
A large frigatebird-like tarsometatarsus from the London Clay of Walton-on-the-Naze may shed light on the affinities of a poorly known early Eocene seabird taxon
GERALD MAYR and ANDREW C. KITCHENER
Mayr, G. and Kitchener, A.C. 2024. A large frigatebird-like tarsometatarsus from the London Clay of Walton-on-the-Naze may shed light on the affinities of a poorly known early Eocene seabird taxon. Acta Palaeontologica Polonica 69 (3): 523–528.
We report a tarsometatarsus and an associated pedal phalanx from the lower Eocene London Clay of Walton-on-the-Naze (Essex, UK). The specimen resembles the tarsometatarsus of the taxon Limnofregata (Fregatidae), but it belongs to a species that is distinctly larger than any other, extinct or extant, frigatebird, from which it also differs in some morphological features. Because of a close stratigraphical and geographical provenance, as well as a similar large size and frigatebird-like morphology, we consider it possible that the fossil belongs to Marinavis longirostris. This large seabird was initially described from the London Clay of Abbey Wood and is based on fragments of the rostrum, which likewise show a resemblance to the Fregatidae. If correctly assigned to the Fregatidae, the fossils would be among the earliest records of frigatebirds and the first fossils of this group of birds from the Paleogene of Europe, but we note that our tentative classification is still afflicted with considerable uncertainty.
Key words: Aves, Fregatidae, Marinavis longirostris evolution, taxonomy.
Gerald Mayr [Gerald.Mayr@senckenberg.de; ORCID: https://orcid.org/0000-0001-9808-748X ], Ornithological Section, Senckenberg Research Institute and Natural History Museum Frankfurt, Senckenberganlage 25, 60325 Frankfurt am Main, Germany.
Andrew C. Kitchener [a.kitchener@nms.ac.uk; ORCID: https://orcid.org/0000-0003-2594-0827] , Department of Natural Sciences, National Museums Scotland, Chambers Street, Edinburgh EH1 1JF, UK and School of Geosciences, University of Edinburgh, Drummond Street, Edinburgh EH8 9XP, UK.
Received 2 May 2024, accepted 19 July 2024, published online 30 September 2024.
Copyright © 2024 G. Mayr and A.C. Kitchener. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Almost half a century ago Harrison and Walker (1977) described a large bird from the Blackheath Member of the London Clay at Abbey Wood (Kent, UK) as Marinavis longirostris. The holotype consists of fragments of a dentary and maxilla, and a referred premaxilla probably belongs to the same individual (Fig. 1B, C; Harrison and Walker 1977: 6). Harrison and Walker (1977) also tentatively assigned a fragmentary carpometacarpus from Abbey Wood to M. longirostris (Fig. 1E), but as detailed by Mayr (2022), this unusually long and narrow bone, which was not found in association with the aforementioned remains, is more likely to be from a species of the Pelagornithidae (bony-toothed birds; Fig. 1F).
Marinavis longirostris had a long bill with a decurved tip, similar to that of the Procellariiformes and some Suliformes (Fregatidae and Phalacrocoracidae). Despite the limited known material, Harrison and Walker (1997) assigned the species to a new family-level taxon Marinavidae, which they classified into the Procellariiformes. However, the material does not allow an unambiguous phylogenetic placement, and the authors also noted similarities to “pelecaniform” birds. Owing to the highly fragmentary nature of the holotype, M. longirostris was largely ignored by subsequent authors, but the fossils certainly represent a large seabird that is distinct from the Pelagornithidae (as evidenced by the lack of pseudoteeth along the tomia of the bill).
