
New Katian bivalves from the Upper Ordovician Xiazhen Formation, Jiangxi Province, China
Yong Wang, Ren-bin Zhan, and Zong-jie Fang
Wang, Y., Zhan, R.-B., and Fang, Z.-J. 2026. New Katian bivalves from the Upper Ordovician Xiazhen Formation, Jiangxi Province, China. Acta Palaeontologica Polonica 71 (1): 133–153.
A bivalve association reported herein was recently discovered from the Xiazhen Formation (upper Katian, Upper Ordovician) at Zhuzhai, southern Yushan County, northeastern Jiangxi Province, East China. Nine genera and ten species are systematically documented including four new species: Palaeoneilo rectus sp. nov. (most abundant), Palaeoneilo cf. constricta, Goniophorina volvens, Modiolopsis elegantulus, Cyrtodontula sp., Cuneamya? sp., Trigonoconcha brevis sp. nov., Similodonta minor sp. nov., Concavodonta varius sp. nov., and Paulinea cf. parva. The association is mainly composed of Protobranchia (Nuculiformii and Nuculaniformii) and subordinate Pteriomorphia, suggesting a relatively low palaeolatitudinal setting. The dominance of infaunal protobranch bivalves is notable, and against the trend of rapid diversification of pteriomorphs during the Late Ordovician. This discrepancy is likely attributable to a cold water tongue extending from then South Pole to South China palaeoplate, and the ecological pressure from those much more abundant brachiopods on the substrate surface, both hindering the development of epifaunal pteriomorphs in the region. Significantly, this bivalve association shares several common genera with its coeval bivalve fauna from the Oslo region, Norway.
Key words: Bivalvia, Protobranchia, Pteriomorphia, epifauna, infauna, Katian, Late Ordovician, Jiangxi, South China.
Yong Wang [yongwang@nigpas.ac.cn; ORCID:https://orcid.org/0000-0002-3253-2242], and Ren-Bin Zhan [rbzhan@nigpas.ac.cn; ORCID: https://orcid.org/0000-0003-3541-9023], State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, China; University of the Chinese Academy of Sciences, Beijing 100049, China.
Zong-Jie Fang [zjfang@nigpas.ac.cn; ORCID: https://orcid.org/0000-0002-0254-5077], State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, China.
Received 11 August 2025, accepted 11 January 2026, published online 26 March 2026.
Copyright © 2026 Y. Wang et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
During the Sandbian (Late Ordovician), the radiation of pteriomorph bivalves took place as part of the Great Ordovician Biodiversification Event (GOBE) (Cope 2004; Fang 2006). The Late Ordovician bivalve faunas were taxonomically diverse especially at low to middle palaeolatitudes (Bretsky 1969, 1970a, b; Toni 1975; Frey 1987; Tunnicliff 1982; Cope 1996; Ebbestad et al. 2022).
The bivalve association reported in this paper occurs in the middle part of the Xiazhen Formation (upper Katian, Upper Ordovician) at the Zhuzhai section of southern Yushan County, northeastern Jiangxi Province, East China (Fig. 1B, C), which was palaeogeographically near the Cathaysian Block (Fig. 1A). The Xiazhen Formation at this section comprises limestone, mudstone, and calcareous mudstone interbeds (Fig. 2). Abundant shelly fossils, such as brachiopods, corals, bryozoans, stromatoporoids and trilobites, have been found and systematically studied by various experts (e.g., Hu 1986; Zhan and Rong 1994, 1995a, b; Zhan et al. 2002; Lee 2013; Zhang et al. 2018; Jeon et al. 2022). Bivalves found in this formation are quite species-rich, although not as many as brachiopods. The bivalve association from the Xiazhen Formation provides an important insight into the Late Ordovician bivalve fauna of the South China palaeoplate, and is a valuable case study for this time interval.
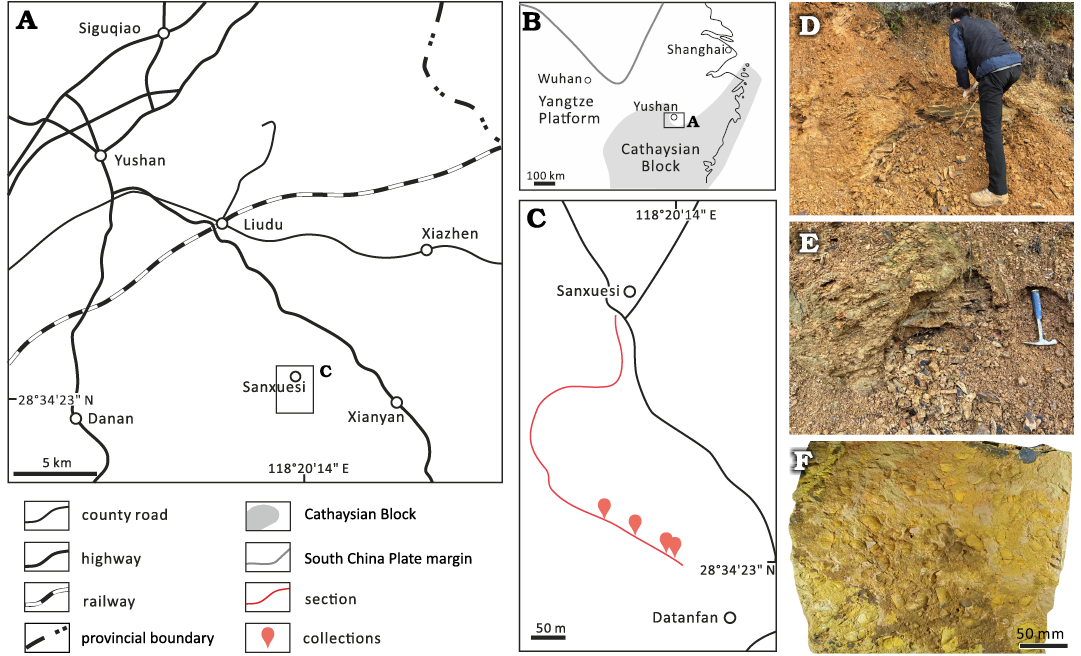
Fig. 1. Map showing geographical (A) and palaeogeographical (B) positions of Yushan during Katian (Late Ordovician). Location of the lower part of the Zhuzhai Section and collection sites (C). Outcrops of the Xiazhen Formation (D) and its front photo (E). Surface of a slab where the bivalves were collected (F), showing the dominant brachiopods of this benthic shelly fauna.
Yushan is located in the Jiangshan-Changshan-Yushan district (abbreviated as JCY area). This is one of the classical areas for the Palaeozoic research and includes two GSSPs, i.e., the base of the Jiangshanian Stage (upper Cambrian) at Duibian of Jiangshan County and the base of the Darriwilian Stage (Middle Ordovician) at Huangnitang of Changshan County respectively. The Zhuzhai section (Fig. 1, GPS of collection JYZ-10: 28°34′23″N, 118°20′14″E) exposes the upper Katian (Upper Ordovician) Xiazhen Formation, one of the best representatives of this formation in JCY area. Zhan and Fu (1994) summarized three contemporaneous lithostratigraphic units for the upper Katian in JCY area, including the Changwu Formation (on the Zhexi slope), the Sanqushan Formation (along the platform margin, or in the transition belt between the Zhe-Gan platform and the Zhexi slope), and the Xiazhen Formation (on the Zhe-Gan platform). The sediments cropping out at Zhuzhai section were deposited in the nearshore side of the Zhe-Gan platform. Zhan and Rong (1995a) recognized four brachiopod dominated communities from the Xiazhen Formation at this site, such as the Protospirifer praecursor Community, the Deloprosopus jiangshanensis Community, the Altaethyrella-Ovalospira Community, and the Sowerbyella-Antizygospira Community. Four communities alternately recur several times throughout the Xiazhen Formation at this section, indicating the fluctuating environmental conditions.
Institutional abbreviations.—NIGP, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, China.
Other abbreviations.—ES, eastern part of the section JCY, Jiangshan-Changshan-Yushan; JYZ, Jiangxi-Yushan-Zhuzhai.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in Zoobank: urn:lsid:zoobank.org:pub:E42CE936-29B9-450D-8997-B16F9C00358E.

Fig. 2. Lithological column of the Xiazhen Formation and the stratigraphical range of the bivalves (modified from Zhan and Fu 1994).
Material and methods
The bivalves studied here were collected from the mudstone of the middle part of the Zhuzhai section, together with the brachiopods of the Sowerbyella-Antizygospira Community, which were thought to inhabit a nearshore, shallow water environment corresponding to muddy substrate with benthic assemblage 2–3 (Rong and Han 1986; Rong and Chen 1987; Zhan and Rong 1995a; Zhan et al. 2002). Bivalves were collected from four horizons: ES-a, ES-b (JYZ-9), JYZ-11 and ES-c (JYZ-10). Among these, ES-c yields the most abundant and diverse bivalve fossils. The Xiazhen Formation at this site is biostratigraphically assigned to the Dicellograptus complexus Biozone (upper Katian) (Chen et al. 1987; Zhan and Cocks 1998; Zhan et al. 2002). Most of the bivalves are disarticulated, and those articulated are mainly from the collection ES-c (JYZ-10). The valves are commonly complete and have no distinct orientation or sorting, indicating minimal transportation. All figured specimens are stored at Nanjing Institute of Geology and Palaeontology (NIGP), Chinese Academy of Sciences. The systematic palaeontology in this work primarily follows Carter et al. (2011).
Systematic palaeontology
Class Bivalvia Linnæus, 1758
Subclass Protobranchia Pelseneer, 1889
Superorder Nuculiformii Dall, 1889
Order Nuculida Dall, 1889
Superfamily Nuculoidea Gray, 1824
Family Praenuculidae McAlester, 1969
Subfamily Praenuculinae McAlester, 1969
Genus Paulinea Cope, 1996
Type species: Paulinea parva Cope, 1996; Ogof Hên Formation, Lower Ordovician, South Wales, UK.
Paulinea cf. parva Cope, 1996
Figs. 3, 4.
cf. 1996 Paulinea parva gen. et sp. nov.; Cope 1996: 985, pl. 1: 11–22.
Material.—Five internal and three external moulds of right valves, two internal and two external moulds of left valves (NIGP208861–208867). All from ES-c (JYZ-10), Yushan County, Jiangxi Province (China); Katian (Late Ordovician).
Description.—Small sub-rectangular shell; equivalve and inequilateral. The length/height ratio (L/H) is approximately 1.5 (Fig. 3; Table 1). Anterior margin round; posterior margin inclined truncate; ventral margin broadly convex, with a slight sinus near posteroventral corner; posteroventral angle approximately 90°. Umbo broad and situated at third anterior of the shell, slightly prosogyrate and projected above the hinge line. Anterior adductor scar small and round with short myophoric buttress developed posteriorly. Hinge line with graded series of peg-like taxodont teeth, at least four anterior and eight posterior, increasing slightly in size distally. Ligament opisthodetic, ligament groove visible (Fig. 4C2). Outer shell surface smooth (Fig. 4A2–E2).
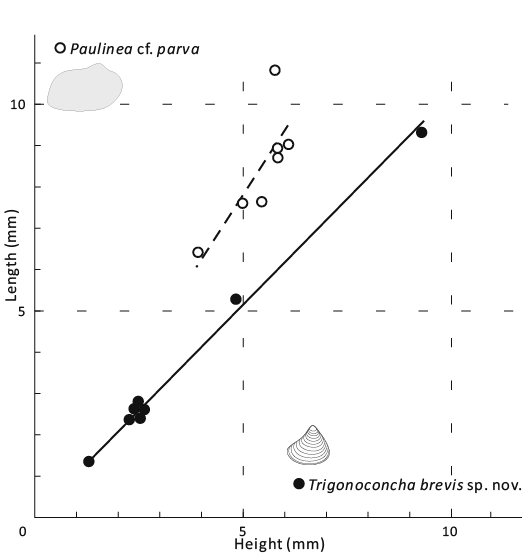
Fig. 3. Bivariate plot of shell morphometrics for praenuculid protobranch bivalve Paulinea cf. parva Cope, 1996 (hollow circles, thick dash line with slope approximately 1.56) and similodontid protobranch Trigonoconcha brevis sp. nov. (solid circles, thick line with slope approximately 1.03).
