
Generic revision of holocystitid blastozoans
CHRISTOPHER R.C. PAUL
Paul, C.R.C. 2025. Generic revision of holocystitid blastozoans. Acta Palaeontologica Polonica 70 (1): 125–141.
Brightonicystis is a valid genus characterised by nine or more oral plates, with two in the B and D ambulacra not reaching the inner mouth edge; five ambulacral facets shared by oral and facetal plates; two generations of smooth thecal plates with buried humatipores. Brightonicystis contains two named species, Brightonicystis gregaria and Brightonicystis salmoensis, with other unnamed taxa known from Canada and Sweden, all of which are Hirnantian. All other holocystitid genera are Silurian. A review of the Holocystitidae confirms the established genera Holocystites and Brightonicystis both characterised by five ambulacra with shared facets, as well as Trematocystis, Pustulocystis, and Paulicystis all characterised by four ambulacra. The hypothesis that the differences between Pentacystis (without orals) and Osgoodicystis (with two or six orals) are taphonomic is reviewed and rejected. The peristome border and food grooves should be absent in Pentacystis if the oral plates on which they occur were lost during preservation. Pentacystis species show both features developed on facetal plates. Osgoodicystis is reinstated as a valid genus with six narrow orals. Moyacystis gen. nov., type species Osgoodicystis cooperi, bears only two orals (O1 and O6) and facetal plates with oral pores in the peristome border. Pentacystis, Osgoodicystis, and Moyacystis gen. nov., all possess five ambulacral facets restricted to single facetal plates and a smooth thecal surface. The Australian genus Austrocystites is a junior synonym of Trematocystis. Holocystitid subfamilies and subgenera introduced by Frest and Strimple (in Frest et al. 2011) are not discussed herein.
Key words: Blastozoa, Diploporita, Holocystitidae, taphonomy, oral plate arrangements.
Christopher R.C. Paul [glcrcp@bristol.ac.uk; ORCID: https://orcid.org/0000-0003-3670-3325 ], School of Earth Sciences, University of Bristol, Wills Memorial Building, Queen’s Road, Bristol, UK.
Received 24 September 2024, accepted 29 January 2025, published online 19 March 2025.
Copyright © 2025 C.R.C. Paul. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Paul (1971: 142–149) originally described the genus Brightonicystis, type species Brightonicystis gregaria Paul, 1971, based on a few specimens from the Hirnantian, Ashgill Shales, near Knock, Cumbria, UK. Other specimens from the Upper Ordovician of Sweden were left under open nomenclature because they did not preserve the critical plating in the oral area. Brightonicystis possesses respiratory structures, called humatipores (Paul 1972), which are confined to the diploporite family Holocystitidae Miller, 1889. It differed from all then known holocystitid genera by having a mouth frame composed of more plates (originally thought to be ten). Species of all other holocystitid genera have a mouth frame composed of six or eight plates (Fig. 1). In none of the original specimens of Brightonicystis gregaria could the full details of the oral area be seen. Searches for additional specimens were unsuccessful. Paul (1971: 70, fig. 24E, repeated in Paul 1973: 13, fig. 8D) introduced an inaccurate diagram of the oral area in Brightonicystis. The two original drawings of the oral and anal area (Paul 1971: figs. 24E and 64; reproduced herein as Fig. 2) are incompatible. This ambiguity was made worse by the omission of one of the plate sutures in a later figure (Paul 1973: 56, fig. 34b).
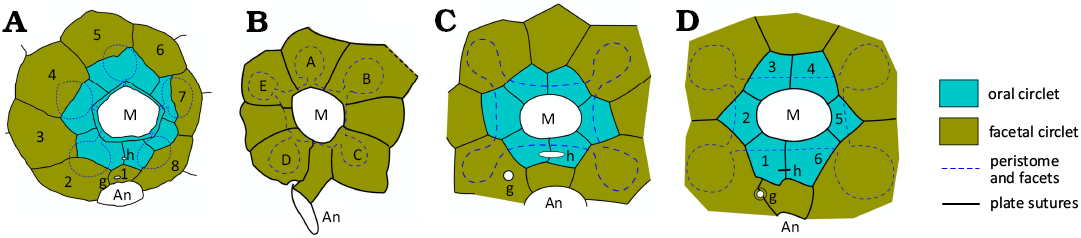
Fig. 1. Interpretive diagrams of oral plating in some holocystitid genera. A. Holocystites Hall, 1864, with six oral plates forming the mouth frame, eight facetal plates and five ambulacral facets shared between plates of both circlets (1–8, facetals). Based on USNM S3070, Holocystites cylindricus (Hall, 1861), from Paul (1971: fig. 26c). B. Pentacystis Paul, 1971, with no orals, eight facetals and five facets. Based on AMNH 20271a, Pentacystis simplex Paul, 1971, from Paul (1971: fig. 42b). C. Trematocystis Jaekel, 1899, with six orals, eight facetals and four facets. Based on UMMP 45862, Trematocystis globosus (Miller, 1878), from Paul (1971: fig. 50a). D. Pustulocystis Paul, 1971, with six orals and facetals plus four facets (1–6, orals). Based on AMNH 20273, Pustulocystis ornatissimus (Miller, 1891), from Paul (1971: fig. 58a). Abbreviations: A–E, Carpenter’s ambulacral designations; An, anus; g, gonopore; h, hydropore; M, mouth. Plate colours and dashed outline of peristome and facets standard for following diagrams. Not to scale.
In the 1970s the New Point Stone Company’s quarry at Napoleon, Indiana, USA was expanded and yielded large numbers of additional holocystitid specimens, including new taxa. Frest (1983: 279) recorded thousands of new diploporite specimens, compared with a total known fauna of about 200 discovered previously from all known localities. The new taxa were finally published posthumously in Frest et al. (2011), but the contents are virtually identical to chapter 7 of Terry Frest’s Ph.D. Thesis (Frest 1983).
Recently, Sheffield (2017: 18), Sheffield and Sumrall (2017: 756; 2019: 743) and Sheffield et al. (2018: 2) have expressed doubts as to the validity of the genera Brightonicystis and Osgoodicystis Frest & Strimple in Frest et al., 2011. They argued that no illustration clearly showed the supposed ten plates forming the oral frame of Brightonicystis (Sheffield 2017: 18; Sheffield and Sumrall 2017: 756) and that the possession of ten oral plates was most unlikely (Sheffield and Sumrall 2019: 743). In addition, as originally defined, Pentacystis Paul, 1971, included species with (e.g., Holocystites wykoffi Miller, 1891) and without (e.g., Pentacystis simplex Paul, 1971) oral plates. Subsequently, Frest (1983: 427; Frest et al. 2011: 92) separated a new genus Osgoodicystis Frest and Strimple in Frest et al., 2011, for those species with any orals. Sheffield and Sumrall (2015) argued that Pentacystis Paul, 1971, was based on a missinterpretation of the oral plating. They regarded absent orals as due to taphonomic loss during preservation, and therefore considered Osgoodicystis as a junior synonym of Pentacystis.
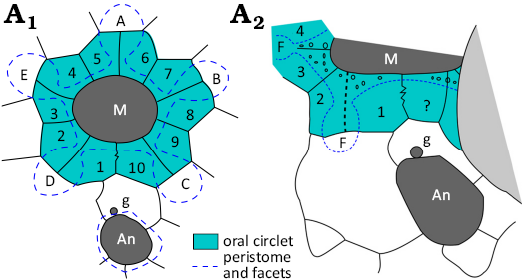
Fig. 2. Original interpretive diagrams of oral plating in Brightonicystis gregaria Paul, 1971. A1, redrawn from Paul 1971: 70, fig. 24a; A2, redrawn from Paul 1971: 146, fig. 64. The dashed suture line above the label F was omitted in the second version of this diagram (Paul 1973: 56, fig. 34b). ? represents a plate not present in A1. Abbreviations: A–E, Carpenter’s (1884, 1891) ambulacral designations; An, anus; F, ambulacral facet; g, gonopore; M, mouth. Pale shading indicates base of another epifaunal diploporite. Not to scale.
Despite their scepticism about Brightonicystis, Sheffield et al. (2018: 4) described a new holocystitid from the Hirnantian (Upper Ordovician) of Anticosti Island, Canada, as Holocystites salmoensis. Their diagram of the oral plating (Sheffield et al. 2018: fig. 5C; redrawn as Fig. 3A herein) shows the circlet immediately surrounding the mouth with nine plates, plus an outer circlet of eight plates, six of which share parts of the five ambulacral facets. An unexpected feature of this diagram is that the B and D ambulacra include plates that reach the outer margin of the peristome, but not the inner margin of the mouth. The unexpected arrangement of oral plates also occurs in the holotype and one paratype of Brightonicystis gregaria, strengthening the suggestion that Holocystites salmoensis is a species of Brightonicystis (Paul 2022). In summary, errors were made in the original description of Brightonicystis. Many new holocystitid specimens have resulted in new taxa being described since 1971, as well as new interpretations of the homology of oral plates in holocystitids and other blastozoans (Sumrall 2008; Sumrall and Waters 2012; Kammer et al. 2013). All of these facts make it appropriate to redescribe Brightonicystis and review other genera in the family Holocystitidae taxonomically. These are the aims of this paper.
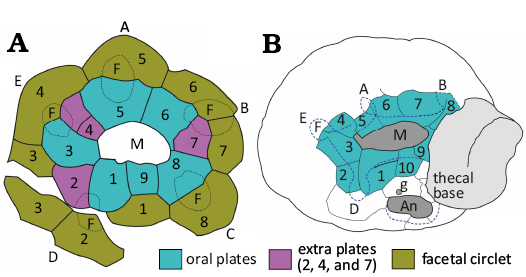
Fig. 3. Oral plating in Brightonicystis species. A. Original interpretation of the shapes and positions of oral and facetal plates in Holocystites salmoensis Sheffield et al., 2018, GSC 126899, coloured and relabelled. The mouth (M) is directly surrounded by a closed circlet of nine plates (1–9), here called orals, two of which (2 and 7) do not reach the inner mouth edge. A second closed circlet of eight plates, here called facetals, surrounds the orals. Ambulacral facets (F) are developed on 2–4 plates of both circlets. Sheffield et al. (2018) interpreted the mouth frame as having only six orals. Facetal 1 was interpreted as oral 7 under the UEH scheme. No explanation was given for the division of O4. B. New interpretation of the oral plating in Brightonicystis gregaria Paul, 1971, holotype, CAMSM A.32813. Note the displacement between orals 4 and 5, and the extra oral (compare with Fig. 3A). Abbreviations: A–E, Carpenter’s (1884, 1891) ambulacral designations; An, anus; F, facets; g, gonopore; M, mouth. Not to scale.
Institutional abbreviations.—AMNH, American Museum of Natural History, New York, USA; CAMSM, Sedgwick Museum, Cambridge, UK; CMCIP, Cincinnati Museum Centre, USA; GSC, Geological Survey of Canada, Ottawa, Canada; HM, Hunterian Museum, Glasgow, UK; MUMG, Miami University, Oxford, USA; SUI, University of Iowa, Iowa City, USA; UMMP, University of Michigan, Museum of Paleontology, Ann Arbor, USA; USNM, United States National Museum, Washington DC, USA; UWM, University of Wisconsin, Milwaukee, USA.