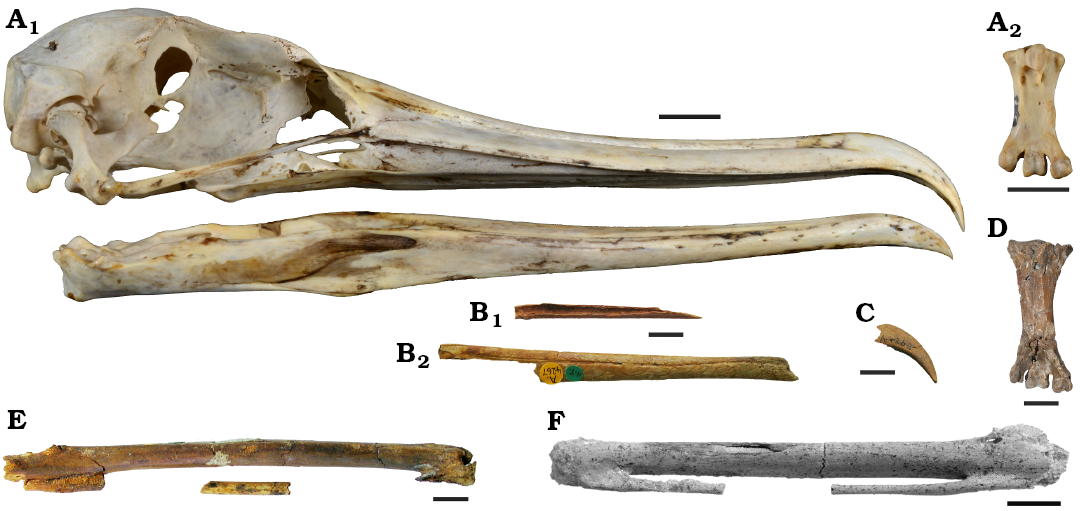
Fig. 1. Comparison of fossils from the London Clay with extant Fregatidae and the extinct Pelagornithidae. A. The extant Fregata magnificens Mathews, 1914 (SMF 5389). A1, skull in right lateral view; A2, tarsometatarsus in plantar view. B, C, E. Marinavis longirostris Harrison & Walker, 1977, from Abbey Wood, Kent, UK; Blackheath Member of the London Clay Formation, lower Eocene. These specimens were described by Harrison and Walker (1977) and are here illustrated to show a size correspondence to the tarsometatarsus from Walton-on-the-Naze. B. NHMUK A 4267, fragments of maxilla (B1) in ventral view and mandible (B2) in right lateral view. C. NHMUK A 4268, premaxilla in right lateral view. E. NHMUK A 4233, partial left carpometacarpus in ventral view; this specimen is here considered to be from a representative of the Pelagornithidae. D. aff. Marinavis longirostris (NMS.Z.2021.40.183) from Walton-on-the-Naze, Essex, UK, Walton Member of the London Clay Formation, lower Eocene; left tarsometatarsus in plantar view. F. The pelagornithid Lutetodontopteryx tethyensis Mayr and Zvonok, 2012 (SMF Av 539) from the middle Eocene of Ukraine; partial right carpometacarpus in ventral view. Scale bars 10 mm.
Here we report a tarsometatarsus and an associated pedal phalanx from the London Clay of Walton-on-the-Naze (Essex, UK), which may belong to Marinavis. The tarsometatarsus shows a resemblance to that of the Fregatidae (frigatebirds), which is notable, because the bill of M. longistrostris is likewise similar to that of frigatebirds in its proportions and the presence of a markedly hooked premaxilla (Fig. 1A1). In their relative size the new tarsometatarsus and the bill fragments of M. longirostris correspond to the bill and tarsometatarsus of extant Fregatidae (Fig. 1A‒D), but these bones belong to a species, which was distinctly larger than any extant frigatebird.
Institutional abbreviations.—LACM, Natural History Museum of Los Angeles County, USA; NHMUK; Natural History Museum, London, UK; NMS, National Museums Scotland, Edinburgh, UK; SMF, Senckenberg Research Institute Frankfurt, Germany; USNM, National Museum of Natural History, Smithsonian Institution, Washington D.C., USA.
Systematic palaeontology
Aves Linnaeus, 1758
?Fregatidae Degland & Gerbe, 1867
Genus aff. Marinavis Harrison & Walker, 1977
aff. Marinavis longirostris Harrison & Walker, 1977
Type species: Marinavis longisristris Harrison & Walker, 1977; Abbey Wood (Kent, UK); Blackheath Member of the London Clay Formation, early Eocene.
Figs. 1, 2.
Material.—NMS.Z.2021.40.183 left tarsometatarsus, pedal phalanx (Fig. 2A); collected in 1977 by Michael Daniels (original collector’s number WN77189) from Walton-on-the-Naze, Essex, UK; Walton Member of the London Clay Formation (previously Division A2; Jolley 1996; Rayner et al. 2009; Aldiss 2012), lower Eocene (lower Ypresian, 54.6‒55 Ma; Collinson et al. 2016).
