
A new proterochampsid archosauriform from the Middle–Upper Triassic of Southern Brazil
RODRIGO T. MÜLLER
Müller, R.T. 2025. A new proterochampsid archosauriform from the Middle–Upper Triassic of Southern Brazil. Acta Palaeontologica Polonica 70 (1): 7–16.
Proterochampsidae is a clade of carnivorous archosauriforms that lived during the Triassic and is characterized by an elongated rostrum with dorsally oriented external nares, and a unique pes configuration. Although the majority of proterochampsids are found in South America, recent phylogenetic studies suggest a broader geographical distribution for this clade. Most proterochampsids are known from cranial remains; if postcranial elements are known they are frequently poorly preserved. This study describes a new proterochampsid, Retymaijychampsa beckerorum gen. et sp. nov., from the Middle–Late Triassic, based on an almost complete and articulated hindlimb. The holotype was excavated at the Linha Várzea 2 (Becker) Site, located in the municipality of Paraíso do Sul, Rio Grande do Sul, Brazil. The fossils from this locality places it within the Dinodontosaurus Assemblage Zone, which is Ladinian to early Carnian in age according to biostratigraphic investigations. The new taxon is the second proterochampsid described from the Pinheiros-Chiniquá Sequence and represents one of the oldest known members of the clade worldwide. An interesting aspect of R. beckerorum gen. et sp. nov. is its phylogenetic position because it is more closely related to Proterochampsa than to rhadinosuchine proterochampsids. Therefore, the discovery of the new proterochampsid partially fills a long ghost lineage extending from the Middle Triassic to the early Late Triassic. Furthermore, the presence of two proterochampsid species within the Pinheiros-Chiniquá Sequence deposits supports the hypothesis of significant ecomorphological diversity among proterochampsians in Triassic environments.
Key words: Archosauromorpha, Proterochampsia, Proterochampsidae, phylogeny, Carnian, Ladinian, South America, Brazil.
Rodrigo T. Müller [rodrigotmuller@hotmail.com; ORCID: https://orcid.org/0000-0001-8894-9875 ], Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia, Universidade Federal de Santa Maria, São João do Polêsine, RS 598, 97230-000, Brazil.
Received 29 August 2024, accepted 2 November 2024, published online 25 February 2025.
Copyright © 2025 R.T. Müller. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Triassic Period witnessed the emergence and extinction of numerous groups of reptiles (Benton 2016), including the Proterochampsidae (Trotteyn et al. 2013). Although this clade was previously considered endemic to South America (Ezcurra 2010; Trotteyn et al. 2013), recent phylogenetic investigations indicate a broader geographical distribution (Ezcurra and Sues 2021; Paes-Neto et al. 2024). The oldest known proterochampsids have been found in the Ladinian/lower Carnian deposits of Argentina and Brazil (Romer 1971; Arcucci 1990; Hsiou et al. 2002; Trotteyn and Ezcurra 2020; Paes-Neto et al. 2024), whereas the most recent records come from the upper Carnian/lower Norian in the same region (Reig 1959; Barberena 1982). Phylogenetic analyses place Proterochampsidae within Proterochampsia, a clade of archosauriforms that is the sister-group to Archosauria (Ezcurra 2016; Ezcurra and Sues 2021; Müller et al. 2022; Paes-Neto et al. 2024). Because of its affinities, the clade is particularly important for understanding the early evolution of archosaurs. In addition, proterochampsids evolved a relatively diverse array of body plans, with some forms exhibiting putative semi-aquatic adaptations, whereas other were adapted to a terrestrial life style (Reig 1959; Sill 1967; Arcucci et al. 2019; Ezcurra et al. 2021; García Marsa et al. 2023). The diversity within the group includes gracile forms, such as Tropidosuchus romeri Arcucci, 1990, as well as species with flat skulls adorned with protuberances (Reig 1959; Barberena 1982). Proterochampsids are characterized by an elongated skull, particularly a long rostrum, dorsally oriented external nares, and a unique configuration of the pes, which is notably asymmetrical (Trotteyn et al. 2013).
Most proterochampsids are known exclusively from cranial remains (Price 1946; Reig 1959; Barberena 1982; Paes-Neto et al. 2024). In addition, details of certain postcranial structures are often poorly preserved in specimens that include postcranial elements (Trotteyn 2011; Trotteyn and Ezcurra 2014). Therefore, new specimens that preserve such remains are essential for clarifying aspects of the anatomy, biology, and evolution of proterochampsids. Since the pelvic girdle and hindlimb play a crucial role in the phylogenetic context of Archosauria and related groups (Sereno 1991; Nesbitt 2011; Ezcurra 2016), these structures are especially important for investigating the evolution of these groups. The extensive diversity in the structure of the pelvic girdle and hindlimb among early archosaurs and related groups reflects a broad range of postures and ecological adaptations (Hutchinson and Gatesay 2000; Li et al. 2016; Müller et al. 2023; Agnolín et al. 2024). In the present study, a new proterochampsid from the Middle–Upper Triassic of Southern Brazil (Fig. 1) is described. The specimen consists of an almost complete and articulated hindlimb, making it one of the few specimens with these bones preserved from Brazil.
