
A new azhdarchoid pterosaur from the Lower Cretaceous of Brazil and the paleobiogeography of the Tapejaridae
GABRIELA M. CERQUEIRA, MATEUS A.C. SANTOS, MAIKON F. MARKS, JULIANA M. SAYÃO, and FELIPE L. PINHEIRO
Cerqueira, G.M. Santos, M.A.C., Marks, M.F., Sayão, J.M., and Pinheiro, F.L. 2021. A new azhdarchoid pterosaur from the Lower Cretaceous of Brazil and the paleobiogeography of the Tapejaridae. Acta Palaeontologica Polonica 66 (3): 555–570.
The Tapejaridae were an apparently worldwide distributed clade of edentulous pterosaurs, being a major component of several Lower Cretaceous terrestrial faunas. Despite their distribution across Gondwana and Laurasia, the oldest tapejarid remains were found in Barremian units from Europe, what led to the assumption that the clade originated in Eurasia and later dispersed southwards. Here we present a new tapejarid pterosaur species (Kariridraco dianae gen. et sp. nov.) from the Lower Cretaceous Romualdo Formation of Brazil. The holotype (MPSC R 1056) comprises an incomplete, three dimensionally preserved skull, lower jaw, and cervical vertebrae. It shows a unique combination of features such as unusually tall and comparatively short nasoantorbital fenestrae, as well as a premaxillary crest forming an angle of about 45° with respect to main skull axis. Phylogenetic analyses recover the new taxon as a Tupuxuara-related Thalassodrominae, a clade of early-diverging tapejarids that were apparently indigenous to central Gondwana. The inclusion of the new taxon in current phylogenetic frameworks, in addition to similarity cluster analyses of Early Cretaceous tapejarid-bearing pterosaur faunas, indicate Gondwana as the most parsimonious origin center for Tapejaridae, and show that pterosaur communities were affected by large scale tectonic-driven vicariant events.
Key words: Pterosauria, Azhdarchoidea, phylogeny, biogeography, Cretaceous, Romualdo Formation, Araripe Basin, Brazil.
Gabriela M. Cerqueira [gabrielamenezes0301@gmail.com], Maikon F. Marks [maikon.fortes.marks@gmail.com] and Felipe L. Pinheiro [felipepinheiro@unipampa.edu.br] (corresponding author), Laboratório de Paleobiologia, Universidade Federal do Pampa, Rua Aluízio Barros Macedo, s/n. BR 290 – km 423. São Gabriel, RS, Brazil.
Mateus A.C. Santos [mateus.costas@ufrgs.br], Programa de Pós-Graduação em Geociências, Instituto de Geociências, Universidade Federal do Rio Grande do Sul, Av. Bento Gonçalves, 9500, 91501-970, Bairro Agronomia, Porto Alegre, Rio Grande do Sul, Brazil.
Juliana M. Sayão [jmsayao@gmail.com], Seção de Museologia, Museu Nacional/Universidade Federal do Rio de Janeiro, Quinta da Boa Vista, s/n, Bairro Imperial de São Cristóvão, 20940-040, Rio de Janeiro, Rio de Janeiro, Brazil.
Received 22 October 2020, accepted 29 March 2021, available online 14 September 2021.
Copyright © 2021 G.M. Cerqueira et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Romualdo Formation of the Araripe Basin (Lower Cretaceous, Northeastern Brazil) is a well-known fossil Konservat-Lagerstätte, especially notorious for the occurrence of some of the best three-dimensionally preserved pterosaur specimens worldwide, as well as for its diverse pterosaur fauna (e.g., Kellner et al. 2002, 2013; Vila Nova et al. 2011; Bantim et al. 2014; Pinheiro and Rodrigues 2017). A major component of the Santana Group pterosaur fauna, the Tapejaridae, is a peculiar clade of toothless animals that draws attention due the extreme morphology observed in some of its representatives. Tapejarids were so far reported in Lower Cretaceous strata from China, Morocco, Spain, England and Brazil (e.g., Kellner 1989; Wellnhofer and Buffetau 1999; Wang and Zhou 2003; Holgado et al. 2011; Vullo et al. 2012; Kellner et al. 2019; Martill et al. 2020a, b). The tapejarid clade Thalassodrominae is thus far exclusive to the Lower Cretaceous of Northeastern Brazil (Kellner and Campos 2007), whereas the Tapejarinae, the most diverse tapejarid clade, had a widespread distribution in Gondwana and Laurasia (Martill et al. 2020a, b).
The Thalassodrominae were originally defined by Kellner and Campos (2007) as those tapejarids presenting high nasoantorbital fenestra and a premaxillary sagittal crest beginning on the anterior part of the skull and expanding posteriorly. The Tapejarinae, the other tapejarid subclade, would allocate those pterosaurs with a downturned rostral end of the maxilla, a premaxillary crest constricted above the nasoantorbital fenestra, as well as for having orbits positioned only slightly below the dorsal margin of the nasoantorbital fenestra (Kellner and Campos 2007, Pinheiro et al. 2011). Despite being consistently recovered as a basal tapejarine (e.g., Kellner 2013; Holgado et al. 2019; Kellner et al. 2019), the Brazilian tapejarid Caupedactylus ybaka shares with the Thalassodrominae a tall, unconstricted premaxillary crest, and orbits positioned very low with respect to the nasoantorbital fenestrae.
Tapejarine pterosaurs have a wider stratigraphic distribution when compared to other tapejarids, and their oldest remains were recovered from Barremian European units (Vullo et al. 2012; Martill et al. 2020a). This led to the conclusion that the Tapejaridae originated in Eurasia, posteriorly spreading southwards to diversify in central Gondwana (Vullo et al. 2012; but see Pêgas et al. 2016).
Here we report a new edentulous pterosaur species from the Romualdo Formation, consisting of an incomplete skull associated to the lower jaws and cervical vertebrae I–IV. Besides adding to the pterosaur diversity of the Araripe Basin, the new taxon contributes with new information on tapejarid biogeography, challenging a Eurasian origin for this clade.
Institutional abbreviations.—AMNH, American Museum of Natural History, New York, USA; CPCA, Centro de Pesquisas Paleontológicas da Chapada do Araripe, Crato, Brazil; IMCF, Iwaki Coal and Fossil Museum, Japan; MCT, DGM, Museu de Ciências da Terra Rio de Janeiro, Brazil; MN, Museu Nacional, Rio de Janeiro, Brazil; MPSC, Museu de Paleontologia Plácido Cidade Nuvens, Santana do Cariri, Brazil; NMSG, Naturmuseum St. Gallen, Switzerland; SMNK, Staatliches Museum für Naturkunde, Karlsruhe, Germany; UFC, Universidade Federal do Ceará, Fortaleza, Brazil.