Measurements (in mm).—Tarsometatarsus: maximum length, 44.4; proximal width, 19.0; distal width, 18.3; pedal phalanx: length, 31.1.
Description.—The tarsometatarsus of NMS.Z.2021.40.183 (Fig. 2A) is characterised by its extreme stockiness and the distal projection of the trochlea metatarsi II. With regard to these features it corresponds to the tarsometatarsus of the Fregatidae (Fig. 2C) and Plotopteridae (Fig. 2D), which are Paleogene foot-propelled diving birds of the North Pacific (e.g., Olson and Hasegawa 1996; Mayr and Goedert 2016, 2022).
In particular, the tarsometatarsus from Walton-on-the-Naze closely resembles that of the stem group frigatebird Limnofregata Olson, 1977, from the lower Eocene of North America (Fig. 2B; Olson 1977; Olson and Matsuoka 2005; Stidham 2015). However, NMS.Z.2021.40.183 is almost twice as long as the tarsometatarsus of Limnofregata azygosternon Olson, 1977 (tarsometatarsus length 44.4 mm versus 26.7‒27.3 mm in L. azygosternon; Olson 1977); the tarsometatarsi of the somewhat larger Limnofregata hasegawai Olson & Matsuoka, 2005, and Limnofregata hutchinsoni Stidham, 2015, are unknown). Extant Fregatidae have an even shorter tarsometatarsus, and this bone has a length of only 22.0 mm in Fregata magnificens Mathews, 1914, one of the largest extant species (identical measurements were obtained from two individuals in the collection of SMF). Furthermore, the tarsometatarsus of extant Fregatidae is stouter and has a wider shaft.
The hypotarsus of NMS.Z.2021.40.183 bears a single wide sulcus, presumably for the tendon of musculus flexor digitorum longus, which is medially bordered by a prominent crista medialis. In extant Fregatidae this sulcus is closed to form a canal, whereas there are two sulci, for the tendons of m. flexor digitorum longus and m. hallucis longus, in L. azygosternon (Olson 1977: fig. 28). In the Plotopteridae there are also two hypotarsal sulci (Fig. 2D3). Smith (2010: 27) noted that suliform birds are characterised by the fact that the cotyla medialis is significantly larger than the cotyla lateralis. The cotylae are poorly preserved in the fossil and the exact size and shape of the cotyla lateralis cannot be determined.
Only the lateral foramen vasculare proximale is clearly visible in NMS.Z.2021.40.183; the area of the medial foramen is damaged. The tuberositas musculi tibialis cranialis has a subovate shape and is medially situated. The plantar surface of the shaft lacks a sulcus flexorius. Owing to damage to the specimen, the foramen vasculare distale is not clearly discernible. On the dorsal surface of the tarsometatarsus, there is no well-defined sulcus extensorius that extends proximal to the foramen vasculare distale. This sulcus is typically present in the species of Suliformes (Fig. 2C1 and Smith 2010: fig. 8), but it is also weakly developed in species of Limnofregata (Fig. 2B1). The fossa metatarsi I is located near the medial margin of the shaft and is proximodistally extensive.
