
On Triassic Murchisonia-like gastropods—surviving the end-Permian extinction to become extinct in the Late Triassic
ALEXANDER NÜTZEL and BARAN KARAPUNAR
Nützel, A. and Karapunar, B. 2023. On Triassic Murchisonia-like gastropods—surviving the end-Permian extinction to become extinct in the Late Triassic. Acta Palaeontologica Polonica 68 (3): 539–559.
High-spired Murchisonia-like slit-band gastropods are an important component of late Paleozoic gastropod faunas. Twenty-seven genera of such gastropods have been reported from the Permian, most of which representing the caenogastropod family Goniasmatidae. Only four genera, Trypanocochlea, Wannerispira, Laschmaspira, and Altadema crossed the Permian/Triassic boundary. Based on the study of newly collected specimens and material from natural history collections, we studied the surviving genera as well as the Triassic recovery of this group. Two new species (Laschmaspira lirata sp. nov. and Altadema hausmannae sp. nov.) and one new subfamily (Cheilotomoninae) are introduced. Murchisonia-like caenogastropods, chiefly Goniasmatidae, were diverse and abundant until the Permian, barely survived the end-Permian extinction, regained a certain generic diversity within the Triassic with the evolution of several new genera but failed by far to regain their Permian generic diversity. This once successful and diverse group shares a similar fate (surviving the end-Permian extinction, a reduced Triassic diversity and extinction during Late Triassic crises) as conodonts, orthoceratids, conulariids, and others. This diversity pattern does not qualify for the “Dead Clade Walking” phenomenon, i.e., the extinction shortly after a major mass extinction event (survival without recovery) because they have survived for ca. 30 Ma (at least until the Norian) and even produced a number of new genera. The exact time of their extinction is unknown but there are no safe Rhaetian occurrences. Their extinction is part of a long-term selective trend against the character “shell-slit”.
Key words: Gastropoda, Goniasmatidae, diversity, recovery, end-Permian extinction, Triassic, Italy, Austria.
Alexander Nützel [nuetzel@snsb.de; https://orcid.org/0000-0002-8852-7688 ], SNSB-Bayerische Staatssammlung für Paläontologie und Geologie, Richard-Wagner-Str. 10, 80333 Munich, Germany; Ludwig-Maximilians-Universität München, Department of Earth and Environmental Sciences, Palaeontology & Geobiology, Richard-Wagner-Str. 10, 80333 Munich, Germany; GeoBio-Center LMU, Richard-Wagner-Str. 10, 80333 Munich, Germany.
Baran Karapunar [karapunar@snsb.de; https://orcid.org/0000-0001-9711-1492 ], SNSB-Bayerische Staatssammlung für Paläontologie und Geologie, Richard-Wagner-Str. 10, 80333 Munich, Germany; Ludwig-Maximilians-Universität München, Department of Earth and Environmental Sciences, Palaeontology & Geobiology, Richard-Wagner-Str. 10, 80333 Munich, Germany; GeoBio-Center LMU, Richard-Wagner-Str. 10, 80333 Munich, Germany; Universität Hamburg, Institute for Geology, Department of Earth System Sciences, Hamburg, Germany.
Received 14 June 2023, accepted 19 August 2023, available online 11 September 2023.
Copyright © 2023 A. Nützel and B. Karapunar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
High-spired gastropods with selenizone (slit-band, closed shell slit) are a typical component of Paleozoic gastropod faunas from the Ordovician onwards. Like many typical Paleozoic gastropod groups, they were severely affected at the end-Permian mass extinction (Erwin 1990; Nützel 2005; Karapunar and Nützel 2021) and became a minor element in the Triassic until their demise in the Late Triassic. Due to their rarity in most of the Triassic faunas, studies on Murchisonia-like gastropods from the Triassic are scarce and the Triassic representatives of the group are less known compared to their Paleozoic representatives. Herein, we revise Triassic Murchisonia-like gastropods to better understand the diversity of this group in the Triassic and their possible relationships to Paleozoic forms.
High-spired slit-bearing gastropods have been traditionally assigned to the family Murchisoniidae or to the superfamily Murchisonioidea (e.g., Knight et al. 1960) and many of such gastropods ranging from the Ordovician to the Triassic were placed in the genus Murchisonia. However, there is a considerable number of Murchisonia-like genera and the composition of the group is quite complex. Murchisonia-like gastropods are considered either as exceptionally high-spired Pleurotomariida (Vetigastropoda) or as slit-bearing Caenogastropoda depending on the larval shell morphology and shell microstructure (Koken 1889; Knight et al. 1960; Nützel 1998; Nützel and Bandel 2000; see also Mazaev 2011). However, these characters are still unknown for many genera and hence their classification is doubtful. Murchisonia, the type genus of the group, has a Devonian type species, M. bilineata (Dechen, 1832), which is a highly variable species (Heidelberger 2001; Heidelberger and Koch 2005). Its early ontogeny, including protoconch morphology, is largely unknown and this hinders a correct taxonomic and systematic assessment of Murchisonia and Murchisoniidae in general. Several late Paleozoic Murchisonia-like gastropods (Goniasmatidae) have been reported with well-preserved early ontogenetic shells possessing caenogastropod-type larval shells (Yoo 1988, 1994; Nützel 1998; Nützel and Bandel 2000; Bandel 2002; Bandel et al. 2002; Pan and Erwin 2002; Nützel and Pan 2005; Karapunar et al. 2022). The specimens from the Pennsylvanian Buckhorn Asphalt deposit (USA, Oklahoma) further revealed that their Carboniferous representatives lack nacre but have an aragonitic crossed lamellar shell microstructure (Bandel et al. 2002). The absence of nacre supports a placement of these gastropods in Caenogastropoda because vetigastropods including Pleurotomariida commonly possess nacre, and only some vetigastropods are known to lack nacre (e.g., Skeneidae, Fissurelloidea; Ponder et al. 2020). By contrast, nacre is unknown in Gastropoda outside Vetigastropoda. As already stated by Koken (1889), Murchisonia-like gastropods commonly develop an anterior siphonal canal, a character that is absent in true Pleurotomariida. This character also brings the entire group closer to caenogastropods. On the other hand, Frýda and Manda (1997) and Frýda et al. (2008) documented well-preserved Murchisonia-like gastropods that have a protoconch typical of Vetigastropoda (i.e., consisting of about one whorl) suggesting placement in Vetigastropoda. The presence of nacre in the Ordovician “Murchisonia” (Mutvei 1983) further corroborates such a placement at least of early members assigned to this group. Murchisonioidea, in its present composition, is likely polyphyletic and holds members of both, Vetigastropoda and Caenogastropoda. There is no modern phylogenetic analysis including shell microstructure and larval shell characters of Murchisonia-like gastropods; hence, the importance of these characters in inferring the phylogeny of Murchisonioidea are not yet assessed.
There are several Triassic holdovers of high-spired, selenizone-bearing gastropods. To date, six of these Murchisonia-like genera comprising a total of 30 nominate species have been reported from the Triassic: “Murchisonia”, Pseudomurchisonia, Cheilotomona, Trypanocochlea, Wannerispira, and Vistilia. Batten (1973) and Hallam and Wignall (1997) stated that Murchisonioidea became extinct at the end-Triassic mass extinction event. Based on new material and the re-study of type material of several species, we discuss the diversity of Triassic Murchisonia-like gastropods, their link to late Paleozoic taxa, and revise their classification.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: http://zoobank.org/urn:lsid:zoobank.org:pub:3B3057AC-62DC-4B94-A557-5A4099947A6F.
Institutional abbreviations.—GBA, Geologische Bundesanstalt, Vienna, Austria; GPIT-PV, former Geologisch-paläontologisches Institut Tübingen (PV, Petrefaktenverzeichnis), now Senckenbergische Sammlungen in Tübingen, Germany; MUSE, Museo delle Scienze, Trento, Italy; NHMW, Naturhistorisches Museum Wien, Vienna, Austria; SNSB-BSPG, Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany.
Systematic palaeontology
Subclass Caenogastropoda Cox in Knight et al., 1960
Family Goniasmatidae Nützel and Bandel, 2000
Included genera: Laschmaspira Mazaev, 2003, and Altadema Kues, 2002; these genera have been placed in Orthonematidae. However, they clearly possess a selenizone and hence they are placed in Goniasmatidae. Pseudomurchisonia Koken, 1896, has only a weak sinus in the upper whorl portion; it is possible that it would develop into a selenizone. Therefore, it is tentatively placed in Goniasmatidae. Murchisonietta Nützel, Kaim, and Grădinaru, 2018, a subjective younger synonym of Wortheniopsis Böhm, 1895, has also been placed tentatively in Goniasmatidae (Nützel et al. 2018).
Genus Laschmaspira Mazaev, 2003
Type species: Laschmaspira rara Mazaev, 2003, Pennsylvanian, Russia.
Remarks.—To date, Laschmaspira has been known only from a few Pennsylvanian and Permian species (Mazaev 2003). Here we place two Late Triassic species in this genus: Laschmaspira euglypha Koken, 1896 comb. nov. (previously Murchisonia euglypha Koken, 1896), and Laschmaspira lirata sp. nov., both from the Feuerkogel, Austria. Laschmaspira lirata sp. nov. and the type species of Laschmaspira have a spiral ornament on the teleoconch whorls. However, Laschmaspira euglypha Koken, 1896, seems to lack spiral ornament.
Stratigraphic and geographic range.—Permian (Asselian) of Southern Fergana (Uzbekistan), middle Carboniferous of Central Russia– Carnian (Upper Triassic).
Laschmaspira lirata sp. nov.
Figs. 1, 2.
Zoobank LCID: urn:lsid:zoobank.org:act:361FE73E-A4F1-4E81-A9A 2-042103EE645F
Etymology: From Latin lirata, ridged; for the spiral lirae on the teleoconch,
Type material: Holotype, NHMW 2023/0080/0001, well preserved shell with protoconch. Paratypes, NHMW 2023/0080/0002–0007, well preserved shells. All from the typical Hallstatt Limestone facies, calcite pseudomorphoses covered by a reddish stain of ferruginous material.
Type locality: Feuerkogel, south slope, near Röthelstein und Tellschen, near Kainisch, Steiermark, Austria.
Type horizon: Hallstatt Limestone, Carnian (Upper Triassic).
Material.—Type material only.
Description.—Shell high-spired; holotype consists of about 9 whorls, 15.4 mm high, 7.9 mm wide; first four whorls smooth, convex, dome-shaped with an apical angle of 70–80°; initial whorl 0.33–0.36 mm wide; clear ontogenetic boundaries (embryonic/larval/teleoconch) not visible due to preservation or possibly gradual; later teleoconch with an apical angle of ca. 30°; teleoconch whorls markedly convex, angulated at borders of selenizone, with a broad ramp; selenizone forming from third whorl onwards as an increasingly deepening sinus of growth lines; selenizone on mature whorls broad, concave with lunulae of dense, distinct growth lines; late teleoconch whorls with numerous, distinct spiral lirae above and below selenizone; suture deep; base convex, anomphalous, demarcated from whorl face by an angulation, with numerous spiral threads.

Fig. 1. Goniasmatid caenogastropod Laschmaspira lirata sp. nov., Carnian, Feuerkogel, Austria. A. NHMW 2023/0080/0001, holotype, in lateral view (A1), detail of last whorls in lateral view (A2), detail adapical whorl face with spiral lirae in lateral view (A3), detail selenizone (slit-band) in lateral view (A4), detail of whorl face and base of last whorl with spiral lirae in lateral view (A5), early whorls in lateral view (A6, A7), early whorls in oblique lateral view (A8, A 9), detail of early whorls in oblique lateral view showing beginning of selenizone as a sinus (A10), early whorls in apical view (A11).

