Genus Marmosa Gray, 1821
Type species: Didelphis murina Linnaeus, 1758, Recent, South America, by monotypy.
?Marmosa sp.
Fig. 6A.
Material.—UFAC-CS 413, part of the stylar shelf (StC–E) of a right ?M2 from the locality PRE 06, sampling point C, Envira River, State of Acre, Brazil; Solimões Formation, Acre Basin, (?lower) Upper Miocene.
Measurements.—StE–ectoflexus = 1.8 mm; Metacone–labial margin = 2.36 mm; Metacone–StE = 2.68 mm.
Description.—The specimen is worn and broken at the level of the StC and the trigon basin. It includes part of the stylar shelf with the StC–E, and the metacone. The metacone is opposite to StD, relatively far from the latter, and lingually oriented. There is a subtle basin between the metacone and the StC and StD. The premetacrista is short and slightly labiolingually curved. The postmetacrista is longer, extending until the StE with an oblique orientation. There is a crest-like structure at the base of the posterior wall of the metacone, however, it is not possible to state to which crest it corresponds, it could also be a product of the fragmentary and worn state of the molar. There is a small crest-like structure lingually to the StC. The labial margin has a steep oblique orientation from the StC to the StE. The StC and StD are subequal in size; both poorly developed but still visible. The StC and StD are not twinned or close to each other. The StE is posteriorly broken, but it is still possible to observe a small and narrow cusp.
Remarks.—Morphological comparisons with similarly sized didelphids indicate that UFAC-CS 413 is smaller than species of Didelphis, the latter having more rounded stylar cusps. UFAC-CS 413 is approximately the same size as species of Lutreolina but has a smaller StC. In the species of Philander, the StC is more developed and morphologically different. UFAC-CS 413 shows closer affinities with the species of Marmosa in morphology, such as the presence of StE, StD and StC (Rossi et al. 2010). UFAC-CS 413 is larger than the teeth of the recent species Marmosa demerarae (Thomas, 1905). Fossil representatives of the genus include Marmosa laventica, the oldest record of Marmosa, which was described for the Middle Miocene deposits of La Venta, Colombia (Marshall 1976a). Marmosa laventica is larger than the living species of Marmosa, among other diagnostic characteristics. However, it was described based only on lower dentition. Later, Goin (1997), when revisiting the diagnosis of M. laventica, assigned it to Micoureus laventicus and added upper molars in its hypodigm. However, recent molecular analyses considered Micoureus as a synonym of Marmosa, being now recognized as a subgenus of the latter (Voss and Jansa 2009). UFAC-CS 413 is larger than the specimens reported by Goin (1997), and, because of its incomplete state, the diagnostic characteristics of M. laventica could not be observed on it. Marshall (1976a) also reported specimens of Marmosa sp. from La Venta, which are smaller than M. laventica, and thus not compatible in size with UFAC-CS 413. The size of UFAC-CS 413 also distinguishes it from Marmosa sp. 1 of Suárez Gómez (2019), which is reported as being ~40% smaller than M. laventica. Antoine et al. (2013) reported a partial M2 of Marmosa (Micoureus) cf. laventica for the lower Middle Miocene locality MD-67 in Peru, which differs from UFAC-CS 413 in being smaller and not possessing StE. Yet, both specimens show a large stylar shelf and StC and StD. Compared to Hesperocynus, Sparassocynus, and Thylatheridium, UFAC-CS 413 does not possess a very long postmetacrista and the metacone does not seem to be a tall cusp, but the latter could be due to the great wear (Goin et al. 2000; Forasiepi et al. 2009; Abello et al. 2015).
Therefore, UFAC-CS 413 was assigned to Marmosini, ?Marmosa sp. due to the morphological resemblance of it with Marmosa. Nevertheless, the incomplete state of the specimen hampers a more precise taxonomical identification.
Tribe Didelphini Gray, 1821
Genus Didelphis Linnaeus, 1758
Type species: Didelphis marsupialis Linnaeus, 1758, Recent, Central and South America and Mexico, by subsequent designation (Thomas 1888).
Didelphis cf. Didelphis solimoensis Cozzuol, Goin, de los Reyes, and Ranzi, 2006
Fig. 6C.
Material.—UFAC-CS 415, left DP3 from the locality PRE 06, sampling point C, Envira River, State of Acre, Brazil; Solimões Formation, Acre Basin, (?lower) Upper Miocene.
Measurements.—L = 3.08 mm; W = 3.22 mm.
Description.—This DP3 is anteroposteriorly elongate in its labial portion and relatively narrow. The protocone is slightly anterior to the paracone, forming a small, triangular-shaped trigon basin, which is deep and anteroposteriorly compressed. The metacone is conic and posteriorly oriented. It is high and well developed. At its dorsolingual base, there is a small metaconule. The centrocrista is well developed and U-shaped. The preparacrista is almost vertical and does not reach the anterior cingulum/StA. There is a very long postmetacrista, which is obliquely oriented towards the StE. The pre- and postprotocrista are well developed, short, and oblique. There is a well-developed anterior cingulum passing along the base of the paracone and joining a small paraconule. At its labial extremity, there is a small StA. A well-marked labial cingulum is present, extending from StA up to StC. The StB is high and fused with the paracone. Next to StB there is a small and rounded StC, which is closer to StD. The StD is wide and slightly shorter than StB, it is located labial to the metacone and followed by a marked crest, which is oriented parallel to the anteroposterior dental axis. At its end, a small StE forms a rounded posteriormost tip.
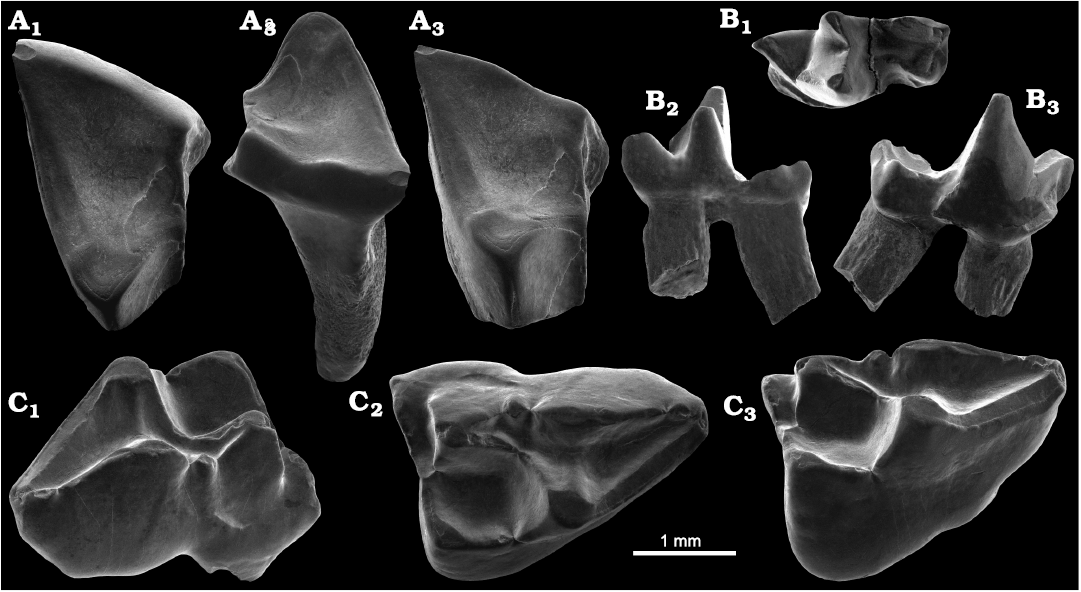
Fig. 6. Didelphids from the locality PRE 06, Envira River, Acre State, Brazil; (?lower) Upper Miocene. A. Marmosini, ?Marmosa sp., UFAC-CS 413, partial right ?M2; in occlusal (A1); approximately labial (A2). and approximately lingual (A3) views. B. Thylamys? colombianus Goin, 1997, UFAC-CS 202, left ?m4; in occlusal (B1), approximately lingual (B2), and approximately labial (B3) views. C. Didelphis cf. D. solimoensis Cozzuol, Goin, de los Reyes, and Ranzi, 2006, UFAC-CS 415, left DP3; in labial (C1), occlusal (C2), and approximately lingual (C3) views.
Remarks.—Based on its morphological characteristics and dimensions, UFAC-CS 415 belongs to a didelphid smaller than species of Didelphis, Lutreolina, Philander, and Chironectes but larger than Marmosa murina (Linnaeus, 1758). Compared with UFAC-CS 413, UFAC-CS 415 is morphologically different from it, and it resembles the DP3 morphology of Didelphis, such as the development and position of the cusps, trigon basin, and stylar cusps, and well-developed anterior cingulum and by being anteroposteriorly elongate. However, UFAC-CS 415 has smaller dimensions than what it is observed in the living species of Didelphis. Didelphis solimoensis, from the Upper Miocene of Acre, is considered the smallest known species of the genus (Cozzuol et al. 2006). UFAC-CS 415 might be slightly smaller than D. solimoensis. However, it should be noted that D. solimoensis is only known from an isolated dentary with p3–m2. Still, given that the occlusal morphology of UFAC-CS 415 resembles the one of Didelphis DP3s and that D. solimoensis is the smallest Didelphis species described in the literature, UFAC-CS 415 was assigned to Didelphis cf. D. solimoensis, pending the discovery of additional specimens of the former, especially preserving the upper dentition.
Tribe Thylamyini Hershkovitz, 1992
Genus Thylamys Gray, 1843
Type species: Didelphis elegans Waterhouse, 1839, Recent, Chile, by monotypy.
Thylamys? colombianus Goin, 1997
Fig. 6B.
Material.—UFAC-CS 202, left ?m4 from the locality PRE 06, sampling point C, Envira River, State of Acre, Brazil; Solimões Formation, Acre Basin, (?lower) Upper Miocene.