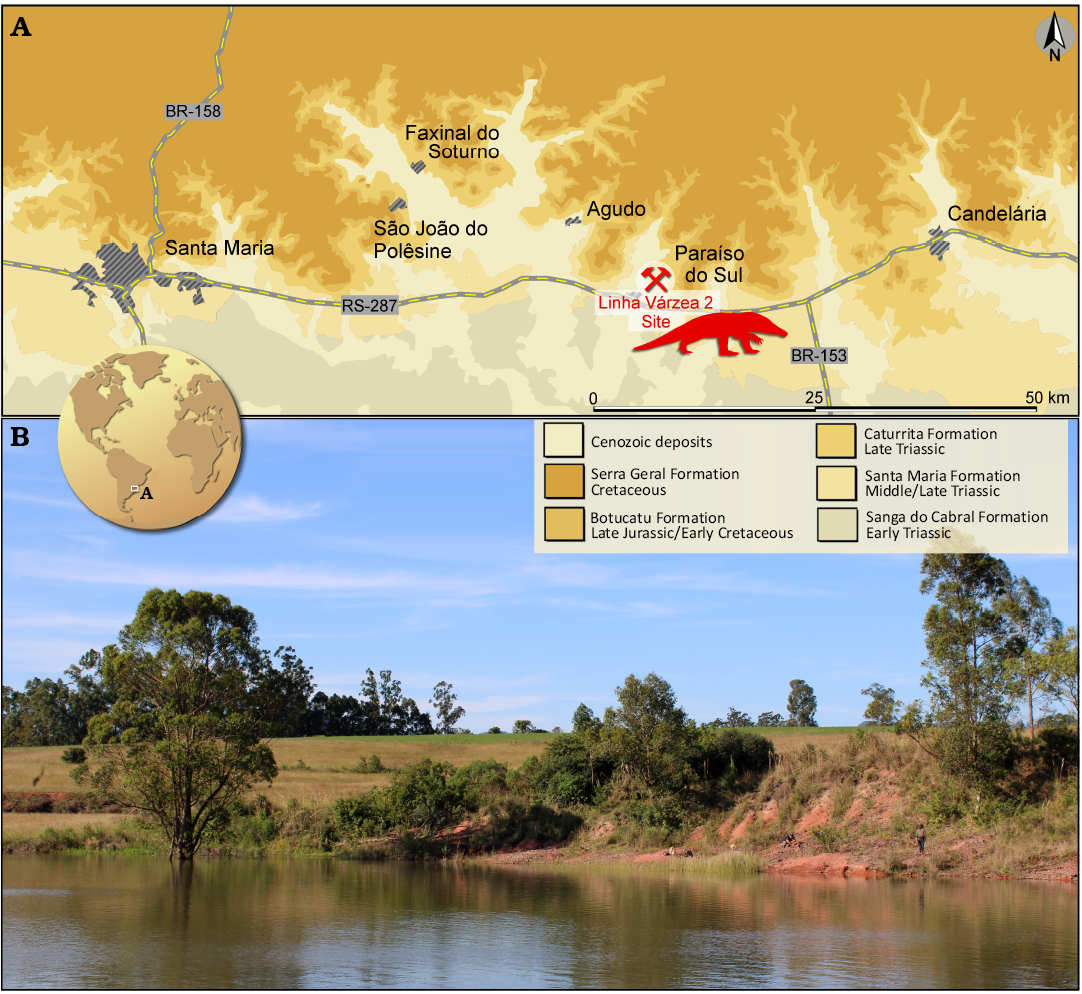
Fig. 1. Provenance of the proterochampsid archosauriform Retymaijychampsa beckerorum gen. et sp. nov. A. Location and geological context of the Linha Várzea 2 (Becker) Site, Paraíso do Sul, Rio Grande do Sul, Brazil; silhouette based on the artwork of Matheus Fernandes Gadelha (Limoeiro do Norte, Brazil). B. General view of the site (taken in August 2024).
Institutional abbreviations.—CAPPA/UFSM, Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia/Universidade Federal de Santa Maria, São João do Polêsine, Rio Grande do Sul, Brazil; MCP, Museu de Ciencias e Tecnología, Pontificia Universidade Catolica, Porto Alegre, Brazil; PVL, Paleontología de Vertebrados, Instituto Miguel Lillo, Tucumán, Argentina; PVSJ, División de Paleontología de Vertebrados del Museo de Ciencias Naturales y Universidad Nacional de San Juan, San Juan, Argentina; UFRGS-PV-T, Triassic Vertebrate Collection of the Museu de Paleontologia da UFRGS “Irajá Damiani Pinto,” Universidade Federal de Rio Grande do Sul, Porto Alegre, Brazil.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the International Code of Zoological Nomenclature. The LSID for this publication is: urn:lsid:zoobank.org:pub:FA6231E7-5E62-4221-AB07-E1B1C42BC107.
Material and methods
The phylogenetic affinities of the new proterochampsid were examined using a new data matrix modified from that published by Sengupta et al. (2024). The former data matrix was constructed to investigate a broader context of archosauromorphs (Ezcurra 2016). Therefore, to investigate the affinities of the new taxon within Proterochampsia, a reduced set of operational taxonomic units (OTUs) was selected, which includes all proterochampsians from the previous study. Additionally, Euparkeria capensis, Gnathovorax cabreirai, Buriolestes schultzi, Gracilisuchus stipanicicorum, Prestosuchus chiniquensis, and Diandongosuchus fuyuanensis were selected to compose the outgroup. Following a similar approach to that employed by Langer et al. (2022), only informative characters that vary within the present sample were selected from the original data matrix. A new character based on the ratio between the width of the proximal end of the tibia and the total length of the bone was included (character number 339). The final data matrix is composed of 401 morphological characters and 26 OTUs (see SOM, Supplementary Online Material available at http://app.pan.pl/SOM/app70-Muller_SOM.pdf).
The most parsimonious trees (MPTs) were reconstructed using equally weighted parsimony analysis with the software TNT v. 1.5 (Goloboff and Catalano 2016). Characters 1, 2, 7, 12, 13, 19–23, 30, 31, 34, 36, 41, 44, 46, 52, 57, 58, 62, 76, 90, 111, 116, 118, 126–130, 146, 151, 165, 167, 169, 181, 207, 216, 219, 222, 224, 227, 235, 240, 241, 243, 247, 249, 255, 260, 262, 268, 269, 278, 283, 288, 290, 291, 293, 294, 296, 300, 301, 306, 310, 312, 313, 315, 316, 325, 330, 338, 339, 343, 352, 359, 360, 364, 370, 373, 374, 385, 388, and 389 were set as ordered following previous iterations of the original data matrix (Ezcurra 2016; Ezcurra et al. 2017, 2020; Müller et al. 2023; Sengupta et al. 2024). Euparkeria capensis was used to root the MPTs, which were recovered using the random addition sequence + tree bisection reconnection (TBR), which included 1000 replicates of Wagner trees (with random seed = 0), TBR and branch-swapping (holding 10 trees saved per replicate). Nodes without synapomorphies were collapsed after the search. Decay indices (Bremer support values) and bootstrap values (1000 replicates) were obtained with TNT v. 1.5.
Systematic palaeontology
Archosauromorpha Huene, 1946 (sensu Dilkes 1998)
Archosauriformes Gauthier et al., 1988
Proterochampsidae Sill, 1967 (sensu Trotteyn 2011)
Clade Proterochampsinae nov.
Type genus: Proterochampsa Reig, 1959.
Definition.—Most inclusive clade including Proterochampsa barrionuevoi Reig, 1959, but not Cerritosaurus binsfeldi Price, 1946, nor Rhadinosuchus gracilis Huene, 1938.
Diagnosis.—Ratio between the maximum width of the skull and length of the presacral vertebral column 0.336–0.440; distinct coarse ornamentation on lateral surface of the surangular and angular; in medial view, the area ventral to the internal mandibular fenestra showing a larger contribution from the angular bone compared to the prearticular, or both bones contribute equally; prearticular with ventral margin posterior to its contact with the splenial straight or ventrally curved on the anterior half of the bone in medial or lateral view; and presence of ventral keel on ninth presacral vertebral centrum.
Genus Retymaijychampsa nov.
ZooBank LSID: urn:lsid:zoobank.org:act:99025E25-552E-47C4-BEC4- C2FCCCEADEEE
Type species: Retymaijychampsa beckerorum gen et sp. nov., monotypic, see below.
Etymology: The combination of Guarani retyma, leg and ijy, strong with Greek champsa, crocodile; referring to the robust constitution of the animal’s hindlimb.
Diagnosis.—Same as for the only known species.
Retymaijychampsa beckerorum sp. nov.
Fig. 3.
ZooBank LSID: urn:lsid:zoobank.org:act:80A4B7E7-6C18-4A81-B668- B623A9D416A4
Etymology: In honor of the family Becker, the owners of the property were the Linha Várzea 2 site is located.
