
The first ornithomimosaur remains from Germany
DENIS THEDA, DARIUS NAU, RENÉ DEDERICHS, and ACHIM H. SCHWERMANN
Theda, D., Nau, D., Dederichs, R., and Schwermann, A.H. 2025. The first ornithomimosaur remains from Germany. Acta Palaeontologica Polonica 70 (3): 543–555.
Ornithomimosauria is a group of coelurosaurs primarily known from the Cretaceous of Asia and North America. The European record is comparatively sparse, with Pelecanimimus from the Lower Cretaceous of Spain being the only unequivocal representative. Here, we present a manual ungual and a distal metatarsal III from a Lower Cretaceous (Barremian to Aptian) karstic fissure fill in Balve, northwestern Germany, which we assign to Ornithomimosauria indet. We also review the literature regarding manual unguals of ornithomimosaurs and confirm previous reports of quite consistent positional variation within Ornithomimosauria, with manual ungual I being the most recurved and bearing the largest flexor tubercle, and the unguals of digits II and III being less recurved and possessing smaller tubercles. The manual ungual from Balve is closest in morphology to manual digit III. The metatarsal has a shaft with a strongly triangular cross-section, marking it as a sub- or fully developed arctometatarsal. This type of specialized third metatarsal occurs in a number of different clades of Coelurosauria (Alvarezsauroidea, Ornithomimosauria, Oviraptorosauria, Troodontidae, Tyrannosauridae). Based on its overall morphology and the rarity (Alvarezsauroidea, Troodontidae) or absence (Oviraptorosauria, Tyrannosauridae) of other clades with arctometatarsals from the fossil record of Europe, we regard it as ornithomimosaurian. This is only the second definitive record of European ornithomimosaurs, after the description of Pelecanimimus polyodon from Spain, and represents the first reported occurrence of this clade in Germany.
Key words: Ornithomimosauria, Theropoda, arctometatarsus, unguals, Lower Cretaceous, Germany.
Denis Theda [theda-paleo@gmx.de; ORCID: https://orcid.org/0009-0003-6984-0219 ], Lippisches Landesmuseum Detmold, Ameide 4, 32756 Detmold, Germany.
Darius Nau [dariusnau@uni-bonn.de; ORCID: https://orcid.org/0009-0000-4343-6830 ], Bonn Institute for Organismic Biology, University of Bonn, Nussallee 8, 53115 Bonn, Germany.
René Dederichs [rededpaleo@gmail.com; ORCID: https://orcid.org/0009-0000-2442-1545 ], Bonn Institute for Organismic Biology, University of Bonn, Nussallee 8, 53115 Bonn, Germany; University of Zurich, Department of Paleontology, Karl Schmid-Strasse 4, CH-8006 Zurich, Switzerland.
Achim H. Schwermann [achim.schwermann@lwl.org; ORCID: https://orcid.org/0000-0001-8928-8773 ], LWL-Museum für Naturkunde, Westfälisches Landesmuseum mit Planetarium, Sentruper Straße 285, Münster 48161, Germany.
Received 30 April 2025, accepted 7 July 2025, published online 10 September 2025.
Copyright © 2025 D. Theda et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
European Ornithomimosaurs.—Ornithomimosauria is a group of maniraptoriform coelurosaurs (Lee et al. 2014; Hendrickx et al. 2015), which first appeared during the Early Cretaceous and went extinct during the K/Pg extinction event. Currently, the stratigraphically oldest ornithomimosaur is Nqwebasaurus thwazi from the Lower Cretaceous Kirkwood Formation (Berriasian to Valanginian) of South Africa, but Hexing qingyi and Shenzhousaurus orientalis from the lower Yixian Formation in China are similar in age (Jin et al. 2012). Notably, it has recently been suggested that the latter is solely Barremian (Zhong et al. 2021). While Early Cretaceous ornithomimosaurs have a rich Asian fossil record (Barsbold and Perle 1984; Ji et al. 2003; Kobayashi and Barsbold 2005; Makovicky et al. 2010; Jin et al. 2012; Hattori et al. 2023), remains of this clade are rare in Lower Cretaceous deposits from Europe.
The first documented European ornithomimosaur, known from a partial anterior skeleton, is Pelecanimimus polyodon from the upper Barremian (Fregenal-Martínez et al. 2017; Cuesta et al. 2022) Las Hoyas Konservatlagerstätte in Spain (Fig. 1B). Remains originating from the Berriasian Angeac-Charente Lagerstätte (Allain et al. 2022) in France (Fig. 1B) have also been referred to Ornithomimosauria (Allain et al. 2014, 2022). Based on comparisons with the same material, Allain et al. (2014) additionally stated potential ornithomimosaurian affinities for the English (Fig. 1B) Early Cretaceous taxa Thecocoelurus daviesi Seeley, 1888 (holotype, cervical vertebra BMNH R181) and Valdoraptor oweni (Lyddeker, 1889) (holotype, left metatarsus BMNH R2559). However, both Thecocoelurus and Valdoraptor, are regarded as nomina dubia (Naish and Martill 2002; Allain et al. 2014, respectively), and are based on extremely fragmentary material. The taxonomic affinities of the Angeac taxon are ambiguous due to the presence of noasaurid abelisauroid characters in the material (Allain et al. 2014, 2022). Moreover, the material was included in a recent phylogenetic analysis in which it was recovered as a member of Abelisauroidea (Cau and Paterna 2025). Hence, we currently regard Pelecanimimus as the only unequivocal European ornithomimosaur. In this article, we add two isolated elements from an Lower Cretaceous (Barremian to Early Aptian) site from Northwestern Germany (Fig. 1) to the hitherto sparse European ornithomimosaur record.
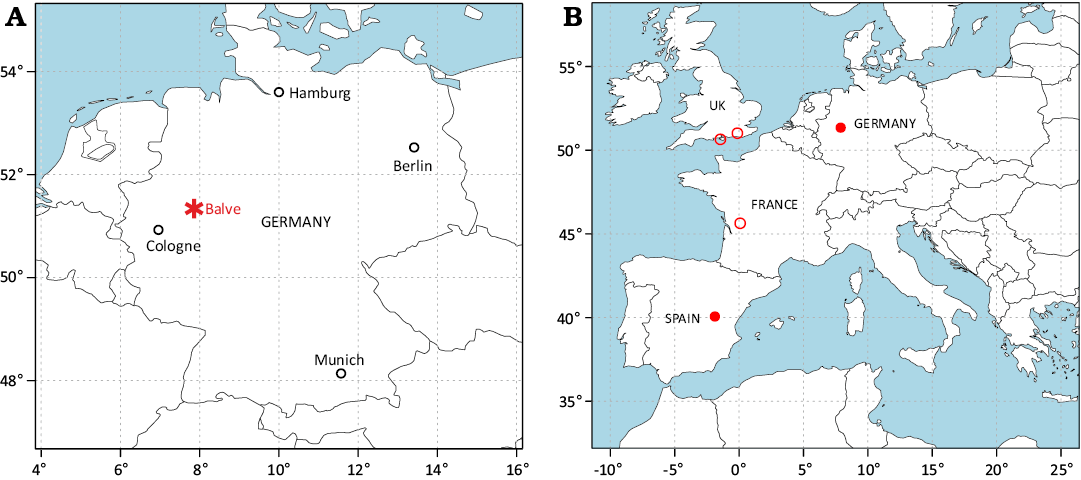
Fig. 1. Maps showing geographical position of the Balve excavation site in Germany (A, asterisk) and European localities (B) that have yielded potential (hollow circles) and definitive (solid circles) ornithomimosaur material. Occurrence data in B sourced from the Paleobiology Database (PaleoDB contributors 2025; citing Seeley 1888; Lydekker 1889; Pérez-Moreno et al. 1994; Néraudeau et al. 2012).
The Balve site.—The fossil-bearing site in Busche quarry near Balve-Beckum (Sauerland, NW Germany, Fig. 1A) is a Lower Cretaceous fissure infill in Devonian limestone. The cavities within the limestone were formed by hydrothermal karstification, beginning at the latest in the Jurassic. This Mesozoic paleokarst phenomenon is generally known from the Sauerland area as well as the Eifel region (Drozdzewski et al. 2017). At different times, the resulting fissures were filled with erosional debris from the surface (Wirth 1970; Richter et al. 2014; Drozdzewski et al. 2017).
In Balve-Beckum, the infilling took place during the Early Cretaceous (Barremian to Aptian; Lanser and Heimhofer 2015). Similar deposits of cave sediments are also known from other nearby sites, for example Wülfrath (Drozdzewski et al. 1998; Hartkopf-Fröder et al. 2012) and Brilon-Nehden (Oekentorp 1984; Norman 1987 ).
Excavations have been carried out by the LWL-Museum of Natural History in Balve-Beckum every year since 2002. The fossil material is dominated by highly fragmentary remains of vertebrates. The fauna identified so far includes cartilaginous and bony fishes, amphibians, small reptiles, turtles, crocodiles, iguanodontid ornithischians, sauropods and theropods, as well as pterosaurs, and mammals (Lanser 2011; Lanser and Heimhofer 2015; Lanser 2015; Skutschas et al. 2020; Martin et al. 2021, 2022a, b; Hornung et al. 2023; Joyce et al. 2023).
Institutional abbreviations.—AMNH, American Museum of Natural History, New York City, USA; BMNH, Natural History Museum, London, UK; CEUM, College of Eastern Utah Prehistoric Museum, Price, USA; FPDM, Fukui Prefectural Dinosaur Museum, Japan; GIN, Paleontological Center of Mongolia, Ulaan Bataar, Mongolia; IZK, Institute of Zoology, Almaty, Kazakhstan; MOR, Museum of the Rockies, Bozeman, USA; UALVP, University of Alberta Laboratory for Vertebrate Paleontology, Edmonton, Canada; UCMZ, University of Calgary, Canada; WMNM, LWL-Museum für Naturkunde Münster, Germany; YPM, Yale Peabody Museum of Natural History, New Haven, USA; ZIN PH, Paleoherpetological Collection, Zoological Institute, Russian Academy of Sciences, Saint Petersburg, Russia; ZPAL, Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland.
Other abbreviations.—CSG, claw sheath groove (= vascular groove, blood groove, vascular furrow, attachment groove); Mt, metatarsal.
Material and methods
The material presented herein comprises two isolated skeletal elements consisting of one manual ungual (WMNM P117116) and one distal metatarsal (Mt) III fragment (WMNM P100817). Both fossils are housed in the collection of the LWL-Museum für Naturkunde in Münster (WMNM). All reported measurements were taken using vernier calipers. For the sake of nomenclatural simplicity, we use the term arctometatarsal as a noun to refer to the third metatarsal with a distinct arctometatarsalian condition (sensu Holtz 1994, 1995).
Systematic palaeontology
Dinosauria Owen, 1842
Theropoda Marsh, 1881
Tetanurae Gauthier, 1986
Coelurosauria von Huene, 1914
Ornithomimosauria Barsbold, 1976
Ornithomimosauria indet.
