
Late Ordovician scolecodonts and chitinozoans from the Pin Valley in Spiti, Himachal Pradesh, northern India
PETRA TONAROVÁ, THOMAS J. SUTTNER, OLLE HINTS, YAN LIANG, MAREK ZEMEK, MICHAL KUBAJKO, TOMÁŠ ZIKMUND, JOZEF KAISER, and ERIKA KIDO
Tonarová, P., Suttner, T.J., Hints, O., Liang, Y., Zemek, M., Kubajko, M., Zikmund, T., Kaiser, J., and Kido, E. 2024. Late Ordovician scolecodonts and chitinozoans from the Pin Valley in Spiti, Himachal Pradesh, northern India. Acta Palaeontologica Polonica 69 (2): 199–215.
The end of the Ordovician witnessed major perturbations in the ecosystem, seriously affecting global marine biodiversity. Nevertheless, some marine organism groups and their crisis-bound palaeogeographic distribution are still understudied. Among the outliers are eunicid polychaetes, even though they flourished and diversified extensively during the Ordovician. A collection of seven genera of jaw-bearing polychaetes, including the new ramphoprionid genus Spitiprion Tonarová, Suttner, & Hints, with type new species of Spitiprion khannai Tonarová, Suttner, & Hints, is described here from Katian (Upper Ordovician) deposits of Spiti, northern India. The new species is preserved as isolated maxillae and a jaw cluster, and 3D models of the maxillary apparatus are reconstructed based on submicron-CT. Along with the scolecodonts, a low-diversity assemblage of chitinozoans was recovered, comprising five genera. The most common chitinozoan species are Acanthochitina cf. cancellata and Spinachitina suecica.
Key words: Polychaeta, Eunicida, Ramphoprionidae, Chitinozoa, organic-walled microfossils, Early Palaeozoic, Gondwana, submicron-CT.
Petra Tonarová [petra.tonarova@geology.cz; ORCID: https://orcid.org/0000-0001-5002-6389 ], Thomas J. Suttner [thomas.suttner@geology.cz; ORCID: https://orcid.org/0000-0002-8803-7611 ], and Michal Kubajko [michal.kubajko @geology.cz; ORCID: https://orcid.org/0009-0005-7096-2584 ], Czech Geological Survey, Klárov 3/131, 11821 Praha 1, Czech Republic.
Olle Hints [olle.hints@taltech.ee; ORCID: https://orcid.org/0000-0003-4670-4452 ], Tallinn University of Technology, Ehitajate tee 5, 19086 Tallinn, Estonia.
Yan Liang [liangyan@nigpas.ac.cn; ORCID: https://orcid.org/0000-0002-2376-8202 ], Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, 39 East Bejing Road, Nanjing, China.
Marek Zemek [Marek.Zemek@ceitec.vutbr.cz; ORCID: https://orcid.org/0000-0002-3236-4111 ], Tomáš Zikmund [tomas.zikmund@ceitec.vutbr.cz; ORCID: https://orcid.org0000-0003-2948-5198 ], and Jozef Kaiser [jozef.kaiser@ceitec.vutbr.cz; ORCID: https://orcid.org/0000-0002-7397-125X ], CEITEC, Central European Institute of Technology, Brno University of Technology, Purkynova 656/123, 612 00 Brno, Czech Republic.
Erika Kido [kidoe@appstate.edu; ORCID: https://orcid.org/0000-0001-5130-8804 ], Department of Geological and Environmental Sciences, Appalachian State University, Boone, NC, 28608-2006, USA.
Received 11 January 2024, accepted 3 April 2024, published online 8 May 2024.
Copyright © 2024 P. Tonarová et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Ordovician Period was a time of prominent biotic changes. The increase in biodiversity during the Ordovician has often been called the “Great Ordovician Biodiversification Event” (GOBE) (Webby et al. 2004; Servais et al. 2010; Ontiveros et al. 2023). However, nowadays, there are various opinions on the characteristics and timing of Ordovician radiation. This widely used term has been re-evaluated as various groups of organisms were characterised by different diversification scenarios, and the process was not a short-lived event (Servais et al. 2023). Nevertheless, without a doubt, the end of the Ordovician was characterised by prominent changes in the atmosphere and oceans, including a period of glaciation (e.g., Brenchley et al. 2003) and a major perturbation in the carbon and sulphur cycling during the early Hirnantian (Schmidt et al. 2016) followed by a warming during the late Hirnantian (Bond and Grasby 2020). Just before the Hirnantian glaciation, another warming episode, the late Katian Boda Event, was documented through the palaeogeographical shifts in shelly faunas and the nature of carbonate precipitation (Fortey and Cocks 2005; Suttner et al. 2007a; Lefebvre et al. 2014; Kröger et al. 2016; Myrow et al. 2019; Lu et al. 2021). However, most of our data on these environmental and biotic events comes from Laurentia and Baltica. More information is needed from other paleogeographic domains, including Gondwana.
The fossil record of soft-bodied invertebrates like polychaetes is rather uneven. Although body fossils have been reported from Palaeozoic strata, many lack diagnostic features of living taxa; hence, their proper taxonomic assignment is often impossible. The bulk of the fossil record of polychaetes is represented by their resistant jaws, the scolecodonts. The oldest finds of scolecodonts date back to the latest Cambrian (Williams et al. 1999) and starting from the mid-Ordovician, they become widespread and diverse (Hints et al. 2017). The majority of preserved Early Palaeozoic jawed polychaetes belong to aciculate polychaetes, the Order Eunicida, a group of the subclass Errantia. They are characterised by the presence of reversible jaw apparatus, comprising ventral mandibles and dorsal maxillae in the ventral muscularised pharynx (Zanol et al. 2021). The Order Eunicida includes seven extant and 17 extinct families (Budaeva and Zanol 2021). Today, they are common in all types of marine environments (Fauchald 2013), inhabiting soft and hard substrates from intertidal to deep sea in all oceans; many burrow into hard corals and calcareous algae or live in their crevices (Hutchings 1986; Zanol et al. 2021).
The present paper aims to report and discuss a new Late Ordovician jawed polychaete fauna from the Takche Formation of the Pin River section near Farka Muth in the Pin Valley, Spiti, northern India. To date, only a few scolecodont maxillary apparatuses and isolated jaws are described from Gondwana, e.g., from the South African Soom Shale Lagerstätte (Whittle et al. 2008) or the Prague Basin (Tonarová et al. 2023). In addition to scolecodonts, we report the assemblage of chitinozoans, which were extracted from the same samples. The discussion of the chitinozoan assemblage contributes to the recently published palynological study from a coeval section near Gechang in the Parahio River Valley, Spiti, by Wang et al. (2021).
Institutional abbreviations.—CEITEC, Central European Institute of Technology, University of Technology, Brno, Czech Republic; CGS, Czech Geological Survey, Prague, Czech Republic.
Other abbreviations.—AL, attachment lamella; CT, computed tomography; M, maxilla.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in Zoobank: urn:lsid:zoobank.org:pub:2CBBF838-A505-4F28-A745-13F78D379733
Geological setting
Spiti, of which the Pin Valley is a part of, lies in the north Indian state of Himachal Pradesh and displays a well-exposed approx. 10 km thick sedimentary sequence ranging in age from the Neoproterozoic to Cretaceous (Bagati 1990; Bhargava and Bassi 1998; Fig. 1). The Ordovician–Silurian interval of the sequence is part of the Sanugba Group (approx. 1500 m thick) and divided into the Thango and Takche formations (compare Bhargava et al. 1991; Bhargava and Bassi 1998). A recently published summary of the lithostratigraphy of the ?Lower Ordovician–lower Silurian interval by Shabbar et al. (2023) shows that historically, the Thango and Takche units have also been called the Shian Formation and the Pin Formation, respectively (Srikantia 1974, 1977; Srikantia and Bhargava 2018 vs. Goel and Nair 1977, 1982; Suttner et al. 2001; Suttner 2003, 2007). To avoid confusion by parallel terminology for the Upper Ordovician–lower Silurian sequence in Spiti, the scheme of Shabbar and colleagues is followed here (Shabbar et al. 2023: fig. 3; the Takche Formation subdivided into the Farka Muth, Pin and Thanam members).
A Late Ordovician (Katian) to early Silurian (Llandovery) age of the middle and upper part of the Takche Formation (Pin and Thanam members) is constrained by conodonts in the Pin River and the Parahio River sections (Suttner 2003; Suttner et al. 2007b; Myrow et al. 2019). The siliciclastic lowermost unit (Farka Muth Member) has not provided conodonts to date. More promising are palynomorph studies on chitinozoans and acritarchs (e.g., Khanna and Sah 1983; Wang et al. 2021) or macrofossils, particularly brachiopods and trilobites (Reed 1912; Bhargava and Bassi 1998; Suttner 2003).
The rich and diverse fossil flora and fauna of Spiti have attracted scientists since the early–mid 19th century. The first stratigraphic discussions and monographs on early Palaeozoic fossils from Spiti were published by Salter and Blanford (1865), Stoliczka (1866), Griesbach (1891), Hayden (1904), and Reed (1912). Detailed stratigraphic documentation and taxonomic studies on calcimicrobes and calcareous algae were provided by Bhargava and Bassi (1986), Kato et al. (1987), Suttner and Hubmann (2005), Suttner et al. (2005), Hubmann and Suttner (2007), Pandey and Parcha (2018), Chaubey et al. (2019) and Vinn et al. (2023). Shabbar et al. (2022) focused on non-calcified warm-water macroalgae. Corals and stromatoporoids were reported by Bhargava and Bassi (1986, 1998), Suttner (2003), Suttner et al. (2007b) and Suttner and Kido (2011). Other fossil groups described include cephalopods (Suttner and Kröger 2006), ostracods (Schallreuter et al. 2008), cornulitids (Shabbar et al. 2023), bryozoans (Suttner and Ernst 2007) and conodonts (Suttner 2003; Suttner et al. 2007b; Myrow et al. 2019). Palaeoscolecids, tentaculitids, gastropods, trilobites, brachiopods, and echinoderms were briefly characterised and figured by Reed (1912), Bhargava and Bassi (1998), Suttner (2003), and Suttner et al. (2007b).
Bhargava and Bassi (1986, 1998) studied five richly fossiliferous Ordovician–Silurian sections in detail. From the northwest to southeast, these are located within the Takche area, where the type section of the Takche Formation is exposed along the Spiti River bank (sensu Srikantia 1974, 1977 and Srikantia and Bhargava 2018), in the Parahio Valley near Gechang, in the Pin Valley near Farka Muth, at Leo and Manchap. Bed-by-bed lithological logs of the Takche Formation were provided only of the Pin River and Parahio River sections by Suttner (2003) and Myrow et al. (2019), respectively. The unit is generally characterised by a mixed siliciclastic-carbonate shallow-marine environment and varies in thickness between 145 and 280 m (Bhargava and Bassi 1986, 1998; Suttner 2003; Suttner et al. 2007b; Myrow et al. 2019; Chaubey et al. 2023).
Following the palaeogeographic reconstructions of Scotese (2000) and Torsvik and Cocks (2009, 2013), the area was a part of the Gondwana palaeocontinent and located between Arabia, Antarctica, and Australia (Fig. 1D). A palaeolatitudinal position at about ~25–30° S for northern India during the Late Ordovician–early Silurian, as suggested by Torsvik and Cocks (2009), is also supported by the occurrence of warm-water fossil groups like dasycladacean algae, stromatoporoids and corals in the Takche Formation.