Measurements.—L = 1.91 mm, W = 0.9 mm.
Description.—UFAC-CS 202 is a small lower molar with high, well-developed trigonid cuspids and a talonid much lower than the trigonid, with scarcely developed cuspids. The paraconid is conspicuous, lower than the metaconid, and slightly labial and distant from it, with a sharp anterior and occlusal tip. The protoconid is large and prominent, being the highest and largest cuspid of the whole molar. The metaconid is opposite to and much lower than the protoconid. The talonid is narrower than the trigonid but is well developed compared to the small didelphids’ condition. The entoconid is laterally compressed and inconspicuous, possibly due to the molar wear, and it seems to be situated more anteriorly than the hypoconid. The hypoconid is short and inconspicuous, being positioned lingual to the protoconid. The hypoconulid is well developed, triangular-shaped, and lingually located, connected to the posthypocristid via a subtle postcristid. The mesiolingual vertical crest of the paraconid forms a keel on the anteriormost part of the molar. The postparacristid is well developed and obliquely oriented, meeting a conspicuous preprotocristid. The postprotocristid is almost transversally oriented, meeting a shorter premetacristid and forming a shallow sulcus. The pre- and post-entocristids are poorly developed, the preentocristid having a labial orientation. The postentocristid connects the entoconid to the hypoconulid. The cristid obliqua is small and almost parallel to the anteroposterior dental axis. A narrow anterobasal cingulid is visible from the base of the paraconid to the base of the protoconid. There is a faint posterior cingulid on the posteriormost wall of the molar. Two roots are preserved, an anterior rounded one and a posterior one, which is larger and ovoid.
Remarks.—We interpret UFAC-CS 202 as being a ?m4 due to the presence of a slender and long talonid, a more lingual position of the hypoconid than what is commonly observed on m3, and the absence of a wear-facet on the distal side of the tooth. The posterior cingulid present on this specimen is a plesiomorphic characteristic since the absence of this structure is considered to be the single unambiguous synapomorphy of Didelphidae. However, some taxa have already been reported to possess this feature (Beck et al. 2022). Compared to basal metatherians that possess a posterior cingulid, UFAC-CS 202 differs from Marmosopsis juradoi Paula Couto, 1962, by having longer talonid and hypoconulid; it differs from Monodelphopsis travassosi Paula Couto, 1952, by showing high trigonid, which is wider than the talonid and entoconid subequal to hypoconid in size and height (Marshall 1987). UFAC-CS 202 is smaller than m4 of the species of Chironectes, Didelphis, Caluromys, Lutreolina, and Philander, but larger than those of Chacodelphys and the extinct didelphid Sairadelphys Oliveira, Nova, Goin, and Avilla, 2011. UFAC-CS 202 differs from the species of Monodelphis in having a longer and narrower talonid. UFAC-CS 202 is smaller than m4 of most species of Marmosa. Morphological differences with the species of Marmosa are an anterolabial cingulid and entoconid less developed and the absence of a labial cingulid. The m4 of the species of Gracilinanus have a larger entoconid and no posterior cingulid, and those of Cryptonanus have m4 with a marked labial cingulid. In the species of Marmosops, the hypoconulid is more posterior, the hypoconid has a more labial position, the metaconid and protoconid are closer and the entoconid is basally larger concerning the condition observed on UFAC-CS 202. UFAC-CS 202 has the same size as m4s of the species of Thylamys; the position of the trigonid cuspids and the entoconid and hypoconulid is also similar. Regarding other fossil species of Thylamys, UFAC-CS 202 is larger than m4 of Thylamys minutus Goin, 1997, from the Middle Miocene of La Venta, Colombia (Goin 1997). Nevertheless, it has approximately the same size as Thylamys colombianus, also from La Venta, which has m3 with W = 1.55 mm and m1? with L = 1.9 mm and W = 0.9 mm (only a partially preserved m4 for this taxon was found; Suárez Gómez 2019). Besides, UFAC-CS 202 also resembles lower molars of T. colombianus in having: (i) a paraconid more mesio-labially displaced, being more distant from other cuspids, (ii) a labio-lingually compressed entoconid, (iii) a long talonid, and (iv) a similar relative entoconid/hypoconulid position. Nevertheless, no well-preserved m4 has been described until now for Thylamys colombianus, hampering a more precise comparison. The m1 of Thylamys cf. T. colombianus from TAR-31 (late Middle Miocene, Peru; Stutz et al. 2022) has a paraconid and a hypoconid more anteriorly and more labially placed, respectively, but with a position of the entoconid/hypoconulid similar to that in UFAC-CS 202. Based on living species, recent gene-based phylogenies suggested that Thylamys diverged from other didelphids later than the presumed age of the PRE 06 deposits (not until the Pliocene, and, for the last common ancestor of Thylamys and Gracilinanus + Cryptonanus, not until ~10 Ma) (Jansa et al. 2014; Beck and Taglioretti 2020). Thus, we chose to consider Thylamys? as a generic assignment, pending a better assessment of the concerned Miocene specimens, whether they belonged to this genus or a stem representative of Thylamyini.
Didelphidae indet.
Fig. 7.
Material.—UFAC-CS 412, right p1; UFAC-CS 414, right ?p2; UFAC-CS 416, right ?P3; UFAC-CS 427, left dp3 from the locality PRE 06, sampling point C, Envira River, State of Acre, Brazil; Solimões Formation, Acre Basin, (?lower) Upper Miocene.
Measurements.—UFSC-CS 412 L = 2.27 mm, W = 1.18 mm; UFAC-CS 414 L = 3.36 mm, W = 1.54 mm; UFAC-CS 416 L = 3.24 mm, L = 1.95 mm; UFAC-CS 427 L = 2.3 mm, W tal = 1.42 mm, W tri = 1.32 mm.
Description.—UFAC-CS 412 (Fig. 7B): small and slender p1 displaying two roots. The crown consists of one single central cuspid, rounded and short (i.e., stocky). It is followed by a subtle cristid running postero-labially. A small cingulid is present posterior to the cuspid, with a minute labially situated accessory cuspid.
UFAC-CS 414 (Fig. 7C): this specimen is larger than UFAC-CS 412. It is robust, with one single cuspid, which is broken and abraded. Posteriorly to this cuspid, there is a tenuous cristid labially oriented. A posterior cingulid is present, with a marked accessory cuspid.
UFAC-CS 416 (Fig. 7A): this specimen is a large upper premolar with two roots. A subtle anterior cingulum is present. There is only one cusp, which is high, robust, and posteriorly oriented. The anterior margin of the cusp is rounded and at the posterior one is a marked, sharp, and centrally oriented crest. There is a posterior cingulum, which is particularly developed in its labial margin. A small accessory cusp is present, occupying a posterior labial position.
UFAC-CS 427 (Fig. 7D): this specimen does not possess any signs of roots in its ventral side, which is open, exposing a broad portion of the tooth’s interior. It presents an oval crown outline and robust molariform pattern, being anteroposteriorly compressed and with a rounded labial margin. The trigonid is much higher than the talonid, especially the proto- and metaconids. Proto-, meta-, hypo-, and entoconid are large and blunt. A small, but individualized paraconid is present, which is labially displaced. The protoconid is a large and high cuspid, connected to the paraconid by a short preprotocristid. The protoconid and metaconid are closely positioned, without an evident postprotocristid. The protoconid is higher than the metaconid, but the latter is also a large cuspid. The talonid is short. The hypoconid appears slightly broken on its dorsal tip and labially displaced, evincing a wide talonid basin. The entoconid is robust and anteroposteriorly long, with marked, but short pre- and postentocristids. A minute hypoconulid can be observed, which is twinned with the entoconid and situated at its posterolabial base. The premetacristid is not visible, but the postmetacristid is strongly developed. The cristid obliqua is marked and obliquely oriented, but short. The posthypocristid is marked, short and slightly oblique in relation to the anteroposterior axis of the tooth. A discrete and short anterolabial cingulid is present.
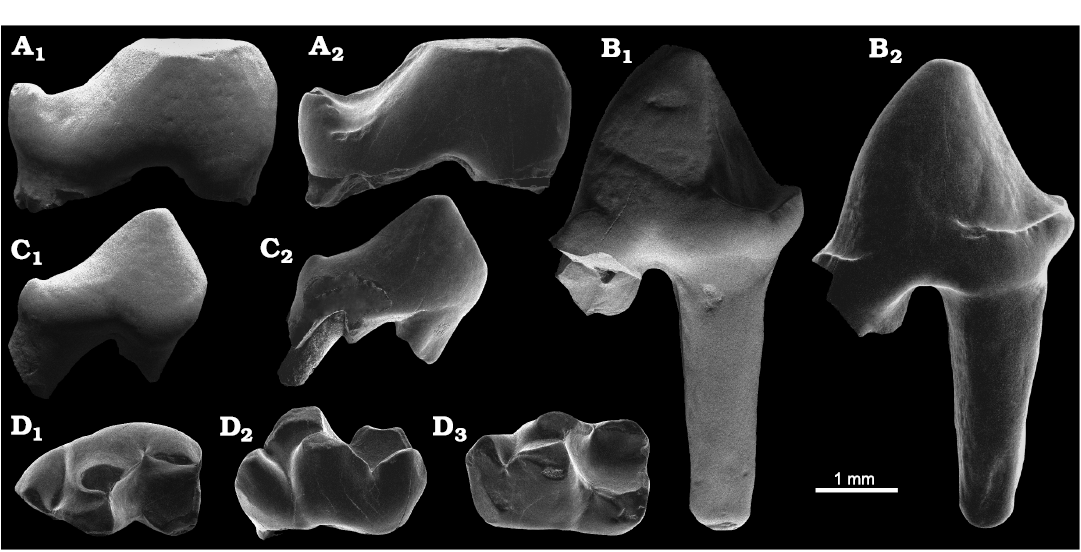
Fig. 7. Didelphidae indet. from the locality PRE 06, Envira River, Acre State, Brazil; (?lower) Upper Miocene. A. UFAC-CS 416, right ?P3; in labial (A1) and lingual (A2) views. B. UFAC-CS 412, right p1; in labial (B1) and lingual (B2) views. C. UFAC-CS 414, right ?p2; in labial (C1) and lingual (C2) views. D. UFAC-CS 427, left dp3; in occlusal (D1), labial (D2), and lingual (D3) views.