Figs. 2, 3.
Material.—One manual ungual (WMNM P117116) and the distal part of a third metatarsal (WMNM P100817) from the Barremian–Aptian fissure filling at Balve-Beckum, Germany.
Description.—Manual ungual: WMNM P117116 is an elongate, low and narrow element with an anteroposterior length of 36 mm. Upon its discovery, it was broken into multiple smaller fragments and was subsequently restored. It lacks the distal tip and a few smaller pieces from the preaxial and postaxial surfaces (Fig. 2).
WMNM P117116 exhibits a slight palmar curvature. It is quite narrow along its dorsal edge, but the palmar surface is expanded into a wider platform (Fig. 2D, E). The dorsal lip is positioned along the centerline of the ungual (Fig. 2D). The preaxial and postaxial margins of WMNM P117116 are sub-parallel in dorsal and palmar view, from just distal of the articular end up to approximately half the distance towards the distal end. At this point, both ungual margins begin to gently curve towards the midline of the ungual up to the point where they would have converged at the missing claw tip in a complete ungual (Fig. 2D, E). The articular surface of WMNM P117116 is moderately concave (in pre- and postaxial view) (Fig. 2A, B) and is divided into a medial and a lateral facet. Both sub-facets are similar in size and shape, with both being more than twice as tall dorsoventrally as they are wide pre- to postaxially (Fig. 2C). The dorsal lip of the articular facet protrudes slightly further proximally than the ventral margin of the articular surface. The claw sheath groove (CSG) is well developed on both the preaxial and the postaxial side. The CSGs on WMNM P117116 are quite similar in depth and shape. Their curvature appears symmetrical in dorsal and distal view (Fig. 2D, F). Both are deeply set into the ungual body and do not become shallower in distal direction. One of the CSGs appears slightly deeper in distal view (Fig. 2F). The flexor tubercle is only a small protrusion on the ventral side of the ungual and is positioned directly between the articular end of the ungual and the more distally situated ventral platform (Fig. 2A, B, E). This flexor tubercle is about 10 mm distal to the ventral border of the articular facet. In pre- and postaxial view, the palmar margin between both is straight (Fig. 2A, B).
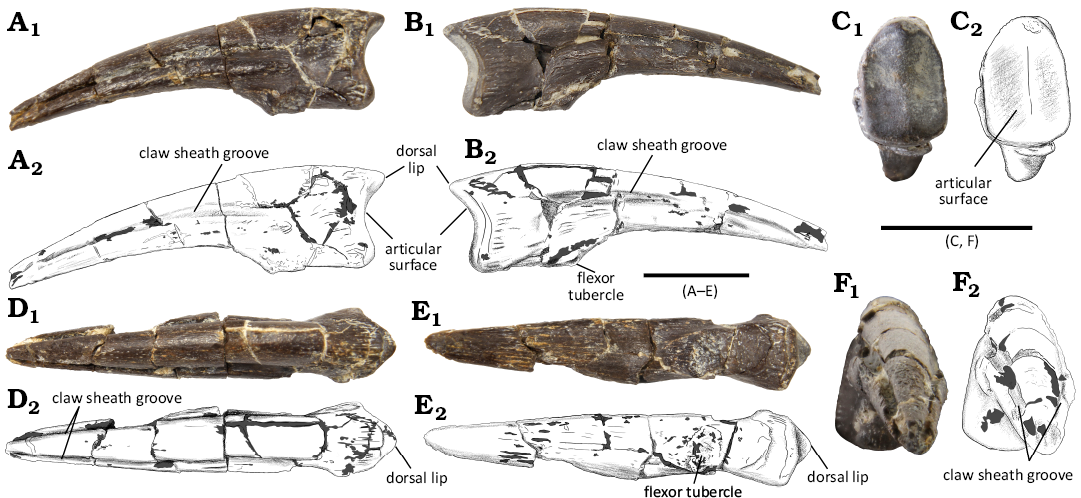
Fig. 2. Manual ungual of Ornithomimosauria indet. (WMNM P117116) from the Lower Cretaceous of Balve-Beckum, Germany, in left (A) and right (B), proximal (C), dorsal (D), palmar (E), and distal (F) views. Photographs (A1–F1) and explanatory drawings (A2–F2). Scale bars 10 mm.
Arctometatarsal: WMNM P100817 is the distal end of a left metatarsal, which was recovered from the field in three pieces that were subsequently reassembled using cyanoacrylate glue. A portion close to the plantar side of the articular surface is missing (Fig. 3C), and the edges of the distal articular facet are abraded. WMNM P100817 is 61 mm in length proximodistally. The distal articular surface is 16 mm wide and the broken, the proximal end is 10 mm wide mediolaterally. The proximal half of the fragmentary metatarsal has a pinched plantar side (Fig. 3C, F). The resulting plantar ridge (pr) along the plantar side of the specimen extends distally to the level of the gap left by the abovementioned missing fragment (Fig. 3C). Distal to the gap it bifurcates, resulting in two ridges extending to and contacting the medial and the lateral margin of the articular facet (Fig. 3C). Proximal to the gap, the ridge first increases in height evenly, before starting to decrease at around the proximodistal midpoint of the specimen (Fig. 3B, D). The plantar ridge divides the articular facet for Mt II and Mt IV. The articular surface for Mt II is larger and more pronounced than the one for Mt IV. The articulation for Mt II has a rough surface that is marked by primarily proximodistally oriented striations (Fig. 3B, D). These run from the broken part of the fossil and extend distally to the point at which the plantar ridge begins to decrease in height. The Mt IV facet differs from the one for Mt II in exhibiting a more rugose surface (Fig. 3B). Unlike in the Mt II facet, this rugosity only covers the total width of the facet for the proximal 12 mm. Distal to this point, the rugose surface rapidly tapers in mediolateral width until it is restricted to a narrow strip on the lateral side of the ridge (Fig. 3B, C). This rugose strip then continues to approximately the level of the gap (missing fragment on plantar side) and subsequently disappears (Fig. 3B, C).
A medial expansion is present at mid-shaft. From proximal to distal, it first expands medially, before curving back towards the shaft. While the medial margin extends anteromedially in proximal view, the lateral margin is more strongly angled in anterior direction. Additionally, the proximal third of the shaft is distinctly concave. In combination, this gives the proximal end of WMNM P100817 a strongly tick-shaped cross-section (Fig. 3F). The anterior surface of the metatarsal gradually becomes less concave and flatter distally, making the distal end of the shaft triangular in cross-section. In anterior view, WMNM P100817 shows a sinusoidal profile, with both the medial and lateral margin first curving laterally in the proximal half of the shaft, before they reverse direction and curve strongly medially (Fig. 3A). In anterior view, the medial margin of the metatarsal appears especially concave (Fig. 3A).
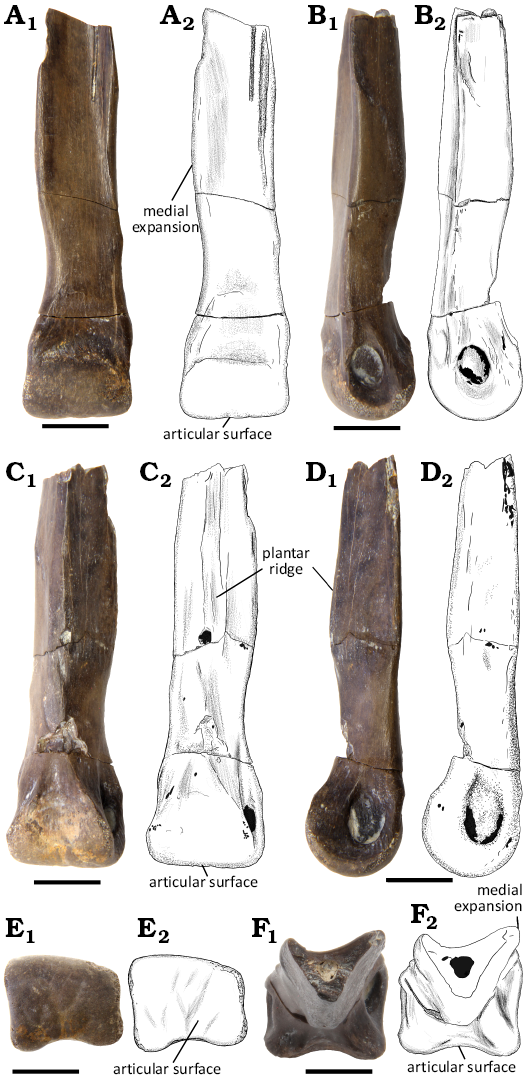
Fig. 3. Metatarsal of Ornithomimosauria indet. (WMNM P100817) from the Lower Cretaceous of Balve-Beckum, Germany, in anterior (A), lateral (B), plantar (C), medial (D), distal (E), and proximal (F) views. Photographs (A1–F1) and explanatory drawings (A2–F2). Scale bars 10 mm.
The distal articular surface is non-ginglymoid. Instead, it exhibits one continuous, smooth articulation for the proximal-most pedal phalanx (Fig. 3E). Only the plantar side shows a slight concavity (Fig. 3C), facilitating a division of the articular surface into a medial and lateral part. However, this divided plantar portion of the facet is also well-rounded and only extends proximally to about half the level of the collateral ligament fossae. The articular surface extends slightly further in proximal direction on the anterior side (compare Fig. 3A, C). The articular facet is not fully symmetrical, as it is slightly proximodistally taller on the medial than on the lateral side in anterior view (Fig. 3A). Additionally, the medial side of the articular end of WMNM P100817 is more anteroplantarly expanded (Fig. 3C).
The collateral ligament pits are different in size. The medial pit (Fig. 3D) is elliptical and proximodistally longer than the lateral pit (Fig. 3B). It also has a less defined proximal margin and instead gradually merges with the shaft (Fig. 3D). The lateral pit is more circular in shape and exhibits a continuously steep and clearly delimited margin (Fig. 3B). The lateral ligament fossa is surrounded by a rhomboid to elliptical concavity (Fig. 3B).
The bean-shaped, centrally positioned extensor fossa, proximal to the dorsal margin of the articular surface, is shallow (Fig. 3A). It is more deeply excavated in its distal part right adjacent to the articular facet. The fossa is not symmetrical in anterior view, extending furthest in proximal direction in its lateral region (Fig. 3A). The area around the extensor fossa has a faintly indented appearance in proximal, medial and lateral direction.
Discussion
Manual ungual WMNM P117116.—The manual unguals of ornithomimosaurs possess a relatively small and distally placed flexor tubercle (compared to other theropod clades; Makovicky et al. 2004) and often tend to have only a weakly developed ventral curvature (Nicholls and Russell 1985: fig. 8; Chinzorig et al. 2018: fig. 6). All of these general characters are present in WMNM P117116 (Figs. 2, 4).