Holotype: CAPPA/UFSM 0430, a complete and articulate right hindlimb.
Type locality: Linha Várzea 2 (= Becker) site (29°44′03″ S, 53°09′07″ W), Paraíso do Sul, Rio Grande do Sul, Brazil (Fig. 1).
Type horizon: Santa Maria Formation (Schultz et al. 2020); Pinheiros-Chiniquá Sequence (Horn et al. 2014) of the Santa Maria Supersequence (Zerfass et al. 2003), Paraná Basin. This third-order sequence is associated with the Dinodontosaurus Assemblage Zone, which is Ladinian to early Carnian in age (Ezcurra et al. 2017; Schultz et al. 2020; Novas et al. 2021; Müller and Garcia 2022).
Material.— Holotype only.
Diagnosis.—Retymaijychampsa beckerorum gen. et sp. nov. differs from all other known proterochampsids with comparable skeletal material based on the following local autapomorphies: absence of an extensor fossa on the anterior surface of the distal portion of the femur, resulting in a straight anterior margin in distal view; fibular condyle of the femur projects more ventrally than tibial condyle, forming an uneven distal surface in anterior view; and markedly robust tibia, with a ratio of 0.37 between the maximum width of the proximal end and the total length of the bone.
Description.—Although the femur lacks the proximal portion, it cleary had a sigmoid shape (Fig. 3A1). The preserved segment of the shaft includes the distal half of the fourth trochanter, which forms an elongated ridge (Fig. 3A6). The distal portion of the trochanter merges smoothly with the shaft. Whereas the femur is not completely preserved, it is evident that the fourth trochanter is distally elongated, similar to the condition seen in most proterochampsids (Trotteyn et al. 2013). Conversely, the fourth trochanter of Stenoscelida aurantiacus Müller et al., 2022 (CAPPA/UFSM 0293; Müller et al. 2022) and Tropidosuchus romeri Arcucci, 1990 (PVL 4601; Arcucci 1990) is short. The distal portion of the femur expands relative to the shaft and is arched posteriorly. The popliteal fossa is not elongated. Unlike other proterochampsids (Trotteyn 2011; Trotteyn and Ezcurra 2020; Müller et al. 2022; Paes-Neto et al. 2024), this specimen lacks an extensor fossa on the anterior surface of the distal portion of the femur, resulting in a straight anterior margin in distal view (Fig. 3A4, A8). In addition, compared to other proterochampsids, the fibular condyle projects more ventrally than the tibial condyle, forming an uneven distal surface in anterior view. The tibiofibular crest is not strongly expanded and there are no distinguishing features between the lateral surface of the tibiofibular crest and the lateral condyle.
The complete tibia (Table 1) is notably robust, with a ratio of 0.37 between the maximum width of the proximal end and the total length of the bone. For comparison, the ratio for the holotype of Stenoscelida aurantiacus (CAPPA/UFSM 0293) is 0.25 and for the holotype of Kuruxuchampsa dornellesi Paes-Neto et al., 2024 (UFRGS-PV-0877-T) is 0.26. In lateral/medial view, the proximal portion of the tibia is expanded both anteriorly and posteriorly relative to the shaft (Fig. 3A5). The anterior projection comprises the cnemial crest, which tapers to a point and is straight (Fig. 3A6). The posterior projection is the medial condyle, which is positioned further posteriorly relative to the lateral condyle. The proximal articular surface is flat. The shaft of the tibia is straight and expands distally, with the expansion predominantly oriented anteroposteriorly. This condition differs the specimen from Tropidosuchus romeri (PVL 4601; Arcucci 1990), in which the distal end of the tibia is not expanded. The posterior margin of the distal end of the tibia of CAPPA/UFSM 0430 is gently concave (Fig. 3A7).
Table 1. Measurements (in mm) of the right hindlimb of Retymaijychampsa beckerorum gen. et sp. nov. (CAPPA/UFSM 0430). * incomplete; ? not preserved or inaccessible.
|
Element |
Length |
Maximum |
Maximum |
|
femur |
66.7* |
? |
16.6 |
|
tibia |
70.5 |
26.3 |
16.5 |
|
fibula |
64.7 |
11.5 |
9.7 |
|
metatarsal I |
18.4 |
8.2 |
4.9 |
|
metatarsal II |
28.7 |
14.4 |
10.7 |
|
metatarsal III |
34.8 |
12.8 |
7.8 |
|
metatarsal IV |
43.7 |
? |
3.4 |
|
metatarsal V |
14.5 |
? |
2.9 |
The fibula is slender, with a shaft that has approximately half the transverse width of the tibial shaft (Fig. 3A6). The fibula is slightly shorter than the tibia (Table 1). The proximal end is expanded posteriorly relative to the shaft and is compressed transversely (Fig. 3A6). The shaft is straight along its length, differing from the arched shaft of the fibula in most pseudosuchians (Nesbitt 2011; Ezcurra 2016). The width of the shaft remains consistent along the entire length of the fibula, whereas in Stenoscelida aurantiacus (CAPPA/UFSM 0293; Müller et al. 2022) the proximal half of the shaft is wider. There is no tubercle or ridge for the attachment of the iliofibularis muscle, such as in Proterochampsa barrionuevoi Reig, 1959 (PVSJ 606; Trotteyn et al. 2013). The distal end expands relative to the shaft, and the distal articular surface is flat.
The astragalus is transversely expanded (Fig. 3A7). The dorsal margin of its medial surface is concave, while its ventral surface of the bone is convex anteroposteriorly. There is no anterior expansion at the anteromedial corner. The posterior margin lacks a dorsal process. The calcaneum is lateromedially shorter than the astragalus (Fig. 3A6, A7). The facet for the fibula is oriented dorsomedially. The calcaneal tuber is well-developed and perpendicular to the main body of the calcaneum (Fig. 3A6). There is a depression between the main body of the bone and the posterior surface of the calcaneal tuber (Fig. 3A6, A7).
The configuration of the metatarsals is consistent with that in other proterochampsids (Table 1): metatarsal I is short; metatarsal II is slightly longer than metatarsal I and the stoutest of the elements; metatarsal III is longer than metatarsal II and more slender; metatarsal IV is quite slender and as long as metatarsal III; and metatarsal V is reduced and tapers distally. Pedal digit I includes two phalanges (Fig. 3A5). The specimen differs from Tropidosuchus romeri (PVL 4601; Arcucci 1990) and Stenoscelida aurantiacus (CAPPA/UFSM 0293; Müller et al. 2022) in that both of the latter taxa have a shorter digit I. Whereas pedal digit II is longer than digit I, it is shorter than digit III (Fig. 3A6). Digit IV preserves one phalanx, which is quite slender (Fig. 2A5, A6). As this phalanx is not an ungual bone, the total number of phalanges in this digit remains uncertain. There are no phalanges in the digit V. This condition is shared with other proterochampsids, whereas a putative vestigial phalanx has been reported for Stenoscelida aurantiacus (CAPPA/UFSM 0293; Müller et al. 2022).