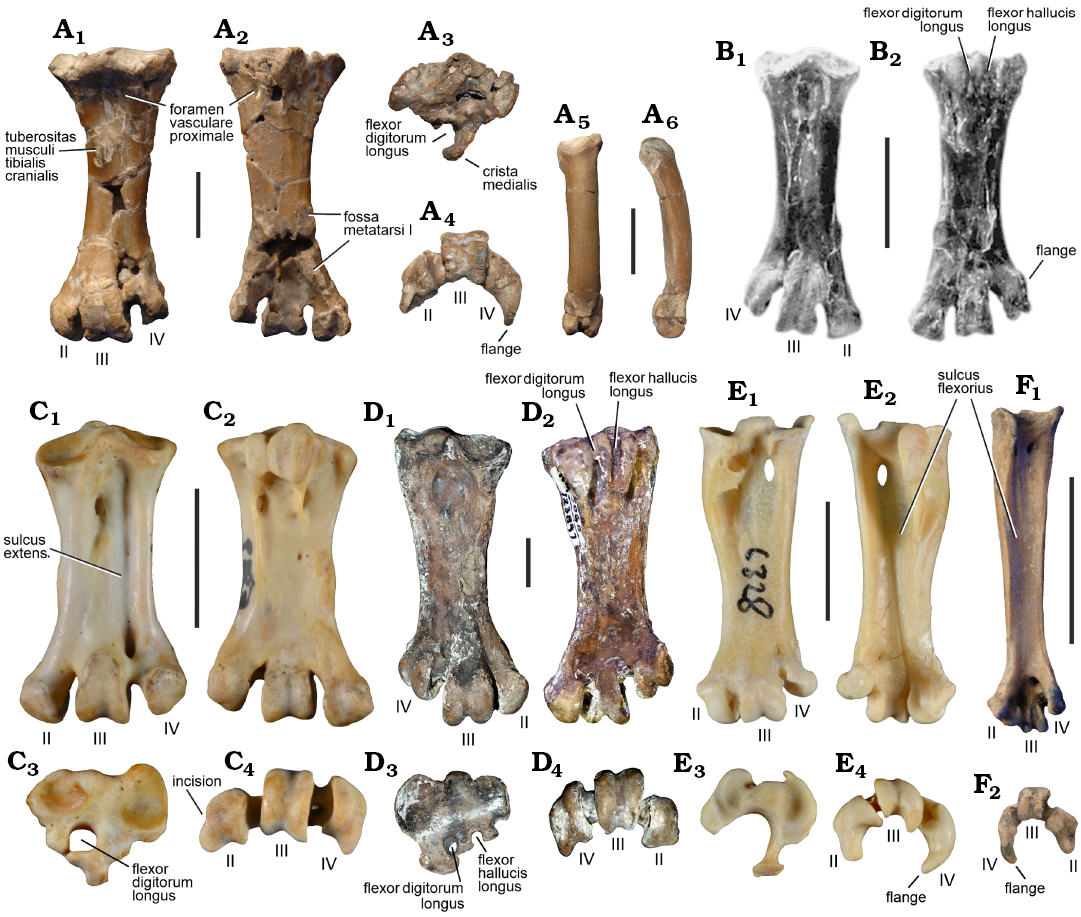
Fig. 2. Comparison of the fossil tarsometatarsus with that of various extant and extinct birds. A. aff. Marinavis longirostris Harrison & Walker, 1977 (NMS.Z.2021.40.183) from Walton-on-the-Naze, Essex, UK, Walton Member of the London Clay Formation, lower Eocene; left tarsometatarsus in dorsal (A1), plantar (A2), proximal (A3), and distal (A4) views, phalanx of the hallux in dorsal (A5) and lateral (A6) views. B. Limofregata azygosternon Olson, 1977 (USNM 22753) from the lower Eocene Green River Formation, Wyoming, USA; right tarsometatarsus in dorsal (B1) and plantar (B2) views (from Olson 1977: fig. 28). C. The extant fregatid Fregata magnificens Mathews, 1914 (SMF 5389); right tarsometatarsus (mirrored) in dorsal (C1), plantar (C2), proximal (C3), and distal (C4) views. D. The plotopterid Phocavis maritimus Goedert, 1988 (holotype, LACM 123897) from the upper Eocene of Oregon, USA; right tarsometatarsus in dorsal (D1), plantar (D2), proximal (D3), and distal (D4) views. E. The extant strigiform Surnia ulula (Linnaeus, 1758) (SMF 6327, the number written on the specimen is incorrect); left tarsometatarsus in dorsal (E1), plantar (E2), proximal (E3), and distal (E4) views. F. The strigiform Ypresiglaux michaeldanielsi Mayr & Kitchener, 2023 (holotype, NMS.Z.2021.40.26) from Walton-on-the-Naze, Essex, UK, Walton Member of the London Clay Formation, lower Eocene; right tarsometatarsus in plantar (F1) and distal (F2) views. The tarsometatarsal trochleae are denoted by Roman numerals. Scale bars 10 mm.