Fig. 2. Goniasmatid caenogastropod Laschmaspira lirata sp. nov., Carnian, Feuerkogel, Austria. A. NHMW 2023/0080/0002, paratype, in lateral view (A1), early whorls in lateral (A2), oblique lateral (A3), and apical (A4, A5) views. B. NHMW 2023/0080/0003, paratype, in lateral view (B1), detail of whorl face with selenizone (slit-band) and ornamentation of spiral lirae in oblique lateral (B2) and lateral (B3) views.
Remarks.—The most similar Triassic species is Laschmaspira euglypha (Koken, 1896), from the same locality (Feuerkogel). However, this species lacks spiral ornament, its whorls are more angulated, and the selenizone starts after the first whorl according to Koken (1896, 1897). Laschmaspira lirata sp. nov. exhibits a marked ontogenetic change regarding apical angle and ornament from broad, smooth, slitless initial whorls to ornamented much more slender and slit-bearing later whorls. However, a clear indication of a larval shell of the planktotrophic type could not be found. The size of the initial whorl (>0.3 mm) is rather large and suggests non-planktotrophic larval development (see Nützel 2014). Laschmaspira lirata sp. nov. can be regarded as a descendant of Paleozoic Goniasmatidae. The Permian Goniasma gerasimovi Mazaev, 2019, resembles Laschmaspira lirata sp. nov. in having an ornament of numerous spiral lirae which is otherwise unusual for Goniasma. However, G. gerasimovi differs in having a submedian angulation as is typical for Goniasma.
Stratigraphic and geographic range.—Type horizon and locality only.
Genus Pseudomurchisonia Koken, 1896
Type species: Pseudomurchisonia insueta Koken, 1896, Austria, Norian (Upper Triassic); subsequent designation by Cossmann (1897).
Remarks.—Pseudomurchisonia was tentatively placed in Purpurinidae (Caenogastropoda) by Wenz (1939: 526) who erroneously considered P. woehrmanni Koken, 1896, to be the type species. Thus, the genus was not included in the Treatise (Knight et al. 1960) which covered only few caenogastropods. Both originally included species, Pseudomurchisonia insueta Koken, 1896, and P. woehrmanni Koken, 1896, come from the Carnian (Upper Triassic) of the Feuerkogel in Austria. Both are moderately high-spired and have a broad selenizone that appears late during ontogeny. Apart from the two originally included species, two Early Triassic species were placed in Pseudomurchisonia: P. extracta (Berger, 1860) and P.? sundaica Krumbeck, 1924, but none from the Paleozoic and the post-Triassic.
The type species Pseudomurchisonia insueta (see below) differs from Goniasma in lacking an angulation, but resembles this genus in having a selenizone low on the whorls and lacking strong ornament. We therefore place this genus tentatively in Goniasmatidae. More knowledge about the early ontogeny of Pseudomurchisonia insueta is needed for a better substantiated systematic placement of this genus.
Pseudomurchisonia extracta (Berger, 1860) (originally placed in Natica) is based on an obviously poorly preserved specimen (probably a steinkern) from the Schaumkalk, a lithological unit of the Muschelkalk in the German Basin. The type is therefore of Anisian age. It shows a markedly gradate spire with a ramp bordered by an edge high on the whorls. This species has been placed in Pseudomurchisonia by Wittenburg (1908a: 284, pl. 5: 7) who illustrated a specimen with evenly rounded whorls and a median selenizone from the Werfen Formation of the Valsugana, Italy. Wittenburg’s (1908a) specimen is certainly not conspecific with Bergers’s (1860) type specimen. The Early Triassic specimen reported by Wittenburg (1908a) could either be a member of Pleurotomariida or Goniasmatidae. Baumgarte and Schulz (1986) and Kramm (2004) placed Pseudomurchisoni extracta in Angularia; their illustration of specimens from Hessen indicates that this assignment is more correct. Pseudomurchisonia extracta is probably a nomen dubium but its type specimens must be studied.
Pleurotomaria triadica Benecke, 1868, and Pleurotomaria euomphala Benecke, 1868, from the Lower Triassic of Mount Zacon at Borgo Valsugana (Italy) were placed in Pseudomurchisonia by Wittenburg (1908b), but need re-study of the types for confirmation.
Pseudomurchisonia schmidti Wittenburg, 1908b, is a nomen nudum.
Pseudomurchisonia? sundaica Krumbeck, 1924, from the Lower Triassic is actually close to the type species Pseudomurchisonia insueta Koken, 1896, and could be congeneric.
Stratigraphic and geographic range.—Lower Triassic, Timor and Carnian (Upper Triassic), Austria.
Pseudomurchisonia insueta Koken, 1896
Fig. 3.
1896 Pseudomurchisonia insueta Koken; Koken 1896: 86, fig. 12.
1897 Pseudomurchisonia insueta Koken; Koken 1897: 106, fig. 31.
2021 Pseudomurchisonia insinueta [sic!] Koken; Karapunar and Nützel 2021: 52.
Material.—Teleoconch fragment, NHMW 1926/0002/0461, Carnian (Upper Triassic), Feuerkogel, Austria.
Remarks.—We illustrate a single specimen (teleoconch fragment) from the Carnian of the Feuerkogel, Austria, representing the type species Pseudomurchisonia insueta, housed at the NHMW. The spire whorls are convex with an edge low on the whorls that is reduced later on. The earliest preserved whorl has neither a sinus nor a selenizone but straight, slightly opisthocyrt growth lines. Then a sinus and eventually a selenizone develops distinctly below mid-whorl. The selenizone is flush with the whorl face and smooth with sinuous growth lines. The whorls are smooth except of comarginal furrows in some whorls. The base is strongly convex and anomphalous. The transition from whorl face to base is evenly rounded.
The present specimen agrees well with the description and illustrations of Pseudomurchisonia insueta as given by Koken (1896, 1897). The apical whorls including the protoconch and the shell microstructure are unknown. Hence the higher classification of Pseudomurchisonia is doubtful. As already noted by Karapunar and Nützel (2021) and as discussed below, Acutitomaria woehrmanni (the other species originally included in Pseudomurchisonia by Koken 1896) clearly does not represent the Pseudomurchisonia.
Stratigraphic and geographic range.—Carnian (Upper Triassic), Feuerkogel, Austria.
Subfamily Cheilotomoninae nov.
Zoobank LCID: urn:lsid:zoobank.org:act:D1C49498-95D9-45A5-AF 5D-12762FA25B99
Included genera: Cheilotomona Strand, 1928, Trypanocochlea Tomlin, 1931, Vistilia Koken, 1896.
Type genus: Cheilotomona Strand, 1928.
Diagnosis.—High-spired shells with slit band and a median whorl angulation; early teleoconch whorls have three spiral cords, which in later whorls become subsutural cord, whorl angulation and basal cord; nodes on whorl angulation before development of selenizone; selenizone develops late in ontogeny (4th–7th whorl); the lower border of selenizone or the selenizone itself situated at whorl angulation.
Remarks.—We place the subfamily Cheilotomoninae in Goniasmatidae due to their high-spired shell with a median selenizone. Species with a multi-whorled protoconch of the planktotrophic type with sinusigera or a paucispiral protoconch with sinusigera (characteristic of goniasmatids) have not been documented in the members of Cheilotomoninae. Cheilotomona has smooth early whorls with fluent transition to the teleoconch and a large initial indicating non-planktotrophic early ontogeny (Karapunar and Nützel 2021, and see below).
Cheilotomona, Trypanocochlea, and Vistilia were previously placed in Murchisoniidae but Murchisonia lacks a prominent whorl crest on which the selenizone is situated, and this brings these genera close to Goniasma and thus they are placed in Gonismatidae. The protoconch of Trypanocochlea cerithioides is unknown and therefore, the possibility that Trypanocochlea belongs to Pleurotomariida and not to the caenogastropod family Goniasmatidae cannot be excluded. On the other hand, general whorl morphology, surface ornament (subsutural cord and strong axial ribs), and early ontogenetic shell development (nodular periphery, subsutural cord) supports its close association to Cheilotomona. Nützel and Nakazawa (2012) showed that a Permian Trypanocochlea species from Japan has a mammilated protoconch of the caenogastropod type supporting a placement in Goniasmatidae.
Genus Cheilotomona Strand, 1928
Type species: Pleurotoma blumi Wissmann in Münster, 1841, Carnian (Upper Triassic), St. Cassian Formation, South Tyrol, Italy; subsequent designation by Diener (1926).
Remarks.—Replacement name for Cheilotoma Koken, 1889, a junior homonym (see also Karapunar and Nützel 2021).
Stratigraphic and geographic range.—Anisian (Middle Triassic)–Carnian (Upper Triassic), Italy, Austria, Romania, Poland, Hungary, China.
Cheilotomona blumi (Wissmann in Münster, 1841)
Fig. 4.
1841 Pleurotoma blumi Wissmann n. sp., Wissmann in Münster: 123, pl. 13: 47a–c
For exhaustive synonymy an chresonymy see Karapunar and Nützel (2021)
Remarks.—A detailed description of this abundant and variable species and other three Cheilotomona species from the St. Cassian Formation was given by Karapunar and Nützel (2021). Cheilotomona blumi has about two smooth initial whorls with an indistinct transition to the teleoconch and a diameter of the first whorl of about 0.3 mm indicating an early ontogeny without planktotrophic larval development. The selenizone develops very late. Mature whorls have an angulated periphery at which the selenizone is situated.
Bandel (2006: 92) stated that “if Cheilotomona blumi was found in Paleozoic deposits, it could be placed with the genus Goniasma” and that “Cheilotomona represents the last known member of the Orthonematidae” (he considered Goniasmatidae and Orthonematidae to be synonymous, but see Nützel and Pan 2005 for a different opinion). Goniasma might be related to but is certainly not synonymous with Cheilotomona which has a strong axial ornament on early teleoconch whorls that is absent in Goniasma. Karapunar and Nützel (2021) placed Cheilotomona in the family Goniasmatidae Nützel and Bandel, 2000 (superfamily Orthonematoidea Nützel and Bandel, 2000).
Stratigraphic and geographic range.—Only known from the St. Cassian Formation, Carnian (Upper Triassic), N Italy.
Genus Vistilia Koken, 1896
Type species: Vistilia klipsteini Koken, 1896, Austria, Carnian (Upper Triassic); subsequent designation by Cossmann (1897).
Stratigraphic and geographic range.—Anisian (Middle Triassic)–Norian (Upper Triassic), Austria, Italy, Turkey.
Vistilia salisburgensis (Schnetzer, 1934)
Fig. 5.
1934 Stuorella salisburgensis n. sp.; Schnetzer 1934: 35, pl. 1: 12, 13.
2021 Vistilia salisburgensis (Schnetzer, 1934); Karapunar and Nützel 2021: 6, 60.
Material.—A total of 8 specimens from the Anisian (Middle Triassic), Saalfelden, Salzburg, Austria: SNSB-BSPG AS XXXIV 557, holotype, original of Schnetzer 1934: pl. 1: 12, teleoconch fragment of ca. three whorls, parts of shell preserved; paratypes: SNSB-BSPG AS XXXIV 558 (original of Schnetzer 1934: pl. 1: 12), SNSB-BSPG AS XXXIV 559 (five specimens), SNSB-BSPG AS XXXIV 560.
Description.—Fragments of a high-spired shell; whorls low with strong angulation above abapical suture; adapical whorl face steeply sloping, slightly concave, largely smooth with fine prosocline growth lines, faint spiral striation, and distinct spiral cord below adapical suture; selenizone concave at adapical border, distinctly convex near abapical border, forming periphery, smooth or with faint lunulae; whorl face below selenizone narrow, concave, sharply inclining adaxially, with prosocyrt growth lines; base flat, with opisthocyrt growth lines, only partly preserved.