Stratigraphic and geographic range.—Type locality and horizon only.
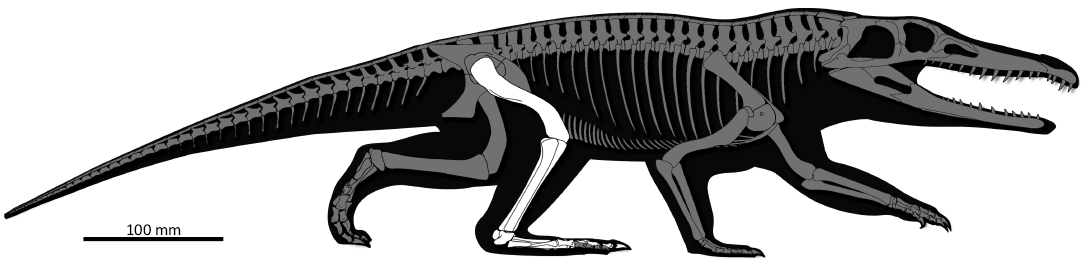
Fig. 2. Reconstruction of the skeleton of Retymaijychampsa beckerorum gen. et sp. nov. depicting the preserved elements; based on the artwork of Mauricio Silva Garcia (São João do Polêsine, Brazil).
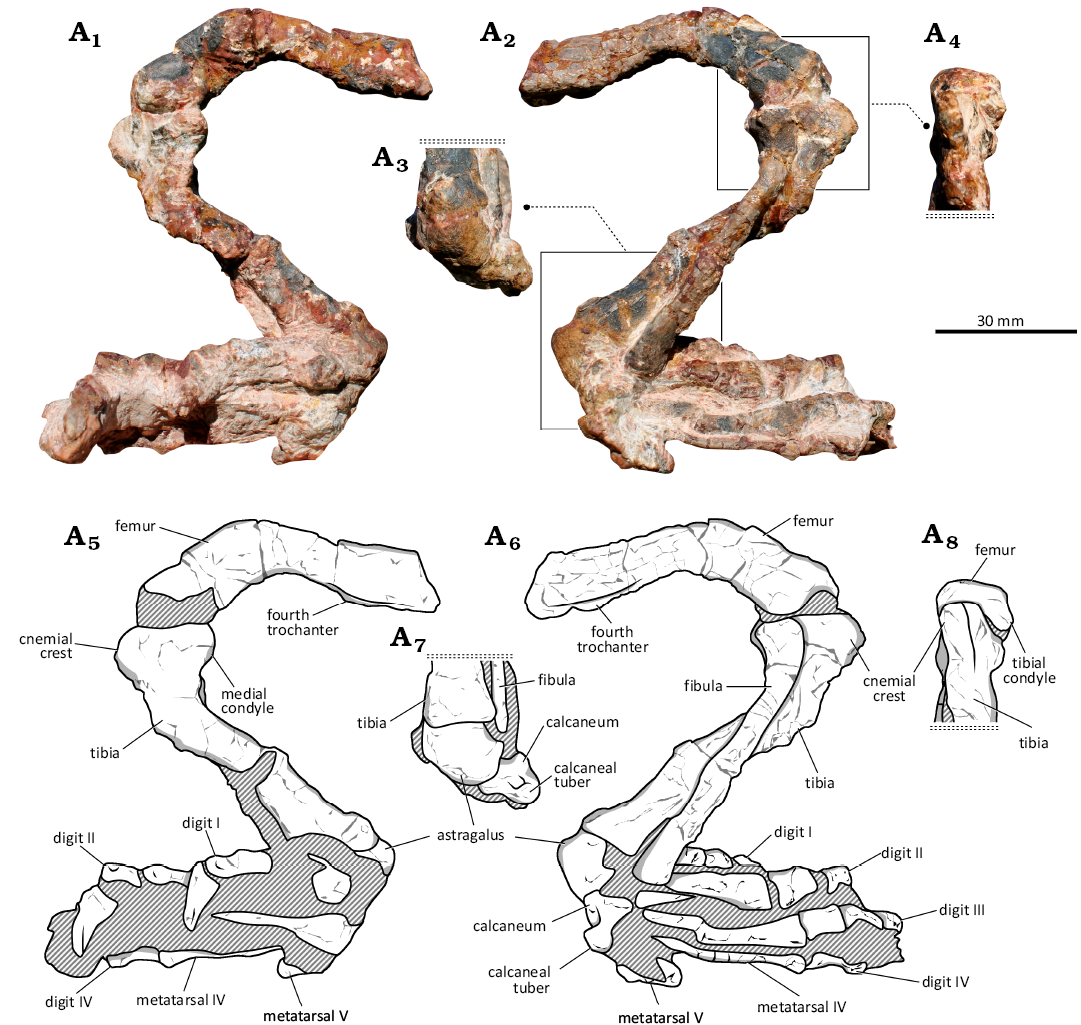
Fig. 3. Holotype of the archosauriform proterochampsid Retymaijychampsa beckerorum gen. et sp. nov. (CAPPA/UFSM 0430) from the Linha Várzea 2 (Becker) Site, Paraíso do Sul, Rio Grande do Sul, Brazil; Ladinian to lower Carnian, Middle to lower Upper Triassic. Right hindlimb in medial (A1, A5), lateral (A2, A6), posterior (A3, A7), and anterior (A4, A8) views. Photographs (A1–A4) and explanatory drawings (A5–A8). The stippled lines associated with A3, A4, A7, and A8 indicate that there is continuity of the bones, but they are not represented in the images.
Phylogenetic analysis
The phylogenetic analysis recovered a single most parsimonious tree (Fig. 4) of 914 steps each, with a consistency index of 0.521 and a retention index of 0.557. Retymaijychampsa beckerorum gen. et sp. nov. is found in a trichotomy with both species of Proterochampsa, based on the elongated fourth trochanter of the femur (ch. 329: 0→1) and the ratio between the maximum width of the proximal end and the total length of the tibia (ch. 339: 2→3). Sphodrosaurus pennsylvanicus is the sister taxon to the node comprising R. beckerorum gen. et sp. nov. plus both species of Proterochampsa. The clade comprising these four OTUs is named here Proterochampsinae and is supported by five synapomorphies: ratio between the maximum width of the skull and length of the presacral vertebral column ranging from 0.336 to 0.440 (ch. 12: 1→3); distinct coarse ornamentation on lateral surface of the surangular and angular (ch. 187: 0→1); in medial view, the area ventral to the internal mandibular fenestra shows a larger contribution from the angular bone compared to the prearticular, or both bones contribute equally (ch. 188: 0→1); prearticular with ventral margin posterior to its contact with the splenial straight or ventrally curved on the anterior half of the bone in medial or lateral view (ch. 189: 1→0); and presence of ventral keel on the centrum of the ninth presacral vertebra (ch. 231: 0→1). Stenoscelida aurantiacus is recovered as the sister taxon to Proterochampsidae. The inner composition of Proterochampsidae and Doswelliidae follows previous analyses of the original data matrix (Paes-Neto et al. 2024).