Other abbreviations.—A–E, Carpenter’s (1884, 1891) ambulacral designations; An, anus; F, FF, facetal plates; g, gonopore; h, hydropore; O, OO, oral plates; UEH, Universal Elemental Homology (Sumrall and Waters 2012).
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in Zoobank: urn:lsid:zoobank.org:pub: C0D24A07-AB52-4AB2-A3A9-1167FAD099C0.
Material and methods
The original type series of Brightonicystis gregaria Paul, 1971, has been re-examined and photographed in the Collections Centre of the Sedgwick Museum, Cambridge, UK. Photographs were taken with an Olympus Tough TG6 camera, using the microscope setting, which allows focusing to within 10 mm of the lens and automatically stacks up to eight images at different focal distances. It saves only the first image, closest to the camera, and the compound, stacked image. A ring light attachment was used for such close-up photography.
In addition, all the relevant literature since the initial revision of the “Holocystites fauna” (Paul 1971) has been reviewed, including chapter 7 of Frest’s Ph.D. Thesis (Frest 1983). Interpretive diagrams in this paper are restricted to those derived by overlaying photographs with outlines of plate sutures, or by using camera lucida drawings. This produces the most accurate results.
Both taxonomy and descriptions of taxa are more reliable if based on observable facts, rather than interpretations. Thus, in the systematic section descriptions of taxa are purely descriptive, especially in discussing the oral plating. The number of plates surrounding the mouth is uncontroversial; either there are six or eight, etc. Most holocystitid genera have a closed circlet of six plates forming the mouth frame, surrounded by a second closed circlet of eight plates, four or five of which bear ambulacral facets. Herein, the plates of the inner circlet are called “orals” and those of the outer circlet “facetals”. No homology is implied by either term. Under Universal Elemental Homology (Sumrall and Waters 2012), one of the plates in the outer circlet is considered to be an oral plate along with the six directly surrounding the mouth. This is based entirely on an interpretation of the plate homology, which, even if it is correct, is one step removed from the observable fact that there are two closed circlets, an inner one of six plates and an outer one of eight. Previously published alternative interpretations are evaluated using photographs, rather than diagrams. Consideration of how echinoderms grew often clarifies how plate patterns came about.
Systematic palaeontology
Phylum Echinodermata Leuckart, 1854
Subphylum Blastozoa Sprinkle, 1973
Class Diploporita Müller, 1854
Order Anambulacralia Paul & Toom, 2021
Diagnosis.—Stemless diploporites with an oral frame composed of either or both radial circumorals and interradial periorals, outside the peristome food grooves lie directly on thecal plates and lack both floor and cover plates (Paul and Toom 2021: 228).
Remarks.—The Anambulacralia includes four families, the Aristocystitidae Neumayr, 1889, Holocystitidae Miller, 1889, Parasphaeronitidae Bockelie, 1984, and Sphaeronitidae Neumayr, 1889. Anambulacralians entirely lack a stem, but were directly attached by an aboral attachment area to solid, or at least firm, substrates. They are distinguished from the other diploporites by the lack of a definite ambulacral structure. All other diploporites, whether directly attached or stem-bearing have ambulacra in which each ambulacral plate supported a single, erect feeding structure (commonly a brachiole). Rows of adambulacral plates either formed part of the thecal wall (mural ambulacra) as in the Protocrinitidae Bather, 1899, or were recumbent on thecal plates, as in the Mesocystidae Bather, 1899.
Family Holocystitidae Miller, 1889
Emended diagnosis.—Anambulacralians with four or five ambulacra each with a single large facet; with humatipores (simplified from Paul 1971).
Remarks.—Sphaeronitids and holocystitids are united by the presence of six inter-radial oral cover plates called palatals (Fig. 4). Holocystitids have pore-structures called humatipores (Fig. 5), whereas sphaeronitids bear diplopores and are characterized by a mouth frame of six orals, five of which are directly radial (Paul 2017: 587, fig. 5.1–3). Holocystitids have variable oral frame plating. Parasphaeronitids include two genera, both with an oral frame of eight plates, four of which are directly radial (Bockelie 1984). Aristocystitids have an elongate mouth surrounded by eight plates and covered by a double series of cover plates, larger outer cover plates and smaller central cover plates (Paul and Gutiérrez-Marco 2022: 154, fig. 1). Nevertheless, Oretanocalix Gutiérrez-Marco, 2000, the only aristocystitid known with five ambulacra, also has six oral cover plates that reach all the way from the edge of the peristome to the median suture. These are thought to be homologous with palatals, suggesting that the palate of holocystitids and sphaeronitids is a derived paedomorphic character. Furthermore, Oretanocalix and some other aristocystitid and sphaeronitid genera have diplopores that were covered by a thin epistereom in life. Thus, their diplopores were closer to the structure of the humatipores of holocystitids.
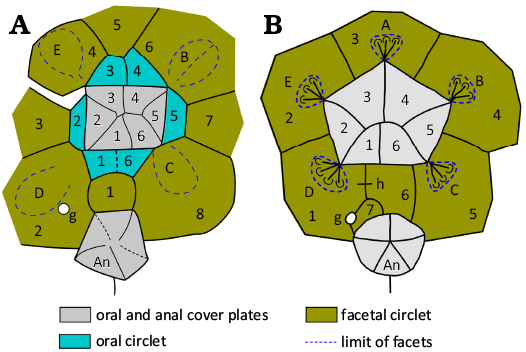
Fig. 4. Oral and anal cover plates in holocystitid (A) and sphaeronitid (B) blastozoans. A. Trematocystis rotundus (Miller, 1878) based on USNM S3063a. Redrawn from Paul (1971: 122, fig. 50c). B. Haplosphaeronis oblonga (Angelin, 1878). Redrawn from Paul (1973: 13, fig. 8b). Abbreviations: A–E, Carpenter’s (1884, 1891) ambulacral designations; An, anal pyramid; g, gonopore; h, hydropore. Not to scale.
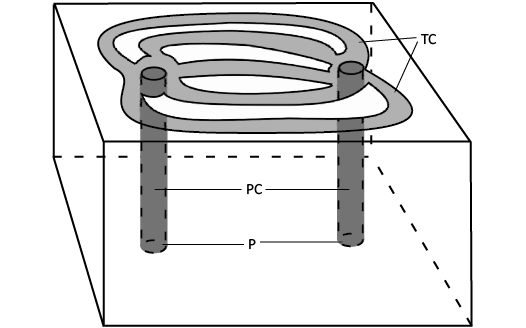
Fig. 5. Diagram of a humatipore. A humatipore consists of two perpendicular canals (PC) opening in pores (P) internally and connected just below the external surface by tangential canals (TC). Body fluids flowed up one perpendicular canal, respiratory gas exchange took place through the stereom over the tangential canals, then fluids flowed down the other tangential canal.
Frest (1983: 576, table 26) and Frest and Strimple in Frest et al. (2011: 69) subdivided the Holocystitidae into three subfamilies: the Holocystitinae, Pentacystinae, and Trematocystinae. These need re-assessment in cladistic terms (Sheffield and Sumrall 2017: 759, fig. 5) and are not considered further here, although the adjective “pentacystine” is useful in discussing relevant genera (see below). Frest’s (1983) subgenera of Holocystites Hall, 1864, are also not discussed further here.
Genus Brightonicystis Paul, 1971
Type species: Brightonicystis gregaria Paul, 1971, by original designation; from the Ashgill Shales (Hirnantian), Cumbria and W Yorkshire, UK.
Species included: Type species and Brightonicystis salmoensis (Sheffield et al., 2018).
Remarks.—Among holocystitid genera only Brightonicystis and Holocystites have their ambulacral facets developed over two to four oral and facetal plates. All other genera have the facets developed within a single facetal plate, except for Paulicystis Frest & Strimple in Frest et al., 2011, where the ambulacra were recumbent on the thecal surface and grew down the theca beyond the facetal circlet (Table 1). Brightonicystis differs from Holocystites in having nine or more, not six orals and in having at least four food grooves (A, B, D, and E) that follow sutures between orals.
Table 1. Key characters in holocystitid genera.
|
Genus /Character |
No. orals |
No. facetals |
No. facets |
Shared facets |
Recumbent ambulacra |
Humatipore type |
Plate generations |
|
Brightonicystis |
9–11 |
8 |
5 |
yes |
no |
buried |
2 |
|
Holocystites |
6 |
8 |
5 |
yes |
no |
raised or buried |
1–3 |
|
Moyacystis gen. nov. |
2 |
8 |
5 |
no |
no |
buried |
1 |
|
Osgoodicystis |
6 |
8 |
5 |
no |
no |
buried |
1 |
|
Paulicystis |
6 |
8 |
4 |
no |
yes |
buried |
1 |
|
Pentacystis |
0 |
8 |
5 |
no |
no |
buried |
1 |
|
Pustulocystis |
6 |
6 |
4 or 5 |
no |
no |
raised |
1 |
|
Trematocystis |
6 |
8 |
4 |
no |
no |
buried |
1 |
Emended diagnosis.—Holocystitids with five ambulacral facets on oral and facetal plates, oral frame composed of nine or more plates, with two in the B and D ambulacra not reaching the inner margin of the mouth, humatipores buried beneath smooth external surface with many tangential canals (emended from Paul 1971: 142).
Stratigraphic and geographic range.—Hirnantian, Upper Ordovician, of Anticosti Island (Canada), northern England (UK), and Kinnekulle (Sweden).
Brightonicystis gregaria Paul, 1971
Figs. 2A2, 6–8.
1966 Trematocystis sp.; Ingham 1966: 502.
1967 “Trematocystis” sp. nov.; Paul 1967: 147.
1967 Trematocystis sp. nov.; Jefferies et al. 1967: 569.
1971 Brightonicystis gregarius sp. nov.; Paul 1971: 143, figs. 62–64.
1972 Brightonicystis gregarius Paul; Paul 1972: 26, pl. 7: 6.
1973 Brightonicystis gregaria Paul; Paul 1973: 56, text-figs. 8D, 34, pl. 8: 4–9, pl. 9: 1.
1983 Brightonicystis gregarius Paul; Frest 1983: 483, fig. 56A.
2011 Brightonicystis gregarius Paul; Frest et al. 2012: 44, 47, 48, text-fig. 29A.
2017 Brightonicystis gregaria Paul; Paul 2017: 587, fig. 5.4.
Type material: Holotype: CAMSM A.32813, the most complete theca, preserved in original calcite showing the humatipores; ambulacrum C obscured by the base of another epifaunal specimen, but otherwise showing the critical oral area. Paratypes: CAMSM A.32811 (Fig. 6C), an almost complete theca free of matrix but with the base of another diploporite attached; CAMSM A.32812 (Fig. 6B), another almost complete theca still partially buried in sediment and with some of the plates in the oral area missing; CAMSM A.32814a, b, part and counterpart of a small area of articulated plates showing the humatopores well. All from the type locality.
Type locality: Banks of Swindale Beck, just upstream of the junction with Great Rundale Beck, near Knock, Cumbria, England, UK (National Grid reference NY688274).
Type horizon: Ashgill Shales, Hirnantian, Upper Ordovician.
Material.—Eight complete or partial thecae, including the holotype, two paratypes, and HM E5880, E5881, E5882, CAMSM A.43229, A.74804, plus numerous isolated plates, including the third paratype, CAMSM A.32814a. For details see Paul (1973: 56).
