
A new Early Devonian antiarch placoderm from Belarus, and the phylogeny of Asterolepidoidei
DMITRY P. PLAX and ERVINS LUKŠEVIČS
Plax, D.P. and Lukševičs, E. 2023. A new Early Devonian antiarch placoderm from Belarus, and the phylogeny of Asterolepidoidei. Acta Palaeontologica Polonica 68 (3): 513–527.
A new asterolepidoid antiarch, Sherbonaspis talimaae sp. nov., is described based on the disarticulated skeletal elements from several boreholes in Belarus, from the Lepel Beds of the Vitebsk Formation, which has been assigned to an Early Devonian, late Emsian age. New information is provided on the structure of the paranuchal plate previously unknown in Sherbonaspis. Data on the remains of the other fossil vertebrates, and the characteristics of the fossil-bearing rocks are provided. These data add information on the taxonomic composition of the late Emsian–early Eifelian fish fauna from the western part of the East European Platform. The new cladistic analysis of the Euantiarcha provides results more reliable in comparison with the analysis of all antiarchs together, and supports traditional subdivision of the group into Bothriolepidoidei and Asterolepidoidei. However, further analysis demonstrates unresolved polytomy close to the base of the Asterolepidoidei, and supports the hypothesis that the family Pterichthyodidae is polyphyletic. Until the phylogenetic relationships of Asterolepidoidei are more clearly resolved, Sherbonaspis is attributed to Pterichthyodidae. Comparison of the sections of the Lepel Beds of Belarus with the age-equivalent deposits in Estonia, Lithuania, and Latvia supports a late Emsian or possibly early Eifelian age for the Lepel Beds.
Key words: Placodermi, Asterolepidoidei, phylogeny, Devonian, Belarus.
Dmitry P. Plax [agnatha@mail.ru; ORCID: https://orcid.org/0000-0003-0990-2353 ], Belarusian National Technical University, Nezavisimosti Avenue, 65, 220013, Minsk, Belarus.
Ervin Lukševičs [ervins.luksevics@lu.lv; ORCID: https://orcid.org/ 0000-0002-5362-7953 ] (corresponding author), Department of Geology, University of Latvia, Rainis Blvd 19, Riga LV-1586, Latvia.
Received 5 April 2023, accepted 25 May 2023, available online 4 September 2023.
Copyright © 2023 D.P. Plax and E. Lukševičs. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The first findings of antiarchs from probable Lower Devonian deposits on the territory of Belarus were reported almost half a century ago (Bessonova et al. 1972; Lyarskaya 1978; Golubtsov et al. 1983). Published data show that unidentifiable individual scales and fragmentary plates of Antiarcha gen. et sp. indet. have been found in the Obol and Lepel Beds of the Vitebsk Regional Stage, probably corresponding to the upper Emsian, together with stromatolites, plant remains, miospores, conchostracans, ostracods, lingulids, various skeletal elements of arthrodires, scales and fin spines of acanthodians, and teeth and scales of sarcopterygians and actinopterygians. However, for over 20 years there were no new published reports on antiarch remains in the upper Emsian deposits on the territory of Belarus.
Dmitry P. Plax started a new study of the upper Emsian–lower Eifelian deposits in Belarus in 2002. Since then, a large number of wells that intersect these deposits have been viewed and studied. In the cores of some of these wells, he managed to find a few remains of antiarchs, represented by separate small fragments of plates from the proximal segment of the pectoral fin, small fragments of articular processes, and small fragmentary unidentifiable plates and scales. All this small, fragile, highly fragmentary and difficult to determine skeletal material was identified as Antiarcha indet. in a series of publications (Plaksa 2007, 2008; Plax 2008, 2015a, b, 2016, 2018). However, after many years of study DPP was at last lucky to find several well-preserved plates of a new representative of antiarchs, for which we provide a detailed description, and also discuss the phylogeny of euantiarchs, and biogeographic and stratigraphic implications of this new antiarch.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:B5470082-7D5C-461A-BD9F-9A02C2ADFA4D.
Institutional abbreviations.—BNTU, Belarusian National Technical University, Minsk, Belarus.
Other abbreviations.—adc; anterior dorsal corner of AVL plate; ADL, anterior dorsolateral plate; AVL, anterior ventrolateral plate; B/L, breadth/length index; c.al, anterior lateral corner of AVL; cf.ADL, area overlapping ADL plate; cf.MV, area overlapping MV plate; cf.MxL, area overlapping MxL plate; cf.PVL, area overlapping PVL plate; cf.Sm, notch for the semilunar plate; cit1, crista transversalis interna anterior; cr.pm, paramarginal crest; dc, dorsal corner of PVL plate; f.ax, axillary foramen; f.ax1, inner axillary foramen; ifc1, infraorbital sensory line canal; L/B, length/breadth index; mpg, middle pit-line groove; MV, median ventral plate; MxL, mixilateral plate; oa.AVL, area overlapped by AVL plate; pdc, posterodorsal corner of AVL plate; Pn, paranuchal plate; pr.br, processus brachialis; ptc, cephalic division of the main lateral line; PVL, posterior ventrolateral plate; ri, ridge on the visceral surface of AVL plate; Sm; semilunar plate; vlr, ventrolateral ridge of trunk armour; RS, Regional Stage.
Material and methods
The holotype of Sherbonaspis talimaae sp. nov. was collected from the grey siltstone of the Lepel Beds reached in the Osipovichi 6 borehole in the central part of Belarus. The other skeletal remains are represented by isolated complete or almost complete armour plates sampled from three wells: Osipovichi 6, Korma 1 and Smol’ki 6п (Fig. 1, Table 1). The specimens were extracted by dissolving rock samples with 9% acetic acid. The preservation of skeletal material is good; it bears no signs of corrosion or abrasion.
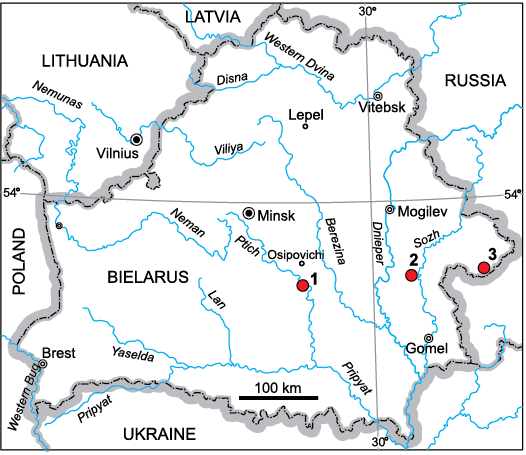
Fig. 1. Map of the location of the borehole sections where the skeletal elements of the antiarch fish Sherbonaspis talimaae sp. nov. were found. Boreholes: 1, Osipovichi 6; 2, Korma 1; 3, Smol’ki 6п.
The plates were measured using a ruler and Vernier caliper. The skeletal elements were examined under an MBS-1 microscope and photographed with a PowerShot SX130 IS and Sony A58 with an Industar-50 lens. The photographs were processed using Adobe Photoshop CC 2019 and the drawings were constructed using CorelDraw Graphics Suite 2019. Cladistic analyses were performed using the heuristic algorithms of PAUP version 4a (169) (Swofford 2003). The resulting most parsimonious trees were resampled over 100 replicates to generate bootstrap supports for each estimated node. The terminology for morphological characters defined and used by Stensiӧ (1948), and modified by Miles (1968) and Young (1988), is adopted here. All specimens are stored at the Belarusian National Technical University in the Department of Mining Works, Paleontological Collection, Minsk.
Table 1. The list of specimens of Sherbonaspis talimaae sp. nov. BNTU 44/2-1 and 44/2-2 were found in the same piece of core. Abbreviations: AVL, anterior ventrolateral plate; Pn, paranuchal plate; PVL, posterior ventrolateral plate.
|
Specimen No. |
Plate |
Borehole |
Depth [m] |
|
BNTU 44/2-1 |
AVL |
Osipovichi 6 |
113.8 |
|
BNTU 44/2-2 |
PVL |
||
|
BNTU 44.1-13a |
Pn |
Osipovichi 6 |
118.8 |
|
BNTU 121/20-1 |
AVL |
Korma-1 |
317.5 |
|
BNTU 158/1-1 |
PVL |
Smol’ki 6п |
474.0 |
Geological settings
All of the described material comes from the upper part of the Vitebsk Formation, corresponding to the Lepel Beds, Vitebsk Regional Stage. These deposits are quite widespread in Belarus. According to the current Devonian stratigraphic chart of Belarus (Obukhovskaya et al. 2010) the Vitebsk RS completely corresponds to the upper Emsian, Lower Devonian (Fig. 2). However, earlier the same authors placed the Emsian/Eifelian boundary approximately in the middle of the Lepel Beds, close to the boundary between the lower and upper members (Kruchek et al. 2001: fig. 5.22). In the territory of Belarus rocks of the Lepel Beds concordantly overlap deposits of the Obol Beds of the Vitebsk RS and are covered everywhere with deposits of the Adrov RS corresponding to the Middle Devonian Eifelian Stage. Deposits of the Lepel Beds correspond to the upper part of the Rhabdosporites mirus–Gneudnaspora divellomedium Miospore Zone, while in terms of conodonts they roughly correspond to the upper part of the Polygnathus costatus patulus Zone of the previous version of conodont zonation (Obukhovskaya et al. 2010). However, as conodonts have not been found in the Vitebsk Formation, correlation with the conodont zonation is uncertain. In the Baltic States, the age analogue of the deposits of Lepel Beds is approximately the upper part of the Rēzekne Formation (Lyarskaya 1978; Lyarskaya and Kleesment 1981; Obukhovskaya et al. 2010). The upper part of the Rēzekne Formation in the modern stratigraphic chart of Latvia corresponds to the lowermost Eifelian (Lukševičs et al. 2010; Lukševičs and Stinkulis 2018), but to the uppermost Emsian in the stratigraphic chart of Estonia (Mark-Kurik and Põldvere 2012). In Central Russia, the upper part of the Novobasovo Beds of the Ryazhsk RS may correspond to the Lepel Beds (Rodionova et al. 1995; Valiukevičius and Kruchek 2000; Vorotnikova et al. 2011). As the exact correlation of the Lepel Beds with the global chronostratigraphic scale remains controversial, broader aspects of the Emsian/Eifelian boundary in the Baltic-Belarusian area are considered in the Discussion section. Here we give more details of the locations of the described skeletal elements, the lithological characteristics of the host rocks and a list of fish taxa found together with Sherbonaspis.
