
Early Permian crinoids from Laurasia and their paleogeographic implications
WILLIAM I. AUSICH, MATIJA KRIŽNAR, KAROLINA PASZCZA, İZZET HOŞGÖR, BARTOSZ J. PŁACHNO, and MARIUSZ A. SALAMON
Ausich, W.I., Križnar, M., Paszcza, K., Hoşgör, İ., Płachno, B.J., and Salamon, M.A. 2024. Early Permian crinoids from Laurasia and their paleogeographic implications. Acta Palaeontologica Polonica 69 (3): 447–466.
A new crinoid fauna is described from the Trogkofel Group of the Southern Karavanke Mountains, Slovenia (lower Permian, Artinskian). This fauna is from a shallow platform or upper slope setting and is comprised primarily of isolated aboral cups. One taxon, Protencrinus baliensis, previously described from the lower Permian of Crete, is the only species known from elsewhere. New taxa from the Trogkofel Group include Karavankecrinus bedici gen. et sp. nov., Moapacrinus dovjensis sp. nov., Parastachyocrinus sloveniaensis sp. nov., and Sinocrinus websteri sp. nov. Several taxa are left in open nomenclature, including Dieuryocrinus? sp., Platycrinitidae indet., Eucladida indet. A, and unidentified Crinoidea. If Dieuryocrinus? sp. belongs to Dieuryocrinus, this would expand the range of this genus, previously known from the Viséan (Mississippian) of the United Kingdom. Other genera are well known as lower Permian taxa. In addition, the species-level systematics of Parastachyocrinus is revised, and one new species from the lower Permian of Timor is described, P. wanneri sp. nov.
Key words: Echinodermata, Crinoidea, Permian, Europe, Southern Calcareous Alps, Slovenia.
William I. Ausich [ausich.1@osu.edu; ORDCID: https://orcid.org/0000-0001-7140-5986 ], School of Earth Sciences, Ohio State University, Columbus, OH, USA.
Matija Križnar [mkriznar@pms-lj.si; ORCID: https://orcid.org/0000-0002-9974-4733 ], Slovenian Museum of Natural History, Ljubljana, Slovenia.
Karolina Paszcza [karolinapaszcza@gmail.com; ORCID: https://orcid.org/0000-0002-7663-7476 ] and Mariusz A. Salamon [mariusz.salamon@us.edu.pl; ORCID: https://orcid.org/ 0000-0001-9399-2798 ] (corresponding author), University of Silesia in Katowice, Faculty of Natural Sciences, Będzińska 60, PL-41-200 Sosnowiec, Poland.
İzzet Hoşgör [İhosgor@tpao.gov.tr; ORCID: https://orcid.org/0000-0002-3886-1784 ], Türkiye Petrolleri A.O-TPAO, Exploration Dept., Ankara, Türkiye.
Bartosz J. Płachno [bartosz.plachno@uj.edu.pl; ORCID: https://orcid.org/0000-0001-5579-5101 ], Jagiellonian University in Kraków, Faculty of Biology, Institute of Botany, Gronostajowa Street 9, PL-30-387 Cracow, Poland.
Received 28 March 2024, accepted 26 June 2024, published online 27 September 2024.
Copyright © 2024 W.I. Ausich et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The knowledge of Permian crinoids as compared to Devonian and Carboniferous crinoids, has expanded in recent years. However, it is still relatively inadequate, at least in some areas. The bulk of Permian crinoid studies (qualitatively and quantitatively) is from Asia (China, India, Oman, Pakistan, Thailand, and Timor; Waagen 1887; Wanner 1916, 1924, 1937; Tien 1926; Kirk 1940; Marez Oyens 1940; Moore and Teichert 1978; Chen and Yao 1993; Jell and Willink 1993; Webster and Jell 1993; Webster and Sevastopulo 2007; Webster and Donovan 2012; Webster et al. 2009a, c, 2022; Webster 2012a; Mao et al. 2018), Australia and Oceania (Australia, Tasmania; M’Coy 1847; Sieverts-Doreck 1942; Teichert 1949, 1954; Webster 1987, 1990; Willink 1978, 1979a, b, 1980; Webster and Jell 1992, 1999; Teichert and Webster 1993), South America (Bolivia; Strimple and Moore 1971; Burke and Pabian 1978) and North America (Canada, Mexico, USA; Weller 1930; Moore 1939; Moore and Plummer 1940; Lane and Webster 1966; Webster and Lane 1967, 2007; Strimple 1971; Pabian and Strimple 1974; Webster et al. 2009b). In addition, Valette (1934), Termier and Termier (1949, 1958, 1977) and Lane (1979) documented two camerates and two mespilocrinid flexibles from the Wordian (Guadalupian) at Djebel Tebaga, Tunisia (North Africa).
In Europe, complete or nearly complete (possessing cups and/or crowns) Permian crinoids have come from Norway, Russia, Italy, and Greece. Holtedahl (1911), based on an articulated cup from lower Permian of Spitsbergen (Norway), erected Platycrinites spitzbergensis. However, Gorzelak et al. (2013) noted that this species is exclusively known from the Carboniferous. From lower Permian of the same area Gorzelak et al. (2013) described a nearly complete specimen assigned to Catacrinidae gen. indet. Yakovlev (1926, 1927, 1930), Yakovlev and Ivanov (1956) and Arendt (1968, 1970) mentioned several upper Permian crinoid taxa from some areas in the Urals of Russia (Krasnoufimsk city, Pechora Basin, Kolva River). Additionally, Moore and Strimple (1978a, 1978b, respectively) indicated the presence of Apographiocrinus and Metallagecrinus in the upper Permian strata of the European part of Russia, although they did not specify the exact locality. Gregorio (1930), Yakovlev (1934), Ramovš and Sieverts-Doreck (1968), and Strimple and Sevastopulo (1982) recorded several crinoids from the upper Permian, including microcrinoids from Italy (Sicily). Webster (2012b) described five new taxa from the lower Permian of Greece (Crete) and concluded that at the generic level the fauna had greater affinity with North America than with the fauna from Timor.
The presence of crinoid ossicles in lower Permian of Slovenia have been noted by Ramovš and Sieverts-Doreck (1968), Ramovš and Kochansky-Devide (1979) and Novak and Skaberne (2009). Ramovš and Sieverts-Doreck (1968) mentioned Palermocrinus jaekeli, Entrochus sp. 1, Entrochus sp. 2, “Poteriocrinus” sp. indet. and identified a new species, Palermocrinus togatus Ramovš & Sieverts-Doreck, 1968 (see also Lach et al. 2013). Unfortunately, all of them were documented from only fragments of stems, and their value has been questioned (see also e.g., Lane and Webster 1980; Holterhoff 1997). Additionally, Gorzelak et al. (2013) noted that such isolated ossicles are common and widespread in Permian rocks. Nonetheless, an artificial (parataxonomic) classification scheme based on isolated columnals and pluricolumnals has been established and used mainly for the biostratigraphic purposes (e.g., Moore and Jeffords 1968; Donovan 1986, 1989, 1995; Głuchowski 2002 and literature cited therein).
Here, for the first time, we systematically evaluate crinoids known from the lower Permian (Cisuralian, Artinskian) of Slovenia. Identified taxa include the following: Dieuryocrinus? sp., Parastachyocrinus sloveniaensis sp. nov., Karavankecrinus bedici gen. et sp. nov., Moapacrinus dovjensis sp. nov., Sinocrinus websteri sp. nov., and Protencrinus baliensis Webster, 2012b. In additional, several other taxa left in open nomenclature are discussed; and the genus Parastachyocrinus Wanner, 1949, is re-evaluated with one new species from Timor described.
The majority of known Permian crinoids are from Timor, where the chronostratigraphic position of crinoid occurrences is basically unresolved (Webster and Jell 1992, 1999; Webster 1998a, b; Charlton et al. 2002). Thus, description of new Permian crinoid faunas from sections with well-defined chronostratigraphy is especially important in order to understand the temporal, palaeoenvironmental, and palaeogeographic history of crinoids leading up to Permian extinctions, during which crinoids nearly underwent total extinction.
Institutional abbreviations.—CrdKap, Geologisch-Paläontologisches Institut, Freiburg im Breisgau, Germany; GMJ: PAL, Upper Sava Museum, Jesenice, Slovenia; PMSLJ, Slovenian Museum of Natural History, Ljubljana, Slovenia; RGM, Naturalis Biodiversity Center, Leiden, Netherlands.
Other abbreviations.—ACH, aboral cup height; ACW, aboral cup width; CaW, calyx width.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:D6FB3782-2A7F-4ABD-97BD-A65FFCAFB872.
Geological and stratigraphical setting
The outcrops of the upper Paleozoic (upper Carboniferous‒lower Permian) fossiliferous shallow marine deposits in the Southern Karavanke Mountains are scattered between Solčava and Kranjska Gora (Fig. 1). The lower Permian beds are best exposed north of Tržič, in the famous fossil site of the Dovžanova soteska and Jelendol Valleys (Novak and Skaberne 2009; Novak et al. 2019). To the west of the Southern Karavanke Mountains, the lower Permian beds are mainly exposed in Javorniški Rovt and Pristava and between Črni vrh and Dovje villages (Novak and Skaberne 2009). On the opposite side of Sava Valley in the Julian Alps, small outcrops are present in the vicinity of Kranjska Gora and also farther west toward the Slovenian-Italian border. In the Trogkofel Beds (after Ramovš and Kochansky-Devide 1979) from some of these localities, Artinskian microfossils have been noted (such as fusulinid foraminifera and dasycladacean algae), as well as ossicles of the crinoid Palermocrinus togatus (Ramovš and Kochansky-Devide 1979; Novak and Skaberne 2009) and some scattered macrofossils.
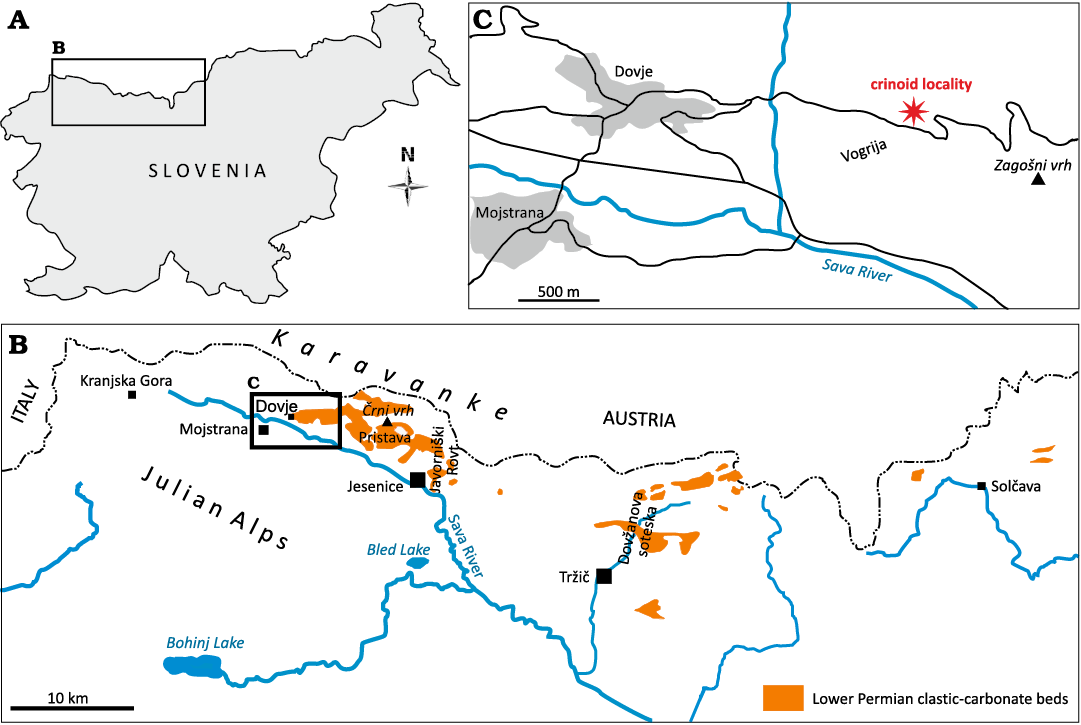
Fig. 1. Location of the crinoid locality within Slovenia (A, C). Index map the outcrops of lower Permian (Trogkofel Group) clastic-carbonate beds in Southern Karavanke Mountains (Slovenian part) study area indicated by the rectangle (B).
The lithostratigraphic subdivision of the lower Permian rocks in the Southern Karavanke Mts. is comprised of the Rattendorf Group and Trogkogel Group (base to top) (Novak and Skaberne 2009; Fig. 2). The Trogkofel Group, with a maximum of thickness 400 meters, consists of carbonate and clastic sediments. The carbonates include light-grey, dark-red, and rose-red reefal fossiliferous limestones and fore-reef limestone breccias in some parts. The Trogkofel Limestone is massive to thick-bedded and fossiliferous. The clastic sediments include the alternation of dark to light-grey shales, siltstones, and sandstones with rare conglomerate beds (Novak and Skaberne 2009). Within this clastic succession, layers or lenses of dark-grey to black biosparitic, biomicritic, and oolitic limestones and limestone breccias also occur with a biota that includes rock-forming algae, fusulinids, brachiopods, rare corals, and crinoids (Novak and Skaberne 2009). The age of the Trogkofel Group (with the Trogkofel Limestone) in Southern Karavanke is middle to late Artinskian (Novak and Skaberne 2009).