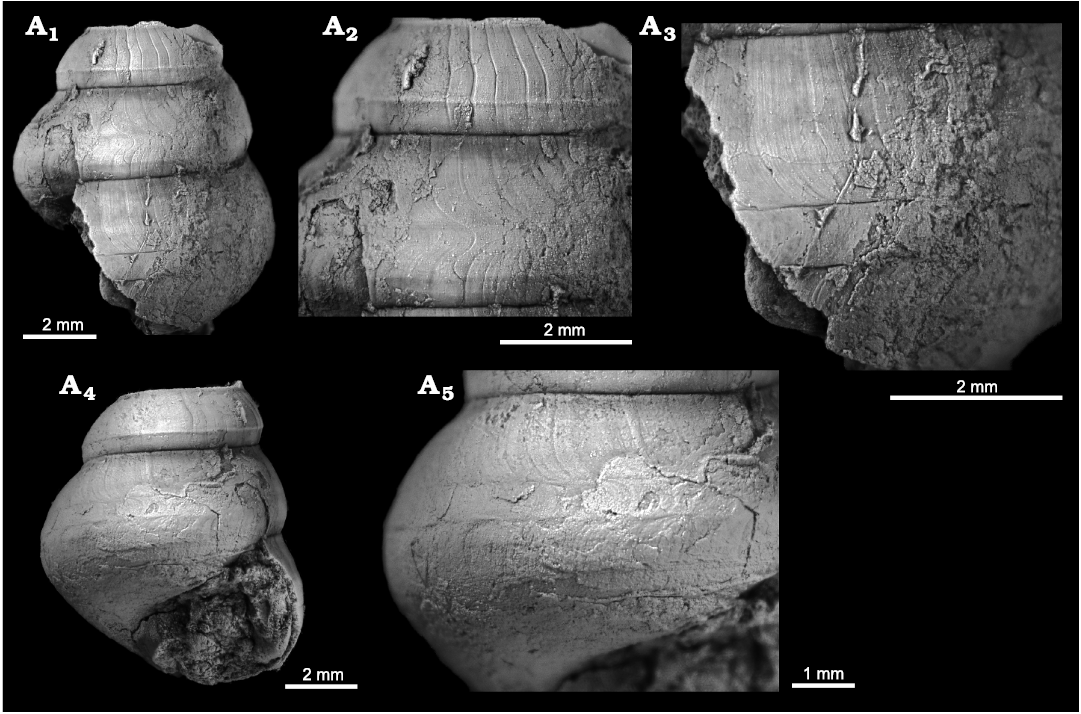
Fig. 3. Goniasmatid caenogastropod Pseudomurchisonia insueta Koken, 1896, Carnian, Feuerkogel, Austria. A. NHMW 1926/0002/0461, in lateral view (A1), detail of spire whorls in lateral view showing lack of selenizone in adapical whorl, and beginning of selenizone in abapical whorl (A2), detail of last part of last preserved whorl with fully developed selenizone (slit-band) in lateral (A3) and apertural lateral (A4) views, detail of last preserved whorl in lateral view (A5).

Fig. 4. Goniasmatid caenogastropod Cheilotomona blumi (Wissmann in Münster, 1841), Carnian, St. Cassian Formation, Italy. A. MB.Ga.4307.2, in lateral view (A1), early whorls in lateral view (A2), detail of whorl face with selenizone (slit-band) on crest in lateral view (A3). B. MB.Ga.4304.1, in lateral view (B1), early whorls including smooth protoconch in lateral view (B2), early whorls including smooth protoconch in oblique lateral view (B3).

Fig. 5. Goniasmatid caenogastropod Vistilia salisburgensis (Schnetzer, 1934), Anisian, Saalfelden, Salzburg, Austria. A. SNSB-BSPG AS XXXIV 557, holotype and original of Schnetzer (1934: pl. 1: 12), in lateral view (A1), detail of last preserved whorl in lateral view (A2, A3), detail of last preserved whorl in lateral view to show growth lines and selenizone (slit-band) on crest (A4). B. SNSB-BSPG AS XXXIV 559, paratype, in lateral view (B1), basal view (B2), detail of last preserved whorl in lateral view to show growth lines and selenizone (slit-band) on crest (B3).
Remarks.—As previously stated by Karapunar and Nützel (2021), Schnetzer’s (1934) original assignment to Stuorella cannot be maintained because this genus does not have a whorl angulation with a convex selenizone. Instead, Stuorella has a selenizone on flat whorl face somewhat above basal edge of the whorls. Vistilia salisburgensis has the characteristic teleoconch morphology of Vistilia with low whorls. However, protoconch and early teleoconch of Vistilia salisburgensis are unknown. Vistilia salisburgensis closely resembles Cheilotomona tristriata (Münster, 1841) from the lower Carnian St. Cassian Formation (Karapunar and Nützel 2021: fig. 86) in general whorl morphology, surface ornament and in having a pronounced subsutural spiral cord. The presence of a single subsutural cord is a feature also shared by Cheilotomona (Fig. 4) and Trypanocochlea (Fig. 6). Vistilia salisburgensis is the oldest known Triassic species of this genus. Vistilia last appeared in the Carnian (Koken 1896).
Stratigraphic and geographic range.—Known from the type locality only, see above.
Genus Trypanocochlea Tomlin, 1931
Type species: Verania cerithioides Koken, 1896, Carnian (Upper Triassic), Austria; original designation.
Remarks.—Trypanocochlea is a replacement for Verania Koken, 1896.
Stratigraphic and geographic range.—Middle Permian, Japan–Norian (Upper Triassic), Austria.
Trypanocochlea cerithioides (Koken, 1896)
Fig. 6.
1896 Verania cerithioides Koken; Koken 1896: 86, text-fig. 11.
1897 Verania cerithioides Koken; Koken 1897: 105, text-fig. 29.
Material.—Lectotype, herein designated, SNSB-BSPG 1878 XI B 478 (original of Koken 1896: text-fig. 11; original of Koken 1897: text-fig. 29), Carnian (Upper Triassic), Röthelstein, Aussee, Austria. NHMW 1858/0047/0088, SNSB-BSPG 2011 XXXIX 78d, Carnian (Upper Triassic), Feuerkogel, Austria.
Description.—Shell high-spired, apical angle ca. 45°; type specimen consists of ca. 8 whorls, 7.8 mm high, 3.4 wide; earliest preserved whorls rounded, smooth, developing an angulation low on whorl face after one-two whorls; whorl face strongly angulated slightly below middle of whorl face; selenizone broad, situated on angulation, forming periphery; selenizone forms late (after the second preserved whorl), ornamented with pronounced node-forming lunulae, and a median spiral cord; lunulae closely spaced in early whorls, widely spaced in later whorls; selenizone bordered by outward bending shell forming sharp edges (spiral cords); whorl face steeply sloping above selenizone, ornamented with strengthened growth lines and spiral thread in subsutural zone; whorl face below selenizone sharply inclined adaxially; transition from whorl face to conical, convex base demarcated by spiral cord, accompanied by one or few spiral threads; aperture trapezoidal; inner lip convex with about 12 spiral lirae (folds).

Fig. 6. Goniasmatid caenogastropod Trypanocochlea cerithioides (Koken, 1896), Carnian, Austria. A. SNSB-BSPG, lectotype, 1878 XI B 478 and original of Koken (1896: text-fig. 11) and Koken (1897: text-fig. 29), Röthelstein, Aussee, in lateral view (A1), early whorls in lateral view (A2), aperture with folds in lateral view (A3). B. SNSB-BSPG 2011 XXXIX 78d, Feuerkogel, in lateral view (B1), early whorls in lateral view (B2). C. NHMW 1858/0047/0088, Feuerkogel, in lateral view (C1), detail of last preserved whorl in lateral view to show growth lines and selenizone on crest (C2), middle whorl in lateral view (C3), detail of last preserved whorl in oblique lateral view to show growth lines and selenizone on crest (C4).
Remarks.—Trypanocochlea cerithioides, type species of Trypanocochlea, is a murchisonimorph gastropod with a strongly ornamented, convex selenizone. Its inner lip has spiral lirae (folds) unlike any other members of Goniasmatidae. A middle Permian representative of Trypanocochlea, has been reported by Nützel and Nakazawa (2012) but that species seemingly lacks folds on the inner lip (maybe due to preservation). Similar gastropods (also lacking folds on the inner lip) have also been reported from the Pennsylvanian/lower Permian of the USA: Taosia crenulata (Girty, 1939) (Kues 1991, 2004a, b). Taosia crenulata differs much from the type species of Taosia, and at first glance this late Paleozoic species resembles Trypanocochlea. However, while the selenizone of Taosia crenulata is situated on upper whorl face above the nodular periphery, the selenizone of Trypanocochlea cerithioides is situated on the angular periphery and the nodes are formed within the selenizone. Cheilotomona subgranulata (Klipstein, 1844) from the St. Cassian Formation develops similar nodular selenizone on whorl periphery, but species of Cheilotomona lack the folds on inner lip. The only other report of Trypanocochlea (an unnamed species) is from the ?upper Norian/Rhaetian (Upper Triassic) of Peru by Haas (1953), who reported absence of folds in the Peruvian species. Due to the unclear age if this probably last occurrence of Trypanocochlea, the exact timing of the exticntion of this genus is unknown.
Stratigraphic and geographic range.—Known from the type locality area only, see above.
Subfamily uncertain
Genus Altadema Kues, 2002
Type species: Altadema convexa Kues, 2002, Pennsylvanian, USA.
Remarks.—The type species of Altadema has a small, moderately high-spired largely smooth shell with strongly convex whorls; its selenizone is situated high on whorl. So far, four Pennsylvanian species have been assigned to this genus (Kues 2002; Mazaev 2003). Herein, we place the Early Triassic species Altadema kokeni (Wittenburg, 1908a) and Altadema hausmannae sp. nov. in Altadema. Altadema hausmannae has a broad slit-band above the periphery in mature whorls whereas Altadema kokeni (Wittenburg, 1908a) has a sinus or selenizone high on the whorls.
The Triassic species Wannerispira shangganensis Kaim and Nützel in Kaim et al. 2010, closely resembles Altadema but has a more pronounced selenizone situated lower on the whorls.
Stratigraphic and geographic range.—Upper Carboniferous, USA and Russia–Lower Triassic, Italy.
Altadema hausmannae sp. nov.
Fig. 7.
2005 Pseudomurchisonia kokeni Wittenburg, 1908; Nützel and Schulbert 2005: 497, fig. 16.
Zoobank LCID: urn:lsid:zoobank.org:act:CA52C5A2-AF63-458F-BB B6-AD0A705260DD
Etymology: Dedicated to Imelda M. Hausmann for her work on Triassic marine biota.
Type material: Holotype, MUSE-PAL 8176, specimen of ca. four whorls lacking apex, with attached rock matrix. Paratypes, nine specimens: MUSE-PAL 8177–8185.
Type locality: 750 m south of Cimirlo, near the road from Cimirlo to Busa del Vent, where road crosses creek in a narrow curve; GPS (WGS 84): 46°03’50.5’’ N, 11°10’58.0’’ E; Trentino, North Italy.
Type horizon: Werfen Formation, Gastropod Oolite Member, Lower Triassic, probably Dienerian/Smithian transition (Hofmann et al. 2015).
Material.—Type material; in addition numerous specimens in rock-forming quantities.
Description.—Largest specimen comprises ca. six whorls (apex missing), 4.9 mm high, 3.0 mm wide, turbiniform with variable apical angle of ca. 60–80°; whorls markedly convex, separated by distinct sutures; whorls with more or less pronounced subsutural shelf so that spire is gradate in most specimens; shell smooth (but studied specimens covered by a reddish crust of iron oxides or hydroxides that may obscure ornament if ornament was present); ramp demarcated by the adapical edge of broad selenizone; transition from ramp to lateral whorl face may be angular; selenizone situated high on the whorls; base convex with an umbilical chink; transition from whorl face to base evenly rounded.