Other abbreviations.—Char., character; OTU, operational taxonomic unit.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:3031C608-123C-422C-820D-249892E8CBA5
Material and methods
Phylogenetic analyses.—Cladistic analyses were performed to test the relationships of Kariridraco dianae gen. et sp. nov. within Pterosauria. The new specimen (MPSC R 1056) was included as an operational taxonomic unit (=OTU) in a modified version of Holgado et al. (2019) dataset, resulting in a data matrix that totalizes 146 characters and 60 OTUs. Updates in Holgado et al. (2019) original dataset are as follows: (i) inclusion of two additional OTUs (Keresdrakon vilsoni and Tupuxuara longicristatus); (ii) corrections on the original coding of several tapejarids for character 53, following first-hand observation of specimens; and (iii) inclusion of two additional characters (Char. 36: premaxillary sagittal crest, when starting at close or at the anterior portion of the skull, tilt angle [0] under 30°, [1] 30–45°, [2] 45–60°, [3] over 60°, character inapplicable for pterosaurs lacking a rostral premaxillary crest; and Char. 146: palate, surface at mid-length [0] flat or concave, [1] convex). Updated scores for K. vilsoni were included following the modified dataset of Holgado and Pêgas (2020). Our preferred analysis excludes a priori this last OTU, as it was detected that its inclusion generates considerable noise during parsing. The data matrix was edited using MESQUITE v.3.51 (Maddison and Maddison 2018), and is included as a SOM, Supplementary Online Material available at http://app.pan.pl/SOM/app66-Cerqueira_etal_SOM.pdf. Heuristic searches were performed using the TNT (Tree analysis using New Technology) version 1.5 (Goloboff and Catalano 2016) software. Searches were conducted using TNT tree bisection reconnection swapping algorithm, with 1000 replications and random seed.
Faunistic similarity indexes.—The Jaccard similarity index is a β-diversity method used to estimate similarity between three or more sample sites using presence/absence data and paired groups. In Jaccard index, 0 indicates a total dissimilarity between a pair of sites, and a 1 value means that both sites are equal (Sales et al. 2018).
Here we use the Jaccard similarity index to quantify overall taxonomic similarity between different tapejarid-bearing pterosaur faunas. Only pterosaur data was considered for each unit, being recorded the presence/absence of different groups (using subfamilies as preferred Linnaean taxonomic hierarchy, see SOM). All sampled units contain tapejarid representatives, but our raw sample is not restricted to this clade. Sampled sedimentary units are distributed both in Laurasia and Gondwana, and they are as follows: Crato, Romualdo, and Goio-Erê formations (Brazil); Jiufotang and Yixian formations (China); Kem Kem Group (Morocco); Wessex Formation (England, UK) and La Huérguina Formation (Spain). As the Hungarian Csehbánya Formation, thus far records only one nominal pterosaur taxon, we choose to prune it a priori from our dataset. The list of sampled units and recorded taxa considered in the present study is displayed in the SOM. To calculate Jaccard indexes, we used the clustering analyses included in the software package PAST v. 3.18 (Hammer et al. 2001) and the data was presented as a dendrogram.
Geological setting
The material described herein comes from a carbonate concretion found in the Romualdo Formation (e.g., Saraiva et al. 2007), one of the units that compose the Araripe Basin’s sedimentary succession. The Romualdo Formation comprises a wide range of lithologies and is characterized by conglomeratic sandstone followed by a sequence of green and black shale (Assine 2007), stratified conglomerates, fine-to medium-grained sandstone, laminated limestone, marl, and coquina levels (Assine et al. 2014; Melo et al. 2020). The black shale is occasionally intercalated with several layers where carbonate concretions are abundant (Fara et al. 2005; Saraiva et al. 2007; Vila Nova et al. 2011; Kellner et al. 2013). These layers probably were deposited following pulses of mass mortality, and the lithologies characterize a coastal/marine environment (Melo et al. 2020). The Romualdo Formation was recently considered as Aptian in age, based on palynomorph, foraminifera and ostracod data (Arai and Assine 2020; Melo et al. 2020). The Romualdo Formation crops out on the slopes of the Araripe Plateau, Northeastern Brazil. The precise locality were the new material was found is, however, unknown.
Systematic palaeontology
Pterosauria Kaup, 1834
Pterodactyloidea Plieninger, 1901
Azhdarchoidea Nesov, 1984 (sensu Unwin 2003)
Tapejaridae Kellner, 1989
Thalassodrominae Kellner and Campos, 2007
Genus Kariridraco nov.
Zoobank LSID: urn:lsid:zoobank.org:act:77359171-69E4-4C18-A52A-285F1CF4294D
Etymology: After the Kariris, former indigenous people that lived in the area of the Araripe Plateau, combined with the Latin draco, dragon.
Type species: Kariridraco dianae gen. et sp. nov., see below.
Diagnosis.—As for the type species, by monotypy.
Kariridraco dianae gen. et sp. nov.
Zoobank LSID: urn:lsid:zoobank.org:act:D311DC9D-B9ED-4D0A-BAC1-6DC36584B9D5
Etymology: In reference to Diana Prince, alter ego of the DC fictional character Wonder Woman.
Holotype: MPSC R 1056 consists of a fairly complete skull, preserving most of its temporal, orbital, and preorbital portions, part of its premaxillary crest, an almost complete lower jaw and four cervical vertebrae (Fig. 1). Mechanical preparation revealed that the original concretion bearing the fossil was a composite, in which the rostral ends of the premaxillae and dentaries of a second pterosaur specimen was glued to the holotype. In order to avoid misinterpretations, we choose to only illustrate those elements that safely belong to a single individual. A photograph of the composite material is depicted in the SOM: fig. 1.
Type locality: The exact location where MPSC R 1056 was found is, unknown, since the specimen was donated to MPSC by local workers. The Romualdo Formation is geographically restricted to the Araripe Plateau, Northeastern Brazil.
Type horizon: Romualdo Formation, Aptian, Lower Cretaceous.
Diagnosis.—Kariridraco dianae gen. et sp. nov. differs from all other pterosaurs for the following combination of characters (characters marked with * represent autapomorphies): presence of a subtle jugal shelf; comparatively short and tall nasoantorbital fenestra; premaxillary crest forming an angle of about 45° with respect to the maxilla*; lacrimal with an anteriorly-directed deep concavity*, and a single elliptic lateral pneumatic foramen in the left lateral surface of cervical IV centrum.
Description.—The holotype (MPSC R 1056) consists of a partially preserved skull, including the premaxilla, maxilla, premaxillary crest, part of the orbital and temporal regions (jugal, lacrimal, quadrate/quadratojugal), in addition to an almost complete lower jaw. Associate postcranial elements include the atlas-axis complex and two mid-cervicals. The specimen lacks most of the rostrum anterior to the nasoantorbital fenestra, as well as the premaxillary crest above the temporal region and part of its skull roof.
MPSC R 1056 presents the typical Romualdo Formation preservation, lacking evidence of lateral distortion. The lower jaw has a preserved length of 205 mm and is still in close association to the skull. Both upper and lower jaws are edentulous, a typical feature within Azhdarchoidea (Unwin 2003; Kellner 2004; Witton 2007; Kellner et al. 2019). The most striking features of the new specimen are the exceptionally tall nasoantorbital fenestra and the well-developed, steep premaxillary crest, this latter having its outermost bony layer eroded and exposing trabecular bone. The orbit is piriform, with a subvertical anterior margin limited by the lacrimal ramus of the jugal and the lacrimal itself. The dorsoposterior margin of the orbits, which would be formed by frontoparietals, are absent. In close association to the orbits, K. dianae gen. et sp. nov. preserves a slit-like lower temporal fenestra. Most of the occiput was lost, but a wide occipital plate is visible posterior to the lower temporal fenestra, being detached from its original position. Cervical vertebrae were preserved articulated, still attached to the skull, but were posteriorly isolated through mechanical preparation.