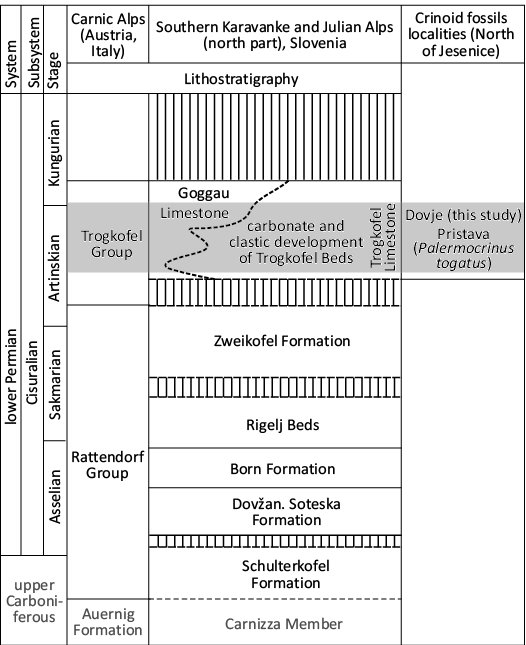
Fig. 2. Upper Permian lithostratigraphy of western part of Southern Karavanke Mountains, correlated with the subdivision and the standard zonation in the Carnic Alps (Austria) and with the stratigraphic position of studied crinoids in Trogkofel Group beds near Dovje (adopted after Novak and Skaberne 2009).
An outcrop exposed near Dovje (Vogrije in local dialect; Fig. 1), consisting of dark-grey limestones and alternating light-grey shales and siltstones, along the forest road between Dovje and Plavški Rovt, has yielded crinoids of Artinskian age (Novak and Skaberne 2009; Križnar 2022; Matijaž Novak, personal communication 2023). The exposed beds are very fossiliferous (fossiliferous cherts) containing disarticulated crinoid remains. The associated fauna includes productid and spiriferid brachiopods, bivalves, gastropods, larger foraminifera (fusulinids), algae (Shamovella–Archaeolithoporella), and bryozoans (fenestellid bryozoans). Rare vertebrate remains of chondrichthyan teeth bases of genus Petalodus was previously described by Križnar (2022) from these beds.
The succession of Trogkofel Group beds in the vicinity of Dovje indicates open marine sedimentary environments. Trogkofel Beds are interpreted as shelf margin reefs, with shallow platform deposits (fossiliferous bedded limestones) and the upper slope platform (with breccia sedimentation) environment (Novak and Skaberne 2009). No complete crinoids have been noted, suggesting that all samples contain material that was transported.
Material
The entire Slovenian collection from the Dovje area (see Fig. 1, Tables 1 and 2) consists of one partial calyx, 11 cups, two isolated basal or infrabasal circlets, two partly preserved arm portions, and several hundreds of columnals and pluricolumnals (only a few of the latter are illustrated herein) (Figs. 3–5).
Table 1. List of crinoidal remains stored in Upper Sava Museum Jesenice collected by Jože Bedič (Jesenice, Slovenia).
|
Specimen number |
Taxon |
Description |
Figures |
|
GMJ:PAL-0000011 |
Karavankecrinus bedici gen. et sp. nov. |
cup on slab |
Fig. 4E |
|
GMJ:PAL-0000002 |
Protencrinus baliensis |
cup on slab |
Fig. 5C |
|
GMJ:PAL-0000014a |
Moapacrinus dovjensis sp. nov. |
cup |
Fig. 4D |
|
GMJ:PAL-0000014b |
Eucladida indet. A |
cup |
Fig. 5A |
|
GMJ:PAL-0000014c |
Parastachyocrinus sloveniaensis sp. nov. |
cup |
Fig. 5D |
|
GMJ:PAL-0000014d |
Crinoidea indet. |
cup |
not illustrated herein |
|
GMJ:PAL-0000369 |
Parastachyocrinus sloveniaensis sp. nov. |
cup |
Fig. 4C |
|
GMJ:PAL-0000371 |
Parastachyocrinus sloveniaensis sp. nov. |
cup on slab |
Fig. 4F |
|
GMJ:PAL-0000373a |
Crinoidea indet. |
basal |
Fig. 5E |
|
GMJ:PAL-0000373b |
Crinoidea indet. |
basal |
Fig. 5F |
|
GMJ:PAL-0000372 |
Crinoidea indet. |
arm portion |
Fig. 4A |
|
GMJ:PAL-0000370 |
Crinoidea indet. |
arm portion |
Fig. 4B |
|
GMJ:PAL-0000368 |
Dieurycrinus? sp. |
calyx |
Fig. 5G |
Two cups and several hundreds of columnals and pluricolumnals are stored in Palaeontological collections of the Slovenian Museum of Natural History (Prirodoslovni muzej Slovenije), Ljubljana, and acronymed as PMSLJ Ba (Table 2). Several columnals and pluricolumnals have not yet been registered.
Table 2. List of crinoidal remains stored in Slovenian Museum of Natural History, Ljubljana, collected by Jožef Batič (Kranj, Slovenia) and unknown collectors.
|
Specimen number |
Taxon |
Description |
Figures |
|
PMSLJ Ba373 |
Sinocrinus websteri sp. nov. |
cup |
Fig. 5B |
|
PMSLJ Ba1744 |
Crinoidea indet. |
cup on slab |
not illustrated herein |
|
PMSLJ Ba370 |
Crinoidea indet. |
columnal |
Fig. 3A |
|
PMSLJ Ba369 |
Crinoidea indet. |
columnal |
Fig. 3B |
|
PMSLJ Ba374 |
Crinoidea indet. |
columnals+pluricolumnals |
Fig. 3D |
|
PMSLJ Ba381 |
Crinoidea indet. |
columnals+pluricolumnals |
Fig. 3F |
|
PMSLJ Ba381 |
Crinoidea indet. |
pluricolumnal |
Fig. 3E |
|
PMSLJ Ba381 |
Crinoidea indet. |
pluricolumnal |
Fig. 3C |
|
PMSLJ Ba381 |
Crinoidea indet. |
pluricolumnal |
Fig. 3G |
Classification and terminology.—Ordinal and superordinal classification of crinoids follows Cole (2017, 2018), Wright (2017a, b), and Wright et al. (2017). Family-level classification of crinoids follows Moore and Teichert (1978). Morphologic terminology for crinoids is from Webster (1974), Ubaghs (1978a), Kammer et al. (2013), Webster and Maples (2008), Ausich et al. (2020), and Ausich and Donovan (2023). A, B, C, D, and E represent echinoderm rays following the Carpenter Ray system (see Ubaghs 1978a: T63).
All measurements in mm; * after a measurement indicates the specimen is crushed or the feature is incomplete.
Systematic palaeontology
Class Crinoidea Miller, 1821
Infraclass Eucamerata Cole, 2017
Subclass Camerata Wachsmuth & Springer, 1885
Order Monobathrida Moore & Laudon, 1943
Suborder Glyptocrinina Ubaghs, 1978b
Superfamily Platycrinitoidea Austin & Austin, 1842
Family Platycrinitidae Austin & Austin, 1842
Platycrinitidae indet.
Fig. 3A.
Material.—PMSLJ Ba381; well-preserved columnal on a bedding surface; Trogkofel Group, Permian (middle to upper Artinskian), near Dovje, Slovenia.
Remark.—On a small limestone piece, crinoid columnals and pluricolumnals, brachiopods, and bryozoans are preserved (Fig. 3A). One columnal on this piece is elliptical and has the articular ridge oriented symmetrically along the widest portion of the columnal facet. This columnal has a maximum diameter of 12.6 mm and a minimum diameter of 7.4 mm, and it belongs to the Platycrinitidae. Such columnals were once regarded as diagnostic for the genus Platycrinites. However, now this basic columnal morphology is recognized as a family character for the Platycrinitidae (Ausich and Kammer 2009). During the Permian, this form of columnal could belong to Platycrinites Miller, 1821; Neoplatycrinus Wanner, 1916; or Pleurocrinus Austin and Austin, 1843.

Fig. 3. Crinoid columnals and pluricolumnals from Dovje, Slovenia, lower Permian, Cisuralian, Artinskian. A. Multiple columnals and one pluricolumnal; Platycrinitidae indet., PMSLJ Ba381. B. PMSLJ Ba381, crinoid pluricolumnal, articulation surface of one columnal (B1), pluricolumnal in lateral view (B2). C. PMSLJ Ba381, pluricolumnal with moldic preservation. D. PMSLJ Ba369, articular facet. E. PMSLJ Ba370, articular facet. F. PMSLJ Ba374, pluricolumnal with associated plates from radices. G. PMSLJ Ba381, crinoid pluricolumnal, articulation surface of one columnal (G1), pluricolumnal in lateral view (G2). Scale bars 10 mm.
Subclass Pentacrinoidea Jaekel, 1894
Infraclass Inadunata Wachsmuth & Springer, 1885
Parvclass Cladida Moore & Laudon, 1943
Magorder Eucladida Wright, 2017a
Superorder Flexibilia Zittel, 1895
Order Sagenocrinida Springer, 1913
Superfamily Sagenocrinoidea Roemer, 1854
Family Euryocrinidae Moore & Strimple, 1973
Genus Dieuryocrinus Wright, 1954
Type species: Euryocrinus duplex Wright, 1942, by original designation. Coplow Knoll, Clitheroe, UK; Mississippian (upper Tournaisian).
Stratigraphic and geographic range.—Viséan, Mississippian, Carboniferous, UK and questionably Artinskian, Cisuralian, Permian, Slovenia.
Dieuryocrinus? sp.
Fig. 4A.
Material.—GMJ:PAL-0000368, well-preserved partial calyx; Trogkofel Group, Permian (middle to upper Artinskian), near Dovje, Slovenia.
Description.—Calyx large in size; flat cone shape; plates flat to gently convex, smooth plate sculpturing. Infrabasal circlet plate and basal plate circlets not preserved and only distal-most portion of two radial plates visible. CD interray unknown. Regular interrays with one relatively small, pentagonal first interradial plate supporting two interradial plates above, regular interray plating 1-2-2-2-2-2-2-?; interray plates above the first interradial plate are hexagonal and diminish in size distally. Oral surface and anal sac unknown. Brachials fixed through at least the fifth secundibrachial. First secundibrachial hexagonal, wider than high; second primibrachial axillary, heptagonal, wider than high. Fixed secundibrachials wider than high. Fixed intrabrachials present between fixed secundibrachials, arranged 1-2-2-2-2-?. Free arms and column unknown.
Measurements.—CaW, 8.9*.

Fig. 4. Crinoids from Dovje, Slovenia, lower Permian, Cisuralian, Artinskian. A. Crinoidea indet. GMJ:PAL-0000373a, infrabasal circlet in basal view. B. Parastachyocrinus sloveniaensis sp. nov., GMJ:PAL-0000014c, aboral cup in basal view. C. Protencrinid Protencrinus baliensis Webster, 2012b, GMJ:PAL-0000002, aboral cup in basal view. D. Crinoidea indet. GMJ:PAL-0000373b, infrabasal circlet in basal view. E. Euryocrinid Dieuryocrinus? sp. GMJ:PAL-0000368, an incomplete calyx in basal view. F. Erisocrinid Sinocrinus websteri sp. nov., holotype, PMSLJ Ba373, aboral cup in basal (F1) and lateral (F2) views. G. Eucladid indet. A, GMJ:PAL-0000014b, aboral cup in basal (G1) and lateral (G2) views. Scale bars 10 mm.
Remarks.—Previously, Dieuryocrinus was only known from Viséan strata of the United Kingdom, so if this questioned generic assignment proves correct, this Slovenian occurrence greatly expands the temporal and geographic range of this genus. The generic assignment is questioned because only a partial crown of Dieuryocrinus? sp. is preserved. Taxonomically useful attributes of this specimen are a low to flat bowl-shaped calyx, fixed calyx plates through at least the proximal tertibrachials, two primibrachials, three or more secundibrachials, absence of patelloid processes on fixed brachial plates, first interradial plate followed by two columns of interradial plates, and fixed intrabrachial plates separating second secundibrachials and higher fixed brachials. Important morphological information missing on this specimen is the nature of the proximal column, plating in the posterior interray, extent of fixed brachials, and characters of the free brachials.
Most Permian flexible crinoids belong to specialized clades, such as the Calycocrinidae, Mespilocrinidae, Palaeoholopidae, and Prophyllocrinidae. This flexible crinoid from Slovenia is more typical of Permian Sagenocrinitoidea with numerous fixed brachials and intrabrachial plates. Previously four sagenocrinitoid genera have been described from Permian strata note, as listed in Webster and Webster (2014), Forbesiocrinus siculus Gregario, 1930, does not belong to the Forbesiocrinus).
The Slovenian flexible crinoid is most similar to two genera in the Euryocrinidae: Caldenocrinus Wright, 1946, and Dieuryocrinus Wright, 1954 (both Viséan, Scotland) that lack patelloid processes and have both interradial and intrabrachial plates. With a double row of interradial plates and several intrabrachial plates, this flexible crinoid is most similar to Dieuryocrinus and is assigned to Dieuryocrinus? sp., which may extend the range of this genus into the lower Permian. The generic assignment is questioned and a species is not named because key morphological characters of this taxon remain unknown.
Superorder Cyathoformes Wright et al., 2017
Cyathoformes incertae sedis “Poteriocrinida” Jaekel, 1918
Superfamily Erisocrinoidea Wachsmuth & Springer, 1886
Family Stachyocrinidae Moore & Strimple, 1973
Remarks.—Moore and Strimple (1973: 22) erected the Stachyocrinidae to include erisocrinoids with a cylindrical to ovoid crown, low bowl to saucer-shaped aboral cups with a shallow basal concavity, proximal tips of the radial plates reaching the basal plate or well above it, primanal not visible from the exterior of the cup in lateral view, ten arms with rectilinear uniserial brachials, and a circular proximal column. They included Stachyocrinus Wanner, 1916, and Parastachyocrinus Wanner, 1937, with Stachyocrinus having aboral cups with the basal plates not visible in lateral view, whereas the basal plates are clearly visible in Parastachyocrinus.