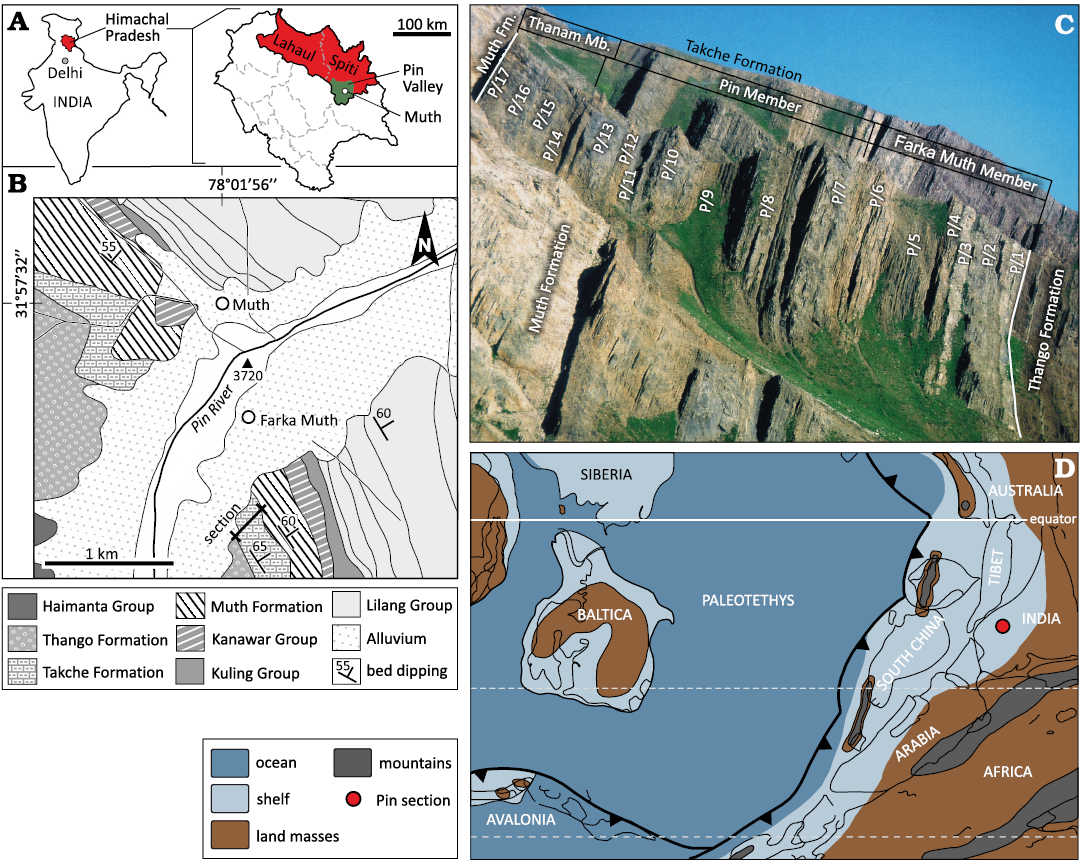
Fig. 1. A. Geographic location of the studied area within India. The state of Himachal Pradesh and an enlarged sketch of it with the Lahaul and Spiti district (red) and the Pin Valley (green). B. Geological map of the outcropping area within the Pin Valley in Spiti with the studied section south of Farka Muth indicated. The age of the lithostratigraphic units largely follows Bhargava and Bassi (1998): Haimanta Group (late Proterozoic to middle Cambrian), Sanugba Group including the Thango Formation (Ordovician) and the Tackche Formation (Upper Ordovician to lower Silurian), Kanawar Group (Lower Devonian to lower Permian) with the Lower Devonian Muth Formation at its base, Kuling Group (Permian), Lilang Group (Lower Triassic to Upper Jurassic). C. Photograph of the studied section including the regional lithostratigraphic subdivision to units P/1–P/17. D. Palaeogeographic reconstruction with the assumed position of the Pin River section during the Late Ordovician (redrawn from Scotese 2000).
The Pin River Section.—The studied section is located in the Pin Valley (Spiti) near the small village of Farka Muth at an altitude between ~3800–4100 m above sea level. The section can be reached by jeep or bus starting from Manali (Kullu) over Kaza, through Sangnam directly to Muth (GPS coordinates: N 31°57’32”, E 78°01’56”) and Farka Muth (GPS coordinates: N 31°56’55”, E 78°01’55”). Two high mountain passes (Rothang 3976 m and Kunzam La 4551 m) must be crossed before arriving at Losar on the border of the district (Suttner 2003). The sedimentary sequence of the Pin Valley was mapped in detail by, e.g., Fuchs (1982) and Bhargava and Bassi (1998). The section comprises early Palaeozoic sediments, including the uppermost portion of the Ordovician purple-coloured Thango Formation, followed by the grey Takche Formation, and succeeded by the white quartzites of the Lower Devonian Muth Formation (Fig. 1). A bed-by-bed lithological log of the 280 m thick Takche Formation at the Pin River section near Farka Muth was produced by Suttner (2003). Based on lithological characters, 17 distinct units (Figs. 1C, 2: P/1–P/17) were discriminated (Suttner 2003, 2007; Suttner et al. 2007b). Units P/1–P/6 consist of siliciclastic deposits and represent the Farka Muth Member (originally established by Suttner 2007). They are succeeded by a siliciclastic-influenced limestone portion (units P/7–P/13) which, according to Shabbar et al. (2022), represents the Pin Member. The upper portion of the Takche Formation (units P/14–P/17) is again siliciclastic and assigned to the Thanam Member (compare Shabbar et al. 2023: fig. 3). For the Pin Member, a Katian age is suggested by the occurrence of the Late Ordovician conodont Amorphognathus ordovicicus (Suttner 2003; Suttner et al. 2007b). Near the base of the Thanam Member, within the unit P/14, an early Silurian conodont fauna was identified (Suttner 2003; Suttner et al. 2007b). These biostratigraphic datings are also supported by a distinct carbon isotope excursion across the section (Suttner et al. 2007b).
Material and methods
Initially, 41 rock samples were collected, processed by acid leaching (5% formic acid), wet sieved in four fractions (sieve mesh width: 63 µm, 125 µm, 250 µm, 500 µm), and dry-picked by TJS in 2000–2003 (Suttner 2003: 46–51, appendix). A further 16 samples (weight between 20–800 g) of the remaining collection stored in the repository of the Department of Palaeontology, University of Vienna, were dissolved to obtain additional scolecodonts. After dissolution in 6% acetic acid, the residues were gently rinsed through 80 and 50 µm sieves. The wet samples were handpicked for organic-walled microfossils from Petri dishes under a stereomicroscope, using a micropipette or an eyelash (wet technique sensu Kielan-Jaworowska 1966).
The insoluble residue contained scolecodonts, chitinozoans, tentaculitids, conodonts, ostracods, brachiopods, crinoids, gastropods, corals, silicified bryozoans, melanosclerites, spicules, and palaeoscolecid remains. Most invertebrate skeletons recovered after acid treatment were silicified or preserved as casts or moulds on clay-rich rock surfaces. In total, seven samples were productive for scolecodonts (P/7/31, P/8/27, P/8/69, P/11/80, P/13/117, P/13/231, P/13/283), and six samples contained chitinozoans (P/7/31, P/8/69, P/8/78, P/12/9, P/13/117, P/13/231) (Fig. 2). The most productive sample for organic-walled microfossils was P/13/231. The preservation of microfossils varies from poor to very good. Some specimens could not be determined even to genus level due to flattening, deformation, or fragmentary preservation. The relatively small number of identifiable specimens did not allow a statistical analysis of the assemblage. The specimens were photographed using a Scanning Electron Microscope FEG-SEM Tescan Mira 3GMU at the Czech Geological Survey, Prague. X-ray micro- and submicron-CT were used for the study of internal structures and 3D reconstruction using the below-specified facilities of CEITEC (Brno). This technique is capable of non-destructive three-dimensional visualisation and analysis of the surfaces and internal structures of objects at a high spatial resolution. In CT, a series of X-ray projections is acquired over a circular arc, and cross-sections of the scanned samples are obtained from these projections by tomographic reconstruction. Several methods of mounting the specimens for the CT were tested. The best results were obtained using the following technique: the isolated specimens were prepared for measurement by placing them in a kapton tube and sealing the tube with parafilm. This preparation allowed the maxillae to be stable during CT measurements without any need for adhesives or other mounting measures. The maxillae were previously studied on SEM and therefore they were gold-coated. It turned out that the coating did not affect the study of internal structures, just there is a bright outer layer in the photos that was induced by the metal coating. In a few places, dark shadows appeared but did not influence the quality of measuring. Our study shows that the gold coating is not an obstacle for CT; it helped to make the specimens more resistant, which is useful when preparing the specimens for measurements. Preparation of the partly exposed scolecodont maxillary apparatus on a tiny piece of rock consisted of glueing it onto a rigid but low-density plastic rod. The cyanoacrylate glue fixed and stabilised the sample during measurement but did not interfere with parts of the sample containing the fossil material of interest.
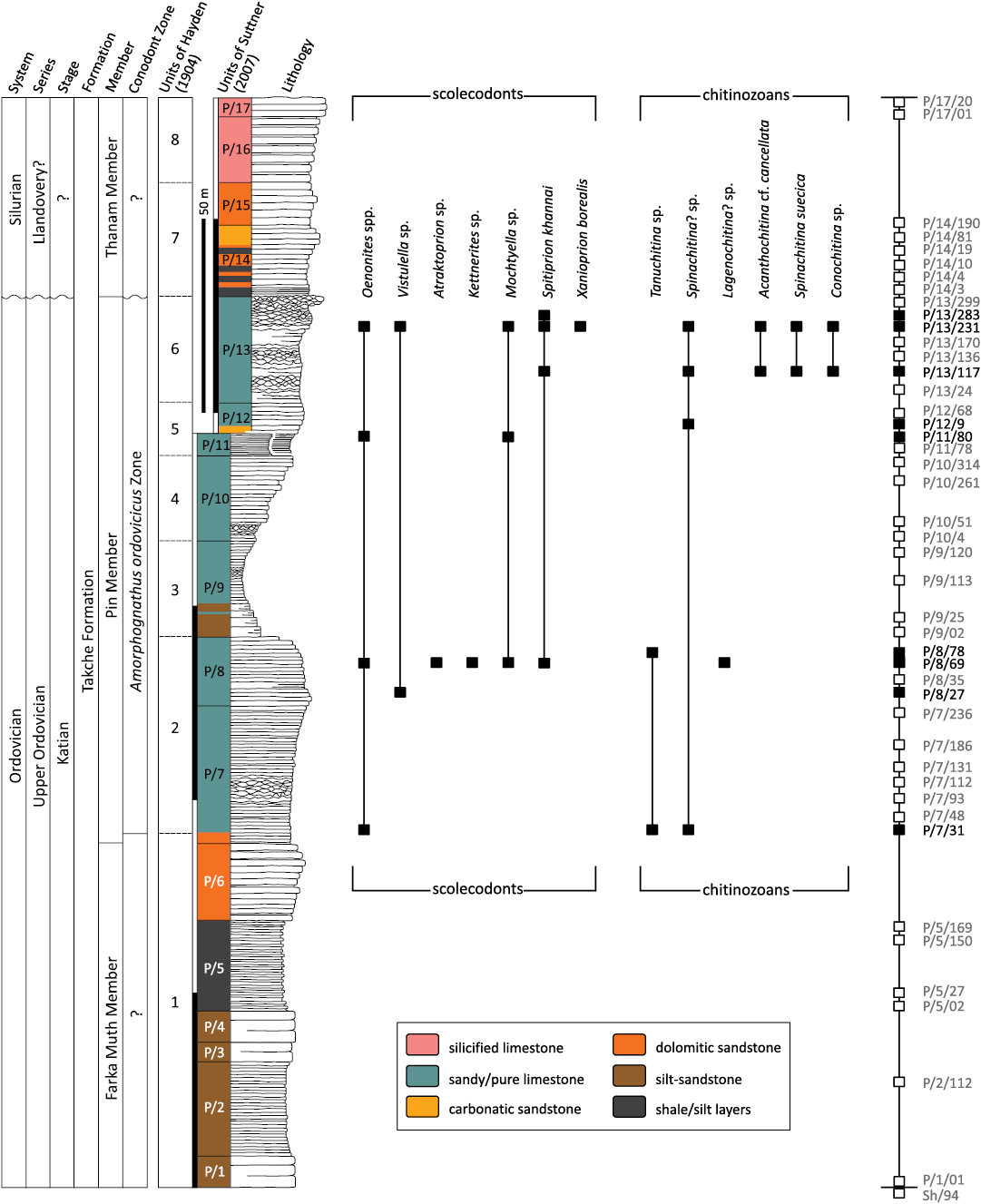
Fig. 2. Simplified lithological log of the Takche Formation at the Pin River section with the position of samples and the distribution of scolecodont and chitinozoan taxa. Sampling log on the right: solid squares denote productive samples for microfossils, whereas white squares indicate samples barren for scolecodonts and chitinozoans.