The distal end of the tarsometatarsus of NMS.Z.2021.40.183 is distinguished from that of extant Fregatidae in that the trochleae are arranged on a curve (in distal view of the bone). The condition in Limnofregata is difficult to assess, because the known specimen with preserved tarsometatarsi are flattened skeletons on slabs. Both incisurae intertrochleares of NMS.Z.2021.40.18 are wide. As in the Fregatidae and most other extant Suliformes, the trochlea metatarsi II projects far distally and exceeds the trochlea metatarsi III in length; owing to damage to its lateroplantar portion, this trochlea appears mediolaterally narrower than it probably was originally. The fossils lacks a medial incision in the trochlea metatarsi II, which was identified as an apomorphy of the Pelecanidae and Suliformes by Smith (2010); this incision is present in a tarsometatarsus of Fregata minor (Gmelin, 1789) shown by Smith (2010: fig. 9G), but it is weakly developed in two specimens of F. magnificens examined by us (SMF 9572 and SMF 5389; Fig. 2C4). The trochlea metatarsi III is proportionally somewhat wider than that of Limnofregata, its plantar articular surface is damaged but does not seem to have tapered proximally. Unlike in extant Fregatidae, the trochlea metatarsi IV bears a pronounced plantar flange, and with regard to this feature the specimen differs from all extant representatives of the Suliformes. The known tarsometatarsi of Limnofregata are from compression fossils on slabs, so that the morphology of the distal end is affected by diagenetic flattening, but the published images suggest a flange to be present (Fig. 2B2). In the Plotopteridae, the distal margin of the trochlea metatarsi IV is more laterally slanted (Fig. 2D2).
NMS.Z.2021.40.183 also includes a long pedal phalanx (Fig. 2A5, A6), which is here identified as the phalanx of the hallux. The bone is curved in lateral view and is stouter than the corresponding phalanx of Limnofregata.
Concluding remarks
The tarsometatarsus of NMS.Z.2021.40.183 (Fig. 2A1‒A4) shows a superficial similarity to that of some stout-legged extant Strigidae (true owls), such as Surnia ulula (Linnaeus, 1758) (Fig. 2E). However, the hypotarsal sulcus is narrower than in strigiform birds, the plantar surface of the shaft lacks a distinct sulcus flexorius, the trochlea metatarsi II is proportionally smaller, the wing-like flange of the trochlea metatarsi IV is much less developed, and the phalanx of the hallux is proportionally longer. Moreover, the bone differs markedly from the tarsometatarsus of early Paleogene Strigiformes, in which the trochlea metatarsi II is either proportionally wider (Berruornis, Primoptynx, Ogygoptynx) or does not reach farther distally than the trochlea metatarsi III (Minerva, Ypresiglaux; Fig. 2F) (Mourer-Chauviré 1994; Mayr et al. 2020; Mayr and Kitchener 2023).
Derived characteristics shared by NMS.Z.2021.40.183 and species of the Fregatidae include the extreme stockiness of the tarsometatarsus and the distal projection of the trochlea metatarsi II, but we could not identify unambiguous tarsometatarsal synapomorphies that are unique to the fossil and frigatebirds. A distally projected trochlea metatarsi II, which exceeds the trochlea metatarsi III in length, was identified as an apomorphy of the Suliformes (Fregatidae, Sulidae, Phalacrocoracidae, and Anhingidae) by Smith (2010). The only other tarsometatarsal character listed as a suliform apomorphy by Smith (2010), a more expansive cotyla medialis relative to the cotyla lateralis, cannot be unambiguously assessed in the fossil, in which the cotylae of the proximal end of the bone are damaged. The single tarsometatarsus feature listed as a synapomorphy of Limnofregata and extant Fregatidae by Smith (2010), a low eminentia intercotylaris, is present in NMS.Z.2021.40.183, but shows much homoplasy in birds.
Within Suliformes, an equally stocky tarsometatarsus also occurs in the Plotopteridae and Anhingidae (darters). In the Anhingidae the trochlea metatarsi II exhibits a pronounced medial incision, which is absent in the fossil (as detailed above, we could not confirm Smith’s (2010) observation that this incision is present in all Fregatidae), the trochlea metatarsi IV is much wider than in the fossil, and the eminentia intercotylaris is more prominent. Plotopterids occurred in the North Pacific and likewise have a wider trochlea metatarsi IV with a more laterally slanted distal margin. In both, Anhingidae and Plotopteridae, the hypotarsus forms a marked sulcus for the tendon of musculus flexor hallucis longus.