The unusually steep premaxillary crest and the fact that the rostrum anterior to this structure was adulterated by fossil dealers led us to raise concerns about the crest authenticity. Careful examination of the fossil and surrounding matrix revealed that, although there are signs of a breakage close to the base of the crest (where it contacts the main body of the premaxillomaxilla), the two resulting counterparts fit together, with no signs of add-ons or other modifications. Bone surface at the base of the crest was eroded off, so that trabecular bone is exposed at that portion. Additionally, the carbonate concretion remained uniform at the surroundings of the breakage, differing, for instance, from the composite rostrum (Fig. 1, SOM: fig. 1), in which the anterior portion of the upper and lower jaws of a second individual was glued together with MPSC R 1056. This second specimen was preserved in a different, much lighter in color, carbonate matrix. Crest base preparation removed uniform rock throughout the whole procedure, while mechanical removal of matrix in composite specimens fatally menages to reveal add-ons (usually epoxy resin) used to glue pieces together. It is also noteworthy that, close to the anteroventral border of the nasoantorbital fenestra, the premaxillomaxilla thickens to delineate the rounded outline of the aperture, showing that, MPSC R 1056 indeed had an unusually short nasoantorbital fenestra. This dorsal expansion of the jugal process of the premaxillomaxilla is compatible with the crest’s position, steepness and shape.
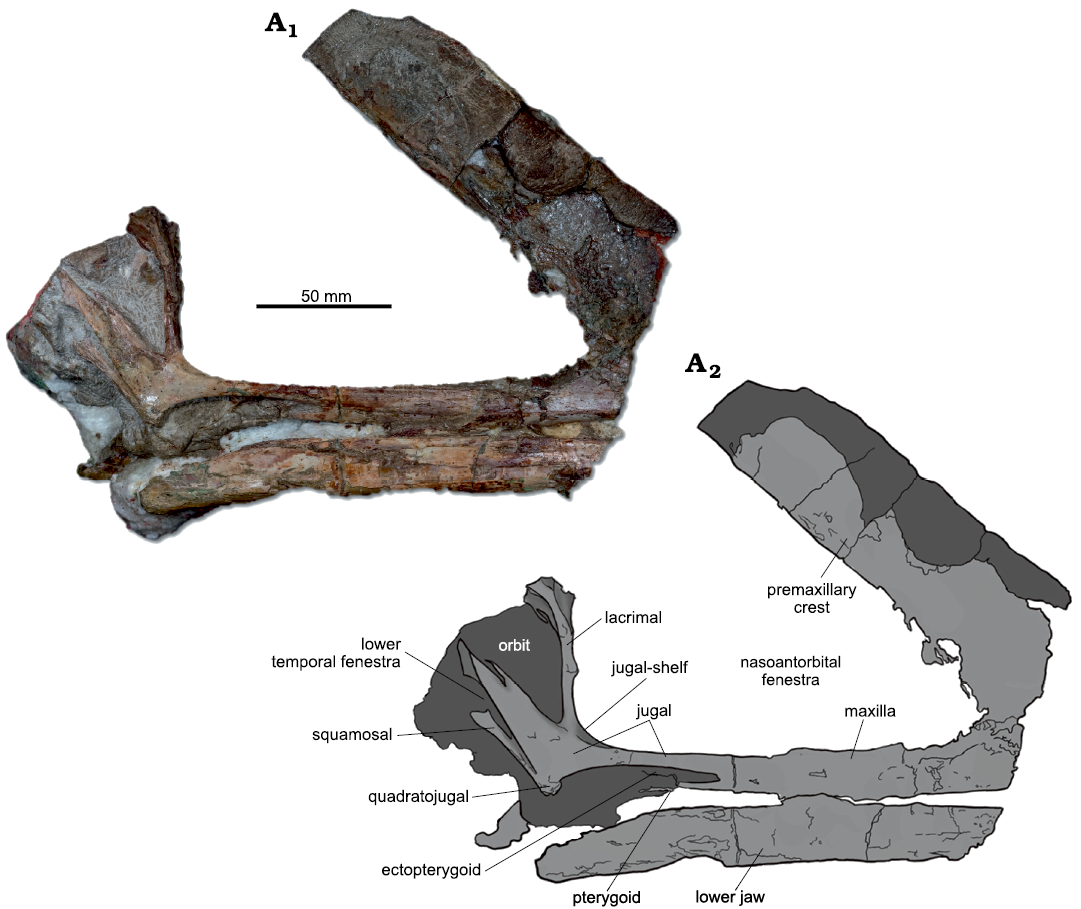
Fig. 1. Azhdarchoid pterosaur Kariridraco dianae gen. et sp. nov. holotype (MPSC R 1056) from Lower Cretaceous of Northeastern Brazil. Skull in right lateral view. Photograph (A1), interpretative drawing (A2). Dark grey areas indicate remains of the carbonate matrix still attachaed to the skull.
Premaxillomaxilla: The most striking feature of the fused premaxilla/maxilla is the well-developed sagittal crest. As preserved, this structure has a total length of 202 mm. The crest is broken posterodorsally, so that it is not possible to estimate its total original length. Parts of the crest are still embedded in carbonate, as its delicate nature prevented total preparation without further damaging, even using standard techniques for Santana crested pterosaurs (Silva and Kellner 2006). The posteroventral margin of the crest is notably straight and forms the preserved anterior borders of the nasoantorbital fenestrae (Fig. 1). The exact spot where the premaxillary crest would contact the frontoparietals is impossible to estimate. Close to its base, the crest widens slightly, so that the anteroventral end of this structure is deeper than the posterodorsal one.
The anterior-most portion of the premaxillomaxillae is not preserved. The premaxillomaxillae anteriorly border the large and wide nasoantorbital fenestrae, the anterior limit of which is unusually steep (about 45° with respect to the horizontal plane) (Fig. 1A2). Most of the anterodorsal and ventral borders of the nasoantorbital fenestrae are also limited by the premaxillomaxillae. These latter contact the jugals posteroventrally, even though the precise suture between both bones cannot be determined with confidence. Ventrally, the premaxillomaxillae bend medially to form broad palatal plates anterior to the choanae (see Ősi et al. 2010; Pinheiro and Schultz 2012). Incomplete preservation prevents the recognition of the entire extent of the premaxillomaxillae, as these bones display an anteroventrally-oriented breakage close to the anterior border of the nasoantorbital opening. Even though the specimen is visible only in the right lateral view, it is clear that the palatal surface of the maxillary palatal plate is strongly convex.
Jugal: Despite being difficult to recognize suture lines with the surrounding bones, the right jugal is apparently triradiate, with three well-developed processes (Fig. 1A2). The jugal is well-preserved (except for a fracture later restored), not showing signs of erosion. Anteriorly, a maxillary process apparently contacts the maxilla close to the ventral midpoint of the nasoantorbital fenestra. The dorsally-oriented lacrimal process detaches itself from the main body of the jugal by the presence of a medial recess, ventrally bordered by a moderately-developed jugal shelf. The lacrimal process is very thin and is positioned at an almost right angle with respect to the maxillary process. Posterodorsally, the jugal tapers to form an acute process that makes most of the preserved dorsoventral orbital margin.
Lacrimal: The lacrimal is exposed in its right lateral view, being its medial surface still embedded in matrix. The tridimensional conformation of this bone was not altered by preservation. The ventral limit of the lacrimal is positioned at the mid orbital height, and this bone broadens dorsally to make the posterodorsal margin of the nasoantorbital opening. Albeit laterally eroded, the anterior margin of the lacrimal, which forms the posterodorsal border of the nasoantorbital fenestra, shows a deep anterior, dorsoventrally oriented groove (Fig. 1). Posteriorly, the lacrimal displays a posteriorly-directed bend, outlining the anterior margin of the pear-shaped orbit.