The isolated maxillae were analysed by a Rigaku Nano3DX submicron-CT scanner (Rigaku Corporation, Japan). X-rays were produced using a MicroMax-007 HF generator operating at 50kV/24mA, with a molybdenum target. Projection images were acquired using a CCD (charge-coupled device) detector equipped with a scintillating crystal and magnifying microscope optics. Optical magnification was used because the quasi-parallel geometry of the nano3DX has very little geometric magnification. The rock piece with the jaw apparatus was studied by a Waygate Phoenix v|tome|x L240 micro-CT scanner (Waygate Technologies, Germany). A nanofocus transmission source at 60kV/120µA was used for X-ray production. Images were enlarged through geometric magnification and captured using a flat panel detector. Scan parameters for the measurements are summarised in Table 1. The acquired data were reconstructed using the respective Rigaku and Waygate reconstruction software to obtain three-dimensional CT volumes. These were then processed using the software VGStudio MAX 2023.1 (Volume Graphics, Germany). Results in videos SOM 1–6 (Supplementary Online Material available at http://app.pan.pl/SOM/app69-Tonarova_etal_SOM.pdf)
The sample code goes back to Suttner (2003) and is as follows: e.g., Pin River section/Unit 8/bed 69 = P/8/69. All figured specimens are housed in the collection of the Czech Geological Survey, collection numbers CGS PT156–170.
Table 1. CT scan parameters.
|
Sample |
P13, isolated maxillae |
Rock piece |
|
Scanner |
nano3DX |
v|tome|x L240 |
|
Tube voltage [kV] |
50 |
60 |
|
Tube current [µA] |
24000 |
120 |
|
Target material |
Mo |
W |
|
Filter |
0.1 mm Al |
none |
|
Linear voxel size [µm] |
0.524 |
1.800 |
|
Scan range |
180° |
360° |
|
Projection no. |
800 |
2000 (3×average + 1 skip) |
|
Exposure [s] |
6 |
0.5 |
Historical background
Most studies on Late Ordovician scolecodonts focused on Baltoscandia (Eriksson et al. 2013). Systematic studies were started already by Kielan-Jaworowska (1962, 1966) and, more recently, Hints (2000), Hints et al. (2010, 2023), and Hints and Tonarová (2023) provided analyses from the Baltica palaeocontinent. They concluded that the jawed polychaetes were diverse at that time, with a prominent component of mochtyellids from the Placognatha group (Pistoprion, Mochtyella, Vistulella) and polychaetaspids from the Labidognatha group (details on the concept of apparatus types can be found in Paxton 2009). The second best-covered area is Laurentia. Late Ordovician scolecodonts from Laurentia were first studied e.g., by Stauffer (1933) and Eller (1942, 1945 and other papers of the author) or, more recently, by Bergman (1991), Eriksson and Bergman (2003), Eriksson et al. (2005), Schröer et al. (2016), or Hints et al. (2016). Overall, the faunas are dominated by labidognath and prionognath taxa, such as polychaetaspids, paulinitids, and atraktoprionids, with placognath taxa being less common than in Baltica. However, starting from the latest Ordovician, Laurentian and Baltic polychaete faunas have become increasingly similar (Eriksson et al. 2013).
There is only a handful of published records on Late Ordovician scolecodonts from Gondwana. These studies were summarised by Hints et al. (2015) and Tonarová et al. (2023). Hints et al. (2015) described a Late Ordovician assemblage from the Qusaiba-1 drill core, Saudi Arabia, and found out that most of the species recovered belong to the families Ramphoprionidae, Paulinitidae, Polychaetaspidae, and Atraktoprionidae, and only a very low proportion of taxa are with placognath type jaw apparatuses. Katian/Hirnantian scolecodonts from the Prague Basin (Levín locality) were studied by Tonarová et al. (2023). Similar to the Qusaiba-1 drill core, the taxa with labidognath and prionognath type maxillary apparatuses predominated in the Prague Basin samples, whereas placognath and ctenognath taxa were relatively rare. Polychaetaspids dominated the Levín assemblage, followed by other families such as ramphoprionids, paulinitids, and atraktoprionids. A similar general pattern is typical for the Laurentian samples but contrasts with the Baltic polychaete faunas.
Concerning India, the only previous report on scolecodonts was published by Khanna (1984), who worked on palynomorphs from lower Palaeozoic deposits of the “Pin Dolomite” (= Takche Formation) at the Parahio River section in Spiti. Khanna (1984) determined 15 single-element genera, but due to poor preservation of the material, proper identification was impossible. However, the presence of paulinitids and possibly mochtyellids can be confirmed based on the specimens illustrated by Khanna (1984). Unfortunately, the work of Ashok Kumar Khanna remained unfinished due to a fatal accident during the fieldwork in 1984.
The Late Ordovician chitinozoans from Gondwana are far better studied than scolecodonts. Here, we mention only some examples of the previous studies that are more relevant to our new material. Firstly, several papers describe chitinozoan faunas from the Middle East, e.g., by Al-Hajri (1995) or Al-Shawareb et al. (2017) from the Katian of Saudi Arabia, Paris et al. (2015) from the Katian and Hirnantian of Saudi Arabia, Ghavidel-Syooki (2001, 2023), Ghavidel-Syooki and Winchester-Seeto (2002), Ghavidel-Syooki and Piri-Kangarshahi (2021) from Iran, among others. The South American chitinozoans were studied by Grahn (1992) in Brazil, de la Puente et al. (2020) and de la Puente and Astini (2023) in Argentina. Other reports come from Northern Africa, from Morocco by Elaouad-Debbaj (1984) and Bourahrouh et al. (2004), and from Libya by Paris (1988) and Thusu et al. (2013). South China is relatively well-studied (e.g. Liang et al. 2023), and the first reports from the Prague Basin were presented by Tonarová et al. (2023).
Within the Himalayas, Sinha et al. (2011) and Sinha and Verniers (2016) discovered Middle to Late Ordovician chitinozoans from the Shiala Formation of the Garhwal-Kumaon subbasin, Uttarakhand, India. Khanna and Sah (1983), Khanna (1984) and Khanna et al. (1985), however, were the first authors to mention the presence of chitinozoans and other palynomorphs in the Spiti area. Chitinozoans from the Parahio River section, representing the time interval before and during the Boda event, were studied by Wang et al. (2021). That section, measured bed-by-bed by Myrow et al. (2019), is located approximately 12 kilometres north of the Pin River section near Farka Muth, as reported in our study. Wang et al. (2021) found 13 species of chitinozoans belonging to seven genera, indicating a late Katian age.
Systematic palaeontology
Scolecodont descriptive terminology is based on that of Kielan-Jaworowska (1966), Jansonius and Craig (1971) and Bergman (1989).
Class Polychaeta Grube, 1850
Order Eunicida Dales, 1963
Family Ramphoprionidae Kielan-Jaworowska, 1966
Genus Spitiprion Tonarová, Suttner, & Hints nov.
Zoobank LSID: urn:lsid:zoobank.org:act:560719CA-4C5D-47EB-B077- 077546969C97
Etymology: After Pin River section Spiti in the Trans-Himalayan region of northern India.
Type species: Spitiprion khannai Tonarová, Suttner, & Hints sp. nov., see below. By monotypy.
Diagnosis.—As for the type species.
Remarks.—The closest known ramphoprionid to Spitiprion is Megaraphoprion Eriksson, 2001. The biggest difference is fully denticulated dentary of the first maxillae of the latter genus and the ramus of right first maxilla (MI) is more slender in Spitiprion. Spitiprion has also much more prominent and slender hooks on the first maxillae than the type species M. magnus. The second maxillae of S. khannai Tonarová, Suttner, & Hints gen. et sp. nov. also differ considerably from the type species of Megaraphoprion. The second maxillae (MII) of M. magnus are sturdy, with wide ramus and blunt teeth whereas the MIIs of S. khannai Tonarová, Suttner, & Hints gen. et sp. nov. are rather slender, with pointed denticles and pointed ramus, reminding the second maxillae of Ramphoprion Kielan-Jaworowska, 1962.
Stratigraphic and geographic range.—Katian, Upper Ordovician of Pin Member (Amorphognathus ordovicicus Conodont Biozone), Takche Formation, Pin Valley, Spiti, India.
Spitiprion khannai Tonarová, Suttner, & Hints sp. nov.
Figs. 3A–G, 4; SOM 1–6.
Zoobank LSID: urn:lsid:zoobank.org:act:7D98ABE7-300E-4D50-9028- 9608403D9ABA
Etymology: In honour of the Indian palaeontologist Ashok K. Khanna (1951–1984), who first described scolecodonts from India.
Holotype: CGS PT_159_4, left first maxilla, from sample P/8/69.
Type locality: Pin River section, Spiti, India.
Type horizon: Amorphognathus ordovicicus Conodont Biozone, Katian, Upper Ordovician.
Material.—Five isolated left MI (CGS PT_159_3, 159_4, 169_1, 169_16, 170_2), 5 isolated right MI (CGS PT_159_5, 168_8, 169_2, 159_17, 170_ 1), 1 MII (CGS PT_159_2), 1 partly preserved apparatus (including left and right MI, MIV) (CGS PT_159_1). All from the type locality and horizon.
Diagnosis.—MI elongated, widest in the posterior third. The dentary spread along the posterior half of the jaw length, terminating with a prominent and bent anterior hook. The right MI with a conspicuous slender ramus of rectangular shape, with a prominent inner wing occupying approximately posterior third of jaw, the wing is ending with a pointed extension. The left MI with a prominent inner wing extending approx. posterior 40% of the jaw, ending by a prominent pointed extension. In the outer margin, slightly below the centre protrudes a prominent extension, symmetrically to the inner wing extension. The dentary of both first maxillae is occupied by 8 to 12 relatively small but sharp denticles. The un-denticulated hook starts at approx. mid-length of the jaw, and it has an almost whip-like ending. The myocoele opening occupies about one-third of the jaw length. The MIIs are expected to be near mirror images of each other, with a slender ramus placed in the anterior-most part, pointed denticles, with the most prominent first denticle. MIV (right confirmed) is slightly longer than wide, occupied with relatively large and sturdy denticles. The basal plate was not found, but most likely it is sub-rectangular, with small denticles and a tapering antero-dextral process.
Description.—Right MI: Length 480–540 µm, width 120–126 µm, width is approx. four times shorter than length (width 0.2–0.3 of jaw length), dorsal view: the 9–12 pointed denticles are bent outwards, with the tip often broken off. The undenticulated hook extends for approx. half of the maxilla length (compared with other ramphoprionids that have denticulated also the anterior part of the dentary), the bend pointed tip is very slender and long. Typical for the species is a prominent extension of the maxilla on the centre of both outer and inner face.
Ventral view: the myocoele opening represents approx. 1/3 of the maxilla. In the ventral view, all denticles are visible through the myocoele opening (i.e., the length of the denticulated dentary approximately matches the length of the myocoele opening).
In the right MI (Fig. 3E) remained attached also an attachment lamella (AL). The ALs are attached to the bases of MI, and when the jaws are retracted, the AL extend anteriorly-posteriorly on either side of the maxillae to near MIII and MIV, therefore they may increase resistance to hyperextension of the primary maxillae that may result from contact of the open jaws with struggling prey or surrounding substrate—this potential function of the AL is consistent with the motile carnivorous lifestyle of Lumbrinereis latreilli Audouin & Milne Edwards, 1834 (Clemo and Dorgan 2017). However, it is supposed that ramphoprionids do not have descendants in the recent oceans (Kielan-Jaworowska 1966), therefore the lifestyle and diet remain speculative.