Overall, the tarsometatarsus from Walton-on-the-Naze most closely resembles that of the early Eocene stem group frigatebird Limnofregata (Fig. 2B), but as in crown group Fregatidae NMS.Z.2021.40.183 has an unusually large hypotarsal sulcus/canal for the tendon of musculus flexor digitorum longus, whereas Limnofregata exhibits the plesiomorphic condition found in other Suliformes, in which there are two hypotarsal sulci or canals for the deep flexor tendons of the toes (Olson 1977; Mayr 2016b). Even though some distinct differences between NMS.Z.2021.40.183 and extant Fregatidae exist (in particular, concerning the arrangement of the trochleae on a curve in distal view of the tarsometatarsus and the marked plantar flange formed by the trochlea metatarsi IV), we were unable to identify character evidence for a more compelling alternative placement of the fossil. However, our tentative classification of NMS.Z.2021.40.183 is afflicted with uncertainty and we caution against using this fossil as a definitive record of the Fregatidae, which has to be corroborated or refuted through the discovery of further fossil material.
NMS.Z.2021.40.183 would be the largest known species of frigatebird and the first Paleogene record from the Old World. With an age of about 54.6 to 55 million years, NMS.Z.2021.40.183, as well as the holotype of Marinavis longirostris from the Blackheath Member of the London Clay, would also be among the oldest records of frigatebirds. Limnofregata hutchinsoni from the Wasatch Formation of Wyoming, USA, which is older than 53 Ma and likely also older than 54 Ma (Stidham 2015), is potentially of comparable age to the specimen from Walton-on-the-Naze. A tentative record of a frigatebird from the Nanjemoy Formation in Virginia, USA, has an age of 53.6 to 52.8 Ma (Mayr 2016a), and the records of Limofregata azygosternon and L. hasegawai from the Green River Formation are even younger, dating to about 48 to 52 million years (Stidham 2015).
Unlike the Limnofregata fossils, which were found in fossil sites of lacustrine origin, NMS.Z.2021.40.183 stems from sediments that were deposited in a marine environment; a tentatively referred carpometacarpus from the North American Nanjemoy Formation likewise stems from marine sediments (Mayr 2016a). If correctly assigned to the Fregatidae, these fossils would suggest a broader habitat preference of Eocene frigatebirds, which was also considered by Stidham (2015). The curvature of the tarsometatarsal trochleae and the plantar flange of the trochlea metatarsi IV indicate that, unlike in extant frigatebirds, the feet of the species from Walton-on-the-Naze had grasping capabilities, even though the absence of a sulcus flexorius along the plantar surface of the tarsometatarsus shaft shows that these were not as strongly developed as in, e.g., strigiform birds. Therefore, the differences in tarsometatarsus morphology to extant Fregatidae suggest that the species from Walton-on-the-Naze was less aerial and, if indeed related to frigatebirds, possibly used its legs to catch prey (presumably fish) from the sea surface.
Owing to the lack of overlap in the known skeletal elements, meaningful morphological comparisons between the previously described material of Marinavis longirostris and the new fossils cannot be performed. However, and as far as comparisons are possible due to the fragmentary nature of the holotype, the shape of the upper bill of M. longirostris matches that of extant Fregatidae (compare Fig. 1A1 and 1B, C). The Blackheath Member of the London Clay has an age of about 55 Ma (Hooker 2010) and therefore corresponds stratigraphically to the Walton Member of the London Clay (Rayner et al. 2009). It is this stratigraphically and geographically close provenance of the new specimen and the M. longirostris holotype, as well as their similar large size and frigatebird-like morphology, which suggest affinities of the fossil from Walton-on-the-Naze to M. longirostris.
If our assignment of NMS.Z.2021.40.183 to Marinavis is correct, the poorly diagnosed taxon Marinavidae Harrison & Walker, 1977, would be a junior synonym of Fregatidae Degland & Gerbe, 1867. However, owing to the limited material available from both Abbey Wood, the type locality of M. longirostris, and Walton-on-the-Naze, we refrain from a definitive taxonomic action. Future specimens will hopefully allow an unambiguous classification of Marinavis and a robust assessment of the affinities between this taxon and the fossil from Walton-the-Naze.
Acknowledgements
We thank Sven Tränkner (Senckenberg Research Institute and Natural History Museum, Frankfurt am Main, Germany) for taking some of the photographs (additional images are by GM). Comments from Zbigniew Bochenski (Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Kraków, Poland) and an anonymous reviewer improved the manuscript
References
Aldiss, D.T. 2012. The stratigraphical framework for the Palaeogene successions of the London Basin, UK. British Geological Survey Open Report OR/12/004: 1–87.