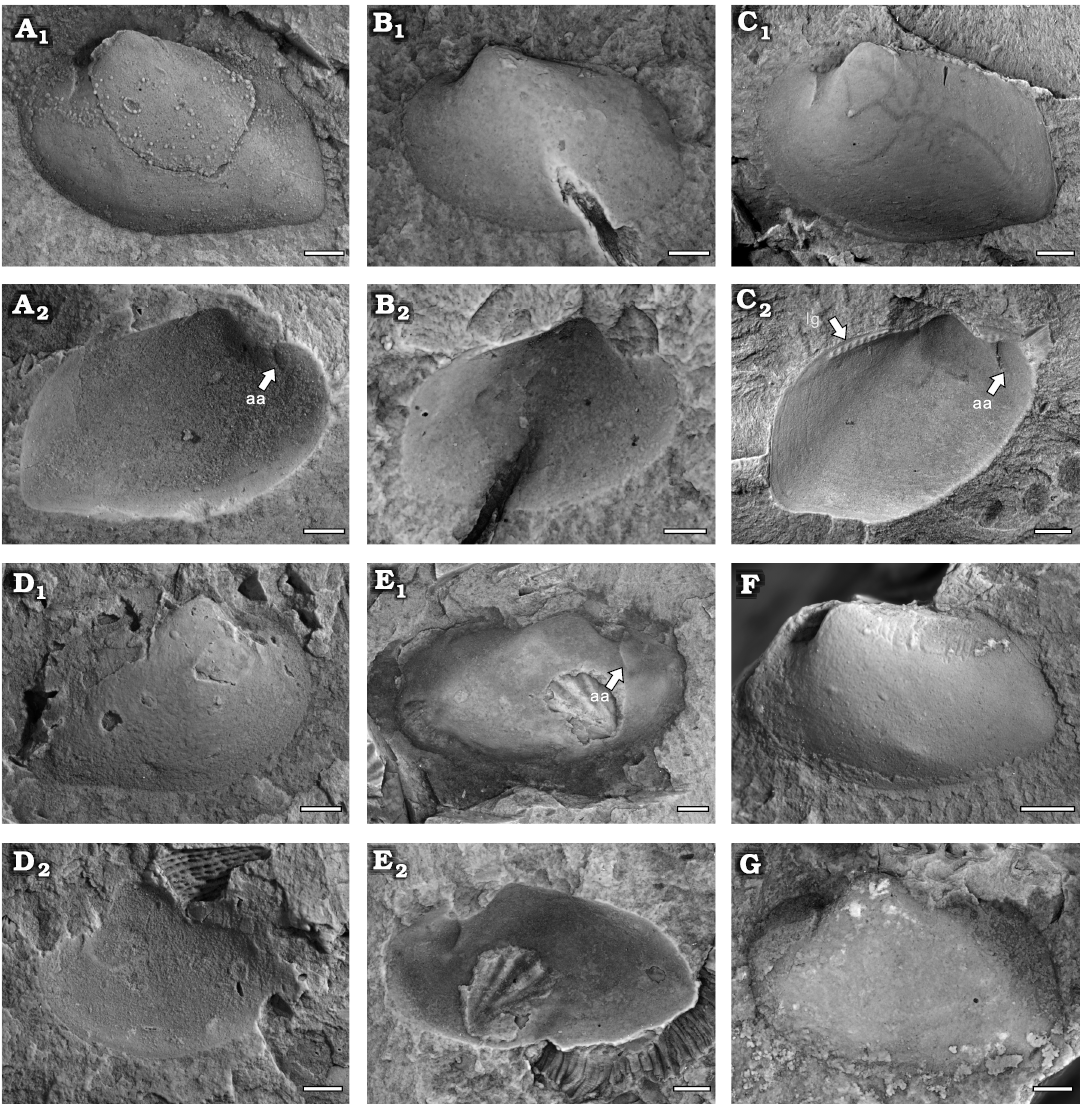
Fig. 4. Praenuculid protobranch bivalve Paulinea cf. parva Cope, 1996, Yushan County, Jiangxi Province (China); Katian (Late Ordovician). A. NIGP208861, left valve, internal (A1) and external (A2) moulds. B. NIGP208862, left valve, internal (B1) and external (B2) moulds. C. NIGP208863, left valve showing the dentition of this species, internal (C1) and external (C2) moulds. D. NIGP208864, right valve, internal (D1) and external (D2) moulds. E. NIGP208865, right valve, slightly compressed internal mould (E1) and external mould (E2). F. NIGP208866, left valve, internal mould. G. NIGP208867, left valve, internal mould. Abbreviations: aa, anterior adductor scar; lg, ligament. Scale bars 1 mm.
Table 1. Shell measurements (in mm) of Paulinea cf. parva Cope, 1996; L/H, length/height ratio (dimensionless).
|
Specimen number |
Length |
Height |
L/H |
|
NIGP208861 |
8.7 |
5.8 |
1.49 |
|
NIGP208864 |
7.6 |
5.4 |
1.40 |
|
NIGP208866 |
6.4 |
3.9 |
1.64 |
|
NIGP208863 |
9.0 |
6.1 |
1.48 |
|
NIGP208862 |
7.6 |
5.0 |
1.52 |
|
NIGP208867 |
8.9 |
5.8 |
1.53 |
|
NIGP208865 |
10.8 |
5.8 |
1.88 |
Remarks.—According to Sánchez’s (1999) definition of the Praenuculinae, most of the genera have convexodont teeth except for Pensarnia (Cope, 1996) and Paulinea. Pensarnia has a more distinct adductor scar and ornamentation than those observed in our specimens. Compared with the type species Paulinea parva, our specimens share a similar shell outline with some individuals of the type species (Cope 1996: pl. 1: 16–17). However, our specimens have a more significantly truncated posterior margin. As the dentition in our specimens is unclear, it seems reasonable to assign our specimens to Paulinea cf. parva, rather than to erect a new species.
Subfamily Concavodontinae Sánchez, 1999
Genus Concavodonta Babin & Melou, 1972
Type species: Nucula ponderata Barrande, 1881, Upper Ordovician, Czech Republic.
Concavodonta varius sp. nov.
Figs. 5, 6.
Zoobank LSID: urn:lsid:zoobank.org:act:9DE10739-775D-4D93-A160- 77A4669542BC.
Etymology: From Latin varius, various, indicating the changes in shell forms within the ontogeny.
Type material: Holotype, NIGP208868 (Fig. 5A), internal and external moulds of articulated valves. Paratypes: NIGP208869 (Fig. 5B); NIGP208870 (Fig. 5C); NIGP208873 (Fig. 5D); NIGP208871 (Fig. 5E); NIGP208872 (Fig. 5F).
Type locality: Yushan County, Jiangxi Province, East China.
Type horizon: Katian, Upper Ordovician.
Material.—Seven internal moulds (four right valves, two left valves, one conjoined valve), one external mould of conjoined valve (NIGP208868–208873). From JYZ-11 and ES-c (JYZ-10), Yushan County, Jiangxi Province (China); Katian (Late Ordovician).
Diagnosis.—Concavodonta with sub-rectangular shell in adults but oval in juveniles.
Description.—Small to medium sub-rectangular shell; equivalve and inequilateral. Anterior margin rounded; posterior margin truncated; ventral margin convex; anterior dorsal margin concave; posterior dorsal margin straight. The L/H varies from 1.24 to 1.86, with juvenile shells being less elongated posteriorly and having a smaller L/H (Fig. 6; Table 2). Broad umbo situated in anterior third of the shell, prosogyrate. Adductor scars ill-defined, a faint oval anterior adductor scar partially in one specimen (Fig. 5F). Taxodont dentition with concavodont teeth: six–seven anterior teeth and 11 posterior teeth in adult shells; three–four anterior teeth and eight posterior teeth in juvenile shells. An edentulous area between anterior and posterior teeth below the umbo. Anterior and posterior dorsal borders meet at angle of 160°. Shell ornamentation of fine commarginal growth lines.
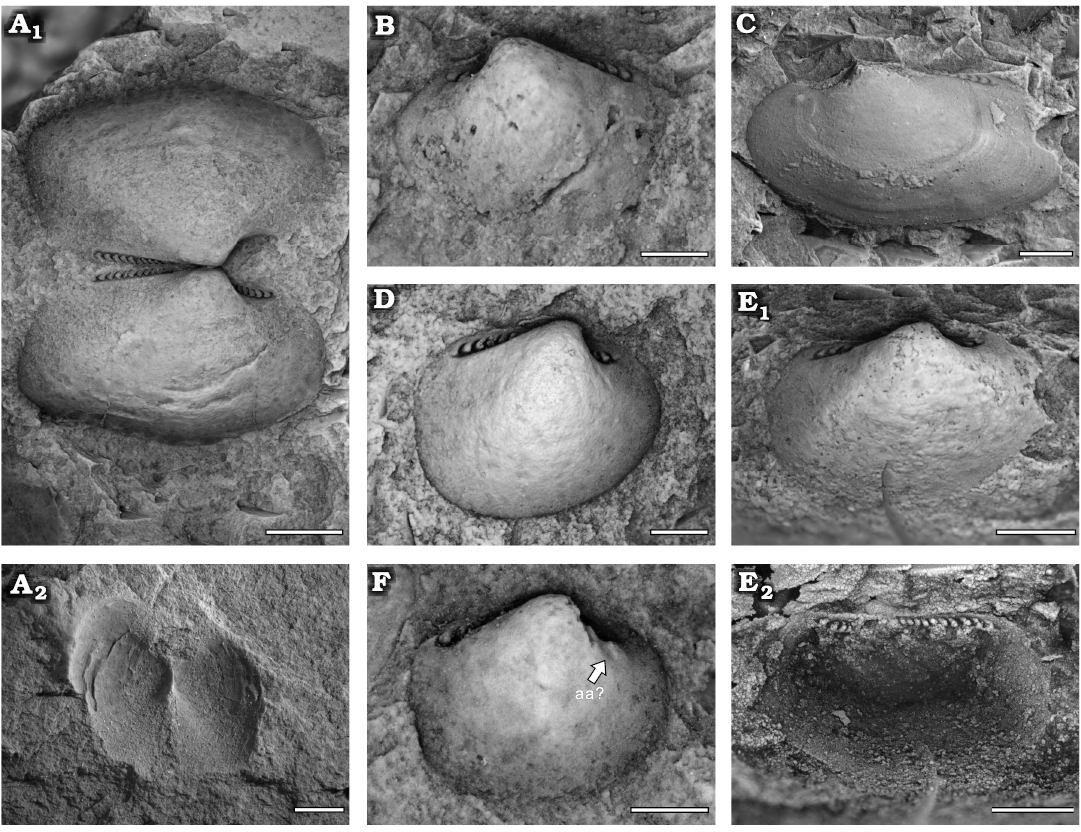
Fig. 5. Praenuculid protobranch bivalve Concavodonta varius sp. nov., Yushan County, Jiangxi Province (China); Katian (Late Ordovician). A. NIGP208868 (holotype), conjoined shell, internal (A1) and incomplete external (A2) moulds. B. NIGP208869 (paratype), left valve, internal mould. C. NIGP208870, paratype, left valve, internal mould. D. NIGP208873 (paratype), right valve, internal mould. E. NIGP208871 (paratype), right valve, internal mould (E1) and its latex cast (E2). F. NIGP208872 (paratype), right valve, internal mould. Abbreviation: aa, anterior adductor scar. Scale bars A, 2 mm; C, E, 1 mm; B, D, F, 0.5 mm.
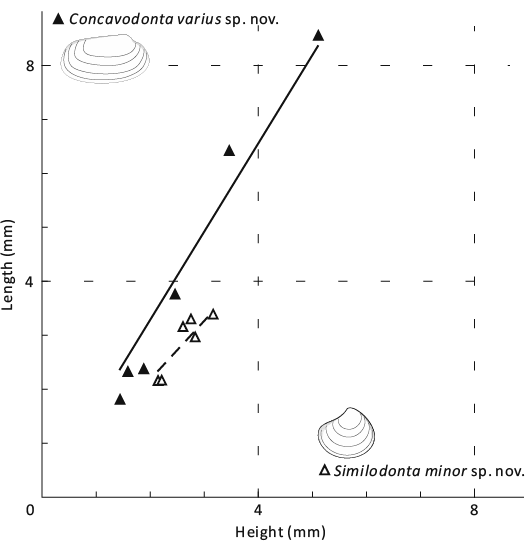
Fig. 6. Bivariate plot of shell morphometrics for praenuculid protobranch bivalve Concavodonta varius sp. nov. (solid triangles, thick line with slope approximately 1.64) and similodontid protobranch bivalve Similodonta minor sp. nov. (hollow triangles, thick dash line with slope approximately 1.09).
Table 2. Shell measurements (in mm) of Concavodonta varius sp. nov.; L/H, length/height ratio (dimensionless).
|
Specimen number |
Length |
Height |
L/H |
|
NIGP208872 |
1.8 |
1.4 |
1.24 |
|
NIGP208873 |
2.4 |
1.9 |
1.26 |
|
NIGP208869 |
2.3 |
1.6 |
1.46 |
|
NIGP208871 |
3.8 |
2.5 |
1.52 |
|
NIGP208868 |
8.5 |
5.1 |
1.67 |
|
NIGP208870 |
6.4 |
3.5 |
1.86 |
Remarks.—In comparison with the type species Concavodonta ponderata, C. varius sp. nov. has a more elongate outline, a straighter dorsal margin, and a greater angle between the anterior and posterior dorsal borders. Concavodonta ovalis (Sánchez, 1999) from the Upper Ordovician of Argentina is similar to our specimens in outline, but its umbo does not project above the hinge line as in our specimens, and is located more centrally on the shell. Concavodonta imbricata (Portlock, 1843) from the Upper Ordovician of Ireland has more posterior teeth, more distinct concentric ornamentation, and a more contracted posterior margin than our specimens.
Stratigraphic and geographic range.—Middle part of the Xiazhen Formation, upper Katian, Upper Ordovician; Yushan County, China.
Superorder Nuculaniformii Carter et al., 2000
Order Cardiolaritida Cope, 2000
Superfamily Tironuculoidea Babin, 1982
Family Similodontidae Carter & Pojeta in Carter et al., 2011
Genus Trigonoconcha Sánchez, 1999
Type species: Trigonoconcha acuta Sánchez, 1999, from the Don Braulio Formation, Upper Ordovician, Argentina.
Trigonoconcha brevis sp. nov.
Figs. 3, 7.
Zoobank LSID: urn:lsid:zoobank.org:act:AE78B449-4021-4736-B87D-3B246010B765.
Etymology: From Latin brevis, short, suggesting the shorter height and bigger L/H ratio than those of the type species T. acuta.
Type material: Holotype, NIGP208881 (Fig. 7H), an internal mould of right valve and its cast. Paratypes: NIGP208874 (Fig. 7A), NIGP208875 (Fig. 7B), NIGP208876 (Fig. 7C), NIGP208879 (Fig. 7E), NIGP208878 (Fig. 7F), NIGP208877 (Fig. 7G).
Type locality: Yushan County, Jiangxi Province, East China.
Type horizon: Katian, Upper Ordovician.
Material.—Two internal moulds and one external mould of right valves, three internal moulds and one external mould of left valves (NIGP208874–208881). Most from JYZ-11, few from ES-c (JYZ-10), Yushan County, Jiangxi Province (China); Katian (Late Ordovician).
Diagnosis.—Trigonoconcha with L/H ratio about 1, anterior and posterior dorsal margins meeting at an angle of around 70°.
Description.—Small to large triangular shell; equivalve and inequilateral. Umbo acute and situated subcentral, orthogyrate or slightly prosogyrate. Anterior adductor scar well marked, elliptical. Anterior and posterior hinge lines meeting below the beak at an angle approximately 70°. Taxodont dentition, containing five–seven posterior teeth and six–nine anterior convexodont teeth in juvenile shells, increasing to up to 12 posterior teeth and 14 anterior teeth in adult shells; anterior teeth slightly larger than posterior teeth. Amphidetic ligament visible (Fig. 7A, C1). Shell ornamentation of concentric lines.