Two specimens (Fig. 5C and F) from the Trogkofel Group are placed in the Stachyocrinidae because they have a very low bowl-shaped aboral cup with infrabasals partly in a shallow basal concavity and not visible in lateral view, modestly convex basal plates, plenary radial facets, and no posterior plates visible in lateral view of the aboral cup. This new material is placed within Parastachyocrinus because the basal plates are not confined to the base of the aboral cup and are visible in lateral view.

Fig. 5. Crinoids from Dovje, Slovenia, lower Permian, Cisuralian, Artinskian. A. GMJ:PAL-0000372, incomplete arms of a eucladid crinoid in lateral view (A1, A2). B. GMJ:PAL-0000370, incomplete arms of a eucladid crinoid in lateral view (B1, B2). C. Stachyocrinid Parastachyocrinus sloveniaensis sp. nov., holotype, GMJ:PAL-0000369, aboral cup in oral (C1), lateral (C2), and basal (C3) views. D. Cromyocrinid Moapacrinus dovjensis sp. nov., GMJ:PAL-0000014a, aboral cup in basal (D1) and lateral (D2) views. E. Stachyocrinid Karavankecrinus bedici gen. et sp. nov., GMJ:PAL-0000011, cup in basal view. F. Stachyocrinid Parastachyocrinus sloveniaensis sp. nov., paratype, GMJ:PAL-0000371, aboral cup in lateral (F1) and basal (F2) view. Scale bars 10 mm.
As presently understood, species-level systematics within Parastachyocrinus is very poorly defined and is revised herein. First, species currently assigned to Parastachyocrinus have a wide range of morphologies that would more typically be indicative of more than one genus. Second, the morphology of the type species requires emphasis.
In the initial description of Erisocrinus malaianus Wanner, 1924, two morphotypes were illustrated (“Exemplar a” and “Exemplar b”). “Exemplar a” has very convex and/or spinose basal and radial plates (Wanner 1924: pl. 18: 25–27), and he referred to this specimen as the holotype (herein “Exemplar a” of Erisocrinus malaianus Wanner, 1924, is regarded as the holotype of the type species of Parastachyocrinus). Wanner (1924: pl. 18: 22–24) illustrated a second morphology as “Exemplar b”. This specimen has an aboral cup of similar shape to “Exemplar a”, but the aboral cup plates are only moderately convex and nodes and spines are absent. Wanner (1924) also described Erisocrinus obliquus and illustrated five specimens (“Exemplar a” to “Exemplar d”). As noted below, Erisocrinus obliquus was subsequently reassigned to Parastachyocrinus.
In 1949, Wanner erected Parastachyocrinus, designated Erisocrinus malaianus Wanner, 1924, as its type species, and listed Wanner (1924: pl. 18: 22–27), both “Exemplar a” and “Exemplar b” to refer to the morphology of Parastachyocrinus malaianus. However, Wanner (1949) did not re-illustrate “Examplar a”. Instead, he illustrated five additional specimens of P. malaianus, “Exemplar c” to “Exemplar g” (Wanner 1949: pl. 3: 10–17). He also introduced P. malaianus var. ornata (Wanner 1949: pl. 3: 18–19), which was subsequently corrected to P. malaianus ornatus by Webster and Webster (2014).
Also in 1949, Wanner reassigned Erisocrinus granulatus Wanner, 1924, and Erisocrinus obliquus Wanner, 1916, to Parastachyocrinus and named the new species P. inflatus Wanner, 1949. Finally, Moore et al. (1978: fig. 475.1) illustrated two specimens of P. malaianus, “Exemplar a” and “Exemplar c”, which are very dissimilar in morphology, but designated “Exemplar a” as the “holotype” in the figure explanation.
In summary, Webster and Webster (2014) recognized five species-level taxa in Parastachyocrinus, and two new species are recognized herein.
Genus Parastachyocrinus Wanner, 1949
Type species: Erisocrinus malaianus Wanner, 1924. Unknown series of Permian, Timor.
Included species: Parastachyocrinus granulatus (Wanner, 1924), Parastachyocrinus inflatus Wanner, 1949; Parastachyocrinus malaianus (Wanner, 1924), Parastachyo crinusobliquus (Wanner, 1916), Parastachyocrinus ornatus Wanner, 1949; Parastachyocrinus wanneri sp. nov.; Parastachyocrinus sloveniaensis sp. nov.
Remarks.—Even with the revisions proposed herein, some morphological variability exists within species of Parastachyocrinus. Species diagnostic characters, as listed below for each species, are aboral cup symmetry, aboral cup shape, aboral cup outline in basal view, aboral cup plate sculpturing, degree of depression along the aboral cup sutures, presence or absence of a basal concavity, whether infrabasal plates are confined to the basal concavity, whether infrabasal plates are visible in lateral view, relative convexity of basal plates, presence or absence of spines or nodes on basal plates, radial plate convexity, and presence or absence of spines or nodes on radial plates.
Stratigraphic and geographic range.—Unknown series of Permian, Timor; Artinskian, Permian, Slovenia.
Parastachyocrinus malaianus (Wanner, 1924)
1924 Erisocrinus malaianus sp. nov. “Exemplar a”; Wanner, 1924: 291, pl. 18, figs. 25–27.
1943 Erisocrinus malaianus Wanner, 1924, in part; Bassler and Moodey 1943: 459.
1948 Erisocrinus malaianus, Wanner, 1924, in part; Branson 1948: 192.
1949 Parastachyocrinus malaianus, (Wanner, 1924), in part; Wanner 1949: 44.
1973 Parastachyocrinus malaianus, (Wanner, 1924), in part; Webster 1973: 195.
1978 Parastachyocrinus malaianus, (Wanner, 1924), in part; Moore et al. 1978: T720, fig. 475.1.
2014 Parastachyocrinus malaianus (Wanner, 1924), in part; Webster and Webster 2014: 2014.
Holotype: RGM.873646, illustrated by Wanner (1924: pl. 18: 25–27.
Type locality: Basleo, West Timor.
Type horizon: Permian (uncertain series).
Emended diagnosis.—Aboral cup symmetrical, aboral cup shape very low bowl, subcircular aboral cup shape in basal view, smooth aboral cup plate sculpturing, aboral cup plate sutures strongly impressed, basal concavity present, infrabasal plates not confined to basal concavity, infrabasal plates not visible in lateral view, basal plates strongly convex, spines or nodes present or absent on basal plates, radial plates strongly convex, spines or nodes present or absent on basal plates.
Stratigraphic and geographic range.—Unknown series of Permian, Timor.
Parastachyocrinus granulatus (Wanner, 1924)
1924 Erisocrinus granulatus sp. nov. “Exemplar a”, Wanner 1924: 290, pl. 18: 28–30; pl. 19: 5.
1943 Erisocrinus granulatus Wanner, 1924; Bassler and Moodey 1943: 459.
1948 Erisocrinus granulatus Wanner, 1924; Branson 1948: 192.
1949 Parastachyocrinus granulatus (Wanner, 1924); Wanner 1949: 42.
1973 Parastachyocrinus granulatus (Wanner, 1924); Webster 1973: 195.
2002 Erisocrinus granulatus Wanner, 1924; Pabian and Rushlau 2002: 23.
2014 Parastachyocrinus granulatus (Wanner, 1924); Webster and Webster 2014: 2014.
Holotype: RGM.873914, illustrated by Wanner (1924: pl. 18: 28–30).
Type locality: Basleo, West Timor.
Type horizon: Permian (uncertain series).
Emended diagnosis.—Aboral cup asymmetrical, aboral cup shape medium bowl, subcircular aboral cup shape in basal view, granulose aboral cup plate sculpturing, aboral cup plate sutures slightly impressed, basal concavity present, infrabasal plates not confined to basal concavity, infrabasal plates not visible in lateral view, basal plates moderately convex, spines or nodes not on basal plates, radial plates moderately convex, spines or nodes not on basal plates.
Stratigraphic and geographic range.—Unknown series of Permian, Timor.
Parastachyocrinus inflatus Wanner, 1949
1949 Parastachyocrinus inflatus sp. nov.; Wanner 1949: 42, pl. 3: 20–21.
1973 Parastachyocrinus inflatus Wanner, 1949; Webster 1973: 195.
2014 Parastachyocrinus inflatus Wanner, 1949; Webster and Webster 2014: 2014.
Holotype: RGM.892856, illustrated by Wanner (1949: pl. 3: 20–21).
Type locality: Basleo, West Timor.
Type horizon: Permian (stage uncertain).
Emended diagnosis.—Aboral cup symmetrical, aboral cup shape very low bowl, pentalobate aboral cup shape in basal view, smooth aboral cup plate sculpturing, aboral cup plate sutures very strongly impressed, basal concavity present, infrabasal plates confined to basal concavity, infrabasal plates not visible in lateral view, basal plates very strongly convex, spines or nodes not on basal plates, radial plates very strongly convex, spines or nodes not on basal plates.
Stratigraphic and geographic range.—Unknown series of Permian, Timor.
Parastachyocrinus obliquus (Wanner, 1916)
1916 Erisocrinus obliquus sp. nov. “Exemplar a”; Wanner 1916: 225, pl. 103(8): 10a–c.
1924 Erisocrinus obliquus Wanner, 1916 “Exemplar b”; Wanner 1924: 288, pl. 18: 20; text-fig. 56.
1924 Erisocrinus obliquus Wanner, 1916 “Exemplar c”; Wanner 1924: 288, pl. 18: 18–19.
1924 Erisocrinus obliquus Wanner, 1916 “Exemplar d”; Wanner 1924: 288, pl. 18: 17.
1924 Erisocrinus obliquus Wanner, 1916 “Exemplar e”; Wanner 1924: 288, pl. 18: 20, 21.
1943 Erisocrinus obliquus Wanner, 1916; Bassler and Moodey 1943: 459.
1948 Erisocrinus obliquus Wanner, 1916; Branson 1948: 192.
1949 Parastachyocrinus obliquus (Wanner, 1916); Wanner 1949: 42.
1964 Erisocrinus obliquus Wanner, 1916; Sant in Sant and Lane: 1964: 65, fig. 23.1
1973 Parastachyocrinus obliquus (Wanner, 1916); Webster 1973: 195.
2014 Parastachyocrinus obliquus (Wanner, 1916); Webster and Webster 2014: 2014–2015.
Holotype: RGM.873661, illustrated by Wanner (1916: pl. 103(8): 10a–c).
Type locality: Basleo, West Timor.
Type horizon: Permian (uncertain series).
Emended diagnosis.—Aboral cup asymmetrical; aboral cup shape low cone, very low bowl, low bowl, or low urn; subpentagonal aboral cup shape in basal view (as known); smooth aboral cup plate sculpturing; aboral cup plate sutures slightly or moderately impressed or not impressed; basal concavity present or absent(?); infrabasal plates not confined to basal concavity; infrabasal plates visible in lateral view; basal plates slightly to moderately convex; spines or nodes not on basal plates; radial plates slightly to moderately convex; spines or nodes not on basal plates.
Stratigraphic and geographic range.—Unknown series of Permian, Timor.
Parastachyocrinus ornatus Wanner, 1949
1949 Parastachyocrinus malaianus var. ornata var. nov.; Wanner 1949: 45, pl. 3: 18–19.
1973 Parastachyocrinus malaianus var. ornata Wanner, 1949; Webster 1973: 195.
2014 Parastachyocrinus malaianus ornatus (nomen correctum) (Wanner, 1949); Webster and Webster 2014: 2014.
Holotype: Holotype not located, well preserved aboral cup illustrated by Wanner (1949: pl. 3: 18–19).
Type locality: Timor.
Type horizon: unknown series of Permian.
Emended diagnosis.—Aboral cup symmetrical, aboral cup shape low bowl, subcircular aboral cup shape in basal view, granulose? aboral cup plate sculpturing, aboral cup plate sutures slightly impressed, basal concavity present, infrabasal plates not confined to basal concavity, infrabasal plates not visible in lateral view, basal plates moderately convex, spines or nodes not on basal plates, radial plates moderately convex, spines or nodes not on basal plates.
Remarks.—The subspecies Parastachyocrinus malaianus ornatus (Wanner, 1949) is elevated herein to species rank Parastachyocrinus ornatus.
Stratigraphic and geographic range.—Unknown series of Permian, Timor.
Parastachyocrinus wanneri sp. nov.
1949 Parastachyocrinus malaianus Wanner, 1924, “Exemplar c”; Wanner 1949: pl. 3: 10–11.
1949 Parastachyocrinus malaianus, “Exemplar d”; Wanner 1949: pl. 3: 12.
1949 Parastachyocrinus malaianus, “Exemplar e”; Wanner 1949: pl. 3: 13.
ZooBank LSID: urn:lsid:zoobank.org:act:95B7DCDE-897F-427E-AE 7A-09C679348E62.
Etymology: In recognition of Johannes Wanner (1878–1959), whose extensive work on Permian crinoids of Timor has been instrumental in our understanding of crinoids from the late Paleozoic.
Holotype: Holotype not located, “Exemplar c” illustrated by Wanner (1949: pl. 3: 10–11).
Type locality: Timor.
Type horizon: Unknown series of Permian.