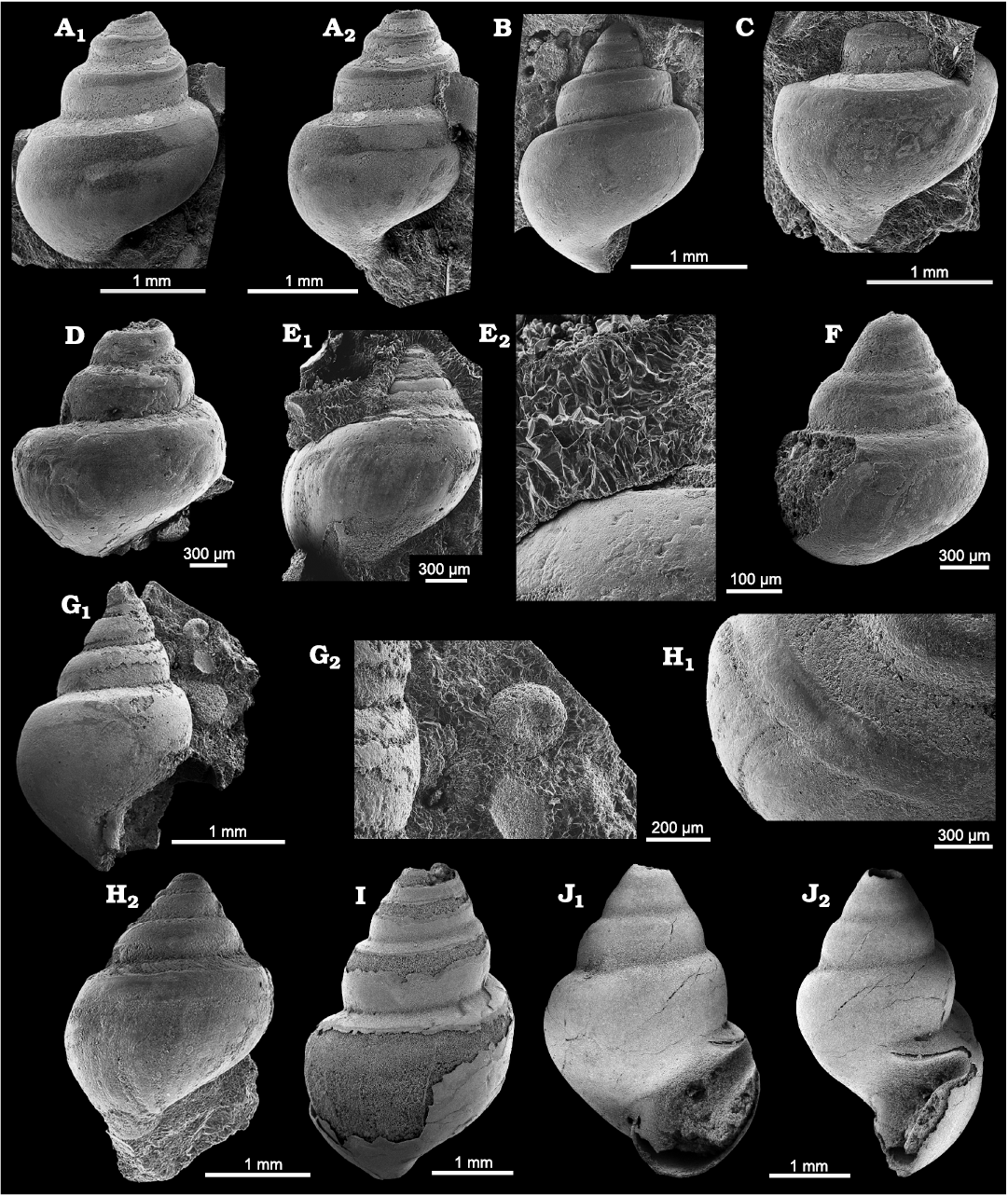
Fig. 7. Goniasmatid caenogastropod Altadema hausmannae sp. nov., Dienerian/Smithian transition, Lower Triassic, Werfen Formation, Gastropod Oolite Member, Italy. A. MUSE-PAL 8176, holotype, lateral view. B. MUSE-PAL 8177, paratype, lateral view. C. MUSE-PAL 8178, paratype, in lateral view. D. MUSE-PAL 8179, paratype, in lateral view. E. MUSE-PAL 8180, paratype, in lateral view (E1), detail (E2). F. MUSE-PAL 8181, paratype, in lateral view. G. MUSE-PAL 8182, paratype, in lateral view (G1), detail (G2). H. MUSE-PAL 8183, paratype, detail to show slit-band, in oblique lateral view (H1) and specimen in lateral view (H2). I. MUSE-PAL 8184, paratype, in lateral view. J. MUSE-PAL 8185, paratype, in apertural (J1) and lateral (J2) views.
Remarks.—Altadema hausmannae sp. nov. occurs in rock-forming quantities at its type locality (Nützel and Schulbert 2005). Nützel and Schulbert (2005) identified this species as Pseudomurchisonia kokeni Wittenburg, 1908a (also from the Lower Triassic Werfen Formation) but a closer examination of the holotype of P. kokeni reveals that they are not conspecific (see below). Altadema kokeni has only a slight sinus, not a selenizone. Otherwise both species resemble each other closely. Altadema hausmannae differs widely from the type species Pseudomurchisonia insueta in whorl morphology and position of selenizone. The other species of Altadema (A. convexa Kues, 2002, A. cryptocarina Mazaev, 2003, A. altadema Mazaev, 2003, A. cryptocarina Mazaev, 2003, and A. lira Mazaev, 2003, all Pennsylvanian) are more slender and the spire is less gradate (having a larger angle between sutural shelf and outer whorl face).
Stratigraphic and geographic range.—Known from the type locality only, see above.
Altadema kokeni (Wittenburg, 1908a) comb. nov.
Fig. 8.
1908a Pseudomurchisonia kokeni n. sp.; Wittenburg 1908a: 16, text-fig. 1; pl. 2: 1.
non 2005 Pseudomurchisonia kokeni Wittenburg, 1908; Nützel and Schulbert 2005: 497, fig. 16.
Material.—The holotype, GPIT-PV-108710 (original of Wittenburg 1908a: pl. 5: 9, 10). Wittenburg (1908a) reported it from the Campiler Schichten (upper Werfen Formation, Lower Triassic) at the Col di Rodella, near the Sella Pass in the Dolomites, North Italy.
Description.—The holotype comprises about four whorls, is 4.8 mm high and 3.5 mm wide. It is turbiniform with an apical angle of ca. 65°. The whorls are markedly convex and separated by deep sutures. The shell is smooth and shiny except for fine growth lines. The growth lines are oblique prosocline below the adapical suture, form a shallow sinus above the periphery and almost orthocline prosocyrt between periphery and abapical suture.

Fig. 8. Goniasmatid caenogastropod Altadema kokeni (Wittenburg, 1908a) and others, Dienerian/Smithian transition, Lower Triassic, Werfen Formation, Gastropod Oolite Member, Italy. A. Crack surface of Gastropod Oolite, near Borgo, Valsugana, Italy, (“Monte Zaccon” locality of Wittenburg 1908b); gastropods (Altadema kokeni and “Polygyrina gracilior”) as well as other clasts covered with iron oxide; Wittenburg’s (1908a, b) material, collection of University of Tübingen (Nützel and Schulbert 2005: fig. 15). B. Altdema kokeni (Wittenburg, 1908a), GPIT-PV-108710, holotype, and original of Wittenburg (1908a: pl. 5: 9, 10), Col di Rodella, in lateral view, not covered by ammonium chloride to show reddish color (B1), detail of last whorl in lateral view to show adapical sinus of growth lines (B2), in lateral view (B3). C. Original drawing of Altdema kokeni from Wittenburg (1908a).
Remarks.—The preservation is good when compared with the otherwise mostly poor preservation of the Werfen gastropods. The shell is replaced by a sparitic, slightly reddish calcite. The specimen is embedded in a hard calcareous rock. It is therefore likely that it comes from the gastropod oolite because other fossiliferous rocks of the Werfen Formation are usually more marly. It resembles Altadema hausmannae sp. nov. in general shape and by having a smooth shell, but it lacks a true selenizone; instead it has a sinus high on the whorls. It is possible that this specimen represents a juvenile shell and that a selenizone would develop later during ontogeny.
Stratigraphic and geographic range.—Known from the type locality only, see above.
?Family Goniasmatidae Nützel and Bandel, 2000
Genus Wortheniopsis Böhm, 1895
[= Murchisonietta Nützel, Kaim, and Grădinaru, 2018]
Type species: Pleurotomaria margarethae Kittl, 1894, Anisian (Middle Triassic), Marmolada Limestone, Italy; type by monotypy.
Emended diagnosis.—Shell moderately high-spired to fusiform; whorls evenly rounded or weakly angulated; whorls embracing somewhat below periphery; surface smooth or with spiral ornament; sinus or shallow slit high on whorl face, which may form a weak angulation; sinus/slit appears very late in ontogeny.
Remarks.—Wortheniopsis was erected by Böhm, 1895, for Worthenia-like shells (placed in Pleurotomariidae) lacking prominent shell angulations. The growth lines are not well-preserved or documented for the type species W. margarethae and hence the presence of a slit-band (selenizone) was only assumed by Kittl (1894) and Böhm (1895). We studied material from the Marmolada Limestone (Figs. 9, 10) and found that Wortheniopsis margarethae (Fig. 9) lacks a true selenizone (slit-band). Instead, growth lines have a backwards projection above mid-whorl of mature whorls forming a shallow sinus. The same pseudo-selenizone has been reported for the Anisian genus Murchisonietta Nützel, Kaim, and Grădinaru, 2018 (Fig. 11) and therefore we consider Wortheniopsis and Murchisonietta to represent synonyms. Similar to Altadema kokeni, Wortheniopsis also develops a sinus high on the whorls in late ontogeny. At this point, the higher classification of Wortheniopsis is unclear because it lacks a true selenizone, its shell microstructure and early whorls including protoconch are unknown.
Wenz (1938: 128) included Wortheniopsis in Pleurotomariidae and Lophospirinae. Knight et al. (1960) placed relatively high-spired taxa (but not as high as murchisoniids) including Wortheniopsis in the murchisonioid family Plethospiridae, which composes taxa ranging from the Ordovician to lower Carboniferous (for instance Platyzona, formerly included in Plethospiridae is regarded as Goniasmatidae, see Karapunar et al. 2022) and is likely a not monophyletic group in its current composition. Nützel et al. (2018) placed the Middle Triassic Murchisonietta (which is considered a junior synonym of Wortheniopsis herein) tentatively in Goniasmatidae and at present, we maintain this arrangement. We consider Wortheniopsis to represent Caenogastropoda, close to Goniasmatidae, but more evidence for that placement is needed, especially from early ontogenetic shell morphology.
There are many erroneous attributions of Permian and Jurassic species to Wortheniopsis.
The species attributed to Wortheniopsis by Szabó (2008) were later correctly removed from that genus and placed in Kericserispira Szabó, 2018 [replacement name for the pre-occupied Faveolaria Szabó, 2016]. Unlike Wortheniopsis, the following two Jurassic species have a clearly developed selenizone and therefore they are removed from Wortheniopsis and placed in Kericserispira Szabó, 2018: Wortheniopsis bakonyensis Szabó, 2016 [Kericserispira bakonyensis (Szabó, 2016) comb. nov.]; Wortheniopsis rakusi Szabó, 2016 [Kericserispira rakusi (Szabó, 2016) comb. nov.]
Branson (1948) listed many Permian species that were assigned to Wortheniopsis and some have been assigned to other genera since then. Wortheniopsis netschajewi Jakowlew, 1899 and Wortheniopsis burtasorum (Golowkinsky, 1868) were placed in Biarmeaspira by Mazaev (2015). Specimens previously identified by Licharew (1967) as Pleurotomaria (Wortheniopsis) sequens Waagen, 1880, were placed in Baylea by Mazaev (2016, 2018). Pleurotomaria (Wortheniopsis?) eulkaiensis Reed, 1927, was placed into Platyzona by Nützel and Nakazawa (2012). Worthenopsis? [sic!] depressa Beede, 1907, is a lenticular shell and was later placed in Cyclites by Knight (1940). Worthenopsis [sic!] sp. by Beede (1907) is a doubtful record and probably does not represent Wortheniopsis.