Remarks.—We identify UFAC-CS 412 as a lower p1 due to its size, and inconspicuous posterior cingulid and accessory cuspid. UFAC-CS 414 is tentatively identified as a p2 because of its size and the development of the posterior cingulid and posterior accessory cuspid. UFAC-CS 416 is considered to likely be a P3 based on its rounded anterior and cutting posterior margins, as observed in didelphids, except for species of Caluromys Allen, 1900, Caluromysiops Sanborn, 1951, Glironia Thomas, 1912, and Hyladelphys Voss, Lunde, and Simmons, 2001 (Voss and Jansa 2009). According to Voss and Jansa (2009), didelphids, together with other marsupials that display plesiomorphic dental traits (such as microbiotheres), have standard lower premolars with only one dominant cuspid, and a distal cingulid frequently producing a posterior accessory cuspid. The P3 of paucituberculatans has a posterior portion higher than the anterior one and palaeothentoids generally have uniradicular and morphologically indistinct p1 and p2 (Abello 2007). Members of the clade Argyrolagidae have reduced premolars nearly styliform (Simpson 1970). Therefore, we preferred to classify the three specimens mentioned above as unidentified Didelphidae due to (i) the lack of diagnostic characteristics on didelphid premolars; (ii) and Didelphidae being the only metatherian family already recognized at the outcrop PRE 06.
UFAC-CS 427 is identified as a left dp3 due to the absence of robust roots and very closely positioned protoconid and metaconid. Compared to other Miocene didelphids, UFAC-CS 427 does not have a compatible size with Thylamys? colombianus and Didelphis cf. D. solimoensis, also found at PRE 06 (see above) or with the two Thylamys spp. from the La Venta deposits (Suárez Gómez 2019). Besides, it is smaller than dp3s of Marmosa laventica Marshall, 1976a, also from La Venta, which possess very different occlusal morphology, with, for instance, well-developed anterolabial cingulid and hypoconulid (Suárez Gómez 2019). UFAC-CS 427 is also different from Lutreolina materdei Goin and de los Reyes, 2011, from the Upper Miocene of Acre River, in Peru, which has large anterolabial cingulid and paraconid and hypoconid not labially displaced, among other distinctive characteristics. However, UFAC-CS 427 and Lutreolina materdei bear a minute hypoconulid (Goin and de los Reyes 2011). When compared to modern didelphids, UFAC-CS 427 stands out by its molariform morphology, with a complete trigonid, differing from the species of Lestodelphys Tate, 1934, Marmosa Gray, 1821, and Thylamys Gray, 1843, which possess a blade-like trigonid on the dp3, and from those of Caluromys, Caluromysiops, and Tlacuatzin Voss and Jansa, 2003, which have a very small paraconid or no paraconid at all. Hyladelphys is another didelphid with a peculiar dp3, which is significantly reduced (Voss and Jansa 2009). Besides, UFAC-CS 427 shows a very discrete anterolabial cingulid, which is a more developed structure in the species of Chironectes Illiger, 1811, Lutreolina Thomas, 1910, Metachirus Burmeister, 1854, Monodelphis Burnett, 1830, and Philander Brisson, 1762. UFAC-CS 427 also possesses labially displaced paraconid and hypoconid, differing from the species of Chironectes, Glironia, and Lutreolina. The protoconid and metaconid are very close in UFAC-CS 427, distinguishing it from the species of Hyperdidelphys Ameghino, 1904, Lutreolina, Metachirus, and Philander. Other distinctive features of UFAC-CS 427 are a short preprotocristid, distinguishing it from the species of Metachirus, which have a long and oblique preprotocristid, a short talonid (long in Didelphis), and a large entoconid (small in Marmosops Matschie, 1916). The most outstanding distinctive feature of UFAC-CS 427 is its extremely reduced hypoconulid compared to the species of Chironectes, Didelphis, Glironia, Lutreolina, Marmosops, Monodelphis, Philander, and Thylophorops Reig, 1952. The species of Cryptonanus Voss, Lunde, and Jansa, 2005, Gracilinanus Gardner and Creighton, 1989, and Chacodelphys Voss, Gardner, and Jansa, 2004, have much smaller teeth than UFAC-CS 427. The corresponding didelphid was approximately Marmosa-sized. Thus, the scarcity of the material and the set of characteristics found in UFAC-CS 427 do not allow its attribution to any of the described didelphid genera, pending the discovery of more specimens.
Order Paucituberculata Ameghino, 1894
Superfamily Palaeothentoidea Sinclair, 1906
Family Palaeothentidae Sinclair, 1906
Subfamily Palaeothentinae Sinclair, 1906
Palaeothentinae indet.
Fig. 8A, B.
Material.—UFAC-CS 65, left M2; UFAC-CS 418 right ?M2 from the locality PRJ 33’ (UFAC-CS 65); and PRJ 25 (UFAC-CS 418), Juruá River, State of Acre, Brazil; Solimões Formation, Acre Basin, ?Middle Miocene.
Measurements.—UFAC-CS 65 L = 1.91 mm, W = 2.2 mm; UFAC-CS 418 L = 1.98 mm, W = 1.91 mm.
Description.—UFAC-CS 65 (Fig. 8A) is an upper molar, quadrangular in occlusal outline, with a much broader base compared to the crown in lateral perspective. Antero-lingually to the StB, at its base, there is a smaller cusp, the paracone, that is poorly developed. The protocone is wide but short and is placed opposite to StB. The metacone is a well-developed cusp, slightly smaller than StC+D, and placed on the anterior portion of the latter. The trigon basin is deep. The metaconule is well developed and wide but not much higher than the talon basin, having approximately the same height as the protocone. It is located opposite to StC+D. At the base of the metacone, there is a sulcus separating it from the metaconule. The postprotocrista is subequal in length to the preprotocrista (Fig. 8A3); dorsal to it there is a shallow entoflexus. The postmetaconular crest is moderately wide and labially oriented, the molar having a shorter outline, thereby differing from the condition that characterizes M1s of other palaeothentids. A narrow anterior cingulum extends from a point immediately anterior to the StB, converging toward the preprotocrista, but oblique and dorsally to the latter. The StA is vestigial or absent. The StB is conical-shaped and represents the highest cusp. The StC+D is well developed but smaller than the StB, from which it is separated by a deeply excavated ectoflexus, with a small style at its base.
UFAC-CS 418 (Fig. 8B) is also a quadrangular-shaped upper molar, appearing worn and abraded due to transport, the labial and lingual sides of the molar having a thicker enamel layer compared to the structures on the occlusal surface. The paracone is small and twinned with the StB, being placed at the antero-lingual slope of the latter. The protocone is well developed, but short. The metacone is twinned with the StC+D, but shorter than the latter and located on its anterior portion. The trigon basin is deep, small, and narrow. Opposite to StC+D there is a well-developed metaconule, preceded by a short premetaconular crest, with a small premetaconular cuspule, and followed posteriorly by a longer postmetaconular crest that runs parallel to the lingual side of the tooth. Both the preprotocrista and postprotocrista are short. A broad but short anterior cingulum is present, running from a well developed StA to almost the dorsolingual base of the protocone. Beneath the metaconule and protocone, there is a deep entoflexus on the lingual margin of the tooth. There is a deep U-shaped ectoflexus on the labial margin of the tooth. The StB is larger and longer than the StC+D. The StC+D is well-developed and the highest cusp of the molar.
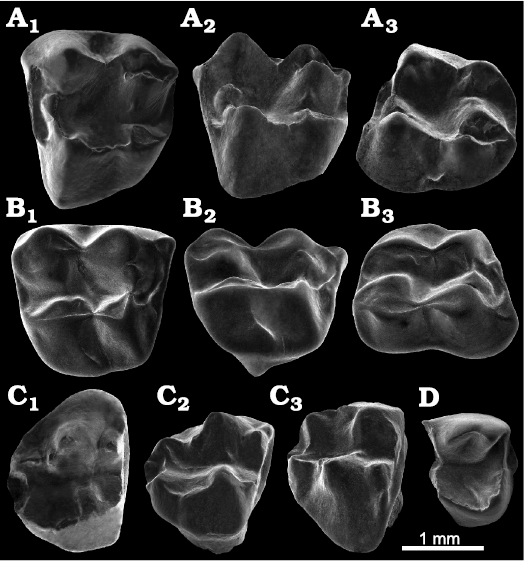
Fig. 8. Paucituberculatans from the upper Juruá River, State of Acre, Brazil, ?Middle Miocene. A. Palaeothentinae indet., UFAC-CS 65, left M2, PRJ 33’, out of stratigraphic context, allochthonous sediments; in occlusal (A1), approximately lingual (A2), and approximately labial (A3) views. B. Palaeothentinae indet., UFAC-CS 418, right ?M2, PRJ 25, out of stratigraphic context, allochthonous sediments; in occlusal (B1), approximately lingual (B2), and approximately labial (B3) views. C, D. Abderitidae indet., PRJ 33, conglomerate level, in situ sediments. C. UFAC-CS 101, right M2; in occlusal (C1), approximately lingual (C2), and approximately labial (C3) views. D. UFAC-CS 109, left fragmentary ?m2; in occlusal view.