An interesting aspect to note is that, prior to the advent of computational phylogenetic analyses, Bonaparte (1971) divided proterochampsians into two main families: “Proterochampsidae” and “Cerritosauridae”. Although far fewer proterochampsian species were known at the time, this subdivision closely aligns with the results obtained in the present analysis.
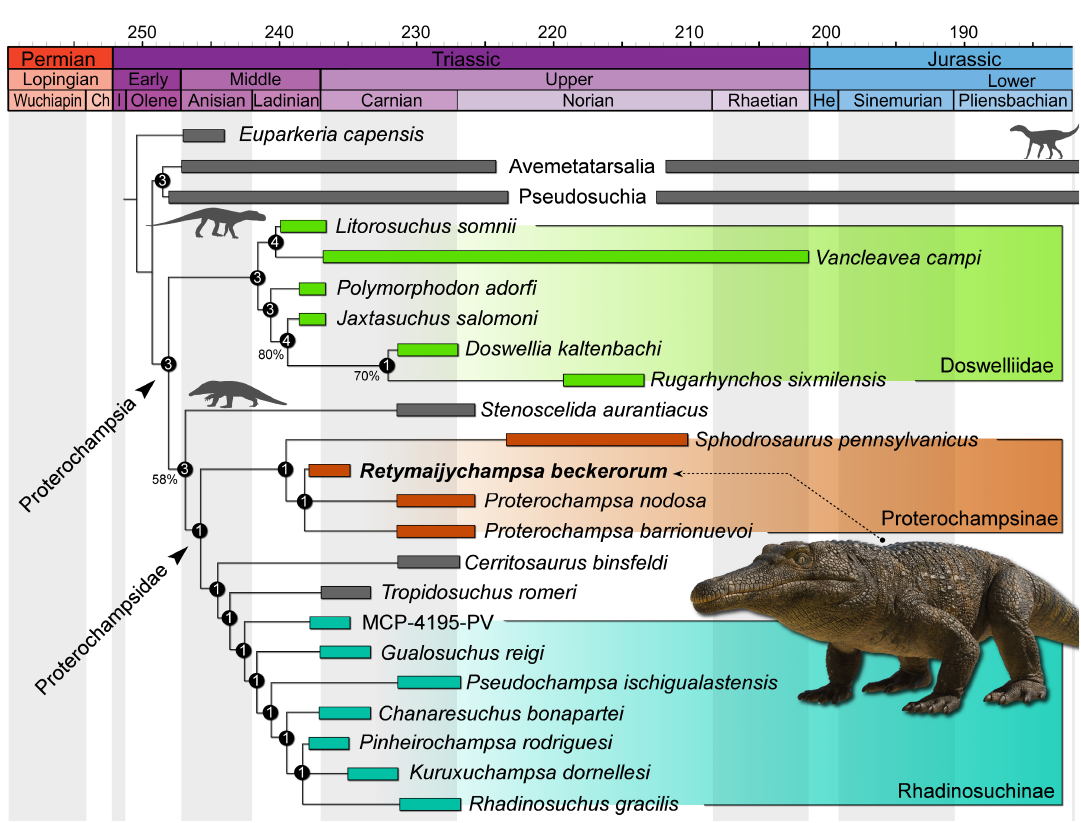
Fig. 4. Time-calibrated most parsimonious tree depicting the phylogenetic position of Retymaijychampsa beckerorum gen. et sp. nov. Number on nodes represent Bremer support values and Bootstrap values higher than 50%. The temporal bars for each OTU represent the maximum and minimum ages of each geological unit. Life reconstruction of Retymaijychampsa beckerorum gen. et sp. nov. by Caio Fantini (Guarulhos, Brazil). Silhouettes based on the artwork of Matheus Fernandes Gadelha (Limoeiro do Norte, Brazil). Abbreviations: Ch, Changhsingian; He, Hettangian; I, Induan.
Discussion
Although most typical traits of proterochampsids concern cranial bones (Ezcurra 2016), Retymaijychampsa beckerorum gen. et sp. nov. is confidently assigned to Proterochampsidae based on the configuration of the pes, which has a stout metatarsal II combined with a rather slender metatarsal IV (e.g., Trotteyn et al. 2013). On the other hand, the new proterochampsid had a unique set of traits that are associated with the robustness of the hindlimb and the shape of the distal end of the femur. The significance of these features is uncertain. Perhaps R. beckerorum gen. et sp. nov. had a posture or mode of locomotion distinct from other proterochampsids. In addition, the holotype (CAPPA/UFSM 0430) represents a small individual. In proterochampsids, the tibia length ranges from approximately 75% (Proterochampsa barrionuevoi; Trotteyn 2011) to 100% (Tropidosuchus romeri; Arcucci 1990) of the total femur length. Based on the tibia length of 70.5 mm and the cited tibia-to-femur ratios in proterochampsids, the estimated femoral length of CAPPA/UFSM 0430 should range between 70.5 mm and approximately 94 mm. According to these values, the length of the skull of CAPPA/UFSM 0430 is estimated between 79.8 to 126.4 mm based on the regression calculated by Müller et al. (2022). Therefore, the holotype of R. beckerorum gen. et sp. nov. was a small-sized proterochampsid when compared to most members of the clade. For comparison, the skull of the holotype of Pinheirochampsa rodriguesi is 276.5 mm in length (Paes-Neto et al. 2024), that of the holotype of Gualosuchus reigi is 330 mm (Romer 1971), and the skulls of some specimens of Proterochampsa barrionuevoi reached approximately 500 mm (Dilkes and Arcucci 2012). Nevertheless, there are some small proterochampsids, such as Tropidosuchus romeri, whose skull was approximately 76 mm in length (Arcucci 1990). Despite the reduced size, a histological investigation suggested that a specimen of Tropidosuchus romeri (PVL 4606) reached sexual and skeletal maturity, though somatic maturity could not be inferred (García Marsa et al. 2023). Therefore, whereas small proterochampsids are present in the fossil record, further investigations and/or specimens are necessary to determine whether CAPPA/UFSM 0430 represents a skeletally immature individual.