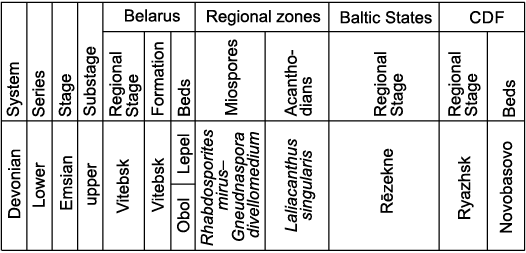
Fig. 2. Stratigraphy of the upper Emsian deposits of Belarus and their correlation with the synchronous deposits from the adjacent territories (according to Obukhovskaya et al. 2010). CDF, Central Devonian Field.
In Osipovichi 6 borehole, drilled within the Bobruisk buried ledge, at a depth of 118.8 m, a right Pn plate of Sherbonaspis talimaae sp. nov. BNTU 44.1-13a was found in a light grey, with greenish tint, dense, platy marl, with interlayers of sandy material. Small indeterminate plate fragments, fused left and right preorbital plates, a right paranuchal plate, incomplete left anterior dorsal lateral plate of Actinolepis zaikai Plax and Newman, 2020, isolated scales of Laliacanthus singularis Karatajūte-Talimaa in Valiukevičius and Karatajūte-Talimaa, 1986, small fragments of fin spines of Acanthodii gen. indet., scales of Porolepis sp., Osteolepididae gen. indet., and teeth of Onychodontidae gen. indet. (Plax 2015b; Plax and Newman 2020) were found together with Sherbonaspis talimaae sp. nov.. In the same borehole, at a depth of 113.8 m, two plates of Sherbonaspis talimaae sp. nov., fragment of the AVL plate BNTU 44/2-1 and left PVL plate BNTU 44/2-2, were found in a light yellowish grey to greenish grey, fine-grained, clayey, well-cemented, dense, platy siltstone with a small amount of carbonate. A small fragmentary plate of Actinolepis zaikai Plax and Newman, 2020, a spinal plate of undetermined arthrodiran fish, a few scales of ?Acanthodes sp., small fragments of fin spines of Acanthodii gen. indet., teeth of Onychodontidae gen. indet. and Sarcopterygii indet., scales of Osteolepididae gen. indet., scales of Cheirolepis sp. and one incomplete impression of caudal part of Actinopterygii indet. (Plax 2015b; Plax and Newman 2020) were collected together with plates of Sherbonaspis talimaae sp. nov. Along with vertebrates, the presence of miospores was established in these rocks. Miospores from this and all the other studied boreholes were studied by Tatyana Obukhovskaya (personal communication); the analysis suggested the correspondence of the spore assemblage with the upper part of the Rhabdosporites mirus–Gneudnaspora divellomedium Miospore Zone.
In the borehole Korma 1, drilled in the territory of the Zhlobin saddle, Gomel’ Region, a left AVL plate of Sherbonaspis talimaae sp. nov. BNTU 121/20-1 was obtained in a greenish grey, dense, platy, silty silt-like clay. Together with it, in this rock and at the same depth, very small indeterminate fragments of Placodermi indet. plates, disarticulated scales of Cheiracanthus sp., Cheiracanthus brevicostatus Gross, 1973, numerous tooth fragments of Onychodontiformes indet., Sarcopterygii indet., and fragmentary scales of Porolepiformes indet. were discovered. In addition to the remains of the fish listed above, miospores were also found here.
Borehole Smol’ki 6п is situated in the territory of the Surazh buried protrusion of the Voronezh Anteclise. The left PVL plate BNTU 158/1-1 has been found in a light grey, fine-grained, well cemented platy quartz sandstone with small admixture of clay. Two small fragmentary plates of psammosteid heterostracan, several scales of ?Acanthodes sp., fragmentary scale of Onychodontiformes indet., some small teeth of Onychodontiformes indet. and Sarcopterygii indet. were found in the same sandstone at the same depth. Miospores were found along with the vertebrates.
Systematic palaeontology
Euantiarcha Janvier and Pan, 1982
Asterolepidoidei Miles, 1968
Pterichthyodidae Stensiӧ, 1948
Genus Sherbonaspis Young and Gorter, 1981
Type species: Sherbonaspis hillsi Young and Gorter, 1981, Goodradigbee valley near Wee Jasper, New South Wales, Australia; ?Emsian/Eifelian, Lower/Middle Devonian.
Sherbonaspis talimaae sp. nov.
Figs. 3–6.
2007 Antiarcha gen. nov.; Plaksa 2007: 7.
2008a Antiarcha gen. nov.; Plaksa 2008a: 84.
2008b Antiarcha gen. nov.; Plaksa 2008b: 136.
2008 Antiarcha gen. nov. Plax; Plax 2008: 70, table 1, p. 76.
2012b Antiarcha gen. nov. Plax; Kruchek et al. 2012b: 95.
2012a Antiarcha gen. nov. Plax; Kruchek et al. 2012a: 100.
2012a Actinolepididae gen. indet.; Kruchek et al. 2012a: 100.
2015a Actinolepididae gen. indet.; Plax 2015a: 171, pl. 1: 5.
2015a Antiarcha gen. nov. Plax; Plax 2015a: 171, pl. 1: 6.
2015b Actinolepididae gen. indet.; Plax 2015b: 21, table (pars), pl. 1: 10.
2015b Antiarcha gen. nov. Plax; Plax 2015b: 21, table.
2018 Antiarcha gen. nov. Plax; Plax 2018: 409.
ZooBank LCID: urn:lsid:zoobank.org:actD051501D-F1DA-4EE9-B 648-537AB4ED18B0
Etymology: In honour of Valentina Karatajūtė-Talimaa (Vilnius, Lithuania), who recently passed away (1930–2022), in appreciation of her works on antiarchs and thelodonts.
Holotype: BNTU 44/2-2, left PVL plate.
Type locality: Borehole Osipovichi 6, 113.8 m deep, close to Osipovichi town, Mogilev Region, Belarus.
Type horizon: Lepel Beds, Vitebsk Regional Stage, probably upper Emsian, Lower Devonian.
Material.—Right Pn plate (BNTU 44/1-13a), left AVL plate (BNTU 121/20-1), fragmentary right AVL plate (BNTU 44/2-1), left PVL plates (BNTU 44/2-2, BNTU 158/1-1). All from type horizon; for localities see Table 1.
Diagnosis.—An asterolepidoid antiarch of small size, with length of the ventral wall of the trunk armour of about 75 mm, and with relatively high trunk armour. Paranuchal plate small, asymmetrical, slightly longer than it is wide. Ventral lateral ridge is well developed along the whole length of the trunk armour. The ventral wall of the trunk armour is flat. The contact face for mixilateral plate on the anterior ventral lateral plate is short. Lateral margin of the ventral lamina of posterior ventral lateral plate almost straight. Length/width index of lateral lamina of posterior ventral lateral plate is about 2.3. Ornamentation generally consists of small tubercles sometimes fused into tuberculate ridges; tubercles tend to form rows perpendicular or parallel to the margins of the plates. The anterior ventral lateral plates bear a fine-meshed network of the shallow pits in the anterior part of the ventral lamina, and small tubercles with sharp tips along the margins in the posterior portion of the plate.
Description.—Only the Pn plate is known from the skull roof of Sherbonaspis talimaae sp. nov. This plate has not been described either in Sherbonaspis hillsi (Young and Gorter 1981) or in Sherbonaspis andreannae (Panteleyev 1993). The right Pn plate BNTU 44/1-13a (Fig. 3C) is subsquare in shape, with the maximum width in its posterior part. It is longer than broad, like in Pterichthyodes milleri (Miller, 1841) (Hemmings 1978: 16), with the B/L index of 0.88, in a sharp contrast to species of Asterolepis (e.g., Lyarskaya 1981) or Walterilepis speciosa (Gross, 1933) (Lukševičs 2021) with the B/L index of 1.12–1.34. The posterior margin of the bone is straight, with a narrow (short) and weakly developed smooth obtected nuchal area. The lateral segment of the bone is narrow, only 0.16 times as wide as the medial segment, in contrast to Walterilepis speciosa with the index of 0.35–0.49. The mpg is well seen and long almost reaching the mesial margin (Fig. 4B) thus differing well from Byssacanthus dilatatus (Eichwald, 1844) (Karatajūte-Talimaa 1960) and Walterilepis speciosa. The infraorbital sensory line canal on the head shield (ifc1) is weakly defined, in contrast to the cephalic division of the main lateral line (ptc) that is well discernible in the posterior third of the lateral margin. The lateral segment is narrowest posteriorly slightly increasing in width anteriorly; this shape significantly differs from that in species of Byssacanthus, Pterichthyodes, and Walterilepis where the lateral segment is narrowest anteriorly (e.g., Lukševičs 2021: fig. 3G1). The internal surface shows a well-defined paramarginal crest (cr.pm) dividing the surface into two regions approximately equal in width. The posterolateral corner of the shallow oticooccipital depression is rounded. The external ornamentation consists of low small tubercles in places fused into radially arranged tuberculate ridges radiating from the posterior lateral angle of the plate.

Fig. 3. Fossil remains of the antiarch placoderm Sherbonaspis talimaae sp. nov.; upper Emsian of Belarus, borehole Korma-1 (A), borehole Osipovichi 6 (B–D), borehole Smol’ki 6п (E). A. BNTU 121/20-1, anterior ventral lateral plate in external (A1) and internal (A2) views. B. BNTU 44/2-1, fragmentary anterior ventral lateral plate in external view. C. BNTU 44/1-13a, paranuchal plate in external view. D. BNTU 44/2-2 (holotype), left posterior ventral lateral plate in external view. E. BNTU 158/1-1, left posterior ventral lateral plate in lateral view.