Remarks.—UFAC-CS 65: the presence of a premetaconular cusp and the metacone situated in the first half of the StC+D allow us to assign this specimen to the family Palaeothentidae according to Abello (2007). Also, according to this same author, the protocone and metaconule directly opposite to StB and StC+D, regarding an axis perpendicular to the anteroposterior one, unambiguously point to Palaeothentinae affinities for UFAC-CS 65. Conversely, decastine palaeothentids are characterized by a protocone posterior to the StB on M1–2, differing from UFAC-CS 65 (Abello 2007). Among Palaeothentinae, Carlothentes chubutensis Bown and Fleagle, 1993, is solely known through lower molars, precluding further comparison with UFAC-CS 65. The upper molars of Palaeopanorthus primus Ameghino, 1902, although having conspicuous paracone and metacone, have a sizeable premetaconular cusp and a moderately developed to large metaconule, differing from UFAC-CS 65 (Abello 2007). In the species of Palaeothentes Ameghino, 1887, a paracone on M2 is only registered in Palaeothentes marshalli Bown and Fleagle, 1993, Palaeothentes migueli Bown and Fleagle, 1993, and Palaeothentes serratus Engelman, Anaya, and Croft, 2017 (Abello 2007; Engelman et al. 2017). Palaeothentes serratus from Quebrada Honda (Middle Miocene of Bolivia), differs from UFAC-CS 65 in possessing a highly reduced metacone. The other species of Palaeothentes from Quebrada Honda, Palaeothentes relictus Engelman, Anaya, and Croft, 2017, is not represented by upper molars (Engelman et al. 2017), hampering comparison with UFAC-CS 65. As for P. migueli and P. marshalli, the molars of P. migueli are smaller than UFAC-CS 65, while those of P. marshalli are closer in size to UFAC-CS 65. Besides, P. marshalli has well-developed metacone and teeth that are more triangular-shaped in occlusal view, as UFAC-CS 65. Nevertheless, the M1–M2 of the species of Palaeothentes do not present many significant differences with the ones of Palaeopanorthus primus (i.e., differences in the proportional size of the paracone, metacone and premetaconular crest; Bown and Fleagle 1993; Abello 2007). Therefore, considering that UFAC-CS 65 is an isolated specimen and that the M2 is not a very distinctive tooth among some Palaeothentinae taxa, we chose an open taxonomic identification, highlighting the closer affinities of UFAC-CS 65 especially with P. marshalli, but also to P. migueli and Palaeopanorthus primus.
UFAC-CS 418 can be considered as belonging to Palaeothentoidea due to the differences in the enamel thickness observed between the lateral and occlusal sides of the molar, and as a representative of Palaeothentidae due to (i) the metacone located on the anterior half of the StC+D, and (ii) the presence of a premetaconular cusp on upper molars (Abello 2007). As mentioned above, for UFAC-CS 65, the M2 of palaeothentines is not very diagnostic; therefore, an open taxonomic identification is again favored. As with UFAC-CS 65, UFAC-CS 418 also has closer affinities with P. marshalli, P. migueli, and Palaeopanorthus primus.
Compared to UFAC-CS 418, UFAC-CS 65 has a less developed anterior cingulum, StA and paracone; a more developed metacone; a more defined triangular outline; and it is larger. Because both are here considered as M2s, these differences would not be related to distinct dental loci but possibly to different taxa. Thus, both UFAC-CS 65 and UFAC-CS 418 are considered to belong to palaeothentines. Yet, a more precise taxonomic assignment would require more associated specimens (especially of other dental loci) and a better knowledge of the morphology of the already described Palaeothentinae.
Family Abderitidae Ameghino, 1889
Abderitidae indet.
Fig. 8C, D.
Material.—UFAC-CS 101, right M2; UFAC-CS 109, left ?m2; from the locality PRJ 33, stratified conglomerate level, Juruá River, State of Acre, Brazil; Solimões Formation, Acre Basin, Middle Miocene.
Measurements.—UFAC-CS 101, L = 1.58 mm, W = 1.9 mm; UFAC-CS 109, W = 1.41 mm, L = 1.08 mm.
Description.—UFAC-CS 101 (Fig. 8C) is a small upper molar, quadrangular in occlusal view, strongly recalling the morphology and outline of an M2. It is wide at its base, especially labio-lingually, with a conspicuous labial slope at the base of the StB. The protocone is robust but relatively short. The metaconule is smaller than the protocone. The postprotocrista meets a crest that runs anterolingually to the metaconule, lingually delimitating a small trigon basin, which is separated from the talon basin. There is a wide anterior cingulum, with a conspicuous parastylar cusp sensu Abello and Rubilar-Rogers (2012) present at its labial margin, which possesses a small projection/style at its labial base. The StB is large and high, being the tallest cusp of the molar. The posterior StB crest presents a small cusp-like structure. This crest and the anterior StC+D crest do not form a V-shaped sulcus but run more parallel to the anteroposterior molar axis. The StC+D is small and is placed quite posteriorly. Posterolingual to the StC+D, there is a short wide basin formed by the postmetaconular crest. Crest-like structures run from the stylar cusps towards the protocone and metaconule.
The UFAC-CS 109 specimen (Fig. 8D) is a fragment of a lower molar, with only the talonid preserved. None of the trigonid cuspids are preserved, and the talonid displays two lingual cuspids and one labial. The two lingual ones correspond to an anterior cuspid-like entocristid, which has a blunt tip, and a posterior entoconid, both labio-lingually compressed. The hypoconid occupies a posterior position, with a cristid-like structure running antero-lingually from its tip to the talonid basin. The talonid basin is relatively narrow. Posteriorly to the entoconid, a postentocristid runs almost perpendicular to the anteroposterior molar axis, joining a postcristid towards the molar midline. Basal to them there is a small hypoconulid. The cristid obliqua is slightly oblique and has a sharp occlusal margin.
Remarks.—The presence of lophodont molars is considered a diagnostic characteristic of the family Abderitidae. Besides, the presence of a parastylar cusp on the M2 is also common among abderitids (Abello 2007). The latter and the absence of both paracone and metacone on this tooth, as well as a cristid obliqua with sharp occlusal margin on the m2–m3, and a hypoconid not anteroposteriorly compressed are characteristics present in UFAC-CS 101 and UFAC-CS 109 which are diagnostic of the genus Abderites Ameghino, 1887, according to Abello and Rubilar-Rogers (2012). Three species of Abderites are currently recognized, Abderites meridionalis Ameghino, 1887, Abderites crispus Ameghino, 1902, and Abderites aisenense Abello and Rubilar-Rogers, 2012. The size of UFAC-CS 101 and UFAC-CS 109 is much smaller than the specimens assigned to any known species of Abderites, the closer of them in size being A. crispus (Abello and Rubilar-Rogers 2012). Several of the characteristics that define each of the species of Abderites are located on other molar loci or regions not preserved in the specimens from Acre. Several synonymies were already considered among the referred species, the differences among them being somewhat subtle (Abello and Rubilar-Rogers 2012). The labial slope seen in UFAC-CS 101 is a peculiar characteristic not found in none of the previously known species of Abderites, which have a sub-vertical labial wall on M1–M3. Besides, compared to another abderitid, Pitheculites Ameghino, 1902, the specimens from Acre are slightly larger than Pitheculites minimus Ameghino, 1902, slightly smaller than Pitheculites rothi Marshall, 1990, and Pitheculites ipururensis Stutz, Abello, Marivaux, Boivin, Pujos, Benites-Palomino, Salas-Gismondi, Tejada-Lara, Custódio, Roddaz, Ventura Santos, Ribeiro, and Antoine, 2022, and much smaller than Pitheculites chenche (Dumont and Bown, 1997) (Marshall 1976b; Suárez Gómez, 2019; Stutz et al. 2022). Furthermore, Pitheculites is characterized by a metacone on M2 and a short cristid obliqua with a rounded occlusal margin (Abello 2007).
Therefore, due to their scarcity and fragmentary state, the open taxonomic assignation Abderitidae indet. is considered more conservative for the specimens UFAC-CS 101 and UFAC-CS 109.
Discussion
Miocene Acre diachronous deposits in the upper Juruá and Envira rivers.—The fossil record of Amazonian metatherians remains scarce, especially when compared to the high latitudes of the South American landmass, and the marsupial faunas are quite distinct in their composition between both areas (Goin et al. 2016). Such a disparity does not allow for a precise taxonomic assignment in most cases for newly-found fossil remains, mainly when they consist of isolated tooth specimens. A review of the Middle–Late Miocene metatherian record from Northern South America was recently proposed by Stutz et al. (2022). In Brazilian Amazonia, only didelphids had so far been described, also from Miocene deposits of Acre (unidentified specimens and D. solimoensis; Czaplewski 1996; Cozzuol et al. 2006). Thus, the new records reported here from the Solimões Formation add three didelphid taxa and another order (Paucituberculata), with two new families documented (Palaeothentidae and Abderitidae) to the marsupial paleodiversity of Acre. Although notable, the absence of the orders Sparassodonta and Microbiotheria is likely related primarily to the low number of specimens recovered, despite the field efforts. Nevertheless, these orders are recorded in other coeval localities in northern South America, sometimes in association (e.g., La Venta, Colombia; Suárez Gómez 2019), and sparassodontans are present and diverse in Quebrada Honda (Bolivia; Engelman et al. 2020). Comparatively, with one didelphid, one palaeothentid, and one abderitid but no sparassodontan or microbiothere, the Middle Miocene (Laventan) TAR-31 locality from Peruvian Amazonia (Stutz et al. 2022) yields the taxonomically-closest marsupial community with the three assemblages described here. The presence of Abderitidae indet., found in the PRJ 33 conglomerate level, suggests a Middle Miocene age for this fossil-bearing locality, an age estimate that would agree with that deriving from chinchilloid (e.g., Microscleromys sp.) and erethizontoid (Nuyuyomys sp.) rodent specimens found on the same level (preliminary identifications made by LM and LK, material under analysis).