Some ornithomimosaur taxa include manual unguals that are substantially different in morphology from WMNM P117116. For instance, many early-branching ornithomimosaurs are quite distinct from the studied specimen. WMNM P117116 has a weaker ventral curvature than the manual unguals of Hexing qingyi (Jin et al. 2012: fig. 26.6.), Nqwebasaurus thwazi (Choiniere et al. 2012: fig. 12; Sereno 2017) and Shenzhousaurus orientalis (Ji et al. 2003: fig. 11). The manual unguals of the paratype (CEUM 5072) of Nedcolbertia justinhofmanni also differ from the WMNM P117116, because manual ungual I of CEUM 5072 has a relatively large flexor tubercle for an ornithomimosaur (Brownstein 2017), while it is very small in its manual ungual II. The flexor tubercle of manual ungual II in CEUM 5072 is also more proximally placed (Kirkland et al. 1998) than in WMNM P117116. Among more derived ornithomimid ornithomimosaurs, WMNM P117116 is clearly distinct morphologically from Gallimimus bullatus with its comparatively short, dorsoventrally deep and more recurved manual unguals (Osmólska et al. 1972; Chinzorig et al. 2018: fig. 6E). The same holds true for the manual unguals of the deinocheirids (Osmólska and Roniewicz 1970) Deinocheirus mirificus (Lee et al. 2014; Chinzorig et al. 2018: fig. 6G) and Paraxenisaurus normalensis (Serrano-Brañas et al. 2020). The manual unguals of the ornithomimids Ornithomimus sp. and Anserimimus planinychus (Barsbold 1988; Chinzorig et al. 2018: fig. 6A) are extremely flat (Makovicky et al. 2004, the authors only refer to the genus Ornithomimus and no particular species; Kobayashi and Barsbold 2006). In Anserimimus planinychus, the manual unguals are also rather wide, as they possess pronounced mediolateral keels, which give them a more triangular profile in dorsal view (Kobayashi and Barsbold 2006: fig. 5; Chinzorig et al. 2018: fig. 5G) than is present in WMNM P117116.
We agree with previous works (e.g., Makovicky et al. 2004; Ikegami 2016; Chinzorig et al. 2018; Macdonald and Currie 2019; Nottrodt 2022), that the manual unguals of ornithomimosaurs exhibit notable positional variation (Fig. 4A). Manual digit I of many ornithomimosaurs has an ungual that is more recurved and possesses a larger flexor tubercle than manual digits II and III (e.g., Makovicky et al. 2004; Chinzorig et al. 2018: fig. 6). This condition is present in the non-ornithomimid ornithomimosaurs Harpymimus okladnikovi (Barsbold and Perle 1984; see also Kobayashi and Barsbold 2005), Nedcolbertia justinhofmanni (Kirkland et al. 1998), Nqwebasaurus thwazi (Choiniere et al. 2012; Sereno 2017), Pelecanimimus polyodon (Cuesta et al. 2022), and Beishanlong grandis (Makovicky et al. 2010). It is also present in the ornithomimids Ornithomimus edmontonicus and sp. respectively (Sternberg 1933; Nottrodt 2022) and Sinornithomimus dongi (Kobayashi and Lü 2003). Material referred to Archaeornithomimus asiaticus encompasses relatively strongly recurved, medioloaterally compressed manual unguals, as well as rather straight ones (Smith and Galton 1990), which also resembles positional variation. With its small flexor tubercle, straight lateral and medial profile and general slenderness, WMNM P117116 shows much closer resemblance to the manual unguals II and III than to manual ungual I of all of these taxa (see Chinzorig et al. 2018: fig. 6).
Several ornithomimosaurs possess manual unguals comparable in morphology to WMNM P117116. Those of Sinornithomimus dongi are generally similar to WMNM P117116, but their articular facets are less concave and exhibit a less developed dorsal lip (Kobayashi and Lü 2003: fig. 18). Another manual ungual that is quite similar to WMNM P117116 is part of specimen ZPAL MgD-I/65 from the Upper Cretaceous Nemegt Formation of Mongolia, which has been as assigned to aff. Anserimimus planinychus (Bronowicz 2011: fig. 5). However, its articular surface is also much less concave than the one of WMNM P117116. Bronowicz (2011) states that the ungual is less recurved than, and lacks the pronounced mediolateral spurs, seen in Anserimimus. Bronowicz (2011) interpreted this as a potential taxonomic signal differentiating ZPAL MgD-I/65 from Anserimimus planinychus. However, the lessened palmar curvature of the ungual fits the general positional variation seen in ornithomimosaurs outlined above, according to which manual ungual III is typically the most slender and unrecurved one (e.g., Harpymimus okladnikovi, Kobayashi and Barsbold 2005: fig. 6.7). We suggest it is possible that the ungual shown in fig. 5 of Bronowicz (2011) might actually just be a manual ungual III of an ornithomimosaur and therefore provide a positional, rather than a taxonomic signal. Moreover, manual ungual III is unknown for the holotype GIN 100/300 of Anserimimus (Barsbold 1988), which means that there is potentially no overlap in the manual unguals of both specimens. However, if the shape of the ungual in Bronowicz (2011: fig. 5) is representative for all referred unguals mentioned in that work, it might indeed be a valid taxonomic feature differentiating Anserimimus planinychus and ZPAL MgD-I/65.
The manual unguals referred to Tyrannomimus fukuiensis (Hattori et al. 2023) from the Aptian of Japan are generally comparable to WMNM P117116. They are relatively straight, symmetrical and have a moderately concave articular facet in medial and lateral view (Hattori et al. 2023: fig. 5). A straight palmar margin between the flexor tubercle and the ventral margin of the articular facet similar to WMNM P117116, is only present in one of the figured manual unguals (Hattori et al. 2023: fig. 5M, FPDM-V-8580).
The single preserved manual ungual (right manual digit II) of UALVP 16182, a specimen referred to Dromiceiomimus brevitertius, also closely mirrors the morphology of WMNM P117116 (Macdonald and Currie 2019). They share the character of a small, distally placed flexor tubercle (Macdonald and Currie 2019). Both are also similar in possessing a straight ventral margin between the flexor tubercle and the ventral edge of the proximal articular facet (see Macdonald and Currie 2019: fig. 13A). WMNM P117116 and the manual ungual of UALVP 16182 also both exhibit a moderately concave articular surface and both have dorsal and ventral lips reaching to about the same level proximally. The slight ventral curvature (more so than in e.g., Shenzhousaurus orientalis, less so than in e.g., Anserimimus planinychus) is also comparable in both.
The straighter ones among the ungual specimens referred to the genus Archaeornithomimus asiaticus (see Smith and Galton 1990: fig. 2W, X, AMNH 21886 and 21887) closely resemble WMNM P117116 in their reduced ventral curvature, gracile ungual body distal to the flexor tubercle and the proximodistally straight palmar margin connecting the ventral margin of the articular surface and the flexor tubercle. WMNM P117116 is also notably similar to manual ungual III of the European, Early Cretaceous Pelecanimimus polyodon in terms of slenderness, straightness and small size of the flexor tubercle (Fig. 4B, C; Cuesta et al. 2022). Also quite close in overall morphology is manual ungual III of Harpymimus okladnikovi (Fig. 4A3, C), with which WMNM P117116 shares the elongate and low preaxial or postaxial profile, strongly reduced ventral curvature, flat ventral surface (Kobayashi and Barsbold 2005: fig. 6.7H) and the roughly rectangular profile of the distally placed flexor tubercle and the straight palmar margin connecting it to the ventral margin of the articular surface (resulting in rectangular preaxial and postaxial profiles of the flexor tubercle and the palmar margin). The articular surfaces of WMNM P117116 and manual ungual III of Harpymimus okladnikovi also appear to be equally concave (Kobayashi and Barsbold 2005: fig. 6.7H). Moreover, the dorsal lip extends proximally to around the level of the ventral margin of the articular facet just as in WMNM P117116 (Kobayashi and Barsbold 2005: fig. 6.7H).
Overall, WMNM P117116 is extremely similar to the manual unguals seen in species of several different ornithomimosaur genera (Archaeornithomimus, Dromiceiomimus, Harpymimus, Pelecanimimus, and Tyrannomimus). Out of these it is most comparable to the straighter and rather gracile unguals of the second, or even more likely, third manual digits. WMNM P117116 is particularly close in appearance to manual ungual III of Harpymimus polyodon and Pelecanimimus okladnikovi (Fig. 4).
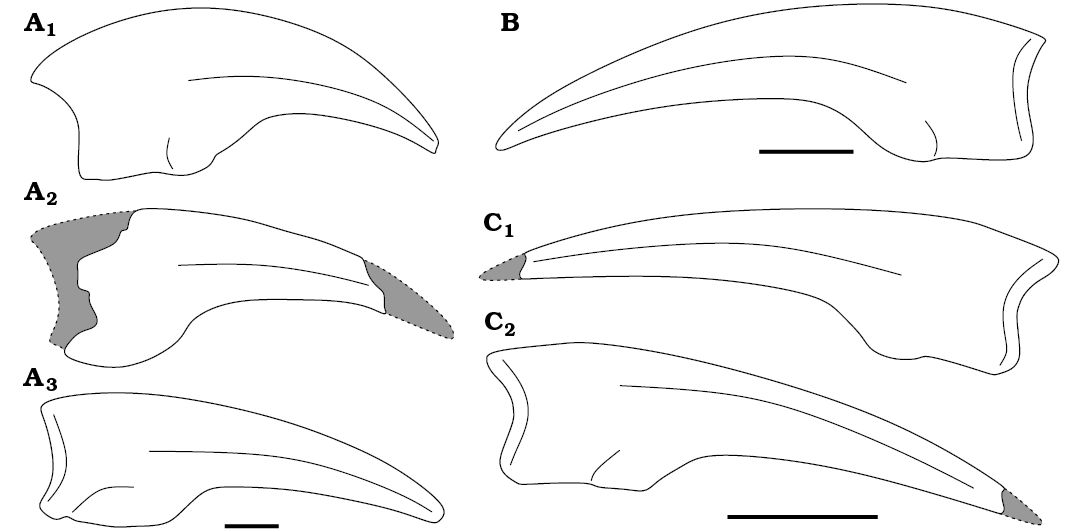
Fig. 4. Outline drawings of manual unguals of different ornithomimosaurs. A. Harpymimus okladnikovi Barsbold & Perle, 1984, ungual I (A1), ungual II (A2), ungual III (A3). B. Pelecanimimus polyodon Pérez-Moreno et al., 1994, ungual III (after Cuesta et al. 2022: fig. 16E). C. Ornithomimosauria indet. (WMNM P117116) in left (C1) and right (C2) views. Scale bars 10 mm.
Metatarsal WMNM P100817.—The triangular outline of the shaft of WMNM P100817 clearly shows that it is the distal half of a (sub-)arctometatarsal (Mt III) of a theropod. Arctometatarsals are also commonly represented by their more robust distal half in the fossil record, as their thin midshaft is susceptible to breakage (Buffetaut et al. 2009).