Retymaijychampsa beckerorum gen. et sp. nov. represents the second proterochampsid from the Pinheiros-Chiniquá Sequence. The other species, Pinheirochampsa rodriguesi, is known solely from cranial remains (Paes-Neto et al. 2024). Therefore, a direct comparison between CAPPA/UFSM 0430 and specimens of Pinheirochampsa rodriguesi is not feasible. Even the size difference between CAPPA/UFSM 0430 and specimens of Pinheirochampsa rodriguesi are an ambiguous difference because the development degree of CAPPA/UFSM 0430 is uncertain. On the other hand, both species belong to different clades according to the phylogenetic analysis performed here. Retymaijychampsa beckerorum gen. et sp. nov. nests within Proterochampsinae, whereas Pinheirochampsa rodriguesi is a derived member of Rhadinosuchinae. Therefore, although more complete specimens are needed to further investigate this taxonomic issue, the cladistic data supports the presence of two phylogenetically distinct proterochampsids in the Pinheiros-Chiniquá Sequence. Actually, the co-occurrence of different proterochampsian species is common in South American deposits (Fig. 5). For instance, in the Massetognathus–Chanaresuchus Assemblage Zone of the Argentinean Chañares Formation there are three coeval species (Ezcurra et al. 2021). In the Hyperodapedon Assemblage Zone of the Brazilian Candelária Sequence, there are four species distributed across two sub-zones (Müller et al. 2022). The co-occurrence of R. beckerorum gen. et sp. nov. and Pinheirochampsa rodriguesi reinforces an ecological model in which proterochampsians were relatively diverse in Middle to early Late Triassic environments (Fig. 6). Moreover, if the size differences between the two species are not due to ontogenetic variation, it is plausible that these species occupied distinct niches, especially if the unique hindlimb morphology of R. beckerorum gen. et sp. nov. is associated with a distinct locomotor strategy compared to other proterochampsians. In fact, archosauriforms evolved a broad range of postures and locomotor strategies during the Triassic (Agnolín et al. 2024).
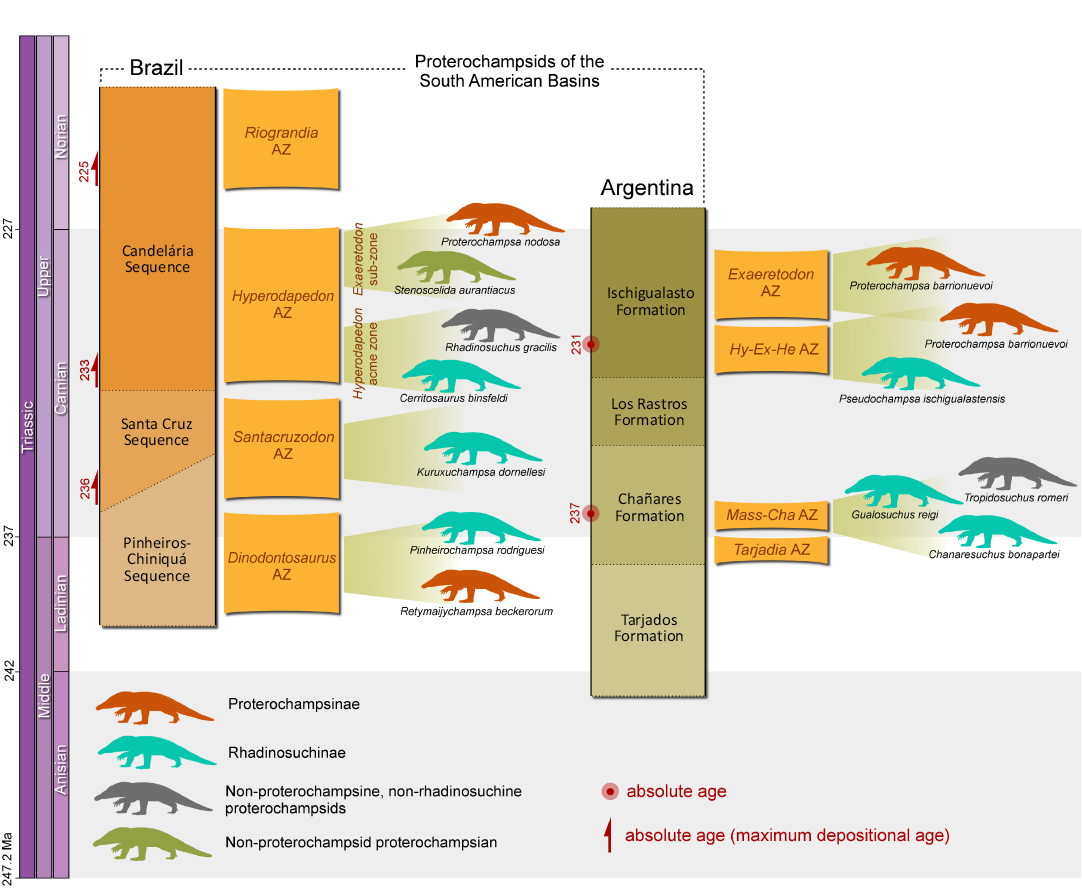
Fig. 5. Stratigraphic distribution of proterochampsids in South American deposits. Radioisotopic data according to Martínez et al. (2011), Marsicano et al. (2016), Ezcurra et al. (2017), Langer et al. (2018), and Philipp et al. (2018). Silhouettes based on the artwork of Matheus Fernandes Gadelha (Limoeiro do Norte, Brazil). Abbreviations: AZ, Assemblage Zone; Hy-Ex-He, Hyperodapedon–Exaeretodon–Herrerasaurus; Mass-Cha, Massetognathus–Chanaresuchus.

Fig. 6. Artistic representation of a Middle–Late Triassic landscape of southern Brazil depicting two individuals of the proterochampsid archosauriform Retymaijychampsa beckerorum gen. et sp. nov. Artwork by Caio Fantini (Guarulhos, Brazil).
Another interesting aspect of the discovery of R. beckerorum gen. et sp. nov. concerns its phylogenetic position. So far, there have been no records of Ladinian to early Carnian proterochampsids more closely related to Proterochampsa (i.e., Proterochampsinae) than to rhadinosuchines, resulting in a ghost lineage extending from the Middle to early Late Triassic (Ezcurra and Sues 2021; Paes-Neto et al. 2024). Retymaijychampsa beckerorum gen. et sp. nov. partially fills this gap and represents the oldest known proterochampsine proterochampsid worldwide. It highlights the phylogenetic importance of postcranial remains in order to further explore the evolutionary history of these archosauriforms. In addition, the composition of Proterochampsinae including three South American taxa along with the North American Sphodrosaurus pennsylvanicus (Paes-Neto et al. 2024; here), suggests a wide geographical distribution of this clade. These affinities add to the growing number of Triassic tetrapod taxa shared between deposits in South America and North America (Irmis et al. 2007; Martínez et al. 2016; Novas et al. 2021; Ezcurra and Sues 2022; Fitch et al. 2023; Müller et al. 2023).
Conclusions
Retymaijychampsa beckerorum gen. et sp. nov. represents the second proterochampsid from the Pinheiros-Chiniquá Sequence (Ladinian to upper Carnian) of Brazil and comprises one of the oldest proterochampsids worldwide. It has a diagnostic set of hindlimb traits and is phylogenetically close to Proterochampsa, partially filling a ghost lineage of proterochampids more closely related to Proterochampsa than to Rhadinosuchinae. Finally, the co-occurrence of R. beckerorum gen. et sp. nov. and Pinheirochampsa rodriguesi provides further support for the idea of high ecomorphological diversity among proterochampsians in Middle to early Late Triassic environments.