Emended diagnosis.—A species of Brightonicystis with at least ten oral plates, a globular theca, two generations of tumid plates with smooth external surfaces; up to nine or ten tangential canals per humatipore (emended from Paul 1971: 143).
Description.—Moderate-sized theca (20–35 mm across), globular or oviform, composed of about 100–150 smooth, gently convex thecal plates with slightly impressed sutures, with pentagonal mouth opposite a relatively large attachment area. Thecal plates of two generations (Fig. 6A3, B), larger primaries arranged in crude circlets, smaller secondaries usually triangular. All plates bear buried humatipores with many tangential canals (Fig. 6A3, B). Peristome pentagonal (Figs. 6C1, 7A1, A2) about 8 mm wide externally surrounding an oval mouth 6 mm wide in the holotype which is slightly crushed in an anterior-posterior direction; with a border up to 1.5 mm wide and with many oral pores (Fig. 6A1). Peristome frame composed of nine plates arranged as in Fig. 7A2. Narrow food grooves pass from the five ambulacral facets across the peristome border (Fig. 7: ambulacra B, D, and E; Fig. 8: ambulacra D, E, and A). Facets are oval, 2×1.7 mm, with a central ridge separating two lateral depressions, and developed on plates of both the oral and facetal circlets (Figs. 6A1, 7A2, 8A3), one facet per ambulacrum, except that paratype CAMSM A.32812 has two in the D ambulacrum (Fig. 8). Feeding appendages unknown. The periproct is polygonal, surrounding a rounded anus about 4 mm across (Figs. 6A3, 8A3). Oral and anal cover plates are unknown. The gonopore is a small circular orifice, 0.4 mm across, in the plate immediately adoral to the periproct and very close to the periproct border (Figs. 6A3, 7A2). A hydropore has not definitely been detected, although there is a groove in O1 of the holotype (Fig. 7A2). Attachment was direct by a large smooth attachment area up to about 50% of the thecal diameter. The holotype (CAMSM A.32813) and paratype (CAMSM A.32811a) both bear the bases of additional examples that attached post mortem (Fig. 6A3, C2).
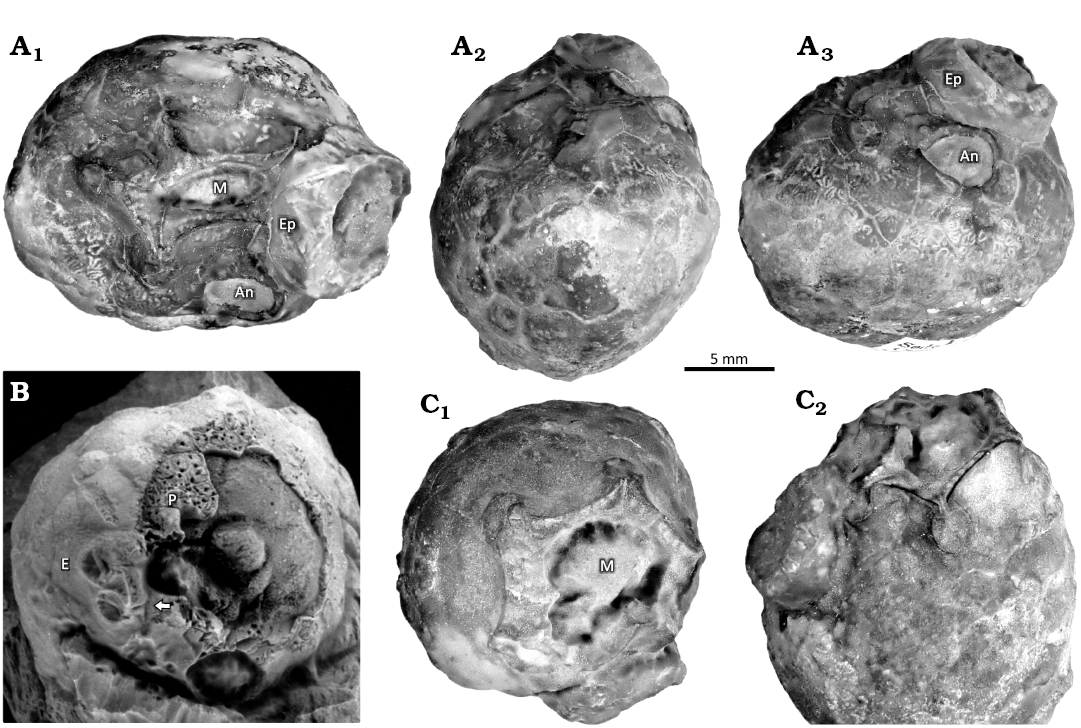
Fig. 6. Photographs of holocystitid blastozoan Brightonicystis gregaria Paul, 1971, from Hirnantian (Upper Ordovician), Knock, Cumbria, England, UK. A. Holotype CAMSM A.32813. A1, oral view showing slightly distorted central mouth (M) and anus (An) with C ambulacral facet and food groove hidden by base of epifaunal theca (Ep), compare with Fig. 7; A2, left lateral view showing thecal outline with anus (upper right); A3, posterior lateral view showing anus, small circular gonopore, some humatipores centrally below the anus, and gently convex thecal plates of two generations. B. Paratype CAMSM A.32812, showing weathered thecal plates with open perpendicular canals of humatipores (P). Note the branched food groove of the D ambulacrum (arrow) and facet of the E ambulacrum (E). Specimen whitened with NH4Cl. C. Paratype CAMSM A.32811a. C1, oral view showing pentagonal peristome and nearly circular mouth; C2, oblique lateral view centred on the B ambulacral facet showing oral pores and food groove.
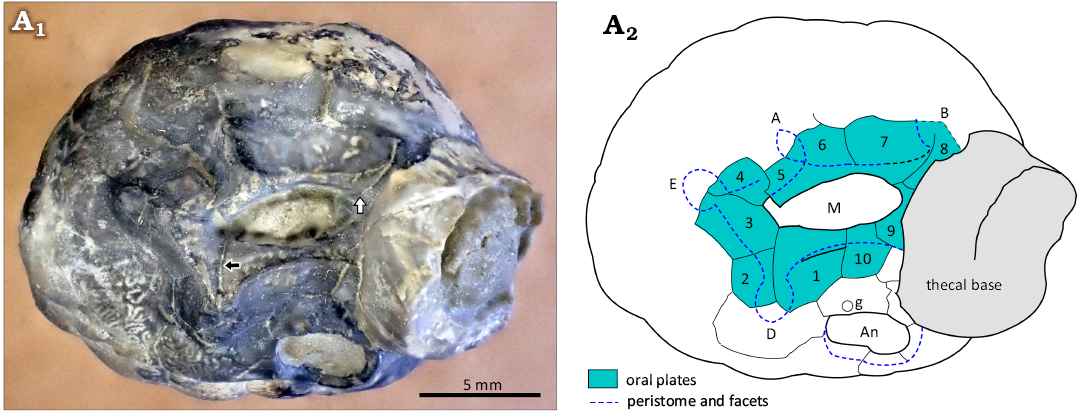
Fig. 7. Holotype of holocystitid blastozoan Brightonicystis gregaria Paul, 1971, CAMSM A.32813, Hirnantian (Upper Ordovician), Knock, Cumbria, England, UK. A1, photograph of specimen in oral view; A2, interpretation of plating. Note the displacement of oral plates 4 and 5; black arrow shows inner limit of oral plate 2, white arrow shows inner limit of oral plate 8. Abbreviations: A, B, D, E, Carpenter’s (1884, 1891) ambulacral designations; An, anus; g, gonopore; M, mouth.
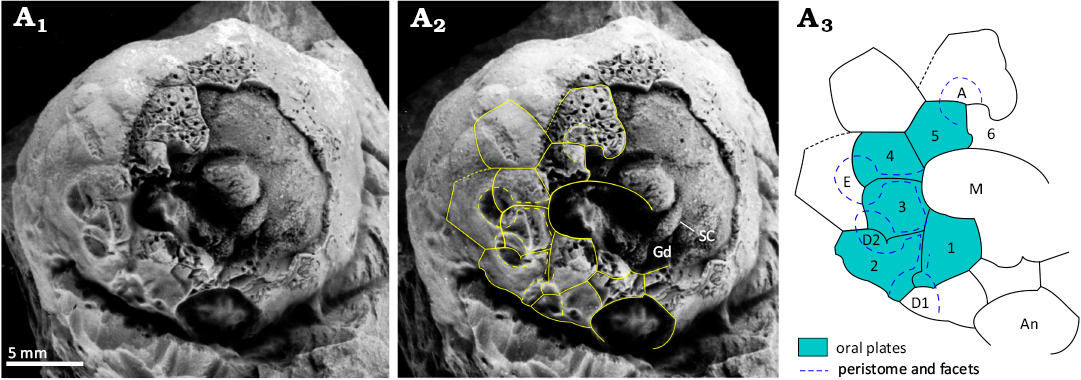
Fig. 8. Paratype of holocystitid blastozoan Brightonicystis gregaria Paul, 1971, CAMSM A.32812, Hirnantian (Upper Ordovician), Knock, Cumbria, England, UK. A1, original photograph of oral area; A2, photograph with outlines of oral plates and orifices superimposed; A3, interpretive diagram of oral plating, not to scale. Note the food groove divides in ambulacrum D, giving rise to two facets, D1 and D2. Abbreviations: A, D1, D2, E, Carpenter’s (1884, 1891) ambulacral designations; An, anus; Gd gonoduct; M mouth; SC stone canal.
Remarks.—Brightonicystis gregaria is characterized by a relatively modest globular theca, with smooth gently convex plates of two generations and humatipores with numerous tangential canals. The mouth is surrounded by at least ten oral plates, two of which in the B and D ambulacra do not reach the inner margin of the peristome. These characteristics are only shared by Holocystites salmoensis Sheffield et al., 2018, with which Brightonicystis gregaria is compared. For further discussion see after the description of Brightonicystis salmoensis.
Stratigraphic and geographic range.—Brightonicystis gregaria comes from the Ashgill Shales (Hirnantian) from several lower Palaeozoic inliers in Cumbria and West Yorkshire, northern England, UK.
Brightonicystis salmoensis (Sheffield et al., 2018) comb. nov.
Figs. 3, 9.
2018 Holocystites salmoensis sp. nov.; Sheffield et al. 2018: 4, fig. 5.
2022 “Holocystites” salmoensis Sheffield, Ausich, and Sumrall; Paul 2022: 470, fig. 1.
Holotype: GSC 126899, an incomplete, weathered theca preserved in original calcite showing the humatipores and oral plating.
Type locality: Road cut along the Salmon-Table Head Road, 800 m east of Macaire Creek, Salmon River Valley, Anticosti Island, Canada.
Type horizon: Ellis Bay Formation, Laframboise Member, Hirnantian, Upper Ordovician.
Material.—Twelve complete or partial thecae, including holotype GSC 126899 and paratypes GSC 126896, 126897, 126898, 126900, 126902, 126903a, 126903b, 126903c, 126906, 126907, 126908.
Emended diagnosis.—A species of Brightonicystis with nine oral plates, relatively large, globular to pyriform theca, two generations of tumid plates with smooth external surfaces; with two-four tangential canals per humatipore (simplified from Sheffield et al. 2018: 4).