The Permian Worthenopsis [sic!] bicarinata Branson, 1930, is herein placed tentatively in Glabrocingulum (Ananias).
The following Permian species have a strongly angulated whorls face and a smooth selenizone. Hence, they are tentatively placed in Baylea Koninck, 1883, herein: Worthenopsis dejactinensis Jakowlew, 1899, Worthenopsis kyschertianaeformis Jakowlew, 1899, Worthenopsis grandicarinata Jakowlew, 1899, Worthenopsis denjatinensis Jakowlew, 1899. The placement of the following species in Wortheniopsis are doubtful; they probably represent Ananias Knight, 1945, or Baylea Koninck, 1883:
Wortheniopsis sequens (Waagen, 1880)
Wortheniopsis orientalis (Krotow, 1885)
Wortheniopsis kyschertiana (Stuckenberg, 1898)
Wortheniopsis permiana (Stuckenberg, 1898)
Wortheniopsis jakovlewi Stuckenberg, 1905
Wortheniopsis pachti Stuckenberg, 1905
Wortheniopsis volgensis Stuckenberg, 1905
Wortheniopsis sysranicus Stuckenberg, 1905
The Triassic Wortheniopsis sp. reported by Sha (1995) and Wortheniopsis budensis Kutassy, 1927, are doubtful records. At present, there are only three accepted species of Wortheniopsis (from the Anisian and Ladinian): Wortheniopsis margarethae (Kittl, 1894), Wortheniopsis quirini (Stoppani, 1860), and Wortheniopsis acuta (Nützel, Kaim, and Grădinaru, 2018) (Fig. 11).
Stratigraphic and geographic range.—Anisian (Middle Triassic)–Norian (Upper Triassic), Italy, Romania, Hungary.
Wortheniopsis margarethae (Kittl, 1894)
Fig. 9.
1894 Pleurotomaria margarethae Kittl n.f.; Kittl 1894: 114, pl. 6: 1–3.
1895 Wortheniopsis margarethae Kittl sp.; Böhm 1895: 220, text-fig. 6; pl. 14: 24.
Material.—Two specimens, NHMW 1884/0006/0872/1, 2, from the Anisian (Middle Triassic), Marmolada Limestone, Italy.
Description.—Shell moderately high-spired, acutely conical, with four preserved whorls (earlier whorls missing), 15 mm high, 10 mm wide; whorls high, with convex to slightly angulated profile and incised suture; whorls above sinus steeply sloping, with prosocline growth lines and faint subsutural spiral cords; sinus situated on faint angulation, high on whorl face; whorl face below sinus with prosocyrt growth lines and wide, faint spiral bands; transition to base rounded; base anomphalous, ornamented with wide imbricated spiral bands.
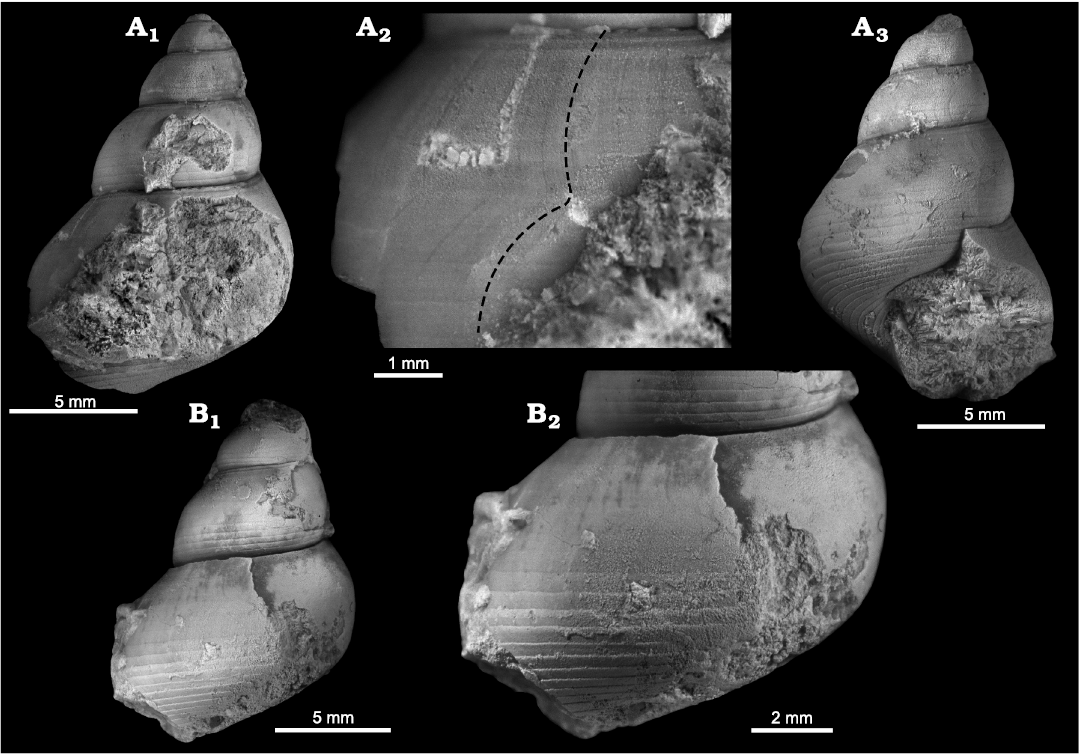
Fig. 9. Goniasmatid? caenogastropod Wortheniopsis margarethae (Kittl, 1894), Ladinian, Marmolada Limestone, Italy. A. NHMW 1884/0006/0872/1, in lateral (A1) and apertural (A3) views, detail of last preserved whorl in lateral view (A2) to show growth lines (course indicated with dashed line). B. NHMW 1884/0006/0872/2, in lateral view (B1), detail of last preserved whorl in lateral view (B2).
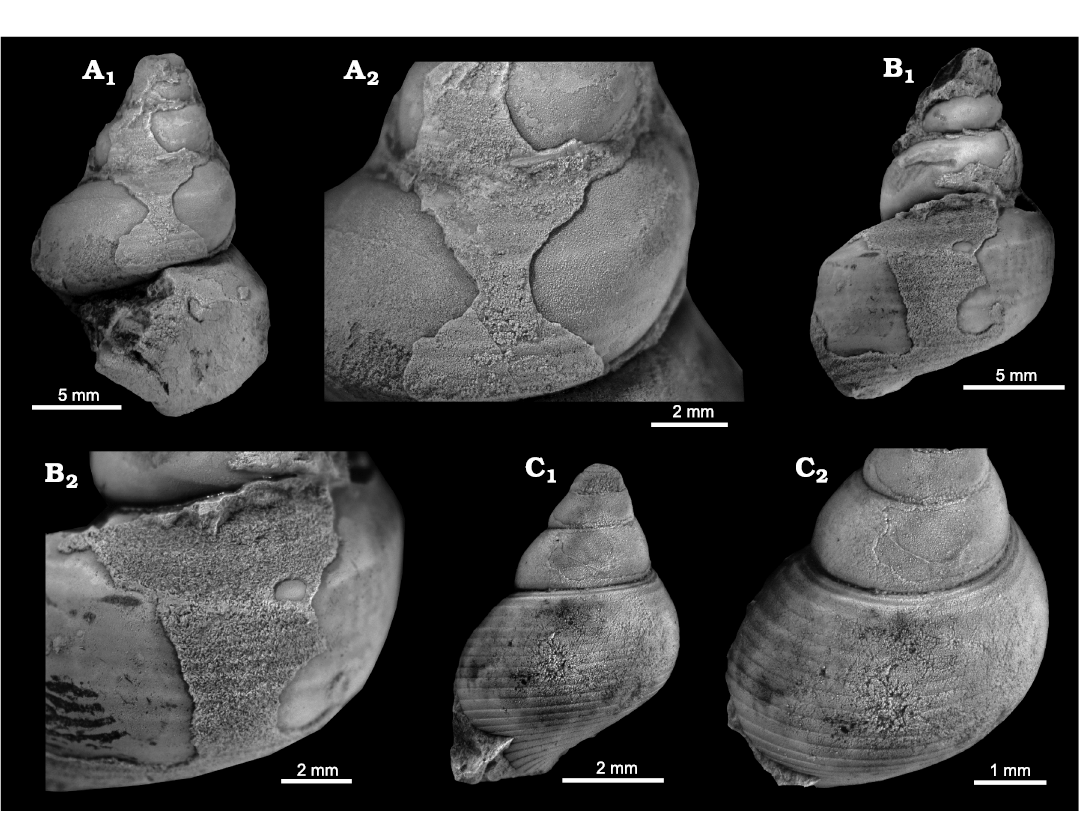
Fig. 10. Goniasmatid? caenogastropod Wortheniopsis quirini (Stoppani, 1860), Ladinian, Marmolada Limestone, Italy. A. NHMW 1969/1102/0000/1, original of Koken (1899, pl. 1: 14), in lateral view (A1), detail of last preserved whorl in lateral view (A2). B. NHMW 1969/1102/0000/2, original of Koken (1899: pl. 1: 15), in lateral view (B1), detail of last preserved whorl in lateral view (B2). C. NHMW 2019/0177/0020, juvenile specimen, in lateral (C1) and oblique lateral (C2) views.
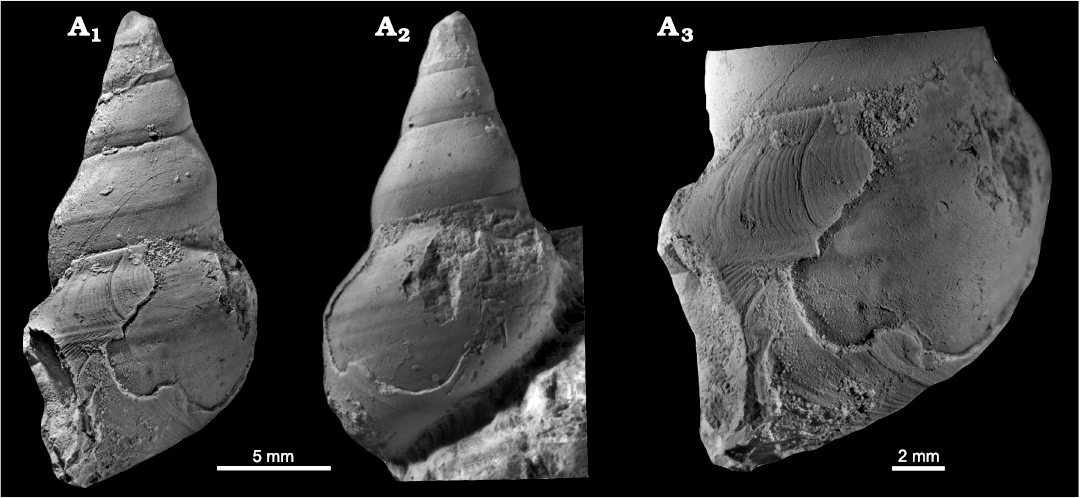
Fig. 11. Goniasmatid? caenogastropod Wortheniopsis acuta (Nützel, Kaim, and Gradinaru, 2018) comb. nov., Anisian, Romania. A. SNSB-BSPG 2015 VI 32, holotype, in lateral view (A1,A2), detail of last preserved whorl in lateral view (A3).
Remarks.—The specimens placed in Wortheniopsis quirini (Stoppani, 1860) by Kittl (1899) are figured herein (Fig. 10). Their whorls are more angulated, and although poorly preserved, they show the characteristic imbricated spiral bands.
Stratigraphic and geographic range.—Ladinian (Middle Triassic), Marmolada, Italy.