Quadratojugal: The quadratojugal is fused to the main corpus of the jugal, forming a thin rod that limits posteroventrally the lower temporal fenestra.
Quadrate: The right quadrate is preserved as a thin bone rod that contributes to the posteroventral margin of the lower temporal fenestra, having contact with the squamosal and quadratojugal. Quadrate inclination with respect to the horizontal plane is of about 130°.
Squamosal: Only a fragment of the right squamosal is preserved, forming a thin process that reaches the middle posterior margin of the orbit. Together with the quadrate and quadratojugal the squamosal also composes the posterior margin of the lower temporal fenestra.
Palate: The palatal surface of Kariridraco dianae gen. et sp. nov. is preserved, except for the anterior most portion and the articular portion for the lower jaw. Only the right pterygoid is visible, being still embedded in matrix and not completely preserved. It connects with the maxilla in its anterior portion, where a fibrous joint marks the limits between both bones (Fig. 1). A thin bone rod adjacent to the pterygoid may be a lateral process to the pterygoid main body. The pterygoid has no preserved connection with the ectopterygoid or jugal. A small portion of the ectopterygoid is also preserved, still attached to the jugal. The proximal portion of this bone rod is, however, still covered by matrix, obscuring the connection between the former and the pterygoid.
Lower jaw: The lower jaw is almost complete and still occluded to the skull. It only lacks the rostral-most portion (anterior to the nasoantorbital fenestra). Bone fusion, however, precludes the exact identification of the posterior mandibular bones. Posterior to the jaw junction, a ventrally-slopping retroarticular process is visible. Albeit displaying a fair preservation, the lower jaw displays a restored fracture spot in its mid-length. There is no evidence of a dentary crest in its preserved portion, but this structure could have been present in the missing rostral part. The lower jaw appears to be completely edentulous (Fig. 1).
Atlas-axis: The atlas-axis complex is completely fused, what is usually displayed by adult individuals (e.g., Bennett 1993; Kellner 2015; Fig. 2). The atlas is fused to the axis by its annular components, located at the base of the vertebra. Above the annular components is the round-shaped neural canal, which is not surrounded by pneumatic foramina anteriorly (Fig. 2A1, A4). In posterior view, however, the neural canal is laterally flanked by two small elliptical pneumatic foramina, one in each side (Fig. 2A2, A5). In both its left and right lateral surfaces, the atlas-axis complex displays a small rounded pneumatic foramen located in the middle portion of the axial centrum (Fig. 2A3, A6).
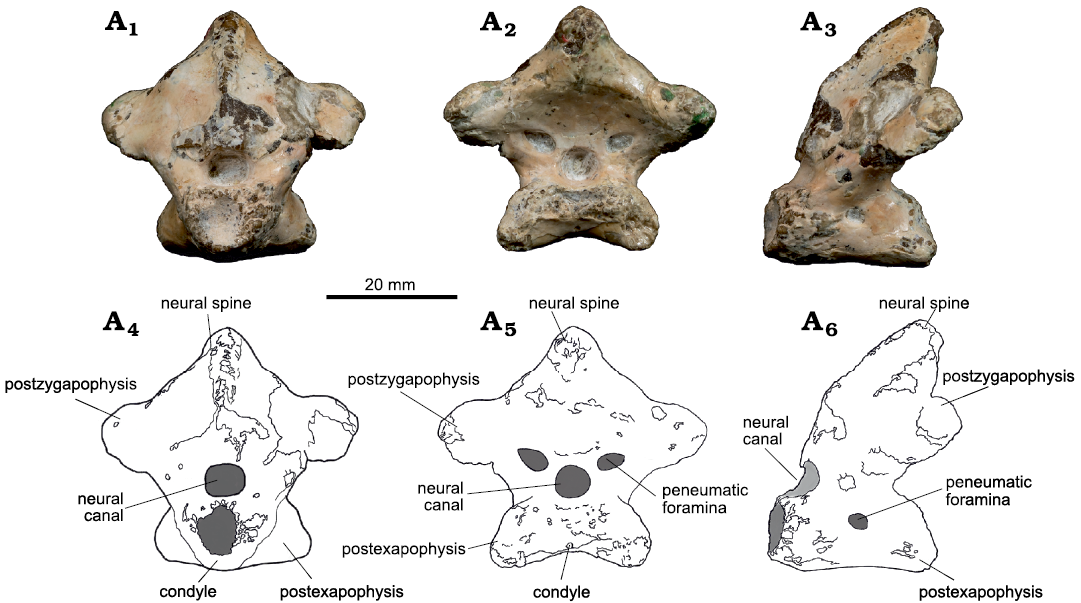
Fig. 2. Azhdarchoid pterosaur Kariridraco dianae gen. et sp. nov. holotype (MPSC R 1056) from Lower Cretaceous of Northeastern Brazil. Atlas-axis complex in anterior (A1, A4), posterior (A2, A5), and left lateral (A3, A6) views. Photographs (A1–A3), interpretative drawings (A4–A6).
The axis presents a well-developed, almost complete neural spine, displaying some erosion in its dorsal and anterior portions (Fig. 2, 3). It binds the well-developed postzygapophyses, which form an angle of 135° between each other. In lateral view, the neural spine forms an angle of about 10° with the vertical plane close to its base, bending backwards to form an angle of 40° in its superior portion. Posteriorly, the axis has well-developed postexapophyses.
The condyle is located dorsal to the postexapophyses. It is elliptical in shape, with its transverse length much larger than its height. It is dorsally followed by a concavity that forms the ventral margin of the neural canal. There is a slight slope lateral to this concavity, and the ventral margin of the centrum between the postexapophyses is convex (Fig. 3A1, A3).
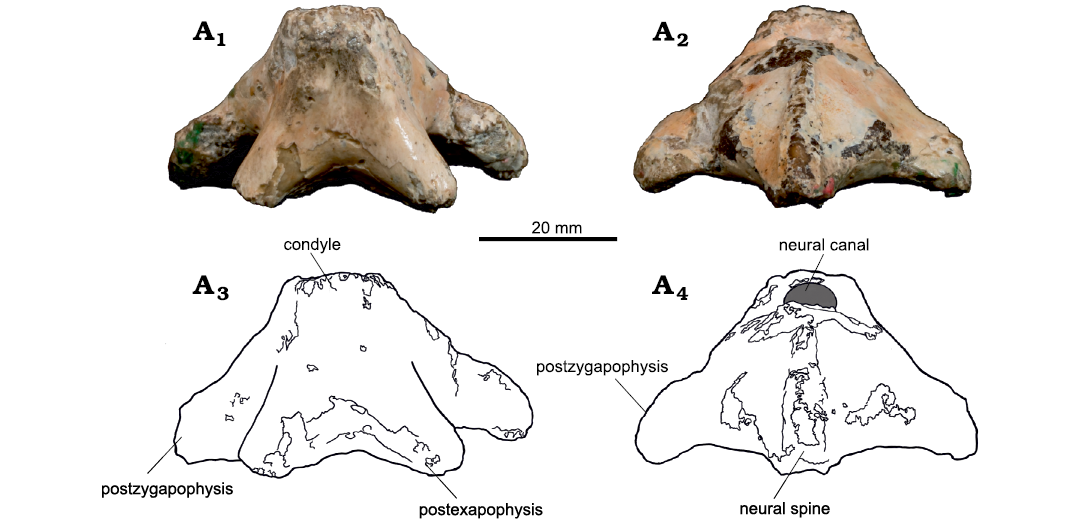
Fig. 3. Azhdarchoid pterosaur Kariridraco dianae gen. et sp. nov. holotype (MPSC R 1056) from Lower Cretaceous of Northeastern Brazil. Atlas-axis complex in ventral (A1, A3) and dorsal (A2, A4) views. Photographs (A1, A2), interpretative drawings (A3, A4).