Description.—Theca globular to pyriform depending on the relative size of the attachment area, composed of over 100 smooth, convex thecal plates with distinctly impressed sutures, with a pentagonal mouth. Thecal plates of two generations in paratype GSC 126906 (Sheffield et al. 2018: fig. 5E), larger primaries arranged in oblique, apparently closed circlets of about ten plates; smaller secondaries usually pentangular inserted between primary circlets. All plates bear buried humatipores with relatively few tangential canals (Fig. 9A1).
Peristome pentagonal (Fig. 9), surrounding an oval mouth. Peristome border deeply weathered in holotype and paratype GSC 126897 (Fig. 9), but with a frame composed of nine plates arranged as in Fig. 9A3, including two in the B and D ambulacra that do not reach the inner margin of the peristome. Feeding appendages unknown.
The periproct is hexagonal, surrounding a rounded anus in GSC 126897 (Sheffield et al. 2018: fig. 5D). Oral and anal cover plates are unknown. The gonopore is a small circular orifice in the plate immediately adoral to the periproct in GSC 126897 (Sheffield et al. 2018: fig. 5D). A hydropore has not definitely been detected. Attachment was direct but the details of the attachment area are unknown.
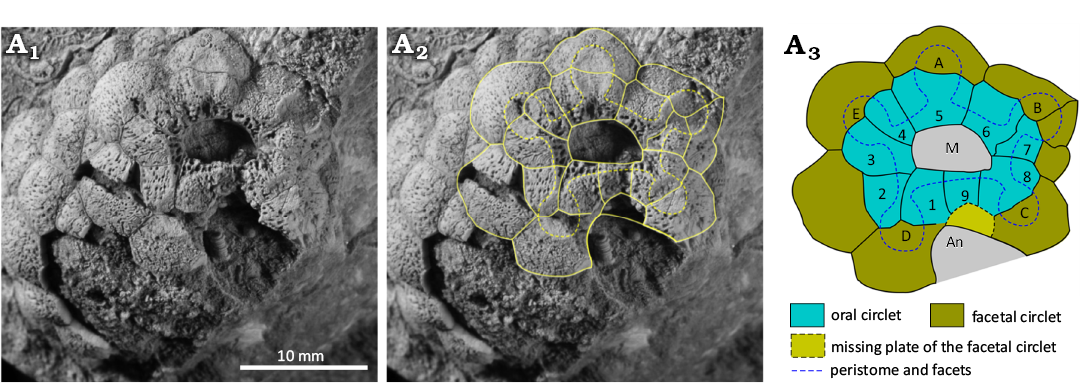
Fig. 9. Holotype of holocystitid blastozoan Holocystites salmoensis Sheffield et al., 2018, GSC 126899, Hirnantian (Upper Ordovician), Anticosti Island, Canada. A1, original photograph of oral area (from Sheffield et al. 2018: 5, fig. 5A); A2, photograph with outlines of oral plates and orifices superimposed; A3, interpretive diagram of oral plating, not to scale. Abbreviations: A–E, Carpenter’s (1884, 1891) ambulacral designations; An, area where anus would be; M, mouth.
Remarks.—Brightonicystis salmoensis is so similar to Brightonicystis gregaria that it is difficult to give a differential diagnosis. It appears to have more tumid thecal plates, arranged in oblique, closed circlets with a larger number of primary plates, and fewer tangential canals per humatipore, than B. gregaria. Both come from similar aged Hirnantian strata but well below the Ordovician/Silurian boundary. Both species attributed to the genus Brightonicystis are characterized by relatively small, globular thecae, composed of two generations of smooth, gently convex plates with many humatipores. The mouth is surrounded by a closed circlet of nine plates in Brightonicystis salmoensis, two of which lying in the B and D ambulacra do not reach the inner margin of the peristome. A second circlet of plates surrounds the oral circlet and five single facets for unknown feeding appendages are shared by plates of both circlets. The critical oral area is only known from a single specimen. Sheffield (2017: 18), Sheffield and Sumrall (2017: 756; 2019: 743) and Sheffield et al. (2018: 2) have expressed doubts as to the validity of the genus Brightonicystis. The oral area of Brightonicystis gregaria is partly known in three specimens. Nevertheless, the details of ambulacrum C are not seen in any of them. Food grooves in the A, B, D and E ambulacra lie along plate sutures and sutures also occur between these ambulacra. If ambulacrum C is the same, the total number of oral plates is eleven, as an additional interradial plate lies in the CD interambulacrum.
The unexpected plate arrangement in which a plate in the B and D ambulacra reaches the edge of the peristome, but not the inner margin of the mouth, can be confirmed in both ambulacra of the holotypes of both Brightonicystis salmoensis and Brightonicystis gregaria. The D ambulacrum of paratype CAMSM A.32812 of Brightonicystis gregaria shows the same arrangement (Fig. 8) but plates of the B ambulacrum are missing. In contrast, the suture in the B ambulacrum of paratype CAMSM A.32811 does not appear to branch (Fig. 6C1). In addition, paratype CAMSM A.32812 has two facets in the D ambulacrum, whereas all other known holocystitids have only a single facet per ambulacrum. Thus, it seems unwise to assume the characters of the unseen facet in ambulacrum C of Brightonicystis gregaria. If all ambulacra are the same, Brightonicystis gregaria has eleven oral plates. If not, the plate arrangement in Brightonicystis gregaria was not fixed, but varied from individual to individual. What is certain, however, is that both Brightonicystis gregaria and Brightonicystis salmoensis have more oral plates and they are more similar to each other than to any other holocystitid genus. The practical solution to this dilemma is to assign them both to the genus Brightonicystis for which a rather unsatisfactory diagnosis of ‘with nine or more orals’ forms part.
The characters discussed above distinguish Brightonicystis from the only other holocystitid genus with ambulacral facets developed on oral and facetal plates, Holocystites. The latter has only six oral plates which have a variable relationship to the overlying facets and food grooves (see below).
Stratigraphic and geographic range.—Brightonicystis salmoensis is only known from the Ellis Bay Formation, Laframboise Member, Hirnantian, Upper Ordovician of NE Anticosti Island, Canada.
Genus Holocystites Hall, 1864
Type species: Caryocystites cylindricus Hall, 1861, by subsequent designation of Miller 1889: 253, from the Racine Dolomite, Sheinwoodian (lower Silurian), Grafton, Wisconsin, USA.
Species included: Type species and Holocystites abnormis Hall, 1864, Holocystites alternatus (Hall, 1861), Holocystites clavus Frest & Strimple in Frest et al., 2011, Holocystites greenevillensis Foerste, 1917, Holocystites ovatus Hall, 1864, Holocystites scutellatus Hall, 1864, Holocystites spangleri Miller, 1891.
Emended diagnosis.—Holocystitids with cylindrical, globular or pyriform thecae, five ambulacral facets shared by oral and facetal plates, six irregular orals with food grooves crossing them, humatipores typically tubercular, but buried beneath smooth plates in at least one species (simplified from Paul 1971: 73).
Remarks.—In Holocystites (Figs. 1A, 10, 11) the theca may be cylindrical, oval or pyriform, often with a small aboral extension. The attachment area is usually small. Thecal plates may be of one to three generations, with primary plates usually in closed circlets of eight plates. In Holocystites alternatus (Hall, 1861) there are circlets of primary, secondary and tertiary plates that alternate regularly in the order primary, tertiary, secondary, tertiary, primary (Fig. 11A). In contrast, Holocystites cylindricus is entirely composed of primary plates (Fig. 11B). The plates usually have tubercular humatipores (Fig. 10A1), but in Holocystites abnormis Hall, 1864, the thecal surface is smooth with buried humatipores. The mouth is surrounded by six oral plates with five concave facets shared by oral and facetal plates (Fig. 10). Food grooves are more often on the oral plates than along the sutures between them, in contrast to all other holocystitid genera in which the food grooves follow oral sutures. Only in the C radius do food grooves cross or lie close to the oral suture. The peristome border is pitted with oral pores and covered by a palate of six cover plates in Holocystites scutellatus Hall, 1864 (CMCIP 75376, Hill 2022: fig. 1.7: 2). Two palatals are preserved in a specimen of Holocystites cylindricus (FMNH 10695, Paul 1971: fig. 25). The oral circlet is surrounded by a facetal circlet of eight plates (Fig. 10A2, A3). The periproct is polygonal, surrounding a relatively large anus, separated from the peristome by three plates (O1, O6, F1, Fig. 10A2, A3). In life, it was covered by a simple anal pyramid, preserved in Holocystites scutellatus (CMCIP 75377, Hill 2022: fig. 1.7: 3). The hydropore is usually a short slit, within an oval tubercle across the O1:O6 suture. The gonopore is a circular pore also set in a tubercle usually in plate F1 or rarely F2.
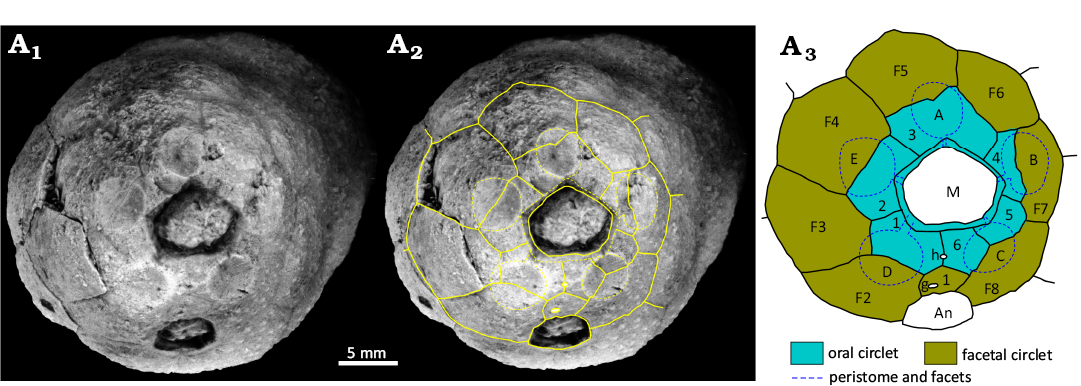
Fig. 10. Holocystitid blastozoan Holocystites cylindricus (Hall, 1861), USNM S3070, Wenlock (Silurian), Newsom, Tennessee, USA. A1, original photograph of oral area; A2, photograph with outlines of oral plates and orifices superimposed; A3, interpretive diagram of oral plating, not to scale. Abbreviations: A–E, Carpenter’s (1884, 1891) ambulacral designations; An, anus; g, gonopore; h, hydropore; M, mouth.

Fig. 11. Plate arrangements in cylindrical species of holocystitid blastozoan Holocystites Hall, 1864, from Racine Dolomite, Sheinwoodian (lower Silurian), Racine (A) and Grafton (B), Wisconsin, USA. A. Holocystites alternatus (Hall, 1861), UWM G20001. B. Holocystites cylindricus (Hall, 1861), lectotype AMNH 2018a.