Triassic gastropods erroneously identified as Murchisonia-like gastropods
Subclass Vetigastropoda Salvini-Plawen, 1980
Order Pleurotomariida Cox and Knight in Knight et al., 1960
Family Lancedelliidae Bandel, 2009
Genus Acutitomaria Karapunar and Nützel, 2021
Type species: Acutitomaria kustatscherae Karapunar and Nützel, 2021, Carnian (Upper Triassic), St. Cassian Formation, South Tyrol, Italy; original designation.
Remarks.—This genus unites exceptionally high-spired Pleurotomariida with late developing selenizone and a pronounced cancellate ornamentation (see Karapunar and Nützel 2021 for emended diagnosis of the family Lancedelliidae).
Acutitomaria woehrmanni (Koken, 1896) comb. nov.
Fig. 12.
1896 Pseudomurchisonia woehrmanni Koken; Koken 1896: 87, fig. 13.
1897 Pseudomurchisonia woehrmanni Koken; Koken 1897: 106, fig. 30, pl. 6: 3.
2021 Pseudomurchisonia woehrmanni Koken; Karapunar and Nützel 2021: 51.
Material.—Four fragments from the GBA, three of which were photographed and might represent Koken’s (1896) illustrated type specimen that is broken (GBA 1897/003/0001). The label says “Unterer Rötelstein”, in the publications (Koken 1896, 1897), the Feuerkogel is given as locality. GBA 1897/003/0001 is designated herein as lectotype.
Description.—Shell moderately high-spired, apical angle ca. 60°; early whorls not well-preserved but earliest whorls seem low trochospiral; whorls distinctly convex, ornamented with strong, densely spaced axial ribs and few spiral cords; axial ribs become knobby when crossing spiral cords in lower part of whorls; axial ribs become visible on third whorl where they are straight prosocline; selenizone appears within 4th whorl, high on whorl face, flush to slightly concave with strong lunulae; in addition, selenizone has a fine ornament of converging spiral threads (so far unknown from this species); base convex with strong spiral cords and axial ribs with small and axially elongated nodes at intersections.
Stratigraphic and geographic range.—Carnian (Upper Triassic), Austria.

Fig. 12. Pleurotomariid vetigastropod Acutitomaria woehrmanni (Koken, 1896) “Unterer Rötelstein”, according to Koken (1896, 1897), Feuerkogel, Austria is given as locality. A. GBA 1897/003/0001, fragmented lectotype, early whorls in lateral view (A1), detail of whorl face in lateral view showing ornament, selenizone with lunulae and converging micro-ornament (A2), early whorls in oblique lateral (A3) and apical (A4) views, last whorl in lateral views (A5, A6).
Discussion
High-spired slit-band gastropods are an important component of late Paleozoic gastropod faunas. A considerable number of Permian (some of them latest Permian) species have been reported in the last decades (Winters 1963; Pan and Erwin 2002; Nützel and Nakazawa 2012; Nützel and Pan 2005; Mazaev 2006, 2015, 2018, 2019; Karapunar et al. 2022; Ketwetsuriya et al. 2016, 2020a, b, 2021). At least 27 Permian Murchisonia-like genera of the family Goniasmatidae have been reported (Table 1).
As has been elaborated in the introduction of this contribution, several of these Murchisonia-like gastropods proved to represent caenogastropods based on their protoconch morphology, the absence of nacre, their high-spired shape, and the presence of siphonal canals. We have shown that this group of caenogastropods apparently survived the end-Permian mass extinction event. On the other hand, high-spired Pleurotomariida were also present in that period of time (Karapunar and Nützel 2021) and Acutitomaria is shown as being a Late Triassic example for that.
Triassic Murchisonia-like gastropods are a small but heterogeneous group as our study has shown. Previously, Triassic murchisonimorph species have been attributed to the genera Murchisonia, Pseudomurchisonia, Cheilotomona, Trypanocochlea, Wannerispira, and Vistilia. We add the genera Laschmaspira, Altadema, and Wortheniopsis (alias Murchisonietta) (Table 1). Trypanocochlea, Wannerispira, Laschmaspira, and Altadema originate in the Paleozoic and persist into the Triassic. Pseudomurchisonia, Wortheniopsis, Cheilotomona, and Vistilia are restricted to the Triassic. However, the four Triassic species assigned to Murchisonia (M. euglypha Koken, 1894, M. sera Böhm, 1895, M. subeuglypha Krumbeck, 1924, and M. timorensis Krumbeck, 1924) likely do not represent this genus. Murchisonia euglypha Koken, 1894, is likely a member of the Laschmaspira (see above), Murchisonia sera Böhm, 1895, is a very slender, small form with a median slit-band and a pronounced ornament of spiral cords. It certainly does not belong in Murchisonia. Its generic assignment needs to be revised, it possibly represents Stegocoelia or an undescribed genus. Murchisonia timorensis also strongly deviates from the type species of Murchisonia, as does M. subeuglypha that as a strong angulation low on the whorls and additional strong spiral cords.
Table 1. Permian and Triassic Murchisonia-like gastropod genera (Goniasmatidae).
| |
Permian |
Triassic |
|
Aclisina Koninck, 1881 |
× |
|
|
Akasakiella Nützel in Nützel and Nakazawa, 2012 |
× |
|
|
Altadema Kues, 2002 (Carboniferous and Triassic) |
× |
× |
|
Arribazona Kues, 1990 |
× |
|
|
Cerithioides Haughton, 1859 |
× |
|
|
Cerithiozone Nützel in Nützel and Nakazawa, 2012 |
× |
|
|
Cheilotomona Strand, 1928 |
|
× |
|
Cibecuia Winters, 1956 |
× |
|
|
Costataenia Nützel in Nützel and Nakazawa, 2012 |
× |
|
|
Donaldospira Batten, 1966 |
× |
|
|
Erwinispira Nützel and Pan, 2005 |
× |
|
|
Goniasma Tomlin, 1930 |
× |
|
|
Helicospira Girty, 1915 |
× |
|
|
Hypergonia Donald, 1892 |
× |
|
|
Kazanella Mazaev, 2015 |
× |
|
|
Laschmaspira Mazaev, 2003 |
× |
× |
|
Loxisonia Batten, 1985 |
× |
|
|
Metorthonema Erwin, 1988 |
× |
|
|
Micheliopsis Termier and Termier in Termier et al., 1977 |
× |
|
|
Peruvispira Chronic, 1949 |
× |
|
|
Platyzona Knight, 1945 |
× |
|
|
?Pseudomurchisonia Koken, 1896 |
|
× |
|
Stegocoelia Donald, 1889 |
× |
|
|
Taosia Girty, 1939 |
× |
|
|
Trypanocochlea Tomlin, 1931 |
× |
× |
|
Vebericochlis Licharew, 1967 |
× |
|
|
Vistilia Koken, 1896 |
|
× |
|
Wannerispira Kaim and Nützel in Kaim et al., 2010 |
× |
× |
|
?Wortheniopsis Böhm, 1895 |
|
× |
The Triassic Wortheniopsis lacks a true slit-band and have instead a more or less pronounced sinus. Therefore its assignment to Goniasmatidae is tentative; it might represent a new caenogastropod family.
Thus, at the generic level, there seems to be a considerable turnover in Murchisonia-like gastropods at the Paleozoic/Mesozoic transition according to the current state of knowledge. At the family level, it seems that only Goniasmatidae survived into the Triassic. Murchisonia-like gastropods have been reported from the Lower Triassic to the Norian but no clear Rhaetian species have been identified (species from the Carnian/Norian of Timor need a better dating; commonly only “Late Triassic” is given as age in the literature) and representatives are absent in the Jurassic. Amongst Murchisonia-like gastropods only Trypanocochlea has been considered by Ferrari and Hautmann (2022) who restricted their analysis of gastropod diversity to genera from the Norian to the Pliensbachian. They found that the number of genera declined considerably at the Triassic/Jurassic boundary. The standing gastropod diversity declined considerably from the Carnian to the Rhaetian and was very low in the Rhaetian (Nützel 2005; Karapunar and Nützel 2021). Murchisonia-like gastropods are absent in all important Rhaetian faunas with the possible exception of Trypanocochlea sp. from the Pucará Group, Peru. The single specimen in question is from Lot 48, and Haas (1953: 307) indicated that this lot has an age from the “Late Norian or Early Rhaetian” to the Rhaetian, thus it is unclear whether that specimen has a Norian or Rhaetian age. Generally, there are only few species-rich Rhaetian gastropod faunas (e.g., Haas 1953; Nützel and Senowbari-Daryan 1999; Nützel et al. 2022). It is well possible that most of the Murchisonia-like gastropods became extinct at the mid-Carnian crisis (Carnian Pluvial Event) and were thus already extinct prior to the end-Triassic event. Slit-bearing vetigastropods (Pleurotomariida) were also selectively affected by the mid-Carnian crisis (Karapunar and Nützel 2021).
Conclusions
Murchisonia-like caenogastropods, chiefly Goniasmatidae, were diverse and abundant until the Permian, barely survived the end Permian extinction, regained a certain generic diversity within the Triassic with the evolution of several new genera but failed by far to regain their Permian generic diversity. Thus, this once successful and diverse group shares a similar fate (surviving the end-Permian extinction, a reduced Triassic diversity and extinction during Late Triassic crises) as conodonts, orthoceratids, conulariids, and others. This diversity pattern does not qualify for the “Dead Clade Walking” phenomenon i.e., the extinction shortly after a major mass extinction event (survival without recovery) as outlined by Jablonski (2002) because they have survived for ca. 30 Ma (at least until the Norian) and even produced a number of new genera. Among gastropods bellerophontids would be a typical example for “Dead Clade Walking”, they survived the end-Permian extinction but eventually became extinct within the Early Triassic, at the end of the Smithian (Kaim and Nützel 2011). In both cases, bellerophontids and Murchisonia-like gastropods, the marginalization at end-Permian extinction event increased the vulnerability of the groups and likelihood for subsequent complete extinction. Both groups are slit-band gastropods and their dwindling that is accentuated by extinction events is part of a long-term overall decline of slit-band gastropods (see also Karapunar and Nützel 2021). Obviously, there prevailed a long-term selective trend against the character “shell-slit”. It is likely that Goniasmatidae and similar forms are related to the ancient caenogastropod clade Cerithioidea (e.g., Nützel 1998; Nützel and Pan 2005) and in so far, this evolutionary lineage still persists until today but this once successful group has ceased at the genus- and family-level. The elimination of shell slits in caenogastropods probably also reflects an evolutionary change of the ventilation of the mantle cavity in these caenogastropods. The exhalant flow went in a lateral or dorsal direction in Murchisonia-like caenogastropods and that changed to a more ventral or posterior direction in modern Cerithioidea.
Acknowledgements
We thank Stefano Monari (Università di Padova, Italy) and Mariel Ferrari (Instituto Patagónico de Geología y Paleontología CCT CONICET-CENPAT, Puerto Madryn, Argentina) for their thoughtful reviews. Ingmar Werneburg (University of Tübingen, Germany) and Irene Zorn (GBA) are acknowledged for giving access to palaeontological collections under their care. Alexander Lukeneder, Andreas Kroh, and Thomas Nichterl (all NHMW) are acknowledged for providing working space, accommodation and access to the NHMW collections. The Deutsche Forschungsgemeinschaft (DFG) is acknowledged for the financial support (DFG NU 96/14-1, DFG NU 96/14-2). This research received support from the SYNTHESYS Project (http://www.synthesys.info/), which is financed by a European Community Research Infrastructure Action under the FP7 “Capacities” Program (AT TAF-1797) and provided a grant to AN for a collection visit to NHMW and GBA.
References
Bandel, K. 2002. Reevaluation and classification of Carboniferous and Permian Gastropoda belonging to the Caenogastropoda and their relation. Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg 86: 81–188. Crossref
Bandel, K. 2006. Families of the Cerithioidea and related superfamilies (Palaeo-Caenogastropoda; Mollusca) from the Triassic to the Recent characterized by protoconch morphology – including the description of new taxa. Freiberger Forschungshefte C 511: 59–138.