Diagnosis.—Aboral cup symmetrical, aboral cup shape low to medium bowl, circular to subcircular aboral cup shape in basal view, smooth aboral cup plate sculpturing, aboral cup plate sutures not impressed, basal concavity absent (as known), infrabasal plates confined(?) or not confined to basal concavity, infrabasal plates visible or not visible in lateral view, basal plates slightly to strongly convex, spines or nodes not on basal plates, radial plates slightly to strongly convex, spines or nodes not on basal plates.
Stratigraphic and geographic range.—Unknown series of Permian, Timor.
Parastachyocrinus sloveniaensis sp. nov.
Figs. 4B, 5C, F.
1924 Erisocrinus malaianus, “Exemplar b”; Wanner 1924: 290, pl. 18: 22–24.
1943 Erisocrinus granulatus Wanner, 1924, in part; Bassler and Moodey 1943: 459.
1948 Erisocrinus granulatus Wanner, 1924, in part; Branson 1943: 192.
1949 Erisocrinus malaianus Wanner, 1924 “Exemplar f”; Wanner 1924: 290, pl. 3: 14.
1949 Erisocrinus malaianus Wanner, 1924 “Exemplar g”; Wanner 1924: 290, pl. 3: 15–17.
1973 Parastachyocrinus malaianus Wanner, 1924, in part; Webster 1973: 195.
2014 Parastachyocrinus malaianus Wanner, 1924, in part; Webster and Webster 1973: 2014.
ZooBank LSID: urn:lsid:zoobank.org:act:105458F9-A6EA-4AAB-BB C8-00640946DDD2.
Etymology: After Slovenia, from where the species is recognized.
Type material: Holotype: GMJ:PAL-0000369, moderately preserved aboral cup. Paratype: GMJ:PAL-0000371, moderately preserved aboral cup.
Type locality: Near Dovje, Slovenia.
Type horizon: Trogkofel Group, Permian, Cisuralian, middle to upper Artinskian.
Material.—Type material and PMSLJ Ba373, moderately preserved aboral cup from the type locality and horizon.
Diagnosis.—Aboral cup symmetrical, aboral cup shape very low bowl, circular to subcircular to subpentalobate aboral cup shape in basal view, smooth aboral cup plate sculpturing, aboral cup plate sutures slightly to moderately impressed, basal concavity present (as known), infrabasal plates not confined to basal concavity (as known), infrabasal plates not visible in lateral view, basal plates slightly to moderately convex, spines or nodes not on basal plates, radial plates slightly to moderately convex, spines or nodes not on radial plates.
Measurements.—GMJ:PAL-0000369: ACH, 19.0; ACW, ~40.0; GMJ:PAL-0000371: ACW, 40.0*.
Description.—Aboral cup symmetrical very low bowl shape, circular to subcircular aboral cup shape in basal view, height width ratio ~0.42; plates broadly convex, smooth sculpturing, plate sutures slightly to moderately impressed. Infrabasal circlet confined to a shallow basal concavity, proximal column covers ~50% of infrabasal circlet, not visible in lateral view; infrabasal plates five, equal in size as known not confined to basal concavity (as known). Basal circlet visible in lateral view; basal plates five, pentagonal, smaller than radial plates, wider than high, slightly to moderately convex, spines or nodes not on basal plates. Radial plates presumably five, pentagonal, ~2.0 times wider than high. Radial facets plenary, horizontally oriented as known, articular ridge probably across full width of facet, but other details of radial facet unknown. No posterior interray plates in external wall of aboral cup. Oral surface, anal sac, and arms unknown. Proximal column circular, lumen circular, other details unknown.
Remarks.—Two specimens are definitely assigned to P. sloveniaensis sp. nov. Both are aboral cups, and most aboral cup characters are known. Parastachyocrinus sloveniaensis sp. nov. is a species of Parastachyocrinus with a symmetrical aboral cup and smooth aboral plate sculpturing. It is differentitated from other Parastachyocrinus species with these two attributes because P. malaianus has an aboral cup shape very low bowl, subcircular aboral cup shape in basal view, aboral cup plate sutures strongly impressed, basal concavity present, infrabasal plates not confined to basal concavity, infrabasal plates not visible in lateral view, basal plates strongly convex, spines or nodes present or absent on basal plates, radial plates strongly convex, spines or nodes present or absent on basal plates; P. inflatus has an aboral cup shape very low bowl, pentalobate aboral cup shape in basal view, aboral cup plate sutures very strongly impressed, basal concavity present, infrabasal plates confined to basal concavity, infrabasal plates not visible in lateral view, basal plates very strongly convex, spines or nodes not on basal plates, radial plates very strongly convex, spines or nodes not on basal plates; and P. wanneri has an aboral cup shape low to medium bowl, circular to subcircular aboral cup shape in basal view, aboral cup plate sutures not impressed, basal concavity absent (as known), infrabasal plates confined(?) or not confined to basal concavity, infrabasal plates visible or not visible in lateral view, basal plates slightly to strongly convex, spines or nodes not on basal plates, radial plates slightly to strongly convex, spines or nodes not on basal plates. In contrast, P. sloveniaensis sp. nov. has an aboral cup shape very low bowl, circular to subcircular to subpentalobate aboral cup shape in basal view, aboral cup plate sutures slightly to moderately impressed, basal concavity present (as known), infrabasal plates not confined to basal concavity (as known), infrabasal plates not visible in lateral view, basal plates slightly to moderately convex, spines or nodes not on basal plates, radial plates slightly to moderately convex, spines or nodes not on radial plates.
The plate structure of the aboral cup in Fig. 4B is uncertain because of the lack of a defined infrabasal circlet with defined infrabasal plates, which was definitely present. It is not clear if individual infrabasal plates existed or if infrabasal plates are fused. A more probable explanation is that an isolated plate from another crinoid specimen is lodged in the basal concavity and obscures the infrabasal plates. Because of the similarity of the basal and radial plates of Fig. 4B, this specimen is questionably assigned to P. sloveniaensis sp. nov.
All three specimens attributed to Parastachyocrinus sloveniaensis sp. nov. appear to have been variously weathered, so the plate sculpturing may be a question. However, plate sculpturing is considered smooth until better specimens can be collected.
Stratigraphic and geographic range.—Trogkofel Group, Permian, Cisuralian, middle to upper Artinskian, near Dovje, Slovenia.
Genus Karavankecrinus nov.
ZooBank LSID: urn:lsid:zoobank.org:act:FC74C6BA-D4BA-42C6-809E-73AAC49BFC92
Type species: Karavankecrinus bedici gen. et sp. nov.; see below.
Etymology: In reference to the Karavanke Mountains, the part of the Alps from which this new crinoid was collected.
Diagnosis.—As for the type species.
Remarks.— Karavankecrinus bedici gen. et sp. nov. is a very distinctive eucladid crinoid because in at least two interrays a basal plate and an interradial plate are in sutural contact, thus separating adjacent radial plates. This occurs in many crinoids in the posterior interray only. However, this condition is only common in Ordovician–Mississippian Rhodocrinitoidea diplobathrid camerates. It is extremely rare in other clades. An exception is the Permian flexible crinoid Trampidocrinus Lane & Webster, 1966 from the Permian of Nevada, USA (see Lane and Webster 1966: fig. 6, pl. 2; Moore 1978: fig. 353.3). A flexible crinoid identification cannot be confirmed because of the relatively few preserved characters, and none are diagnostic for the Flexibilia. Almost uniformly, flexible crinoids have three infrabasal plates (one small and two large). In Karavankecrinus gen. nov., the number and position of infrabasal plates is unclear. Radiating “lines” that appear to be sutures are present, but more than three (and not five) potential sutures exist; and unlike the expectation these “lines” are not oriented in the typical fashion in which they bisect the superjacent basal plate. Thus, it is questioned whether these “lines” represent sutures. Also, Trampidocrinus and many other flexible crinoids have well-developed patelloid processes between the radial plate and first primibrachial plate, as well as between higher brachial plates. A patelloid process is absent on the two, well-preserved radial plates on K. bedici gen. et sp. nov. We are unaware of any eucladid crinoid that has the radial plates separated in any interray except in the posterior interray. Although the specimen is incompletely known, this unique character justifies designation of a new genus and species: K. bedici.
Stratigraphic and geographic range.—Permian, Cisuralian, Artinskian, Slovenia.
Karavankecrinus bedici sp. nov.
Figs. 5E, 6B.
2013 Codiacrinacea Bather, 1890; Lach et al. 2013: 32, fig. 1.
ZooBank LSID: urn:lsid:zoobank.org:act:0BD46C5C-FCEC-4115-B43 B-93E5BFA30881.
Etymology: In honour of Jože Bedič (1923–2002) (Jesenice, Slovenia), who collected the holotype of this new species as well as some other specimens considered in this study.
Holotype: GMJ:PAL-0000011; moderately preserved aboral cup.
Type locality: Near Dovje, Slovenia.
Type horizon: Trogkofel Group, Permian, Cisuralian, middle to upper Artinskian.
Diagnosis.—Aboral cup very low to flat cone, infrabasal circlet almost entirely covered by proximal columnal, in at least two interrays adjacent radial plates are separated by a basal plate and first interradial plate in sutural contact.
Measurements.—ACW, 40.1.
Description.—Aboral cup very low to flat cone shape; plates gently convex, smooth sculpturing. Infrabasal circlet almost completely covered by proximal columnal, in shallow basal concavity, not visible in lateral view. Individual infrabasal plates very poorly defined. Basal circlet slightly visible in lateral view; basal plates five, hexagonal or heptagonal CD basal plate, smaller than radial plates, ~1.2 times wider than high; each radial plate in sutural contact with an interradial plate above (Fig. 6B). Radial plates five, pentagonal, ~2.0 times wider than high, separated in all interrays. Radial facets plenary, declivate, details of the radial facets unknown. CD interray plating presumably with only the radianal plate in the aboral cup. Radianal plate is directly above the CD basal plate along a wide suture and separates the C and D radial plates. In addition to the CD interray, plates in the radial circlet interrupted in other interrays with the basal plate in sutural contact with an interradial plate. Oral surface, anal sac, and arms unknown. Proximalmost columnal circular, lumen pentalobate, other details of column unknown.
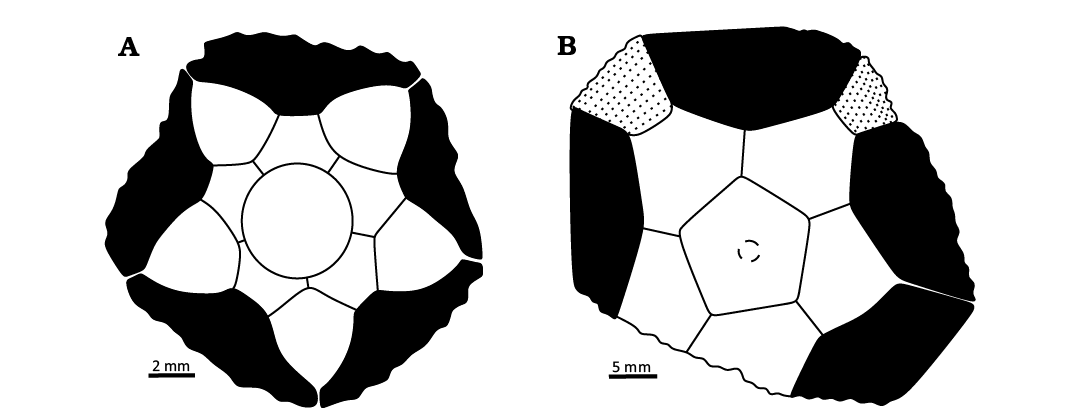
Fig. 6. Plate drawings of the basal view of aboral cups. A. Protencrinid Protencrinus baliensis, GMJ:PAL-0000002, note the suture connection between the infrabasal and radial plates that separate all adjacent basal plates. B. Stachyocrinid Karavankecrinus bedici gen. et sp. nov., PMSLJ Ba381, in two interrays, not{ e?} the suture connection between the basal plate and the first interradial plate that separate all adjacent radial plates.
Remarks.—The plating of this crinoid is highly unusual with the radial circlet interrupted in at least two interrays. One is presumed to be the CD interray, and if that is correct, the other definite example is in the BC interray (Figs. 5E, 6B).
Stratigraphic and geographic range.—Trogkofel Group, Permian, Cisuralian, middle to upper Artinskian, near Dovje, Slovenia.
Family Cromyocrinidae Bather, 1890
Genus Moapacrinus Lane & Webster, 1966
Type species: Moapacrinus rotundatus Lane & Webster, 1966. Nevada, USA; Permian, Sakmarian?.
Included species: Moapacrinus cuneatus Webster & Jell, 1999; M. dovjensis sp. nov.; M. inornatus Webster & Lane, 1967; M. rotundatus Lane & Webster, 1966.
Remarks.—Moapacrinus was originally erected to include, among other things, eucladids with a very low bowl-shaped aboral cup, plenary radial facets, and one anal plate (radianal) partially in the aboral cup. Moapacrinus elexensis Pabian & Strimple, 1993, has two posterior plates in the aboral cup and is not considered, herein, a species Moapacrinus. Also, Webster et al. (2009b) described Moapacrinus sp. nov. 1 and Moapacrinus? sp. nov. 2 based on uncollectable specimens from British Columbia, Canada. Neither specimen is sufficiently preserved to confidently place them in Moapacrinus; and, further, Moapacrinus? sp. nov. 2 appears to have two posterior plates in the aboral cup. Because of morphological differences and poor preservation, M. elexensis Pabian & Strimple, 1993; Moapacrinus sp. nov. 1; and Moapacrinus? sp. nov. are not considered in the discussion of M. dovjensis sp. nov. below.