Theropod (sub-)arctometatarsals: Arctometatarsals (i.e., a proximally strongly pinched Mt III that is fully covered anteriorly by Mt II and Mt IV, fully obscuring Mt III in anterior view) have convergently evolved several times within coelurosaurian theropods as an adaptation to a cursorial lifestyle (Snively et al. 2004; White 2009). The proximal pinch and distally triangular cross-section of the shaft results in medial rotation of pedal digit II and a lateral rotation of digit IV. In contrast, a subarctometatarsalian condition refers to such metatarsals that are merely mediolaterally compressed proximally, but not fully covered anteriorly by Mt II and Mt IV (White 2009). Such sub-arctometetarsals can still exhibit a triangular distal cross section, just like fully developed arctometatarsals (e.g., Xu et al. 2018; Samathi 2024). True arctometatarsals occur in Tyrannosauridae, Troodontidae, Oviraptorosauria, Ornithomimosauria (Holtz 1994, 1995; Snively et al. 2004) and Alvarezsauroidea (Snively et al. 2004). The sub-arctometatarsalian condition has been documented in alvarezsauroids (Choiniere et al. 2010; Xu et al. 2018), oviraptorosaurs (e.g., He et al. 2008; Qiu et al. 2019) and ornithomimosaurs (e.g., Buffetaut et al. 2009). Due to the isolated nature and missing proximal section of WMNM P100817, it is not possible to fully determine whether the preserved element had the sub-arctometatarsalian or the fully arctometatarsalian condition. Therefore, the comparison in this work focuses on generally comparing WMNM P100817 with third metatarsals of taxa with either of these states.
Tyrannosauridae: Due to tyrannosaurids being primarily known from Asia and North America and only appearing during Late Cretaceous (Holtz 2004), we exclude the possibility of these elements stemming from a member of this theropod group. By contrast, all other of the abovementioned clades could at least theoretically be expected in Barremian to Aptian-aged deposits from Europe. The non-tyrannosaurid tyrannosauroid Eotyrannus lengi occurs in roughly coeval strata from England, but lacks the arctometatarsalian condition (Naish and Cau 2022).
Alvarezsauroidea: Early-branching alvarezsauroids tend to exhibit the sub-arctometatarsalian condition (e.g., Haplocheirus sollers). The distal part of Mt III can still be triangular in cross section in these sub-arctometatarsals (e.g., Xiyunykus pengi), as is seen in WMNM P100817. In alvarezsauroids belonging to Parvicursorinae (Karhu and Rautian 1996), a fully arctometatarsalian Mt III is present (e.g., Hone et al. 2013 and references therein). The non-ginglymoid distal articular facet of WMNM P100817 is similar to the one in alvarezsauroids (Fig. 5C). Although Mononykus olecranus was described to possess a “trochlea” at its distal end (Perle et al. 1994), we note it seems to have a rather smooth appearance (Perle et al. 1994: fig. 18G–I). Corroborating the presence of a non-ginglymoid condition in the third metatarsals of alvarezsauroids is the fact that smooth distal articular surfaces are also definitively present in Mt III of Kol ghuva (Turner et al. 2009: figs. 1, 2), Linhenykus monodactylus (Xu et al. 2013: fig. 2i) and Jaculinykus yaruui (Kubo et al. 2023: fig. 16E, G). Parvicursor remotus exhibits an only weakly ginglymoid condition (Averianov and Lopatin 2022).
In Mononykus olecranus (Perle et al. 1994: fig. 18), Xixianykus zhangi (Xu et al. 2010: fig. 7T) and Linhenykus monodactylus (Xu et al. 2011b: fig. 2I) Mt III displays a concave anterior surface with a more anterolaterally oriented lateral margin than the medial margin of the shaft, as is also present in WMNM P100817. By contrast, the third metatarsals of Kol ghuva (Turner et al. 2009: figs. 1–3) and of Jaculinykus yaruui (Kubo et al. 2023: fig. 16E) lack this concave anterior surface. The extensor surface is slightly convex in Parvicursor remotus (Averianov and Lopatin 2022). Moreover, Mt III of all of these alvarezsauroids is different from WMNM P100817 in consistently possessing a relatively straight distal shaft in anterior view as opposed to the slightly sinusoidal anterior profile present in WMNM P100817 (Figs. 3A, 5C). Therefore, we regard alvarezsauroid affinities as unlikely for WMNM P100817. Moreover, to our knowledge, alvarezsauroid remains are absent from the Lower Cretaceous of Europe. The group had appeared in Asia by the early Late Jurassic (e.g., Choiniere et al. 2010, 2014), but only occurs in Europe during the Late Cretaceous (Naish and Dyke 2004).
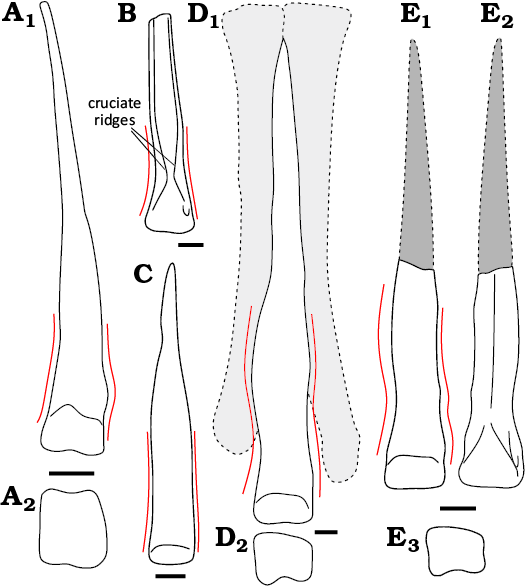
Fig. 5. Outline drawings of left Mt III of theropod dinosaurs (A–D) and Ornithomimosauria indet. (E). A. Talos sampsoni Zanno et al., 2011 (Troodontidae), in anterior (A1) and distal (A2) views (modified after Zanno et al. 2011: fig. 11J, I, respectively). B. Citipes elegans Parks, 1933 (Oviraptorosauria), in plantar view (after Funston et al. 2016: fig. 4A). C. Kol ghuva Turner et al., 2009 (?Alvarezsauridae), in anterior view (after Turner et al. 2009: fig. 1). D. Ornithomimus velox Marsh, 1890 (Ornithomimidae), in anterior (D1) and distal (D2) views (Mt II and IV shaded, modified after Claessens and Loewen 2016). E. Ornithomimosauria indet. (WMNM P100817), in anterior (E1), distal (E2), and plantar (E3) views. Red lines indicate distal shaft outline in anterior view. Scale bars 10 mm.
Troodontidae: Early-branching troodontids exhibit a sub-arctometatarsalian Mt III (e.g., Sinovenator changii, Xu et al. 2002), but later taxa also evolve a fully arctometatarsalian condition (e.g., Snively et al. 2004). WMNM P100817 differs from Mt III of troodontids. For one, it lacks a large proximoplantar extension of the distal articular surface and the strongly asymmetrical articular surfaces for Mt II and Mt IV (and associated sharp, oblique ridge dividing both) seen in specimens referred to Troodon formosus (Snively et al. 2004: fig. 6; Varricchio et al. 2025: fig. 6, MOR553S-7-29-92-113). A longitudinal groove on the anterior surface of Mt III as seen in Linhevenator tani (Xu et al. 2011a) and Philovenator curriei (Xu et al. 2012) is absent in WMNM P100817. Additionally, the distal articular surface of Mt III tends to be slightly ginglymoid (Fig. 5A1) in some troodontids, such as Tochisaurus nemegtensis (Kurzanov and Osmólska 1991), Talos sampsoni (Zanno et al. 2011), Harenadraco prima (Lee et al. 2024) and a specimen assigned to Troodon formosus (Varricchio et al. 2025: fig. 6, MOR553S-7-29-92-113). According to Norell et al. (2009), Saurornithoides mongoliensis has a non-ginglymoid Mt III, but the element is not well preserved (Norell et al. 2009: fig. 20). Moreover, in troodontids for which we found Mt III figured in distal view, namely Talos sampsoni and Harenadraco prima, the ginglymoid distal articular end of Mt III has a square-shaped outline (Fig. 5A2) in distal view (Zanno et al. 2011: fig. 11I; Lee et al. 2024, respectively) compared to the more sub-rectangular outline in WMNM P100817 (Fig. 3E). An Mt III referred to Troodon formosus (MOR553S-7-29-92-113), exhibits a distal articular surface with a less sub-rectangular outline that instead is anteroposteriorly longer than mediolaterally wide (Varricchio et al. 2025: fig. 6). This is in strong contrast to the more mediolaterally wide than anteroposteriorly long WMNM P100817. We therefore regard WMNM P100817 as likely not belonging to a troodontid.
Troodontids have an extensive record for the Lower Cretaceous of Asia (Russell and Dong 1993; Xu et al. 2002, 2017; Xu and Wang 2004; Shen et al. 2017; Kubota et al. 2024). By contrast, their remains are a rare element of the European fossil record (Sellés et al. 2021). The stratigraphically oldest troodontid teeth from Europe stem from the Cenomanian of France (Vullo et al. 2007) and bony remains appear in the fossil record only during the Maastrichtian of Spain (Sellés et al. 2021).
Oviraptorosauria: A sub-arctometatarsalian condition with a compressed proximal shaft is often present in early-branching oviraptorosaurs (e.g., He et al. 2008; Qiu et al. 2019) and some are even fully arctometatarsalian, such as Yuanyanglong bainian (Hao et al. 2025). An issue with comparing WMNM P100817 to Early Cretaceous oviraptorosaurs lies in an either two-dimensional preservation (He et al. 2008), or, in the case of Yuanyanglong bainian a lack of a preserved distal end of Mt III (Hao et al. 2025).
Three-dimensional, well-preserved arctometatarsals are available for some avimimids, such as some specimens from the Iren Dabasu Formation in China (Funston et al. 2019). This material is comparable to the WMNM P100817 in having a shallow extensor fossa and a mostly smooth, undivided articular facet (Funston et al. 2019: fig. 8I), but lacks the marked, bifurcated plantar ridge of WMNM P100817 and the distal articular facet extends further anteroproximally (e.g., Funston et al. 2019: fig. 8) in the Balve specimen (Fig. 3E). WMNM P100817 also differs from caenagnathid arctometatarsals in several regards. For instance, it lacks the “paired cruciate ridges” (Currie et al. 2016: 154) on the plantar side of Mt III (Currie et al. 2016; Funston et al. 2016; Yu et al. 2018; Funston 2020; Atkins-Weltman et al. 2024) (Fig. 5B, E3). Caenagnathid third metatarsals also tend to be anteroplantarly flattened (Funston et al. 2016) as opposed to WMNM P100817 with its strongly triangular cross section. One exception is Anomalipes zhaoi which has an anteroplantarly deeper than mediolaterally wide Mt III (Yu et al. 2018).