Among holocystitid genera only Brightonicystis and Holocystites have their ambulacral facets developed over multiple plates of the oral and facetal circlets. Holocystites differs from Brightonicystis in having an oral frame composed of six plates which are irregularly arranged with respect to the food grooves and ambulacral facets (Fig. 10). The key characters of Holocystites are the presence of six oral plates, surrounded by a closed circlet of eight facetal plates and five ambulacral facets shared between orals and facetals but with a variable relationship to the underlying thecal plates. Food grooves cross individual orals. Brightonicystis also has five ambulacral facets developed on both oral and facetal plates, but has a larger number of oral plates and food grooves that usually follow oral sutures. So far, all species of Hirnantian holocystitids (which are currently attributed to Brightonicystis) have buried humatipores beneath smooth thecal plates, whereas Holocystites may have tubercular or buried humatipores (Paul 1971: 73; Frest 1983: 325; Frest et al. 2011: 32 and later). All other genera of holocystitids have ambulacral facets developed exclusively on plates of the facetal circlet and all except Pentacystis and Moyacystis cooperi have food grooves along sutures between orals (see below).
Stratigraphic and geographic range.—Sheinwoodian–Homerian (lower Silurian), Midwest USA (Wisconsin, Illinois, Indiana, Ohio, Kentucky, Tennessee).
Genus Pentacystis Paul, 1971
Type species: Pentacystis simplex Paul, 1971, by original designation, from the Lewisburg and Massie formations, Sheinwoodian (lower Silurian), SE Indiana, USA.
Species included: Type species and Pentacystis gibsoni Frest & Strimple in Frest et al., 2011, Pentacystis sphaeroidalis (Miller, 1891).
Emended diagnosis.—Holocystitids with oral frame composed of eight facetal plates, orals absent (simplified from Paul 1971: 102).
Remarks.—In Pentacystis (Figs. 1B, 12B) the theca is inverted pyriform or clavate with a relatively large attachment area (Frest and Strimple in Frest et al. 2011: 19, fig. 13). Thecal plates are smooth and humatipores buried with as few as two tangential canals. The mouth is rounded and lacks the oral circlet (Fig. 12B). The five ambulacral facets are entirely on individual facetal plates. As there are eight facetals, the facet-bearing plates are grouped in an anterior triplet in contact (ambulacra E, A, and B), separated by plates F2 and F6 from the posterior pair of facets (ambulacra C and D), which in turn are separated from each other by plate F1 (Fig. 12B). Food grooves from the facets pass across the peristome border to the inner edge of the mouth (Fig. 12B). The theca has a relatively large attachment area.
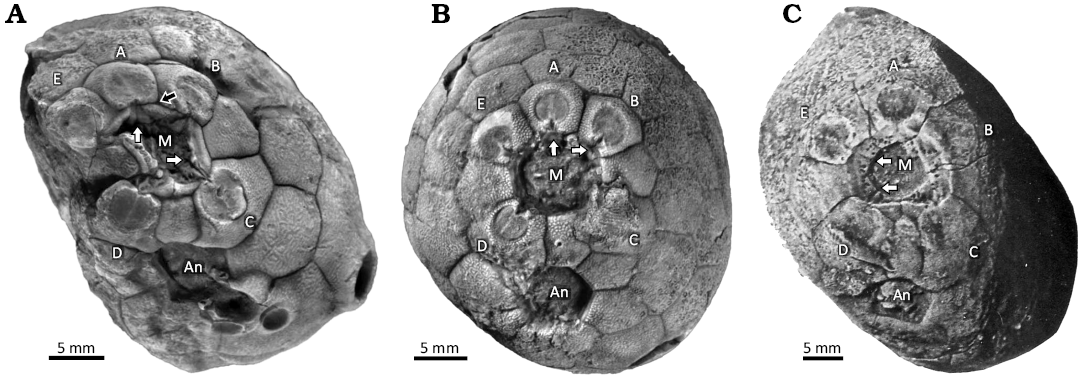
Fig. 12. Photographs of oral areas in holocystitid blastozoans, showing the mouth (M) and anus (An) and the five ambulacral facets (A–E), Massie Formation, Sheinwoodian, Napoleon, Indiana, USA. A. Osgoodicystis bissetti Frest & Strimple in Frest et al., 2011, SUI 48166; black arrow points to the suture between the A ambulacrum facetal and the orals, white arrows point to the oral ends of the A and C food grooves, respectively, both of which follow sutures between orals. B. Pentacystis gibsoni Frest & Strimple in Frest et al., 2011, SUI 46316; white arrows point to the oral ends of the A and B food grooves, respectively, both of which lie entirely on their respective facetals. C. Moyacystis cooperi (Frest & Strimple in Frest et al., 2011), SUI 48167; arrows point to food grooves of the D and E ambulacra. The E food groove crosses the entire peristome margin on a facetal plate. A and B from Sheffield and Sumrall (2017: fig. 6.5 and 6.3, respectively); C from Frest et al. (2011: pl. 14: 1).
Paul (1971: 102) attributed three species to Pentacystis, the type species of which, Pentacystis simplex Paul, 1971, entirely lacks orals. Frest and Strimple (Frest et al. 2011) described three new species of pentacystines and distinguished those with any orals as the genus Osgoodicystis. The type species of Osgoodicystis, Osgoodicystis bissetti has all six orals, but Osgoodicystis cooperi has only the two posterior orals present. It also differs in having oral pores developed in the facetal plates that contribute to the peristome border. To me these are generic characters and so I distinguish Osgoodicystis cooperi as type species of the new genus Moyacystis (see below). Frest (1983: 419) and Frest and Strimple (Frest et al. 2011: 88) clearly considered the same action.
Stratigraphic and geographic range.—Sheinwoodian (lower Silurian), Indiana, USA. Pentacystis ranges from the lower limestone of the Lewisburg Formation into the base of the Massie Formation (Frest et al. 2011: 14, fig. 10; Brett et al. 2012).
Genus Osgoodicystis Frest & Strimple in Frest et al., 2011
Type species: Osgoodicystis bissetti Frest & Strimple in Frest et al., 2011, by original designation, from the Lewisburg and Massie formations, Sheinwoodian (lower Silurian), SE Indiana, USA.
Species included: Type species and Holocystites wykoffi Miller, 1891.
Emended diagnosis.—Holocystitids with six narrow orals; with five food grooves along oral sutures (simplified from Frest and Strimple in Frest et al. 2011: 92).
Remarks.—In Osgoodicystis (Figs. 12A, 13, 14) the thecal outline may be cylindrical or inverted pyriform (Frest and Strimple in Frest et al. 2011: 19, fig. 13). The thecal plates are always smooth with buried humatipores. The mouth is rounded with a reduced oral circlet (Fig. 13) even though all six oral plates are present. Oral pores are present in the oral plates (Fig. 13A1). The five ambulacral facets are confined to individual facetal plates and arranged in the same manner as in Pentacystis. The food grooves pass along the sutures between orals (Fig. 14). The theca has a relatively large attachment area.
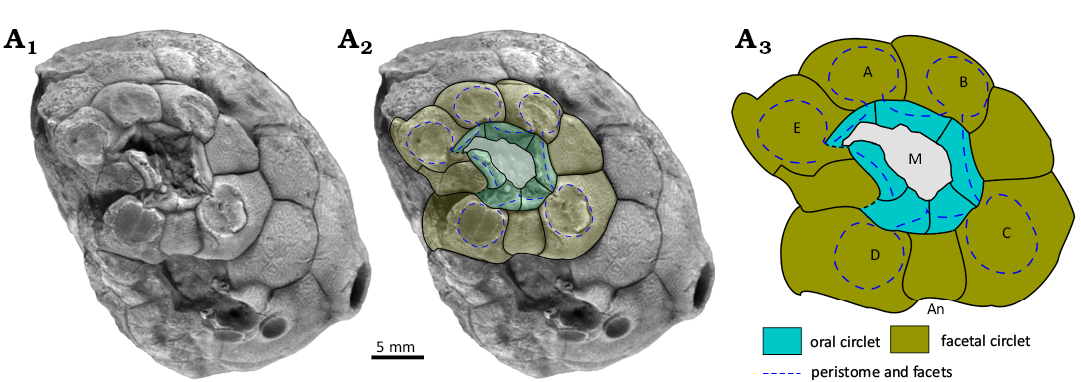
Fig. 13. Oral plating in holocystitid blastozoan Osgoodicystis bissetti Frest & Strimple in Frest et al., 2011, SUI 48166, Massie Formation, Sheinwoodian (lower Silurian), Napoleon, Indiana, USA. A1, original photograph of holotype (from Frest et al. 2011: pl. 12: 1); A2, photograph with outlines of plates, facets and food grooves superimposed; A3, interpretive diagram, not to scale. Abbreviations: A–E, Carpenter’s (1884, 1891) ambulacral designations; An, anus; M, mouth.

Fig. 14. Facet of ambulacrum C in holocystitid blastozoan Osgoodicystis bissetti Frest & Strimple in Frest et al., 2011, SUI 48166, Massie Formation, Sheinwoodian (lower Silurian), Napoleon, Indiana, USA. A1, original photograph (from Frest et al. 2011: pl. 4: 5); A2, photograph with outlines of plates, facet and food grooves superimposed; A3, interpretive diagram. Abbreviations: C, Carpenter’s (1884, 1891) ambulacral designation; FG, food groove from facet to mouth (M); O5 and O6, oral plates 5 and 6; rings in O6 within peristome, oral pores; PO, perioral plates. Note that the food groove follows the suture between the orals.
Stratigraphic and geographic range.—Sheinwoodian (lower Silurian), Indiana, USA. Osgoodicystis ranges from the lower limestone of the Lewisburg Formation into the base of the Massie Formation (Frest et al. 2011: 14, fig. 10; Brett et al. 2012).
Genus Moyacystis nov.
ZooBank LSID: urn:lsid:zoobank.org:act:2091DCE0-0B58-47FD-8823-5E02DF990FFE.
Etymology: Named for my wife Moya and the standard ending cystis.
Type species: Osgoodicystis cooperi Frest & Strimple in Frest et al., 2011, monotypic; Lewisburg Formation, Sheinwoodian (lower Silurian), Ripley County, Indiana, USA.
Species included: Type species only.
Diagnosis.— Holocystitids with only the two posterior orals (O1 and O6), with oral pores in facetals that contribute to the peristome margin.
Remarks.—Moyacystis gen. nov. (Figs. 12C, 15) has a large more or less cylindrical theca, composed of granular plates of a single generation arranged in seven or more circlets of 10–14 plates each. The mouth frame is composed of the two posterior orals (O1 and O6) plus at least parts of all the facetals except F1 (Fig. 15A3). Oral pores are present all around the peristome border (Fig. 15A1). Humatipores are rounded with few (2–3) tangential canals. The gonopore is in a low tubercle in F2. Hydropore and attachment area unknown.