Stratigraphic and geographic range.—Carboniferous, Pennsylvanian, Kasimovian and Permian, USA; Permian, uppermost Artinskian or lowermost Roadian, Australia; Permian, Cisuralian, middle Sakmarian?, Oman; Permian, Cisurialian, Asselian to Sakmarian, Slovenia.
Moapacrinus dovjensis sp. nov.
Fig. 5D.
1978 cup of sea lily; Ramovš 1978: 74, fig. 37.1.
ZooBank LSID: urn:lsid:zoobank.org:act:6FBF99D0-50F9-4A83-9D DF-14BF50D84A5F.
Etymology: In reference to the village of Dovje, Southern Karavanke Mountains, Slovenia, which is close to the locality where this specimen was collected.
Holotype: GMJ:PAL-0000014a; moderately preserved aboral cup.
Type locality: Near Dovje, Slovenia.
Type horizon: Trogkofel Group, Permian, Cisuralian, middle to upper Artinskian.
Diagnosis.—Very low bowl-shaped aboral cup shape, outline of aboral cup in basal view slightly elliptical; smooth? aboral cup plate sculpturing; aboral cup plate sutures slightly depressed; presence or absence of basal concavity unknown; infrabasal circlet pentagonal; infrabasal plates not visible in lateral view; basal plates slightly higher than wide, orientation of distal basal plates angled upward, position of the maximum width of the basal plates at base of aboral cup; radial plate >2.5 times wider than high; radianal plate large, pentagonal, ~50% below top of radial plates; CD basal plate-radianal plate suture at an angle, arms unknown.
Measurements.—ACH. 19.7; ACW, 35.8.
Description.—Aboral cup very low bowl-shaped, height width ratio ~2.9, very slightly elliptical in outline from a basal view; plates broadly convex, smooth sculpturing, aboral cup plate sutures slightly impressed; presence or absence of basal concavity unknown. Infrabasals circlet pentagonal, not visible in lateral view, presumably five infrabasal plates. Basal plates five, hexagonal, smaller than radial plates, slightly higher than wide, orientation of distal basal plates angled upward, position of maximum width of basal plates at the base of the aboral cup. Radial plates presumably five, pentagonal, ~2.5 times wider than high. Radial facets not known. CD interray plating with only radianal in aboral cup; radianal plate large, pentagonal, ~50% below top of radial plates, angled suture between radianal and CD interray plates. Oral surface, anal sac, arms, and column unknown.
Remarks.—Moapacrinus dovjensis sp. nov. is known from a single aboral cup with the infrabasal circlet missing. However, the aboral cup shape and a single large posterior interray plate align this specimen with Moapacrinus. This new species is distinguished from species recognized herein as members of Moapacrinus because M. cuneatus has an outline of the aboral cup in basal view unknown; coarsely nodose aboral cup plate sculpturing; aboral cup plate sutures moderately to deeply impressed; basal concavity shallow; infrabasal circlet shape unknown; infrabasal plates not visible in lateral view; basal plates ~1.3 times wider than high, orientation of distal basal plates angled upward, position of the maximum width of the basal plates near base of aboral cup; radial plate ~1.8 times wider than high; radianal plate large, hexagonal, ~50% below top of radial plates; CD basal plate-radianal plate suture at an angle; distal primibrachial and first secundibrachial do not form a projection; smooth brachial sculpturing; brachials cuneate uniserial; M. inornatus has an outline of the aboral cup in basal view slightly elliptical; smooth aboral cup plate sculpturing; aboral cup plate sutures very slightly impressed; basal concavity narrow, deep; infrabasal circlet pentalobate; distal tips of infrabasal plates visible in lateral view; basal plates slightly wider than high or equidimensional, orientation of distal basal plates nearly vertical, position of maximum width of the basal plates well above base of aboral cup; radial plate >2.0 times wider than high; radianal plate large, hexagonal, ~67% below top of radial plates; CD basal plate-radianal plate suture horizontal; distal primibrachial and first secundibrachial form a projection; smooth brachial sculpturing; and brachials rectilinear uniserial; and M. rotundatus has an outline of the aboral cup in basal view circular; aboral cup plate sculpturing variable, nearly smooth, coarse nodes, or irregular wavy ridges; aboral cup plate sutures moderately impressed; basal concavity narrow and deep; infrabasal circlet pentagonal; infrabasal plates not visible in lateral view; basal plates wider than high, nearly vertical orientation of distal basal plates, position of the maximum width of the basal plates above the base of aboral cup; radial plate >2.0 times wider than high; radianal plate large, pentagonal, ~50% below top of radial plates; CD basal plate-radianal plate suture horizontal; distal primibrachial and first secundibrachial do not form a projection; transverse sculpturing on brachial; and brachials rectilinear uniserial. In contrast, M. dovjensis sp. nov. has an outline of the aboral cup in basal view slightly elliptical; smooth? aboral cup plate sculpturing; aboral cup plate sutures slightly impressed; presence or absence of basal concavity unknown; infrabasal circlet pentagonal; infrabasal plates not visible in lateral view; basal plates slightly higher than wide, orientation of distal basal plates angled upward, position of the maximum width of the basal plates at base of aboral cup; radial plate ~2.5 times wider than high; radianal plate large, pentagonal, ~50% below top of radial plates; CD basal plate-radianal plate suture at an angle; arms unknown.
Stratigraphic and geographic range.—Trogkofel Group, Permian, Cisuralian, middle to upper Artinskian, near Dovje, Slovenia.
Superfamily Erisocrinoidea Wachsmuth & Springer, 1886
Family Erisocrinidae Wachsmuth & Springer, 1886
Included genera: Aikyshicrinus Hashimoto, 1995; Eperisocrinus Burke, 1977; Erisocrinus Meek and Worthen, 1865; Exaetocrinus Strimple and Watkins, 1969; and Sinocrinus Tien, 1926.
Genus Sinocrinus Tien, 1926
Type species: Sinocrinus microgranulatus Tien, 1926 = Sinocrinus granulatus (Wanner, 1924); see discussion in Sheffield (2015); Sakmarian, Permian, China.
Included species: Sinocrinus asymmetricus Strimple, 1976; S. websteri sp. nov.; Sinocrinus cernuus (Trautschold, 1867); Sinocrinus granulatus (Wanner, 1924); Sinocrinus houkouensis Tien, 1924; Sinocrinus lichengensis Tien, 1924; Sinocrinus nodosus Tien, 1924; Sinocrinus obliquus (Wanner, 1916); Sinocrinus sheareri Strimple & Watkins, 1969; Sinocrinus websteri sp. nov.
Diagnosis.—Aboral cup shape variable (low bowl, very low bowl, low cone), granulose plate sculpturing, circular to subpentalobate basal view outline of aboral cup, basal concavity present, infrabasal plates confined or not confined to basal concavity, infrabasal plates visible or not visible in lateral view, large basal plates, planate to slightly declinate radial facets, no posterior interray plates in aboral cup, distal spinose projection from axillary first primibrachial present or absent, second primibrachial and above biserial, circular proximal columnal.
Stratigraphic and geographic range.—Pennsylvanian, Moscovian, Permian, Cisuralian, Artinskian; China, Russia, Slovenia, Timor, USA.
Sinocrinus websteri sp. nov.
Fig. 4F.
ZooBank LSID: urn:lsid:zoobank.org:act:44E00651-5794-41FF-9452-899180B7D977.
Etymology: In honour of Gary D. Webster (1934–2021), whose work greatly advanced our understanding of Permian crinoids around the world.
Holotype: PMSLJ Ba 373; moderately preserved aboral cup.
Type locality: Near Dovje, Slovenia.
Type horizon: Trogkofel Group, Permian, Cisuralian, middle to upper Artinskian.
Diagnosis.—Low bowl-shaped aboral cup, aboral cup plate sculpturing unknown, aboral cup circular in basal view, basal concavity present, proximal column cicatrix >60% of infrabasal circlet diameter, infrabasal circlet not confined to basal concavity, height of all infrabasal plates equal, basal plates gently convex, basal plates wider than high, arms unknown, moderately developed crenulae in the proximal column cicatrix, proximal columnal circular.
Measurements.—ACH, 19.2; ACW~34.0.
Description.—Aboral cup low bowl shape, height width ratio ~0.5, circular in basal view, basal concavity present; plates convex, sculpturing not certain due to weathering, impressed sutures. Infrabasal circlet not visible in lateral view, not confined to basal concavity column cicatrix ~77% of the width of the infrabasal circlet; infrabasal plates presumably partially fused into one plate with very faint sutures, height of all infrabasal plates equal. Basal plates five, pentagonal, gently convex, smaller than radial plates, ~1.7 times wider than high. Radial plates presumably five, pentagonal, ~1.6 times wider than high. Radial facets plenary, horizontally oriented, articular ridge probably across full width of facet, but other details of radial facet not known. No posterior interray plates in CD interray. Oral surface, anal sac, and arms unknown. Moderately developed crenulae in proximal columnal cicatrix, proximal column circular, other aspects of column unknown.
Remarks.—Other Permian species of Sinocrinus include S. granulatus, S. lichengensis, S. obliquus, and S. sheareri (see Sheffield 2015; Mao et al. 2018). These species are differentiated on the shape of the aboral cup, aboral cup plate sculpturing, aboral cup outline in basal view, width of the basal concavity on the infrabasal circlet, relative sizes of infrabasal plates, nodose projection on first primibrachials, column impression on infrabasal circlet, and shape of the proximal columnal. Sinocrinus granulatus has a low cone-shaped aboral cup, fine granulose aboral cup plate sculpturing, circular calyx outline in basal view, basal concavity occupies <60% of infrabasal circlet diameter, all infrabasal plates are of equal height, nodose projections on distal side of first primibrachials, pronounced impression of proximal column crenulae on infrabasal circlet, and proximal column circular; S. lichengensis has a low cone-shaped aboral cup, faintly granulose aboral cup plate sculpturing, slightly pentalobate calyx outline in basal view, basal concavity occupies <60% of infrabasal circlet diameter, individual infrabasal plates of varying heights, first primibrachials unknown, pronounced impression of proximal column crenulae on infrabasal circlet, and proximal column pentalobate; S. obliquus has a low conical aboral cup shape, coarse irregular granulose aboral cup plate sculpturing, circular calyx outline in basal view, basal concavity occupies <60% of infrabasal circlet diameter, individual infrabasal plates of varying heights, first primibrachials unknown, pronounced impression of proximal column crenulae on infrabasal circlet, and proximal column circular; and S. sheareri has a very low bowl aboral cup shape, smooth aboral cup plate sculpturing, circular calyx outline in basal view, basal concavity occupies 100% of infrabasal circlet diameter, all infrabasal plates are of equal height, first primibrachials unknown, moderately developed impression of proximal column crenulae on infrabasal circlet, and proximal column circular. In contrast, S. websteri sp. nov. has a low bowl-shaped aboral cup, unknown aboral cup plate sculpturing, circular calyx outline in basal view, basal concavity occupies >60% of infrabasal circlet diameter, all infrabasal plates are of equal height, nodose projections on distal side of first primibrachials, moderately developed impression of proximal column crenulae on infrabasal circlet, and proximal column circular.
Pennsylvanian Sinocrinus were cosmopolitan with a distribution including China, Spain, and USA. However, Permian occurrences of Sinocrinus include now Sinocrinus granulatus and S. obliquus in the Artinskian of Timor, S. lichengensis in the Sakmarian of Crete, Greece, S. shearieri from the Asselian of the North China Block, and S. websteri sp. nov. in the middle to upper Artinskian of Slovenia.
Stratigraphic and geographic range.—Trogkofel Group, Permian, Cisuralian, middle to upper Artinskian, near Dovje, Slovenia.
Family Protencrinidae Knapp, 1969
Genus Protencrinus Jaekel, 1918
Type species: Protencrinus moscoviensis Jaekel, 1918; Pennsylvanian (Moscovian), Russia.
Included species: Protencrinus atoka (Strimple, 1961); Protencrinus baliensis Webster, 2012b; Protencrinus lobatus Yakovlev, 1948; Protencrinus moscoviensis Jaekel, 1918; Protencrinus mutabilis Knapp, 1969; Protencrinus? sp. indet. in Hashimoto, 1984.
Remarks.—Sutural contact between the infrabasal plates with radial plates is a very unusual character in crinoids. This character is present on all rays or present in some rays in all species of Protencrinus. It also occurs in one species of Prolobocrinus, i.e., P. striatus Wanner, 1937. Protencrinus and Prolobocrinus are distinguished by, among other things, the height of the crown, morphology of the radial facets, the size and position of the basal and radial plates, the aboral cup plate sculpturing, and the anal sac.
At present five Protencrinus species are named, and one species indeterminate is designated. Species of Protencrinus are differentiated on the basis of outline of the aboral cup in oral view, relative depth of the basal concavity, presence and/or absence of sutural contact between the infrabasal circlet and radial plate (and following whether basal plates are separated laterally), if present the relative width of the infrabasal sutural contact between the infrabasal plate and radial plate, and relative size of the basal plates.
Stratigraphic and geographic range.—Lower Permian, Crete (Greece), West Timor; Permian, Cisuralian, Artinskian, Slovenia.
Protencrinus baliensis Webster, 2012b
Figs. 4C, 6A.
Type material: Holotype: CrdKap12, aboral cup without arms or column. Paratype: CrdKap11, moderately preserved aboral cup.
Type locality: Crete, Greece.
Type horizon: Galinos Beds, Permian (Cisuralian).
Material.—GMJ PAL-0000002, aboral cup without arms or column from Permian, Cisuralian, middle to upper Artinskian, Slovenia.