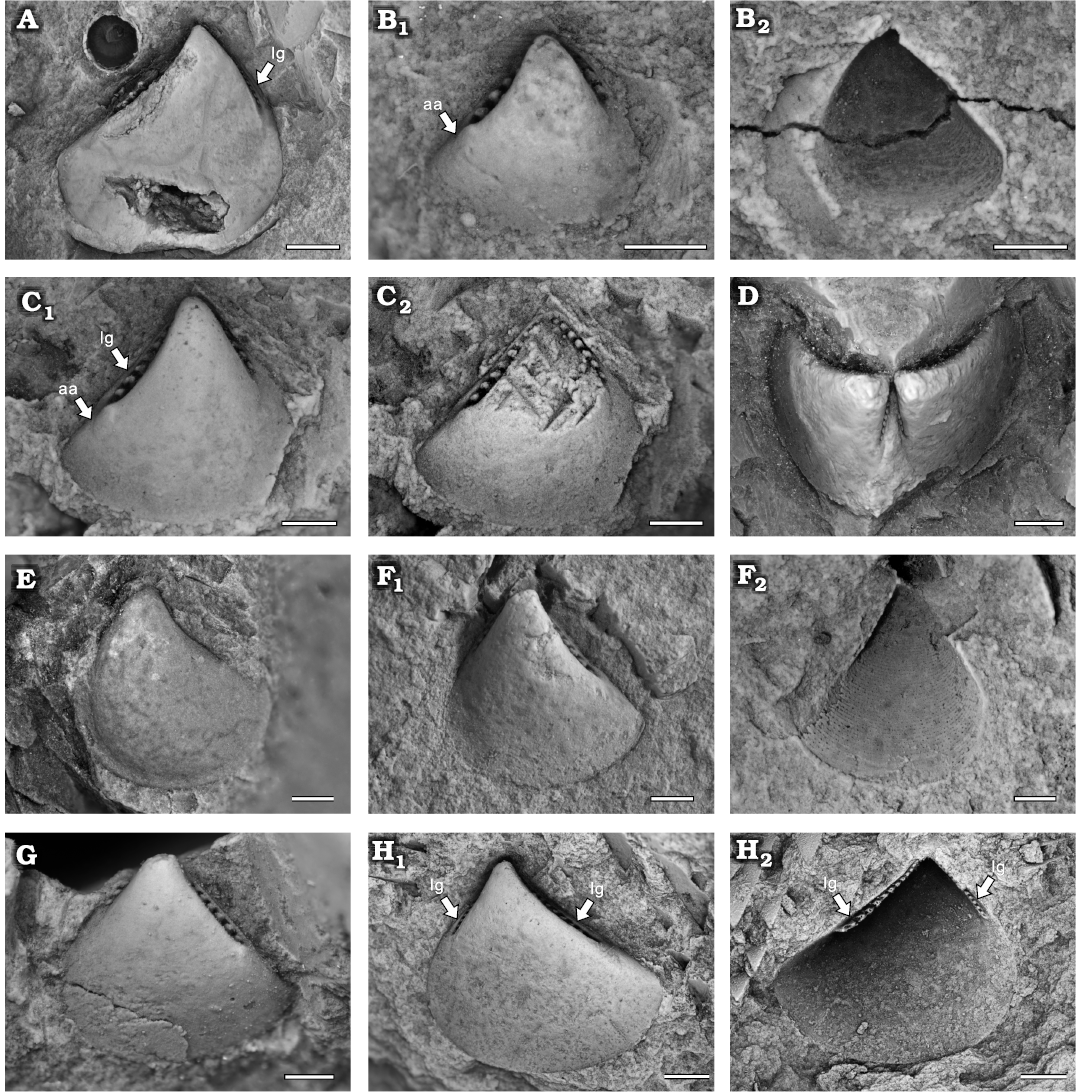
Fig. 7. Similodontid protobranch bivalve Trigonoconcha
brevis sp. nov., Yushan County, Jiangxi Province (China);
Katian (Late Ordovician) A. NIGP208874
(paratype), left valve, internal mould. B.
NIGP208875 (paratype), left valve showing concentric ornamentation,
internal (B1) and external
(B2) moulds. C.
NIGP208876 (paratype), left valve, internal mould (C1)
and dentition after removing umbo (C2).
D. NIGP208880, conjoined shell, internal
mould.
E. NIGP208879 (paratype), right valve,
internal mould. F. NIGP208878 (paratype),
right valve, internal (F1)
and external (F2) moulds. G. NIGP208877 (paratype), right valve, internal
mould. H. NIGP208881 (holotype), right
valve, internal mould (H1)
and its latex cast (H2).
Abbreviations: aa, anterior adductor scar; lg, ligament. Scale bars A,
2 mm; D, H, 1 mm; B, C, E–G, 0.5 mm.
Table 3. Shell measurements (in mm) of Trigonoconcha brevis sp. nov.; L/H, length/height ratio (dimensionless).
|
Specimen number |
Length |
Height |
L/H |
|
NIGP208874 |
9.3 |
9.3 |
1.00 |
|
NIGP208875 |
1.4 |
1.3 |
1.04 |
|
NIGP208876 |
2.4 |
2.3 |
1.04 |
|
NIGP208877 |
2.6 |
2.4 |
1.10 |
|
NIGP208878 |
2.6 |
2.6 |
0.99 |
|
NIGP208879 |
2.4 |
2.5 |
0.95 |
|
NIGP208881 |
5.3 |
4.8 |
1.09 |
Remarks.—The type species was determined by Sánchez (1999) primarily on the base of the small angle between its anterior and posterior dorsal margins (usually less than 70°). Compared with T. acuta, our specimens have fewer anterior and posterior teeth, a higher L/H ratio of approximately 1 (Fig. 3; Table 3), and a greater angle between the anterior and posterior dorsal margins.
Stratigraphic and geographic range.—Middle part of the Xiazhen Formation, upper Katian, Upper Ordovician; Yushan County, China.
Genus Similodonta Soot-Ryen, 1964
Type species: Tellinomya similis Ulrich, 1892, Richmond Group, Upper Ordovician, Minnesota, USA.
Similodonta minor sp. nov.
Figs. 6, 8.
Zoobank LSID: urn:lsid:zoobank.org:act:030A0A6E-177D-4F14-B300-C780E46DC3D2.
Etymology: From Latin minor, less, indicating fewer teeth than other species of Similodonta.
Type material: Holotype, NIGP208882 (Fig. 8A), an internal mould of right valve. Paratypes: NIGP208883 (Fig. 8B), NIGP208923 (Fig. 8D), NIGP208885 (Fig. 8E), NIGP208887 (Fig. 8F).
Type locality: Yushan County, Jiangxi Province, East China.
Type horizon: Katian, Upper Ordovician.
Material.—Three internal moulds of right valves, four internal moulds of left valves (NIGP208882–208887, NIGP208923). Most from JYZ-11, few from ES-c (JYZ-10) or ES-b (JYZ-9), Yushan County, Jiangxi Province (China); Katian (Late Ordovician).
Diagnosis.—Similodonta with less than 30 teeth and an opisthogyrate umbo.
Description.—Small sub-triangular shell; equivalve and inequilateral. Length 2.2–3.4 mm, height 2.2–3.2 mm (Fig. 6; Table 4). Anterior, posterior and ventral margin rounded; anterior and posterior dorsal margin meet at an angle of about 90°. Umbo broad, sub-central to anterior, slightly opisthogyrate. Taxodont dentition, consisting of 13–15 anterior and 10–11 posterior teeth; teeth grading from convexodont teeth away from the umbo to peg-like teeth close to the umbo; anterior teeth slightly larger than posterior teeth. Anterior adductor scar rounded. Posterior adductor scar elliptical, slightly larger than anterior adductor scar.
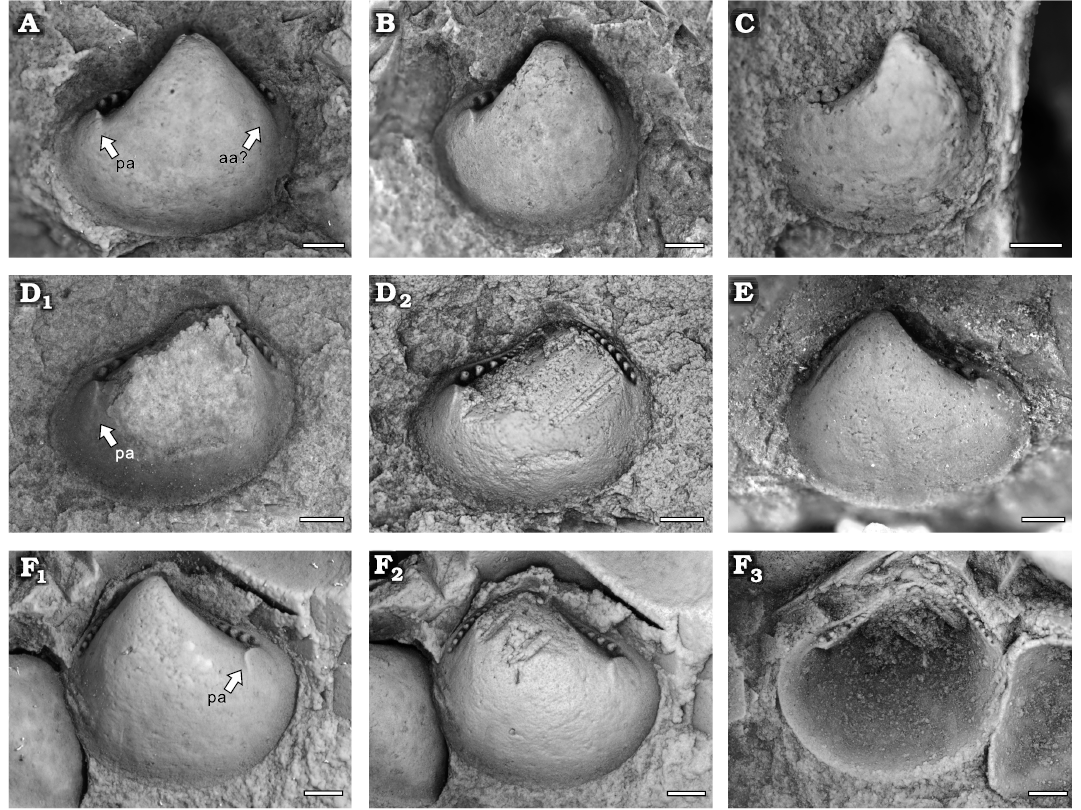
Fig. 8. Similodontid protobranch bivalve Similodonta
minor sp. nov., Yushan County, Jiangxi Province (China);
Katian (Late Ordovician). A. NIGP208882
(holotype), right valve, internal mould. B.
NIGP208883 (paratype), right valve, internal mould. C.
NIGP208884, right valve, internal mould. D.
NIGP208923 (paratype) right valve, incomplete internal mould (D1)
and dentition after removing umbo (D2).
E. NIGP208885 (paratype), left valve,
internal mould.
F. NIGP208887 (paratype), left valve,
internal mould (F1),
dentition after removing umbo (F2),
and latex cast (F3).
Abbreviatons: aa, anterior adductor scar; pa, posterior adductor scar.
Scale bars 0.5 mm.
Table 4. Shell measurements (in mm) of Similodonta minor sp. nov.; L/H, length/height ratio (dimensionless).
|
Specimen number |
Length |
Height |
L/H |
|
NIGP208882 |
3.3 |
2.8 |
1.20 |
|
NIGP208883 |
3.0 |
2.8 |
1.05 |
|
NIGP208884 |
2.2 |
2.2 |
1.00 |
|
NIGP208885 |
3.2 |
2.6 |
1.21 |
|
NIGP208886 |
2.2 |
2.2 |
0.97 |
|
NIGP208887 |
3.4 |
3.2 |
1.07 |
Remarks.—Our specimens are similar in their triangular outline to Similodonta collina (Reed, 1946) from the Upper Ordovician of Scotland, S. spjeldnaesi (Soot-Ryen & Soot-Ryen, 1960) from the Upper Ordovician of Norway, and the type species S. similis. However, the present new species has significantly fewer teeth. Compared with S. minor, S. ceryx (Cope, 1999) from the Middle Ordovician of Wales can be distinguished by its prosogyrate umbo and more elongate posterior. Similodonta wahli (Isakar, 1991) from the Upper Ordovician of Estonia differs from our species in its smaller angle between anterior and posterior hinges.
Stratigraphic and geographic range.—Middle part of the Xiazhen Formation, upper Katian, Upper Ordovician; Yushan County, China.
Order Nuculanida Carter et al., 2000
Superfamily Malletioidea Adams & Adams, 1858
Family Cucullellidae Fischer, 1886
Subfamily Palaeoneilinae Babin, 1966
Genus Palaeoneilo Hall & Whitfield, 1869
Type species: Nuculites constricta Conrad, 1842, Hamilton Group, Middle Devonian, New York, USA.
Palaeoneilo rectus sp. nov.
Figs. 9–11.
Zoobank LSID: urn:lsid:zoobank.org:act:1598E00A-4CF5-454C-9B9B-CEC8561B7BED.
Etymology: From Latin rectus, straight, suggesting the sub-straight hinge line.
Type material: Holotype, NIGP208907 (Fig. 9H), internal and external moulds of left valve. Paratypes: NIGP208888–208906, NIGP208908–208910, NIGP208924–208926 (Figs. 9A–G, I–K, 10A–O).
Type locality: Yushan County, Jiangxi Province, East China.
Type horizon: Katian, Upper Ordovician.
Material.—52 internal moulds and 15 external moulds of right valves, 32 internal moulds and 11 external moulds of left valves, 23 internal moulds and 13 external moulds of conjoined shells (NIGP208888–208911, NIGP208924–208926). From all collections, but mostly from JYZ-11, Yushan County, Jiangxi Province (China); Katian (Late Ordovician).
Diagnosis.—The Palaeoneilo with a sub-straight hinge line and tilted posterior orthodont or slightly concavodont teeth.