Thorax: The left AVL plate BNTU 121/20-1 is partially preserved bone showing only the ventral lamina; almost all of the lateral lamina and processus brachialis are missing (Fig. 3A); this specimen is very thin, extremely fragile, and relatively small (only 41 mm in length). The right AVL plate BNTU 44/2-1 is even less complete, comprising approximately half of the posterior part of both ventral and lateral laminae (Fig. 3B). Length of the ventral lamina in BNTU 44/2-1 is estimated at about 20 mm. Almost all edges of the ventral lamina of the AVL plate in BNTU 121/20-1 are well preserved, except the slightly damaged middle portion of the mesial edge. In contrast, most edges of the lateral lamina of the AVL in BNTU 121/20-1 are damaged, except the ventral lateral ridge and a narrow strip of bone along the ridge. The angle between the ventral and lateral laminae cannot be measured because the lateral lamina is almost absent in BNTU 121/20-1, and because both ventral and lateral laminae in BNTU 44/2-1 form too obtuse angle due to deformation of the specimen. In its squarish outline the ventral lamina resembles that of Byssacanthus dilatatus (Eichwald, 1844), Sherbonaspis hillsi Young and Gorter, 1981, Sherbonaspis andreannae Panteleyev, 1993, or Stegolepis jugata Malinovskaya, 1973 (Karatajūte-Talimaa 1960; Malinovskaya 1973; Young and Gorter 1981; Panteleyev 1993), and differs from species of Asterolepis (e.g., Lyarskaya 1981). It is moderately elongated with a B/L index of about 0.5, resembling that in Sherbonaspis hillsi, being larger than the index in Stegolepis jugata or Walterilepis speciosa (Malinovskaya 1973; Young and Gorter 1981; Lukševičs 2021). The ventral lamina is almost flat, as in Byssacanthus dilatatus, Stegolepis jugata, Sherbonaspis hillsi, and Walterilepis speciosa (Karatajūte-Talimaa 1960; Malinovskaya 1973; Young and Gorter 1981; Lukševičs 2021), and in contrast to Gerdalepis rhenana (Beyrich, 1877) and Gerdalepis jesseni Friman, 1982 (Gross 1941; Friman 1982) or Wurungulepis Young, 1990 (Young 1990), where it is convex. The subcephalic division is relatively short and comprises about 21% of the length of the ventral lamina similarly as in Sherbonaspis hillsi (Young and Gorter 1981); the subcephalic division is rather short also in Stegolepis jugata (Malinovskaya 1973: 75). This division is only slightly shorter than the subcephalic division in Walterilepis speciosa (Lukševičs 2021) and significantly shorter than that in Pterichthyodes milleri (Miller, 1841 ex Agassiz MS) (Hemmings 1978: 30). The anterior margin is well preserved in BNTU 121/20-1, which shows a distinct anterolateral corner (c.al), as is the notch for the semilunar plate (Fig. 4A2: cf.Sm). The anterior margin of the AVL is rounded; it is formed of two almost equal parts: the anterior lateral part is almost straight forming about 135° wide angle with a rounded anterior mesial part of this margin. The right AVL overlies the left one similarly as in Pterichthyodes milleri, Sherbonaspis hillsi, and Stegolepis jugata, but in contrast to Walterilepis (Malinovskaya 1973; Hemmings 1978; Young and Gorter 1981; Lukševičs 2021). The lateral margin is almost straight, it is only slightly convex in the anterior part. It is easy to discern that the mesial margin is subdivided into three portions for contact with the semilunar plate anteriorly (cf.Sm), with the opposite AVL plate in the middle (oa.AVL), and with the median ventral plate posteriorly (cf.MV). The shape of the notch for the semilunar plate (cf.Sm; Fig. 4A1) suggests a narrow trapezoidal shape for this plate, and not a quadrilateral shape as in Stegolepis and Pterichthyodes (e.g., Malinovskaya 1973; Hemmings 1978) or in at least one AVL of Sherbonaspis hillsi (Young and Gorter 1981: fig. 18C); the semilunar plate is about as long as it is wide. However, the semilunar plate remains unknown, hence it is unclear whether it is paired as in Pterichthyodes milleri or not. The contact area with the opposite AVL plate is rather strongly convex, it is about three times longer that the notch for the semilunar plate (cf.Sm), and about 1.8 times longer than the overlapping area of the MV plate (cf.MV). The area overlapping the median ventral plate is moderately wide. The area overlapping the PVL plate (cf. PVL) is rather long; the ventrolateral corner is rounded, but well discernible particularly on the visceral surface. The ventral division of the crista transversalis interna anterior (cit1) is rather low laterally, and decreases in height mesially. It is not subdivided mesially, similarly as in Sherbonaspis hillsi (Young and Gorter 1981: fig. 18), but in contrast to species of Asterolepis, Pterichthyodes, and Walterilepis (e.g., Hemmings 1978: 23; Lyarskaya 1981; Lukševičs 2021). A low ridge stretches from the lateral end of the crista transversalis interna anterior to the anterolateral corner, similarly as in Sherbonaspis hillsi (Young and Gorter 1981: fig. 18A, C).
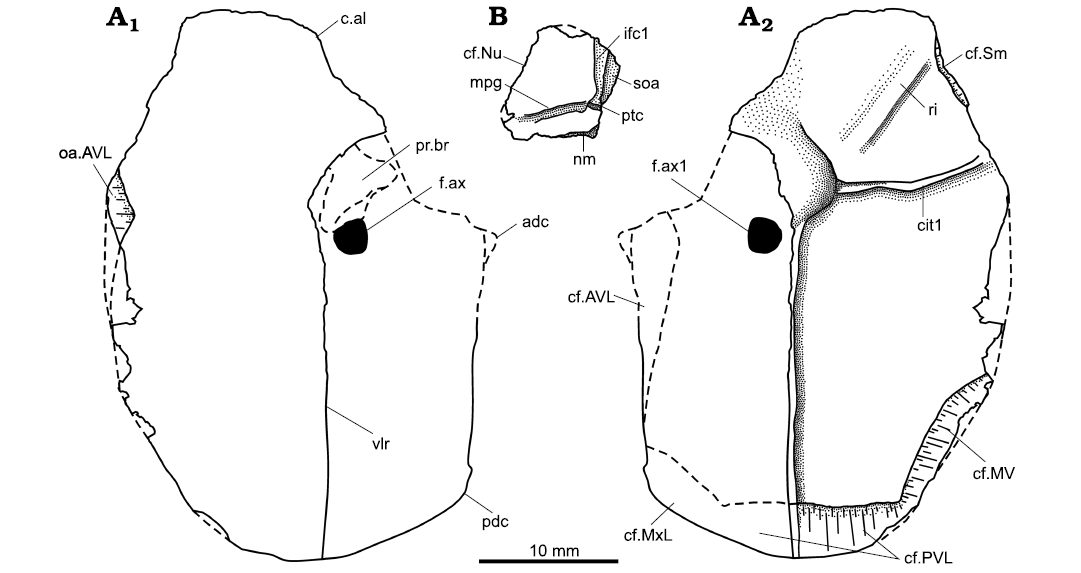
Fig. 4. Drawings of the plates of the antiarch placoderm Sherbonaspis talimaae sp. nov.; upper Emsian of Belarus. A. BNTU 121/20-1, AVL from Korma-1 borehole, plate in external (A1) and internal (A2) view. B. BNTU 44/1-13a from Osipovichi 6 borehole, Pn plate in external view. Abbreviations: adc, anterior dorsal corner; c.al, anterior lateral corner; cf.ADL, area overlapping the ADL plate; cf.MV, area overlapping the MV plate; cf.MxL, area overlapping the MxL plate; cf.Nu, area overlapping the nuchal plate; cf.PVL, area overlapping the PVL plate; cf.Sm, area overlapping the Sm plate; cit1, crista transversalis interna anterior; f.ax, axillary foramen; f.ax1, inner axillary foramen; nm, obtected nuchal area; ifc1, infraorbital sensory line canal; mpg, middle pit-line groove; oa.AVL, area overlapped by the opposite AVL plate; pdc, posterior dorsal corner; pr.br, brachial process; ptc, cephalic division of the main lateral line; ri, ridge on the visceral surface of the AVL plate; soa, subobstantic margin; vlr, ventral lateral ridge.
The lateral lamina is reconstructed at about 2.4–2.8 times as long as high, resembling proportions of the lateral lamina in Sherbonaspis hillsi and Stegolepis jugata (Malinovskaya 1973; Young and Gorter 1981), being proportionately higher than in Byssacanthus dilatatus (Karatajūte-Talimaa 1960: pl. 3: 2), but significantly lower than in Wurungulepis denisoni and Walterilepis speciosa (Young 1990: fig. 12; Lukševičs 2021: fig. 6). Judging from the outline of the dorsal margin in BNTU 44/2-1, it could be highest at the posterodorsal corner (pdc), similarly to Pterichthyodes milleri (Hemmings 1978: figs. 13, 14), Stegolepis jugata (Malinovskaya 1973: 77), and Sherbonaspis hillsi (Young and Gorter 1981: 106, fig. 18) but in contrast to Walterilepis speciosa (Lukševičs 2021), where it is highest at the anterodorsal corner. The contact face for the MxL is shorter than in Byssacanthus dilatatus, Pterichthyodes milleri, and Sherbonaspis hillsi. The shape of the foramen axillare (f.ax) is not known since it is not preserved in both available specimens.
The ornamentation is not well seen on the photographs of the specimen BNTU 121/20-1 due to glue covering the surface; however, it is well seen on the surface of the specimen. AVL plates are ornamented with a fine-meshed network of the shallow pits in the anterior part of the ventral lamina, and small tubercles with sharp tips along the margins in the posterior portion of the plate, which are well seen in BNTU 44/2-1 (Fig. 3), as well as in BNTU 121/20-1 under oblique lighting. In places these sharp small tubercles are fused by their bases in rows forming narrow ridges distributed almost perpendicularly to the margins of the plate. The lateral lamina in specimen BNTU 44/2-1 shows similar ornamentation consisting of small tubercles.
Left PVL plate BNTU 44/2-2 is well preserved despite being flattened so that the angle between the ventral and lateral laminae appears very obtuse (Fig. 3D). Another left PVL plate BNTU 158/1-1 is less complete, particularly the ventral lamina is fairly damaged; however, the lateral lamina is well preserved (Fig. 3E). The PVL plate is rather small, reaching only 18.7 mm in length in the specimen BNTU 44/2-2 and about 30 mm in BNTU 158/1-1. Almost all edges of the ventral lamina of the PVL plate in BNTU 44/2-2 are well preserved, except the middle and posterior portion of the mesial edge, which are represented by the impression on the surface of the rock, and the anterior margin coinciding with the area overlapped by the AVL plate. In contrast, most edges of the ventral lamina in BNTU 158/1-1 are damaged. Almost all edges of the lateral lamina of the PVL are well preserved in both specimens, except small portion along the anterior dorsal margin in BNTU 158/1-1. The angle between the ventral and lateral laminae cannot be measured with confidence due to deformation of both specimens of the PVL plate; the measured angle is about 145° wide, which is too large to be undeformed. The ventral lamina is moderately elongated with a B/L index of about 0.44, resembling that as it is restored in Sherbonaspis hillsi, with the B/L index of 0.47 (Young and Gorter 1981: fig. 15), and being relatively longer than that in Sherbonaspis andreannae, with the B/L index of 0.4 (Panteleyev 1993: 66). The index is larger than the B/L index in Stegolepis jugata (Malinovskaya 1973: pl. 9: 6; B/L index of 0.4), and slightly smaller than that in Pterichthyodes milleri (Hemmings 1978: text-fig. 15; B/L index 0.47–0.52, 0.5 on the average). The subanal division is relatively short and comprises about 21% of the total length of the ventral lamina similarly as in Sherbonaspis hillsi (Young and Gorter 1981), which is less than the corresponding proportion in Sherbonaspis andreannae (Panteleyev 1993), Stegolepis jugata, and Pterichthyodes milleri. The lateral margin is almost straight, being gently convex in the anterior part; it bears small closely set prominent tubercles along the ventrolateral ridge and the whole lateral edge. The area overlapping the MV plate is rather long (Fig. 5B) indicating a relatively large MV plate (Fig. 6), as in Stegolepis, Byssacanthus, and Sherbonaspis, and in contrast to Pterichthyodes and Gerdalepis (Gross 1941; Karatajūte-Talimaa 1960; Malinovskaya 1973; Hemmings 1978; Young and Gorter 1981). The left PVL overlies the right one similarly as in species of Pterichthyodes, Sherbonaspis, and Stegolepis. The ventral lamina is about 1.5 times broad as the lateral lamina is high. The length/width index for the lateral lamina is 2.3 in BNTU 44/2-2, similarly as in Byssacanthus dilatatus, proportionally higher than in Stegolepis jugata or species of Asterolepis, but proportionally lower than in Sherbonaspis hillsi, Wurungulepis denisoni and Gerdalepis jesseni (Karatajūte-Talimaa 1960: pl. 3: 4; Malinovskaya 1973: pl. 9: 6; Young and Gorter 1981: 107; Friman 1987: fig. 5; Young 1990: fig. 12). The dorsal margin of the lateral lamina is almost straight anteriorly from the prominent dorsal corner (dc). The posterior margin of the lateral lamina is concave; overall shape of the lateral lamina resembles that in Sherbonaspis hillsi and Sherbonaspis andreannae (Young and Gorter 1981: fig. 15D; Panteleyev 1993: fig. 4a), but differs from that in Pterichthyodes milleri, Byssacanthus dilatatus, Wurungulepis denisoni, or Stegolepis jugata. The external surface of the posterior portion of the PVL plate BNTU 44/2-2 has been lost, but the slightly elevated area in the impression of the visceral surface indicates a broad transverse thickening on the ventral lamina. The PVL plates are ornamented with small tubercles sometimes fused into tuberculate ridges; tubercles tend to form rows usually perpendicular or in places parallel to the margins of the plate.