Diagnosis.—Aboral cup subpentagonal in outline, shallow basal concavity, smooth sculpturing, relative width of suture between infrabasal plates and radial plates wide, basals not in lateral contact, basal plates relatively large.
Measurements.—ACW, ~21.1.
Description.—Aboral cup flat(?) bowl shape, subpentagonal in outline, shallow basal concavity, plates gently convex, smooth sculpturing. Infrabasal circlet extends well beyond proximal columnal; five infrabasal plates, wider than high, in sutural contact with radial plates distally. Basal circlet visible in lateral view, interrupted in all rays (Fig. 6A); basal plates five, tetragonal with each side convex, as wide as high, smaller than radial plates. Radial circlet not interrupted in CD interray; radial plates five, hexagonal, much wider than high. Radial facets plenary, other details of the radial facets unknown. No posterior interray plates in aboral cup. Oral surface, anal sac, arms, and column unknown.
Remarks.—Protencrinus baliensis is diagnosed by having an oral cup subpentagonal in outline, shallow basal concavity, smooth sculpturing, relative width of suture between infrabasal plates and radial plates wide, basals not in lateral contact, basal plates relatively wide. In contrast, P. atoka has an oral cup subcircular in outline, shallow? basal concavity, presumably smooth sculpturing, relative width of suture between infrabasal plates and radial plates narrow or absent, lateral contact between radial and basal plates present or absent, basal plates relatively wide; P. lobatus has an oral cup pentalobate in outline, deep basal concavity, smooth plate sculpturing, relative width of suture between infrabasal plates and radial plates narrow, basals not in lateral contact, basal plates relatively small; P. moscoviensis has an oral cup circular in outline, shallow basal concavity, relative width of suture between infrabasal plates and radial plates narrow, basals not in lateral contact, basal plates relatively wide; P. mutabilis has an oral cup subcircular in outline, deep basal concavity, relative width of suture between infrabasal plates and radial plates narrow or absent, lateral contact between basal plates present or absent, basal plates relatively narrow; and Protencrinus? sp. indet. (in Hashimoto 1984) has an oral cup subpentagonal in outline, shallow basal concavity, suture between infrabasal plates and radial plates wide narrow or absent, lateral contact between basal plates present or absent, basal plates relatively wide.
Stratigraphic and geographic range.—Lower Permian, series undesignated, Crete, Greece; Permian, Cisuralian, Asselian, Dajian Member, Taiyuan Formation, North China; Trogkofel Group, Permian, Cisuralian, middle to upper Artinskian, near Dovje, Slovenia.
Eucladida indet. A
Fig. 4G.
Material.—GMJ:PA-0000014b, isolated aboral cup with radial facets broken from radial plates, from lower Permian, West Timor; Permian, Cisuralian, middle to upper Artinskian, Slovenia.
Measurements.—ACH, 16.7; ACW, ~36.0.
Description.—Aboral cup low bowl shape, height width ratio ~0.55; plates convex, smooth sculpturing. Infrabasal circlet very small, presumably hidden beneath proximal columnal. Basal plate circlet large; basal plates appear to be fused but five basals separated by faint sutures. Radial plates five, pentagonal, slightly wider than high. Radial facets plenary, horizontally oriented, articular ridge probably across full width of facet, but other details of radial facet not known. Posterior interray, oral surface, anal sac, and arms unknown. Proximal columnal circular; other aspects of the column unknown.
Remarks.—Eucladida indet. A (Fig. 4G) has a very distinctive morphology with a very small infrabasal circlet completely covered by the proximal columnal and a large, convex basal circlet in which the plate sutures are so faint that the basal plates appear to be fused into a single plate. This appears to be a unique morphology for Permian crinoids, but the non-preservation of the posterior interray and arms preclude naming a new taxon.
The unusual character of the basal circlet is similar to species of Bronaughocrinus Strimple, 1951; Pentaramicrinus Sutton and Winkler, 1940; Cryphiocrinus Kirk, 1929; and Contocrinus Knapp, 1969—all of which are Carboniferous in age.
Crinoidea indet.
Figs. 3, 4B, D, E, 5A, B.
Material.—PMSLJ Ba369, Ba370, Ba374, Ba381, Ba1744, GMJ:PAL-0000014d, 0000370, 0000372, 000373a, b, from Permian, Cisuralian, middle to upper Artinskian, Slovenia.
Remarks.—Based on incompletely preserved arms, aboral cups, isolated infrabasal circlets, and columnals and pluricolumnals, crinoids named above are only a small part of the overall crinoid diversity in the Trogkofel Group. Fig. 5A is an incomplete set of arms that are broad and flat to gently convex, and the brachials are chisel biserial. Fig. 5B is also an incomplete set of arms with chisel biserial brachials. These incomplete sets of arms belong to two taxa. The individual brachials in Fig. 5A are flat to gently convex, whereas those in Fig. 5B are moderately convex with a nodose appearance. Without knowing the structure of the aboral cup it is not possible to identify these arms to a genus.
Infrabasal circlets for a cladid crinoid (Fig. 4D, E) and are most likely distinct taxa, but the specimens are too incomplete or too poorly preserved to assign to a genus or species. Fig. 4D is 18.4 mm in diameter. It is an infrabasal circlet with a very wide column attachment covering ~50% of the diameter of the circlet with a narrow crenularium and a wide areola. Five infrabasal plates are in this circlet. Similarly, Fig. 4E is an infrabasal circlet that cannot be assigned to a genus. It is 31.6 mm in diameter, and it contrasts with Fig. 4D by having a very small column attachment covering only ~18% of the circlet width. It has a raised rim around the column attachment and radially arranged ridges and grooves.
In addition to the Platycrinitidae columnal discussed above, many isolated columnals and pluricolumnals are present in the Trogkofel Group fauna (Fig. 3). They are preserved both in calcite (e.g., Fig. 3D) or in moldic preservation (Fig. 3C). Sizes of these columnals and pluricolumnals vary considerably with the largest column diameter ~45 mm and the largest pluricolumnal height 80.2 mm. Fig. 3F is probably the distal, holdfast portion of a column. In addition to a few columnals, numerous plates from the partially disarticulated radices are preserved. With the exception of the Platycrinitidae columnals noted above, isolated columnals and pluricolumnals cannot be linked to crown-based taxa.
Discussion
Taphonomy.—With the possible exception of Dieuryocrinus, all of the crinoids known from the Trogkofel Group belong to clades with few aboral cup/calyx plates that are not firmly cemented together during life. Even Dieuryocrinus with multiple calyx plates has calyx plates that were not firmly cemented during life, hence the superorder name Flexibilia. Meyer (1971) determined that a dead crinoid lying on the seafloor would disarticulate within a few days. Consequently, in order to be preserved with arms and column, a crinoid must be buried intact rapidly and permanently. None of the specimens described herein were preserved with arms, calyx, and the proximal column. Preservation of complete specimens is typically the result of burial by a tempestite (storm deposit) (Lewis 1980; Donovan 1991; Taylor and Brett 1996; Brett et al. 1997; Ausich and Sevastopulo 1994; Ausich 2001; among others). Furthermore, once buried, fossil echinoderms with complete or nearly complete preservation were not exhumed by a subsequent high-turbulence event.
Ausich and Sevastopulo (1994) considered crinoid preservation along a Mississippian palaeoslope in southern Ireland. They concluded that the middle slope setting was optimal for preservation of complete specimens. High-turbulence events that buried living organisms would have been less common along the middle slope than along the upper slope, and the majority of storm events did not reach deep into storm wave base to exhume buried crinoids. In contrast, in an upper shelf setting, the frequency and depth of sediment disturbance was high and disarticulated most echinoderms prior to final burial. At the other extreme, deep shelf settings were well below storm wave base and had few to no disturbances of the sea floor. Crinoids that lived in deep shelf settings typically disarticulated on the seafloor prior to burial.
Novak and Skaberne (2009) interpreted the Trogkofel Group as a shallow platform or upper slope platform palaeoenvironmental setting, where the surface of the sediment would have been subjected to relatively common high-turbulence events. Thus, the lack of complete crinoids in the Trogkofel Group is consistent with predictions of Ausich and Sevastopulo (1994). In this setting, the preservation of common complete specimens is not anticipated.
Stratigraphic and palaeogeographic distribution of Trogkofel Group crinoids.—By the middle of the 20th century, the vast majority of described Permian crinoids were from Timor (e.g., Wanner 1916, 1924, 1937, 1949; Marez Oyens 1940), but the dating of this Permian material was uncertain. As noted above, our understanding of Permian crinoids has expanded greatly since 1940s and the fauna of the Trogkofel Group adds to this understanding. Permian crinoids are especially significant because the class Crinoidea came very close to total extinction at the end of the Paleozoic. Thus, delineating the temporal history of crinoids through the Permian is critical.
A genus-level comparison of Permian crinoids from Slovenia versus the temporal and geographic distribution of other occurrences of these genera is given in Fig. 7. The Trogkofel Group crinoids are Artinksian, Cisuralian, and all other occurrences of these Trogkofel Group genera are Cisuralian: either Asselian, Sakmarian, Artinskian, or in an age range that overlaps with the Artinskian. The only Trogkofel Group species known elsewhere is Protencrinus baliensis, which occurs of the lower Permian of Crete and China (Webster 2012b; Mao et al. 2018). At the genus level, co-occurrences include Australia, Canada, China Crete (Greece), Oman, Russia, Sicily (Italy), USA, and West Timor. Most co-occurrences are from Australia, Crete, Sicily, and West Timor (Fig. 7).

Fig. 7. Range chart comparing Permian crinoid genera from Slovenia with occurrences of these genera elsewhere in the world. Black, range known within one stage; dark gray, range known with two stages; light gray, lower Permian but stage unknown. Black, range known within one stage; dark gray, range known with two stages; light gray, lower Permian but stage unknown.
Conclusions
The lower Permian (Artinskian) Trogkofel Group of the Julian Alps in Slovenia was deposited in shallow marine conditions, shallow platform or upper slope settings. In carbonate and clastic sediments of this group, the fauna is diverse and abundant. The crinoid fauna includes crinoid cups of Protencrinus baliensis, which was previously only described from lower Permian sediments of Crete and China. The remaining crinoids are represented by one new genus and four new species, designated as Moapacrinus dovjensis sp. nov., Parastachyocrinus sloveniaensis sp. nov., Sinocrinus websteri sp. nov., and Karavankecrinus bedici gen. et sp. nov. Fragmentary aboral cups, infrabasal circlets, incompletely preserved arms, columnals and pluricolumnals are also recorded; but due to their fragmentary nature, are left in open nomenclature. These include Dieuryocrinus? sp., Platycrinitidae indet., Eucladida indet. A, and Crinoidea indet. It must be noted that all crinoids, with the possible exception of Dieuryocrinus? sp., were from clades in which calyx plates were typically not firmly cemented together during their life cycle. The state of preservation of Trogkofel Group crinoids is consistent with preservation in relatively shallow water settings influenced by frequent storms.
Acknowledgements
We thank Jan Bohatý (Hessian State Office for the Preservation of Historic Monuments and Sites, Germany) and Przemysław Gorzelak (Institute of Paleobiology PAS, Warsaw, Poland), who provided careful reviews that helped us improve this paper. Wojciech Krawczyński (University of Silesia, Katowice, Poland) is acknowledged for preparing macrophotographs, Marko Mugerli (Upper Sava Museum, Jesenice, Slovenia) for loaning some crinoids for current study, and Natasji den Ouden (RGM) helped with original specimens described by Johannes Wanner. This research project was supported by the “Projekty Badawcze 2023” (Faculty of Natural Sciences, University of Silesia in Katowice) to MAS.
References
Arendt, Yu.A. 1968. Pirasocrinids from Krasnoufimsk [in Russian]. Paleontological Journal 4: 99–101.
Arendt, Yu.A. 1970. The sea lilies cyrtocrinids [in Russian]. Akademiâ Nauk SSSR, Trudy Palentologičeskogo Instituta 128: 3–220.
Ausich, W.I. 2001. Echinoderm taphonomy. In: M. Jangoux and J.M. Lawrence (eds.), Echinoderm Studies. Volume 6, 171–227. A. Balkema, Rotterdam.
Ausich, W.I. and Donovan, S.K. 2023. Part T. Revised, Volume 1, Chapter 7: Glossary of crinoid morphological terms. Treatise Online 167: 1–26. Crossref
Ausich, W.I. and Kammer, T.W. 2009. Generic concepts in the Platycrinitidae Austin and Austin (Class Crinoidea). Journal of Paleontology 83: 694–717. Crossref
Ausich, W.I. and Sevastopulo, G.D. 1994. Taphonomy of Lower Carboniferous crinoids from the Hook Head Formation, Ireland. Lethaia 27: 245–256. Crossref
Ausich, W.I., Wright, D.F., Cole, S.R., and Sevastopulo, G.D. 2020. Homology of posterior interray plates in crinoids: A review and new perspectives from phylogenetics, the fossil record, and development. Palaeontology 63: 525–454. Crossref
Austin, T. and Austin, T. 1842. Proposed arrangement of the Echinodermata, particularly as regards the Crinoidea, and a subdivision of the Class Adelostella (Echinidae). Annals and Magazine of Natural History 10: 106–113. Crossref
Austin, T. and Austin, T. 1843. Description of several new genera and species of Crinoidea. Annals and Magazine of Natural History 11: 195–207. Crossref
Bassler, R.S. and Moodey, M.W. 1943. Bibliographic and faunal index of Paleozoic pelmatozoan echinoderms. Geological Society of America Special Paper 45: 1–734. Crossref
Bather, F.A. 1890. British fossil crinoids. II. The classification of the Inadunata. Annals and Magazine of Natural History 5: 310–334. Crossref
Branson, C.C. 1948. Bibliographic index of Permian invertebrates. Geological Society of America, Memoir 26: 1–1049. Crossref
Brett, C.E., Moffat, H.A., and Taylor, W.L. 1997. Echinoderm taphonomy, taphofacies, and lagerstätten. In: J.A. Waters and C.G. Maples (eds.), Geobiology of Echinoderms. The Paleontological Society Papers 3: 147–190. Crossref
Burke, J.J. 1977. Eperisocrinus new genus (Crinoidea, Inadunata): type species Delocrinus missouriensis Miller and Gurley, 1890. Ohio Journal of Science 77: 174–178.