Vertebrae III and IV: The third cervical vertebra has a total length of 45.79 mm and a maximum width of 44.13 mm. It displays a good preservation, except for its left ventral surface, which shows a circular erosion and slight fragmentation. The vertebra has a subcircular neural canal, flanked by two lateral pneumatic foramina both in its anterior and posterior portions (Fig. 4).
Cervical III centrum apparently lacks lateral pneumatic foramina (Fig. 4A3, A6). While its left surface is incomplete, the right one is well-preserved and not pierced by any aperture. The neural spine of the third vertebra is incomplete but was apparently moderately tall. It expands laterally in its base to form the pre- and postzygapophyses (Fig. 5A1, A3). These latter are dorsally limited by dorsomedially directed laminae, which connects the tip of the zygapophyses to the neural spine. Albeit better preserved, the prezygapophyses are less developed than the postzygapophyses.
The hypapophysis is a small bone protuberance positioned ventrally to the cotyle, while the postexapophyses are well developed and almost complete (Fig. 5). Their ventral portions show slight slopes, also observable in lateral view. Such slopes, however, do not interfere with the overall flat ventral surface of the centrum. The condyle is dorsal to the postexapophyses. Its dorsal portion has a concavity that connects to the neural canal, forming its ventral margin. Its dorsal portion presents slight lateral slopes on both sides. The condyle is directed horizontally with relation to the main axis of the vertebra. The neural canal is flanked by two highly asymmetric pneumatic foramina (Fig. 4A2, A5).
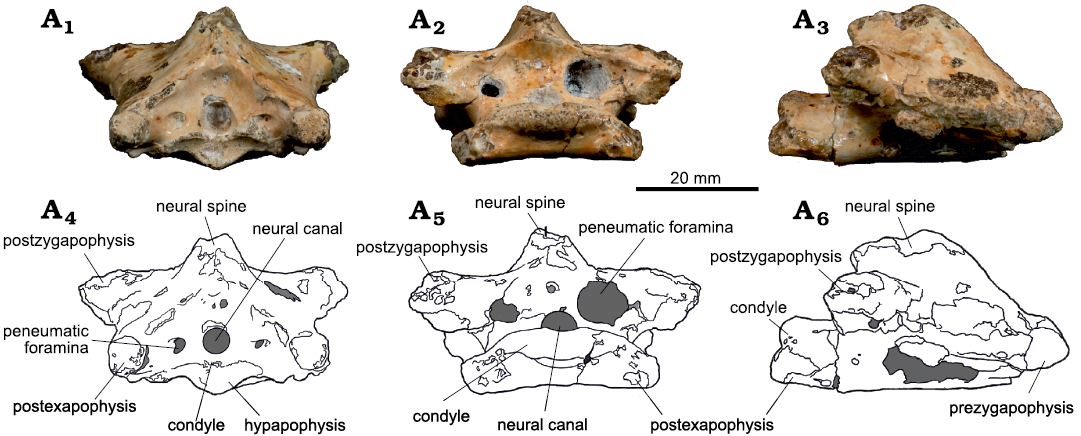
Fig. 4. Azhdarchoid pterosaur Kariridraco dianae gen. et sp. nov. holotype (MPSC R 1056) from Lower Cretaceous of Northeastern Brazil. Cervical vertebra III in anterior (A1, A4), posterior (A2, A5), and right lateral (A3, A6) views. Photographs (A1–A3), interpretative drawings (A4–A6).
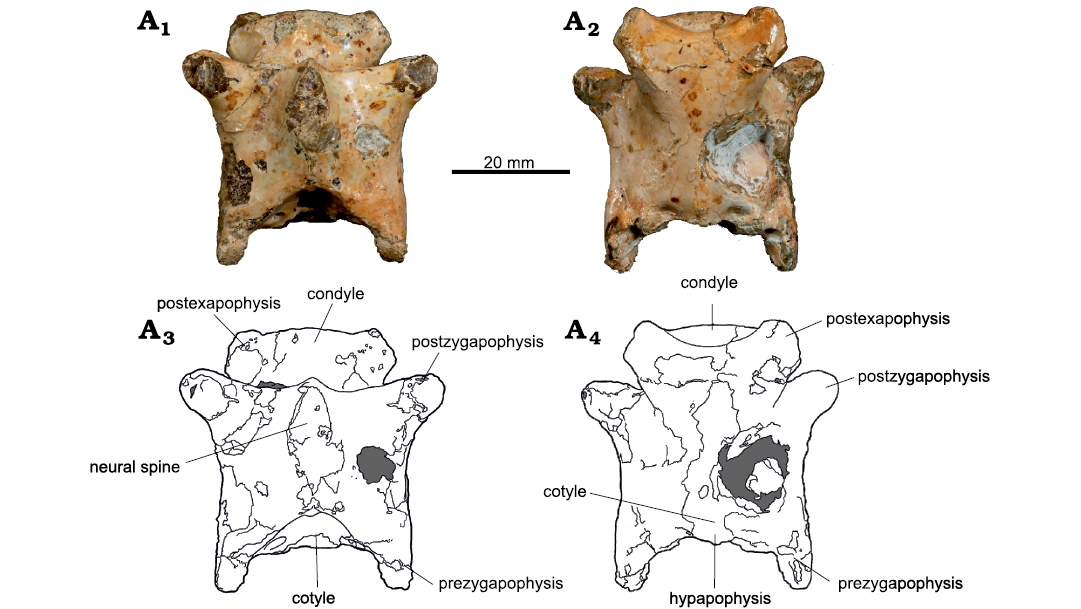
Fig. 5. Azhdarchoid pterosaur Kariridraco dianae gen. et sp. nov. holotype (MPSC R 1056) from Lower Cretaceous of Northeastern Brazil. Cervical vertebra III in dorsal (A1, A3) and ventral (A2, A4) views. Photographs (A1, A2), interpretative drawings (A3, A4).
Cervical IV is poorly preserved. It has its dorsal surface almost completely eroded. The neural canal, as well its adjacent structures, is absent. This is also the case for the neural spine, condyle, postzygapophyses and postexapophyses (Fig. 6). There is a small pneumatic foramen on the ventral left surface of the centrum, close to where the left postzygapophysis would originate (Fig. 6A3, A6). The vertebra IV has a slight concavity in its ventral surface (Fig. 6A3, A6), which is more prominent than what is displayed by cervical III. The hypapophysis is fragmented, but reveals to be better developed when compared to what is observed in cervical III. The right prezygapophysis is better preserved than the left one, which lacks its anterior portion. The right prezygapophysis presents its original conformation and is well pronounced (Fig. 7).
Stratigraphic and geographic range.—Type locality and horizon only.