Acknowledgements
I thank Pedro Lucas Porcela Aurélio (Cachoeira do Sul, Brazil) for the discovery of the material described here, Becker family (Paraíso do Sul, Brazil) for kindly allowing the palaeontological prospection and fieldwork at their property, Federico Agnolín (Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Buenos Aires, Argentina) and Hans-Dieter Sues (Smithsonian Institution, Washington DC, USA) for their valuable comments and suggestions, Caio Fantini (Guarulhos, Brazil) for the artistic representation of the new taxon, and Dilson Vargas Peixoto (Santa Maria, Brazil) for the linguistic assistance in the creation of the new taxon’s name. I also thank the Willi Hennig Society, for making the TNT software freely available. This work was carried out with aid of the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq 404095/2021-6, 303034/2022-0, and 406902/2022-4).
Editor: Daniel Barta.
References
Agnolín, F.L., Novas, F.E., Ezcurra, M.D., Miner, S., and Müller, R.T. 2024. Comments on the pelvic girdle anatomy of Lagerpeton chanarensis Romer, 1971 (Archosauria) and its implications on the posture and gait of early pterosauromorphs. The Anatomical Record 307: 1001–1010. Crossref
Arcucci, A. 1990. Un nuevo Proterochampsidae (Reptilia-Archosauriformes) de la fauna local de Los Chañares (Triásico Medio), La Rioja, Argentina. Ameghiniana 27: 365–378.
Arcucci, A., Previtera, E., and Mancuso, A. C. 2019. Ecomorphology and bone microstructure of Proterochampsia from the Chañares Formation. Acta Palaeontologica Polonica 64: 157–170. Crossref
Barberena, M.C. 1982. Uma nova espécie de Proterocampsa (Proterocampsa nodosa sp. nov.) do Triássico do Brasil. Anais da Academia Brasileira de Ciências 54: 127–141.
Benton, M.J. 2016. The Triassic. Current Biology 26: R1214–R1218. Crossref
Bonaparte, J.F. 1971. Cerritosaurus binsfeldi Price, Tipo de una Nueva Familia de Tecodontes (Pseudosuchia–Proterochampsia). Anais da Academia Brasileira de Ciências 43: 417–422.
Dilkes, D.W. 1998. The Early Triassic rhynchosaur Mesosuchus browni and the interrelationships of basal archosauromorph reptiles. Philosophical Transactions of the Royal Society of London B Biological Sciences 353: 501–541. Crossref
Dilkes, D. and Arcucci, A. 2012. Proterochampsa barrionuevoi (Archosauriformes: Proterochampsia) from the Late Triassic (Carnian) of Argentina and a phylogenetic analysis of Proterochampsia. Palaeontology 55: 853–885. Crossref
Ezcurra, M.D. 2010. Biogeography of Triassic tetrapods: evidence for provincialism and driven sympatric cladogenesis in the early evolution of modern tetrapod lineages. Proceedings of the Royal Society B: Biological Sciences 277: 2547–2552. Crossref
Ezcurra, M.D. 2016. The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriforms. PeerJ 4: e1778. Crossref
Ezcurra, M.D. and Sues, H.D. 2021. A re-assessment of the osteology and phylogenetic relationships of the enigmatic, large-headed reptile Sphodrosaurus pennsylvanicus (Late Triassic, Pennsylvania, USA) indicates archosauriform affinities. Journal of Systematic Palaeontology 19: 1643–1677. Crossref
Ezcurra, M.D., Fiorelli, L.E., Martinelli, A.G., Rocher, S., von Baczko, M.B., Ezpeleta, M., Taborda, J.R.A., Hechenleitner, E.M., Trotteyn, M.J., and Desojo, J.B. 2017. Deep faunistic turnovers preceded the rise of dinosaurs in southwestern Pangaea. Nature Ecology and Evolution 1: 1477–1483. Crossref
Ezcurra, M.D., Montefeltro, F.C., Pinheiro, F.L., Trotteyn, M.J., Gentil, A.R., Lehmann, O.E., and Pradelli, L.A. 2021. The stem-archosaur evolutionary radiation in South America. Journal of South American Earth Sciences 105: 102935. Crossref
Ezcurra, M.D., Nesbitt, S.J., Bronzati, M., Dalla Vecchia, F.M., Agnolin, F.L., Benson, R.B.J., Brissón Egli, F., Cabreira, S.F., Evers, S.W., Gentil, A.R., Irmis, R.B., Martinelli, A.G., Novas, F.E., Silva, L.R., Smith, N.D., Stocker, M.R., Turner, A.H., and Langer, M.C. 2020. Enigmatic dinosaur precursors bridge the gap to the origin of Pterosauria. Nature 588: 445–449. Crossref
Fitch, A.J., Haas, M., C’Hair, W., Ridgley, E., Ridgley, B., Oldman, D., Reynolds, C., and Lovelace, D.M. 2023. A new rhynchosaur taxon from the Popo Agie Formation, WY: Implications for a northern Pangean Early–Late Triassic (Carnian) Fauna. Diversity 15: 544. Crossref
García Marsa, J.A., Ponce, D.A., Agnolin, F., and Novas, F.E. 2023. Histovariability and lifestyle in Proterochampsidae Romer, 1966 (Archosauriformes) from the Chañares Formation (Late Triassic), northwestern Argentina. Comptes Rendus Palevol 22: 605–622. Crossref
Gauthier, J.A., Kluge, A.G., and Rowe, T. 1988. Amniote phylogeny and the importance of fossils. Cladistics 4: 105–209. Crossref
Goloboff, P.A. and Catalano, S.A. 2016. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32: 221–238. Crossref
Hsiou, A.S., Abdala, F., and Arcucci, A. 2002. Novo registro de proterocampsídeo (Reptilia-Archosauriformes) do Triássico médio-superior do Brasil. Revista Brasileira de Paleontologia 3: 48–55.
Horn, B.L.D., Melo, T.M., Schultz, C.L., Philipp, R.P., Kloss, H.P., and Goldberg, K. 2014. A new third-order sequence stratigraphic framework applied to the Triassic of the Paraná Basin, Rio Grande do Sul, Brazil, based on structural, stratigraphic and paleontological data. Journal of South American Earth Sciences 55: 123–132. Crossref
Huene, F. von 1938. Die fossilen Reptilien des südamerikanischen Gondwanalandes. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, Abteilung B 3: 142–151.
Huene, F. von 1946. Die grossen Stämme der Tetrapoden in den geologischen Zeiten. Biologische Zentralblatt 65: 268–275.