Bandel, K. 2009. The slit bearing nacreous Archaeogastropoda of the Triassic tropical reefs in the St. Cassian Formation with evaluation of the taxonomic value of the selenizone. Berliner paläobiologische Abhandlungen 10: 5–47.
Bandel, K., Nützel, A., and Yancey, T.E. 2002. Larval shells and shell microstructures of exceptionally well-preserved Late Carboniferous gastropods from the Buckhorn Asphalt deposit (Oklahoma, USA). Senckenbergiana letheae 82: 639–690.
Batten, R.L. 1966. The Lower Carboniferous gastropod fauna from the Hotwells Limestone of Compton Martin, Somerset, Part 2. Palaeontographical Society Monographs 120 (513): 53–109 Crossref
Batten, R.L. 1973. The vicissitudes of the gastropods during the interval of the Guadalupian-Ladinian time. In: A. Logan and L.V. Hill (eds.), The Permian and Triassic Systems and their Mutual Boundary. Canadian Society of Petroleum Geologists Memoir 2: 596–607.
Batten, R.L. 1985. Permian gastropods from Perak, Malaysia. Part 3. The murchisoniids, cerithiids, loxonematids, and subulitids. American Museum Novitates 2829: 1–40.
Baumgarte, D. and M. Schulz. 1986. Stratigraphie und Fauna des Unteren und Mittleren Wellenkalkes (Unteranis/ Pelson) von Müs (Bl. 5423 Großenlüder). Geologisches Jahrbuch Hessen 114: 69–94.
Beede, J.W. 1907. Invertebrate paleontology of the Upper Permian red beds of Oklahoma and the panhandle of Texas. Kansas University Science Bulletin 4 (3): 113–171. Crossref
Benecke, E.W. 1868. Ueber einige Muschelkalk-Ablagerungen der Alpen. Geognostisch-Paläontologische Beiträge 2 (1): 1–67.
Berger, H.A.C. 1860. Die Versteinerungen des Schaumkalks am Thüringer-Wald. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde 1860: 196–206.
Böhm, J. 1895. Die Gastropoden des Marmolatakalkes. Palaeontographica 42: 211–308.
Branson, C.C. 1930. Paleontology and Stratigraphy of the Phosphoria Formation. The University of Missouri; A Quaterly of Research 5 (2): 5–66, 90–99.
Branson, C.C. 1948. Bibliographic index of Permian invertebrates. Geological Society of America Memoir 26: 1–1049. Crossref
Chronic, B.J. 1949. Part 2. Invertebrate paleontology (excepting fusulinids and corals). In: N.D. Newell, B.J. Chronic, and T.G. Roberts (eds.),Upper Paleozoic of Peru, 46–173. Columbia University Press, New York. [Reissued as Geological Society of America Memoir 58, 1953].
Cossmann, M. 1897. Die Gastropoden der Trias um Hallstatt, von E. Koken (1). Revue critique Paléozoologie 1: 139–143.
Dechen, H. von 1832. Handbuch der Geognosie von H.T. De La Beche. Nach der zweiten Auflage des Engl. Originals bearbeitet von H. von Dechen. 612 pp. Mit 23 eingedrückten Holzschnitten. Duncker & Humblot, Berlin.
Diener, C. 1926. Fossilium Catalogus, I: Animalia, Pars 34, Glossophora triadica. 242 pp. W. Junk, Berlin.
Donald, J. 1889. Descriptions of new species of Carboniferous gastropods. Quarterly Journal of the Geological Society of London 51: 210–234. Crossref
Donald, J. 1892. Notes on some new and little-known species of Carboniferous Murchisonia. Quarterly Journal of the Geological Society of London 68: 562–575. Crossref
Erwin, D.H. 1988. Permian Gastropoda of the Southwestern United States: Cerithiacea, Acteonacea, and Pyramidellacea. Journal of Paleontology 62: 566–575. Crossref
Erwin, D.H. 1990. Carboniferous–Triassic gastropod diversity patterns and the Permo-Triassic mass extinction. Paleobiology 16: 187–203. Crossref
Ferrari, M. and Hautmann, M. 2022. Gastropods underwent a major taxonomic turnover during the end-Triassic marine mass extinction event. Plos One 17 (11): e0276329. Crossref
Frýda, J. and Manda, S. 1997. A gastropod faunule from the Monograptus uniformis graptolite biozone (Early Lochkovian, Early Devonian) in Bohemia. Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg 80: 59–121.
Frýda, J., Nützel, A., and Wagner, P.J. 2008. Paleozoic Gastropoda. In: W.F. Ponder and D.R. Lindberg (eds.), Phylogeny and Evolution of the Mollusca, 239–270. University of California Press, Berkely. Crossref
Girty, G.H. 1915. The fauna of the Wewoka Formation of Oklahoma. United States Geological Survey Bulletin 544: 1–353.
Girty, G.H. 1939. Certain pleurotomariid gastropods from the Carboniferous of New Mexico and Texas. Journal of the Washington Academy of Sciences 29 (1): 21–36.
Golowkinsky, N.A. [Golovkinski, N.A.] 1868 On the Permian formation in the central part of the Kama-Volga Basin [in Russian]. Materialy dlâ geologii Rossii 1: 1–146.
Haas, O. 1953. Mesozoic invertebrate faunas of Peru. Bulletin of the American Museum of Natural History 101: 1–328.
Hallam, A. and Wignall, P.B. 1997. Mass Extinctions and Their Aftermath. 320 pp. Oxford University Press, Oxford.
Haughton, S. 1859. On some fossil Pyramidellidae from the Carboniferous Limestone of Cork and Clonmel. Proceedings of the Dublin University Zoology and Botany Association 1: 281–283.
Heidelberger, D. 2001. Mitteldevonische (givetische) Gastropoden (Mollusca) aus der Lahnmulde (südliches Rheinisches Schiefergebirge). Geologische Abhandlungen Hessen 106: 1–291.
Heidelberger, D. and Koch, L. 2005. Gastropoda from the Givetian „Massenkalk“ of Schwelm and Hohenlimburg (Sauerland, Rheinischel Schiefergebirge, Germany). Geologica et Palaeontologica, Sonderband 4: 1–63.
Hofmann, R., Hautmann, M., and Bucher, H. 2015. Recovery dynamics of benthic marine communities from the Lower Triassic Werfen Formation, northern Italy. Lethaia 48: 474–496. Crossref
Jablonski, D. 2002. Survival without recovery after mass extinctions. PNAS 99: 8139–8144. Crossref
Jakowlew, N. 1899. Die Fauna einiger oberpalaeozoischer Ablagerungen Russlands. I. Die Cephalopoden und Gastropoden. Memoires du Comite Geologique 15 (3): 1–140.
Kaim, A. and Nützel, A. 2011. Dead bellerophontids walking—The short Mesozoic history of the Bellerophontoidea (Gastropoda). Palaeogeography, Palaeoclimatology, Palaeoecology 308: 190–199. Crossref
Kaim, A., Nützel, A., Bucher, H., Brühwiler, T., and Goudemand, N. 2010. Early Triassic (Late Griesbachian) gastropods from South China (Shanggan, Guangxi). Swiss Journal of Geosciences 103: 121–128. Crossref
Karapunar, B. and Nützel, A. 2021. Slit-band gastropods (Pleurotomariida) from the Upper Triassic St. Cassian Formation and their diversity dynamics in the Triassic. Zootaxa 5042 (1): 1–165. Crossref
Karapunar, B., Nützel, A., and Ketwetsuriya, C. 2022. A low-diversity Peruvispira-dominated gastropod assemblage from the Permian Ratburi Group of Central Thailand. Alcheringa: 147–155. Crossref
Karapunar, B., Nützel, A., Seuss, B., and Mapes, R.H. 2022. Taxonomy and diversity of slit-band gastropods (order Pleurotomariida) and some slit bearing Caenogastropoda from the Pennsylvanian of the USA. Papers in Palaeontology 8 (2): e1417. Crossref
Ketwetsuriya, C., Cook, A. G., and Nützel, A. 2020a. Permian gastropods from the Ratchaburi Limestone, Khao Phrik, Central Thailand. PalZ 90: 53–77. Crossref
Ketwetsuriya, C., Hausmann, I.M., and Nützel, A. 2021. Diversity patterns of Middle Permian gastropod assemblages from the Tak Fa Formation, Central Thailand. Palaeobiodiversity and Palaeoenvironments 101: 907–925. Crossref
Ketwetsuriya, C., Karapunar, B., Charoentitirat, T., and Nützel, A. 2020b. Middle Permian (Roadian) gastropods from the Khao Khad Formation, Central Thailand: Implications for palaeogeography of the Indochina Terrane. Zootaxa 4766 (1): 1–47. Crossref
Ketwetsuriya, C., Nützel, A., and Kanjanapayont, P. 2016. Permian gastropods from the Tak Fa Limestone, Nakhon Sawan, Northern Thailand. Bulletin of Geosciences 91: 481–513. Crossref
Kittl, E. 1894. Die triadischen Gastropoden der Marmolata und verwandter Fundstellen in den weißen Riffkalken Südtirols. Jahrbuch der kaiserlich-königlichen geologischen Reichsanstalt 44: 99–182.
Kittl, E. 1899. Die Gastropoden der Esinokalke, nebst einer Revision der Gastropoden der Marmolatakalke. Annalen des kaiserlich königlichen naturhistorischen Hofmuseums 14: 1–237.
Klipstein, A. von 1843–1845. Beitrag zur geologischen Kenntnis der östlichen Alpen. 1–144, pls. 1–9 (1843); 145–240, pls. 10–15 (1844); 241–311, pls. 16–20 (1845). G.F. Heyer’s Verlag, Gießen.
Knight, J.B. 1940. Gastropods of the Whitehorse Sandstone. Geological Society of America Bulletin 51: 302–315.
Knight, J.B. 1945. Some new genera of Paleozoic Gastropoda. Journal of Paleontology 19: 573–587.
Knight, J.B., Cox, L.R., Keen, A.M., Batten, R.L., Yochelson, E.L., and Robertson, R. 1960. Systematic descriptions. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology, Part 1, Mollusca 1, I169–I310. Geological Society of America and University of Kansas Press, Kansas.
Koken, E. 1889. Ueber die Entwicklung der Gastropoden vom Cambrium bis zur Trias. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, Beilageband 6: 440–463.
Koken, E. 1894. Die Gastropoden der Schichten mit Arcestes studeri. Jahrbuch der kaiserlich-königlichen geologischen Reichsanstalt 44: 441–458.
Koken, E. 1896. Die Gastropoden der Trias um Hallstatt. Jahrbuch der kaiserlich-königlichen geologischen Reichsanstalt 46: 37–126.
Koken, E. 1897. Die Gastropoden der Trias um Hallstatt. Abhandlungen der kaiserlich-königlichen geologischen Reichsanstalt 17 (4): 1–111.
Koninck, L.G. de 1881. Faune du calcaire carbonifère de la Belgique. Troisième partie, Gastéropodes. Annales du Musée Royal d´Histoire naturelle de Belgique 6: 1–170.
Koninck, L.G. de 1883. Faune du calcaire carbonifère de la Belgique. Quatrième partie, Gastéropodes (suite et fin). Annales du Musée Royal d’Histoire naturelle de Belgique 8: 1–240.
Kramm, E. 2004. Fauna der unteren Schaumkalkbank (Trias, Anis) von Rodges (Hessen). Beiträge zur Naturkunde in Osthessen 39: 73–82.
Krotow, P. 1885. Artinskische Etage. Geologisch-Palaeontologische Monographie des Sandsteines von Artinsk. Trudy, Obŝestvo Estestvoispuitatelei pri Imperatorskom Kazanskom Universitete 13: 1–314, pls. 1–4. [1–297 in Russian, 298–314 German summary]
Krumbeck, L. 1924. Die Brachiopoden, Lamellibranchiaten und Gastropoden der Trias von Timor II. Paläontologie von Timor 13: 38–90.