Traditionally, the vertebrate fauna of the Solimões Formation is often estimated to be Late Miocene in age (Tortonian, Huayquerian SALMA, see Hsiou et al. 2022 for a recent review) and similar in age to the Urumaco fauna (Venezuela) and Ituzaingó Formation (Argentina) (Cozzuol 2006; Carrillo et al. 2015; Kerber et al. 2017). However, some faunal elements from the Solimões Formation present similarities with La Venta, Fitzcarrald and Quebrada Honda faunas, which are considered as Middle Miocene in age (after the native ungulates and rodents; Goillot et al. 2011; Ribeiro et al. 2013; Tejada-Lara et al. 2015). Furthermore, a Middle to Late Miocene interval is recorded through palynostratigraphy in the Solimões Formation (near Cruzeiro do Sul, Acre; Silva-Caminha et al. 2020). Several elements of the upper Juruá River region suggest the presence of still older stratigraphic units, with taxa closely related to those from the Oligocene Santa Rosa, La Cancha (Argentina), and Salla (Bolivia) faunas (Ribeiro et al. 2013; Kerber et al. 2017).
The distinct marsupial assemblages in the two rivers studied here, upper Juruá and Envira (Table 1), indicate that the concerned deposits do not document the same age and/or environmental context. At PRE 06 (Envira River), considered to be (?early) Late Miocene in age (according to the rodent fauna; Fontoura et al. 2022), solely didelphids were recorded, as in classical Acre localities. Late Miocene records of didelphids are much more abundant than Middle Miocene ones. Besides, it is considered as a period of significant diversification and colonization of new areas by this family (Castro et al. 2021). Conversely, only Palaeothentoidea paucituberculatans were found at PRJ 25 and PRJ 33. This superfamily became extinct by the end of the Miocene (Abello et al. 2020). Nevertheless, there is no Late Miocene record of palaeothentoids in Northern South America. However, this group had great abundance and diversity, as well as a broad geographical range in Early–Middle Miocene low-latitude localities (e.g., Quebrada Honda, MD-61, MD-67, CTA-45, TAR-31, and La Venta localities; e.g., Antoine et al. 2013, 2016, 2017; Engelman et al. 2017; Stutz et al. 2022). Based on these recorded occurrences, the extinction of Palaeothentoidea was hypothesized not to have happened as a long, gradual decline but as an abrupt event (Engelman et al. 2017), or perhaps two events instead (Middle–Late Miocene transition at low latitudes; Late Miocene at mid- and high latitudes). Consequently, the presence of palaeothentoids at PRJ 25 and PRJ 33 furthers the hypothesis of the upper Juruá River fossil-bearing localities being older than their Envira River (and classical Acre) counterparts, and most likely Middle Miocene in age. On the other hand, the species-rich didelphid assemblage at PRE 06, with no paucituberculatan specimens, further matches the (?early) Late Miocene age as recently proposed based on rodent biostratigraphy (Fontoura et al. 2022), more in line with the classical Huayquerian Acre faunas. Unfortunately, the lack of a precise geological context for the PRJ 25 specimens and part of the material from PRJ 33 does not help in this context.
Table 1. Marsupial assemblages by the upper Juruá (PRJ 25 and PRJ 33 localities, Middle Miocene) and Envira rivers (PRE 06 locality, [?lower] Upper Miocene), Acre, Brazil.
|
Taxa |
Localities |
||
|
PRJ 25 |
PRJ 33 |
PRE 06 |
|
|
Age |
Middle Miocene |
(?early) Late Miocene |
|
|
Paucituberculata |
|||
|
Palaeothentidae |
|||
|
Palaeothentinae indet. |
× |
× |
|
|
Abderitidae |
|||
|
Abderitidae indet. |
|
× |
|
|
Didelphimorphia |
|||
|
Didelphidae |
|||
|
Marmosini, ?Marmosa sp. |
|
|
× |
|
Didelphis cf. D. solimoensis |
|
|
× |
|
Thylamys? colombianus |
|
|
× |
|
Didelphidae indet. |
|
|
× |
Miocene metatherian diversity, ecology, and paleobiogeography in the Acre region.—Marmosini is a monophyletic tribe of didelphids, which includes Marmosa, Tlacuatzin, and Monodelphis. Marmosa is a complex genus with five subgenera and 25 living species, broadly distributed in Central and South America and occurring in many tropical and subtropical habitats (Rossi et al. 2010; Voss 2022). The phylogenetic pattern of Marmosa was shown to be closely related to the Andes and its orogeny, with a trans-Andean clade of species (mostly in Central America and Western South America) and a cis-Andean group (Patterson et al. 2012). As mentioned above, fossils documenting Marmosa are reported from the La Venta deposits, Middle Miocene of Colombia, with Marmosa laventica, Marmosa sp., and Marmosa sp. 1 (Marshall 1976a; Goin 1997; Suárez Gómez 2019). Besides, Antoine et al. (2013) mentioned the presence of Marmosa (Micoureus) cf. laventica at MD-67, Middle Miocene of Peru. Additional fossil specimens of Marmosa have been reported only from the Chapadmalal Formation (Pliocene) in Argentina (Marshall 1976a). The new possible record of Marmosini, ?Marmosa sp. from PRE 06 highlights the broad presence of close allies of this taxon in the Western Amazonian region during the Miocene.
The genus Didelphis today comprises six species, which occur from North to South America, occupying almost all the tropical and subtropical biomes (except deserts) of the Americas. Nevertheless, its species-level taxonomy is puzzling (Voss and Jansa 2009; Voss 2022). Didelphis was previously recognized in the Upper Miocene of Acre, with Didelphis solimoensis at the Patos locality, on the Acre River (Cozzuol et al. 2006). Moreover, two extinct species of Didelphis were described from the upper Pliocene–lower Pleistocene of Argentina (Goin and Pardiñas 1996; Cozzuol et al. 2006). The record of Didelphis cf. D. solimoensis at PRE 06 possibly adds new specimens of this taxon, which is known solely for its holotype thus far (only lower dentition).
Thylamys is considered a monophyletic genus, with two subgenera and nine species, occurring solely in South America. It is considered a marker of open vegetation environments (Giarla and Jansa 2014). Fossil occurrences of the genus include the extinct T. minutus and T. colombianus from La Venta, Colombia (Goin 1997), and “Thylamys” cf. “T.” colombianus from Peruvian Amazonia (TAR-31; Stutz et al. 2022), both Middle Miocene in age, as well as Late Miocene and Pliocene records in Argentina (Gardner 2007). The record of Thylamys? colombianus at PRE 06 expands the range of this taxon east and southwards and allow for considering its survival in Late Miocene times in Western Amazonia.
Thus, the record of these three didelphid taxa from PRE 06 highlights the diversity and distribution of the Didelphidae during the (?early) Late Miocene in Western Amazonia. As pointed out by Goin (1997) and Suárez Gómez (2019) for the La Venta deposits, this area would have functioned both as a high-diversification rate region, as well as a place with low extinction rates (see also Meseguer et al. 2022). Castro et al. (2021) emphasized the importance of Amazonia for the early diversification of didelphids, with the La Venta and Madre de Dios records (MD-67, Peru), as well as the records in Argentinean Patagonia (Goin et al. 2007) as the first reliable ones of crown representatives of Didelphidae.
Since a more precise taxonomic attribution could not be achieved for Marmosini, ?Marmosa sp. from PRE 06, it is impossible to correlate this taxon to specific paleoenvironmental conditions. Indeed, species of Marmosa inhabit a wide range of habitats. Most species, however, occur in lowland rainforests, and marmosines are generally associated with a more insectivorous-carnivorous niche (Goin 1997; Voss and Jansa 2009). Didelphis species are also distributed in almost all types of habitats, and are considered to have omnivorous and opportunistic behavior, thus, not a helpful indicator for paleoenvironmental considerations either (Cozzuol et al. 2006; Gardner 2007; Voss 2022). On the other hand, species of Thylamys occur today in open environments of subequatorial South America (none in low latitudes), having insectivorous-omnivorous habits (Gardner 2007; Cáceres 2012). As to the fossil Thylamys, Suárez Gómez (2019) suggested that T. colombianus would have had insectivorous/faunivorous habits. Besides, this Thylamys diversity recorded in Amazonia would suggest a much widespread geographical distribution in the past, but the question remains as to whether these Miocene specimens belong to stem Thylamyini or to Thylamys sensu stricto. Interestingly, the ancestral area of Thylamyini was suggested to be Amazonia, with stem thylamyin inhabiting closed-canopy forests and later diversifying and colonizing open habitats east of the Andes, prior to the Late Miocene (Jansa et al. 2014; Castro et al. 2021)
Palaeothentes is a speciose genus that was recovered as monophyletic in the most recent phylogenies considering a comprehensive taxonomic sampling of paucituberculatans, but excluding “Palaeothentes” primus, reassigned to Palaepanorthus (Abello 2007; Engelman et al. 2017). The unidentified palaeothentine(s) from the upper Juruá (PRJ 25 and PRJ 33’) have the closest affinities with Palaeothentes marshalli and Palaeothentes migueli (both being closely related) and with Palaeopanorthus primus (phylogenetically remote) (Engelman et al. 2017; Abello et al. 2020). Palaepanorthus primus is known from the Lower Miocene of Argentinean Patagonia, it had a medium size compared to other palaeothentines, and its dietary habits are not known (Bown and Fleagle 1993; Abello 2007). Palaeothentes is widespread in Miocene localities all over South America, and Northern Amazonia seems to have played a significant role in the evolutionary history of its superfamily, which needs to be more investigated (Stutz et al. 2022 and references therein).