While on morphological grounds we do not strictly exclude the possibility of WMNM P100817 being referable to an early-branching oviraptorosaur, we regard it as unlikely at this time, as the presence of oviraptorosaurs in Europe as a whole remains a subject of debate at the time of writing. Similar to troodontids, early oviraptorosaurs were widespread in Asia by the Early Cretaceous (e.g., Qiang et al. 1998; Xu et al. 2002; He et al. 2008; Qiu et al. 2019; Hao et al. 2025) and possibly present in North America by the Late Jurassic (Chapelle et al. 2021), but this is not reflected by the European record. The holotype cervical vertebra of Thecocoelurus daviesi (BMNH R181) from the Lower Cretaceous of the Isle of Wight was considered as referable to Oviraptorosauria by Naish and Martill (2002), but it has since also alternatively been proposed to belong to a therizinosaur, a noasaurid, or an ornithomimosaur (Allain et al. 2014 and references therein).
Ornithomimosauria: Despite not fully ruling out the possibility of WMNM P100817 belonging to one of the other mentioned theropod groups with (sub-)arctometatarsalian Mt IIIs (such as early-branching oviraptorosaurs), its overall morphology is most consistent with third metatarsals of the ornithomimosaur taxa we were able to compare it to through the literature. The distal articular facet of WMNM P100817 is non-ginglymoid, as is generally the case in ornithomimosaurs (Snively et al. 2004) and exceptions are only very weakly ginglymoid (Arkansaurus fridayi, Hunt and Quinn 2018; Tototlmimus packardensis, Serrano-Brañas et al. 2016; also slightly in the Dromiceiomimus brevitertius [UALVP 16182], Macdonald and Currie 2019: fig. 16C; Qiupalong henanensis, Xu et al. 2011a). Moreover, WMNM P100817 possesses a shallow extensor fossa as is present in ornithomimosaurs (Snively et al. 2004).
Similar to that in the other theropod groups discussed above, the arctometatarsalian condition of Mt III within Ornithomimosauria is developed to varying degrees. Early-branching ornithomimosaurs such as Nqwebasaurus thwazi (shaft only triangular in distal half, Sereno 2017), or the deinocheirids Deinocheirus mirificus and Paraxenisaurus normalensis are non-arctometatarsalian (Lee et al. 2014; Serrano-Brañas et al. 2020). More derived non-ornithomimid ornithomimosaurs exhibit intermediate, sub-arctometatarsalian conditions. Among these sub-arctometatarsalian taxa are Arkansaurus fridayi, Beishanlong grandis, Garudimimus brevipes, Harpymimus okladnikovi, and Nedcolbertia justinhofmanni (Hunt and Quinn 2018; Makovicky et al. 2010; Buffetaut et al. 2009; Brownstein 2017). The cross-section of the distal shaft of WMNM P100817 differs from the D-shaped cross-section observed in some of the early sub-arctometatarsalian ornithomimosaurs, such as Garudimimus brevipes and Harpymimus okladnikovi (Buffetaut et al. 2009). The fully arctometatarsalian condition, i.e., shaft triangular in cross section which tapers extremely in proximal direction and proximal end of Mt III fully covered by Mt II and Mt IV in anterior view, is present in ornithomimid ornithomimosaurs, such as Aepyornithomimus tugrikinensis (Chinzorig et al. 2017), Anserimimus planinychus (Barsbold 1988), Archaeornithomimus asiaticus (Smith and Galton 1990), Gallimimus bullatus (Osmólska et al. 1972), Ornithomimus velox (e.g., Claessens and Loewen 2016), Qiupalong henanensis (Xu et al. 2011a), Rativates evadens (McFeeters et al. 2016) and Sinornithomimus dongi (Kobayashi and Lü 2003).
The third metatarsals of certain ornithomimosaurs differ strongly from WMNM P100817. In the Asian ornithomimid Qiupalong henanensis, the lateral condyle is slightly larger than in WMNM P100817. The entire distal articulation is mediolaterally wider than in most other arctometatarsalian ornithomimosaurs (Xu et al. 2011a: fig. 6). The slightly ginglymoid condition of Qiupalong (Xu et al. 2011a: fig. 6a, c) is also absent in WMNM P100817, resulting in a marked difference in overall appearance. Mt III of the Early Cretaceous Arkansaurus fridayi also differs strongly from WMNM P100817. The distal articulation of Mt III also has a generally very slender appearance in Arkansaurus fridayi (Hunt and Quinn 2018: fig. 3), as opposed to the wider distal articular facet in WMNM P100817. The anterior surface is convex in Mt III of Arkansaurus fridayi (Hunt and Quinn 2018), instead of concave as in WMNM P100817. The flat anterior surface of Mt III of the ornithomimids Anserimimus planinychus (Barsbold 1988), and Rativates evadens (McFeeters et al. 2016) also differs from WMNM P100817. This concave anterior surface is one of the most distinct features of the preserved shaft section of WMNM P100817. A similar structure is present in the Dromiceiomimus brevitertius (UALVP 16182), which also exhibits an anterolaterally bent lateral margin (Macdonald and Currie 2019: fig. 16C) comparable to WMNM P100817.
The third metatarsals of multiple ornithomimosaurs are quite comparable to WMNM P100817. For instance, WMNM P100817 is very similar in overall morphology to the arctometatarsal of the Mongolian Late Cretaceous Aepyornithomimus tugrikinensis (Chinzorig et al. 2017). Both have a concave anterior surface on the shaft of the metatarsal. They also both exhibit a distal articular facet that has a rectangular cross section (although with rounded corners) in anterior view. The articular facet is non-ginglymoid with barely any intercondylar groove being recognizable in both. However, the extensor fossa is more pronounced in WMNM P100817 than in Aepyornithomimus tugrikinensis. Additionally, the medial margin of the articular facet of WMNM P100817 is concave in distal view, differing from the straight margin in Aepyornithomimus tugrikinensis. The concave medial margin between the articular end and the medial expansion of WMNM P100817 (visible in anterior view) also differentiates it from the straight margin in Mt III of Aepyornithomimus tugrikinensis (Chinzorig et al. 2017: fig. 3a). This concave medial margin of WMNM P100817 situated between the distal articular surface and the medial expansion of the shaft of Mt III is more reminiscent of the condition in the sub-arctometatarsalian ornithomimosaur Garudimimus brevipes, and, at least somewhat, in Harpymimus okladnikovi (Kobayashi and Barsbold 2005: fig. 6.10B). WMNM P100817 is also similar to Mt III of the Early Cretaceous Kinnareemimus khonkaenensis from Thailand (Buffetaut et al. 2009; Samathi 2024). For instance, they share a mid-shaft with a triangular cross-section and a marked medial expansion (Buffetaut et al. 2009; Samathi 2024). The distal articulation of WMNM P100817 (Fig. 3) and Mt III of Kinnareemimus khonkaenensis (Samathi 2024: fig. 4) are non-ginglymoid. The articular surface of WMNM P100817 also has a similar outline to Kinnareemimus khonkaenensis in distal view, being mediolaterally wider than anteroplantarly long (Samathi 2024: fig. 4K). It is also anteroplantarly longer on its medial side than on its lateral side in both, WMNM P100817 (Fig. 3E) and Kinnareemimus khonkaenensis (Samathi 2024: fig. 4K). However, the flat anterior shaft surface of Mt III of Kinnareemimus khonkaenensis (Buffetaut et al. 2009; Samathi 2024, fig. 4) is markedly different from the anteriorly concave mid-shaft of WMNM P100817 (Fig. 3A). The distinct sinusoidal anterior profile of the WMNM P100817 (Fig. 3A) is also absent in metatarsals of Kinnareemimus khonkaenensis (Samathi 2024: fig. 4). Finally, the central, plantar ridge bifurcates very close to the distal articulation in WMNM P100817 (Fig. 3C) and appears to do so much further proximally in Kinnareemimus khonkaenensis (Samathi 2024: fig. 4H).
WMNM P100817 is morphologically even closer to Mt III of the holotype (YPM 542) of Ornithomimus velox (Marsh 1890). Both have a distal articular surface in which the medial half is more anteroplantarly expanded than the lateral half (see Fig. 5D2, E2; Claessens and Loewen 2016: figs. 4H, 8N). Both metatarsals also exhibit a concave anterior surface of the shaft with a more anteriorly inclined lateral margin (Claessens and Loewen 2016: fig. 4). This condition is also present in Struthiomimus altus (AMNH 5385, UCMZ 1980.1, McFeeters 2015: fig. 2.20B, C). WMNM P100817 and Mt III of Ornithomimus velox share that the anterior inclination of the lateral margin is strongest at about the level of the greatest medial extent of the medial expansion (Fig. 5D1, E1). In Struthiomimus altus, this anterior inflexion is greatest proximal to the medial expansion (McFeeters 2015: fig. 2.20B, C). While more strongly developed in WMNM P100817, the concave medial margin (in anterior view) between the distal articular surface and the medial expansion is also somewhat present in Ornithomimus velox (YPM 542, Claessens and Loewen 2016: figs. 4H, 8E). This gives Ornithomimus velox a slightly sinusoidal anterior outline, albeit less than in Struthiomimus altus and WMNM P100817 (Fig. 5D1, E1), in both of which this condition is quite pronounced (McFeeters 2015). The extensor fossa of WMNM P100817 is more conspicuous than in most other ornithomimosaurs, in which it is normally a shallow structure (e.g., Snively et al. 2004). However, a shallow extensor fossa is clearly visible in Qiupalong henanensis (Xu et al. 2011a: fig. 6a) and in Rativates (McFeeters, personal communication 2025; McFeeters et al. 2016: fig. 10; note that Hunt and Quinn 2018: fig. 6 additionally shows a shallow extensor fossa in Mt III of Beishanlong grandis and Harpymimus okladnikovi). A shallow, yet clearly demarcated extensor fossa is also present in Ornithomimus velox (Claessens and Loewen 2016: fig. 4H, 8E) and Struthiomimus altus (McFeeters 2015: fig. 2.20B, C). The latter shows the greatest resemblance to WMNM P100817, with a clearly demarcated, but shallow and bean-shaped extensor fossa that very gradually decreases in depth proximally. Additionally, the distal articular surface extends further proximally on the medial side (in anterior view) in both, WMNM P100817 and Mt III of Struthiomimus altus (McFeeters 2015: fig. 2.20). WMNM P100817 is also morphologically similar (despite being much smaller) to a distal Mt III from the Upper Cretaceous of Kazakhstan (IZK 1432/2012) referred to Ornithomimidae by Averianov et al. (2017), as both possess a bifurcated central ridge on the plantar side and have a comparable general articular facet shape (non-ginglymoid, deeper anteroplantarly on the medial side in distal view, Averianov et al. 2017: fig. 4).
WMNM P100817 is not an exact match for a third metatarsal of any known ornithomimosaur and instead exhibits a combination of features from other taxa. What is of note here is that the distal Mt fragment is very close in morphology to the ornithomimids Aepyornithomimus tugrikinensis and Dromiceiomimus brevitertius (UALVP 16182), considering the concave anterior surface of the shaft and well-rounded and non-ginglymoid articular surface, as well as some non-ornithomimid ornithomimosaurs (Garudimimus brevipes, Harpymimus okladnikovi, Kinnareemimus khonkaenensis). However, WMNM P100817 actually most closely resembles the arctometatarsals of the derived North American, Late Cretaceous ornithomimids Ornithomimus velox and Struthiomimus altus as all of them have very similar articular facets (especially in distal and anterior view), a concave anterior side of the shaft and a sinusoidal outline in anterior view.