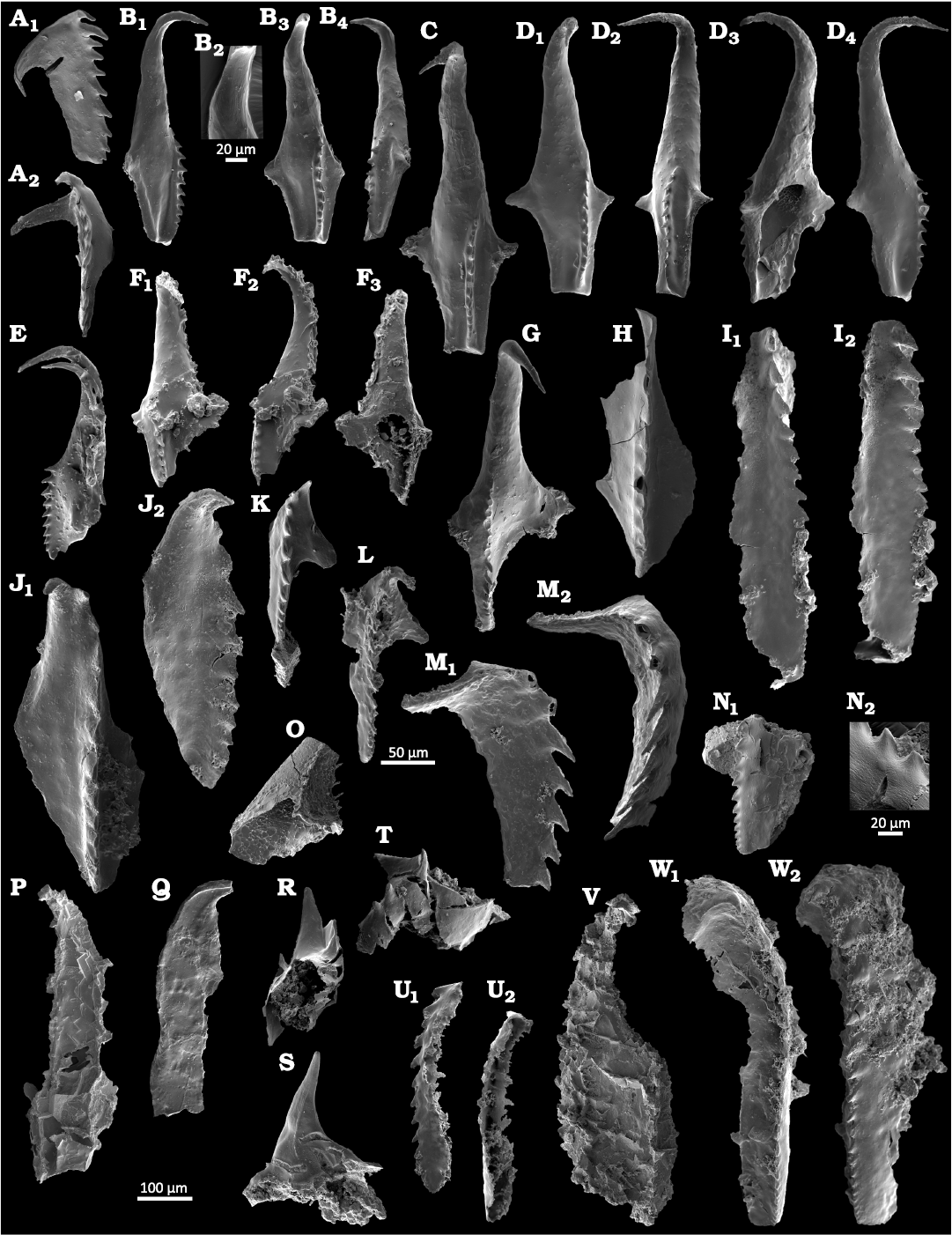
Fig. 3. Photomicrographs of eunicid polychaete scolecodonts from the Katian, Upper Ordovician of the Pin River section (Pin River Valley, Spiti, Himalaya, India). A–G. Spitiprion khannai gen. et sp. nov. A. CGS PT159_2 (sample P/8/69), left MII in lateral (A1) and dorsal (A2) views. B. CGS PT159_3 (sample P/8/69), left MI in lateral (B1, B4) and dorsal (B3) views, a close-up of the anterior part (B2). C. CGS PT159_4, holotype (sample P/8/69), left MI in dorsal view. D. CGS PT169_1 (sample P/13/231), left MI in dorsal (D1), dorsolateral (D2), and lateral (D4) views, ventral view (D3), note the polygonal structure in the myocoele opening. E. CGS PT170_1 (sample P/13/283), right MI and lateral tooth in lateral view. F. CGS PT169_2 (sample P/13/231), right MI in dorsal (F1), lateral (F2), and ventral (F3) views. G. CGS PT159_5 (sample P/8/69), right MI in dorsal view. H. Oenonites? sp., CGS PT159_6 (sample P/8/69), broken dorsal part of left MI in dorsal view. I. Mochtyella? sp., CGS PT169_3 (sample P13/231), right MI in dorsolateral view (I1), and in lateral view (I2). J–L. Oenonites sp. J. CGS PT169_4 (sample P/13/231), left MI in dorsal (J1) and lateral (J2) views. K. CGS PT159_7 (sample P/8/69), right MI in dorsal view. L. CGS PT169_5 (sample P/13/231), right MI in dorsal view. M. Xanioprion borealis Kielan-Jaworowska, 1962, CGS PT169_6 (sample P13/231), left MII in lateral (M1) and dorsal (M2) views. N, O, R–U. Scolecodonta indet. N. CGS PT159_8 (sample P/8/69), fragment of an undetermined maxilla in lateral view (N1) and detail of the dentary (N2). O. CGS PT159_9 (sample P8/69), fragment of the lateral tooth. R. CGS PT159_12 (sample P/8/69), single lateral tooth. S. CGS PT159_13 (sample P/8/69), single lateral tooth. T. CGS PT159_14 (sample P/8/69), single lateral tooth. U. CGS PT169_7 (sample P/13/231), placognath maxilla in lateral (U1) and ventral (U2) views. P, Q. Kettnerites sp. P. CGS PT159_10 (sample P/8/69), right MI in dorsal view. Q. CGS PT159_11 (sample P/8/69), left MI in dorsal view. V. Atraktoprion sp., CGS PT159_15 (sample P/8/69), left MI in dorsal view. W. Vistulella sp., CGS PT169_7 (sample P/13/231), left MI in dorsal (W1) and dorsolateral (W2) views. The scale bar refers to all specimens except for B2, L, and N2, where a separate bar is indicated.
Left MI: Length 430–580 µm, width 126–138 µm, width is approx. 4 times shorter than length (width 0.2–0.3 of jaw length), dorsal view: the shape of the maxilla is rectangular in the posterior area and tapers anteriorly to the un-denticulated anterior part that extends for half of the maxilla length and ends with a prominent hook-like slender fang. The dentary consists of ten pointed denticles that are situated only in the posterior half of the dentary, the denticles are bent outwards, and decrease in size toward the posterior end, with the smallest first two and last two denticles. Posteriormost denticles are tightly packed and continue into the un-denticulated short ridge. The posterior margin is rather short, it extends 14% of the jaw length. Among other ramphoprionids the posterior margin represents 20 to 30% of the jaw length in Protarabellites species, 15 to 20% of the jaw length in Ramphoprion species, and 19 to 25% in Megaramphoprion magnus Eriksson, 2001. In ventral view, the myocoele opening extends a little more than 1/3 of the maxilla (approx. 35–40% of the jaw length), which is more closed than in other ramphoprionid species—where it is usually representing from 0.4 (Ramphoprion bialatus Hints, 1998) to 0.8 (Protarabellites triangularis) of the maxilla length.
Left MII: Length 300 µm, width 180 µm (0.6 of the jaw length). The dentary is prominent, strongly elevated, gently curved, occupying the entire jaw length, and equipped with ten denticles of similar size as in the first maxillae. The first two biggest teeth are followed by two smaller denticles and then a row of teeth of similar size. The ramus is prominent but very narrow, triangular in shape. In the ventral view, the myocoele is almost gaping.
Right MIV: Jaw is about as high as long, it is quite flat, with an almost square outline. The left side is occupied by seven to eight sturdy denticles of more or less similar size, except for the first bigger denticle. The preservation (flattening) of MIV may have influenced the appearance of the maxilla and may be modified when more material is at hand.
Other jaws presently unknown.
Remarks.—The ramphoprionids, known solely by their jaws (Eriksson 2002), emerged in the Middle Ordovician (Kielan-Jaworowska 1962, 1966; Szaniawski 1996) and their youngest known record is late Ludfordian (Ludlow, Silurian) in age (Eriksson 2001). Up to now, there were four genera included in the family, the type genus Ramphoprion Kielan-Jaworowska, 1962, Protarabellites Stauffer, 1933, Pararamphoprion Männil & Zaslavskaya, 1985, and Megaramphoprion Eriksson, 2001. Eriksson (2002) suggested that the Silurian ramphoprionids were inhabiting only regions close to the equator, but Tonarová et al. (2012) confirmed their presence in the Ludfordian of the Prague Basin, which was located between 25 and 30 degrees south during the Ludfordian (Tasáryová et al. 2014).
The family Ramphoprionidae was considered basal and fairly primitive among labidognath families (Eriksson 2001). However, their jaw apparatuses exhibit characters considered both primitive, such as denticulation in first maxillae and advanced, such as enclosed to sometimes near strongly enclosed myocoeles. Therefore, ramphoprionids were presumably more advanced than the polychaetaspids, а group with similar jaw apparatus architecture, although the latter apparently have а longer (documented) stratigraphic range (cf. Kielan-Jaworowska 1966; Szaniawski 1996).
However, the first maxillae of Spitiprion khannai Tonarová, Suttner, & Hints gen. et sp. nov. differ from other ramphoprionids by the reduced denticulation to only half of the maxillae. Eriksson (2001: figs. 9.28, 9.31–44, and 45, and 46, respectively) reported from Gorstian and Ludfordian (Ludlow, Silurian) of Gotland a decrease in denticulation of the species Ramphoprion gotlandensis in the anterior part of the first maxillae dentaries, but that was reflected by a paucidate dentary, i.e., usually blunt denticles with longer gaps in between. The loss of denticulation in the first maxillae is observed in several extant polychaete groups, e.g., in the families Eunicidae or Onuphidae (Beesley et al. 2000; Paxton 2009). However, the denticulation can be variable, e.g., in the labidognath family Paulinitidae Lange, 1947 (Langeites lublinensis Szaniawski and Wrona, 1973) and in the extant prionognath family Oenonidae (Drilonereis Claparède, 1870). Palaeozoic paulinitids were considered directly ancestral to modern labidognaths by Kielan-Jaworowska (1966).
Stratigraphic and geographic range.—Katian, Upper Ordovician, Pin Member (Amorphognathus ordovicicus Conodont Biozone), Takche Formation, Pin Valley, Spiti, India.
Results
Scolecodonts.—The collection of scolecodonts from Katian deposits of the Takche Formation at the Pin River section near Farka Muth in Spiti includes approx. 60 maxillae (counting only first or second maxillae, which can be determined at least to the generic level). These were assigned to seven genera belonging to six families: Polychaetaspidae, Paulinitidae, Ramphoprionidae, Mochtyellidae, Xanioprionidae, and Atraktoprionidae (Figs. 2, 3). The maxillae are fragile and commonly fragmentarily preserved. Some of the jaws also reflect scars of mineral growth. All these conditions impede species-level identification. The two richest samples were P/8/69 and P/13/231, both contained representatives of five different polychaete genera (Fig. 2).
Ramphoprionids are represented by a new genus and species Spitiprion khannai Tonarová, Suttner, & Hints. Representatives of the new species occur in units P/8 and P/13 of the Takche Formation at the Pin River section. Additionally, a maxillary apparatus consisting of four jaws (paired MI, MII and MIV) partly embedded in a piece of rock, was found in sample P/8/69 and studied using micro-CT (Fig. 4). The isolated maxillae of the new species from the sample P/13/231 were studied using a submicron-CT (Table 1, SOM 1–6; same specimens are on Fig. 3D and F, respectively; for a detailed description see Systematic palaeontology).

Fig. 4. Photomicrographs of the eunicid polychaete Spitiprion khannai gen. et sp. nov. apparatus preserved on an acetic acid-etched rock surface, CGS PT159_1 (sample P/8/69) from the Katian, Upper Ordovician of the Pin River section (Pin River Valley, Spiti, Himalaya, India). A1, optical microscope photograph; A2, SEM photomicrograph; A3, 3D reconstruction based on images from X-ray micro-computed tomography. The affinity of the maxilla (orange) is yet unknown (could be the second maxilla but diagnostic features are missing). A4, sketch of the maxillary apparatus of the new species of eunicid polychaete Spitiprion khannai. The maxillae (white) with dotted lines are assumed according to closely related species but have not been found yet. MI, first maxilla; MII, second maxilla; MIV, fourth maxilla; AL, attachment lamella; BP, basal plate. Scale bars 100 µm.
Jaws of polychaetaspids were found in three samples (P/7/31, P/8/69, P/11/80, P/13/231), and all of them are assigned to Oenonites Hinde, 1879, with at least two species present. Because of poor preservation, determination at the species level was impossible; thus, the specimens were assigned to Oenonites spp. (Fig. 3H, J–L).