Regarding body size, Palaeothentinae indet. from the upper Juruá (PRJ 25 and PRJ 33’) can be considered to have been small to medium-sized when compared to other palaeothentines, being much smaller than the very large Palaeothentes aratae and Titanothentes simpsoni (>500 g; Abello et al. 2020), and slightly smaller than Palaeothentes serratus (~100 g) and P. relictus (~150 g; Engelman et al. 2017). The absence of m2 in the studied material hinders the dietary reconstruction of the unidentified palaeothentine(s), following Dumont et al. (2000) protocol. Nonetheless, Palaeothentes marshalli and Palaeothentes migueli were unambiguously interpreted as frugivorous (Engelman et al. 2017). Palaeothentids have been related to various feeding habits and different kinds of habitats (mixed vegetational cover and seasonal aridity at Quebrada Honda, Bolivia, Engelman et al. 2017; at Santa Cruz and Pinturas formations, Argentina, markedly seasonal, not dry reconstructed paleoenvironment, Kramarz and Bellosi 2005; Spradley et al. 2019). In Western Amazonia, palaeothentids were abundantly recorded at the Shapaja section in early Oligocene deposits, which do record a drier period after the Eocene–Oligocene transition (Antoine et al. 2021; Stutz et al. 2021; and material under analysis by the authors) as well as in La Venta, which is suggested as having hosted a tropical rainforest by the Middle Miocene (Spradley et al. 2019) and at TAR-31, where the paleoenvironmental conditions are suggested to have been humid, warm, with heterogenous landscape (Stutz et al. 2022).
The record of Abderitidae indet. at PRJ 33 broadens the geographical range of this family and adds a new occurrence of it in South American low latitudes. The only abderitid recognized in Northern South America is Pitheculites, with Pitheculites chenche in La Venta and Pitheculites ipururensis in TAR-31, Peru (Middle Miocene; Suárez Gómez 2019; Stutz et al. 2022), whereas Parabderites and Abderites are classically restricted to the southern part of South America (Argentina and Chile) and spanning the Early–Middle Miocene interval (Colhuehuapian–Colloncuran SALMAs; Abello and Rubilar-Rogers 2012). Because a more precise taxonomic assignment could not be achieved, it is not possible to state whether these specimens represent the occurrence of another abderitid genus in the Amazon region. Abderitids are considered to have been dietary generalists, with a derived dental complexity allowing them to process a wide range of food items (Abello et al. 2020). They thrive in wet and humid paleoenvironments (Ortiz Jaureguizar 2003; Engelman et al. 2017).
Conclusions
The upper Juruá and Envira marsupials described here highlight the role of Western Amazonia regarding Middle and Late Miocene didelphids and palaeothentoids, especially for palaeothentines and abderitids, which were previously unknown in Brazilian Amazonia. Strong affinities are revealed with Middle–early Late Miocene-aged localities of Peruvian Amazonia (MD-67 in Madre de Dios; TAR-31 near Tarapoto; CTA-44 and CTA-43 near Contamana), Colombia (La Venta) and Quebrada Honda (Bolivia). Hence, it reinforces the hypothesis of a unique and homogeneous biogeographical province for land mammals in Western Amazonia over the Middle Miocene times, and probably slightly before and after, possibly in close relation with the Pebas Mega-Wetland System (Stutz et al. 2022). However, the upper Juruá and Envira rivers outcrops need to be better constrained stratigraphically and in terms of geochronological data. Furthermore, much remains to be investigated in terms of fossil marsupials from Northern South America: no pre-Pleistocene fossils are known for the current Amazon region of Ecuador and Colombia (Antoine et al. 2017). As pointed out by Goin et al. (2016), this scarcity of fossil data makes it challenging to understand the South American marsupials’ phylogenetic and paleobiogeographic relationships, not only at tropical latitudes but also for the whole continent.
Acknowledgements
The authors would like to thank all the people who participated in the field campaigns and the staff of the Centro de Microscopia e Microanálise, Universidade Federal do Rio Grande do Sul, Brazil and Laboratório Central de Microscopia e Microanálise, Pontifícia Universidade Católica do Rio Grande do Sul, Brazil for access to SEM facilities. We would also like to thank Paulo Simões Lopes, Jorge J. Cherem, Maurício Graipel (all UFSC), and Victor Pacheco (Museo de Historia Natural of the Universidad Nacional Mayor San Marcos, Lima, Peru) for the access to the scientific collections, and Francisco J. Goin (Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata, Argentina) and one anonymous reviewer for their relevant contribution. This work was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) Ph.D. grants (CNPq 140773/2019-3, CAPES-COFECUB nº 88881.143095/2017-01) (NSS); Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul (FAPERGS, 21/2551-0000781-8) and grants: CNPq 310023/2021-1 and CNPq/MCTI/CONFAP-FAPS PROTAX 22/2020 441626/2020-3 to AMR; CNPq 309414/2019-9 to LK; and CNPq 310948/2021-5 to ASH; Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP 2011/14080-0 and 2019/14153-0 to ASH); Ministério de Ciência e Tecnologia e Inovação (01200.001631/2010-32 and 01200.001631/2010-32); and the French Agence Nationale de la Recherche financed ANR in the framework of the LabEx CEBA (ANR-10-LABX-25-01), through the EMERGENCE project to NSS, LM, POA, FP. This is ISEM publication n°: 2023-186 Sud.
References
Abello, M.A. 2007. Sistemática y Bioestratigrafía de los Paucituberculata (Mammalia, Marsupialia) del Cenozoico de América del Sur. 381 pp. Unpublished Ph.D. Thesis, Universidad Nacional de La Plata, La Plata.
Abello, M.A. and Candela, A.M. 2010. Postcranial skeleton of the Miocene marsupial Palaeothentes (Paucituberculata, Palaeothentidae): paleobiology and phylogeny. Journal of Vertebrate Paleontology 30: 1515–1527. Crossref
Abello, M.A. and Rubilar-Rogers, D. 2012. Revisión del género Abderites Ameghino, 1887 (Marsupialia, Paucituberculata). Ameghiniana 49: 164–184. Crossref
Abello, M.A., De Los Reyes, M., Candela, A.M., Pujos, F., Voglino, D., and Quispe, B.M. 2015. Description of a new species of Sparassocynus (Marsupialia: Didelphoidea: Sparassocynidae) from the late Miocene of Jujuy (Argentina) and taxonomic review of Sparassocynus heterotopicus from the Pliocene of Bolivia. Zootaxa 3937: 147–160. Crossref
Abello, M.A., Toledo, N., and Ortiz-Jaureguizar, E. 2020. Evolution of South American Paucituberculata (Metatheria: Marsupialia): adaptive radiation and climate changes at the Eocene–Oligocene boundary. Historical Biology 22: 476–493. Crossref
Antoine, P.-O., Abello, M.A., Adnet, S., Altamiro Sierra, A.J., Baby, P.; Billet, G., Boivin, M., Calderón, Y., Candela, A., Chabain, J., Corfu, F., Croft, D.A., Ganerød, M., Jaramillo, C., Klaus, S., Marivaux, L., Navarrete, R.E., Orliac, M.J., Parra, F., Pérez, M.E., Pujos, F., Rage, J.C., Ravel, A., Robinet, C., Roddaz, M., Tejada-Lara, J.V., Vélez-Juarbe, J., Wesselingh, F.P., and Salas-Gismondi, R. 2016. A 60-million-year Cenozoic history of western Amazonian ecosystems in Contamana, Eastern Peru. Gondwana Research 31: 30–59. Crossref
Antoine, P.-O., Roddaz, M., Brichau, S., Tejada-Lara, J., Salas-Gismondi, R., Altamirano, A., Louterbach, M., Lambs, L., Otto, T., and Brusset, S. 2013. Middle Miocene vertebrates from the Amazonian Madre de Dios Subandean Zone, Perú. Journal of South American Earth Sciences 42: 91–102. Crossref
Antoine, P.-O., Salas-Gismondi, R., Pujos, F., Ganerød, M., and Marivaux, L. 2017. Western Amazonia as a hotspot of mammalian biodiversity throughout the Cenozoic. Journal of Mammalian Evolution 24: 5–17. Crossref
Antoine, P.-O., Yans, J., Aliaga Castillo, A., Stutz, N., Abello, M.A., Adnet, S., Andriolli Custódio, M., Benites-Palomino, A., Billet, G., Boivin, M., Herrera, F., Jaramillo, C., Mártinez, C., Moreno, F., Navarrete, R.E., Negri, F.R., Parra, F., Pujos, F., Rage, J.-C., Ribeiro, A.M., Robinet, C., Roddaz, M., Tejada-Lara, J.V., Varas-Malca, R., Ventura Santos, R., Salas-Gismondi, R., and Marivaux, L. 2021. Biotic community and landscape changes around the Eocene‒Oligocene transition at Shapaja, Peruvian Amazonia: regional or global drivers? Global and Planetary Change 202: 103512. Crossref
Beck, R.M.D. and Taglioretti, M.L. 2020. A Nearly Complete Juvenile Skull of the Marsupial Sparassocynus derivatus from the Pliocene of Argentina, the Affinities of Sparassocynids, and the Diversification of Opossums (Marsupialia; Didelphimorphia; Didelphidae). Journal of Mammalian Evolution 27: 385–417. Crossref
Beck, R.M.D., Voss, R.S., and Jansa, S.A. 2022. Craniodental morphology and phylogeny of marsupials. Bulletin of the American Museum of Natural History 457: 1–350. Crossref
Bergqvist, L.P., Ribeiro, A.M., and Bocquentin, J. 2002. Primatas, roedores e liptoternas do Mio/Plioceno da Amazônia Sul-Ocidental (Formação Solimões, Bacia do Acre), Brasil. Geologia Colombiana 23: 19–29.