Conclusions
The isolated postcranial material described herein is referred to Ornithomimosauria indet. While we cannot fully exclude other affinities for the distal (sub-)arctometatarsal WMNM P100817, we consider it to be morphologically most similar to ornithomimid ornithomimosaurs. In fact, WMNM P100817 particularly resembles the arctometatarsals of the Late Cretaceous ornithomimids Ornithomimus spp. and Struthiomimus altus. We regard the very distinctly shaped manual ungual WMNM P117116 as the main evidence for the presence of ornithomimosaurs at the Balve site. Due to the similar pattern of positional variation in most ornithomimosaurs, the straight shape and narrowness identify it as a second, or even more likely, third manual ungual. Despite only being able to provide a very general taxonomic identification here, our findings nevertheless strengthen the evidence for ornithomimosaurs as an established faunal component of European Early Cretaceous ecosystems, and extend their known range, for that stratigraphic time interval, from Iberia (Pelecanimimus polyodon) to the Central European Rise. Since the excavation at the Balve site continues annually, more dinosaur material is uncovered every year, which makes us optimistic to be able to expand upon the ornithomimosaurs from the Balve site in the future.
Acknowledgements
The excavations in Balve have been ongoing since 2002 with the kind permission of Lhoist Company. AHS sincerely appreciates Lhoist’s excellent cooperation and active support. From 2002 to 2016, the excavations were conducted by Klaus-Peter Lanser (formerly WMNM), along with preparators and students. Since 2017, AHS has continued the work with the assistance of WMNM preparators, as well as numerous dedicated student volunteers, interns, trainees, and other volunteers. We extend our heartfelt gratitude to all of them, with special thanks to Jerome Gores (WMNM) for his outstanding contributions to the success of recent excavations. Moreover, we would like to sincerely thank Christian Foth (Museum für Naturkunde Berlin, Germany) for his input, as he, independently of us, came to the conclusion that the manual ungual WMNM P117116 is ornithomimosaurian. We also thank Elena Cuesta (Ludwig-Maximilians-Universität, Munich, Germany) for correspondence. Finally, we are grateful for suggestions made by Bradley McFeeters (Carleton University, Ottawa, Canada) and an anonymous reviewer, which helped greatly with improving the quality of the manuscript.
Editor: Daniel Barta
References
Allain, R., Vullo, R., Le Loeuff, J., and Tournepiche, J.-F. 2014. European ornithomimosaurs (Dinosauria, Theropoda): an undetected record. Geologica Acta 12: 127–135.
Allain, R., Vullo, R., Rozada, L., Anquetin, J., Bourgeais, R., Goedert, J., Lasseron, M., Martin, J.E., Pérez-García, A., Peyre de Fabrègues, C., Royo-Torres, R., Augier, D., Bailly, G., Cazes, L., Despres, Y., Gailliègue, A., Gomez, B., Goussard, F., Lenglet, T., Vacant, R., Mazan, and Tournepiche, J.-F. 2022. Vetrebrate paleobiodiversity of the Early Cretaceous (Berriasian) Angeac-Charente Lagerstätte (southwestern France): implications for continental faunal turnover at the J/K boundary. Geodiversitas 44: 683–752. Crossref
Atkins-Weltman, K.L., Simon, D.J., Woodward, H.N., Funston, G.F., and Snively, E. 2024. A new oviraptorosaur (Dinosauria: Theropoda) from the end-Maastrichtian Hell Creek Formation of North America. PLOS ONE 19 (1): e0294901. Crossref
Averianov, A.O. and Lopatin, A.V. 2022. A re-appraisal of Parvicursor remotus from the Late Cretaceous of Mongolia: implications for the phylogeny and taxonomy of alvarezsaurid theropod dinosaurs. Journal of Systematic Palaeontology 19: 1097–1128. Crossref
Averianov, A.O., Sues, H.-D., Dyke, G., and Bayshashov, B. 2017. Hind limb bones of an ornithomimid dinosaur from the Upper Cretaceous Bostobe Formation, northeastern Aral Sea region, Kazakhstan. Palaeoworld 26: 194–201. Crossref
Barsbold, R. 1976. On the evolution and systematics of the late Mesozoic carnivorous dinosaurs [in Russian]. Paleontologiâ i biostratigrafiâ Mongolii. Trudy, Sovmestnaâ Sovetsko-Mongol’skaâ paleontologičeskaâ èkspediciâ 3: 68–75.
Barsbold, R. 1988. A new Late Cretaceous ornithomimid from the Mongolian People’s Republic [in Russian]. Paleontologičeskij žurnal 1988 (1): 122–125.
Barsbold, R. and Perle, A. 1984. On first new find of a primitive ornithomimosaur from the Cretaceous of the MPR [in Russian]. Paleontologičeskij žurnal 1984 (2): 121–123.
Bronowicz, R. 2011. New material of a derived ornithomimosaur from the Upper Cretaceous Nemegt Formation of Mongolia. Acta Palaeontologica Polonica 56: 477–488. Crossref
Brownstein, C.D. 2017. Description of Arundel Clay ornithomimosaur material and a reinterpretation of Nedcolbertia justinhofmanni as an “Ostrich Dinosaur”: biogeographic implications. PeerJ 5: e3110. Crossref
Buffetaut, E., Suteethorn, V., and Tong, H. 2009. An early ‘Ostrich Dinosaur’ (Theropoda: Ornithomimosauria) from the Early Cretaceous Sao Khua Formation of NE Thailand. Geological Society, London, Special Publications 315: 229–243. Crossref
Cau, A. and Paterna, A. 2025. Beyond the Stromer’s Riddle: the impact of lumping and splitting hypotheses on the systematics of the giant predatory dinosaurs from northern Africa. Italian Journal of Geosciences 144 (2): 1–24. Crossref
Chapelle, K.E., Norell, M., Ford, D.P., Hendrickx, C., Radermacher, V.J., Balanoff, A., Zanno, L.E., and Choiniere, J.N. 2021. A ct-based revised description and phylogenetic analysis of the skull of the basal maniraptoran Ornitholestes hermanni Osborn 1903. 81st Society of Vertebrate Paleontology Annual Meeting (virtual). [available onlin, https://vertpaleo.org/wp-content/uploads/2021/10/SVP_2021_VirtualBook_final.pdf]
Chinzorig, T., Kobayashi, Y., Khishigjav, T., Currie, P.J., Takasaki, R., Tanaka, T., Iijimaa, M., and Barsbold, R. 2018. Ornithomimosaurs from the Nemegt Formation of Mongolia: manus morphological variation and diversity. Palaeogeography, Palaeoclimatology, Palaeoecology 494: 91–100. Crossref
Chinzorig, T., Kobayashi, Y., Tsogtbaatar, K., Currie, P.J., Watabe, M., and Barsbold, R. 2017. First ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia. Scientific Reports 7: 5835. Crossref
Choiniere, J.N., Clark, J.M., Norell, M.A., and Xu, X. 2014. Cranial Osteology of Haplocheirus sollers Choiniere et al., 2010 (Theropoda: Alvarezsauroidea). American Museum Novitates 3816: 1–44. Crossref
Choiniere, J.N., Forster, C.A., and e Klerk, W.J. 2012. New information on Nqwebasaurus thwazi, a coelurosaurian theropod from the Early Cretaceous Kirkwood Formation in South Africa. Journal of African Earth Sciences 71: 1–17. Crossref
Choiniere, J.N., Xu, X., Clark, J.M., Forster, C.A., Guo, Y., and Han, F. 2010. A basal alvarezsauroid theropod from the Early Late Jurassic of Xinjiang, China. Science 327: 571–574. Crossref
Claessens, L.P.A.M. and Loewen, M.A. 2016. A redescription of Ornithomimus velox Marsh, 1890 (Dinosauria, Theropoda). Journal of Vertebrate Paleontology 36 (1): e1034593. Crossref
Cuesta, E., Vidal, D., Ortega, F., Shibata, M., and Sanz, J.L. 2022. Pelecanimimus (Theropoda: Ornithomimosauria) postcranial anatomy and the evolution of the specialized manus in Ornithomimosaurs and sternum in maniraptoriforms. Zoological Journal of the Linnean Society 194: 553–591. Crossref
Currie, P.J., Funston, G.F., and Osmólska, H. 2016. New specimens of the crested theropod dinosaur Elmisaurus rarus from Mongolia. Acta Palaeontologica Polonica 61: 143–157. Crossref
Drozdzewski, G., Hartkopf-Fröder, C., Lange, F.G., Oestreicher, B., Ribbert, K.H., Voigt, S., and Vrede, V. 1998. Vorläufige Mitteilung über unterkretazischen Tiefenkarst im Wülfrather Massenkalk (Rheinisches Schiefergebirge). Mitteilungen des Verbandes der deutschen Höhlen- und Karstforscher 44 (2): 53–66.
Drozdzewski, G., Richter, D.K., and Wrede, V. 2017. Hydrothermalkarst im nördlichen Rheinischen Schiefergebirge. Jahrbuch Karst und Höhle 2015–2017: 1–88.
Fregenal-Martínez, M., Meléndez, N., Muñoz-García, M.B., Elez, J., and de la Horra, R. 2017. The stratigraphic record of the Late Jurassic–Early Cretaceous rifting in the Alto Tajo-Serranía de Cuenca region (Iberian Ranges, Spain): genetic and structural evidences for a revision and a new lithostratigraphic proposal. Revista de la Sociedad Geológica de España 30: 113–142.
Funston, G. 2020. Caenagnathids of the Dinosaur Park Formation (Campanian) of Alberta, Canada: anatomy, osteohistology, taxonomy, and evolution. Vertebrate Anatomy Morphology Palaeontology 8: 105–153. Crossref
Funston, G.F., Currie, P.J., and Burns, M.E. 2016. New elmisaurine specimens from North America and their relationship to the Mongolian Elmisaurus rarus. Acta Palaeontologica Polonica 61: 159–173. Crossref
Funston, G.F., Currie, P.J., Ryan, M.J., and Dong, Z.-M. 2019. Birdlike growth and mixed-age flocks in avimimids (Theropoda, Oviraptorosauria). Scientific Reports 9: 18816. Crossref
Gauthier, J.A. 1986. Saurischian monophyly and the origin of birds. Memoirs of the California Academy of Sciences 8: 1–55.