Description.—Small to medium elongated-elliptical shell; equivalve and inequilateral. Average L/H ratio around 1.8 (Fig. 11, Table 5), Anterior margin rounded, with an auricle near the anterior dorsal margin; posterior margin elongated and slightly contracted; ventral and dorsal margins convex. The umbo situated in anterior third of the shell, projected above the hinge line, orthogyrate or prosogyrate. Ovate anterior adductor scar well preserved in some specimens, close to the umbo, with a small pedal retractor scar dorsally (Figs. 9D2, H1, 10C, L). Hinge line nearly straight, half length of the dorsal margin, with 4 concavodont anterior teeth and 14 peg-like posterior teeth in adult shell (8 in juvenile shell). The posterior teeth tilted towards the umbo. No distinct discontinuity between the anterior and posterior teeth series, and the teeth beneath the umbo are smaller. Outer shell surface smooth.
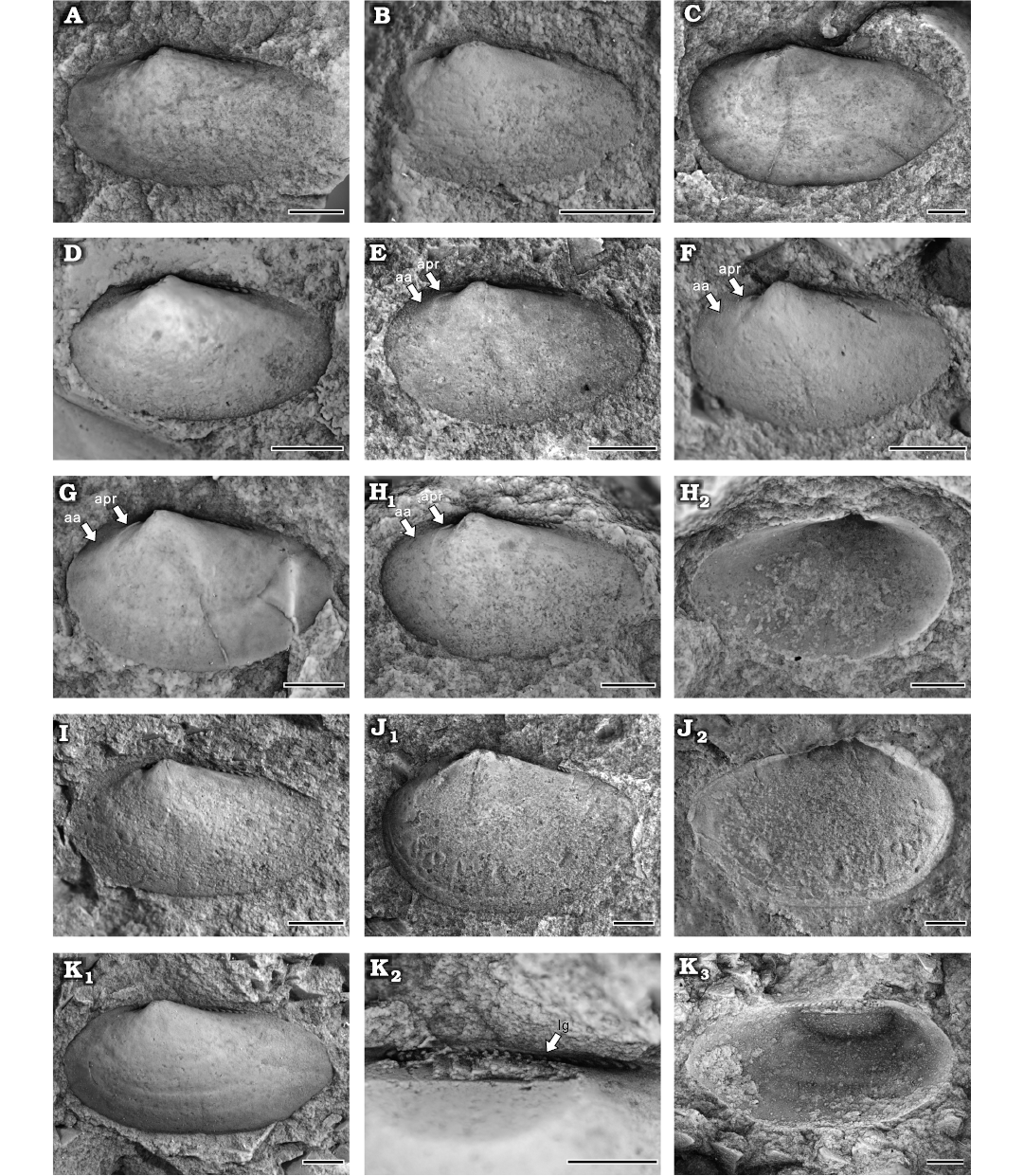
Fig. 9. Cucullellid protobranch Palaeoneilo rectus sp. nov., Yushan County, Jiangxi Province (China); Katian (Late Ordovician). A. NIGP208901 (paratype), left valve, internal mould. B. NIGP208924 (paratype), left valve, internal mould. C. NIGP208902 (paratype), internal mould, left valve. D. NIGP208903 (paratype), internal mould, left valve. E. NIGP208904 (paratype), left valve, internal mould. F. NIGP208905 (paratype), left valve, internal mould with a faint depression. G. NIGP208910 (paratype), left valve, internal mould. H. NIGP208907 (holotype), left valve, internal (H1) and external (H2) mould with an auricle. I. NIGP208908 (paratype), left valve, internal mould. J. NIGP208909 (paratype), left valve, internal mould (J1) and external mould (J2) with an auricle. K. NIGP208906 (paratype), left valve, internal mould (K1), dentition after removing umbo (K2), latex cast (K3). Abbreviations: aa, anterior adductor scar; apr, anterior pedal retractor scar; lg, ligament. Scale bars 1 mm.
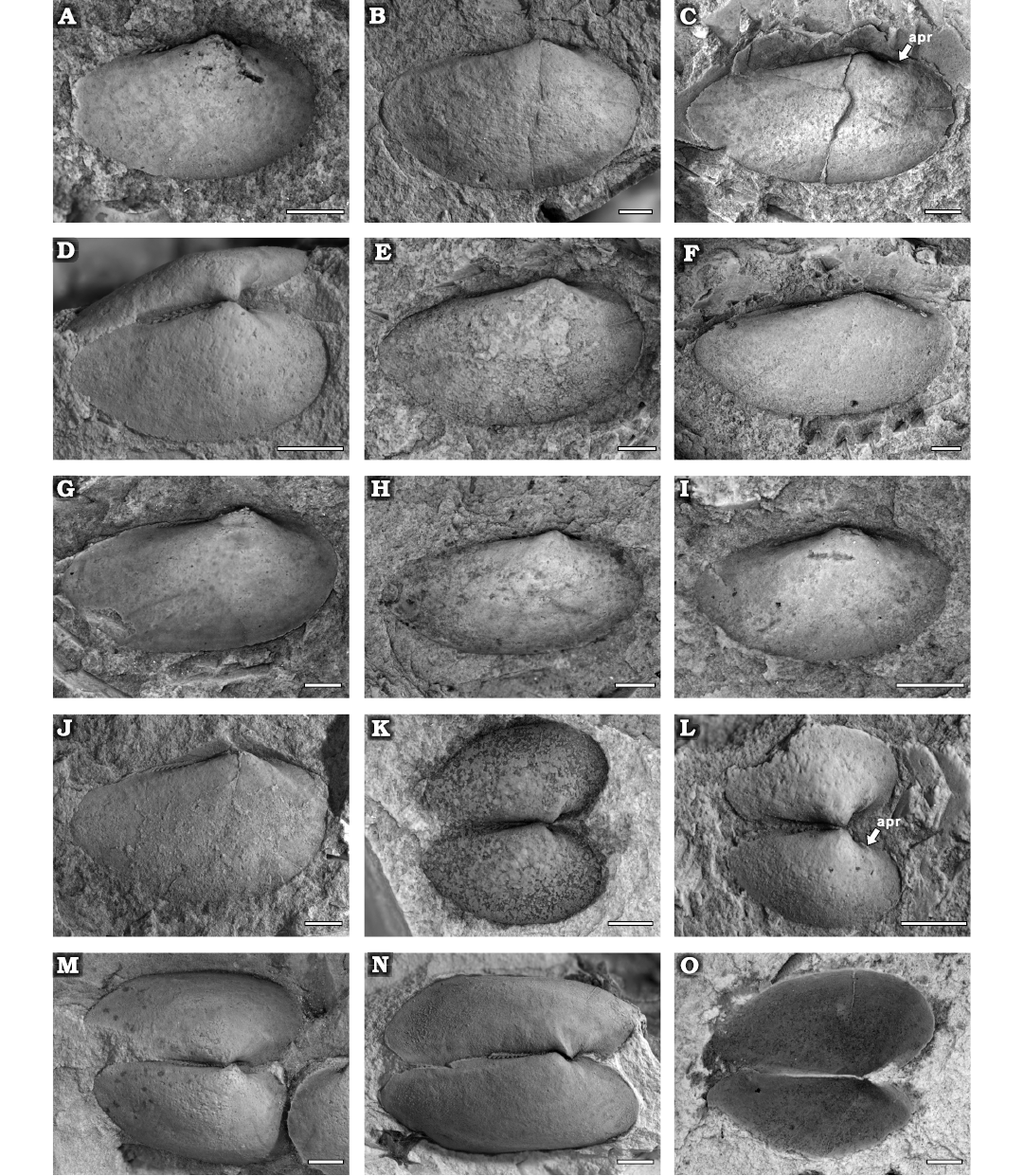
Fig. 10. Cucullellid protobranch Palaeoneilo rectus sp. nov., Yushan County, Jiangxi Province (China); Katian (Late Ordovician). A. NIGP208888 (paratype), right valve, internal mould. B. NIGP208889 (paratype), right valve, internal mould. C. NIGP208890 (paratype), internal mould, right valve. D. NIGP208891 (paratype), right valve of a conjoint valve, internal mould. E. NIGP208892 (paratype) , right valve, internal mould. F. NIGP208893 (paratype), right valve, internal mould. G. NIGP208894 (paratype), right valve, internal mould. H. NIGP208895 (paratype), right valve, internal mould. I. NIGP208896 (paratype), right valve, internal mould. J. NIGP208897 (paratype), right valve, internal mould. K. NIGP208898 (paratype), conjoint valve, internal mould. L. NIGP208899 (paratype), conjoint valve, internal mould. M. NIGP208925 (paratype), conjoint valve, internal mould. N. NIGP208926 (paratype), conjoint valve, internal mould. O. NIGP208900 (paratype), conjoint valve, external mould. Abbreviation: apr, anterior pedal retractor scar. Scale bars 1 mm.
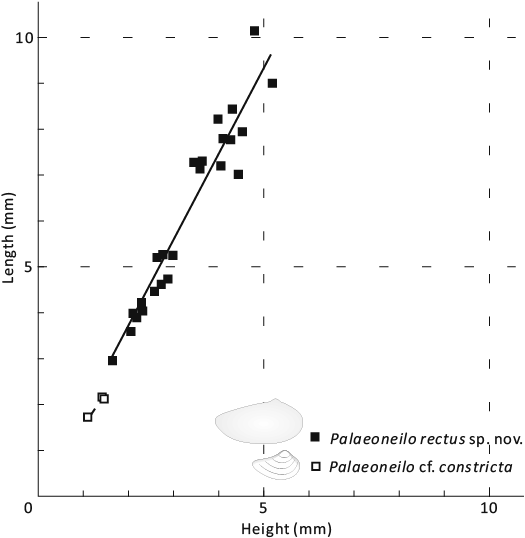
Fig. 11. Bivariate plot of shell morphometrics for cucullellid protobranch Palaeoneilo rectus sp. nov. (solid squares, thick line with slope approximately 1.86) and cucullellid protobranch Palaeoneilo cf. constricta Conrad, 1842 (hollow squares, thick dash line with slope approximately 1.51).
Table 5. Shell measurements (in mm) of Palaeoneilo rectus sp. nov.; L/H, length/height ratio (dimansionless).
|
Specimen number |
Length |
Height |
L/H |
|
NIGP208888 |
4.6 |
2.7 |
1.70 |
|
NIGP208889 |
9.0 |
5.2 |
1.74 |
|
NIGP208890 |
8.2 |
4.0 |
2.07 |
|
NIGP208891 |
4.2 |
2.3 |
1.84 |
|
NIGP208892 |
7.8 |
4.3 |
1.82 |
|
NIGP208893 |
10.2 |
4.8 |
2.12 |
|
NIGP208894 |
8.4 |
4.3 |
1.97 |
|
NIGP208895 |
7.3 |
3.4 |
2.11 |
|
NIGP208896 |
4.0 |
2.1 |
1.89 |
|
NIGP208897 |
8.0 |
4.5 |
1.76 |
|
NIGP208898 |
4.5 |
2.6 |
1.73 |
|
NIGP208899 |
3.0 |
1.6 |
1.79 |
|
NIGP208900 |
7.2 |
3.6 |
2.00 |
|
NIGP208901 |
5.3 |
2.8 |
1.91 |
|
NIGP208902 |
7.8 |
4.1 |
1.90 |
|
NIGP208903 |
3.9 |
2.2 |
1.79 |
|
NIGP208904 |
4.0 |
2.3 |
1.76 |
|
NIGP208905 |
3.6 |
2.0 |
1.75 |
|
NIGP208906 |
7.3 |
3.6 |
2.01 |
|
NIGP208907 |
5.2 |
3.0 |
1.76 |
|
NIGP208908 |
5.2 |
2.6 |
1.98 |
|
NIGP208909 |
7.0 |
4.4 |
1.58 |
|
NIGP208910 |
4.7 |
2.9 |
1.65 |
|
NIGP208911 |
7.2 |
4.0 |
1.78 |
Remarks.—The umbonal-posteroventral depression was cited as a key diagnostic feature of Palaeoneilo (Babin 1966; McAlester 1969). However, because the depression is not preserved in all assigned species, it may not be considered as the diagnostic feature for the genus.
Compared with the type species Palaeoneilo constricta, P. rectus has a more rounded and elongate posterior margin. This feature distinguishes it from P. smithi (Pojeta & Gilbert-Tomlinson, 1977) from the Middle Ordovician of Australia. The new species differs from Palaeoneilo filosa (Conrad, 1842) and P. muta (Hall, 1870) from the Devonian of USA in having shorter hinge and weaker ornamentations. Other species, such as P. emarginata (Conrad, 1841), P. angusta (Hall, 1885), P. perplana (Hall & Whitfield, 1869) and P. truncata (Hall, 1883) from the Devonian of USA could be distinguished in having much more distinct umbonal posterior depressions.