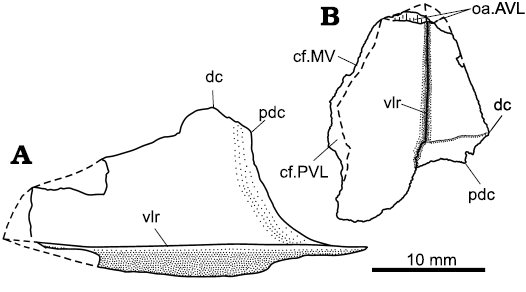
Fig. 5. Drawings of the left ventral lateral plates of the antiarch placoderm Sherbonaspis talimaae sp. nov.; upper Emsian of Belarus. A. BNTU 158/1-1 from Smol’ki 6п borehole, in lateral view. B. BNTU 44/2-2 from Osipovichi 6 borehole, in external view. Abbreviations: cf.MV, area overlapping the MV plate; cf.PVL, area overlapping the opposite PVL plate; dc, dorsal corner; oa.AVL, area overlapped by the AVL plate; pdc, posterior dorsal corner; vlr, ventral lateral ridge.
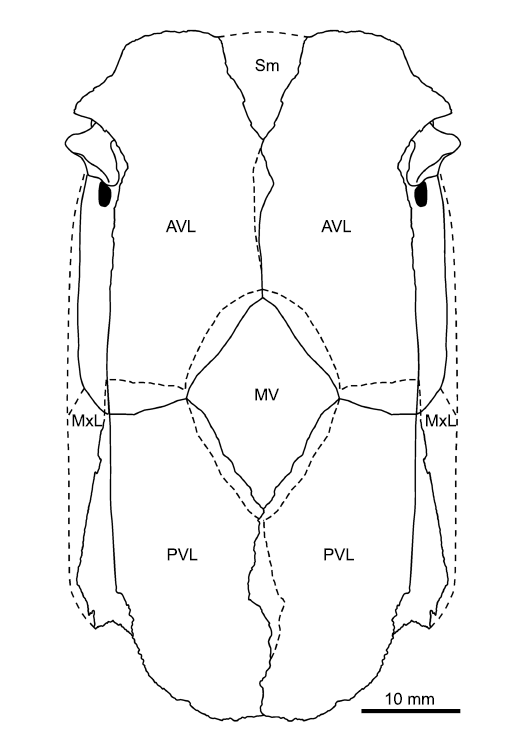
Fig. 6. Reconstruction of the ventral wall of the trunk armour of the antiarch placoderm Sherbonaspis talimaae sp. nov., based on BNTU 44/2-1, 44/2-2, 121/20-1, and 158/1-1; upper Emsian of Belarus. Abbreviations: AVL, anterior ventral lateral plate; MV, median ventral plate; MxL, mixilateral plate; PVL, posterior ventral lateral plate; Sm, semilunar plate.
Remarks.—Sherbonaspis talimaae sp. nov. differs well from such asterolepidoids as Wurungulepis denisoni, Byssacanthus dilatatus, asterolepidids, Pterichthyodes milleri, Gerdalepis rhenana and Gerdalepis jesseni, and Walterilepis speciosa by many features. Sherbonaspis talimaae sp. nov. resembles Sherbonaspis hillsi in (i) small size; (ii) B/L index of ventral lamina of AVL; (iii) short subcephalic division; (iv) semilunar plate which is as long as it is broad; (v) semilunar plate with a narrow trapezoidal shape; (vi) crista transversalis interna anterior (cit1) which is not subdivided mesially; (vii) proportions and shape of the lateral lamina of AVL; (viii) shape and proportions of the ventral lamina of PVL; (ix) short subanal division; (x) tubercular ornamentation. Sherbonaspis talimaae sp. nov. differs from Sherbonaspis hillsi in (i) that the lateral margin of the ventral lamina of PVL is almost straight; (ii) L/B index of lateral lamina of PVL. Sherbonaspis andreannae is a somewhat larger fish; however, it resembles Sherbonaspis talimaae sp. nov. in various features, but differs in: (i) B/L index of the ventral lamina of PVL; (ii) shorter subanal division; (iii) L/B index of lateral lamina of PVL; (iv) more ridged ornamentation. Sherbonaspis talimaae sp. nov. resembles Stegolepis jugata in (i) the short subcephalic division; (ii) proportions of the lateral lamina of AVL; (iii) tuberculate ornamentation. However, it differs from Stegolepis jugata in (i) B/L index of ventral lamina of AVL; (ii) shape and (iii) proportions of the semilunar plate; (iv) L/B index of lateral lamina of PVL; (v) less ridged ornamentation.
Stratigraphic and geographic range.—Upper Emsian/lower Eifelian, Lower/Middle Devonian; eastern part of Belarus.
Discussion
Taxonomic remarks.—The Antiarcha is currently considered as the most basal group of jawed vertebrates (Brazeau 2009; Zhu et al. 2012, 2013; Pan et al. 2017) comprising yunnanolepidoids, Minicrania, sinolepids and euantiarchs (Zhu 1996; Zhu and Janvier 1996). Euantiarchs, consisting of two suborders, the Bothriolepidoidei and the Asterolepidoidei, are defined by the well-developed brachial process (Young and Gorter 1981; Janvier and Pan 1982; Zhu 1996; Zhu and Janvier 1996; Lukševičs 2001). However, some authors treat these two groups as the orders Bothriolepiformes and Asterolepiformes (Moloshnikov 2010, 2012; Van der Laan 2018). At least six families were established within the Asterolepidoidea: Asterolepididae Traquair, 1888, Remigolepididae Stensiö, 1931, Pterichthyodidae Stensiö, 1948, Lepadolepididae Berg, 1955, Pambulaspidae Young, 2010, and Asperaspididae Moloshnikov, 2012.
Young and Gorter (1981) originally attributed the genus Sherbonaspis to the family Pterichthyodidae. They also provided a shortened definition of the Pterichthyodidae from the diagnosis proposed by Hemmings (1978) and attributed Stegolepis to this family, following Malinovskaya (1973). Later Panteleyev (1993) followed this attribution establishing another species of Sherbonaspis, namely Sherbonaspis andreannae. Previously Karatajūte-Talimaa (1960, 1963) and Obruchev (1964) had included Pterichthyodes, Byssacanthus, and Gerdalepis in the family Pterichthyodidae. Berg (1955) established a separate family Lepadolepididae containing Gerdalepis, Grossaspis, and Lepadolepis, whilst Gross (1965) suggested to unite Gerdalepis, Grossaspis, and Lepadolepis into a subfamily Gerdalepidinae within the Pterichthyodidae. Zhu (1996) provided a cladistic analysis of all antiarchs and demonstrated that the Pterichthyodidae as defined earlier appears polyphyletic. The most comprehensive recent analysis of antiarch relationships based on 42 ingroup taxa and 79 characters (Wang and Zhu 2018) demonstrated the monophyly of the Asterolepidoidei. However, the analysis shows difficulties to define smaller, family level groups except the family Asterolepididae uniting Asterolepis, Pambulaspis, and Remigolepis, and relatively weakly defined group of the three genera, which is treated as the family Lepadolepididae by some authors (e.g., Moloshnikov 2012). Surprisingly, the family Jiangxilepidae with Jiangxilepis, Ningxialepis, and Kirgisolepis, appeared in the cladogram as belonging to the Asterolepidoidei, despite similarities with Dianolepis. Jiangxilepis and Ningxialepis were usually treated as belonging to the Bothriolepidoidea (Jia et al. 2010; Young and Lu 2020). Kirgisolepis was attributed to the Bothriolepidoidei, the family Dianolepididae (Moloshnikov 2012) or argued to be closely related to Byssacanthus and placed among the basal asterolepidoids (Young 2010). According to the cladograms (Wang and Zhu 2018), both the strict-consensus tree and the 50% majority-rule consensus trees show a polytomy consisting of five genera (Wurungulepis, Byssacanthus, Stegolepis, Pterichthyodes, and Sherbonaspis) and two sister-groups, Asterolepididae and Lepadolepididae of some authors, thus demonstrating Pterichthyodidae as a polyphyletic group. However, some taxa of antiarchs described before 2018 were not included in the analysis of Wang and Zhu (2018), e.g., Asperaspis carinata Panteleyev, 1993, from the Givetian of Kazakhstan, and Merimbulaspis meemannae Young, 2010, from the ?Givetian/Frasnian of New South Wales, Australia.
Moloshnikov (2012) attributed Asperaspis to a separate family Asperaspididae containing only one genus and thus being a redundant taxonomic unit. Asperaspis differs well from other asterolepidoid antiarchs in the absence of the median ventral plate of the trunk armour, and in short subcaudal region of the ventral wall of trunk shield. It differs from Asterolepis in the well-developed separate PDL plate and PL plate, as well as in the unpaired semilunar plate. It differs also from Remigolepis in the unpaired semilunar plate. However, Asperaspis demonstrates the low, relatively long trunk armour, thus more closely resembling Asterolepis and Remigolepis, and not pterichthyodids or lepadolepidids with generally short and high trunk armour.