Paulinitid maxillae were found only in one sample (P/8/69). The specimens have been determined as Kettnerites Žebera, 1935. However, the first maxillae of Kettnerites sp. (Fig. 3P, Q) are so distinctive that there is no doubt about their assignment.
Placognath taxa are relatively rare in the samples. Their saw-like maxillae tend to break, and therefore taphonomy may have biased the abundance quite significantly. The sample P/13/231 contains most of the placognath forms, but it is also the most productive sample overall. Among mochtyellids, two genera were recorded: Vistulella sp. (Fig. 3W) and Mochtyella sp. (Fig. 3I). Xanioprionids are also very rare in the Pin River section—only a single maxilla was recovered and identified as Xanioprion borealis Kielan-Jaworwoska, 1962 (Fig. 3M).
Additionally, a single maxilla from sample P/8/69 was assigned to Atraktoprion sp. (Fig. 3V), showing close similarity to Atraktoprion sp. A of Eriksson et al. (2005).
Chitinozoans.—The collection of chitinozoans from the Pin River section includes approx. 50 specimens, which are assigned to five genera (Fig. 2). Generally, the specimens are characterised by their black vesicles. The appendices are commonly broken off, and the ornamentation is difficult to recognise. Most abundant are specimens with a medium conical test of about 300–400 µm in length, characterised by distinct, web-like, regularly distributed spines (Fig. 5C) suggesting mesh-like ornamentation. These specimens are assigned to Acanthochitina cf. cancellata Martin, 1983 (Fig. 5A–G). The second most common genus is Spinachitina, specimens of which are here assigned to Spinachitina suecica (Laufeld, 1967) (Fig. 5H–M). Because of poor preservation, several other chitinozoan specimens are kept under open nomenclature within the genera Spinachitina (Fig. 5N), Conochitina (Fig. 5O), Lagenochitina? (Fig. 5P), and Tanuchitina (Fig. 5Q–R).
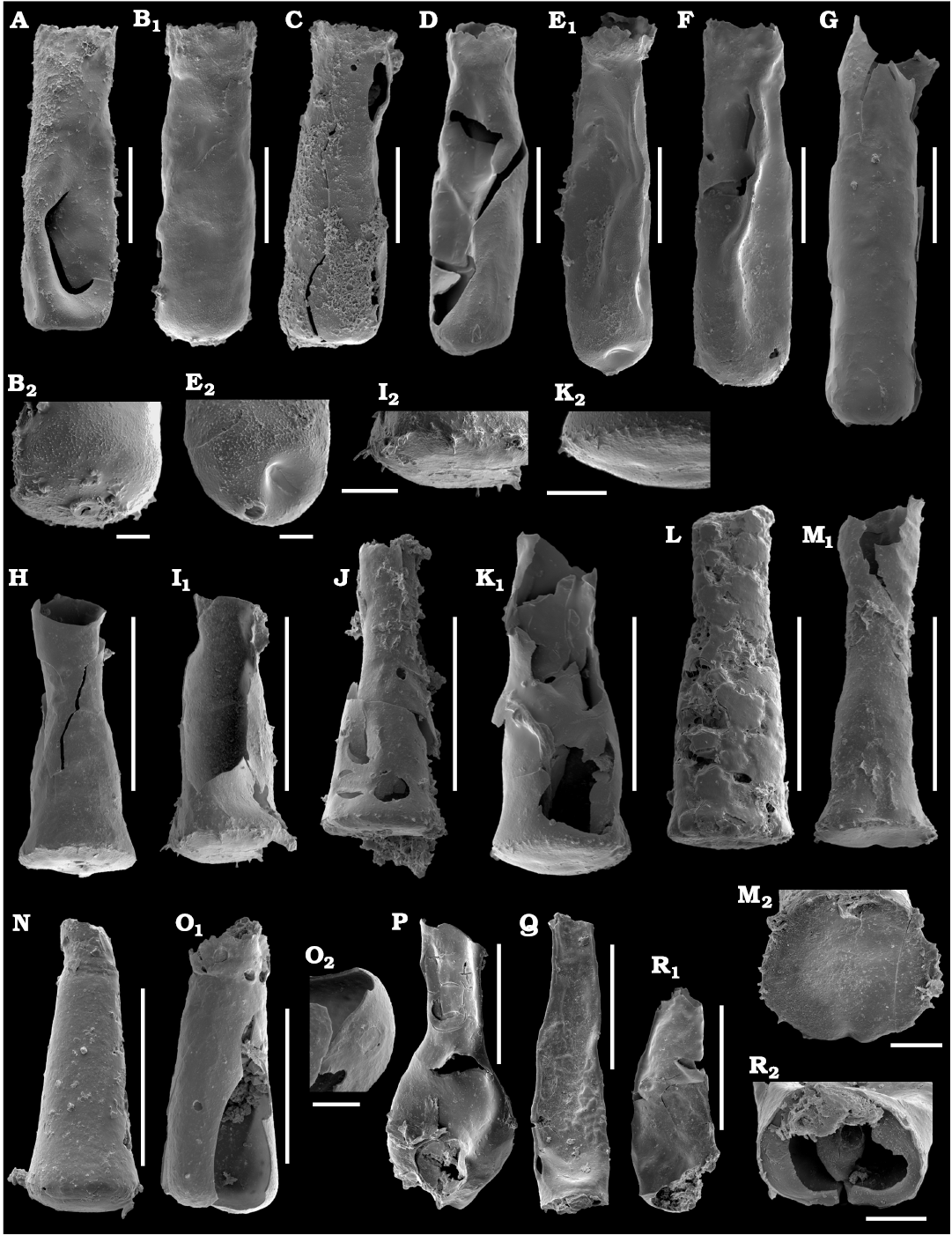
Fig. 5. Photomicrographs of selected chitinozoans from the Katian, Upper Ordovician of the Pin River section (Pin River Valley, Spiti, Himalaya, India). A–G. Acanthochitina cf. cancellata Martin, 1983. A. CGS PT169_9 (sample P/13/231). B. CGS PT169_10 (sample P/13/231), general view (B1), close-up showing the base in detail (B2). C. CGS PT169_11 (sample P/13/231). D. CGS PT169_12 (sample P/13/231). E. CGS PT169_13 (sample P/13/231), general view (E1), close up showing the base in detail (E2). F. CGS PT169_14 (sample P/13/231). G. CGS PT168_1 (sample P/13/117). H–M. Spinachitina suecica (Laufeld, 1967). H. CGS PT168_2 (sample P/13/117). I. CGS PT168_3 (sample P/13/117), general view (I1), close up showing the base in detail (I2). J. CGS PT168_4 (sample P/13/117). K. CGS PT168_5 (sample P/13/117), general view (K1), close up showing the base in detail (K2). L. CGS PT169_15 (sample P/13/231). M. CGS PT168_6 (sample P/13/117), general view (M1), close up showing the base in detail (M2). N. Spinachitina? sp., CGS PT156_1 (sample P/7/31). O. Conochitina sp., CGS PT168_7 (sample P/13/117), general view (O1), close-up of the base of the specimen with a mucron (O2). P. Lagenochitina? sp., CGS PT156_16 (sample P/8/69). Q, R. Tanuchitina sp., Q. CGS PT156_2 (sample P/7/31). R. CGS PT156_3 (sample P/7/31), general view (R1), close-up showing the base in detail and the short carina (R2). Scale bars 100 µm, except for enlargements B2, E2, I2, K2, M2, O2, R2 20 µm.
Discussion
The regional fossil record and facies of the Takche Formation indicate shallow-water conditions within the subtropical–tropical realm (Bhargava and Bassi 1986, 1998; Bordoloi 2002; Suttner 2003; Suttner et al. 2007b; Chaubey et al. 2023) that usually offer ideal conditions for jawed polychaetes (Eriksson et al. 2004). The two most productive samples were P/8/69 and P/13/231, both of which are on top of a transgressive cycle (sensu Suttner et al. 2007b). The preservation of maxillae allowed recognition of a general pattern only, which shows that, e.g., the labidognath taxa prevail compared to placognaths. As computed tomography (CT) has become a major method in palaeontology for non-destructive analysis and 3D reconstruction of fossil invertebrate and vertebrate soft and hard tissue (Abel et al. 2012), we decided to apply the method to the specimens herein.
There is only a handful of papers on CT studies of scolecodonts. Clemo and Dorgan (2017) undertook an analysis of Recent jawed polychaetes, mainly focusing on the maxillary apparatuses. The resulting 3D models are based on computed tomography data produced for analysis of the principles of the functionality of the jaw apparatuses. Recently, Eriksson et al. (2017) analysed fossil jaws using CT scanning for the reconstruction of maxillae that were embedded in sedimentary rock. They studied the oldest bobbit worm from the Lower–Middle Devonian of Ontario, Canada. However, the maxillae were over one centimetre in length, which is extreme among scolecodonts. Parry et al. (2019) employed micro-CT to reconstruct the maxillary apparatus of Plumulites (machaeridian) and confirmed a long-disputed position of machaeridians within the crown group of the Phyllodocida (sister group of Eunicida). Thanks to micro-CT analysis, Gueriau et al. (2023) resolved a genuine affinity of Gilsonicaris rhenanus from the Lower Devonian Hunsrück Slate, Germany and assigned the specimen to eunicidan polychaetes.
We have chosen the new genus and species Spitiprion khannai Tonarová, Suttner, & Hints to be documented using micro-CT and submicron-CT, respectively. It is also the most characteristic species among the entire scolecodont assemblage obtained. Its maxillae are relatively large and sturdy, even though their length varies around 500 µm. For the isolated maxillae, the resolution of micro-CT turned out to be too low; therefore the submicron-CT was applied (parameters in Table 1; results in videos SOM 1–6). The micro-CT was applied for the partly preserved apparatus, embedded in the piece of rock (sample P/8/69, Fig. 4). The CT analysis allowed us not only to document the outer surface of the maxillae but also to study the myocoele cavity and observe tiny cavities in the maxillae walls. The polygonal internal structure is also visible.
In comparison to other regions, the diversity of ramphoprionids is much lower, with just one genus present. Elsewhere, the family is represented by the genera Ramphoprion Kielan-Jaworowska, 1962, and Protarabellites Stauffer, 1933 (see, e.g., Hints 2000). However, to date all Late Ordovician Gondwanan assemblages studied bear specific endemic taxa among ramphoprionids. In Spiti, it is Spitiprion khannai Tonarová, Suttner, & Hints gen. et sp. nov., in Saudi Arabia “Gen. et sp. indet. 1” of Hints et al. (2015) and Ramphoprion sp. according to Tonarová et al. (2023) in the Prague Basin. Even though, the last species is rather similar to the Laurentian species Ramphoprion cf. deflexus.
Except for the endemic Spitiprion, the Indian assemblage shares a general pattern with other localities from Gondwana. There are recorded polychaetaspids, a very common family in Ordovician and Silurian samples worldwide (Eriksson et al. 2013). Even though the diversity of at least two species of the genus Oenonites Hinde, 1879, is lower than observed in coeval strata (Hints 2000; Tonarová et al. 2023). On the Gonwanan shelf, paulinitids of the genus Kettnerites are first known from the uppermost Katian of Saudi Arabia (Hints et al. 2015), and in the lowermost Hirnantian in the Prague Basin (Tonarová et al. 2023), which is corroborated by the occurrence of Kettnerites sp. in the sample P/8/69. Placognaths, represented by Vistulella and Mochtyella, are relatively rare in the Pin River section. Placognath mochtyellids were also described from the Hirnantian Soom Shale Lagerstätte in South Africa by Whittle et al. (2008), who raised a new genus and species Synaptogenys rietvleiensis. On the other hand, an otherwise common genus in the Ordovician of Baltica, Pistoprion Kielan-Jaworowska, 1966, is missing in Indian samples, similarly as in the Prague Basin (Tonarová et al. 2023), where it first appeared during the Llandovery (Tonarová et al. 2019). Xanioprionids are very rare in Spiti. However, Whittle et al. (2008) described Xanioprion? n. sp. from the Hirnantian Soom Shale Lagerstätte in South Africa, and the family is a common component of Late Ordovician assemblages in Baltica (Hints 2000; Hints et al. 2010).