Kues, B.S. 1990. New and little known Pennsylvanian gastropods from the Flechado Formation, Taos City, New Mexico. In: P.W. Bauer, S.G. Lucas, C.K. Mawer, and W.C. McIntosh (eds.), Tectonic Development of the Southern Sangre de Cristo Mountains, New Mexico, 41st Annual Fall Field Conference Guidebook, 251–258. New Mexico Geological Society, Socorro . Crossref
Kues, B.S. 1991. Some gastropods from the lower Wolfcampian (Basal Permian) Laborcita Formation, Sacramento Mountains, New Mexico. In: J.M. Barker, B.S. Kues, G.S. Austin, and S.G. Lucas (eds.), Geology of the Sierra Blanca, Sacramento, and Capitan Ranges, New Mexico, 42nd Annual Fall Field Conference Guidebook, 221–230. New Mexico Geological Society, Socorro. Crossref
Kues, B.S. 2002. New genera and species of Middle Pennsylvanian gastropods from West Texas. Journal of Paleontology 76: 52–62. Crossref
Kues, B.S. 2004a. Marine invertebrate assemblages from the Late Pennsylvanian (Virgilian) Holder Formation, Dry Canyon, Sacramento Mountains, south-central New Mexico. New Mexico Geology 26: 43–53. Crossref
Kues, B.S. 2004b. Stratigraphy and brachiopod and molluscan paleontology of the Red Tanks Formation (Madera Group) near the Pennsylnanian-Permian boundary, Lucero uplift, west-central New Mexico. New Mexico Museum of Natural History and Science Bulletin 25: 143–210.
Kutassy, A. 1927. Beiträge zur Stratigraphie und Paläontologie der alpinen Triasschichten in der Umgebung von Budapest. Magyar Királyi Földtani Intézet Évkönyve 27: 107–175.
Licharew, B.K. [Ličarev, B.K.] 1967. Scaphopods and gastropods—Archaeogastropoda (excluding subord Bellerophontina and suborder Neritopsina) from the Upper Paleozoic of South Fergana [in Russian]. Trudy Vsesoûznogo Naučno-Issledovatel’skogo Geologičeskogo Instituta, Biostratigrafičeskij sbornik, Novaâ Seriâ 116: 1–115.
Mazaev, A.V. 2003. The family Orthonemidae (Gastropoda) from Middle and Upper Carboniferous of Central part of Russian Plate. Ruthenica 13: 89–100.
Mazaev, A.V. 2006. Permian gastropods from Kulogory Formation of northern Moscow Syneclise. Paleontological Journal 40: 42–53. Crossref
Mazaev, A.V. 2011. Pennsylvanian gastropods of the suborders Murchisoniina Cox et Knight, 1960 and Sinuspirina Mazaev subordo nov. from the central regions of the Russian Platform: Morphology, taxonomy, and phylogeny. Paleontological Journal 45: 1533–1599. Crossref
Mazaev, A.V. 2015. Upper Kazanian (Middle Permian) Gastropods of the Volga-Urals Region. Paleontological Journal 49: 869–986. Crossref
Mazaev, A.V. 2016. Evolution of the genus Baylea (Gastropoda) in the Kazan Paleobasin (Middle Permian, Volga-Ural Region). Paleontological Journal 50: 585–600. Crossref
Mazaev, A.V. 2018. Early Kazanian (Middle Permian) gastropods. Paleontological Journal 52: 796–827. Crossref
Mazaev, A.V. 2019. Lower Permian gastropods of Shakhtau (Asselian–Sakmarian Boundary Beds, Southern Cisuralia). Paleontological Journal 53: 1237–1345. Crossref
Münster, G. von 1841. Beschreibung und Abbildung der in den Kalkmergelschichten von St. Cassian gefundenen Versteinerungen. In: H.L. Wissmann, G. von Münster, and K.F. Braun (eds.), Beiträge zur Geologie und Petrefacten-Kunde des südöstlichen Tirol’s vorzüglich der Schichten von St. Cassian, 25–152. Buchner, Bayreuth.
Mutvei, H. 1983. Ultrastructural evolution of molluscan nacre. In: P. Westbroek and E.W. de Jong (eds.), Biomineralization and Biological Metal Accumulation, 267–271. Reidel Publishing Company, Dordrecht. Crossref
Nützel, A. 1998. Über die Stammesgeschichte der Ptenoglossa (Gastropoda). Berliner geowissenschaftliche Reihe E 26: 1–229.
Nützel, A. 2005. Recovery of gastropods in the Early Triassic. Comptes Rendus Palevol 4: 501–515. Crossref
Nützel, A. 2014. Larval ecology and morphology in fossil gastropods. Palaeontology 57: 479–503. Crossref
Nützel, A. and Bandel, K. 2000. Goniasmidae and Orthonemidae: two new families of the Palaeozoic Caenogastropoda (Mollusca, Gastropoda). Neues Jahrbuch für Geologie und Paläontologie Monatshefte 2000 (9): 557–569. Crossref
Nützel, A. and Nakazawa, K. 2012. Permian (Capitanian) gastropods from the Akasaka Limestone (Gifu Prefecture, Japan). Journal of Systematic Palaeontology 10: 103–169. Crossref
Nützel, A. and Pan, H.-Z. 2005. Late Paleozoic evolution of the Caenogastropoda: larval shell morphology and implications for the Permian/Triassic mass extinction event. Journal of Paleontology 79: 1175–1188. Crossref
Nützel, A. and Schulbert, C. 2005. Gastropod lagerstätten in the aftermath of the end-Permian mass extinction—Diversity and facies of two major Early Triassic occurrences. Facies 51: 495–515. Crossref
Nützel, A. and Senowbari-Daryan, B. 1999. Gastropods from the Upper Triassic (Norian–Rhaetian) Nayband Formation of central Iran. Beringeria 23: 93–132.
Nützel, A., Kaim, A., and Grădinaru, E. 2018. Middle Triassic (Anisian, Bithynian) gastropods from North Dobrogea (Romania) and their significance for gastropod recovery from the end-Permian mass extinction event. Papers in Palaeontology 4: 477–512. Crossref
Nützel, A., Nose, M., Hautmann, M., and Hochleitner, R. 2022. Latest Triassic (Sevatian–Rhaetian) reef carbonates from the Northern Calcareous Alps (Austria), their mollusc dwellers, and their fate at the end-Triassic extinction event. PalZ 97: 265–309. Crossref
Pan, H.-Z. and Erwin, D.H. 2002. Gastropods from the Permian of Guangxi and Yunnan provinces, South China. Journal of Paleontology. The Paleontological Society Memoir 76 (56): 1–49. Crossref
Ponder, W.F., Lindberg, D.R., and Ponder, J.M. 2020. Biology and Evolution of the Mollusca. Vols. 1–2. 870 pp. CRC Press, Boca Raton. Crossref
Reed, F.R.C. 1927. Palaeozoic and Mesozoic fossils from Yunnan. Memoirs of the Geological Survey of India. Palaeontologia Indica, New Series 10: 1–331.
Salvini-Plawen, L. von 1980. A reconsideration of systematics in the Mollusca. Malacologia 19: 249–278.
Schnetzer, R. 1934. Die Muschelkalkfauna des Öfenbachgrabens bei Saalfelden. Palaeontographica A 81: 1–159.
Sha, J. 1995. Palaeontology of the Hoh Xil region, Qinghai [in Chinese with English summary]. In: S. Jingeng (ed.), The Series of Comprehensive Scientific Expedition to the Hoh Xil Region. The Comprehensive Scientific Expedition to the Hoh Xil Region, 69–81. Science Press, Beijing.
Stoppani, A. 1858–60. Les pétrifications d’Ésino, ou Description des fossiles appartenant au dépôt triasique supérieur des environs d’Ésino en Lombardie. 151 pp. Joseph Bernardoni, Milan.
Strand, E. 1928. Miscellanea nomenclatorica zoologica et palaeontologica. Archiv für Naturgeschichte 92A: 30–75.
Stuckenberg, A. 1898. Allgemeine geologische Karte von Russland. Blatt 127. Memoires du Comite Geologique 16: 1–361.
Stuckenberg, A. 1905. Die Fauna der obercarbonischen Suite des Wolgadurchbruches bei Samara. Trudy Geologičeskogo Komiteta, novaâ seriâ 23: 11–140.
Sun, X., Qiu, X., Tihelka, E., Yang, H., Sun, D., Tong, J., and Tian, L. 2021. A diverse gastropod fauna from the shallow marine carbonate platform of the Yangou section (south China) in the immediate aftermath of the Permian–Triassic mass extinction. Geological Journal 56: 6199–6209. Crossref
Szabó, J. 2008. Gastropods of the Early Jurassic Hierlatz Limestone Formation; part 1: a revision of the type collections from Austrian and Hungarian localities. Fragmenta Palaeontologica Hungarica 26: 1–108.
Szabó, J. 2016. Gastropods of the Lower Jurassic Hierlatz Limestone Formation, part 2. Some new archaic type slit-bearing components from the fauna of the Hierlatz Alpe (Hallstatt, Austria) and the Bakony Mts (Hungary). Fragmenta Palaeontologica Hungarica 33: 3–30. Crossref
Szabó, J. 2018. Kericserispira nomen novum, a new substitute name instead of Foveolaria Szabó, 2017 (non Foveolaria Busk, 1884). Fragmenta Palaeontologica Hungarica 35: 85. Crossref
Termier, H., Termier, G., and Vachard, D. 1977. Monographie paléontologique des affleurements Permiens du Djebel Tébaga (sud Tunisien). Palaeontographica A 156: 1–109.
Tomlin, J.R.L.B. 1930. Some preoccupied generic names. Journal of Molluscan Studies 19: 22–24. Crossref
Tomlin, J.R.L.B. 1931. Some preoccupied generic names. Journal of Molluscan Studies 19 (4): 174–175. Crossref
Waagen, W. 1880. Salt-Range Fossils. I. Productus Limestone Fossils. 2. Pisces-Cephalopoda: Supplement Gasteropoda. Memoirs of the Geological Survey of India (Palaeontologia Indica) 13: 1–183.
Wenz, W. 1938–1944. Gastropoda. Teil I: Allgemeiner Teil und Prosobranchia. In: O.H. Schindewolf (ed.), Handbuch der Paläozoologie, Band 6, xii + 1639. Lieferung 1, 1–240 [March 1938]; 3, 241–480 [October 1938]; 4, 481–720 [July 1939]; 6, 721–960 [August 1940]; 7, 961–1200 [October 1941]; 8, 1201–1506 [October 1943]; 9, 1507–1639, i-xii [November 1944]. Gebrüder Borntraeger, Berlin.
Winters, S.S. 1956. New Permian gastropod genera from eastern Arizona. Journal of the Washington Academy of Sciences 46 (2): 44–45
Winters, S.S. 1963. Supai Formation (Permian) of Eastern Arizona. The Geological Society of America 89: 1–99. Crossref
Wittenburg, P. von 1908a. Einige neue Fossilien aus den Werfener Schichten Südtirols. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 1: 16–21.
Wittenburg, P. von 1908b. Neue Beiträge zur Geologie und Paläontologie der Werfener Schichten Südtirols, mit Berücksichtigung der Schichten von Wladiwostok. Centralblatt für Mineralogie, Geologie und Paläontologie 2: 67–89.
Yoo, E.K. 1988. Early Carboniferous Mollusca from Gundy, Upper Hunter, New South Wales. Records of the Australian Museum 40: 233–264. Crossref
Yoo, E.K. 1994. Early Carboniferous Gastropoda from the Tamworth Belt, New South Wales, Australia. Records of the Australian Museum 46: 63–110.Crossref
Acta Palaeontol. Pol. 68 (3): 539–559, 2023
https://doi.org/10.4202/app.01087.2023