Many species of Palaeoneilo have convexodont teeth in both anterior and posterior hinge lines, such as P. constricta, P. smithi, P. hopensacki (Verneuil & Barrande, 1855) and P. beirensis (Sharpe, 1853). Some others have orthodont teeth beneath the umbo, like P. fecunda. The dentition of P. rectus consists of concavodont anterior teeth and orthodont posterior teeth. The posterior teeth tilt toward the umbo, thicken and grade into concavodont teeth posteriorly (Fig. 9K2).
Stratigraphic and geographic range.—Middle part of the Xiazhen Formation, upper Katian, Upper Ordovician; Yushan County, China.
Palaeoneilo cf. constricta Conrad, 1842
Figs. 11, 12.
cf. 1842 Nuculites constricta sp. nov.; Conrad, 1842: 249, pl. 15: 8.
Material.—Two right valve internal moulds, four left valve internal moulds, five incomplete external moulds (NIGP208912–208914). From ES-b (JYZ-9), ES-c (JYZ-10) and JYZ-11, Yushan County, Jiangxi Province (China); Katian (Late Ordovician).
Description.—Small nuculaniform shell; equivalve and inequilateral. Anterior and ventral margin convex; posterior dorsal margin concave; posterior margin forming a short rostrum. A ventral sinus is situated at the posterior ventral margin. The umbo broad, situated subcentral to anterior, orthogyrate or slightly opisthogyrate. Adductor scars well preserved (Fig. 12B). Anterior adductor scar rounded, smaller than elongated elliptical posterior adductor scar, and positioned closer to the dorsal margin. Taxodont dentition, including 8 anterior teeth and 12 posterior teeth; anterior and posterior teeth show similar size. The shell surface ornamented with commarginal lines. The intervals between the lines are much wider than lines themselves.
Remarks.—The distinct ventral sinus and the constricted posterior margin suggest the specimens are close to the type species Palaeoneilo constricta. The shape is variable on the L/H ratio (Hall 1885; McAlester 1962, 1968). In our specimens, the L/H ratio is around 1.5 (Table 6), similar to those figured by Conrad (1842) and Hall (1885: pl. 48: 2, 4). The ornamentation differs slightly from that of previously documented specimens by having much wider intervals between the concentric lines.
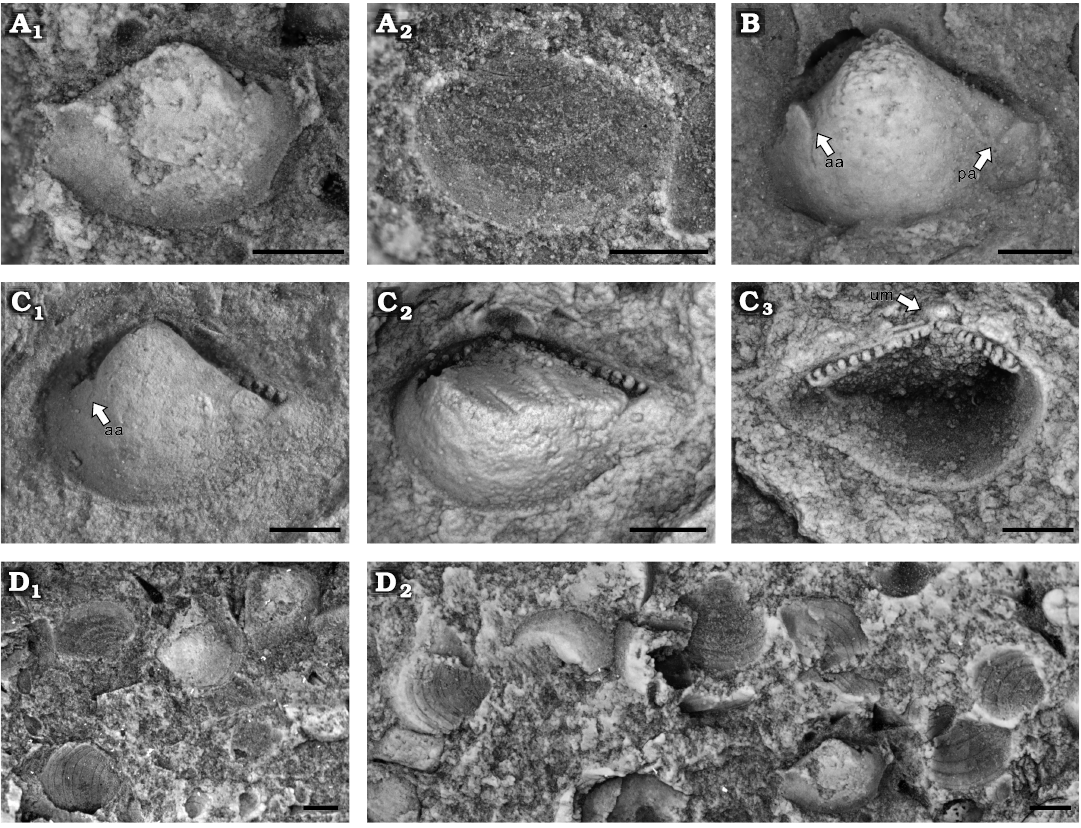
Fig. 12. Cucullellid protobranch Palaeoneilo cf. constricta Conrad, 1842, Yushan County, Jiangxi Province (China); Katian (Late Ordovician). A. NIGP208912, right valve showing ornamentation, internal (A1) and external (A2) moulds. B. NIGP208913, left valve, internal mould. C. NIGP208914, left valve, internal mould (C1), dentition after removing umbo (C2) and latex cast (C3). D. Block with the stack of numerous shells, bearing external (D1) and internal (D2) mould of NIGP208912. Abbreviations: aa, anterior adductor scar; pa, posterior adductor scar; um, umbo. Scale bars A, 0.2 mm; B, C, 0.5 mm; D, 1 mm.
Table 6. Shell measurements (in mm) of Palaeoneilo cf. constricta Conrad, 1842; L/H, length/height ratio (dimensionless).
|
Specimen number |
Length |
Height |
L/H |
|
NIGP208912 |
1.7 |
1.1 |
1.57 |
|
NIGP208913 |
2.1 |
1.5 |
1.46 |
|
NIGP208914 |
2.2 |
1.4 |
1.53 |
This represents the first record of material closely comparable to P. constricta from the Ordovician. However, all previously confirmed occurrences of P. constricta are from the Devonian, representing a significant stratigraphic gap with our Ordovician material. Therefore, we propose the more conservative designation of P. cf. constricta for these specimens.
Subclass Autobranchia Grobben, 1894
Infraclass Pteriomorphia Beurlen, 1944
Order Mytilida Férussac, 1822
Superfamily Modiolopsoidea Fischer, 1886
Family Goniophorinidae Sánchez, 2006
Genus Goniophorina Isberg, 1934
Type species: Goniophorina volvens Isberg, 1934, Kallholn Formation, Silurian, Dalarna, Sweden.
Goniophorina volvens Isberg, 1934
Figs. 13, 14A, B.
1934 Goniophorina volvens sp. nov.; Isberg 1934: 205, pl. 27: 6a–b; pl. 28: 1a–c.
1969 Goniophorina volvens Isberg, 1934; LaRocque and Newell 1969: N395, figs. D2, 10.
1975 Goniophorina volvens Isberg, 1934; Toni 1975: 150, fig. 3K.
Material.—One internal and one external mould of a right valve, one internal and one external mould of a left valve (NIGP208915–208916). All from ES-c (JYZ-10), Yushan County, Jiangxi Province (China); Katian (Late Ordovician).
Description.—Small to medium sized shell, posteriorly elongate sub-elliptical, equivalve, inequilateral. Anterior margin rounded; posterior margin broad and elongated ventrally, with the posteroventral angle less than 90°. Ventral margin straight, with a faint sinus centrally. Broad and prosogyrate umbo is situated in anterior fourth of the shell, strongly curved and projecting above the hinge line. Well-defined carina extending from the beak to the posteroventral angle, slightly curving dorsally and delimiting the posterior slope. Edentulous. Ornamentation consisting of fine commarginal lines.
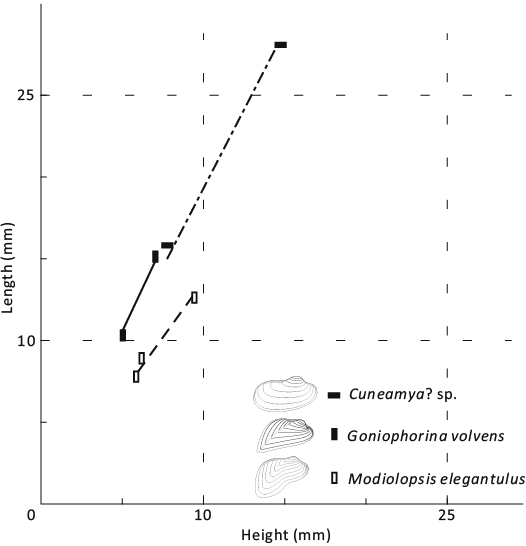
Fig. 13. Bivariate plot of shell morphometrics for goniophorinid pteriomorph bivalve Goniophorina volvens Isberg, 1934 (solid vertical rectangles, thick line with slope approximately 2.11), grammysiid pholadomyioid Cuneamya? sp. (solid horizontal rectangles, thick dash-dotted line with slope approximately 1.93), and modiolopsid pholadomyioid Modiolopsis elegantulus Sharpe, 1853 (hollow vertical rectangles, thick dash line with slope approximately 1.36).
Remarks.—The carina in Goniophorina volvens described by Isberg (1934) curves ventrally, differing from our specimens. In contrast, the specimen described by Toni (1975) possesses a straight or slightly curved carina. It is worthy to note that the carina of Goniophora contraria (Mansuy, 1912) from the Silurian of China is similar to that of our specimens, curved toward dorsal margin. However, as the internal details of Goniophora contraria are not preserved, it is difficult to confirm whether it is edentulous. Compared with other species, our specimens differ from G. tenuicostata (Harrington, 1938) from the Lower Ordovician of Argentina and G. extensa (Cope, 1996) from the Upper Ordovician of UK in lacking the radial ornamentation. Some species reported in China, such as G. mangbuensis (Guo, 1985) from the Lower Ordovician, G. dianensis (Guo, 1985) from the Silurian and G. contracta (Fang & Cope, 2004) from the Middle Ordovician differ from our specimens in having a weaker carina.
Stratigraphic and geographic range.—Upper Ordovician, Oslo, Norway, Dalarna, Sweden, and Yushan County, China.
Family Modiolopsidae Fischer, 1886
Genus Modiolopsis Hall, 1847
Type species: Pterinea modiolaris Conrad, 1838, Pulaski beds, Upper Ordovician, New York, USA.
Modiolopsis elegantulus Sharpe, 1853
Figs. 13, 15A, B.
1853 Modiolopsis elegantulus sp. nov.; Sharpe 1853: 152, pl. 9: 15a, b.
1991 Modiolopsis? elegantulus Sharpe, 1853; Babin and Gutiérrez-Marco 1991: 123, pl. 5: 1.
2003 Modiolopsis elegantulus Sharpe, 1853; Babin and Beaulieu 2003: 201, pl. 4: 4–6, 8.
Material.—One internal mould and two external moulds of right valves, two internal moulds and one external mould of left valves (NIGP208917–208919). All from ES-b (JYZ-9), Yushan County, Jiangxi Province (China); Katian (Late Ordovician).
Description.—Small to middle modioliform shell; equivalve and inequilateral. Anterior margin rounded; posterior elongated; a sinus in the anterior third of the ventral margin. Umbo situated anterior fourth of the shell, prosogyrate. A posterior umbonal ridge distinct near the umbo, fading towards the postero-ventral angle. Edentulous. Anterior adductor scar small and ill-defined, elliptical (Fig. 15A1). Shell thin, the fine commarginal lines clearly visible on the internal moulds.
Remarks.—Our specimens are very similar to those from the Upper Ordovician of Portugal (Sharpe 1853). In contrast, the specimens from the Middle Ordovician of Spain (Babin and Gutiérrez-Marco 1991) have a thinner shell and stronger ornamentations than ours. Our specimens differ from the type species Modiolopsis modiolaris and other species, such as M. drabovensis (Barrande, 1881) from the Upper Ordovician of Czech Republic, M. pojetai (Jokobsen et al., 2016) from the Middle Ordovician of Australia, and M. cuyana (Sánchez, 1990) from the Upper Ordovician of Argentina, in having a distinct ventral sinus. M. aragonensis (Babin & Hammann, 2001) from the Lower Ordovician of Spain has a stronger umbonal ridge than our specimens.
Stratigraphic and geographic range.—Darriwilian (Middle Ordovician)–lower Silurian. Bussaco, Portugal, Iberian Cordillera, Spain, Maine-et-Loire, France, and Yushan County, China.
Order Cyrtodontida Scarlato & Starobogatov in Nevesskaja et al., 1971
Superfamily Cyrtodontoidea Ulrich, 1894
Family Cyrtodontidae Ulrich, 1894
Subfamily Cyrtodontinae Ulrich, 1894
Genus Cyrtodontula Tomlin, 1931
Type species: Whitella obliquata Ulrich, 1890, from the Cincinnati Group, Upper Ordovician, Ohio, USA.
Cyrtodontula sp.
Fig. 14C, D.
Material.—One internal mould and one external mould of the right valve (NIGP208920) from ES-c (JYZ-10), Yushan County, Jiangxi Province (China); Katian (Late Ordovician).
Description.—Shell large, subquadrate, tumid, and inequilateral, with dimensions of approximately 61 mm in length and 49 mm in height. Anterior and posterior margins rounded; dorsal margin straight; ventral margin broadly rounded. Umbo strongly projected above the dorsal margin, situated in anterior third of the shell, prosogyrate. Shell surface ornamented with fine growth lines (Fig. 14C, D1). A possible elongated posterior lateral tooth visible on the internal mould (Fig. 14C, D2). Ligament and adductor scars not preserved.