In summary, the generic composition is rather similar to that in other Gondwanan assemblages (Whittle et al. 2008; Hints et al. 2015; Tonarová et al. 2023), supporting the closer affinity of the Gondwanan assemblage to Laurentia. On the other hand, two families, Hadoprionidae and Kalloprionidae, that are present in Laurentia, have not been recorded in the Indian samples so far. Differences with Laurentia are also among the placognath and ctenognath taxa that were not present in the Upper Ordovician Sylvan Shale in Oklahoma, USA (Eriksson et al. 2005) but were found in the Upper Ordovician of the Cincinnatian region (Eriksson and Bergman 2003) and abundantly on Baltica (Hints 2000).
The dark colour of chitinozoan vesicles supplementary to a regional conodont colour alteration index (CAI) of 4–5 (Suttner 2003) and 4 (Myrow et al. 2019), suggests a considerable thermal exposure (Tricker et al. 1992). That supports the suggestion of Sinha and Verniers (2016), who described also very dark vesicles of chitinozoans from the Kumaon region and interpreted the state of preservation as a result of intense tectonic activity during the Himalayan orogenic movement, which resulted in elevated burial temperatures.
Apart from the contribution to the regional thermal history, chitinozoans are useful climate markers, as shown e.g., by Vandenbroucke et al. (2010), who reconstructed palaeoclimatic belts based on the palaeogeographic distribution of Late Ordovician chitinozoan assemblages. At the Pin River section, five chitinozoan genera were identified and compared regionally to the findings from the closely located Parahio River section by Wang et al. (2021). Our results show a less diverse assemblage than that of Wang et al. (2021), lacking the genera Euconochitina, Cyathochitina, and Ancyrochitina. Another two genera missing in the Pin River assemblage are Angochitina and Belonechitina, which were described from the Shiala Formation in Kumaon by Sinha et al. (2011) and Sinha and Verniers (2016). On the other hand, representatives of Conochitina were documented from both Spiti and Kumaon.
Specimens of Acanthochitina cf. cancellata most closely resemble Acanthochitina? cancellata of Martin (1983) from lower Katian of Quebec in test outline and size. However, the type specimen of Acanthochitina? cancellata, together with the subsequent reports from Quebec (Achab 1978; Asselin et al. 2004), is photographed using an optical microscope; thus, fine morphological details and the ornamentation of this species have remained poorly known.
The second common species is Spinachitina suecica, which, according to previous publications, is mainly reported from the Sandbian to lower Katian. The type specimen comes from the middle Sandbian upper Dalby Limestone of Sweden (Laufeld 1967). Later, S. suecica was reported from the Macrourus siltstone from the Öland erratics (middle Caradocian according to Grahn 1981), the upper Kukruse to Nabala Baltic regional stages (lower and middle Katian) of Gotland, Sweden (Grahn 1982), the uppermost Uhaku–middle Haljala regional stages (uppermost Darriwilian–mid Sandbian; Middle–Upper Ordovician) of Estonia (Nõlvak and Bauert 2006). The material recovered in the present study shows some morphological variability, from a typical shape of the holotype (Fig. 5H) to a more slender form (Fig. 5M). The slender form also resembles Spinachitina sp. aff. coronata from the Ancyrochitina merga biozone (middle upper Katian) of Central Anti-Atlas, Morocco (Bourahrouh et al. 2004). However, the type material of Spinachitina coronata recovered in Baltica is characterised by a long and slender test of around 300 microns and a poorly developed flexure. Generally, the chitinozoan assemblage from the Takche Formation indicates a Late Ordovician age.
Conclusions
•
Scolecodonts and chitinozoans from the Upper Ordovician Pin Member of
the Takche Formation (Katian, Amorphognathus
ordovicicus Conodont Biozone) at the Pin
River section, Pin Valley, Spiti, northern India, are reported.
• A collection of approx.
60 scolecodonts, assigned to Oenonites, Atraktoprion, Vistulella,
Kettnerites, Mochtyella,
Xanioprion, and the new genus Spitiprion,
and approx. 50 chitinozoans belonging to Acanthochitina,
Conochitina,
Lagenochitina?, Spinachitina,
and Tanuchitina were recovered.
• A new ramphoprionid genus
and species, Spitiprion khannai Tonarová,
Suttner, & Hints is described based on a partly preserved maxillary
cluster and isolated maxillae. To date, it is endemic to the Himalayan
region.
• Comparison with other Gondwanan jawed polychaete faunas, e.g., from the Hirnantian Soom Shale in South Africa, indicates that both areas, even though located on opposite sides of the Gondwanan shelf, have representatives belonging to Mochtyellidae and Xanioprionidae in common. Geographically closer assemblages from Saudi Arabia share besides placognath jaws even more taxa at the genus level including Atraktoprion, Kettnerites, and Oenonites.
• This study is the first to present submicron-CT data on scolecodonts. We have confirmed the usefulness of this technique on further jawed polychaete studies, contributing to the establishment of 3D virtual scolecodont type collections.
Acknowledgements
The authors gratefully acknowledge the thorough reviews by Graciela Susana de la Puente CONICET, Buenos Aires, Argentina) and Anthony Butcher (University of Portsmouth, UK) that greatly improved the manuscript. We would like to thank Zuzana Heřmanová (National Museum in Prague, Czech Republic) and Victory Jaques (CEITEC) for the initial testing of computed tomography techniques. JK thanks to the support of grant FSI-S-23-8389 provided by Brno University of Technology and support of the Faculty of Mechanical Engineering at the Brno University of Technology. This paper is a contribution to IGCP project 735 “Rocks and the Rise of Ordovician Life: Filling knowledge gaps in the Early Palaeozoic Biodiversification”. This paper is also a contribution to the Strategic Research Plan of the Czech Geological Survey (DKRVO/ČGS 2023–2027, internal project No. 311410). This research was supported by the Czech Science Foundation through project 23-05944K, and Estonian Research Council project PRG1701. We acknowledge CzechNanoLab Research Infrastructure supported by MEYS CR (LM2023051).
Authors’ contributions
Conceptualisation, funding acquisition, and project administration PT; resources and data curation PT, TJS; formal analysis PT, OH, YL; investigation PT, OH, TJS, YL; methodology and software PT, MZ, MK, TZ, JK; visualization PT, MZ, MK, TZ, JK, TJS; writing: original draft, review and editing PT, TJS, OH, MZ, MK, YL, TZ, JK, EK.
References
Abel, R.L., Laurini, C.R., and Richter, M. 2012. A palaeobiologist’s guide to “virtual” micro-CT preparation. Palaeontologia Electronica 15 (2): 15.2.6T. Crossref
Achab, A. 1978. Sur quelques chitinozoaires de la Formation de Vauréal et de la Formation de Macasty (Ordovicien supérieur), Ile d’Anticosti, Québec, Canada. Review of Palaeobotany and Palynology 25: 295–314. Crossref
Al-Hajri, S. 1995. Biostratigraphy of the Ordovician chitinozoa of northwestern Saudi Arabia. Review of Palaeobotany and Palynology 89: 27–48. Crossref
Al-Shawareb, A., Miller, M., and Vecoli, M. 2017. Late Ordovician (Katian) chitinozoans from northwest Saudi Arabia: Biostratigraphic and paleoenvironmental implications. Revue de Micropaléontologie 60: 333–369. Crossref
Asselin, E., Achab, A., and Soufiane, A. 2004. Biostratigraphic significance of lower Paleozoic microfaunas from eastern Canada. Canadian Journal of Earth Sciences 41: 489–505. Crossref
Audouin, J.V. and Milne Edwards, H. 1833. Classification des Annélides et description de celles qui habitent les côtes de la France. Annales des sciences naturelles, Paris. séries 1: 187–247. Crossref
Bagati, T.N. 1990. Lithostratigraphy and facies variation in the Spiti basin (Tethys), Himachal Pradesh, India. Journal of Himalayan Geology 1: 35–47.
Beesley, P., Ross, G.J.B., and Glasby, C.J. (eds.) 2000. Polychates and Allies: The Southern Synthesis. Vol. 4A: Polychaeta, Myzostomida, Pogonophora, Echiura, Sipuncula. 420 pp. CSIRO Publishing, Melbourne.
Bergman, C.F. 1989. Silurian paulinitid polychaetes from Gotland. Fossils and Strata 25: 1–128. Crossref
Bergman, C.F. 1991. Revision of some Silurian paulinitid scolecodonts from western New York. Journal of Paleontology 65: 248–254. Crossref
Bhargava, O.N. and Bassi, U.K. 1986. Silurian reefal buildups, Spiti-Kinnaur, Himachal Himalaya, India. Facies 15: 35–52. Crossref
Bhargava, O.N. and Bassi, U.K. 1998. Geology of Spiti-Kinnaur Himachal Himalaya. Memoir Geological Survey of India 124: 1–210.
Bhargava, O.N., Srivastava, R.N., and Gadhoke, S.K. 1991. Proterozoic–Palaeozoic Spiti sedimentary basin. In: S.K. Tandon, C.C. Pant, and S.M. Casshyap (eds.), Sedimentary Basins of India: Tectonic Context. 236–260. Gyanodaya Prakashan, Nainital.
Bond, D.P.G. and Grasby, S.E. 2020. Late Ordovician mass extinction caused by volcanism, warming, and anoxia, not cooling and glaciation. Geology 48: 777–781. Crossref
Bordoloi, S. 2002. Petrography of Pin Formation, Spiti, Tethyan Himalaya, Himachal Pradesh. 43 pp. Unpublished M.Sc. Thesis, University of Delhi, Delhi.
Bourahrouh, A., Paris, F., and Elaouad-Debbaj, Z. 2004. Biostratigraphy, biodiversity and palaeoenvironments of the chitinozoans and associated palynomorphs from the Upper Ordovician of the Central Anti-Atlas, Morocco. Review of Palaeobotany and Palynology 130: 17–40. Crossref
Brenchley, P.J., Carden, G.A., Hints, L., Kaljo, D., Marshall, J.D., Martma, T., Meidla, T., and Nõlvak, J. 2003. High-resolution stable isotope stratigraphy of Upper Ordovician sequences: Constraints on the timing of bioevents and environmental changes associated with mass extinction and glaciation. Geological Society of America Bulletin 115: 89–104. Crossref
Budaeva, N. and Zanol, J. 2021. Eunicida Dales, 1962. In: G. Purschke, W. Westheide, and M. Böggemann (eds.), Handbook of Zoology Annelida. Volume 3: Sedentaria III, Errantia I, 353–360. De Gruyter, Berlin. Crossref
Chaubey, R.S., Singh, B.P., Vinn, O., Bhargava, O.N., Prasad, S.K., and Sati, M. 2023. Integrated lithofacies, microfacies and sequence stratigraphic framework of the Takche Formation (Ordovician–Early Silurian), Pin Valley, Spiti Himalaya, India. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 307: 51–64. Crossref
Chaubey, R.S., Vinn, O., Singh, B.P., Bgargava, O.N., Prasad, S.K., and Kishore, N. 2019. Warm-water Dasycladaceae algae from the Late Ordovician of the Parahio Valley, Spiti, India. Estonian Journal of Earth Sciences 68: 45–53. Crossref
Claparède, É. 1870. Les Annélides Chétopodes du Golfe de Naples. Supplément. Mémoires de la Société de physique et d’histoire naturelle de Genève 20: 365–542. Crossref
Clemo, W.C. and Dorgan, K.M. 2017. Functional morphology of eunicidan (Polychaeta) jaws. The Biological Bulletin 233: 227–241. Crossref
Dales, R.P. 1963. Annelids. 200 pp. Hutchinson, London.