Hao, M., Li, Z., Wang, Z., Wang, S., Ma, F., Qing, G., King, J.L., Pei, R., Zhao, Q., and Xu, X. 2025. A new oviraptorosaur from the Lower Cretaceous Miaogou Formation of western Inner Mongolia, China. Cretaceous Research 167: 106023. Crossref
Hartkopf-Fröder, C., Rust, J., Wappler, T., Friis, E.M., and Viehofen, A. 2012. Mid-Cretaceous charred fossil flowers reveal direct observation of arthropod feeding strategies. Biology Letters 8: 295–298. Crossref
Hattori, S., Shibata, M., Kawabe, S., Imai, T., Nishi, H., and Azuma, Y. 2023. New theropod dinosaur from the Lower Cretaceous of Japan provides critical implications for the early evolution of ornithomimosaurs. Scientific Reports 13 (1): 13842. Crossref
He, T., Wang, X.-L., and Zhou, Z.-H. 2008. A new genus and species of caudipterid dinosaur from the Lower Cretaceous Jiufotang Formation of Western Liaoning, China. Vertebrata Palasiatica 46 (3): 178–189.
Hendrickx, C., Hartman, S.A., and Mateus, O. 2015. An overview of non-avian theropod discoveries and classification. PalArch’s Journal of Vertebrate Palaeontology 12 (1): 1–73.
Holtz, T.R.Jr. 1994. The phylogenetic position of the Tyrannosauridae: Implications for theropod systematics. Journal of Paleontology 68: 1100–1117. Crossref
Holtz, T.R., Jr. 1995. The arctometatarsalian pes, an unusual structure of the metatarsus of Cretaceous Theropoda (Dinosauria: Saurischia). Journal of Vertebrate Paleontology 14: 480–519. Crossref
Holtz, T.R., Jr. 2004. Tyrannosauroidea. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria, 111–136. University of California Press, Berkeley. Crossref
Hone, D.W.E., Choiniere, J.N., Tan, Q., and Xu, X. 2013. An articulated pes from a small parvicursorine alvarezsauroid dinosaur from Inner Mongolia, China. Acta Palaeontologica Polonica 58: 453–458. Crossref
Hornung, J.H., Sachs, S., and Schwermann, A.H. 2023. The first record of sauropod dinosaurs from a palaeotopographical upland environment and its implications for megaherbivorous dinosaur faunal turnover in the Early Cretaceous of northwestern Europe. Geologie und Paläontologie in Westfalen 97: 1–34.
von Huene, F. 1914. Beiträge zur Geschichte der Archosaurier. Geologische und Paläontologische Abhandlungen 13 (7): 1–56.
Hunt, R.K. and Quinn, J.H. 2018. A new ornithomimosaur from the Lower Cretaceous Trinity Group of Arkansas. Journal of Vertebrate Paleontology 38 (1): e1421209. Crossref
Ikegami, N. 2016. The first record of an ornithomimosaurian dinosaur from the Upper Cretaceous of Japan. Historical Biology 28: 264–269. Crossref
Ji, Q., Norell, M.A., Makovicky, P.J., Gao, K.-Q., Ji, S., and Yuan, C. 2003. An early ostrich dinosaur and implications for ornithomimosaur phylogeny. American Museum Novitates 3420: 1–19. Crossref
Jin, L., Chen, C., and Godefroit, P. 2012. A new basal ornithomimosaur (Dinosauria: Theropoda) from the early Cretaceous Yixian formation, Northeast China. In: P. Godefroit (ed.), Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems, 467–505. Indiana University Press Bloomington, Indiana.
Joyce, W.G., Evers, S.W., Ren, S., Rollot, Y., and Schwermann, A.H. 2023. The helochelydrid turtle Helochelydra nopcsai from the Early Cretaceous (late Barremian–early Aptian) fissure fills of Balve, North Rhine-Westphalia, Germany, including a large sample of granicones. Fossil Record 26: 117–133. Crossref
Karhu, A.A. and Rautian, A.S. 1996. A new family of Maniraptora (Dinosauria: Saurischia) from the Late Cretaceous of Mongolia. Paleontological Journal 30: 583–592.
Kirkland, J.I., Britt, B.B., and Whittle, C.H. 1998. A small coelurosaurian theropod from the Yellow Cat Member of the Cedar Mountain Formation (Lower Cretaceous, Barremian) of eastern Utah. New Mexico Museum of Natural History and Science Bulletin 14: 239–248.
Kobayashi, Y. and Barsbold, R. 2005. Anatomy of Harpymimus okladnikovi Barsbold and Perle 1984 (Dinosauria; Theropoda) of Mongolia. In: K. Carpenter (ed.), The Carnivorous Dinosaurs, 97–126. Indiana University Press, Bloomington.
Kobayashi, Y. and Barsbold, R. 2006. Ornithomimids from the Nemegt Formation of Mongolia. Journal of the Paleontological Society of Korea 22: 195–207.
Kobayashi, Y. and Lü, J.C. 2003. A new ornithomimid dinosaur with gregarious habits from the Late Cretaceous of China. Acta Palaeontologica Polonica 48: 235–259.
Kubo, K., Kobayashi, Y., Chinzorig, T., and Tsogtbaatar, K. 2023. A new alvarezsaurid dinosaur (Theropoda, Alvarezsauria) from the Upper Cretaceous Baruungoyot Formation of Mongolia provides insights for bird-like sleeping behavior in non-avian dinosaurs. PLOS ONE 18 (11): e0293801. Crossref
Kubota, K., Kobayashi, Y., and Ikeda, T. 2024. Early Cretaceous troodontine troodontid (Dinosauria: Theropoda) from the Ohyamashimo Formation of Japan reveals the early evolution of Troodontinae. Scientific Reports 14: 16392. Crossref
Kurzanov, S.M. and Osmólska, H. 1991. Tochisaurus nemegtensis gen. et sp. n., a new troodontid (Dinosauria, Theropoda) from Mongolia. Acta Palaeontologica Electronica 36: 69–76.
Lanser, K.-P. 2011. Raubsaurier aus einer Verkarstung im devonischen Massenkalk im Hönnetal bei Balve. Archäologie In Westfalen-Lippe 2010: 18–20.
Lanser, K.-P. 2015. Nachweise von Pterosauriern aus einer unterkreidezeitlichen Karstfüllung im nördlichen Sauerland (Rheinisches Schiefergebirge, Deutschland). Geologie und Paläontologie in Westfalen 87: 93–117.
Lanser, K.-P. and Heimhofer, U. 2015. Evidence of theropod dinosaurs from a Lower Cretaceous karst filling in the northern Sauerland (Rhenish Massif, Germany). Paläontologische Zeitschrift 89: 79–94. Crossref
Lee, S., Lee, Y.-N., Park, J.-Y., Kim, S.-H., Badamkhatan, Z., Idersaikhan, D., and Tsogtbaatar, K. 2024. The first troodontid (Dinosauria: Theropoda) from the Upper Cretaceous Baruungoyot Formation of Mongolia. Journal of Vertebrate Paleontology 43 (6): e2364746. Crossref
Lee, Y.-N., Barsbold, R., Currie, P.J., Kobayashi, Y., Lee, H.-J., Godefroit, P., Escuillié, F., and Chinzorig, T. 2014. Resolving the long-standing enigmas of a giant ornithomimosaur Deinocheirus mirificus. Nature 515: 257–260. Crossref
Lydekker, R. 1889. Note on some points in the nomenclature of fossil reptiles and amphibians, with preliminary notices of two new species. Geological Magazine (Decade 3) 6: 325–326. Crossref
Macdonald, I. and Currie, P.J. 2019. Description of a partial Dromiceiomimus (Dinosauria: Theropoda) skeleton with comments on the validity of the genus. Canadian Journal of Earth Sciences 56: 129–157. Crossref
Makovicky, P.J., Kobayashi, Y., and Currie, P.J. 2004. Ornithomimosauria. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria, 137–150. University of California Press, Berkeley. Crossref
Makovicky, P.J., Li, D., Gao, K.-Q., Lewin, M., Erickson, G.M., and Norell, M.A. 2010. A giant ornithomimosaur from the Early Cretaceous of China. Proceedings of the Royal Society B: Biological Sciences 277: 191–198. Crossref
Marsh, O.C. 1881. Principal characters of American Jurassic dinosaurs. Part V. The American Journal of Science and Arts, Series 3 21: 417–423. Crossref
Marsh, O.C. 1890. Description of new dinosaurian reptiles. The American Journal of Science, Series 3 39: 81–86. Crossref
Martin, T., Averianov, A.O., Schultz, J.A., and Schwermann, A.H. 2021. First multituberculate mammals from the Lower Cretaceous of Germany. Cretaceous Research 119: 104699. Crossref
Martin, T., Averianov, A.O., Schultz, J.A., and Schwermann, A.H. 2022a. A stem therian mammal from the Lower Cretaceous of Germany. Journal of Vertebrate Paleontology 42 (6): e2224848. Crossref
Martin, T., Averianov, A., Schultz, J., Schellhorn, R., and Schwermann, A. 2022b. First spalacotheriid and dryolestid mammals from the Cretaceous of Germany. Acta Palaeontologica Polonica 67: 155–175. Crossref
McFeeters, B. 2015. Evolution and Diversity of Ornithomimid Dinosaurs in the Upper Cretaceous Belly River Group of Alberta. 253 pp. Master’s thesis, Carleton University Ottawa, Ontario.
McFeeters, B., Ryan, M.J., Schröder-Adams, C., and Cullen, T.M. 2016. A new ornithomimid theropod from the Dinosaur Park Formation of Alberta, Canada. Journal of Vertebrate Paleontology 36 (6): e1221415. Crossref
Naish, D. and Cau, A. 2022. The osteology and affinities of Eotyrannus lengi, a tyrannosauroid theropod from the Wealden Supergroup of southern England. PeerJ 10: e12727. Crossref
Naish, D. and Dyke, G.J. 2004. Heptasteornis was no ornithomimid, troodontid, dromaeosaurid or owl: the first alvarezsaurid (Dinosauria: Theropoda) from Europe. Neues Jahrbuch für Geologie und Paläontologie-Monatshefte 2004 (7): 385–4001. Crossref
Naish, D. and Martill, D.M. 2002. A reappraisal of Thecocoelurus daviesi (Dinosauria: Theropoda) from the Early Cretaceous of the Isle of Wight. Proceedings of the Geologists’ Association 113: 23–30. Crossref
Néraudeau, D., Allain, R., Ballèvre, M., Batten, D.J., Buffetaut, E., Colin, J.P., Dabard, M.P., Daviero-Gomez, V., El Albani, A., Gomez, B., Grosheny, D., Le Lœuff, J., Leprince, A., Martín-Closas, C., Masure, E., Mazin, J.-M., Philippe, M., Pouech, J., Tong, H., Tournepiche, J.F., and Vullo, R. 2012. The Hauterivian–Barremian lignitic bone bed of Angeac (Charente, south-west France): Stratigraphical, palaeobiological and palaeogeographical implications. Cretaceous Research 37: 1–14. Crossref
Nicholls, E.L. and Russell, A.P. 1985. Structure and function of the pectoral girdle and forelimb of Struthiomimus altus (Theropoda: Ornithomimidae). Palaeontology 28: 619–628.