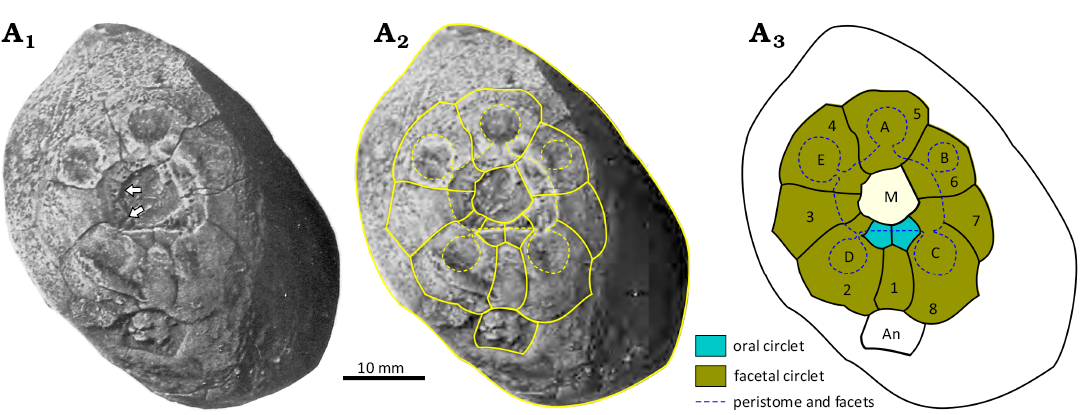
Fig. 15. Oral plating in holocystitid blastozoan Moyacystis cooperi (Frest & Strimple in Frest et al., 2011), SUI 48167, Massie Formation, Sheinwoodian (lower Silurian), Napoleon, Indiana, USA. A1, original photograph of oral area of the holotype, arrows point to the food grooves in ambulacra D and E which pass right across the peristome border to the edge of the mouth (from Frest et al. 2011: pl. 14: 1); A2, photograph with outlines of oral plates and orifices superimposed; A3, interpretive diagram of oral plating. Abbreviations: A–E, Carpenter’s (1884, 1891) ambulacral designation; An, anus; M, mouth.
Moyacystis gen. nov., Osgoodicystis Frest & Strimple in Frest et al., 2011, and Pentacystis Paul, 1971, are all very similar, differing only in the presence of some or all the oral plates. Moyacystis cooperi has a more or less cylindrical theca, but otherwise even the thecal shapes of species in all three genera are similar. Sheffield and Sumrall (2015: 161, fig. 1B, C; 2017: 758, fig. 3) have argued that the variable number of oral plates seen in the three pentacystine genera is purely taphonomic. They considered that well-preserved examples have all the orals, as in Osgoodicystis wykoffi (Miller, 1891) and Osgoodicystis bissetti Frest & Strimple in Frest et al., 2011, whereas more disarticulated examples have only two orals, as in Moyacystis cooperi (Frest & Strimple in Frest et al., 2011), or none at all, as in Pentacystis sphaeroidalis (Miller & Gurley, 1895), Pentacystis simplex Paul, 1971, and Pentacystis gibsoni Frest & Strimple in Frest et al., 2011. The diagram to support this idea (Sheffield and Sumrall 2015: fig. 1b, c) implies a complex and unlikely taphonomic history. The orals had to remain in place while the theca filled completely with sediment. Then they detached and moved across the mouth to accumulate on one side on top of the sediment fill.
This is not the only point against their interpretation. To illustrate their interpretation, Sheffield and Sumrall (2017: fig. 6.5) figured Osgoodicystis bissetti, with all six orals preserved and Pentacystis gibsoni, without any orals (their fig. 6.3). The orals of Osgoodicystis bissetti (Sheffield and Sumrall 2017: fig. 6.5) include the inner margin of the mouth, the peristome border (which in plate O2 clearly bears oral pores; Fig. 13A1), food grooves running along the sutures between the oral plates (clearly seen in ambulacrum C; Fig. 14A1), and the outer peristome border with a recess for the insertion of the cover plates (Figs. 12A, 14A3). Under Sheffield and Sumrall’s (2015) hypothesis, all these features should be missing in Pentacystis gibsoni because they are all developed on the (missing) orals. The oral opening would appear as an angular, roughly hexagonal hole defined by the outline of the oral plate circlet (Fig. 16).
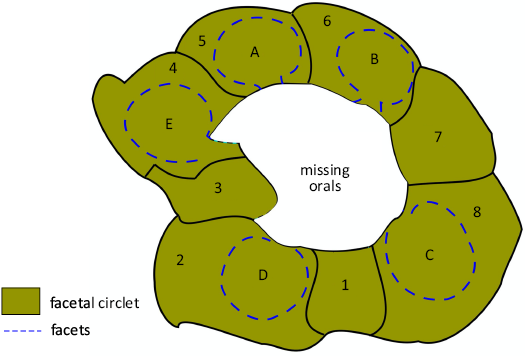
Fig. 16. Interpretive diagram of how the oral area of holocystitid blastozoan Osgoodicystis bissetti Frest & Strimple in Frest et al., 2011, would look if the entire oral circlet were missing. Note no peristome border, oral pores or food grooves are present because they are developed entirely on the orals. The facetals (1–8) would surround the oral opening truncated by the missing orals.
Sheffield and Sumrall (2017: 764) argued that the orals of Pentacystis gibsoni detached prior to burial, as the oral cover plates undoubtedly have. Their photograph of Pentacystis gibsoni (Sheffield and Sumrall 2017: fig. 6.3) shows parts of the rounded inner margin of the mouth, a clear peristome border across which food grooves pass from the adoral edge of the facets to the inner margin of the mouth, and an outer peristome border with a recess for the insertion of cover plates (Fig. 12B). In short, all the features that should be missing except the oral pores in the peristome border. Frest et al. (2011: pl. 14: 1) show the oral area of Moyacystis cooperi (Frest & Strimple in Frest et al., 2011) which has oral pores in the peristome border formed by the facetal plates (Fig. 12C). Thus, in different pentacystine species with two or no oral plates, all the features that should be missing under Sheffield and Sumrall’s (2015) hypothesis are preserved. Altogether, I prefer to accept Frest’s (1983) interpretation of pentacystine holocystitids, reinstate Osgoodicystis and recognize the species with only two orals as a new genus, Moyacystis.
The remaining holocystitid genera have four ambulacra except for the unique type specimen of Pustulocystis pentax Paul, 1971 (Fig. 17B) which has five. Trematocystis, and Paulicystis have six orals, eight facetals and buried humatipores but Paulicystis has recumbent ambulacra. Pustulocystis has only six facetals and typically tubercular humatipores (Table 1).
Stratigraphic and geographic range.—Lewisburg Formation, Sheinwoodian (lower Silurian), Ripley County, Indiana, USA.
Genus Trematocystis Jaekel, 1899
= Austrocystites Brown, 1963
Type species: Holocystites subglobosus Miller, 1889 (= Holocystites globosus Miller, 1878), by original designation, from the Osgood Formation, Sheinwoodian (lower Silurian), Jefferson County, Indiana, USA.
Species included: Type species and Trematocystis branagani (Brown, 1963), Trematocystis magniporatus Frest & Strimple in Frest et al., 2010, Trematocystis rotundus (Miller, 1879), Trematocystis wetherbyi (Miller, 1878)
Emended diagnosis.—Holocystitids with six inter-radial orals and eight facetals, four ambulacral facets confined to a single facetal plate, food grooves along O:O sutures, humatipores buried beneath smooth external plate surface (after Paul 1971: 115).
Remarks.—In Trematocystis (Figs. 1C, 4A) the theca is inverted pyriform, with a relatively large attachment area and smooth thecal surface composed of only primary plates. The oral area is characterized by six interradial orals, surrounded by eight facetals only four of which bear ambulacral facets confined to a single facetal plate (Figs. 1C, 4A). Ambulacrum A of Carpenter (1884, 1891) always lacks a facet. USNM S3063a preserves the six palatal plates over the mouth and five anals over the periproct (Fig. 4A). A small oval hydropore is shared by O1 and O6; a small circular gonopore may occur in either F1 or F2 (Figs. 1C, 4A). Buried humatipores have 4–6 tangential canals. Trematocystis wetherbyi (Miller, 1878) has a theca with five circlets of 14–16 primary plates. All other species, including the Australian one, have eight or nine plates per circlet.
Species of Trematocystis are distinguished on the position of the gonopore and details of the humatipores. Brown (1963: 388) described a new diploporite, Austrocystites branagani Brown, 1963, on a unique internal mould from the Silurian of Yass, New South Wales. The specimen showed a single generation of relatively large plates, arranged in three circlets up to the level of the periproct (Brown 1963: 389, fig. 2), the lower two circlets with eight plates each. All the plates bore casts of perpendicular canals of pore structures not arranged in rhombic patterns and with numerous pores in each plate. Brown correctly interpreted the specimen as a diploporite, but the internal mould lacked diagnostic characters on which to make a more precise identification. Brown thought Austrocystites was allied to the sphaeronitid diploporite Eucystis Angelin, 1878.
More recently, Jell (2010: 35) described and illustrated a second internal mould as Eucystis branagani (Brown, 1963). This specimen clearly shows that the casts of the perpendicular canals are arranged in pairs (Jell 2010: 36, fig. 6e). A latex cast of a small area of the external mould (Jell 2010: fig. 6d) shows an entirely smooth external surface to parts of three plates in the second circlet up from the base of the theca. In typical diplopores the perpendicular canals pass right through the thecal plates and open in pits in the external surface called peripores (Paul 1971: 19, fig. 6a; 1972: 6–7, figs. 6, 7). Jell (2010: 37) explained the absence of any trace of the peripores on the external surface around the supposed diplopores as being due to wear or weathering, but no amount of wear would eliminate the perpendicular canals of diplopores. Alternatively, if the pore structures were humatipores buried beneath smooth thecal plates, a cast of the inner surface would show paired pits (or tubercles on an internal mould), but an undamaged external surface would show no trace of pores (Paul 1971: fig. 6b; 1972: 8, fig. 9b).
Immediately following his description of Eucystis branagani, Jell (2010: 37) described the holocystitid Trematocystis wrighti Jell, 2010, which not only bears humatipores beneath smooth thecal plates, but also seems to have the thecal plates arranged in circlets of eight, as do many American holocystitids. The morphological similarity, plus the same locality and horizon suggest to me that Austrocystites is not a sphaeronitid, but a holocystitid and a junior synonym of Trematocystis. Thus, the correct name should be Trematocystis branagani (Brown, 1963). Peter Jell (personal communication 2025) agrees with this interpretation.
Stratigraphic and geographic range.—Sheinwoodian (lower Silurian), Indiana, USA; uppermost Ludfordian (upper Silurian), Yass Basin, New South Wales, Australia. Trematocystis in the USA ranges from the upper Osgood Formation into the base of the Massie Formation (Frest et al. 2011: 14, fig. 10; Brett et al. 2012) in Indiana, and in Jefferson County, Kentucky, just across the Ohio River. It is also recorded in Australia from the Rainbow Hill Marl Member of the Rosebank Shale Formation, uppermost Ludlovian (Jell 2010). This record is the youngest known occurrence of any holocystitid.
Genus Pustulocystis Paul, 1971
Type species: Holocystites ornatissimus Miller, 1891, by original designation, from the Osgood Formation, Sheinwoodian (lower Silurian), Jefferson County, Indiana, USA.
Emended diagnosis.— Holocystitids with six interradial oral plates and six facetals, four or rarely five ambulacral facets confined to a single facetal plate, food grooves along O:O sutures, humatipores typically in raised tubercles (modified after Paul 1971: 131).
Species included: Type species and Pustulocystis pentax Paul, 1971.
Remarks.—Pustulocystis (Figs. 1D, 17, 18) has a fusiform to globular theca composed of a single generation of distinctly pustular plates (Fig. 18) . The oval mouth is surrounded by the usual six interradial orals, with a closed circlet of six facetals beyond, four of which bear facets (Fig. 17A). The hydropore is in a tubercle across the O1:O6 suture and the gonopore in a rounded tubercle in variable positions usually across a suture between facetals, or facetal and oral plates and close to the D ray facet (Fig. 17). The periproct is rounded. Cover plates of both mouth and anus are unknown. Attachment was direct, by a relatively small attachment area compared with Trematocystis. The most distinctive feature of Pustulocystis is the prickly appearance of the theca due to the prominent tubercular humatipores (Fig. 18).