Young (2010) described Merimbulaspis from a small collection of plates with distinctive ornamentation, separate PDL plate, similar overlap relationships between the dorsolateral plates of the trunk armour and the location of the lateral line sensory groove on a separate PL plate, thus indicating a close relation to Pambulaspis. These similarities were used to erect a separate family Pambulaspidae (Young 2010).
Despite the polyphyletic condition having been demonstrated for the family Ptericthyodidae, some authors continue using this grouping. Moloshnikov (2012) attributed Sherbonaspis to the family Pterichthyodidae divided into two subfamilies: Pterichthyodinae with Pterichthyodes, Sherbonaspis, and Wurungulepis, and Byssacanthidinae containing Byssacanthus and Stegolepis. Lukševičs (2021) provided a new description of Walterilepis speciosa (Gross, 1933), based on the type specimen and new material from the Frasnian deposits of Latvia. Previously known material of Walterilepis consisted only of the holotype, comprising articulated nuchal and postpineal plates. New material includes various plates of the head shield and the trunk armour demonstrating characters of asterolepidoids with a high and short armour, thus Walterilepis was attributed to Pterichthyodidae.
New cladistic analyses were performed within the frame of this study using the heuristic algorithms of PAUP. The antiarch species Walterilepis speciosa, Asperaspis carinata, and Merimbulaspis meemannae were added to the list of ingroup taxa of Wang and Zhu (2018). The first approach is based on the same 79 characters offered by Wang and Zhu (2018) and one additional character: the PDL plate overlaps the ADL plate in dorsal part and is overlapped in ventral part (0 absent, 1 present). In total 45 antiarch taxa were included in the analysis in the first approach (see SOM, Supplementary Online Material available at http://app.pan.pl/SOM/app68-Plax_Luksevics_SOM.pdf); the “actinolepid” arthrodire Kujdanowiaspis and the “acanthothoracid” Romundina were used as the outgroup taxa. In the second approach, only 31 taxa of euantiarchs were involved in the analysis; Parayunnanolepis and Grenfellaspis were utilised as outgroup taxa.
The strict consensus topology of 185 best trees of 45 antiarch taxa matches well the results of Wang and Zhu (2018), surprisingly demonstrating inability to clearly separate Bothriolepidoidei and Asterolepidoidei (Fig. 7). For instance, Ningxialepis from the Famennian of China and Kirgisolepis from the Upper Devonian of Kyrgyzstan are shown as the sister group of Byssacanthus from the Eifelian/Givetian of the Baltic Devonian Basin, most probably due to the dorsal spine on the AMD plate. However, the dorsal spine in Byssacanthus is situated almost in the middle of the AMD plate (Karatajūte-Talimaa 1960), whereas in Ningxialepis and Kirgisolepis it appears on both the AMD and PMD plates (Jia et al. 2010; Panteleyev 1992). In most previous analyses Ningxialepis and Kirgisolepis were attributed to Bothriolepidoidei, being closely related to Jiangxilepis (Jia et al. 2010; Moloshnikov 2012). Moloshnikov (2012) has attributed Kirgisolepis along with Dianolepis, Jiangxilepis, and Ningxialepis to the family Dianolepididae, subfamily Dianolepidinae.
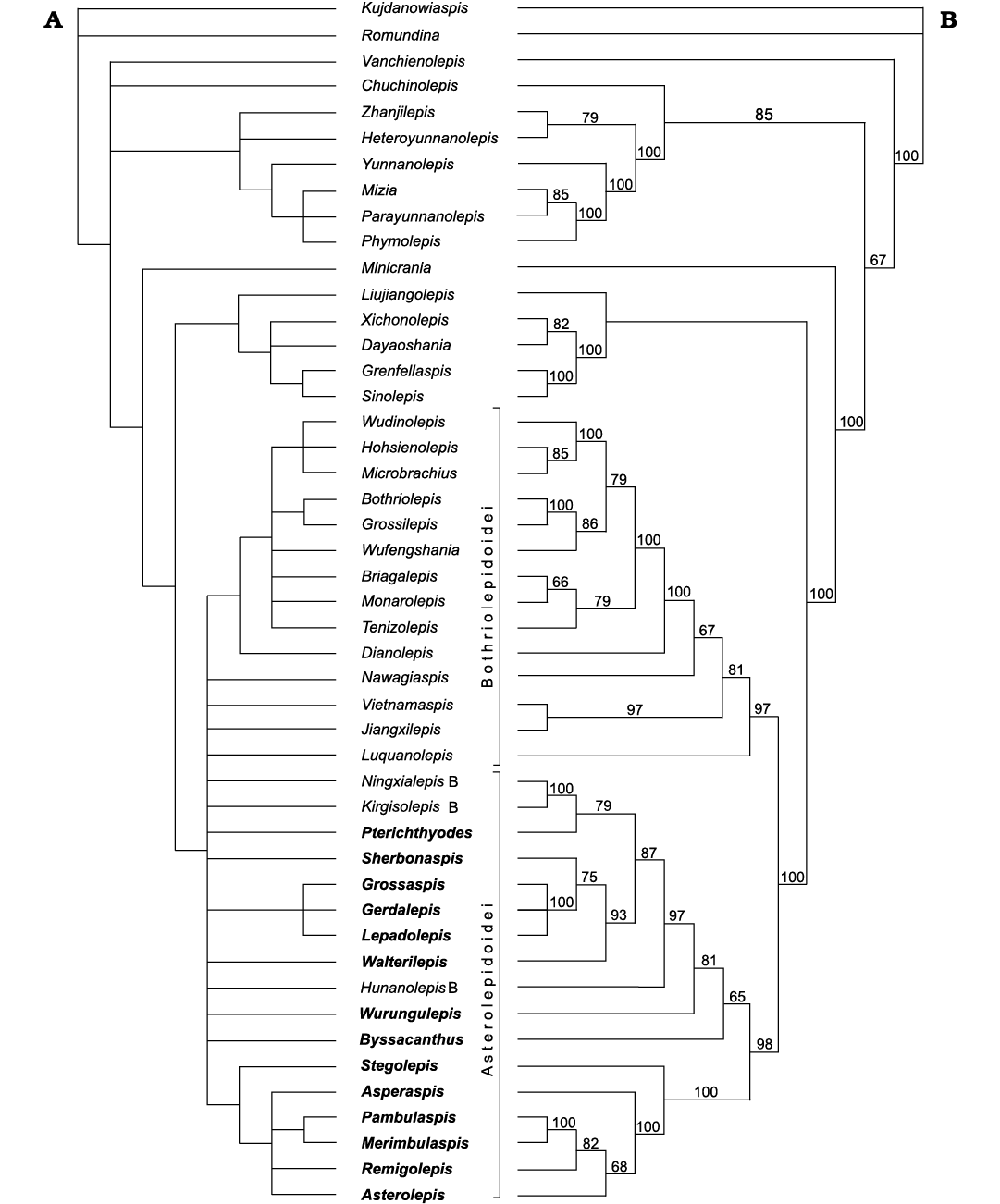
Fig. 7. Phylogeny of 45 taxa of Antiarcha. A. Strict consensus tree of 185 most parsimonious trees based on the data-set from Wang and Zhu (2018), with addition of one character (SOM: character 80) and three antiarchs species (Asperaspis carinata, Walterilepis speciosa and Merimbulaspis meemannae). B. 50% majority-rule consensus tree of the 185 most parsimonious trees. Numbers on branches indicate the percentage of most parsimonious trees that contain a particular clade. Legend: B, taxa traditionally attributed to Bothriolepidoidea. Taxa in bold traditionally attributed to Asterolepidoidea.
The strict consensus tree of all antiarch taxa shows that Asperaspis might be attributed to the family Asterolepididae, or it is a sister taxon of the Asterolepididae + Pambulaspidae in accordance with the 50% majority-rule consensus tree. The other family-level groups are the Pambulaspididae and Lepadolepididae. All the other Asterolepidoidei are forming a wide group of relatively small fishes including Byssacanthus, Hunanolepis, Pterichthyodes, Sherbonaspis, Stegolepis, Walterilepis, and Wurungulepis. The 50% majority-rule consensus tree, however, allow to separate two groups of asterolepidoids. One group consists of Byssacanthus, Wurungulepis, Hunanolepis, and two more crown-ward placed clades. The first contains Pterichthyodes, Ningxialepis, and Kirgisolepis, and the second Sherbonaspis and Walterilepis, which are more closely related to Lepadolepididae. The second group contains Stegolepis, Asperaspis, and Asterolepididae + Pambulaspidae.
However, the cladistics analysis of Euantiarcha alone (31 ingroup taxa) demonstrates a slightly different situation (Fig. 8). Both the strict consensus tree and the 50% majority-rule tree calculated from the 124 best trees shows relatively stable cladogram and clear distinction of Bothriolepidoidei (including Ningxialepis and Kirgisolepis, which appear as a sister group of Jiangxilepis) and Asterolepidoidei with Hunanolepis as one of the basal-most asterolepidoids. In accordance with the strict consensus tree and the 50% majority-rule tree Sherbonaspis is closely related to Pterichthyodes, Wurungulepis, and Walterilepis, as well as to the Lepadolepididae. Two genera, Stegolepis and Byssacanthus, form a small clade situated between the Asperaspis + Asterolepididae + Pambulaspidae, and all the other small asterolepidoids. Bootstrap values indicate strong support (79) for monophyly of Pambulaspidae, moderately strong support (62) for a monophyly of Ningxialepis and Kirgisolepis, the same value of support (62) for Lepadolepididae. Moderately strong support (50) suggests close relationships of Pambulaspidae with Asterolepis and Remigolepis. The other grouping all are weakly supported. However, these results demonstrate that the phylogenetic analysis of the Euantiarcha alone provides more reliable results if compare with the analysis of all antiarchs together. This is presumably because the number of characters that are uncodable for some taxa increases in case of all antiarchs, therefore certain homology judgements become less reliable. These results also clearly support the point of view of Young (2010) that there are too many gaps in our knowledge of antiarch morphology, therefore any phylogenetic scheme still should be treated as very tentative.

Fig. 8. Phylogeny of Euantiarcha. A. Strict consensus tree of 124 the most parsimonious trees based on the shortened and revised data-set from Wang and Zhu (2018). B. 50% majority-rule consensus tree of the most parsimonious trees. Numbers on branches indicate the percentage of most parsimonious trees that contain a particular clade (100% unless otherwise indicated). Taxa in bold traditionally attributed to Asterolepidoidea.