De la Puente, S.G. and Astini, R.A. 2023. Ordovician chitinozoans and review on basin stratigraphy, biostratigraphy and paleobiogeography of northern Argentina along the Proto-Andean margin. Geobios 81: 199–226. Crossref
De la Puente, G.S., Paris, F., and Vaccari, N.E. 2020. Latest Ordovician–earliest Silurian chitinozoans from the Puna region, north-western Argentina (Western Gondwana). Bulletin of Geosciences 95: 391–418. Crossref
Elaouad-Debbaj, Z. 1984. Chitinozoaires ashgilliens de l’Anti-Atlas (Maroc). Geobios 17: 45–75. Crossref
Eller, E.R. 1942. Scolecodonts from the Erindale Upper Ordovician, at Sreetville, Ontario. Annals of the Carnegie Museum 29: 241–270. Crossref
Eller, E.R. 1945. Scolecodonts from the Trenton Series (Ordovician) of Ontario, Quebec, and New York. Annals of the Carnegie Museum 30: 119–212. Crossref
Eriksson, M. 2001. Silurian ramphoprionid polychaetes from Gotland, Sweden. Journal of Paleontology 75: 993–1015. Crossref
Eriksson, M. 2002. The palaeobiogeography of Silurian ramphoprionid polychaete annelids. Palaeontology 45: 985–996. Crossref
Eriksson, M. and Bergman, C.F. 2003. Late Ordovician jawed polychaete faunas of the type Cincinnatian Region, U.S.A. Journal of Paleontology 77: 509–523. Crossref
Eriksson, M.E., Bergman, C.F., and Jeppsson, L. 2004. Silurian scolecodonts. Review of Palaeobotany and Palynology 131: 269–300. Crossref
Eriksson, M.E., Hints, O., Paxton, H., and Tonarová, P. 2013. Chapter 18 Ordovician and Silurian polychaete diversity and biogeography. Geological Society, London, Memoirs 38: 265–272. Crossref
Eriksson, M.E., Leslie, S.A., and Bergman, C.F. 2005. Jawed polychaetes from the Upper Sylvan Shale (Upper Ordovician), Oklahoma, USA. Journal of Paleontology 79: 486–496. Crossref
Eriksson, M.E., Parry, L.A., and Rudkin, D.M. 2017. Earth’s oldest “Bobbit worm”—gigantism in a Devonian eunicidan polychaete. Scientific Reports 7 (1): 43061. Crossref
Fauchald, K. 2013. Worms, Annelida. In: S.A. Levin (ed.), Encyclopedia of Biodiversity, Second Edition, 408–419. Elsevier. Crossref
Fortey, R.A. and Cocks, L.R.M. 2005. Late Ordovician global warming—The Boda event. Geology 33: 405–408. Crossref
Fuchs, G. 1982. The geology of the Pin Valley in Spiti, H. P., India. Jahrbuch der Geologischen Bundesanstalt 124: 325–359.
Ghavidel-Syooki, M. 2001. Biostratigraphy and palaeogeography of Late Ordovician and Early Silurian chitinozoans from the Zagros Basin, Southern Iran. Historical Biology 15: 29–39. Crossref
Ghavidel-Syooki, M. 2023. Biostratigraphy and palaeogeographic implications of Ordovician and Silurian chitinozoa from the High Zagros Mountains, Northern Persian Gulf, Iran. Palynology 47: 2149631. Crossref
Ghavidel-Syooki, M. and Piri-Kangarhahi, M.H. 2021. Biostratigraphy of acritarchs, chitinozoans, and miospores from Upper Ordovician sequences in Kuh-e Boghou, southwest of Kashmar, eastern central Iran: Stratigraphic and paleogeographic implications. Review of Palaeobotany and Palynology 284: 104337. Crossref
Ghavidel-Syooki, M. and Winchester-Seeto, T. 2002. Biostratigraphy and palaeogeography of Late Ordovician chitinozoans from the northeastern Alborz Range, Iran. Review of Palaeobotany and Palynology 118: 77–99. Crossref
Goel, R.K. and Nair N.G.K. 1977. The Spiti Ordovician–Silurian succession. Journal of Geological Society of India 18: 47–48.
Goel, R.K. and Nair N.G.K. 1982. Lower Palaeozoic sequence in the Pin Valley, Spiti. Geological Survey of India Miscellaneous Publications 41: 13–15.
Grahn, Y. 1981. Middle Ordovician Chitinozoa from Öland. Sverige geologiska undersökning. Serie C 784: 1–51.
Grahn, Y. 1982. Caradocian and Ashgillian Chitinozoa from the Subsurface of Gotland. Sveriges geologiska undersökning, Serie C 788: 1–66.
Grahn, Y. 1992. Ordovician chitinozoa and biostratigraphy of Brazil. Geobios 25: 703–723. Crossref
Griesbach, C.L. 1891. Geology of the central Himalayas. Memoirs of the Geological Survey of India 23: 1–232.
Grube, A.E. 1850. Die Familien der Anneliden. Archiv für Naturgeschichte, Berlin 16: 249–364
Gueriau, P., Parry, L.A., and Rabet, N. 2023. Gilsonicaris from the Lower Devonian Hunsrück slate is a eunicidan annelid and not the oldest crown anostracan crustacean. Biology Letters 19 (8): 20230312. Crossref
Hayden, H.H. 1904. The geology of Spiti with parts of Bashahr and Rupshu. Memoirs of the Geological Survey of India 36: 1–129.
Hinde, G.J. 1879. On Annelid jaws from the Cambro-Silurian, Silurian, and Devonian Formations in Canada and from the Lower Carboniferous in Scotland. Quarterly Journal of the Geological Society 35: 370–389. Crossref
Hints, L., Hints, O., Kaljo, D., Kiipli, T., Männik, P., Nõlvak, J., and Pärnaste, H. 2010. Hirnantian (latest Ordovician) bio- and chemostratigraphy of the Stirnas-18 core, western Latvia. Estonian Journal of Earth Sciences 59: 1–24. Crossref
Hints, O. 1998. Late Viruan (Ordovician) polychaete jaws from North Estonia and the St. Petersburg Region. Acta Palaeontologica Polonica 43: 471–516.
Hints, O. 2000. Ordovician eunicid polychaetes of Estonia and surrounding areas: review of their distribution and diversification. Review of Palaeobotany and Palynology 113: 41–55. Crossref
Hints, O., Ainsaar, L., Lepland, A., Liiv, M., Männik, P., Meidla, T., Nõlvak, J., and Radzevičius, S. 2023. Paired carbon isotope chemostratigraphy across the Ordovician–Silurian boundary in central East Baltic: Regional and global signatures. Palaeogeography, Palaeoclimatology, Palaeoecology 624: 111640. Crossref
Hints, O. and Tonarová, P. 2023. A diverse Hirnantian scolecodont assemblage from northern Estonia and resilience of polychaetes to the end-Ordovician mass extinction. Estonian Journal of Earth Sciences 72: 46–49. Crossref
Hints, O., Paris, F., and Al-Hajri, S. 2015. Late Ordovician scolecodonts from the Qusaiba-1 core hole, central Saudi Arabia, and their paleogeographical affinities. Review of Palaeobotany and Palynology 212: 85–96. Crossref
Hints, O., Tonarová, P., and Desrochers, A. 2016. Late Ordovician jaw-bearing polychaetes from Anticosti Island, eastern Canada, and their biogeographic significance. Melchin, M. (ed.), Canadian Journal of Earth Sciences 53: 731–738. Crossref
Hints, O., Tonarová, P., Eriksson, M.E., Rubinstein, C. V., and de la Puente, G.S. 2017. Early Middle Ordovician scolecodonts from north-western Argentina and the emergence of labidognath polychaete jaw apparatuses. Palaeontology 60: 583–593. Crossref
Hutchings, P.A. 1986. Biological destruction of coral reefs. Coral Reefs 4: 239–252. Crossref
Hubmann, B. and Suttner, T. 2007. Calcimicrobes and calcareous algae of the Pin Formation (NW Himalayas, India): a contribution to the “Ordovician Flora”. Acta Palaeontologica Sinica (Special Issue) 46 (Supplement): 188–193.
Jansonius, J. and Craig, J.H. 1971. Scolecodonts: I. Descriptive terminology and revision of systematic nomenclature; II. Lectotypes, new names for homonyms, index of species. Bulletin of Canadian Petroleum Geology 19: 251–293.
Kato, M., Goel, R.K., and Srivastava, S.S. 1987. Ordovician Algae from Spiti, India. Journal of the Faculty of Science, Hokkaido University, Series IV 22: 313–323.
Khanna, A.K. 1984. Scolecodonts from Late Ordovician–Early Silurian sequence of Spiti Valley, Tethys Himalaya. Proceedings of Fifth Indian Geophytological Conference, 24–28. The Palaeobotanical Society, Lucknow.
Khanna, A.K. and Sah, S.C.D. 1983. Chitinozoa of the Pin Dolomite, Tethys Himalaya. Himalayan Geology 11: 256–262.
Khanna, A.K., Sinha, A.K., and Sah, S.C.D. 1985. Yong Limestone of Tethys Himalaya—its stratigraphic status and palynological fossils. Journal of the Geological Society of India 26: 191–198.
Kielan-Jaworowska, Z. 1962. New Ordovician genera of polychaete jaw apparatuses. Acta Palaeontologica Polonica 7: 291–332.
Kielan-Jaworowska, Z. 1966. Polychaete jaw apparatuses from the Ordovician and Silurian of Poland and a comparison with modern forms. Palaeontologia Polonica 16: 1–152.
Kröger, B., Ebbestad, J.O., and Lehnert, O. 2016. Accretionary mechanisms and temporal sequence of formation of the Boda Limestone Mud-Mounds (Upper Ordovician), Siljan District, Sweden. Journal of Sedimentary Research 86: 363–379. Crossref
Lange, F.W. 1947. Annelidos poliquetos dos folhelhos devonianos do Parana. Arquivos do Museu Paranaese 6 (5): 161–230.
Laufeld, S. 1967. Caradocian Chitinozoa from Dalarna, Sweden. Geologiska Föreningen i Stockholm Förhandlingar 89: 275–349. Crossref
Lefebvre, B., Makhlouf, Y., Nardin, E., Nedjari, A., and Touzeau, A. 2014. Impact of the Boda Event (Late Ordovician) on high-latitude Peri-Gondwanan faunas: An Echinoderm Perspective. In: R. Rocha, J. Pais, J.C. Kullberg, and S. Finney (eds.), STRATI 2013, First International Congress on Stratigraphy at the Cutting Edge of Stratigraphy, Conference Proceedings, 49–52. Springer, Cham. Crossref
Liang, Y., Tang, P., Wang, G.-X., Yan,
G.-Z., and Wang, Q. 2023. Middle–Late Ordovician chitinozoans from
Songliang of Qiaojia, western South China, and their biostratigraphic
implications. Palaeoworld 32: 287–302.
Crossref
Lu, Y., Hao, F., Yan, D., and Lu, Y. 2021. Volcanism-induced late Boda warming in the Late Ordovician: Evidence from the Upper Yangtze Platform, South China. Palaeogeography, Palaeoclimatology, Palaeoecology 578: 110579. Crossref
Martin, F. 1983. Chitinozoaires et acritarches ordoviciens de la plate-forme du Saint-Laurent (Québec et Sud-Est de l’Ontario). Geological Survey of Canada, Bulletin 310: 1–59. Crossref
Männil, R. and Zaslavskaya, N. 1985. Silurijskie polihety severa Sibiri. Trudy Instituta Geologii i Geofiziki SO AN SSSR 615: 98–119.
Myrow, P.M., Fike, D.A., Malmskog, E., Leslie, S. A., Zhang, T.R., Singh, B.P., Chaubey, R.S., and Prasad, S.K. 2019. Ordovician–Silurian boundary strata of the Indian Himalaya: Record of the latest Ordovician Boda event. GSA Bulletin 131: 881–898. Crossref
Nõlvak, J. and Bauert, G. 2006. Distribution of Ordovician chitinozoans. Estonian Geological Sections Bulletin 7: 9–11.
Ontiveros, D.E., Beaugrand, G., Lefebvre, B., Markussen Marcilly, C., Servais, T., and Pohl, A. 2023. Impact of global climate cooling on Ordovician marine biodiversity. Nature Communications 14: 6098. Crossref
Pandey, S. and Parcha, S.K. 2018. Calcareous algae from the Ordovician succession (Thango Formation) of the Spiti Basin, Tethys Himalaya, India. Acta Palaeobotanica 58: 97–106. Crossref
Paris, F. 1988. Late Ordovician and early Silurian chitinozoans from central and southern Cyrenaiea (Libya). In: A. El Arnauti, B. Owens, and B. Thusu (eds.), Subsurface Palynostratigraphy of Northeast Libya, 61–71. Garyouis University Publishing, Benghazi.