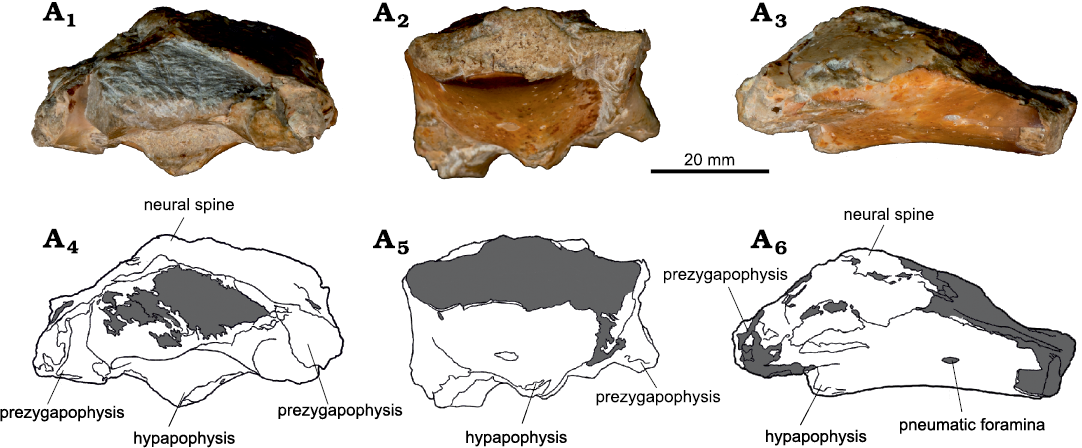
Fig. 6. Azhdarchoid pterosaur Kariridraco dianae gen. et sp. nov. holotype (MPSC R 1056) from Lower Cretaceous of Northeastern Brazil. Cervical vertebra IV in anterior (A1, A4), posterior (A2, A5), and left lateral (A3, A6) views. Photographs (A1–A3), interpretative drawings (A4–A6).

Fig. 7. Azhdarchoid pterosaur Kariridraco dianae gen. et sp. nov. holotype (MPSC R 1056) from Lower Cretaceous of Northeastern Brazil. Cervical vertebra IV in dorsal (A1, A3) and ventral (A2, A4) views. Photographs (A1, A2), interpretative drawings (A3, A4).
Phylogenetic results
Our phylogenetic assessment of Kariridraco dianae gen. et sp. nov. using the modified version of Holgado et al. (2019) dataset resulted in 6 most parsimonious trees (MPTs) of 345 steps each. The strict consensus tree (Fig. 8) recovers a monophyletic Tapejaridae (sensu Kellner and Campos 2007). Kariridraco dianae gen. et sp. nov. is recovered as the sister taxon of Tupuxuara (T. leonardii + T. longicristatus). Thalassodromeus sethi Kellner and Campos, 2002 stands as the sister taxon to (Kariridraco + Tupuxuara) and, as such, Thalassodrominae was recovered as monophyletic. The Tapejarinae was also recovered and, within it, Caupedactylus ybaka Kellner, 2013, and Aymberedactylus cearensis Pêgas, Leal, and Kellner, 2016, are successive sister taxa to a clade formed by Eopteranodon lii Lü and Zhang, 2005, “Huaxiapterus” benxiensis Lü, Gao, Xing, Li, and Ji 2007, “Huaxiapterus” corollatus Lü, Jin, Unwin, Zhao, Azuma, and Ji, 2006, Sinopterus dongi Wang and Zhou, 2003, Tapejara wellnhoferi Kellner, 1989, Europejara olcadesorum Vullo, Marugán-Lobón, Kellner, Buscalioni, Gomez, Fuente, and Moratalla, 2012, Caiuajara dobruskii Manzig, Kellner, Weinschütz, Fragoso, Vega, Guimarães, Godoy, Liccardo, Ricetti, and Moura 2014, and Tupandactylus imperator Campos and Kellner, 1997. Notably, Chinese tapejarines form a monophyletic group, whereas Brazilian representatives of this clade are dispersed in the topology, being either recovered as early-diverging tapejarines (Caupedactylus ybaka and Aymberedactylus cearensis) or forming a clade together with the Laurasian Europejara. The clade formed by Kariridraco and Tupuxuara species is held by a single unambiguous synapomorphy (Char. 146, palatal surface convex at mid-length). Our second analysis, including Keresdrakon vilsoni resulted in 15 equally parsimonious trees of 348 steps. Although the consensus topology displays Keresdrakon as the sister-taxon to the Tapejaridae (in agreement with Kellner et al. 2019), tapejarid taxa relationships are mainly unresolved. Only the Thalassodrominae (Tupuxuara spp. + Thalassodromeus sethi + Kariridraco dianae gen. et sp. nov.) was recovered as a clade, being placed in a polytomy together with Aymberedactylus cearensis, Caupedactylus ybaka and a clade formed by the remaining tapejarines (Tapejara wellnhoferi, Europejara olcadesorum, Caiuajara dobruskii, Tupandactylus imperator, Sinopterus dongi, “Huaxiapterus” spp., and Eopteranodon lii) (SOM: fig. 2).
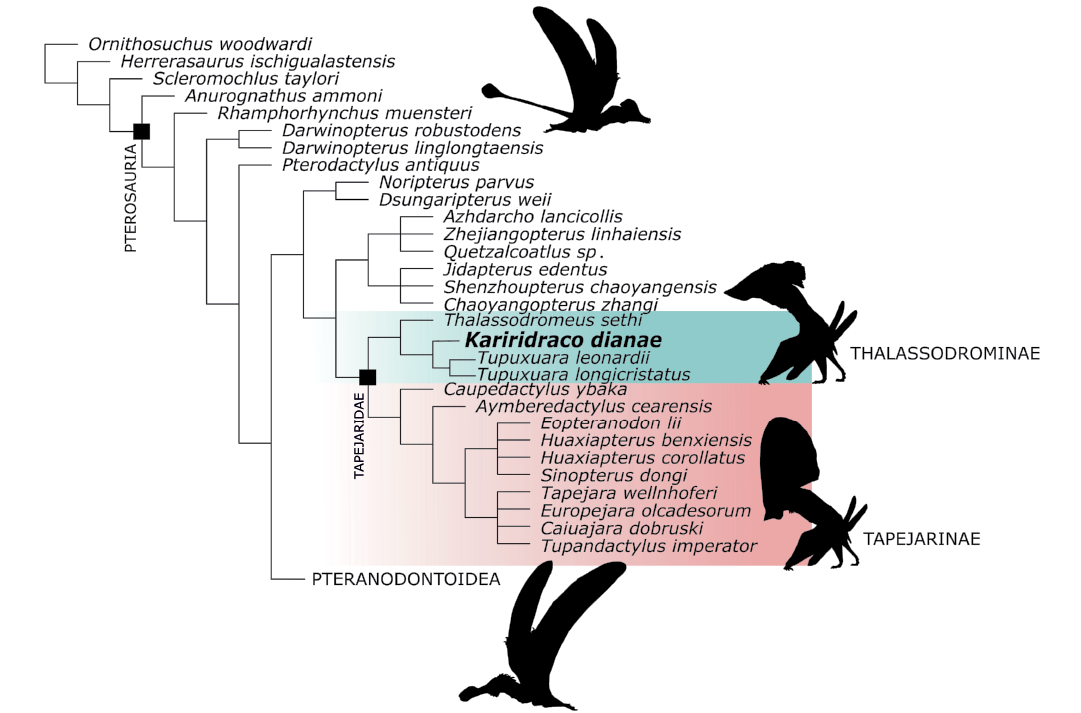
Fig. 8. Simplified strict consensus of the 6 most parsimonious trees recovered in the preferred cladistic analysis. Silhouettes based on drawings by Felipe A. Elias (São Paulo, Brazil).