Palaeogeography and stratigraphic record.—Antiarchs are among the most widely distributed placoderm fishes of the middle Palaeozoic (e.g., Zhao and Zhu 2010; Dupret et al. 2023), particularly such genera as Bothriolepis and Remigolepis. However, several groups of antiarchs show a high degree of endemism. Members of Asterolepidoidei (including poorly known species of Venezuelepis) have hitherto been reported from Antarctica, Australia, Central Asia (Kazakhstan and Kirgisia), China, Euramerica, and South America (e.g., Young and Moody 2002; Zhao and Zhu 2010; Moloshnikov 2012). The most ancient asterolepidoids occur in the Emsian from the Georgina Basin in Central Australia (Young 1984), where a small possibly juvenile asterolepidoid slightly resembling Pterichthyodes was found, and the Emsian/Eifelian of New South Wales, Australia, producing Sherbonaspis hillsi (Fig. 9). The age of the fossil bearing deposits of Sherbonaspis hillsi originally was reported as the early Eifelian (Young and Gorter 1981), but later was re-evaluated as probably late Emsian or earliest Eifelian (Hunt and Young 2012). Another early asterolepidoid, Wurungulepis denisoni Young, 1990, is known from the Eifelian of Queensland, Australia (Young 1990). Two more species of Sherbonaspis, namely Sherbonaspis andreannae has been recorded in the Givetian of Kazakhstan (Panteleyev 1993), and Sherbonaspis talimaae sp. nov. in the ?upper Emsian of Belarus. Prior to this study, the oldest asterolepidoids from Euramerica were Byssacanthus from Eifelian–Givetian of the Baltic Devonian Basin, an unnamed antiarch probably related to Byssacanthus and Gerdalepis (Otto 1999), and Gerdalepis jesseni Friman, 1982, from the middle Eifelian of Germany. Pterichthyodes milleri comes from Eifelian–lower Givetian of Scotland (Hemmings 1978). The known evidence suggests that asterolepidoids were apparently absent in the Early Devonian of China. Instead, the earliest members of asterolepidoid group occur in Gondwana, and appear slightly later in Belarus, later almost simultaneously elsewhere on the territory of the East European Platform, in Germany, China, and Kazakhstan. This agrees with the suggestion of Young (1984; see also Young et al. 2010) that the primary distribution of asterolepidoid antiarchs did not include South China and consideration that asterolepidoids originated elsewhere but not in South China. However, it is difficult to decipher the dispersion routes of early asterolepidoids, particularly taking into account the small size and possible swimming abilities of these bottom-dwelling fishes.
All the other species of asterolepidoid antiarchs are recorded in the younger strata, from the Givetian up to the Famennian (Fig. 9). Some of these antiarchs show almost cosmopolitan distribution, e.g., several species of Remigolepis are known from Australia, Belgium, China, Central Russia, East Greenland, and Kazakhstan, as well as possible Remigolepis reported from Scotland (Andrews 1978; Johanson 1997; Lukševičs 1991; Olive 2015; Pan et al. 1987; Panteleyev 1992).
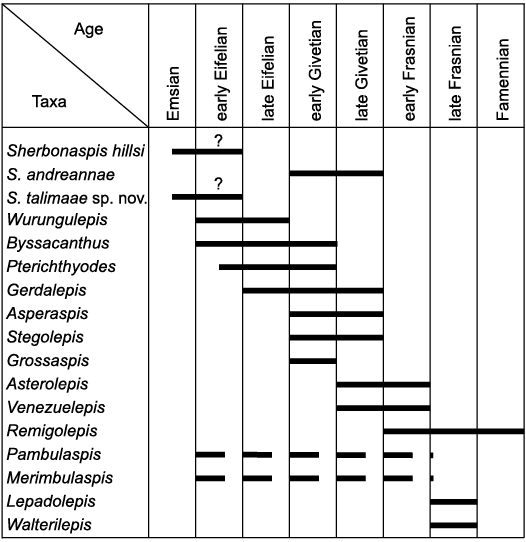
Fig. 9. Ranges of the Asterolepidoidei (based on data from Andrews 1978; Friman 1982; Gross 1941; Hemmings 1978; Johanson 1997; Karatajūte-Talimaa 1960; Lukševičs 1991, 2021; Lyarskaya 1981; Malinovskaya 1973; Moloshnikov 2012; Olive 2015; Pan et al. 1987; Panteleyev 1992, 1993; Stensiö 1931; Zhao and Zhu 2010; Young 1984, 1990; Young and Gorter 1981; Young and Moody 2002).
Stratigraphy.—All of the material of Sherbonaspis talimaae sp. nov. described above comes from the deposits of the upper part of the Vitebsk Formation, corresponding to the Lepel Beds, Vitebsk RS. In accordance with the current Devonian stratigraphical chart of Belarus (Obukhovskaya et al. 2010) the Vitebsk RS corresponds to the upper Emsian. Rocks of the Lepel Beds usually overlap deposits of the Obol Beds of the same Vitebsk RS or lie on the Precambrian basement. Lepel Beds are covered everywhere with deposits of the Adrov RS corresponding to the Middle Devonian Eifelian Stage. The Lepel Beds contain various ostracods, lingulate brachiopods, and a rich assemblage of fishes including acanthodians, placoderms and other fishes from the Laliacanthus singularis Acanthodian Zone and the Skamolepis fragilis Thelodont Zone (Mark-Kurik 2000; Valiukevičius and Kruchek 2000). As it already mentioned above, deposits of the Lepel Beds correspond to the Diaphanospora inassueta Miospore Zone (Valiukevičius and Kruchek 2000), or the upper part of the Rhabdosporites mirus–Gneudnaspora divellomedium Miospore Zone, while in terms of conodonts they roughly correspond to the upper part of the Polygnathus costatus patulus Zone of the previous version of conodont zonation (Obukhovskaya et al. 2010). However, conodonts have not been found in the deposits of the Vitebsk Formation. Recently Vodrážková et al. (2011) granted Polygnathus patulus and P. partitus full species status. Hence, the Upper Polygnathus patulus Zone (now Polygnathus partitus Zone) subsequently defines the Emsian/Eifelian boundary: Polygnathus patulus Zone corresponds to the uppermost Emsian and Polygnathus partitus Zone corresponds to the lowermost Eifelian in a modern conodont zonation (e.g., Aboussalam et al. 2015). In the Baltic States, almost all the authors have treated the upper part of the Rēzekne Formation as the approximate age analogue of the deposits of Lepel Beds of Belarus (Lyarskaya 1978; Lyarskaya and Kleesment 1981; Mark-Kurik 2000; Valiukevičius and Kruchek 2000; Obukhovskaya et al. 2010). However, the Emsian/Eifelian boundary in the Baltic States still is a matter of discussion (Fig. 10). The Laliacanthus singularis Acanthodian Zone is common for both the Rēzekne and Pärnu formations; this zone supposedly marks the beginning of a Middle Devonian level in acanthodian phylogenetical development (Valiukevičius and Kruchek 2000). From another aspect, the age of the Laliacanthus singularis Zone is different in accordance with the acanthodian and miospore zonation. According to miospores, the lower part of the Rēzekne Formation is late Emsian in age, whereas the upper two members of the Rēzekne Formation correspond to the Periplecotriletes tortus Zone and are dated as earliest Eifelian (Valiukevičius and Kruchek 2000). Attribution of the Rēzekne Formation to the upper Emsian by Mark-Kurik (e.g., Mark-Kurik 2000; Mark-Kurik and Põldvere 2012) is based on the distribution of placoderms Wijdeaspis and the Skamolepis fragilis Thelodont Zone. However, acanthodians found together with Wijdeaspis correspond to an older assemblage than that of the Laliacanthus singularis Zone (Valiukevičius and Kruchek 2000). It seems that the data on miospore distribution and their correlation to the conodont zones remain controversial. Thus, Mark-Kurik (2000: 312) stated that “it is reasonable for the time being to preserve the boundary in the Baltic between the Rēzekne and Pärnu formations”. However, analysis of distribution of various vertebrates in the Middle Devonian of the East European Platform (Lukševičs et al. 2010) supported the view of Valiukevičius and Kruchek (2000) that the Laliacanthus singularis Zone might correspond in part to the upper Emsian, and in part to the lower Eifelian. Thus, the lower part of the Rēzekne Formation would correspond to the upper Emsian, and the two upper members to the lower Eifelian, with the upper part of the Rēzekne Formation in the modern stratigraphic scheme of Latvia corresponding to the lowermost Eifelian (Lukševičs et al. 2010; Lukševičs and Stinkulis 2018). If the correlation of the Lepel Beds with the upper part of the Rēzekne Formation is correct, then an early Eifelian age for Sherbonaspis talimaae sp. nov. cannot be excluded.
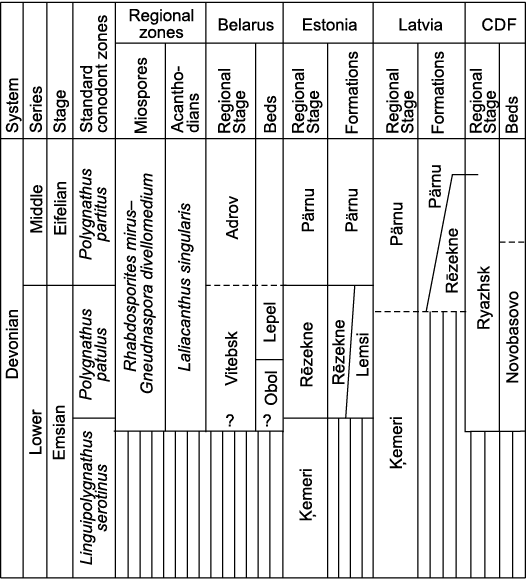
Fig. 10. Stratigraphy and possible correlation of the Emsian–Eifelian deposits of Belarus, Estonia, Latvia, and Central Russia (CDF) (data from Lukševičs and Stinkulis 2018; Mark-Kurik 2000; Mark-Kurik and Põldvere 2012; Obukhovskaya et al. 2010; Valiukevičius and Kruchek 2000).
Conclusions
The new antiarch from the Vitebsk Formation, Lepel Beds, Sherbonaspis talimaae sp. nov., is based on the specimens collected from three boreholes in the central and eastern parts of Belarus. This new species shows independent PVL and PL plates, which constitutes one of apomorphic characters of Asterolepidoidei (Wang and Zhu 2018). Sherbonaspis talimaae sp. nov. resembles Sherbonaspis hillsi in the shape and proportions of the AVL and PVL plates, the shape of the notch for semilunar plate, and the character of ornamentation. The result of a new cladistic analysis of the Euantiarcha supports the traditional subdivision of the group into Bothriolepidoidei and Asterolepidoidei; however, polyphyly of Pterichthyodidae is in agreement with the hypothesis of Zhu (1996). Until the phylogenetic relationships of Asterolepidoidei are better resolved, Sherbonaspis is attributed to Pterichthyodidae. The comparison of the sections of the Lepel Beds of Belarus with the age-equivalent deposits from Estonia and Latvia shows the necessity of further stratigraphic studies including such age-markers as conodonts and spores to clarify a late Emsian or possibly early Eifelian age for the Lepel Beds. The new antiarch taxon is a significant taxonomic addition to the Emsian/Eifelian fish assemblage from the western part of the East European Platform.