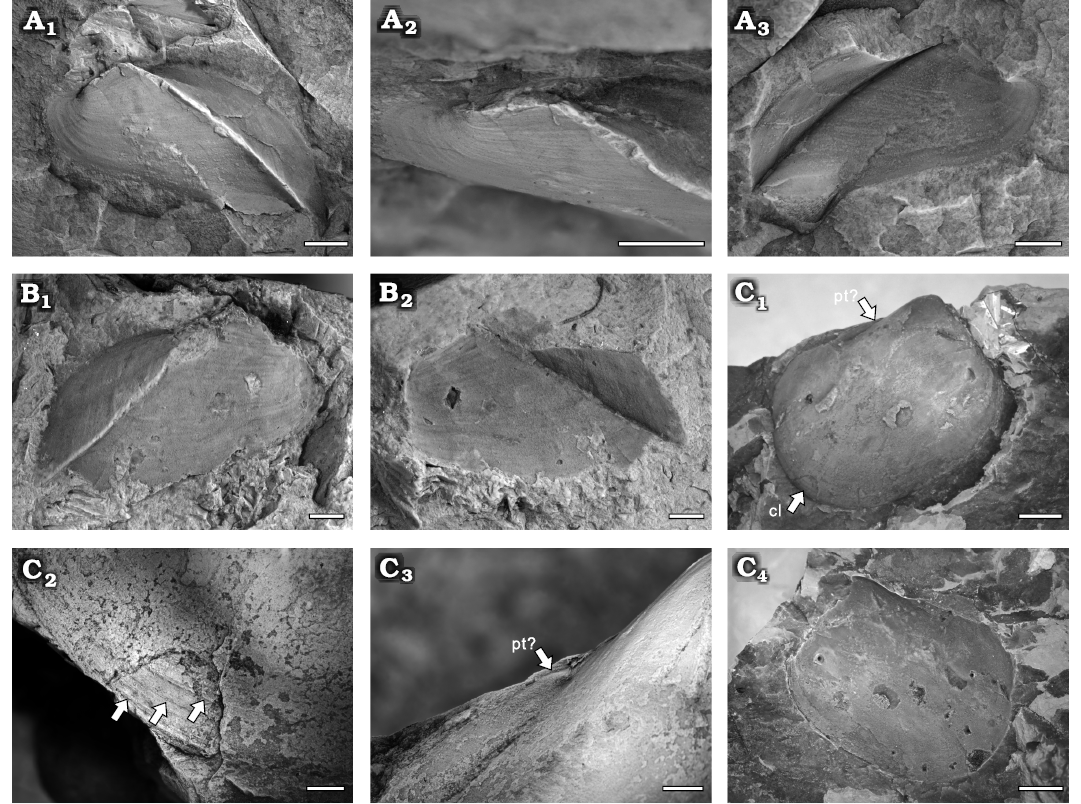
Fig. 14. A, B.
Goniophorinid pteriomorph bivalve Goniophrina
volvens Isberg, 1934, Yushan County, Jiangxi Province
(China); Katian (Late Ordovician).
A. NIGP208915, right valve, internal mould
(A1), close-up of dorsal
side (A2), and external
mould (A3). B.
NIGP208916, left valve, internal (B1)
and external (B2) moulds. C. Cyrtodontid pholadomyoid Cyrtodontula
sp.; NIGP208920, right valve, internal mould (C1),
close-up of ventral side (C2,
arrows point to the commarginal lines), close-up of umbo (C3),
and external mould (C4).
Abbreviations: cl, commarginal lines; pt, posterior tooth. Scale bars
A, C2, C3,
2 mm; B, 1 mm; C1, C4
10 mm.
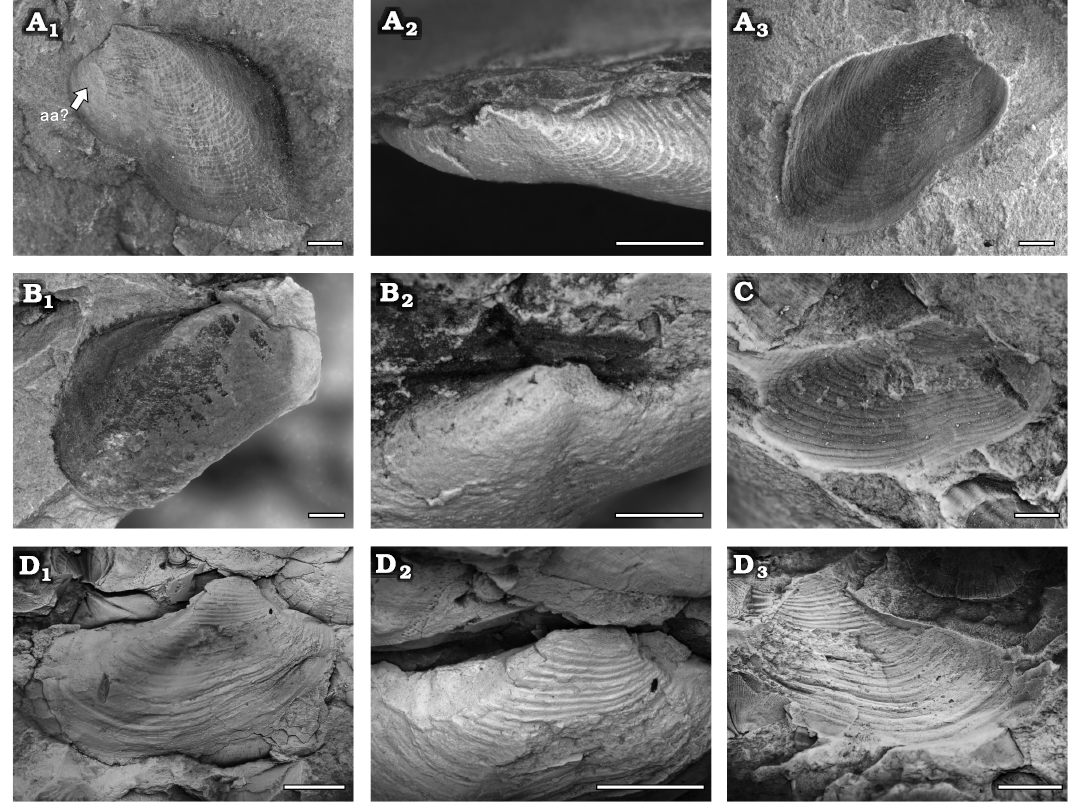
Fig. 15. A, B. Modiolopsid pholadomyoid Modiolopsis elegantulus Sharpe, 1853, Yushan County, Jiangxi Province (China); Katian (Late Ordovician). A. NIGP208918, left valve, internal mould (A1), close-up of dorsal side (A2), and external mould with ornamentation (A3). B. NIGP208917, right valve, internal mould (B1) and close-up of dorsal side (B2). C, D. Grammysiid pholadomyoid Cuneamya? sp. C. NIGP208922, left valve, external mould. D. NIGP208921, right valve, internal mould (D1), close-up of dorsal side (D2), external mould (D3). Abbreviation: aa, anterior adductor scar. Scale bars A, B, 1 mm; C, 2 mm; D, 5 mm.
Remarks.—The large subquadrate shell with its prominent umbo and a possible posterior lateral tooth suggest the affinity to Cyrtodontula. Due to its solitary occurrence, we propose to assign the specimen under open nomenclature. This specimen is similar to the type species C. obliquata, but its dentition is unclear. Compared with C. hadzeli (Pojeta & Gilbert-Tomlinson, 1977) from the Lower Ordovician of Australia, the posterior part of our specimen is not elongate. Furthermore, compared with C. sohli (Pojeta & Gilbert-Tomlinson, 1977) from the Upper Ordovician of Australia and C. scofieldi (Ulrich, 1890) from the Upper Ordovician of USA, our specimen lacks the prominent umbonal ridge.
Infraclass Heteroconchia Hertwig, 1895
Cohort Cardiomorphi Férussac, 1822
Order Pholadomyida Newell, 1965
Superfamily Pholadomyoidea King, 1844
Family Grammysiidae Miller, 1877
Subfamily Cuneamyinae Morris et al.1991
Genus Cuneamya Hall &Whitfield, 1875
Type species: Cuneamya miamiensis Hall &Whitfield, 1875, Trenton Limestone, Upper Ordovician, New York, USA.
Cuneamya? sp.
Figs. 13, 15C, D.
Material.—One internal mould and one external mould of a right valve, one external mould of a left valve (NIGP208921–208922). All from ES-b (JYZ-9), Yushan County, Jiangxi Province (China); Katian (Late Ordovician).
Description.—Medium sized elongated elliptical shell; equivalve and inequilateral. Anterior margin short and rounded; posterior margin elongated and rounded; ventral margin slightly convex with a sinus central to anterior of the margin; posterior dorsal margin concave. The umbo situated in anterior fourth of the shell, strongly incurved and projected above the dorsal margin, orthogyrate or prosogyrate. A broad umbonal ridge extended from the umbo, but faded toward the posteroventral angle. Dentition should be edentulous. Shell thin, ornamented with strong commarginal wrinkles.
Remarks.—The strongly incurved umbo and uniform concentric ornamentation may suggest an affinity to Cuneamya. The type species C. miamiensis has two lateral sulci: one prominent, extending from the umbo to central of the ventral margin, and another one shallower, located behind the edge of the lunule. These sulci are also well shown in C. grandis (Hind, 1910) from the Upper Ordovician of United Kingdom. In contrast, other Ordovician species lack the second shallow sulcus (Runnegar 1974), such as the species from the Upper Ordovician of Norway (Toni 1975), C. multistriata (Soot-Ryen & Soot-Ryen, 1960) from the Middle Ordovician of Norway, and the specimens described in this paper. Compared with Cuneamya multistriata, our specimens have much more elongate posterior. C. truncatula (Ulrich, 1894) and C. oblonga (Ulrich, 1894) from the Upper Ordovician of USA are both similar to our specimens in having subequal anterior and posterior heights, but C. truncatula and C. oblonga have much narrower anterior part. Cuneamya catilloides (Barrande in Perner, 1903) from the Upper Ordovician of Bohemia differs from our specimens in its oval shell shape.
Discussion
The bivalve association recognized here consists of 10 species dominated by Palaeoneilo rectus; therefore, it can be named as the Palaeoneilo rectus association. On the basis of Stanley’s (2015) conclusion that nuculoids are deposit feeders, we interpret Paullinea, Trigonoconcha, Similodonta, and Concavodonta as the infaunal deposit feeders. Palaeoneilo with elongate constricted shells also might be an infaunal deposit feeder (Hodges 2000; Fang 2006; Stanley 2015). Cuneamya with strong commarginal wrinkles is likely a slow infaunal burrower (Pojeta 1971; Polechová 2019). Modiolopsis, Cyrtodontula and Goniophorina are probably semi-infaunal endobyssate bivalves (Pojeta 1971; Liljedahl 1994; Fang 2006; Stanley 2015). Therefore, the Palaeoneilo rectus association recognized here primarily includes infaunal burrowers with few epifaunal or semi-infaunal filter feeders. The bottom of the epicontinental sea was predominantly occupied by epifaunal brachiopods and their dominance likely inhibited the diversification of epifaunal bivalves owing to the competition for the same eco-space.
Cope and Kříž (2013) proposed that the composition of Ordovician bivalve faunas shows a close correlation with the palaeolatitudes. Generally, Ordovician heteroconch bivalves preferred higher palaeolatitudes, while pteriomorph bivalves prefer lower palaeolatitudes, and Ordovician protobranchs (Nuculiformii and Nuculaniformii) have much higher diversities in lower palaeolatitudes. However, owing to the lack of data on high palaeolatitude bivalve faunas, the correlation between the palaeolatitude and the composition of the Late Ordovician bivalve fauna is complicated (Polechová 2022). Nevertheless, based on their known composition, three different Katian bivalve faunas can be recognized:
– The fauna dominated by pteriomorphs (Fig. 16A–C), such as the faunas from Morocco (Ebbestad et al. 2022), Norway (Toni 1975), and USA (Bretsky 1970a). However, the Moroccan fauna is an exception among these faunas, because it occurs in high latitudes during the Katian, where heteroconch bivalves typically prevail. This suggests that the dominance of pteriomorphs might have been affected by the Boda Warming Event (Fortey and Cocks 2005; Loi et al. 2010; Colmenar et al. 2018), which allowed the lower latitude faunas, to spread to higher latitudes during the Late Ordovician.
– The fauna dominated by nuculiform bivalves (Fig. 16D, E), including those from Northern Ireland and Scotland (Tunnicliff 1982; Cope 1996). These Nuculiformii bivalves are interpreted as inhabiting in a deeper-water environment, which is also confirmed by their contemporaneous brachiopods (Harper 2001; Candela 2006). Polechová (2022) suggested that protobranch bivalves (Nuculiformii and Nuculaniformii) can be dominant in the deeper shelf environments regardless of palaeolatitudes.
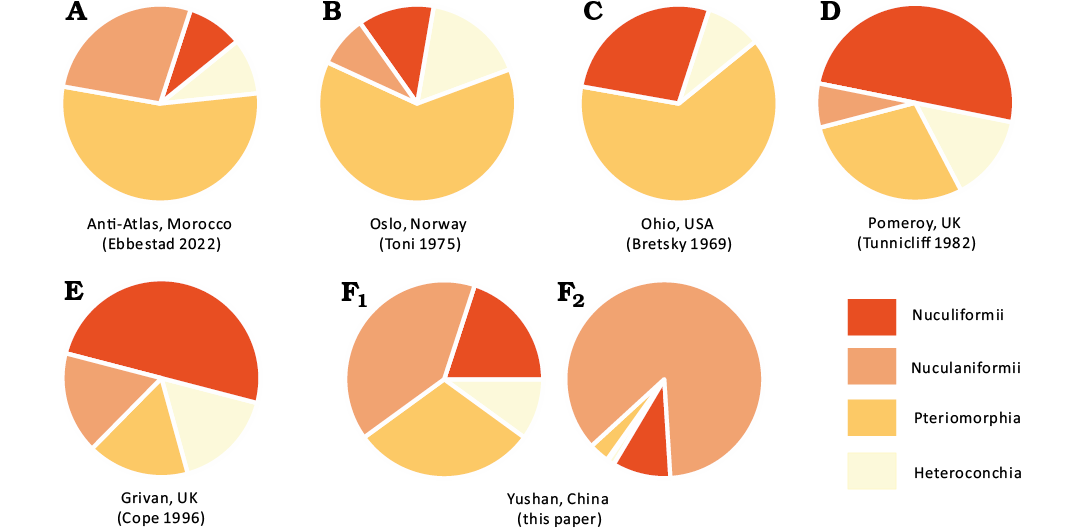
Fig. 16. Pie diagram of the faunal composition of six Katian bivalve associations from Anti-Atlas, Morocco (A), Oslo, Norway (B), Ohio, USA (C), Pomeroy, UK (D), Grivan, UK (E), and the Xiazhen Formation, Yushan, northeastern Jiangxi Province, East, China (F). A–E, F1, the percentage of species number of four main bivalve taxa from different places; F2, the percentage of specimen number of four main bivalve taxa.