Burke, J.J. and Pabian, R.K. 1978. Two crowns of Cibolocrinus patriciae (Crinoidea, Flexibilia) from the lower Permian of Bolivia. Journal of Paleontology 52: 1065–1069.
Charlton, T.R., Barber, A.J., Harris, R.A., Barkham, S.T., Bird, P.R., Archbold, N.W., Morris, N.J., Nicoll, R.S., Owen, H.G., Owens, R.M., Sorauf, J.E., Taylor, P.D., Webster, G.D., and Whittaker, J.E. 2002. The Permian of Timor: stratigraphy, palaeontology and palaeogeography. Journal of Asian Earth Sciences 20: 719–774. Crossref
Chen, Z-t., and Yao, J-h. 1993. Palaeozoic Echinoderm Fossils of Western Yunnan, China. 102 pp. Geological Publishing House, Beijing.
Cole, S.R. 2017. Phylogeny and morphologic evolution of the Ordovician Camerata (Class Crinoidea, Phylum Echinodermata). Journal of Paleontology 91: 815–828. Crossref
Cole, S.R. 2018. Phylogeny and evolutionary history of diplobathrid crinoids (Echinodermata). Palaeontology 62: 357–373. Crossref
Donovan, S.K. 1986. Pelmatozoan columnals from the Ordovician of the British Isles, Part 1. Monograph of the Palaeontographical Society 138: 1–68. Crossref
Donovan, S.K. 1989. Pelmatozoan columnals from the Ordovician of the British Isles, Part 2. Monograph of the Palaeontographical Society 142: 69–114. Crossref
Donovan, S.K. 1991. The taphonomy of echinoderms. calcareous multi-element skeletons in the marine environment. In: S.K. Donovan (ed.), The Process of Fossilization, 241–269. Belhaven Press, London.
Donovan, S.K. 1995. Pelmatozoan columnals from the Ordovician of the British Isles, Part 3. Monograph of the Palaeontographical Society 149: 115–193. Crossref
Głuchowski, E. 2002. Crinoids from the Famennian of the Holy Cross Mountains, Poland. Acta Palaeontologica Polonica 47: 319–328.
Gorzelak, P., Błażejowski, B., Uchman, A., and Hanken, N. 2013. First record of catacrinid crinoid from the lower Permian of Spitsbergen. Polish Polar Research 34: 139–150. Crossref
Gregorio, A., de 1930. Sul Permiano di Sicilia. Annales de Géologie et de Paléontologie 52: 1–67.
Holtedahl, O. 1911. Zur Kenntnis der Karbonablagcrungen des westlichen Spitzbergens I. Eine Fauna der Moskauer Stufe. Videnskabernes Selskabs Skrifter 10: 1–89.
Holterhoff, P.F. 1997. Filtration models, guilds, and biofacies: crinoid paleoecology of the Stanton Formation (Upper Pennsylvanian), midcontinent, North America. Palaeogeography, Palaeoclimatology, Palaeoecology 130: 177–208. Crossref
Hashimoto, K. 1984. Preliminary study of Carboniferous crinoid calyces from the Akiyoshi Limestone Group, southwest Japan. Bulletin of the Yamaguchi Prefectural Yamaguchi Museum 10: 1–53.
Hashimoto, K. 1995. Akiyoshicrinus, a new erisocrinid crinoid genus from the Upper Carboniferous of the Akiyoshi Limestone Group, southwest Japan (Crinoid studies of Akiyoshi-1). Bulletin of the Akiyoshi-dai Museum of Natural History 30: 1–14.
Jaekel, O. 1894. Über die Morphogenie und Phylogenic der Crinoiden. Sitzungsberichten der Gesellschaft Naturforschender Freunde, Jahrgang 1894 4: 101–121.
Jaekel, O. 1918. Phylogenie und System der Pelmatozoen. Paläontologische Zeitschrift 3: 1–128. Crossref
Jell, P.A. and Willink, R.J. 1993. Early Permian cladid crinoids from the Gharif Formation of Oman. In: P.A. Jell (ed.), Palaeontological studies in honour of Ken Campbell. Memoir Association Australasian Palaeontologists 15: 305–312.
Kammer, T.W., Sumrall, C.D., Zamora, S., Ausich, W.I., and Deline, B. 2013. Oral region homologies in Paleozoic crinoids and other plesiomorphic pentaradiate echinoderms. PLoS ONE 8: e7798. Crossref
Kirk, E. 1929. Cryphiocrinus, a new genus of free-swimming crinoids. American Journal of Science 17: 153–161. Crossref
Kirk, E. 1940. A redescription of Lageniocrinus de Koninck. American Journal of Science 238: 129–139. Crossref
Knapp, W.D. 1969. Declinida, a new order of late Paleozoic inadunate crinoids. Journal of Paleontology 43: 340–391.
Križnar, M. 2022. Lower Permian (Artinskian) chondrichthyan tooth remains (Petalodontidae) from Dovje (Karavanke Mts., NW Slovenia). Geologija 65: 217–224. Crossref
Lach, R., Gale, L., Križnar, M., and Novak, M. 2013. Nagromadzenia enigmatycznych permskich liliowców z Dovje (Karawanki, północna Słowenia). XXII Konferencja Naukowa Sekcji Paleontologicznej Polskiego Towarzystwa Geologicznego, Aktualizm i antyaktualizm w paleontologii, 27–30.09.2013, Tyniec, 32–33. Polskie Towarzystwo Geologiczne, Tyniec.
Lane, N.G. 1979. Upper Permian crinoids from Djebel Tebaga, Tunisia. Journal of Paleontology 53: 121–132.
Lane, N.G. and Webster, G.D. 1966. New Permian crinoid fauna from southern Nevada. University of California Publications in Geological Sciences 63: 1–60.
Lane, N.G. and Webster, G.D. 1980. Crinoidea. In: J.A. Waters and T.W. Broadhead (eds.), Echinoderms: Notes for a Short Course. University of Tennessee Studies in Geology 3: 144–157. Crossref
Lewis, R. 1980. Taphonomy. In: T.W. Broadhead and J.A. Waters (eds.), Echinoderms, Notes for a Short Course. University of Tennessee, Studies in Geology 3: 27–39. Crossref
M’Coy, F. 1847. On the fossil botany and zoology of the rocks associated with the coal of Australia. Annales and Magazine of Natural History 20: 226–236. Crossref
Mao, Y., Webster, G.D., Ausich, W.I., Li, Y., Wang, Q., and Reich, M. 2018. A new crinoid fauna from the Taiyuan Formation (early Permian) of Henan, North China. Journal of Paleontology 92: 1066–1080. Crossref
Marez Oyens, F.A.H.W., de 1940. Neue permische Krinoiden von Timor, mit Bemerkungen über deren Vorkommen in Basleogebiet. Geological expdition to the Lesser Sunda Islands under leadership of H.A. Brouwer. N.V. Noord-Holl Uitveg Mijnbouw, Amsterdam 1: 285–348.
Meek, F.B. and Worthen, A.H. 1865. Remarks on the Carboniferous and Cretaceous rocks of eastern Kansas and Nebraska. American Journal of Science 39: 157–174. Crossref
Meyer, D.L. 1971. Post-mortem disarticulation of Recent crinoids and ophiuroids under natural conditions. Geological Society of America Abstracts with Programs 3: 645.
Miller, J.S. 1821. A Natural History of the Crinoidea, or Lily-shaped Animals; with Observations on the Genera, Asteria, Euryale, Comatula and Marsupites. 150 pp. C. Frost, Bristol. Crossref
Moore, R.C. 1939. New crinoids from Upper Pennsylvanian and Lower Permian rocks of Oklahoma, Kansas, and Nebraska. Denison University Bulletin. Journal Sciences Laboratory 34: 171–279.
Moore, R.C. 1978. Flexibilia. In: R.C. Moore and C. Teichert (eds.), Treatise on Invertebrate Paleontology. Part T Echinodermata 2, T759–T812. Geological Society of America and University of Kansas Press, Boulder.
Moore, R.C. and Jeffords, R.M. 1968. Classification and nomenclature of fossil crinoids based on studies of dissociated parts of their columns. University of Kansas Paleontological Contributions 46: 1–86.
Moore, R.C. and Laudon, L.R.: 1943. Evolution and classification of Paleozoic crinoids. Geological Society of America Special Paper 46: 1–154. Crossref
Moore, R.C. and Plummer, F.B. 1940. Crinoids from the upper Carboniferous and Permian strata in Texas. University of Texas Publication 468: 209–313.
Moore, R.C. and Strimple, H.L. 1973. Lower Pennsylvanian (Morrowan) crinoids from Arkansas, Oklahoma, and Texas. University of Kansas Paleontological Contributions, Article 60, Echinodermata 12: 1–84.
Moore, R.C. and Strimple, H.L. 1978a. Superfamily Allagecrinacea Carpenter and Etheridge, 1881. In: R.C. Moore and C. Teichert (eds.), 1978, Treatise on Invertebrate Paleontology. Part T Echinodermata 2, T537–T548. Geological Society of America and University of Kansas Press, Boulder.
Moore, R.C. and Strimple, H.L. 1978b. Superfamily Apographiocrinacea Moore and Laudon, 1943. In: R.C. Moore and C. Teichert, C. (eds.), 1978, Treatise on Invertebrate Paleontology. Pt T Echinodermata 2, T720–T722. Geological Society of America and University of Kansas Press, Boulder.
Moore, R.C. and Teichert, C. (eds.) 1978. Treatise on Invertebrate Paleontology. Part T Echinodermata 2. 1027 pp. Geological Society of America and University of Kansas Press, Boulder.
Moore, R.C., Strimple, H.L., and Lane, N.G. 1978. Superfamily Erisocrinacea. In: R.C. Moore and C. Teichert (eds.), 1978, Treatise on Invertebrate Paleontology. Part T Echinodermata 2, T704–T720. Geological Society of America and University of Kansas Press, Boulder.
Novak, M. and Skaberne, D. 2009. Zgornji karbon in spodnji perm (Upper Carboniferous and Lower Permian). In: M. Pleničar, B. Ogorelec, and M. Novak (eds.), Geologija Slovenije, 99–136. Geološki zavod Slovenije, Ljubljana.
Novak, M., Forke, H.C., and Schönlaub, H.P. 2019. The Pennsylvanian–Permian of the Southern Alps (Carnic Alps/Karavanke Mts.), Austria/Italy/Slovenia—fauna, facies, and stratigraphy of a mixed carbonate-siliciclastic shallow marine platform along the northwestern Palaeotethys margin. Field Trip C3. 19th International Congress on the Carboniferous and Permian, July 29–August 2, 2019, 251–302. University of Cologne, Cologne.
Pabian, R.K. and Rushlau, W.J. 2002. Taphonomic analysis and systematic descriptions of some Late Pennsylvanian and Early Permian crinoids from southeastern Nebraska, eastern Kansas and southwestern Iowa. Conservation and Survey Division Institute of Agriculture and Natural Resources Geological Survey Paper 20: 1–45.
Pabian, R.K. and Strimple, H.L. 1974. Crinoid studies part I. Some Pennsylvanian crinoids from Nebraska. Part II. Some Permian crinoids from Nebraska, Kansas, and Oklahoma. Bulletins of American Paleontology 64: 247–337.
Pabian, R.K. and Strimple, H.L. 1993. Taxonomy, paleoecology and biostratigraphy of the crinoids of the South Bend Limestone (Late Pennsylvanian-Missourian, ?Virgilian) in southeastern Nebraska and southeastern Kansas. Conservation and Survey Division. University of Nebraska-Lincoln, Professional Paper 1: 1–55.
Ramovš, A. 1978. Okamnelo življenje v jeseniškem prostoru: vodnik po paleontološki zbirki Tehniškega muzeja Železarne Jesenice. 80 pp. Tehniški muzej Železarne Jesenice, Jesenice.
Ramovš, A. and Kochansky-Devide, V. 1979. Karbonske in permske plasti v severnih Julijskih Alpah. Geologija 22: 21–54.
Ramovš, A. and Sieverts-Doreck, H. 1968. Interessante Mittelperm-Crinoiden in Slowenien, NW Jugoslavien. Geološki vjesnik 21: 191–206.
Roemer, C.F. 1852–1854. Erste Periode, Kohlen-Gebirge. In: H.G. Bronn (ed.), Lethaea Geognostica, 3rd Edition, Vol. 2, 1–788. E. Schweizerbart, Stuttgart.
Sant, J.F. van 1964. Crawfordsville crinoids. In: J.F. van Sant and N.G. Lane (eds.), Crawfordsville (Indiana) Crinoid Studies, Article 7 Echinodermata, 34–136. University of Kansas Paleontological Contributions, {town}.