Paris, F., Verniers, J., Miller, M.A., Al-Hajri, S., Melvin, J., and Wellman, C. 2015. Late Ordovician–earliest Silurian chitinozoans from the Qusaiba-1 core hole (North Central Saudi Arabia) and their relation to the Hirnantian glaciation. Review of Palaeobotany and Palynology 212: 60–84. Crossref
Parry, L. A., Edgecombe, G.D., Sykes, D., and Vinther, J. 2019. Jaw elements in Plumulites bengtsoni confirm that machaeridians are extinct armoured scaleworms. Proceedings of the Royal Society B: Biological Sciences 286 (1907): 20191247. Crossref
Paxton, H. 2009. Phylogeny of Eunicida (Annelida) based on morphology of jaws. Zoosymposia 2: 241–264. Crossref
Reed, F.R.C. 1912. Ordovician and Silurian fossils from the central Himalayas. Memoirs of the Geological Survey of India. Palaeontologia Indica, Ser. 15, 7 (2): 1–168.
Salter, J.W. and Blanford, H.F. 1865. Palaeontology of Niti in the Northern Himalayas Being Descriptions and Figures of the Palaeozoic and Secondary Fossils Collected by Colonel Richard Strachey. 112 pp. O.T. Cutter, Military Orphan Press, Calcutta.
Schallreuter, R., Hinz-Schallreuter, I., and Suttner, T. 2008. New Ordovician ostracodes from Himalaya and their palaeobiological and palaeogeographical implications. Revue de Micropaléontologie 51 (3): 191–204. Crossref
Schmidt, A., Skeffington, R.A., Thordarson, T., Self, S., Forster, P.M., Rap, A., Ridgwell, A., Fowler, D., Wilson, M., Mann, G.W., Wignall, P.B., and Carslaw, K.S. 2016. Selective environmental stress from sulphur emitted by continental flood basalt eruptions. Nature Geoscience 9: 77–82. Crossref
Schröer, L., Vandenbroucke, T.R.A., Hints, O., Steeman, T., Verniers, J., Brett, C.E., Cramer, B.D., and McLaughlin, P.I. 2016. A Late Ordovician age for the Whirlpool and Power Glen formations, New York. Canadian Journal of Earth Sciences 53: 739–747. Crossref
Scotese, C.R. 2000. Paleomap Project [available on line, http://www.scotese.com]
Servais, T., Cascales-Miñana, B., Harper, D.A.T., Lefebvre, B., Munnecke, A., Wang, W., and Zhang, Y. 2023. No (Cambrian) explosion and no (Ordovician) event: A single long-term radiation in the early Palaeozoic. Palaeogeography, Palaeoclimatology, Palaeoecology 623: 111592. Crossref
Servais T., Owen A.W., Harper D.A.T., Kröger B., and Munnecke A. 2010 The Great Ordovician Biodiversification Event (GOBE): The palaeoecological dimension. Palaeogeography, Palaeoclimatology, Palaeoecology 294: 99–119. Crossref
Shabbar, H., Saxena, A., Gupta, S., Singh, K.J., and Goswami, S. 2022. The first record of cornulitids tubeworms from the early Late Ordovician of Spiti, Tethyan Himalaya, India. Historical Biology 34: 176–187. Crossref
Shabbar, H., Saxena, A., Tinn, O., Gupta, S., and Singh, K.J. 2023. Non-calcified warm-water marine macroalgae from the Ordovician strata of Spiti, Tethys Himalaya, India. Palaeoworld 32: 396–410. Crossref
Sinha, H.N. and Verniers, J. 2016. Discovery of the chitinozoans Belonechitina capitata from the Shiala Formation of northeastern Garhwal-Kumaon Tethys Himalaya, Pithoragarh District, Uttrakhand, India. Geoscience Frontiers 7: 859–864. Crossref
Sinha, H.N., Vandenbroucke, T.R.A. and Verniers, J. 2011. First Ordovician chitinozoans from Indian Gondwana—new evidence from the Shiala Formation. Review of Palaeobotany and Palynology 167: 117–122. Crossref
Srikantia, S.V. 1974. Geology of Part of Lahaul and Spiti, H.P. With Preliminary Appraisal of Phosphorite Occurrence. Unpublished Report. 30 pp. Geological Survey of India, Lucknow.
Srikantia, S.V. 1977. Sedimentary cycles in the Himalaya and their significance on the orogenic evolution of the mountain belt. Colloques International Centre Naturalle Recherche Scientifique 268: 395–408.
Srikantia, S.V. and Bhargava, O.N. 2018. Stratigraphic nomenclature of early Palaeozoic in the Spiti Himalaya: cobwebs cleared. Journal Palaeontological Society of India 63: 233–241.
Stauffer, C.R. 1933. Middle Ordovician Polychaeta from Minnesota. Bulletin of the Geological Society of America 44: 1173–1218. Crossref
Stoliczka, F. 1866. Geological section across the Himalayan Mountains, from Wangtu bridge on the River Sutlej to Sungdo on the Indus: with an account of the formations of Spiti, accompanied by a revision of all known fossils from that district. Memoirs of the Geological Survey of India 5: 1–173.
Suttner, T.J. 2003. Die Pin-Formation (Altpaläozoikum) von Muth, Spiti (Indischer Himalaya): Stratigraphie und Fazies. 81 pp. Unpublished Diploma Thesis, University of Vienna, Vienna.
Suttner, T.J. 2007. The Upper Ordovician to Lower Silurian Pin Formation (Farka Muth, Pin Valley, north India)—a formal discussion and redefinition of its controversial type-section. Acta Palaeontologica Sinica 46 (Supplement): 460–465. Crossref
Suttner, T.J. and Ernst, A. 2007. Upper Ordovician bryozoans of the Pin Formation (Spiti Valley, northern India). Palaeontology 50: 1485–1518.
Suttner, T. and Hubmann, B. 2005. Calcimicrobes and Green algae from Spiti Area, Northern India (Northern Gondwana Low Latitude Shelf Areas). Climatic and evolutionary controls on Palaeozoic reefs and bioaccumulations, Abstract volume, 53–54. Muséum National d’Histoire Naturelle, Paris.
Suttner, T. and Kido, E. 2011. Upper Ordovician and Lower Silurian corals from the Pin Formation (Spiti Valley, Northern India). In: M. Aretz, S. Delcuée, and J. Denazer (eds.), 11th Symposium on Fossil Cnidaria and Porifera, Liege, August 19–29, 2011, Abstracts. Kölner Forum für Geologie und Paläontologie 19: 169–172.
Suttner, T.J. and Kröger, B. 2006. Late Ordovician to Early Silurian Gondwanan cephalopods of the Pin Formation (Spiti, northern India). Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 2006 (10): 595–612. Crossref
Suttner, T., Draganits, E., and Krystyn, L. 2001. Lithostratigraphy and biostratigraphy of the Pin formation type section (Spiti, NW India). Abstract Volume. Journal of Asian Earth Sciences 19 (3a): 65–66.
Suttner, T., Hubmann, B., and Bhargava, O.N. 2005. Late Ordovician Calcimicrobes and Green-algae from Spiti (Himalayas, N-India). In: B. Hubmann and W.E. Piller (eds.), 75. Jahrestagung der Paläontologischen Gesellschaft – Beitragskurzfassungen. Berichte des Institutes für Erdwissenschaften der Karl-Franzens-Universität Graz 10: 130–131.
Suttner, T., Hubmann, B., and Bhargava, O. 2007a. Patch reefs and fringing bioaccumulations, India. In: E. Vennin, M. Aretz, F. Boulvain, and A. Munnecke (eds.), Facies from Palaeozoic Reefs and Bioaccumulations. Chapter II: Ordovician. Mémoires du Muséum national d’Histoire naturelle 195: 95–97.
Suttner, T.J., Lehnert, O., Joachimski, M., and Buggisch, W. 2007b. Recognition of the Boda Event in the Pin Formation of northern India based on new δ13C and conodont data. Acta Palaeontologica Sinica 46 (Supplement): 466–470.
Szaniawski, H. 1996. Scolecodonts. In: J. Jansonius and D.C. McGregor (eds.), Palynology Principles and Applications Vol. 1, 337–354. American Association of Stratigraphic Palynologists Foundation.
Szaniawski, H. and Wrona, R.M. 1973. Polychaete jaw apparatuses and scolecodonts from the Upper Devonian of Poland. Acta Palaeontologica Polonica 18: 223–267.
Tasáryová, Z., Schnabl, P., Čížková, K., Pruner, P., Janoušek, V., Rapprich, V., Štorch, P., Manda, Š., Frýda, J., and Trubač, J. 2014. Gorstian palaeoposition and geotectonic setting of Suchomasty Volcanic Centre (Silurian, Prague Basin, Teplá-Barrandian Unit, Bohemian Massif). GFF 136: 262–265. Crossref
Thusu, B., Rasul, S., Paris, F., Meinhold, G., Howard, J.P., Abutarruma, Y., and Whitham, A.G. 2013. Latest Ordovician–earliest Silurian acritarchs and chitinozoans from subsurface samples in Jebel Asba, Kufra Basin, SE Libya. Review of Palaeobotany and Palynology 197: 90–118. Crossref
Tonarová, P., Eriksson, M., and Hints, O. 2012. A jawed polychaete fauna from the late Ludlow Kozlowskii event interval in the Prague Basin (Czech Republic). Bulletin of Geosciences 87: 713–732. Crossref
Tonarová, P., Vodrážková, S., Hints, O., Männik, P., Kubajko, M., and Frýda, J. 2019. Llandovery microfossils and microfacies of the Hýskov section, Prague Basin. Fossil Imprint 75: 25–43. Crossref
Tonarová, P., Vodrážková, S., Hints, O., Nõlvak, J., Kubajko, M., and Čáp, P. 2023. Latest Ordovician jawed polychaetes, chitinozoans and depositional environments of the Levín section, Prague Basin, Czech Republic. Geobios 81: 179–198. Crossref
Torsvik, T.H. and Cocks, L.R.M. 2009. The Lower Palaeozoic palaeogeographical evolution of the northeastern and eastern peri-Gondwanan margin from Turkey to New Zealand. Geological Society, London, Special Publications 325: 3–21. Crossref
Torsvik, T.H. and Cocks, L.R.M. 2013. Gondwana from top to base in space and time. Gondwana Research 24: 999–1030. Crossref
Tricker, P.M., Marshall, J.E.A., and Badman, T.D. 1992. Chitinozoan reflectance: a Lower Palaeozoic thermal maturity indicator. Marine and Petroleum Geology 9: 302–307. Crossref
Vandenbroucke, T.R.A., Armstrong, H.A., Williams, M., Paris, F., Sabbe, K., Zalasiewicz, J.A., Nõlvak, J., and Verniers, J. 2010. Epipelagic chitinozoan biotopes map a steep latitudinal temperature gradient for earliest Late Ordovician seas: implications for a cooling Late Ordovician climate. Palaeogeography Palaeoclimatology and Palaeoecology 294: 202–219. Crossref
Vinn, O., Chaubey, R.S., Singh, B.P., Bhargava, O.N., and Prasad, S.K. 2023. The first record of calcitarchs from the Takche Formation (Ordovician–Silurian), Himalaya (India). Proceedings of the Geologists’ Association 134: 590–598. Crossref
Wang, W., Gao, S., Servais, T., Singh, B.P., and Myrow, P. 2021. Preliminary palynological study of the Upper Ordovician Pin Formation in northern Indian Himalaya. Palynology 45: 301–319. Crossref
Webby, B.D., Paris, F., Droser, M.L., and Percival, I.G. (eds.) 2004. The Great Ordovician Biodiversification Event. 484 pp. Columbia University Press, New York. Crossref
Whittle, R., Gabbot, S., Aldridge, R., and Theron, J. 2008. Late Ordovician (Hirnantian) scolecodont clusters from the Soom Shale Lagerstätte, South Africa. Journal of Micropalaeontology 27: 147–159. Crossref
Williams, H.S., Nowlan, G.S., Barnes, C.R., and Batten, R.S.R. 1999. The Ledge section at Cow Head, western Newfoundland as a GSSP candidate for the lower boundary of the second stage of the Ordovician System: new data and discussion of the graptolite, conodont and chitinozoan assemblages. A Report to the IUGS/ICS Subcommission on Ordovician Stratigraphy. June 1999, 1–30.
Zanol, J., Carrera-Parra, L.F., Steiner, T.M., Amaral, A.C.Z., Wiklund, H., Ravara, A., and Budaeva, N. 2021. The Current State of Eunicida (Annelida) Systematics and Biodiversity. Diversity 13 (2): 74. Crossref
Žebera, K. 1935. Les Conodontes et les Scolécodontes du Barrandien. Bulletin international du Academie tchéque 36: 88–96.
Acta Palaeontol. Pol. 69 (2): 199–215, 2024
https://doi.org/10.4202/app.01135.2024