– The fauna dominated by nuculaniform bivalves (Fig. 16F), which known only from the Palaeoneilo rectus association in the Xiazhen Formation. The palaeoenvironment of the Xiazhen Formation at Zhuzhai was interpreted as nearshore shallow-water setting (Zhan et al. 2002). This interpretation was based on the dominance of brachiopods and the presence of associated taxa like corals, trilobites, bryozoans, and stromatoporoids (Zhan and Rong 1995a; Lee 2013; Joen et al. 2022). Within the Palaeoneilo rectus association, the pteriomorph bivalves are rare in abundance and low in diversity, despite the Yangtze Platform being in low latitude during the Late Ordovician, which is a setting typically favorable for them. The equatorial cold-water tongue during the Late Ordovician (Jin et al. 2018) affected South China palaeoplate, as it transported the cold-water mass from then South Pole to the subequatorial (or even equatorial) Yangtze Platform and created an unsuitable environment for pteriomorph bivalves, which might be the main reason for the rarity of pteriomorph bivalves in the Palaeoneilo rectus association. The cold-water tongue enabled the cool-water favoring Foliomena brachiopod fauna to be widespread on the Yangtze Platform during the Sandbian and Katian (Late Ordovician). Conversely, Foliomena brachiopod fauna was almost absent in the epicontinental seas of Laurentia, where warm-water brachiopods were flourishing (Jin et al. 2018). A similar difference is also evident in the bivalve associations, and Late Ordovician warm-water pteriomorph bivalves are primarily found in Laurentia but rare in South China (Fig. 16C, F). The dominance of nuculiform bivalves in Ireland and Scotland may also be attributed to the similar cold-water mass, as these regions lay directly in the path of the cold-tongue.
The Palaeoneilo rectus association has some elements in common (Similodonta, Modiolopsis, Cyrtodontula, Goniophorina, Cuneamya) with its contemporaneous bivalve fauna from the Oslo region, Norway (Toni 1975). However, the Oslo fauna exhibits a significantly higher diversity. The lecithotrophic larvae of bivalves are thought to have appeared in the Ordovician (Babin 1995; Sánchez and Babin 2003), and could migrate along deep shelves or between separate terranes (Havlíček et al. 1994; Fatka and Mergl 2009). Both the poleward faunal migrations associated with the Boda Warming Event (Cocks and Torsvik 2002) and the migration corridor from pole to equator provided by High-Latitude Peri-Gondwana Cold Current (Jin et al. 2018), enabled interchange between these two bivalve associations.
Conclusions
Acknowledgements
We express our sincere thanks to Chi Xiangri, Liu Congying, Gao Jiahua, Li Fengyan, Ruan Ting, Song Siyu, Yang Jingwen, Yu Xinmiao, Yue Chaosheng, Wang Ting, Wu Shouhan, and Zhu Xiuping (all NIGP) for their kind help in the field, to Li Wenjie (NIGP) for his generous sharing of his samples, and to David A.T. Harper (Durham University, UK) for his help in linguistic check of our manuscript. We are deeply grateful to Marika Polechová (Czech Geological Survey, Czech Republic) and an anonymous reviewer for their insightful comments and suggestions which have substantially improved the manuscript. Financial support for this study was provided by the National Key Research and Development Program of China (2023YFF0803602), the Ministry of Science and Technology of China (2021FY200102) and the State Key Laboratory of Palaeobiology and Stratigraphy (NIGP). This is also a contribution to the IGCP 735: Rocks and the rise of Ordovician life.
Editor: Krzysztof Hryniewicz
References
Adams, H. and Adams, A. 1858. The Genera of Recent Mollusca, Arranged According to their Organization. Vol. Ⅱ. 661 pp. John Van Voorst. London. Crossref
Babin, C. 1966. Mollusques Bivalves et Céphalopodes du Paléozoique Armoricain. 497 pp. Imprimerie Commerciale et Administrative, Brest.
Babin, C. 1982. Mollusques Bivalves et Rostroconches. In: C. Babin, R. Courtessole, M. Melou, J. Pillet, D. Vizcaino, and E.L. Yochelson (eds.), Brachiopodes (Articulés) et Mollusques (Bivalves, Rostroconches, Monoplacophores, Gastropodes) de l’Ordovicien inférieur (Trémadocien-Arenigien) de la Montagne Noire (France Méridionale), 37–49. Société des Études Scientifiques de l’Aude, Carcassonne.
Babin, C. 1995. The initial Ordovician bivalve mollusc radiations on the western Gondwanan shelves. In: J.D. Cooper, M.L. Droser, and S.C. Finney (eds.), Ordovician Odyssey: Short Papers for the Seventh International Symposium on the Ordovician System, 491–498. Pacific Section Society for Sedimentary Geology, Fullerton.
Babin, C. and Beaulieu, G. 2003. Les Mollusques Bivalves de l’Ordovicien de Saint-Clément-de-la-Place (Maine-et-Loire, sud-est du Massif Armoricain). Bulletin de la Société des sciences naturelles de l’Ouest de la France, nouvelle série 25(4): 177–206.
Babin, C. and Gutiérrez-Marco, J.C. 1991. Middle Ordovician bivalves from Spain and their phyletic and palaeogeographic significance. Palaeontology 34: 109–147.
Babin, C. and Hammann, W. 2001. Une nouvelle espèce de Modiolopsis (Bivalvia) Dans L’ Arenig (Ordovicien Inferieur) de Daroca (Aragon, Espagne); réflexions sur la denture des bivalves primitifs. Spanish Journal of Palaeontology 16: 269–282. Crossref
Babin, C. and Melou, M. 1972. Mollusques Bivalves et Brachiopodes des “schistes de Raguenez” (Ordovicien supérieur du Finistère); conséquences stratigraphiques et paléobiogéographiques. Annales de la Société Géologique du Nord 92: 79–94. Crossref
Barrande, J. 1881. Systême silurien du centre de la Bohême, Vol. VI. Classe des mollusques. Order des Acéphalés. 576 pp. Chez Pauteur er diteuré, Paris.
Beurlen, K. 1944. Beiträge zur Stammesgeschichte der Muscheln. Mathematisch-Naturwissenschaftlichen Abteilung der Bayerischen Akademie der Wissenschaften zu München, Sitzungsberichte 1944: 133–145.
Bretsky, P.W. 1969. Central Appalachian Late Ordovician Communities. Geological Society of America Bulletin 80: 193–212. Crossref
Bretsky, P.W. 1970a. Late Ordovician Benthic Marine Communities in North-central New York. New York State Museum and Science Service Bulletins 414: 1–34.
Bretsky, P.W. 1970b. Upper Ordovician ecology of the central Appalachians. Peabody Museum of Natural History Bulletin 34: 1–150.
Candela, Y. 2006. Late Ordovician brachiopod faunas from Pomeroy, Northern Ireland: a palaeoenvironmental synthesis. Transactions of Royal Society of Edinburgh: Earth Sciences 96: 317–325. Crossref
Carter, J.G., Altaba, C.R., Anderson, L.C., Araujo, R., Biakov, A.S., Bogan, A.E., Campbell, D.C., Campbell, M., Chen, J.H., Cope, J.C.W., Delvene, G., Dijkstra, H.H., Fang, Zong-jie, Gardner, R.N., Gavrilova, V.A., Goncharova, I.A., Harries, P.J., Hartman, J.H., Hautmann, M., Hoeh, W.R., Hylleberg, J., Jiang B.Y., Johnston, P., Kirkendale, L., Kleemann, K., Koppka, K., Kříž, J., Machado, D., Malchus, N., Márquez-Aliaga, A., Masse, J.P., McRoberts, C.A., Middelfart, P.U., Mitchell, S., Nevesskaja, L.A., Özer, S., Pojeta J.J., Polubotko, I.V., Pons, J.M., Popov, S., Sánchez, T., Sartori, A.F., Scott, R.W., Sey, I.I., Signorelli, J.H., Silantiev, V.V., Skelton, P.W., Steuber, T., Waterhouse, J.B., Wingard, G.L., and Yancey, T. 2011. A synoptical classification of the Bivalvia (Mollusca). Paleontological Contributions 4: 1–47.
Carter, J.G., Campbell, D.C., and Campbell, M.R. 2000. Cladistic perspectives on early bivalve evolution. In: E.M. Harper, J.D. Taylor, J.A. Crame (eds.), The Evolutionary Biology of the Bivalvia. The Geological Society of London, Special Publication 177: 47–79. Crossref
Chen, X., Rong, J.Y., Qiu, J.S., Han, N.R., Li, L.Z., and Li, S.J. 1987. A preliminary study of the stratigraphy, sedimentary characteristics and environment of the Late Ordovician in Zhuzhai, Yushan, Jiangxi. Journal of Stratigraphy 11: 23–34. [in Chinese, with English abstract].
Cocks, L.R.M. and Torsvik, T.H. 2002. Earth geography from 500 to 400 million years ago: A faunal and palaeomagnetic review. Journal of the Geological Society, London 159: 631–644. Crossref
Colmenar, J., Villas, E., and Rasmussen, C.M.Ø. 2018. A synopsis of Late Ordovician brachiopod diversity in the Anti-Atlas, Morocco. In: A.W. Hunter, J.J. Álvaro, B. Lefebvre, P. Van Roy, and S. Zamora (eds.), The Great Ordovician Biodiversification Event: Insights from the Tafilalt Biota, Morocco. Geological Society, London, Special Publications 485: 153–163. Crossref
Conrad, T.A. 1838. Report on the palaeontological department of the Survey. Documents of the assembly of the state of New York, sixty-first session 4 (200): 107–119.
Conrad, T.A. 1841. Fifth annual report on the palaeontology of the State of New York. Documents of the assembly of the state of New York, sixty-fourth session 5 (150): 25–57.
Conrad, T.A. 1842. Observations on the Silurian and Devonian systems of the United States, with descriptions of new organic remains. Journal of the Academy of Natural Sciences of Philadelphia 8: 228–280.
Cope, J.C.W. 1996. Bivalves. In: D.A.T. Harper and A.W. Owen (eds.), Fossils of Upper Ordovician, 95–115. The Palaeontological Association, London.
Cope, J.C.W. 1999. Middle Ordovician bivalves from Mid‐Wales and the Welsh Borderland. Palaeontology 42: 467–499. Crossref
Cope, J.C.W. 2000. A new look at early bivalve phylogeny. In: E.M. Harper, J.D. Taylor, J.A. Crame (eds.), The Evolutionary Biology of the Bivalvia. Geological Society of London, Special Publications 177: 81–95. Crossref
Cope, J.C.W. 2004. Bivalve and Rostroconch mollusks. In: B.D. Webby, F. Paris, M.L. Droser, and I.G. Percival (eds.), The Great Ordovician Biodiversification Event, 196–208. Columbia University Press, Oakland. Crossref
Cope, J.C.W. and Kříž J. 2013. The Lower Palaeozoic palaeobiogeography of Bivalvia. In: D.A.T. Harper and T. Servais (eds.), Early Palaeozoic Biogeography and Palaeogeography. Geological Society, London, Memoirs 38: 221–241. Crossref
Dall, W.H. 1889. On the hinge of pelecypods and its development, with an attempt toward a better subdivision of the group. American Journal of Science and Arts (Series 3) 38: 445–462. Crossref
Ebbestad, J.O.R., Polechová, M., Kröger, B., and Gutiérrez-Marco, J.C. 2022. Late Ordovician molluscs of the central and eastern Anti-Atlas, Morocco. Geological Society, London Special Publications 485: 237–296.Crossref
Fang, Z.-J. 2006. An introduction to Ordovician bivalves of southern China, with a discussion of the early evolution of the Bivalvia. Geological Journal 41: 303–328. Crossref
Fang, Z.-J. and Cope, J.C.W. 2004. Early Ordovician bivalves from Dali, West Yunnan, China. Palaeontology 47: 1121–1158. Crossref
Fatka, O. and Mergl, M. 2009. The “microcontinent” Perunica: status and story 15 years after conception. In: M.G. Bassett (ed.), Early Palaeozoic Peri-Gondwanan Terranes: New Insights from Tectonics and Biogeography. Geological Society of London, Special Publications 325: 65–102. Crossref
Férussac, A.E.J.P.J.F. 1821–1822. Tableaux Systèmatiques des Animaux Mollusques Classés en Familles Naturelles, dans Lesquels on a établi la Concordance de tous les Systèmes; Suivis d’un Prodrome Général pour tous les Mollusques Terrestres ou Fluviatiles, Vivants ou Fossiles. 110 pp. Chez Arthus Bertrand, Paris.
Fischer, P. 1886. Manuel de Conchyliologie et de Paléontologie Conchyliologique, ou Histoire Naturelle des Mollusques Vivants et Fossiles suivi d’un Appendice sur les Brachiopodes. 1569 pp. Librairie F. Savy, Paris. Crossref
Fortey, R.A. and Cocks, L.R.M. 2005. Late Ordovician global warming—The Boda event. Geology 33: 405–408. Crossref
Frey, R.C. 1987. The Occurrence of Pelecypods in Early Paleozoic Epeiric-Sea Environments, Late Ordovician of the Cincinnati, Ohio Area. Palaios 2: 3–23. Crossref
Gray, J.E. 1824. Shells. In: W.E. Parry (ed.), A Supplement to the Appendix of Captain Perry’s Voyage for the Discovery of a North West Passage, in the Years 1819–1820, Containing an Account of the Subjects of Natural History. Appendix X, Natural History, 240–246. W. Clowes, John Murray, London.
Grobben, C. 1894. Zur Kenntniss der Morphologie, der Verwandtschaftsverhältnisse und des Systems der Mollusken. Kaiserliche Akademie der Wissenschaften (Mathematisch-Naturwissenschaftlichen Classe), Sitzungsberichte 103(1): 61–86. .