Collinson, M.E., Adams, N.F., Manchester, S.R., Stull, G.W., Herrera, F., Smith, S.Y., Andrew, M.J., Kenrick, P., and Sykes, D. 2016. X-ray micro-computed tomography (micro-CT) of pyrite-permineralized fruits and seeds from the London Clay Formation (Ypresian) conserved in silicone oil: a critical evaluation. Botany 94: 697–711. Crossref
Harrison, C.J.O. and Walker, C.A. 1977. Birds of the British Lower Eocene. Tertiary Research Special Paper 3: 1–52. Crossref
Hooker, J.J. 2010. The mammal faunas of the early Eocene Blackheath Formation of Abbey Wood, London. Monograph of the Palaeontographical Society, London 164: 1–162. Crossref
Jolley, D.W. 1996. The earliest Eocene sediments of eastern England: an ultra-high resolution palynological correlation. Geological Society of London, Special Publications 101: 219–254. Crossref
Mayr, G. 2016a. The world’s smallest owl, the earliest unambiguous charadriiform bird, and other avian remains from the early Eocene Nanjemoy Formation of Virginia (USA). Paläontologische Zeitschrift 90: 747–763. Crossref
Mayr, G. 2016b. Variations in the hypotarsus morphology of birds and their evolutionary significance. Acta Zoologica 97: 196–210. Crossref
Mayr, G. 2022. Paleogene Fossil Birds, Second Edition. 239 pp. Springer, Cham. Crossref
Mayr, G. and Goedert, J.L. 2016. New late Eocene and Oligocene remains of the flightless, penguin-like plotopterids (Aves, Plotopteridae) from western Washington State, U.S.A. Journal of Vertebrate Paleontology 36: e1163573. Crossref
Mayr, G. and Goedert, J.L. 2022. New late Eocene and Oligocene plotopterid fossils from Washington State (USA), with a revision of “Tonsala” buchanani (Aves, Plotopteridae). Journal of Paleontology 96: 224–236. Crossref
Mayr, G. and Kitchener, A.C. 2023. Early Eocene fossil illuminates the ancestral (diurnal) ecomorphology of owls and documents a mosaic evolution in the strigiform stem lineage. Ibis 165: 231–247. Crossref
Mayr, G., Gingerich, P.D., and Smith, T. 2020. Skeleton of a new owl from the early Eocene of North America (Aves, Strigiformes) with an accipitrid-like foot morphology. Journal of Vertebrate Paleontology 40: e1769116. Crossref
Mourer-Chauviré, C. 1994. A large owl from the Palaeocene of France. Paleontology 37: 339–348.
Olson, S.L. 1977. A Lower Eocene frigatebird from the Green River Formation of Wyoming (Pelecaniformes: Fregatidae). Smithsonian Contributions to Paleobiology 35: 1–33. Crossref
Olson, S.L. and Hasegawa, Y. 1996. A new genus and two new species of gigantic Plotopteridae from Japan (Aves: Plotopteridae). Journal of Vertebrate Paleontology 16: 742–751. Crossref
Olson, S.L., and Matsuoka, H. 2005. New specimens of the early Eocene frigatebird Limnofregata (Pelecaniformes: Fregatidae), with the description of a new species. Zootaxa 1046: 1‒15. Crossref
Rayner, D., Mitchell, T., Rayner, M., and Clouter, F. 2009. London Clay Fossils of Kent and Essex. 228 pp. Medway Fossil and Mineral Society, Rochester, Kent.
Smith, N.D. 2010. Phylogenetic analysis of Pelecaniformes (Aves) based on osteological data: Implications for waterbird phylogeny and fossil calibration studies. PLoS ONE 5 (10): e13354. Crossref
Stidham, T.A. 2015. A new species of Limnofregata (Pelecaniformes: Fregatidae) from the Early Eocene Wasatch Formation of Wyoming: implications for palaeoecology and palaeobiology. Palaeontology 58: 239–249. Crossref
Acta Palaeontol. Pol. 69 (3): 523–528, 2024
https://doi.org/10.4202/app.01169.2024