Norell, M.A., Makovicky, P.J., Bever, G.S., Balanoff, A.M., Clark, J.M., Barsbold, R., and Rowe, T. 2009. A Review of the Mongolian Cretaceous Dinosaur Saurornithoides (Troodontidae: Theropoda). American Museum Novitates 3654: 1–63. Crossref
Norman, D.B. 1987. A mass-accumulation of vertebrates from the Lower Cretaceous of Nehden (Sauerland), West Germany. Proceedings of the Royal Society of London B. Biological Sciences 230: 215–255. Crossref
Nottrodt, R.E. 2022. First articulated ornithomimid specimens from the upper Maastrichtian Scollard Formation of Alberta, Canada. Journal of Vertebrate Paleontology 41 (5): e2019754. Crossref
Oekentorp, K. 1984. Die Saurierfundstelle Brilon-Nehden (Rheinisches Schiefergebirge) und das Alter der Verkarstung. Kölner Geographische Arbeiten 45: 293–315.
Osmólska, H. and Roniewicz, E. 1970. Deinocheiridae, a new family of theropod dinosaurs. Palaeontologica Polonica 21: 5–19.
Osmólska, H., Roniewicz, E., and Barsbold, R. 1972. A new dinosaur, Gallimimus bullatus n. gen., n. sp. (Ornithomimidae) from the Upper Cretaceous of Mongolia. Palaeontologica Polonica 27: 103–143.
Owen, R. 1842. Report on British fossil reptiles, part II. Report of the British Association for the Advancement of Science 11: 60–204.
PaleobioDB contributors 2025. The Paleobiology Database [downloaded from https://paleobiodb.org/ on 17 April 2025].
Parks, W.A. 1933. New species of dinosaurs and turtles from the Upper Cretaceous formations of Alberta. University of Toronto Studies, Geological Series 34: 3.
Pérez-Moreno, B.P., Luis Sanz, J., Buscalioni, A.D., Moratalla, J.J., Ortega, F., and Rasskin-Gutman, D. 1994. A unique multitoothed ornithomimosaur dinosaur from the Lower Cretaceous of Spain. Nature 370: 363–367. Crossref
Perle, A., Chiappe, L.M., Barsbold, R., Clark, J.M., and Norell, M.A. 1994. Skeletal morphology of Mononykus olecranus (Theropoda: Avialae) from the Late Cretaceous of Mongolia. American Museum Novitates 3105: 1–29.
Qiang, J., Currie, P.J., Norell, M.A., and Shu-An, J. 1998. Two feathered dinosaurs from northeastern China. Nature 393: 753–761. Crossref
Qiu, R., Wang, X., Wang, Q., Li, N., Zhang, J., and Ma, Y. 2019. A new caudipterid from the Lower Cretaceous of China with information on the evolution of the manus of Oviraptorosauria. Scientific Reports 9: 6431. Crossref
Richter, D.K., Götte, T., Stritzke, R., and Mutterlose, J. 2014. Eine miozäne Hohlraumfüllung im Massenkalk der östlichen Iserlohner Kalksenke. Geologie und Paläontologie in Westfalen 86: 5–17.
Russell, D.A. and Dong, Z.-M. 1993. A nearly complete skeleton of a new troodontid dinosaur from the Early Cretaceous of the Ordos Basin, Inner Mongolia, People’s Republic of China. Canadian Journal of Earth Sciences 30: 2163–2173. Crossref
Samathi, A. 2024. Phylogenetic position of Kinnareemimus khonkaenensis (Dinosauria: Theropoda: Ornithomimosauria) from the Lower Cretaceous of Thailand. Zootaxa 5448: 67–84. Crossref
Seeley, H.G. 1888. On Thecospondylus daviesi (Seeley), with some remarks on the Classification of the Dinosauria. Quarterly Journal of the Geological Society of London 44: 79–87. Crossref
Sellés, A.G., Vila, B., Brusatte, S.L., Currie, P.J., and Galobart, À. 2021. A fast-growing basal troodontid (Dinosauria: Theropoda) from the latest Cretaceous of Europe. Scientific Reports 11 (1): 4855. Crossref
Sereno, P.C. 2017. Early Cretaceous ornithomimosaurs (Dinosauria: Coelurosauria) from Africa. Ameghiniana 54: 576–616. Crossref
Serrano-Brañas, C.I., Espinosa-Chávez, B., Augusta Maccracken, S., Gutiérrez-Blando, C., de León-Dávila, C., and Flores Ventura, J. 2020. Paraxenisaurus normalensis, a large deinocheirid ornithomimosaur from the Cerro del Pueblo Formation (Upper Cretaceous), Coahuila, Mexico. Journal of South American Earth Sciences 101: 102610. Crossref
Serrano-Brañas, C.I., Torres-Rodríguez, E., Reyes-Luna, P.C., González-Ramírez, I., and González-León, C. 2016. A new ornithomimid dinosaur from the Upper Cretaceous Packard Shale formation (Cabullona Group) Sonora, México. Cretaceous Research 58: 49–62. Crossref
Shen, C., Gao, C., Lü, J., and Kundrát, M. 2017. A new troodontid dinosaur (Liaoningvenator curriei gen. et sp. nov.) from the Early Cretaceous Yixian Formation in Western Liaoning Province. Acta Geoscientica Sinica 38: 359–371.
Skutschas, P.P., Kolchanov, V.V., and Schwermann, A.H. 2020. First salamander from the Lower Cretaceous of Germany. Cretaceous Research 116: 104606. Crossref
Smith, D. and Galton, P. 1990. Osteology of Archaeornithomimus asiaticus (upper Cretaceous, Iren Dabasu Formation, People’s Republic of China). Journal of Vertebrate Paleontology 10: 255–265. Crossref
Snively, E., Russell, A.P., and Powell, G.L. 2004. Evolutionary morphology of the coelurosaurian arctometatarsus: descriptive, morphometric and phylogenetic approaches. Zoological Journal of the Linnean Society 142: 525–553. Crossref
Sternberg, C.M. 1933. A new Ornithomimus with complete abdominal cuirass. The Canadian Field-Naturalist 47 (5): 79–83. Crossref
Turner, A.H., Nesbitt, S.J., and Norell, M.A. 2009. A large alvarezsaurid from the Late Cretaceous of Mongolia. American Museum Novitates 3648: 1–14. Crossref
Varricchio, D.J., Hogan, J.D., and Gardner, J.D. 2025. Troodontid specimens from the Cretaceous Two Medicine Formation of Montana (USA) and the validity of Troodon formosus. Journal of Paleontology 99: 219–240. Crossref
Vullo, R., Neraudeau, D., and Lenglet, T. 2007. Dinosaur teeth from the Cenomanian of Charentes, Western France: evidence for a mixed Laurasian–Gondwanan assemblage. Journal of Vertebrate Paleontology 27: 931–943. Crossref
White, M.A. 2009. The subarctometatarsus: Intermediate metatarsus architecture demonstrating the evolution of the arctometatarsus and advanced agility in theropod dinosaurs. Alcheringa 33: 1–21. Crossref
Wirth, W. 1970. Eine tertiärzeitliche Karstfüllung bei Eisborn im Sauerland. Fortschritte in der Geologie von Rheinland und Westfalen 7: 403–420.
Xu, L., Kobayashi, Y., Lü, J., Lee, Y.-N., Liu, Y., Tanaka, K., Zhang, X., Jia, S., and Zhang, J. 2011a. A new ornithomimid dinosaur with North American affinities from the Late Cretaceous Qiupa Formation in Henan Province of China. Cretaceous Research 32: 213–222. Crossref
Xu, X. and Wang, X. 2004. A new troodontid (Theropoda: Troodontidae) from the Lower Cretaceous Yixian Formation of Western Liaoning, China. Acta Geologica Sinica 78: 22–26. Crossref
Xu, X., Choiniere, J., Tan, Q., Benson, R.B.J., Clark, J., Sullivan, C., Zhao, Q., Han, F., Ma, Q., He, Y., Wang, S., Xing, H., and Tan, L. 2018. Two Early Cretaceous fossils document transitional stages in alvarezsaurian dinosaur evolution. Current Biology 28: 2853–2860. Crossref
Xu, X., Currie, P.J., Pittman, M., Xing, L., Meng, Q., Lü, J., Hu, D., and Yu, C. 2017. Mosaic evolution in an asymmetrically feathered troodontid dinosaur with transitional features. Nature Communications 8: 14972. Crossref
Xu, X., Norell, M.A., Wang, X., Makovicky, P.J., and Wu, X. 2002. A basal troodontid from the Early Cretaceous of China. Nature 415: 780–784. Crossref
Xu, X., Sullivan, C., Pittman, M., Choiniere, J.N., Hone, D.W.E., Upchurch, P., Tan, Q., Xiao, D., Tan, L., and Han, F. 2011b. A monodactyl nonavian dinosaur and the complex evolution of the alvarezsauroid hand. Proceedings of the National Academy of Sciences 108: 2338–2342. Crossref
Xu, X., Upchurch, P., Ma, Q., Pittman, M., Choiniere, J.N., Sullivan, C., Hone, D.W.E., Tan, Q., Tan, L., Xiao, D., and Han, F. 2013. Osteology of the Late Cretaceous alvarezsauroid Linhenykus monodactylus from China and comments on alvarezsauroid biogeography. Acta Palaeontologica Polonica 58: 25–46.
Xu, X., Wang, D.-Y., Sullivan, C., Hone, D.W.E., Han, F.-L., Yan, R.-H., and Du, F.-M. 2010. A basal parvicursorine (Theropoda: Alvarezsauridae) from the Upper Cretaceous of China. Zootaxa 2413: 1–19. Crossref
Xu, X., Zhao, Q., Sullivan, C., Tan, Q.-W., Sander, M., and Ma, Q.-Y. 2012. The taxonomy of the troodontid IVPP V 10597 reconsidered. Vertebrata Palasiatica 50 (2): 140–150.
Yu, Y., Wang, K., Chen, S., Sullivan, C., Wang, S., Wang, P., and Xu, X. 2018. A new caenagnathid dinosaur from the Upper Cretaceous Wangshi Group of Shandong, China, with comments on size variation among oviraptorosaurs. Scientific Reports 8: 5030. Crossref
Zanno, L.E., Varricchio, D.J., O’Connor, P.M., Titus, A.L., and Knell, M.J. 2011. A new troodontid theropod, Talos sampsoni gen. et sp. nov., from the Upper Cretaceous Western Interior Basin of North America. PLoS ONE 6 (9): e24487. Crossref
Zhong, Y., Huyskens, M.H., Yin, Q.-Z., Wang, Y., Ma, Q., and Xu, Y.-G. 2021. High-precision geochronological constraints on the duration of ‘Dinosaur Pompeii’ and the Yixian Formation. National Science Review 8 (6): nwab063. Crossref
Acta Palaeontol. Pol. 70 (3): 543–555, 2025
https://doi.org/10.4202/app.01262.2025