Authors’ contributions
D. Plax, investigation, visualization, writing: original draft; E. Lukševičs, formal analysis, software, visualization, writing: review and editing.
Acknowledgements
The authors are grateful to Gavin C. Young (Australian National University, Canberra, Australia) and Per E. Ahlberg (Uppsala University, Sweden) for fruitful discussion. We thank Per E. Ahlberg also for the accurate reading and correction of the manuscript, and Dmitry Stepanenko (Minsk, Belarus) for the photography. The authors thank the editors and both reviewers Vincent Dupret (Uppsala University, Sweden) and Gavin C. Young (Australian National University, Canberra, Australia) for their fruitful remarks.
References
Aboussalam, Z.S., Becker, R.T., and Bultynck, P. 2015. Emsian (Lower Devonian) conodont stratigraphy and correlation of the Anti-Atlas (Southern Morocco). Bulletin of Geosciences 90: 893–980. Crossref
Andrews, S.M. 1978. A possible occurrence of Remigolepis in the topmost Old Red Sandstone of Berwickshire. Scottish Journal of Geology 14: 311–315. Crossref
Bessonova, V.Y., Sinichka, A.M. [Sinička, A.M.], Golubtsov, V.K. [Golubcov, V.K.], and Medvedev, M.G. 1972. Opornye skvažiny SSSR. Opornaâ skvažina №2 Oršanskoj vpadiny. 232 pp. Nauka i tehnika, Minsk.
Berg, L.S. 1955. Classification of fishes and fish-like animals, living and fossil [in Russian]. Trudy Zoologičeskogo Instituta Akademiia Nauk SSSR 20: 1–286.
Beyrich, E. 1877. Uber einen Pterichthys von Gerolstein. Zeitschrift der Deutschen Geologischen Gesellschaft 29 (4): 751–756.
Brazeau, M.D. 2009. The braincase and jaws of a Devonian ‘acanthodian’ and modern gnathostome origins. Nature 457: 305–308. Crossref
Dupret, V., Byrne, H.M., Castro, N., Hammer, Ø., Higgs, K.T., Long, J.A., Niedźwiedski, G., Qvarnström, M., Stössel, I., and Ahlberg, P.E. 2023. The Bothriolepis (Placodermi, Antiarcha) material from the Valentia Slate Formation of the Iveragh Peninsula (middle Givetian, Ireland): Morphology, evolutionary and systematic considerations, phylogenetic and palaeogeographic implications. PLoS ONE 18 (2): e0280208. Crossref
Eichwald, E. 1844. Über die Fische des devonischen Systems in der Gegend von Pawlowsk. Bulletin of the Moscow Society of Naturalists. Geological Series 17: 824–843.
Friman, L. 1982. Ein mitteldevonischer Antiarche, Gerdalepis jesseni n. sp., aus der Sötenicher Mulde, NordEifel (Rheinisches Schiefergebirge). Paläontologische Zeitschrift 56: 229–234. Crossref
Golubtsov, V.K. [Golubcov, V.K.], Kedo, G.I., and Kruchek, S.A. [Kruček, S.A.] 1983. Eifelian Stage of Belarus [in Russian]. In: V. Dubatolov (ed.), Nižnij ârus srednego devona SSSR. Trudy instituta geologii i geofiziki 562: 51–56. Crossref
Gross, W. 1933. Die Fische des Baltischen Devons. Palaeontographica 79: 1–74.
Gross, W. 1941. Neue Beobachtungen an Gerdalepis rhenana (Beyrich). Palaeontographica 93A: 193–214.
Gross, W. 1965. Über die Placodermen-gattungen Asterolepis und Tiaraspis aus dem Devon Belgiens und einen fraglichen Tiaraspis-rest aus dem Devon Spitsbergens. Bulletin Institut royal des Sciences naturelles de Belgique 41 (16): 1 –19.
Gross, W. 1973. Kleinschuppen, flossenstacheln und zähne von fischen aus Europäischen und Nordamerikanischen bonebeds des Devons. Palaeontographica 142: 51–155.
Hemmings, S.K. 1978. The Old Red Sandstone antiarchs of Scotland: Pterichthyodes and Microbrachius. Palaeontographical Society (Monographs) 131: 1–64. Crossref
Hunt, J.R. and Young, G.C. 2012. Depositional environment, stratigraphy, structure and paleobiology of the Hatchery Creek Group (Early–?Middle Devonian) near Wee Jasper, New South Wales. Australian Journal of Earth Sciences 59: 355 –371. Crossref
Janvier, P. and Pan, J. 1982. Hyrcanaspis bliecki n. g. n. sp., a new primitive euantiarch (Antiarcha, Placodermi) from the Middle Devonian of Northeastern Iran, with a discussion on antiarch phylogeny. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 164: 364–392. Crossref
Jia, L.-T., Zhu, M., and Zhao, W.-J. 2010. A new antiarch fish from the Upper Devonian Zhongning Formation of Ningxia, China. Palaeoworld 19: 136–145. Crossref
Johanson, Z. 1997. New Remigolepis (Placodermi; Antiarchi) from Canowindra, New South Wales, Australia. Geological Magazine 134: 813–846. Crossref
Karatajūtė-Talimaa, V. 1960. Byssacanthus dilatatus (Eichw.) from the Middle Devonian of the U.S.S.R. In: V. Gudelis (ed.), Collectania Acta Geologica Lithuanica, 293–305. Geological and Geographical Institute, Academy of Sciences of the Lithuanian SSR, Vilnius.
Karatajūtė-Talimaa, V. 1963. Genus Asterolepis from the Devonian of the Russian Platform [in Russian with English summaries]. In: A. Grigelis and V. Karatajūtė-Talimaa (eds.), Voprosy geologii Litvy, 65–169. Akademiâ nauk Litovskoj SSR. Institut geologii i geografii, Vilnius.
Kruchek, S.A. [Kruček, S.A.], Makhnach, A.S. [Mahnač, A.S.], Golubtsov, V.K. [Golubcov, V.K.] and Obukhovskaya, T.G. [Obuhovskaâ, T.G.] 2001. Devonian System. In: A.S. Makhnač, R.G. Garecki, and A.V. Matveev (eds.), Geologiâ Belarusi, 186–236. Institut geologičeskih nauk Nacional’noj akadmii nauk Belarusi, Minsk.
Kruchek, S.A., Obukhovskaya, V.Y., and Plax, D.P. 2012a. A typical section of the Devonian of the Bobruysk Buried Ridge. In: A.M. Kovkhuto (ed.), Geological Science and Innovation. Proceedings of the International Scientific and Practical Conference dedicated to the 85th Anniversary of the Republican Unitary Enterprise “Belarusian Research Geological Exploration Institute”, 99–103. State enterprise “BelNIGRI”, Minsk.
Kruchek, S.A., Obukhovskaya, V.Y., and Plax, D.P. 2012b. On the boundary of Emsian–Eifelian deposits of Belarus and the problem of their correlation. In: A.M. Kovkhuto (ed.), Geological Science and Innovation. Proceedings of the International Scientific and Practical Conference dedicated to the 85th Anniversary of the Republican Unitary Enterprise “Belarusian Research Geological Exploration Institute”, 94–98. State enterprise “BelNIGRI”, Minsk.
Lukševičs, E. 1991. New Remigolepis (Pisces, Antiarchi) from the Famennian deposits of the Central Devonian Field (Russia, Tula region) [in Russian with summaries in Latvian and English]. Daba un muzejs 3: 51–56.
Lukševičs, E. 2001. Bothriolepid antiarchs (Vertebrata, Placodermi) from the Devonian of the north-western part of the East European Platform. Geodiversitas 23: 489–609.
Lukševičs, E. 2021. Revision of asterolepidoid antiarch remains from the Ogre Formation (Upper Devonian) of Latvia. Estonian Journal of Earth Sciences 70: 3–17. Crossref
Lukševičs, E. and Stinkulis, Ģ. 2018. Stratigraphic subdivision, structure, composition and regional disparities of sedimentary cover [in Latvian]. In: O. Nikodemus, M. Kļaviņš, Z. Krišjāne, and V. Zelčs (eds.), Latvija: Zeme, Daba, Tauta, Valsts, 38–58. Academic Publisher of University of Latvia, Riga.
Lukševičs, E., Lebedev, O.A., and Zakharenko, G.V. 2010. Palaeozoogeographic zonation of the Middle–Late Devonian vertebrate communities of the Baltica Province. Part I. Eifelian–Givetian. Palaeoworld 19: 94–107. Crossref
Lyarskaya, L.A. [Lârskaâ, L.A.] 1978. The Rēzekne Formation and its age equivalents [in Russian]. In: V.S. Sorokin (ed.), Stratigrafiâ fanerozoâ Pribaltiki, 22–43. Zinātne, Riga.
Lyarskaya, L.A. [Lârskaâ, L.A.] 1981. Baltic Devonian Placodermi. Asterolepididae [in Russian, with English summary]. 152 pp. Zinātne, Riga.
Lyarskaya, L.A. and Kleesment, A. 1981. Rēzekne Formation [in Russian]. In: V.S., Sorokin, L.A., Lârskaya, L.S. Savvaitova, V. Narbutas, S. Žeiba, E. Mark-Kurik, A. Kleesment, H. Vijding, K. Kajak, G.I. Kedo, T.G. Obuhovskaâ, V. Grāvītis, V. Kuršs, V.J. Nenastjeva, I.A. Polivko, V. Karatajūte-Talimaa, G. Vaitekuniene, and E.I. Vorobyeva (eds.), Devon i Karbon Pribaltiki, 66–80. Zinātne, Riga.
Malinovskaya, S. [Malinovskaâ, S.] 1973. New Middle Devonian genus Stegolepis (Antiarchi, Placodermi) from the Central Kazakhstan [in Russian]. Paleontologičeskij žurnal 1973 (2): 71–82.
Mark-Kurik, E. 2000. The Middle Devonian fishes of the Baltic States (Estonia, Latvia) and Belarus. Courier Forschungsinstitut Senckenberg 223: 309–324.
Mark-Kurik, E. and Põldvere, A. 2012. Devonian stratigraphy in Estonia: current state and problems. Estonian Journal of Earth Sciences 61 (1): 33–47. Crossref
Miles, R. 1968. The Old Red Sandstone antiarchs of Scotland: Family Bothriolepididae. Palaeontographical Society, London 1968: 1–130. Crossref
Miller, H. 1841. The Old Red Sandstone or New Walks in an Old Field. 1st Edition. 275 pp. John Johnstone, Edinburgh.