Bissaro Júnior, M.C., Kerber, L., Crowley, J.L., Ribeiro, A.M., Guilardi, R.P., Guilherme, E., Negri, F.P., Souza-Filho, J.P., and Hsiou, A.S. 2019. Detrital zircon U-Pb geochronology constrains the age of Brazilian Neogene deposits from Western Amazonia. Palaeogeography, Palaeoclimatology, Palaeoecology 516: 64–70. Crossref
Boonstra, M., Troelstra, S.R., Lammertsma, E.I., Ramos, M.I.F., Antoine, P.-O., and Hoorn, C. 2015. Marine connections of Amazonia: Evidence from foraminifera and dinoflagellate cysts (early to middle Miocene, Colombia/Peru). Palaeogeography, Palaeoclimatology, Palaeoecology 417: 176–194. Crossref
Bown, T.M. and Fleagle, J.G. 1993. Systematics, biostratigraphy, and dental evolution of the Palaeothentidae, later Oligocene to early–middle Miocene (Deseadan–Santacrucian) caenolestoid marsupials of South America. Journal of Paleontology 67 (Supplement 2): 1–76. Crossref
Cáceres, N.C. (ed) 2012. Os marsupiais do Brasil. 2ª ed. 498 pp. Editora UFMS, Campo Grande.
Carrillo, J.D., Forasiepi, A., Jaramillo, C., and Sánchez-Villagra, M.R. 2015. Neotropical mammal diversity and the Great American Biotic Interchange: spatial and temporal variation in South America’s fossil record. Frontiers in Genetics 5: 451. Crossref
Castro, M.C., Dahur, M.J., and Ferreira, G.S. 2021. Amazonia as the origin and diversification area of Didelphidae (Mammalia: Metatheria), and a review of the fossil record of the clade. Journal of Mammalian Evolution 28: 583–598. Crossref
Cozzuol, M.A. 2006. The Acre vertebrate fauna: Age, diversity, and geography. Journal of South American Earth Sciences 21 (3): 185-203. Crossref
Cozzuol, M.A., Goin, F., de los Reyes, M., and Ranzi, A. 2006. The oldest species of Didelphis (Mammalia, Marsupialia, Didelphidae), from the late Miocene of Amazonia. Journal of Mammalogy 87: 663–667. Crossref
Croft, D.A., Carlini, A.A., Ciancio, M.R., Brandoni, D., Drew, N.E., Engelman, R.K., and Anaya, F. 2016. New mammal faunal data from Cerdas, Bolivia, a middle-latitude Neotropical site that chronicles the end of the Middle Miocene Climatic Optimum in South America. Journal of Vertebrate Paleontology 36 (5): e1163574. Crossref
Cunha, P.R.C. 2007. Bacia do Acre. Boletim de Geociências da Petrobrás 15 (2): 207–215.
Czaplewski, N.J. 1996. Opossums (Didelphidae) and bats (Noctilionidae and Molossidae) from the late Miocene of the Amazon Basin. Journal of Mammalogy 77: 84–94. Crossref
Dumont, E., Strait, S., and Friscia, A. 2000. Abderitid marsupials from the Miocene of Patagonia: an assessment of form, function, and evolution. Journal of Paleontology 74: 1161–1172. Crossref
Engelman, R.K., Anaya, F., and Croft, D.A. 2017. New palaeothentid marsupials (Paucituberculata) from the middle Miocene of Quebrada Honda, Bolivia, and their implications for the palaeocology, decline and extinction of the Palaeothentoidea. Journal of Systematic Palaeontology 15: 787–820. Crossref
Engelman, R.K., Anaya, F., and Croft, D.A. 2020. Australogale leptognathus, gen. et sp. nov., a second species of small sparassodont (Mammalia: Metatheria) from the Middle Miocene locality of Quebrada Honda, Bolivia. Journal of Mammalian Evolution 27: 37–54. Crossref
Fontoura, E., Negri, F.R., Ribeiro, A.M., and Kerber, L. 2022. A new Late Miocene hystricognath rodent assemblage (Caviidae, Dinomyidae, and Neoepiblemidae) from northern Brazil and its biostratigraphic relevance. Historical Biology [available online, https://doi.org/10.1080/08912963.2022.2132858]. Crossref
Forasiepi, A.M., Goin, F., and Martinelli, A.G. 2009. Contribution to the knowledge of the Sparassocynidae (Mammalia, Metatheria, Didelphoidea), with comments on the age of the Aisol Formation (Neogene), Mendoza Province, Argentina. Journal of Vertebrate Paleontology 29: 1252–1263. Crossref
Gardner, A.L. 2007. Mammals of South America. Vol. 1. 669 pp. The University of Chicago Press, Chicago.
Giarla, T.C. and Jansa, S.A. 2014. The role of physical geography and habitat type in shaping the biogeographical history of a recent radiation of Neotropical marsupials (Thylamys: Didelphidae). Journal of Biogeography 41: 1547–1558. Crossref
Goillot, C., Antoine, P.-O., Tejada Lara, J., Pujos, F., and Salas-Gismondi, R. 2011. Middle Miocene Uruguaytheriinae (Mammalia, Astrapotheria) from Peruvian Amazonia and a review of the astrapotheriid fossil record in northern South America. Geodiversitas 33: 331–345. Crossref
Goin, F.J. 1997. New clues for understanding Neogene marsupial radiations. In: R.F. Kay, R.H. Madden, R.L. Cifelli, and J.J. Flynn (eds.), Vertebrate Paleontology in the Neotropics. The Miocene fauna of La Venta, Colombia, 187–206. Smithsonian Institution Press, Washington.
Goin, F.J. and Candela, A.M. 2004. New Palaeogene marsupials from the Amazon Basin of Eastern Peru. In: K.E. Campbell (ed.), The Paleogene Mammalian Fauna of Santa Rosa, Amazonian Peru. Natural History Museum of Los Angeles County, Science Series 40: 15–60.
Goin, F.J. and de los Reyes, M. 2011. Contribución al conocimiento de los representantes extintos de Lutreolina Thomas, 1910 (Mammalia, Marsupialia, Didelphidae). Historia Natural 1(2): 15–25.
Goin, F.J. and Pardiñas, U.F.J. 1996. Revisión de las especies del género Hyperdidelphys Ameghino, 1904 (Mammalia, Marsupialia, Didelphidae). Su significación filogenética, estratigráfica y adaptative en el Neógeno del cono sur sudamericano. Estudios Geológicos 52: 327–359. Crossref
Goin, F.J., Abello, A., Bellosi, E., Kay, R., and Madden, R. 2007. Los Metatheria sudamericanos de comienzos del Neógeno (Mioceno Temprano, Edad-mamífero Colhuehuapense). Parte 1: Introducción, Didelphimorphia y Sparassodonta. Ameghiniana 44: 29–71.
Goin, F.J., Montalvo, C.I., and Visconti, G. 2000. Los marsupiales (Mammalia) del Mioceno Superior de la Formación Cerro Azul (Provincia de La Pampa, Argentina). Estudios Geológicos 56: 101–126. Crossref
Goin, F.J., Woodburne, M.O., Zimicz, A.N., Martin, G.M., and Chornogubsky, L. 2016. A Brief History of South American Metatherians. 237 pp. Springer Earth System Sciences, New York. Crossref
Hoorn, C., Bogotá-A, G.R., Romero-Baez, M., Lammertsma, E.I., Flantua, S.G., Dantas, E. L., Dino, R., do Carmo, D.A., and Chemale Jr, F. 2017. The Amazon at sea: Onset and stages of the Amazon River from a marine record, with special reference to Neogene plant turnover in the drainage basin. Global and Planetary Change 153: 51–65. Crossref
Hoorn, C., Wesselingh, F.P., Hovikoski, J., and Guerrero, J. 2010a. The development of the Amazonian mega-wetland (Miocene; Brazil, Colombia, Peru, Bolivia). In: C. Hoorn and F.P. Wesselingh (eds.), Amazonia, Landscape and Species Evolution: A Look into the Past, 123–142. Blackwell-Wiley, Chichester. Crossref
Hoorn, C., Wesselingh, F.P., Ter Steege, H., Bermudez, M.A., Mora, A., Sevink, J., Sanmartín, I., Sanchez-Meseguer, A., Anderson, C.L., Figueiredo, J.P., Jaramillo, C., Riff, D., Negri, F.R., Hooghiemstra, H., Lundberg, J., Stadler, T., Särkinen, T., and Antonelli, A. 2010b. Amazonia through time: Andean uplift, climate change, landscape evolution, and biodiversity. Science 330: 927–931. Crossref
Hsiou, A.S., Bissaro-Júnior, M.C., Muniz, F.P., Negri, F.R., Ribeiro, A.M., and Kerber, L. 2022. Bacia do Acre: Evidências das paisagens da proto-Amazônia há 10 milhões de anos, com base em vertebrados fósseis da Formação Solimões. In: L. Corecco (ed.), Paleontologia do Brasil: Paleoecologia e Paleoambientes, 29–64. Interciência, Rio de Janeiro.
Jansa, S.A., Barker, F.K., and Voss, R.S. 2014. The early diversification history of didelphid marsupials: a window into South America’s “splendid isolation”. Evolution 68: 684–695. Crossref
Kerber, L., Bissaro Júnior, M.C., Negri, F.R., Souza-Filho, J.P., Guilherme, E., and Hsiou, A.S. 2018. A new rodent (Caviomorpha: Dinomyidae) from the upper Miocene of southwestern Brazilian Amazonia. Historical Biology 30: 985–993. Crossref
Kerber, L., Candela, A.M., Ferreira, J.D., Pretto, F.A., Bubadué, J., and Negri, F.R. 2022. Postcranial Morphology of the extinct rodent Neoepiblema (Rodentia: Chinchilloidea): insights into the paleobiology of neoepiblemids. Journal of Mammalian Evolution 29: 207–235. Crossref
Kerber, L., Ferreira, J.D., and Negri, F.R. 2019a. A reassessment of the cranial morphology of Neoepiblema acreensis (Rodentia: Chinchilloidea), a Miocene rodent from South America. Journal of Morphology 280: 1821–1838. Crossref
Kerber, L., Negri, F.R., and Sanfelice, D. 2019b. Morphology of cheek teeth and dental replacement in the extinct rodent Neoepiblema Ameghino, 1889 (Caviomorpha, Chinchilloidea, Neoepiblemidae). Journal of Vertebrate Paleontology 38 (6): e1549061. Crossref
Kerber, L., Negri, F.R., Ribeiro, A.M., Nasif, N., Souza-Filho, J., and Ferigolo, J. 2017. Tropical fossil caviomorph rodents from the Southwestern Brazilian Amazonia in the context of the South American faunas: systematics, biochronology, and paleobiogeography. Journal of Mammalian Evolution 24: 57–70. Crossref
Kerber, L., Negri, F.R., Ribeiro, A.M., Vucetich, M.G., and Souza-Filho, J.P. 2016. Late Miocene potamarchine rodents from southwestern Amazonia, Brazil—with description of new taxa. Acta Palaeontologica Polonica 61: 191–203.