Similarity indexes for tapejarid-bearing pterosaur faunas
Our results demonstrate a high similarity index between Wessex and La Huérguina formations (England and Spain, respectively), even though pterosaur specimens are comparatively rare in both units (Fig. 9B). The resulted dendrogram also evidences that the pair formed by La Huérguina and Wessex formations is more akin to the other two Laurasian units sampled here (Yixian and Jiufotang formations) than to the Gondwanan Crato, Romualdo, Goiô-Erê formations (Brazil) and the Kem Kem beds (Morocco). On the other hand, the Kem Kem beds and Crato Formation cluster with the Romualdo Formation, being a higher similarity detected between Crato Formation and the Kem Kem beds. The recovered position of Goio-Erê Formation may be a sampling bias, as only two pterosaur taxa were so far recovered from this unit.
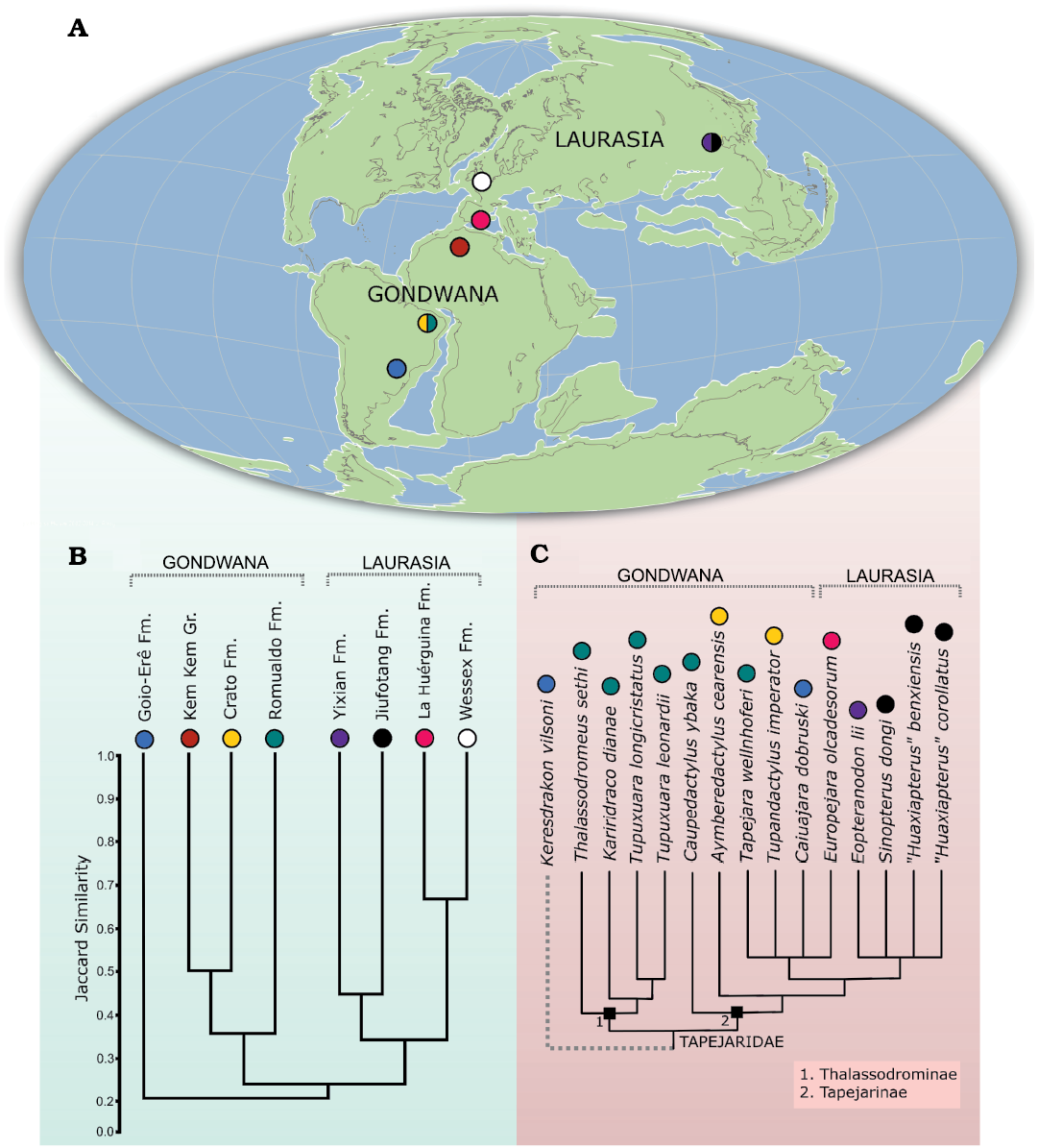
Fig. 9. Paleobiogeography of the Tapejaridae. A. Cretaceous paleomap showing the distribution of tapejarid-bearing localities (colored circles refer to sedimentary units depicted in B). B. Jaccard similarity index applied to different tapejarid-bearing pterosaur faunas. C. Tapejarid phylogeny recovered in this paper, highlighting locality data for each species.
Discussion
Comparisons.—As preserved, Kariridraco dianae gen. et sp. nov. jaws are entirely edentulous, and empty alveoli are absent. Among Romualdo Formation pterosaurs, the lack of teeth is only displayed by the Tapejaridae (e.g., Kellner 2004). A different condition is seen in the well-represented anhanguerians (Rodrigues and Kellner 2013).
Although edentulous chaoyangopterid pterosaurs have been previously reported for the Araripe Basin, they are so far restricted to the lower Crato Formation. The most informative putative chaoyangopterid thus far reported for the Crato beds is Lacusovagus magnificens (Witton 2008), which is represented by a partially complete skull, preserving its anterior rostrum (SMNK PAL 4325). Kariridraco dianae gen. et sp. nov. clearly differs from L. magnificens by the presence of a well-developed premaxillary crest. Comparisons between palatal bones are impossible because L. magnificens holotype (SMNK PAL 4325) is still partially embedded by matrix. More recently, Leal et al. (2018) described mid-cervical vertebrae (IV–VII) putatively attributable to L. magnificens (UFC-721). Direct comparisons between UFC-721 and K. dianae gen. et sp. nov. are also not possible because the latter preserves only the first four vertebrae. We should note, however, that K. dianae gen. et sp. nov. cervicals display well-developed neural spines, whereas Leal et al. (2018) describe low neural spines for UFC-721. Also, Leal et al. (2018) report the absence of lateral pneumatic foraminae and presence of pronounced postexapophyses. Albeit K. dianae gen. et sp. nov. also displays well-developed postexapophyses, lateral pneumatic foraminae are present in cervical IV. UFC-721 also differs from K. dianae gen. et sp. nov. in having the typical centrum elongation of chaoyangopterids (Zhou 2010; Leal et al. 2018), azhdarchids (Suberbiola et al. 2003; Ösi et al. 2005; Vremir et al. 2013) and ctenochasmatids (Andres and Ji} 2008).