Moloshnikov, S. [Mološnikov, S.] 2010. Middle Devonian bothriolepiform antiarchs (Pisces, Placodermi) from Central Kazakhstan and their implication for the antiarch system and phylogeny [in Russian with English summary]. Paleontologičeskij žurnal 2010 (2): 70–82. Crossref
Moloshnikov, S. 2012. Middle–Late Devonian Placoderms (Pisces: Antiarchi) from Central and Northern Asia. Paleontological Journal 46 (10): 1097–1196. Crossref
Obruchev, D.V. [Obručev, D.V.] 1964. Class Placoderms (Placodermi) [in Russian]. In: D.V. Obručev (ed.), Osnovy paleontologii. Beŝelûstnye i ryby, 118–174. Nauka, Moskva.
Obukhovskaya, T.G. [Obuhovskaâ, T.G.], Kruchek, S.A. [Kruček, S.A.], Pushkin, V.I. [Puškin, V.I.], Nekryata, N.S. [Nekrâta, N.S.], Plax, D.P., Sachenko, T.Ph. [Sačenko, T.F.], Obukhovskaya, V.Y. [Obuhovskaâ, V.Y.] and Antipenko, S.V. 2010. Devonskaâ sistema. Stratigrafičeskie shemy dokembrijskih i fanerozojskih otloženij Belarusi: Poâsnitel’naâ zapiska. 282 pp. Gosudarstvennoe predpriâtie “BelNIGRI”, Minsk.
Olive, S. 2015. Devonian antiarch placoderms from Belgium revisited. Acta Palaeontologica Polonica 60: 711–731.
Otto, M. 1999. Antiarchi-Funde aus der Brandenberg-Gruppe (Mitteldevon,Eifelium) des nordlichen Sauerlandes. Mainzer Geowissenschaftliche Mit-teilungen 28: 41–62.
Pan, J., Huo, F.C., Cao, J.X., Gu, Q.C., Liu, S.Y., Wang, J.Q., Gao, L.D., and Liu, C. 1987. Continental Devonian system of Ningxia and its biotas [in Chinese, with English abstract]. 237 pp. Geological Publishing House, Beijing.
Pan, Z., Zhu, M., Zhu, Y., and Jia, L. 2017. A new antiarch placoderm from the Emsian (Early Devonian) of Wuding, Yunnan, China. Alcheringa 42 (2):1–12. Crossref
Panteleyev, N. 1992. New remigolepids and high armoured antiarchs of Kirgizia. In: E. Mark-Kurik (ed.), Fossil Fishes as Living Animals. Palaeozoic Fishes, 2nd Colloquium, 185–192. Academy of Sciences, Tallinn.
Panteleyev, N. [Pantelejev, N.] 1993. New antiarchs (Placodermi) from the Middle Devonian of Central Kazakhstan [in Russian with English summary]. Paleontologičeskij žurnal 1993 (2): 62–71.
Plaksa, D.P. 2007. Devonskaâ (pozdneemssko-franskaâ) ihtiofauna Belarusi i eë stratigrafičeskoe značenie [in Russian with Belarusian and English summary]. 23 pp. Candidate Thesis, Institut geohimii i geofiziki Nacional’noj akademii nauk Belarusi, Minsk.
Plaksa, D.P. 2008. Introduction of vertebrate zonal scales into the Stratigraphic Chart of Devonian deposits of Belarus [in Russian]. Doklady Nacional'noj Akademii nauk Belarusi 52 (4): 83–88.
Plax, D.P. 2008. Devonian fish fauna of Belarus [in Russian]. Lithosphere 29 (2): 66–92.
Plax, D.P. 2015a. Late Emsian placoderms of Belarus. In: V.N. Gubin, V.I. Zui, O.V. Lukashev, and A.F. Sanko (eds.), Topical Questions of Engineering Geology, Hydrogeology and Rational Use of Earth’s Interior. Proceedings of the IX University Geological Readings (Minsk, April, 3, 2015), 171–173. Belarusian State University, Minsk.
Plax, D.P. 2015b. Stratigraphic ichthyofauna assemblages of the Devonian deposits in the east and southeast of Belarus. Lithosphere 42 (1): 20–44.
Plax, D.P. 2016. Stratigraphic ichthyofauna assemblages of the Devonian deposits in the Vileyka Buried Ridge of the Belarusian Anteclise. Natural Resources 2: 14–44.
Plax, D.P. 2018. Antiarch fishes from the Devonian deposits of Belarus. In: S.V. Kharitonchik, A.M. Malyarevich, A.S. Kalinichenko (eds.), Proceedings of the 16 International Scientific and Technical Conference «Science for Education, Production and Economy Purposes». Vol. 1, 409. Belarusian National Technical University, Minsk.
Plax, D.P. and Murashko, O.V. 2021. Stratigraphy and ichthyofauna of the Upper Emsian-Eifelian deposits in the Bykhov 1 and Korma 1 borehole sequences in the East of Belarus. Natural Resources 1: 40–67.
Plax, D.P. and Newman, M.J. 2020. New Early Devonian (Late Emsian) placoderms from Belarus. Journal of Paleontology 94 (4): 773–787. Crossref
Rodionova, G.D., Umnova, V.T., and Kononova, L.I. 1995. Devon Voronežskoj anteklizy i Moskovskoj sineklizy. 265 pp. Central'nyj regional'nyj geologičeskij centr, Moskva.
Stensiö, E. 1931. Upper Devonian vertebrates from East Greenland, collected by the Danish Greenland Expeditions in 1929 and 1930. Meddelelser om Grønland 86 (1): 1–212.
Stensiӧ, E. 1948. On the Placodermi of the Upper Devonian of East Greenland. 2. Antiarchi: subfamily Bothriolepidae. Palaeozoologica Grønland 2: 1–622.
Swofford, D.L. 2003. PAUP: Phylogenetic Analysis Using Parsimony (and Other Methods), Version 4.0b 10. Sinauer Associates, Sunderland.
Traquair, R.H. 1888. On the structure and classification of the Asterolepidae. Annals and Magazine of Natural History (Series 6) 2 (12): 485–504. Crossref
Valiukevičius, J. and Kruchek, S. 2000. Acanthodian biostratigraphy and interregional correlations of the Devonian of the Baltic States, Belarus, Ukraine and Russia. Courier Forschungsinstitut Senckenberg 223: 271–289.
Van der Laan, R. 2018. Family-group names of fossil fishes. European Journal of Taxonomy 466: 1–167. Crossref
Vodrážková, S., Klapper, G., and Murphy, M.A. 2011. Early Middle Devonian conodont faunas (Eifelian, costatus-kockelianus zones) from the Roberts Mountains and adjacent areas in central Nevada. Bulletin of Geosciences 86 (4): 737–764. Crossref
Vorotnikova, G.V., Gavryushova, E.A., Drutskoy, S.V., and Zhuravlev, A.V. 2011. State Geological Map of the Russian Federation. Scale 1: 1 000 000 (third generation). Central European Series. Sheet N-36 (M-36) Smolensk. Explanatory Note. 267 pp. VSEGEI Cartographic Factory, St. Petersburg.
Wang, Y. and Zhu, M. 2018. Redescription of Phymolepis cuifengshanensis (Antiarcha: Yunnanolepididae) using high-resolution computed tomography and new insights into anatomical details of the endocranium in antiarchs. PeerJ 6:e4808. Crossref
Young, G.C. 1984. An asterolepidoid antiarch (placoderm fish) from the Early Devonian of the Georgina Basin, central Australia. Alcheringa 8: 65–80. Crossref
Young, G.C. 1988. Antiarchs (placoderm fishes) from the Devonian Aztec Siltstone, southern Victoria Land, Antarctica. Palaeontographica A 202: 1–125.
Young, G.C. 1990. New antiarchs (Devonian placoderm fishes) from Queensland, with comments on placoderm phylogeny and biogeography. Memoirs of the Queensland Museum 28 (1): 35–50.
Young, G.C. 2010. A new antiarch (placoderm fish: Devonian) from the south coast of New South Wales, Australia. In: D.K. Elliott, J.G. Maisey, X. Ju, and D. Miao (eds.), Morphology, Phylogeny and Palaeogeography of Fossil Fishes, 85–100. Dr. Friedrich Pfeil Verlag, München.
Young, G.C. and Gorter, J.D. 1981. A new fish fauna of Middle Devonian age from the Taemas Wee Jasper region of New South Wales. BMR Bulletin 209: 83–147.
Young, G.C. and Lu, J. 2020. Asia-Gondwana connections indicated by Devonian fishes from Australia: palaeogeographic considerations. Journal of Palaeogeography 9 (8): 1–22. Crossref
Young, G.C. and Moody, J.M. 2002. A Middle–Late Devonian fish fauna from the Sierra de Perijá, western Venezuela, South America. Mitteilungen aus dem Museum für Naturkunde in Berlin, Geowissenschaftlige Reihe 5: 155–206. Crossref
Young, G.C., Burrow, C.J., Long, J.A., Turner, S., and Choo, B. 2010. Devonian macrovertebrate assemblages and biogeography of East Gondwana (Australasia, Antarctica). Palaeoworld 19: 55–74. Crossref
Zhao, W.-J. and Zhu, M. 2010. Siluro-Devonian vertebrate biostratigraphy and biogeography of China. Palaeoworld 19: 4–26. Crossref
Zhu, M. 1996. The phylogeny of the Antiarcha (Placodermi, Pisces), with the description of Early Devonian antiarchs from Qujing, Yunnan, China. Bulletin du Muséum national d’Histoire naturelle, Paris, 4e série 18 (2–3): 233–437.
Zhu, M. and Janvier, P. 1996. A small antiarch, Minicrania lirouyii gen. et sp. nov., from the Early Devonian of Qujing, Yunnan (China), with remarks on antiarch phylogeny. Journal of Vertebrate Paleontology 16: 1–15. Crossref
Zhu, M., Yu, X.B., Ahlberg, P.E., Choo, B., Lu, J., Qiao, T., Qu, Q.M., Zhao, W.J., Jia, L.T., Blom, H., and Zhu, Y.A. 2013. A Silurian placoderm with osteichthyan-like marginal jaw bones. Nature 502: 188–193. Zhu, M., Yu, X.B., Choo, B., Wang, J.Q., and Jia, L.T., 2012. An antiarch placoderm shows that pelvic girdles arose at the root of jawed vertebrates. Biology Letters 8: 453–456. Crossref
Acta Palaeontol. Pol. 68 (3): 513–527, 2023
https://doi.org/10.4202/app.01075.2023