Kloster, A., Gnaedinger, S., and Adami-Rodrigues, K. 2017. Nuevos registros de maderas del Mioceno de la Formación Solimões, cuenca de Acre, Amazonia, Brasil. Revista Brasileira de Paleontologia 20: 383–404. Crossref
Kramarz, A.G. and Bellosi, E.S. 2005. Hystricognath rodents from the Pinturas Formation, Early–Middle Miocene of Patagonia, biostratigraphic and paleoenvironmental implications. Journal of South American Earth Sciences 18: 199–212. Crossref
Lacerda, M.B.S., Romano, P.S.R., Bandeira, K.L.N., and Souza, L.G. 2021. Georeferencing fossiliferous localities from Solimões and Acre Basins (Brazil)—what we know so far about Solimões Formation and future perspectives. Anais da Academia Brasileira de Ciências 93 (Supplement 2): e20201642. Crossref
Linares, O.J. 2004. Bioestratigrafia de la fauna de mamiferos de las Formaciones Socorro, Urumaco y Codore (Mioceno Medio-Plioceno Temprano) de la region de Urumaco, Falcon, Venezuela. Paleobiologia Neotropical 1: 1–26.
Marivaux, L., Aguirre-Diaz, W., Benites-Palomino, A., Billet, G., Boivin, M., Pujos, F., Salas-Gismondi, R., Tejada-Lara, J.V., Varas-Malca, R.M., and Antoine, P.-O. 2020. New record of Neosaimiri (Cebidae, Platyrrhini) from the late Middle Miocene of Peruvian Amazonia. Journal of Human Evolution 146: 102835. Crossref
Marivaux, L., Negri, F.R., Antoine, P.-O., Stutz, N.S., Condamine, F.L., Kerber, L., Pujos, F., Ventura Santos, R., Alvim, A.M.V., Hsiou, A.S., Bissaro Jr., M.C., Adami-Rodrigues, K., and Ribeiro A.M. 2023. An eosimiid primate of South Asian affinities in the Paleogene of Western Amazonia and the origin of New World monkeys. Proceedings of the National Academy of Sciences of the United States of America 120 (28): e2301338120. Crossref
Marshall, L.G. 1976a. New Didelphine marsupials from the La Venta fauna (Miocene) of Colombia, South America. Journal of Paleontology 50: 402–418.
Marshall, L.G. 1976b. Revision of the South American subfamily Abderitinae (Mammalia, Caenolestidae). Publicaciones del Museo Municipal de Ciencias Naturales de Mar del Plata Lorenzo Scaglia 2 (3): 57–90.
Marshall, L.G. 1987. Systematics of Itaboraian (middle Paleocene) age opossum-like marsupials from the limestone quarry at São José de Itaboraí, Brazil. In: M. Archer (ed.), Possums and Opossums: Studies in Evolution, 91–160. Surrey Beatty and Sons and the Royal Zoological Society of New South Wales, Chipping Norton.
Meseguer, A., Michel A. J., Fabre, P.-H., Pérez-Escobar, O.A., Chomicki, G., Riina, R., Antonelli, A., Antoine, P.-O., Delsuc, F., and Condamine, F.L. 2022. Diversification dynamics in the Neotropics through time, clades, and biogeographic regions. Elife 11: e74503. Crossref
Negri, F.R. 2004. Os Tardigrada (Mammalia, Xenarthra) do Neógeno, Amazônia Sul-Ocidental, Brasil. 156 pp. Unpublished. Ph.D. Thesis, Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre.
Negri, F.R., Bocquentin Villanueva, J., Ferigolo, J., and Antoine, P.-O. 2010. 16. A review of Tertiary mammal faunas and birds from western Amazonia. In: C. Hoorn and F.P. Wesselingh (eds.), Amazonia, Landscape and Species Evolution: A Look into the Past, 245–258. Blackwell-Wiley, Chichester. Crossref
Ortiz-Jaureguizar, E. 2003. Relaciones de similitud, paleoecología y extinción de los Abderitidae (Marsupialia, Paucituberculata). Coloquios de Paleontología, Extra Volume 1: 475–498.
Patterson, B. and Costa, L. 2012. Bones, Clones, and Biomes: The History and Geography of Recent Neotropical Mammals. 432 pp. The University of Chicago Press, Chicago. Crossref
Patterson, B.D., Solari, S., and Velazco, P.M. 2012. The Role of the Andes in the Diversification and Biogeography of Neotropical Mammals. In: B.D. Patterson and L. Costa (eds.), Bones, Clones, and Biomes: The History and Geography of Recent Neotropical Mammals, 351–378. The University of Chicago Press, Chicago. Crossref
Ribeiro, A.M., Madden, R.H., Negri, F.R., Kerber, L., Hsiou, A.S., and Rodrigues, K.A. 2013. Mamíferos fósiles y biocronología en el suroeste de la Amazonia, Brasil. In: D. Brandoni and J.I. Noriega (eds.), El Neógeno de la Mesopotamia Argentina, 207–221. Asociación Paleontológica Argentina, Buenos Aires.
Rossi, R.V., Voss, R.S., and Lunde, D.P. 2010. A revision of the didelphid marsupial genus Marmosa. Part 1. The species in Tate’s ‘mexicana’ and ‘mitis’ sections and other closely related forms. Bulletin of the American Museum of Natural History 334: 1–83. Crossref
Sánchez-Villagra, M.R., Aguilera, O.A., Sánchez, R., and Carlini, A.A. 2010. The fossil vertebrate record of Venezuela of the last 65 million years. In: M.R. Sánchez-Villagra, O.A. Aguilera, and A.A. Carlini (eds.), Urumaco & Venezuelan Paleontology, 19–51. Indiana University Press, Bloomington.
Silva-Caminha, S.A.F., D’Apolito, C., Jaramillo, C., Espinosa, B.S., and Rueda, M. 2020. Palynostratigraphy of the Ramon and Solimões formations in the Acre Basin, Brazil. Journal of South American Earth Sciences 103: 102720. Crossref
Simpson, G.G. 1970. The Argyrolagidae, extinct South American marsupials. Bulletin of the Museum of Comparative Zoology 139 (1): 1–86.
Spradley, J.P., Glazer, B.J., and Kay, R.F. 2019. Mammalian faunas, ecological indices, and machine-learning regression for the purpose of paleoenvironment reconstruction in the Miocene of South America. Palaeogeography, Palaeoclimatology, Palaeoecology 518: 155–171. Crossref
Stutz, N.S., Abello, M.A., Marivaux, L., Boivin, M., Custódio, M.A., Benites-Palomino, A.M., Salas-Gismondi, R., Pujos, F., Tejada-Lara, J.V., Ribeiro, A.M., and Antoine, P.-O. 2021. Neotropical metatherian diversity around the Eocene–Oligocene transition: the Shapaja section, Peruvian Amazonia. In: J.C. Fernicola, L. Balarino, L.E. Cruz, and D.G. Lazo (eds.), 14 CAPA, Symposium Mamíferos del Paleógeno de América del Sur, Libro de Resúmenes, 123. Asociación Paleontológica Argentina, Buenos Aires.
The meeting (14 CAPA) was organized by the following committee: Fernicola, J.C., Balarino L., Cruz, L.E., and Lazo D.G.
Stutz, N.S., Abello, M.A., Marivaux, L., Boivin, M., Pujos, F., Benites-Palomino, A.M., Salas-Gismondi, R., Tejada-Lara, J.V., Custódio, M.A., Roddaz, M., Santos, R.V., Ribeiro, A.M., and Antoine, P.-O. 2022. Late middle Miocene Metatheria (Mammalia: Didelphimorphia and Paucituberculata) from Juan Guerra, San Martín Department, Peruvian Amazonia. Journal of South American Earth Sciences 118: 103902. Crossref
Suárez, C., Forasiepi, A., Goin, F.J., and Jaramillo, C. 2016. Insights into the Neotropics prior to the Great American Biotic Interchange: new evidence of mammalian predators from the Miocene of Northern Colombia. Jounal of Vertebrate Paleontology 36 (1): e1029581. Crossref
Suárez Gómez, S.C. 2019. Estudios taxonómicos y paleobiológicos sobre los Metatheria (Mammalia) del Mioceno medio de La Venta, Colombia. 292 pp. Unpublished Ph.D. Thesis, Universidad Nacional de la Plata, La Plata.
Tejada-Lara, J., Salas-Gismondi, R., Pujos, F., Baby, P., Benammi, M., Brusset, S., Franceschi, D., Espurt, N., Urbina, M., and Antoine, P.-O. 2015. Life in Proto-Amazonia: Middle Miocene mammals from the Fitzcarrald Arch (Peruvian Amazonia). Palaeontology 58: 341–378. Crossref
Voss, R.S. 2022. An annotated checklist of recent opossums (Mammalia: Didelphidae). Bulletin of the American Museum of Natural History 455: 1–76. Crossref
Voss, R.S. and Jansa, S.A. 2009. Phylogenetic relationships and classification of didelphid marsupials, an extant radiation of New World metatherian mammals. Bulletin of the American Museum of Natural History 322: 1–177. Crossref
Acta Palaeontol. Pol. 68 (3): 457–475, 2023
https://doi.org/10.4202/app.01057.2023