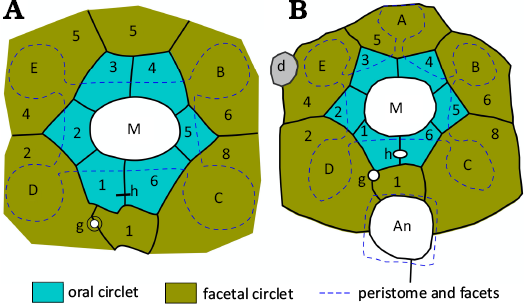
Fig. 17. Interpretive diagrams of oral plating in holocystitid blastozoans Pustulocystis ornatissimus (Miller, 1891), AMNH 20273 (A) and Pustulocystis pentax Paul, 1971, MUMG-T 226 (B). Note only six facetal plates occur. Abbreviations: A–E, Carpenter’s (1884, 1891) ambulacral designation; An, anus; d, damaged area; g, gonopore; h, hydropore; M, mouth. Redrawn, coloured, and relabelled from Paul (1971: 137, fig. 58a) and Paul (1971: 140, fig. 60), respectively. Not to scale.

Fig. 18. Lateral view of the holotype of holocystitid blastozoan Pustulocystis ornatissimus (Miller, 1891), FMNH 8760, Osgood Formation, Sheinwoodian (lower Silurian), Jefferson County, Indiana, USA, showing the thecal outline, circlets of many primary plates and the characteristic pustular ornament enclosing the humatipores which are visible on damaged pustules.
All but one specimen attributed to Pustulocystis bear only four ambulacral facets. The single exception, the holotype of Pustulocystis pentax Paul, 1971, has five (Fig. 17B). With only one example it is impossible to tell whether this is an anomalous specimen or a distinct species. Paul (1971: 140) opted for the latter because its thecal shape was unknown in the more common Pustulocystis ornatissimus (Miller, 1891) and because it came from an uncertain stratigraphic horizon at Newsom, Tennessee, over 300 km from all other known examples in Indiana. A third possible species was left under open nomenclature. It had a thecal shape closer to Pustulocystis ornatissimus, but smooth plates and apparently seven facetals; F7 is definitely absent, but the oral area is not well preserved.
Stratigraphic and geographic range.—Sheinwoodian (lower Silurian), SE Indiana, Jefferson County, Kentucky, and Pegram and Newsom, Tennessee, USA. In Indiana, Pustulocystis ornatissimus has only been recorded from Foerste’s (1897) “main cystoid bed” 23–37 cm below the top of the Osgood Formation as restricted by Brett et al. (2012).
Genus Paulicystis Frest & Strimple in Frest et al., 2011
Type species: Paulicystis densus Frest & Strimple in Frest et al., 2011, by original designation, from the Massie Formation, Sheinwoodian (lower Silurian), Napoleon, Ripley County, Indiana, USA.
Species included: Type species and Paulicystis sparsus Frest & Strimple in Frest et al., 2011.
Emended diagnosis.—Holocystitids with six inter-radial orals and eight facetals, four facets for recumbent ambulacra that extend below the facetal circlet, food grooves along O:O sutures, humatipores below smooth surface of tumid thecal plates in circlets of up to 20 plates, attachment area large (simplified from Frest et al. 2011: 83).
Remarks.—Paulicystis (Figs. 19, 20) has a barrel-shaped theca with a large attachment area, several circlets of smooth tumid plates with deep sutures below the surface of which the humatipores lie (Fig. 19A1). The rounded mouth is surrounded by the usual six interradial orals, with a facetal circlet of eight plates around that. Four of the facetals bear the adoral ends of elongate concave grooves that extend radially beyond the facetal circlet (Fig. 19). The grooves are divided along their length and cross-wise (Fig. 20) indicating that the attached ambulacra were biserial. An oval hydropore lies across the O1:O6 suture and a circular gonopore in plate F1 above the rounded anus (Fig. 19A3). Humatipores are well-developed with four to six tangential canals. According to Frest’s diagrams (Frest 1983: 525, fig. 77C, D; Frest et al. 2011: 81, fig. 45), pairs of perpendicular canals from different humatipores are united by the same set of tangential canals. The attachment area appears to be characterized by a central heptagonal plate, surrounded by a closed circlet of seven basals (Frest 1983: pl. 10: 9; Frest et al. 2011: pl. 20: 9; Thomka and Brett 2014a: 182, fig. 4).
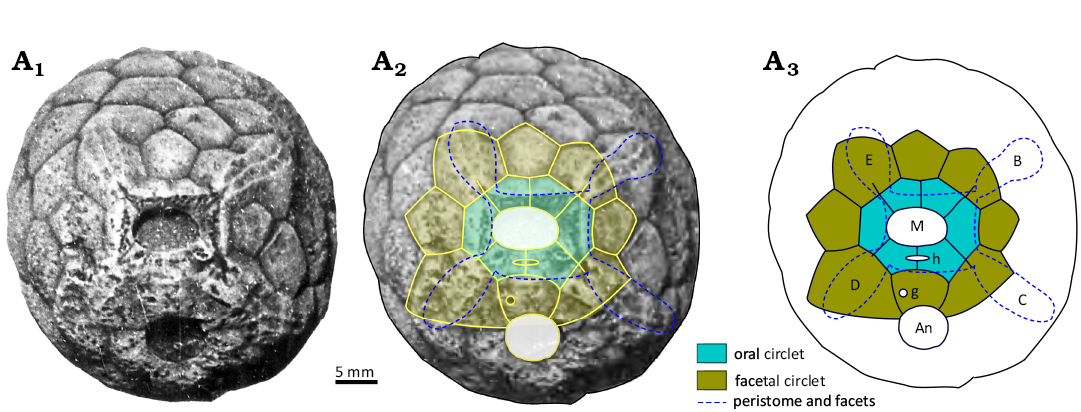
Fig. 19. The oral area of the holotype holocystitid blastozoan Paulicystis sparsus Frest & Strimple in Frest et al., 2011, SUI 48164, Massie Formation, Sheinwoodian (lower Silurian), Napoleon, Indiana, USA. A1, original photograph (from Frest et al. 2011: pl. 11: 1); A2, photograph with interpretation of plating superimposed; A3, interpretive diagram of oral plating. Abbreviations: B–E, Carpenter’s (1884, 1891) ambulacral designation; An, anus; g, gonopore; h, hydropore; M mouth.
Paulicystis (Figs. 19, 20) is unique among holocystitid genera in having four ambulacra that extended radially over the theca (Fig. 19A1). The ambulacral channels are biserial (Fig. 20), so it is assumed that the ambulacra were also. Holocystitid ambulacral appendages have never been preserved, so it is unknown if the recumbent ambulacral gave rise to erect brachioles. Nevertheless, this feature alone is sufficient to distinguish Paulicystis from other holocystitid genera.
Stratigraphic and geographic range.—Sheinwoodian (lower Silurian), Indiana, USA, where Paulicystis has only been recorded from the base of the Massie Formation as restricted by Brett et al. 2012. Thecal bases occur on a hardground surface capping the Lewisburg Formation (Thomka and Brett 2014a, b, 2015), and detached specimens from the lower part of the overlying Massie Formation.
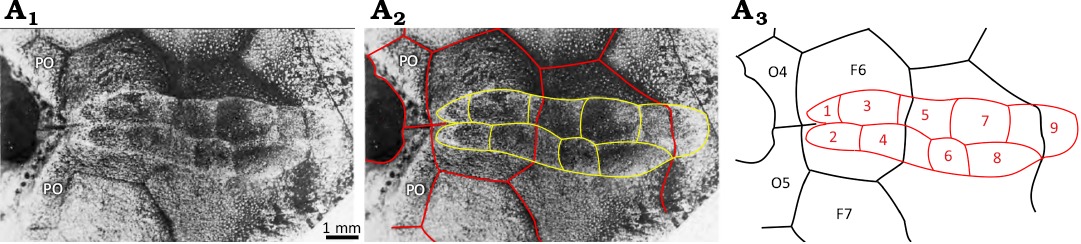
Fig. 20. Facet in ambulacrum B of the holotype holocystitid blastozoan Paulicystis sparsus Frest & Strimple in Frest et al., 2011, SUI 48164, Massie Formation, Sheinwoodian (lower Silurian), Napoleon, Indiana, USA. A1, original photograph (from Frest et al. 2011: pl. 4: 2); A2, photograph with outlines of thecal plates and pits in the facet superimposed; A3, interpretive diagram. Abbreviations: F6, F7, facetals; O4, O5, orals; 1–9 impressions of ambulacral plates in the facet.
Discussion
The family Holocystitidae includes the genera Brightonicystis, Holocystites, Pentacystis, Osgoodicystis, Moyacystis gen. nov., Trematocystis, Pustulocystis, and Paulicystis. They can be distinguished on the number of ambulacral facets (four or five), whether the facets are developed on single or multiple thecal plates, the number of orals, and the number of facetals (eight or six). In addition, Brightonicystis, Pentacystis, Moyacystis gen. nov., Osgoodicystis, Trematocystis, and Paulicystis are characterized by humatipores buried beneath smooth thecal plates. Pustulocystis typically has three-dimensional humatipores raised in distinct tubercles on pustular plates (Fig. 18), although a single specimen provisionally assigned to the genus has a smooth theca. Most species of Holocystites have pustular humatipores but some are smooth, so this character would seem to have evolved independently repeatedly.
Most Silurian holocystitid genera have ambulacral facets developed on single facetal plates. Holocystites, and the Ordovician genus Brightonicystis have facets shared by at least two plates from the oral and facetal circlets. This fact alone indicates that holocystitid facet-bearing plates are not part of the ambulacral system, but thecal plates co-opted to support (unknown) erect feeding structures. In terms of the Extraxial Axial Theory (EAT; Mooi and David 1997; David and Mooi 1998), facet-bearing plates are part of perforate extraxial skeleton. Holocystites oral plating appears to be more variable because the thecae were unstable, frequently fell over but continued to grow, with the erect feeding structures re-orientated to grow up into the water column. Settling the detailed homologies of holocystitid oral plating requires review of at least the oral plating in the Aristocystitidae, Parasphaeronitidae, and Sphaeronitidae which is beyond the scope of this paper. Nevertheless, if as contended here, the oral plating of holocystitids is perforate extraxial, addition of extra plates in the oral area would be expected.
Conclusions
Brightonicystis is a valid genus characterized by nine or more oral plates, with two in the B and D ambulacra not reaching the inner edge of the mouth; five ambulacral facets shared by oral and facetal plates; two generations of smooth thecal plates with buried humatipores. Brightonicystis contains two named species, Brightonicystis gregaria and Brightonicystis salmoensis, with other unnamed taxa known from Canada and Sweden, all of which are Hirnantian.