Guo, F.X. 1985. Fossil Bivalves of Yunnan. 319 pp. Yunnan Science and Technology Publishing House, Kunming. [in Chinese].
Hall, J. 1847. Palaeontology of New York, V. I. Containing Description of the Organic Remains in the Lower Division of the New York System. 338 pp. C. Van Benthuysen, Albany.
Hall, J. 1883–1884. Palaeontology: Vol. V. Part I. Lamellibranchiata I. Plates and Explanations. Containing descriptions and figures of the Monomyaria of the upper Helderberg, Hamilton and Chemung Groups. 268 pp. Charles Van Benthuysen and Sons, Albany.
Hall, J. 1885. Palaeontology: Vol. V. Part I. Lamellibranchiata II. Text and Plates. Containing descriptions and figures of the Dimyaria of the upper Helderberg, Hamilton and Chemung Groups. 561 pp. Charles Van Benthuysen and Sons, Albany.
Hall, J. and Whitfield, R.P. 1869–1870. Preliminary notice of the lamellibranchiate shells of the upper Helderberg, Hamilton and Chemung groups, with Others from the Waverly Sandstones, Part 2. 80 pp. New York State Museum, Albany.
Hall, J. and Whitfield, R.P. 1875. Descriptions of Silurian Fossils. In: J.S. Newberry, E. B. Andrews, E. Orton, T.G. Wormley, and F.B. Meek, Report of the Geological Survey of Ohio. Vol. II. Geology and Palaeontology. Part II. Palaeontology, 65–161. Nevins and Myers, Columbus.
Harper, D.A.T. 2001. Late Ordovician brachiopod biofacies of the Girvan district, SW Scotland. Transactions of Royal Society of Edinburgh: Earth Sicences 91: 471–477. Crossref
Harrington, H.J. 1938. Sobre las faunas del Ordoviciano Inferior del Norte Argentino. Revista del Museo de La Plata. Nueva Serie. Sección Paleontología 1: 109–289.
Havlíček, V., Vaněk, J., and Fatka, O. 1994. Perunica microcontinent in the Ordovician (its position within the Mediterranean Province, series division, benthic and pelagic associations). Sborník Geologických Věd, Geologie 46: 23–56.
Hertwig, R. 1895. Lehrbuch der Zoologie. 599 pp. Gustav Fischer, Jena.
Hodges, P. 2000. The Early Jurassic Bivalvia from the Hettangian and Lower Sinemurian of South-West Britan. Monographs of the Palaeontographical Society 154 (614): 1–71. Crossref
Hind, W. 1910. The Lamellibranchs of the Silurian Rocks of Girvan. Transactions of the Royal Society of Edinburgh 47: 479–548. Crossref
Hu, Z.X. 1986. Late Ordovician bryozoans from Yushan County, Jiangxi Province. Acta Micropalaeontologica Sinica 8: 167–183. [in Chinese, with English abstract].
Isberg, O. 1934. Studien über Lamellibranchiaten des Leptaenakalkes in Dalarna. Beitrag zu einer Orientierung über die Muschelfauna im Ordovicium und Silur. 492 pp. Håkan Ohlssons Buchdruckerei, Lund.
Isakar, M. 1991. Harjuan (Late Ordovician) new Bivalves and a new Gastropod from North Estonia. Acta et Commentationes Universitatis Tartuensis 934: 43–60.
Jeon, J.W., Liang, K., Park, J., Kershaw, S., and Zhang, Y.D. 2022. Diverse labechiid stromatoporoids from the Upper Ordovician Xiazhen Formation of South China and their paleobiogeographic implications. Journal of Paleontology 96: 513–538. Crossref
Jin, J.S., Zhan, R.B., Wu, R.C. 2018. Equatorial cold-water tongue in the Late Ordovician. Geology 46: 759–762. Crossref
Jokobsen, K.G., Brock, G.A., Nielsen, A.T., and Harper, D.A.T. 2016. A Darriwilian (Middle Ordovician) bivalve-dominated molluscan fauna from the Stairway Sandstone, Amadeus Basin, central Australia. Acta Palaeontologica Polonica 61: 897–924. Crossref
King, W. 1844. On a new genus of Palaeozoic shells. The Annals and Magazine of Natural History 14: 313–317. Crossref
LaRocque, A. and Newell, N.D. 1969. Family Modiomorphidae Miller, 1877. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology, Part N, Mollusca 6, Bivalvia, vol.1, N393–N399. Geological Society of America, Boulder and University of Kansas, Lawrence.
Lee, D.C. 2013. Late Ordovician trilobites from the Xiazhen Formation in Zhuzhai, Jiangxi Province, China. Acta Palaeontologica Polonica 58: 855–882.
Liljedahl, L. 1994. Silurian nuculoid and modiomorphid bivalves from Sweden. Fossils and Strata 33: 1–89. Crossref
Linnæus, C.A. 1758. Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis. Tomus I, Regnum Animale. 823 pp. Impensis Laurentii Salvii, Stockholm. Crossref
Loi, A., Ghienne, J.F., Dabard, M.P., Paris, F., Botquelen, A., Christ, N., Elaouad-Debbaj, Z., Gorini, A., Vidal, M., Videt, B., and Destombes, J. 2010. The Late Ordovician glacio-eustatic record from a high-latitude storm-dominated shelf succession: The Bou Ingarf section (Anti-Atlas, Southern Morocco). Palaeogeography, Palaeoclimatology, Palaeoecology 296: 332–358. Crossref
Mansuy, H. 1912. Mémoires du Service Géologique de L’indochine. Vol. 1. Fas. 2. E’tude geologique du Yunnan Oriental, 2th partie, Paleontologie. 146 pp. Imprimerie d’Extreme Orient, Hanot-Haiphong.
McAlester, A.L. 1962. Upper Devonian Pelecypods of the New York Chemung Stage. Bulletin of the Peabody Museum of Natural History, Yale University 16: 1–88.
McAlester, A.L. 1968. Type Species of Paleozoic Nuculoid Bivalve Genera. Geological Society of America Memoirs 105: 1–140. Crossref
McAlester, A.L. 1969. Superfamily Nuculacea and Family Praenuculidae. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology, Part N. Mollusca 6, Bivalvia, Vol. 1, N229–N230. Geological Society of America, Boulder and University of Kansas, Lawrence.
Miller, S.A. 1877. The American Palaeozoic Fossils: A Catalog of the Genera and Species with Names of Authors, Dates, Places of Publication, Groups of Rocks in which found, and the Etymology and Signification of the Words, and an Introduction devoted to the Stratigraphical Geology of the Palaeozoic Rocks. 253 pp. Published by author, Cincinnati. Crossref
Morris, N.J., Dickins, J.M., and Astafieva-Urbajtis, K. 1991. Upper Palaeozoic Anomalodesmatan Bivalvia. British Museum (Natural History), Bulletin (Geology Series) 47: 51–100.
Newell, N.D. 1965. Classification of the Bivalvia. American Museum Novitates 2206: 1–25.
Nevesskaja, L.A., Scarlato, O.A., Starobogatov, Y.I., and Eberzin, A.G. 1971. Novye predstavleniia o sisteme dvustvorchatykh molliuskov. Paleontologicheskii Zhurnal 1971(2): 3–20. [in Russian].
Pelseneer, P. 1889. Sur la classification phylogenetique de pelecypods (communication preliminaire). Bulletin Scientifique de la France et de la Belgique 3: 27–52.
Perner, J. 1903. Systême silurien du centre de la Bohême par Joachim Barrande. 1st partie: Recherches Paléontologiques, Vol. IV. Gastéropodes. 164 pp. Imprimerie de Charles Bellmann, Prague.
Pojeta, J.J. 1971. Review of Ordovician pelecypods. Geological Survey Professional Paper 695. 46 pp. United States Government Printing Office, Washington. Crossref
Pojeta, J.J. and Gilbert-Tomlinson J. 1977. Australian Ordovician Pelecypod Molluscs. 64 pp. Australian Government Publishing Service, Canberra.
Polechová, M. 2019. The bivalve Cuneamya from the Late Ordovician of Bohemia. Fossil Imprint 75: 120–127. Crossref
Polechová, M. 2022. The bivalve fauna from the Letná Formation (Upper Ordovician) of Bohemia: Significance for palaeobiogeography, palaeoecology and diversification of bivalves. Geobios 70: 55–73. Crossref
Portlock, J.E. 1843. Report on the geology of the County of Londonderry, and of parts of Tyrone and Fermanagh. 784 pp. Milliken, Hodges and Smith and Longmans, Dublin.
Reed, F.R.C. 1946. Notes on some lamellibranchs from Quarrel Hill, Girvan. Geological Magazine 83: 201–205. Crossref
Rong, J.Y. and Han, N.R. 1986. Preliminary report on Upper Ordovician (Mid-Ashgillian) brachiopods from Yushan, northeastern Jiangxi (Eastern China). 485–491. In: P.R. Rachebceuf and C. Emig (eds.), Les Brachiopodes fossiles et actuels. Les actes du 1er Congrès international sur les Brachiopodes, Brest, 1985. Biostratigraphie du Paléozoïque 4: 1–500.
Rong, J.Y. and Chen, X. 1987. Faunal differentiation, biofacies and lithofacies pattern of Late Ordovician (Ashgillian) in South China. Acta Palaeontologica Sinica 26: 507–535. [in Chinese, with English introduction].
Runnegar, B. 1974. Evolutionary history of the Bivalve subclass Anomalodesmata. Journal of Paleontology 48: 904–939.
Sánchez, T.M. 1990. Bivalvos del Ordovicico Medio-Tardio de la Precordillera de San Juan (Argentina). Ameghiniana 27: 251–261.
Sánchez, T.M. 1999. New Late Ordovician (Early Caradoc) bivalves from the Sierra de Villicum (Argentina Precordillera). Journal of Paleontology 73: 66–76. Crossref
Sánchez, T.M. 2006. Taxonomic position and phylogenetic relationships of the bivalve Goniophorina Isberg, 1934, and related genera from the early Ordovician of northwestern Argentina. Ameghiniana 43: 113–122.
Sánchez, T.M. and Babin, C. 2003. Distribution paléogéographique des mollusques bivalves durant l’Ordovicien. Geodiversitas 25: 243–259.
Sharpe, D. 1853. Descriptions of the new species of Zoophyta and Mollusca. Quarterly Journal of the Geological Society of London 9: 143–158.
Soot-Ryen, H. 1964. Nuculoid pelecypods from the Silurian of Gotland. Arkiv för Mineralogi och Geologi Series 3 28: 489–519.
Soot-Ryen, H. and Soot-Ryen, T. 1960. The middle Ordovician of the Oslo region, Norway. Pelecypoda. Norsk Geologisk Tidsskrift 40: 81–122.
Stanley, S.M. 2015. Evolutionary ecology of the Bivalvia. Treatise Online Part N, Revised, Volume 1 72: 1–48.
Tomlin, L.R. 1931. Some preoccupied generic names. Proceedings of the Malacological Society of London 19 (4): 174–175.
Toni, R.T. 1975. Upper Ordovician bivalves from the Oslo Region, Norway. Norsk Geologisk Tidsskrift 55: 135–156.
Tunnicliff, S.P. 1982. A revision of Late Ordovician bivalves from Pomeroy Co., Tyrone, Ireland. Palaeontology 25: 43–88.
Ulrich, E.O. 1890. New Lamellibranchiata. In: S. Calvin, E.W. Claypole, J. Eyerman, P. Frazer, A. Lakes, F.W. Cragin, A.C. Lawson, C. Herrick, E.O. Ulrich, I.C. White, A. Winchell, and N.H. Winchell (eds.), The American Geologist A monthly journal of geology and allied sciences 6: 173–181.
Ulrich, E.O. 1892. New lower Silurian Lamellibranchiata chiefly from Minnesota rocks. Minnesota Geological and Natural History Survey 19: 211–248.
Ulrich, E.O. 1894. The Lower Silurian Lamellibranchiata of Minnesota. In: E.O. Ulrich, W.H. Scofield, J.M. Clarke, and N.H. Winchell (eds.), Final Report of the Geological and Natural History Survey of Minnesota 3: 475–628. Crossref
Verneuil, E. and Barrande, J. 1855. Description des fossiles trouvés dans les terrains siluriens et dévoniens d’Almaden, de la Sierra Morena et des monta gnes de Tolède. Bulletin de la Société Géologique de France 12: 964–1025.
Zhan, R.B. and Cocks, L.R.M. 1998. Late Ordovician brachiopods from the South China Plate and their palaeogeographical significance. Special Papers in Palaeontology 59: 5–70.
Zhan, R.B. and Fu, L.P. 1994. New observations on the Upper Ordovician stratigraphy of Zhejiang-Jiangxi border region, E. China. Journal of Stratigraphy 18: 267–274. [in Chinese, with English abstract].
Zhan, R.B. and Rong, J.Y. 1994. Tashanomena, A new strophomenoid genus from Middle Ashgill rocks (Ordovician) of Xiazhen, Yushan, NE. Jiangxi, East China. Acta Palaeontologica Sinica 33: 416–428. [in Chinese, with English abstract].
Zhan, R.B. and Rong, J.Y. 1995a. Distribution pattern of Late Ordovician brachiopod communities in Zhejiang-Jiangxi border. Chinese Science Bulletin 40: 932–935. [in Chinese].
Zhan, R.B. and Rong, J.Y. 1995b. Four new Late Ordovician brachiopod genera from the Zhejiang-Jiangxi border region, East China. Acta Palaeontologica Sinica 34: 549–574. [in Chinese, with English abstract].
Zhan, R.B., Rong, J.Y., Jin, J.S., and Cocks, L.R.M. 2002. Late Ordovician brachiopod communities of southeast China. Canadian Journal of Earth Sciences 39: 445–468. Crossref
Zhang, M., Xia, F.S., Taylor, P.D., Liang, K., and Ma, J.Y. 2018. Upper Ordovician bryozoans from the Xiazhen Formation of Yushan, northeastern Jiangxi, East China. Palaeoworld 27: 343–359. Crossref
Acta Palaeontol. Pol. 71 (1): 133–153, 2026
http://doi.org/10.4202/app.01281.2025