Hutchinson, J.R. and Gatesy, S.M. 2000. Adductors, abductors, and the evolution of archosaur locomotion. Paleobiology 26: 734–751. Crossref
Irmis, R.B., Nesbitt, S.J., Padian, K., Smith, N.D., Turner, A.H., Woody, D., and Downs, A. 2007. A Late Triassic dinosauromorph assemblage from New Mexico and the rise of dinosaurs. Science 317: 358–361. Crossref
Langer, M.C., Marsola, J.C., Müller, R.T., Bronzati, M., Bittencourt, J.S., Apaldetti, C., and Ezcurra, M.D. 2022. The early radiation of sauropodomorphs in the Carnian (Late Triassic) of South America. In: A. Otero, J.L. Carballido, and D. Pol (eds.), South American Sauropodomorph Dinosaurs: Record, Diversity and Evolution, 1–49. Springer International Publishing, Cham. Crossref
Langer, M.C., Ramezani, J., and Da-Rosa, Á.A. 2018. U-Pb age constraints on dinosaur rise from south Brazil. Gondwana Research 57: 133–140. Crossref
Li, C., Wu, X.C., Zhao, L.J., Nesbitt, S.J., Stocker, M.R., and Wang, L.T. 2016. A new armored archosauriform (Diapsida: Archosauromorpha) from the marine Middle Triassic of China, with implications for the diverse life styles of archosauriforms prior to the diversification of Archosauria. The Science of Nature 103: 1–23. Crossref
Martínez, R.N., Apaldetti, C., Correa, G.A., and Abelín, D. 2016. A Norian lagerpetid dinosauromorph from the Quebrada del Barro Formation, northwestern Argentina. Ameghiniana 53: 1–13. Crossref
Martinez, R.N., Sereno, P.C., Alcober, O.A., Colombi, C.E., Renne, P.R., Montañez, I.P., and Currie, B.S. 2011. A basal dinosaur from the dawn of the dinosaur era in southwestern Pangaea. Science 3316: 206–210. Crossref
Marsicano, C.A., Irmis, R.B., Mancuso, A.C., Mundil, R., and Chemale, F. 2016. The precise temporal calibration of dinosaur origins. Proceedings of the National Academy of Sciences 113: 509–513. Crossref
Müller, R.T. and Garcia, M. S. 2022. Oldest dinosauromorph from South America and the early radiation of dinosaur precursors in Gondwana. Gondwana Research 107: 42–48. Crossref
Müller, R.T., Ezcurra, M.D., Garcia, M.S., Agnolin, F.L., Stocker, M.R., Novas, F.E., Soares, M.B., Kellner, A.W.A., and Nesbitt, S.J. 2023. New reptile shows dinosaurs and pterosaurs evolved among diverse precursors. Nature 620: 589–594. Crossref
Müller, R.T., Garcia, M.S., and Fonseca, A.D. O. 2022. A new proterochampsid (Archosauriformes: Proterochampsia) from the Late Triassic of southern Brazil and the emergence of archosaurian hind limb traits. Journal of Systematic Palaeontology 20: 1–19. Crossref
Nesbitt, S.J. 2011. The early evolution of archosaurs: relationships and the origin of major clades. Bulletin of the American Museum of Natural History 352: 1–292. Crossref
Novas, F.E., Agnolin, F.L., Ezcurra, M.D., Müller, R.T., Martinelli, A.G., and Langer, M.C. 2021. Review of the fossil record of early dinosaurs from South America, and its phylogenetic implications. Journal of South American Earth Sciences 110: 103341. Crossref
Paes-Neto, V.D., Lacerda, M.B., Ezcurra, M.D., Raugust, T., Trotteyn, M.J., Soares, M.B., Schultz, C.L., Pretto, F.A., Francischini H., and Martinelli, A.G. 2024. New rhadinosuchine proterochampsids from the late Middle–early Late Triassic of southern Brazil enhance the diversity of archosauriforms. The Anatomical Record 307: 851–889. Crossref
Philipp, R.P., Schultz, C.L., Kloss, H.P., Horn, B.L., Soares, M.B., and Basei, M.A. 2018. Middle Triassic SW Gondwana paleogeography and sedimentary dispersal revealed by integration of stratigraphy and U-Pb zircon analysis: the Santa Cruz Sequence, Paraná Basin, Brazil. Journal of South American Earth Sciences 88: 216–237. Crossref
Price, L.I. 1946. Sobre um novo pseudosúquio do Triássico Superior do Rio Grande do Sul. Boletim Divisão de Geologia e Mineralogia 120: 1–38.
Reig, O.A. 1959. Primeros datos descriptivos sobre nuevos reptiles arcosaurios del Triássico de Ischigualasto (San Juan, Argentina). Revista de la Asociación Geológica Argentina 13: 257–270.
Romer, A.S. 1971. The Chañares (Argentina) Triassic reptile fauna. XI. Two new long-snouted thecodonts, Chanaresuchus and Gualosuchus. Museum of Comparative Zoology Bulletin 142: 387–421.
Schultz, C.L., Martinelli, A.G., Soares, M.B., Pinheiro, F.L., Kerber, L., Horn, B.L.D., Pretto, F.A., Müller, R.T., and Melo, T.P. 2020. Triassic faunal successions of the Paraná Basin, southern Brazil. Journal of South American Earth Sciences 104: 102846. Crossref
Sengupta, S., Ezcurra, M.D., and Bandyopadhyay, S. 2024. The redescription of Malerisaurus robinsonae (Archosauromorpha: Allokotosauria) from the Upper Triassic lower Maleri Formation, Pranhita-Godavari Basin, India. The Anatomical Record 307: 1315–1365. Crossref
Sereno, P.C. 1991. Basal archosaurs: phylogenetic relationships and functional implications. Journal of Vertebrate Paleontology 11: 1–53. Crossref
Sill, W.D. 1967. Proterocampsa barrinuevoi and the early evolution of the Crocodilia. Bulletin of the Museum of Comparative Zoology 135: 415–446.
Trotteyn, M.J. 2011. Material postcraneano de Proterochampsa barrionuevoi Reig 1959 (Diapsida: Archosauriformes) del Triásico Superior del centro-oeste de Argentina. Ameghiniana 48: 424–446. Crossref
Trotteyn, M.J. and Ezcurra, M.D. 2014. Osteology of Pseudochampsa ischigualastensis gen. et comb. nov. (Archosauriformes: Proterochampsidae) from the early Late Triassic Ischigualasto Formation of northwestern Argentina. PLOS ONE 9: e111388. Crossref
Trotteyn, M.J. and Ezcurra, M.D. 2020. Redescription of the holotype of Chanaresuchus bonapartei Romer, 1971 (Archosauriformes: Proterochampsidae) from the Upper Triassic rocks of the Chañares Formation of north-western Argentina. Journal of Systematic Palaeontology 18: 1415–1443. Crossref
Trotteyn, M.J., Arcucci, A.B., and Raugust, T. 2013. Proterochampsia: an endemic archosauriform clade from South America. Geological Society, London, Special Publications 379: 59–90. Crossref
Zerfass, H., Lavina, E.L., Schultz, C.L., Garcia, A.J.V., Faccini, U.F., and Chemale, F. Jr. 2003. Sequence stratigraphy of continental Triassic strata of Southernmost Brazil: a contribution to Southwestern Gondwana palaeogeography and palaeoclimate. Sedimentary Geology 161: 85–105. Crossref
Acta Palaeontol. Pol. 70 (1): 7–16, 2025
https://doi.org/10.4202/app.01204.2024