Sheffield, S.L. 2015. A brief history and revision of the systematics of the cladid crinoid Sinocrinus. Palaeoworld 25: 460–469. Crossref
Sieverts-Doreck, H. 1942. Crinoiden aus dem Perm Tasmanien. Zentralblatt für Mineralogie etc. (B: Geologie und Paläontologie) 1942: 222–231.
Springer, F. 1913. Crinoidea. In: K.A. von Zittel (ed.), Text-book of Paleontology (translated and edited by C.R. Eastman), 2nd Edition, Vol. 1, 173–243. Macmillan & Co., Ltd., London.
Strimple, H.L. 1951. New Carboniferous crinoids. Journal of Paleontology 25: 669–676.
Strimple, H.L. 1961. New Paradelocrinus from Oklahoma. Oklahoma Geology Notes 21: 225–229.
Strimple, H.L. 1971. A Permian crinoid from Coahuila, Mexico. Journal of Paleontology 45: 1040–1042.
Strimple, H.L. 1976. Petalambicrinus, a new inadunate crinoid genus from the Missourian of Texas. Journal of Paleontology 50: 865–868.
Strimple, H.L. and Moore, R.C. 1971. A crinoid crown from d’Orbigny’s famous fossil locality at Yaurichampi. University of Kansas Paleontological Contributions, Paper 56: 33–35.
Strimple, H.L. and Sevastopulo, G.D. 1982. A Permian microcrinoid from Sicily. Journal of Paleontology 56: 1451–1452.
Strimple, H.L. and Watkins, W.T. 1969. Carboniferous crinoids of Texas with stratigraphic implications. Palaeontographica Americana 6: 139–275.
Sutton, A.H. and Winkler, V.D. 1940. Mississippian Inadunata—Eupachycrinus and related forms. Journal of Paleontology 14: 544–567.
Taylor, W.L. and Brett, C.E. 1996. Taphonomy and paleoecology of echinoderm Lagerstätten from the Silurian (Wenlockian) Rochester Shale. Palaios 11: 118–140. Crossref
Teichert, C. 1949. Permian crinoid Calceolispongia. Memoirs of the Geological Society of America 34: 1–132. Crossref
Teichert, C. 1954. A new Permian crinoid from Western Australia. Journal of Paleontology 28: 70–75.
Teichert, C. and Webster, G.D. 1993. Crinoids. In: S.K. Skwarko (ed.), Palaeontology of the Permian of Western Australia. Geological Survey of Western Australia, Bulletin 136: 68–72, 322–339; microfiche sheet 8, 1–26.
Termier, H. and Termier, G. 1949. Hierarchie et correlations des caracteres chez les crinoides fossiles. Bulletin Service de la Carte géologique Algerie, 1st Series, Paleontologie 10: 1–91.
Termier, H. and Termier, G. 1958. Les echinodermes permiens du Djebel Tebaga (extreme sud Tunisien). Bulletin de la Société géologique de France 6: 51–64. Crossref
Termier, H. and Termier, G. 1977. Monographie paleontologique des affleurements Permiens du Djebel Tebaga (Sud Tunisien). Palaeontographica 156: 1–109.
Tien, C.C. 1926. Crinoids from the Taiyuan Series of North China. Palaeontologica Sinica, ser. B 5: 1–58.
Trautschold, H. 1867. Einige crinoideen und andere Thierreste des Jungeren Bergkalks im Gouvernment Moskau. Bulletin de la Societie Imperial Naturalistes de Moscou 15: 1–49.
Ubaghs, G. 1978a. General morphology. In: R.C. Moore and C. Teichert (eds.), Treatise on Invertebrate Paleontology. Part T Echinodermata 2, T58–T216. Geological Society of America and University of Kansas Press, Boulder.
Ubaghs, G. 1978b. Subphylum Camerata. In: R.C. Moore and C. Teichert (eds.), Treatise on Invertebrate Paleontology. Partt T Echinodermata 2, T410–T519. Geological Society of America and University of Kansas Press, Boulder.
Valette, D.A. 1934. Le Permien marin de l’extrême-sud de la Tunisien. III. Les Crinoides permiens du sud de la Tunisie. Memoir Service Carte Geologie, Tunisie, n.s 1: 91–101.
Waagen, W. 1887. Salt-Range fossils. I. Productus-Limestone fossils. 5. Bryozoa-Annelida-Echinodermata. Palaeontologica Indica 13: 771–834.
Wachsmuth, C. and Springer, F. 1880–1886. Revision of the Palaeocrinoidea. Proceedings of the Academy of Natural Sciences of Philadelphia Pt. I. The families Ichthyocrinidae and Cyathocrinidae (1880), 226–378 (separate repaged, 1–153). Pt. II. Family Sphaeroidocrinidae, with the sub-families Platycrinidae, Rhodocrinidae, and Actinocrinidae (1881), 177–411, (separate repaged, 1–237. Pt. III, Sec. 1. Discussion of the classification and relations of the brachiate crinoids, and conclusion of the generic descriptions (1885), 225–364 (separate repaged, 1–138). Pt. III, Sec. 2. Discussion of the classification and relations of the brachiate crinoids, and conclusion of the generic descriptions (1886), 64–226 (separate repaged, 139–302).
Wanner, J. 1916. Die Permischen echinodermen von Timor, I. Teil. Palaontologie von Timor 11: 1–329.
Wanner, J. 1924. Die permischen Krinoiden von Timor. Jaarbook van net Mijnwezen Nederlandes Oost-Indie 1921: 1–348.
Wanner, J. 1937. Neue Beiträge zur Kenntnis der permischen Echinodermen von Timor, VIII–XIII. Palaeontographica 4 (Supplement): 1–212.
Wanner, J. 1949. Neue Beiträge zur kenntnis der Permischen Echinodermen von Timor, Poteriocrinidae 16, Teil 4. Palaeontographica 4 (Supplement): 1–56.
Webster, G.D. 1973. Bibliography and index of Paleozoic crinoids, 1942–1968. Geological Society of America, Memoir 137: 1–341.
Webster, G.D. 1974. Crinoid pluricolumnal noditaxis patterns. Journal of Paleontology 48: 1283‒1288.
Webster, G.D. 1987, Permian crinoids from the type-section of the Callytharra Formation, Callytharra Springs. Western Australia. Alcheringa 11: 95–135. Crossref
Webster, G.D. 1990. New Permian crinoids from Australia. Palaeontology 33: 49–73.
Webster, G.D. 1998a. Distortion in the stratigraphy and biostratigraphy of Timor, a historical review with an analysis of the crinoid and blastoid faunas. Proceedings of the Royal Society of Victoria 110: 45–72.
Webster, G.D. 1998b. Palaeobiogeography of Tethys Permian crinoids. Proceedings of the Royal Society of Victoria 110: 289–308.
Webster, G.D. 2012a. A canted-cup Permian crinoid Exotikocrinus n. gen. (Crinoidea, Dichocrinidae) from Timor with comments on canted or inclined radial summits. Palaeoworld 21: 64–68. Crossref
Webster, G.D. 2012b. An early Permian crinoid fauna from Crete. In: A. Kroh and M. Reich (eds.), Echinoderm Research 2010. Proceedings of the Seventh European Conference on Echinoderms. Göttingen Germany, 2–9 October 2010. Zoosymposia 7: 101–110. Crossref
Webster, G.D. and Webster, D.W. 2014. Bibliography and Index of Paleozoic Crinoids, Coronates, and Hemistreptocrinoids, 1758–2012 [available online, http://crinoids.azurewebsites.net/, accessed 6 February 2023].
Webster, G.D. and Donovan, S.K. 2012. Revision of two species of Ulocrinus and a new pelecocrinid crinoid from West Timor. Palaeoworld 21: 108–115. Crossref
Webster, G.D. and Jell, P.A. 1992. Permian echinoderms from Western Australia. Memoirs of the Queensland Museum 32: 311–373.
Webster, G.D. and Jell, P.A. 1993. Early Permian inadunate crinoids from Thailand. Memoirs of the Queensland Museum 33: 349–359.
Webster, G.D. and Jell, P.A. 1999. New Permian crinoids from eastern Australia. Memoirs of the Queensland Museum 43: 279–340.
Webster, G.D. and Lane, N.G. 1967. Additional Permian crinoids from southern Nevada. University of Kansas Paleontological Contributions, Paper 27: 1–32.
Webster, G.D. and Lane, N.G. 2007. New Permian crinoids from the Battleship Wash patch reef in southern Nevada. Journal of Paleontology 81: 951–965. Crossref
Webster, G.D. and Maples, C.G. 2008. Cladid crinoid radial facets, brachials, and arm appendages: a terminology solution for studies of lineage, classification, and paleoenvironment. In: W.I. Ausich and G.D. Webster (eds.), Echinoderm Paleobiology, 196–226. Indiana University Press, Bloomington.
Webster, G.D. and Sevastopulo, G.D. 2007. Paleogeographic significance of early Permian crinoids and blastoids from Oman. Paläontologische Zeitschrift 81: 399–405. Crossref
Webster, G.D., Angiolini, L., and Tintori, A. 2009a. Permian crinoids from the Saiwan and Khuff formations, southeastern Oman. Rivista Italiana di Paleontologia e Stratigrafia 115: 27–48.
Webster, G.D. and Webster, D.W. 2014. Bibliography and Index of Paleozoic Crinoids, Coronates, and Hemistreptocrinids, 1758–2012 [available online, http://crinoids.azurewebsites.net]
Webster, G.D., Heward, A.P., and Ausich, W.I. 2022. First crinoid crown from the Permian Khuff Formation (Wordian) of Oman. Proceedings of the Geologists’ Association 133: 154–161. Crossref
Webster, G.D., Haggart, J.W., Saxifrage, C., Saxifrage, B., Gronau, C., and Douglas, A. 2009b. Globally significant early Permian crinoids from the Mount Mark Formation in Strathcona Provincial Park, Vancouver Island, British Columbia-preliminary analysis of a disappearing fauna. Canadian Journal of Earth Science 46: 663–674. Crossref
Webster, G.D., Waters, J.A., and Chen, X.Q. 2009c. Revision of the Chen and Yao Devonian to Permian crinoids from WesternYunnan. Palaeobiodiversity and Palaeoenvironments 89: 119–160. Crossref
Weller, J.M. 1930. A group of larviform crinoids from lower Pennsylvanian strata of the Eastern Interior Basin. Ill. Geological Survey Report of Investigations 21: 1–42. Crossref
Willink, R.J. 1978. Catillocrinids from the Permian of eastern Australia. Alcheringa 2: 83–102. Crossref
Willink, R.J. 1979a. Some conservative and some highly-evolved Permian crinoids from eastern Australia. Alcheringa 3: 117–134. Crossref
Willink, R.J. 1979b. The crinoid genera Tribrachyocrinus McCoy, Calcelispongia Etheridge, Jimbacrinus Teichert and Meganotocrinus n. gen. in the Permian of eastern Australia. Palaeontographica, Abteilung A 165: 137–194.
Willink, R.J. 1980. A new coiled-stemmed camerate crinoid from the Permian of eastern Australia. Journal of Paleontology 54: 15–34.
Wright, D.F. 2017a. Bayesian estimation of fossil phylogenies and the evolution of early to middle Paleozoic crinoids (Echinodermata). Journal of Paleontology 91: 799–814. Crossref
Wright, D.F. 2017b. Phenotypic innovation and adaptive constraints in the evolutionary radiation of Palaeozoic crinoids. Scientific Reports 7: 13745. Crossref
Wright, D.F., Ausich, W.I., Cole, S.R., Rhenberg, E.C., and Peter, M.E. 2017. Phylogenetic taxonomy and classification of the Crinoidea (Echinodermata). Journal of Paleontology 91: 829–846. Crossref
Wright, J. 1946. New species of Taxocrinus and Synbathocrinus and other rare crinoids from the Carboniferous limestone of Coplow Knoll, Clitheroe. Geological Magazine 83: 121–128. Crossref
Wright, J. 1942. New British Carboniferous crinoids. Geological Magazine 79: 269–283. Crossref
Wright, J. 1954. The British Carboniferous Crinoidea. Palaeontographical Society, Monograph, 1 (5), 149–190; 2 (1), 191–254. Crossref
Yakovlev, N.N. 1926. Echinoderm fauna from the Permo-Carboniferous of Krasnoufimsk in the Urals, I [in Russian]. Izvestiâ Geologičeskogo Komiteta 45: 51–57.
Yakovlev, N.N. 1927. Echinoderm fauna from the Permo-Carboniferous of Krasnoufimsk in the Urals, II [in Russian]. Izvestiâ Geologičeskogo Komiteta 46: 181–192.
Yakovlev, N.N. 1930. Echinoderm fauna from the Permo-Carboniferous of the Urals at Krasnooufimsk, III [in Russian]. Izvestiâ Geologičeskogo Komiteta 46: 1019–1027.
Yakovlev, N.N. 1934. Crinoidi permiani di Sicilia. Palaeontographica Italica 34: 26–283.
Yakovlev, N.N. 1948. New Permian crinoids from northern Timan [in Russian]. Izvestiâ Akademii Nauk USSR, Seriâ Biologicheskaâ Izvestiâ 1: 119–122.
Yakovlev, N.N. and Ivanov, A.P. 1956. Marine crinoids and blastoids of the Carboniferous and Permian deposits of Russia [in Russian]. Trudy Vsesoûznogo Naučno-issledoavatel’skogo Geologičeskogo Instituta 11: 1–142.
Zittel, K.A. von 1895. Grundzüge der Palaeontologie (Palaeozoologie). 1st Edition. 971 pp. R. Oldenbourg, München.
Acta Palaeontol. Pol. 69 (3): 447–466, 2024
https://doi.org/10.4202/app.01159.2024