A review of the Holocystitidae confirms the established genera Holocystites and Brightonicystis both characterised by five ambulacra with shared facets, as well as Trematocystis, Pustulocystis, and Paulicystis all characterised by four ambulacra. Pentacystis, Frest and Strimple’s (in Frest et al. 2011) proposed genus Osgoodicystis and Moyacystis gen. nov. are also considered valid. They are characterized by reduced or absent orals and five ambulacral facets developed on single facetal plates. The differences are not due to taphonomy. Moyacystis gen. nov., type species Osgoodicystis cooperi Frest & Strimple in Frest et al., 2011, is characterized by only two orals (O1 and O6) and facetal plates with oral pores in the peristome border.
Acknowledgements
I am grateful to Matthew Riley (CAMSM) for access to the original specimens of Brightonicystis gregaria, and my daughters Julia and Kathryn Paul for help in getting me to Cambridge. Bertrand Lefebvre (Université de Lyon 1, France) and Martina Nohejlova (Czech Geological Survey, Prague, Czech Republic), provided helpful reviews that improved the original manuscript.
Editor: Andrzej Kaim.
References
Angelin, N.P. 1878. Iconographia crinoideorum in stratis sueciae siluricis fossilium. 62 pp. Samson and Wallin, Holmiae.
Bather, F.A. 1899. A phylogenetic classification of the Pelmatozoa. Report of the British Association for the Advancement of Science 68: 916–923.
Bockelie, J.F. 1984. The Diploporita of the Oslo Region, Norway. Palaeontology 27: 1–68.
Brett, C. E., Cramer, B.D., McLaughlin, P.I., Kleffner, M.A., Showers, W.J., and Thomka, J.R. 2012. Revised Telychian–Sheinwoodian (Silurian) stratigraphy of the Laurentian mid-continent: building uniform nomenclature along the Cincinnati Arch. Bulletin of Geosciences 87: 733–753. Crossref
Brown, I.A. 1963. A new cystoid (Pelmatozoa, Echinodermata) from the Silurian of New South Wales. Journal and Proceedings of the Linnean Society of New South Wales 88: 386–391.
Carpenter, P.H. 1884. Report upon the Crinoidea collected during the voyage of HMS Challenger during the years 1873–76, part 1. General morphology with descriptions of the stalked crinoids. Reports of the Scientific Results of the Voyage of HMS Challenger, Zoology 11: 1–442. Crossref
Carpenter, P.H. 1891. On certain points of the morphology of the Cystidea. Journal of the Linnean Society (Zoology) 34: 1–52. Crossref
David, B. and Mooi, R. 1998. Major events in the evolution of echinoderms viewed in the light of embryology. In: R. Mooi and M. Telford (eds.), Echinoderms, San Francisco, 21–28. Balkema, Rotterdam.
Foerste, A.F. 1897. A report on the geology of the middle and upper Silurian rocks of Clark, Jefferson, Ripley, Jennings and southern Decatur counties, Ind. Report of the Indiana Department of Geology and Natural Resources 21: 213–288.
Foerste, A.F. 1917. Notes on fossils from Ohio and other central states. Ohio Journal of Science 17: 187–204.
Frest, T.J. 1983. Studies of Silurian echinoderms. Chapter VII. Systematics and paleocecology of new Holocystites fauna taxa (Echinodermata, Blastozoa: Diploporita) from the Wenlock of Indiana. 602 pp. Unpublished Ph.D. Thesis, University of Iowa, Iowa City.
Frest, T.J., Strimple, H.L., and Paul, C.R.C. 2011. The North American Holocystites fauna (Echinodermata: Blastozoa: Diploporita): paleobiology and systematics. Bulletins of American Paleontology 380: 1–141.
Gutiérrez-Marco, J.C. 2000. Revisión taxonómica de “Echinosphaerites” murchisoni Verneuil y Barrande, 1855 (Echinodermata, Diploporita) del Ordovícico Medio centroibérico (España). Geogaceta 27: 83–86.
Hall, J. 1861. Descriptions of new species of fossils; from the investigations of the survey. In: Report of the Superintendent of the Geological Survey [of Wisconsin] Exhibiting the Progress of the Work, 9–52. Galkins & Sons, Madison, Wisconsin.
Hall, J. 1864. Account of some new or little known species of fossils from rocks of the age of the Niagara Group. Report of the New York State Cabinet of Natural History 18: 1–16.
Hill, S.A. 2022. Functional Morphology and Taphonomy of Ordovician–Silurian Diploporan Echinoderms. 58 pp. Unpublished M.Sc. Thesis, University of South Florida, Tampa.
Ingham, J. K. 1966. The Ordovician rocks in the Cautley and Dent districts of Westmorland and Yorkshire. Proceedings of the Yorkshire Geological Society 35: 455–505. Crossref
Jaekel, O. 1899. Stammesgeschichte der Pelmatozoen. 1. Thecoidea und Cystoidea. x + 442 pp. Julius Springer, Berlin. Crossref
Jefferies, R.P.S., Joysey, K.A., Paul, C.R.C., and Ramsbottom, W.H.C. 1967. Cyclocystoidea, Eocrinoidea, Rhombifera, Diploporita and Paracrinoidea. In: W.B. Harland, C.H. Holland, M.R. House, N.F. Hughes, A.B. Reynolds, M.J.S. Rudwick, G.E. Satterthwaite, L.B.H. Tarlo, and E.C. Willey (eds.), The Fossil Record, 566–570. Geological Society, London.
Jell, P.A. 2010. Late Silurian echinoderms from the Yass Basin, New South Wales—the earliest holothurian body fossil and two diploporitan cystoids (Sphaeronitidae and Holocystitidae). Memoirs of the Association of Australasian Palaeontologists 39: 27–41.
Kammer, T.W., Sumrall, C.D., Zamora, S., Ausich, W.I., and Deline, B. 2013. Oral region homologies in Paleozoic crinoids and other plesiomorphic pentaradial echinoderms: PLOS ONE 8 (11): e77989. Crossref
Leuckart, R. 1854. Bericht über die Leistungen in der naturgeschichte der niederen Thiere während der Jahre 1848–1853. Archiv für Naturgeschichte 20: 289–473.
Miller, S.A. 1878. Descriptions of eight new species of Holocystites from the Niagara group. Journal of the Cincinnati Society of Natural History 1: 129–136.
Miller, S.A. 1879. Descriptions of twelve new fossil species and remarks upon others. Journal of the Cincinnati Society of Natural History 2: 104–118.
Miller, S.A. 1889. North American Geology and Palaeontology. 664 pp. Western Methodist Book Concern, Cincinnati.
Miller, S.A. 1891. Advance Sheets from the Seventeenth Report of the Geological Survey of Indiana, Palaeontology. 94 pp. Published by the author, Indianapolis.
Miller, S.A. and Gurley, W.F.E. 1895. Description of new species of Paleozoic Echinodermata. Bulletin of the Illinois State Museum 6: 1–62. Crossref
Mooi, R. and David, B. 1997. Skeletal homologies of echinoderms. In: J.A. Waters and C.G. Maples (eds.), Geobiology of Echinoderms. Paleontological Society Papers 3: 305–335. Crossref
Müller, J. 1854. Über den Bau der Echinodermen. Abhandlungen der königlichen preussischen Akademie der Wissenschaften 1854: 123–219.
Neumayr, M. 1889. Die Stämme des Thierreiches. 1. Wirbellose Thiere. 603 pp. Tempsky, Vienna.
Paul, C.R.C. 1967. New records of cystoids from Britain. Proceedings of the Geological Society of London 1967: 147.
Paul, C.R.C. 1971. Revision of the Holocystites fauna (Diploporita) of North America. Fieldiana: Geology 24: 1–166. Crossref
Paul, C.R.C. 1972. Morphology and function of exothecal pore-structures in cystoids. Palaeontology 15: 1–28.
Paul, C.R.C. 1973. British Ordovician cystoids. Part 1, Diploporita. Monographs of the Palaeontographical Society 127: 1–64. Crossref
Paul, C.R.C. 2017. Testing for homologies in the axial skeleton of primitive echinoderms. Journal of Paleontology 91: 582–603. Crossref
Paul, C.R.C. 2022. Reply to Bauer et al. (2022). Acta Palaeontologica Polonica 67: 469–473. Crossref
Paul, C.R.C. and Gutiérrez-Marco, J.-C. 2022. Pentaradiate diploporites (Echinodermata) from the Spanish Middle Ordovician and their taxonomic significance. Spanish Journal of Palaeontology 37: 157–176. Crossref
Paul, C.R.C. and Toom, U. 2021. The diploporite blastozoan Glyptosphaerites (Echinodermata: Blastozoa) and the origin of diplopores. Estonian Journal of Earth Sciences 70: 224–239. Crossref
Sheffield, S.L. 2017. The Homology and Phylogeny of the Diploporita (Blastozoa: Echinodermata). 173 pp. Unpublished Ph.D. Thesis, University of Tennessee, Knoxville.
Sheffield, S.L. and Sumrall, C.D. 2015. A new interpretation of oral plating patterns of the Holocystites fauna (Diploporita, Echinodermata), In: S. Zamora and I. Rábano (eds.), Progress in Echinoderm Palaeobiology, 159–162, Instituto Geológico y Minero de España, Madrid.
Sheffield, S.L. and Sumrall, C.D. 2017. Generic revision of the Holocystitidae of North America (Diploporita, Echinodermata) based on universal elemental homology. Journal of Paleontology 91: 755–766. Crossref
Sheffield, S.L. and Sumrall, C.D. 2019. The phylogeny of the Diploporita: a polyphyletic assemblage of blastozoan echinoderms. Journal of Paleontology 93: 740–752. Crossref
Sheffield, S.L., Ausich, W.I., and Sumrall, C.D. 2018. Late Ordovician (Hirnantian) diploporitan fauna of Anticosti Island, Canada: implications for evolutionary and biogeographic patterns. Canadian Journal of Earth Sciences 55: 1–7. Crossref
Sprinkle, J. 1973. Morphology and Evolution of Blastozoan Echinoderms Special Publication. 284 pp. Museum of Comparative Zoology, Harvard University, Harvard. Crossref
Sumrall, C.D. 2008. The origin of Lovén’s law in glyptocystitoid rhombiferans and its bearing on the plate homology and heterochronic evolution of the hemicosmitoid peristomial border. In: W.I. Ausich and G.D. Webster (eds.), Echinoderm Paleobiology, 228–241. Indiana University Press, Bloomington, Indiana.
Sumrall, C.D. and Waters, J.A. 2012. Universal elemental homology in glyptocystitoids, hemicosmitoids, coronoids and blastoids: steps towards echinoderm phylogenetic reconstruction in derived Blastozoa. Journal of Paleontology 86: 956–972. Crossref
Thomka, J.R. and Brett, C.E. 2014a. Diploporite (Echinodermata, Blastozoa) thecal attachment structures from the Silurian of southeastern Indiana. Journal of Paleontology 88: 179–186. Crossref
Thomka, J.R. and Brett, C.E. 2014b. Taphonomy of diploporite (Echinodermata) holdfasts from a Silurian hardground, southeast Indiana, United States: palaeoecologic and stratigraphic significance. Geological Magazine 151: 649–665. Crossref
Thomka, J.R. and Brett, C.E. 2015. Paleoecology of pelmatozoan attachment structures from a hardground surface in the middle Silurian Massie Formation, southeastern Indiana. Palaeogeography, Palaeoclimatology, Paleoecology 420: 1–12. Crossref
Acta Palaeontol. Pol. 70 (1): 125–141, 2025
https://doi.org/10.4202/app.01212.2024