Kariridraco dianae gen. et sp. nov. displays typical tapejarid features (Fig. 10), such as a nasoantorbital fenestra corresponding to more than 45% of the total skull size (estimated); orbit showing an inverted pear shape; long sagittal crest formed mainly by the premaxilla and extending posteriorly, and a thin subvertical lacrimal process of jugal (Kellner 2004; Kellner and Campos 2007). The monophyly of Tapejaridae was questioned by some authors, such as Unwin and Lü (1997) and Unwin (2003), who agree in that Tapejaridae sensu Kellner (2003) would be paraphyletic with respect to Azhdarchidae. According to those, Tupuxuara and Thalassodromeus would share a series of characters in common with azhdarchids, while absent in Tapejara-like taxa. This hypothesis was later supported by, for instance, Martill and Naish (2006) and Lü et al. (2006). Pinheiro et al. (2011), however, recovered Tapejaridae sensu Kellner and Campos (2007) as monophyletic, in agreement with Kellner (2004) and other previous works (Wang et al. 2005, 2008, 2009; Andres and Ji 2008).
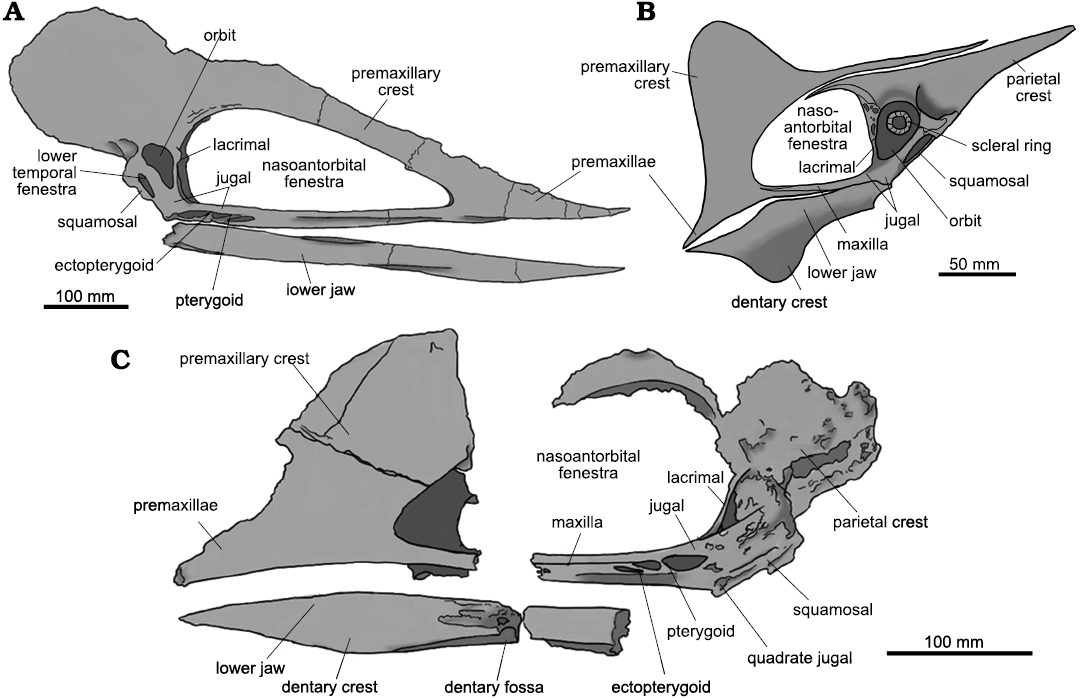
Fig. 10. Interpretative drawings of skulls. A. Tupuxuara leonardii (Thalassodrominae), IMCF 1052, in right lateral view. B. Tapejara wellnhoferi (Tapejarinae), reconstruction based mostly on AMNH 24440, in left lateral view. C. Caupedactylus ybaka (Tapejarinae), MN 4726-V, in left lateral view.
Kellner and Campos (2007) divide the Tapejaridae in two independent clades with sister-taxa relationship. The Tapejarinae would include those tapejarids with a downturned rostrum, premaxillary crest constricted above the nasoantorbital fenestra, low posterior expansion of the cranial crest and orbits positioned on a level only slight below the dorsal margin of the nasoantorbital fenestra (Kellner and Campos 2007; Pinheiro et al. 2011; Fig. 10B). Tapejarinae includes Tapejara wellnhoferi, Tupandactylus imperator, Tupandactylus navigans Frey, Martill, and Buchy 2003, Sinopterus dongi, “Huaxiapterus” jii Lü and Yuan, 2005, “Huaxiapterus” corollatus, “Huaxiapterus” benxiensis, Eopteranodon lii, Europejara olcadesorum, Caupedactylus ybaka, Caiuajara dobruskii, Aymberedactylus cearensis, Sinopterus lingyuanensis Lü, Teng, Sun, Shen, Li, Gao, and Liu, 2016, Sinopterus atavismus (Lü, Teng, Sun, Shen, Li, Gao, and Liu, 2016), and Wightia declivirostris Martill, Green, Smith, Jacobs, and Winch, 2020. By their turn, the Thalassodrominae are characterized by having very high nasoantorbital fenestrae and by a high premaxillary crest which is not constricted above the nasoantorbital openings (Fig. 10A). This clade comprises pterosaurs so far indigenous to the Early Cretaceous of southern Gondwana (Brazil), such as Thalassodromeus sethi, Thalassodromeus oberlii (Headden and Campos, 2015), Tupuxuara longicristatus Kellner and Campos, 1988, Tupuxuara leonardii Kellner and Campos, 1994, and putatively “Tupuxuara” deliradamus. It is noteworthy that, albeit being consistently recovered as an early-diverging tapejarine, Caupedactylus ybaka shares with thalassodromines a high premaxillary crest that is unconstricted above the nasoantorbital openings.
With respect to the Tapejarinae, Kariridraco dianae gen. et sp. nov. differs from Tapejara wellnhoferi by several features, such as the triangular shape of the nasoantorbital fenestra (sub-oval in Tapejara wellnhoferi, based in the specimen AMNH 24440) and the overall morphology of the sagittal crest (Kellner 1989; Wellnhofer and Kellner 1991). It also differs from Tupandactylus by the triangular shape of its nasoantorbital fenestra, the presence of a jugal shelf and an angle of 150° between the quadrate with respect to the jugal bar. Tupandactylus has an elliptical nasoantorbital fenestra, apparently lacks a jugal shelf and has an anteriorly high sagittal crest that constricts itself above the nasoantorbital openings (based on specimens MCT 1622-R and CPCA 3590) (Campos and Kellner 1997; Pinheiro et al. 2011). Kariridraco dianae gen. et sp. nov. resembles Caupedactylus ybaka in having a steep anterior margin of the nasoantorbital fenestra (steeper in K. dianae gen. et sp. nov.); orbits positioned low in the skull; an unconstricted premaxillary crest (the last two characters being also displayed by thalassodromines); and a jugal shelf (better developed in Caupedactylus ybaka). However, K. dianae gen. et sp. nov. is easily distinguishable from C. ybaka for having a much lower premaxillary crest, a convex palate below the nasoantorbital fenestrae (mainly flat in C. ybaka), a more vertical lacrimal process of the jugal and a much less developed jugal shelf.
As for the Thalassodrominae, Kariridraco dianae gen. et sp. nov. shares with Thalassodromeus sethi (holotype DGM 1476 R), a somewhat triangular nasoantorbital fenestra, a 90° angle between the lacrimal and the maxillary processes of the jugal and a jugal shelf. Despite that, the nasoantorbital fenestra is higher in K. dianae gen. et sp. nov. than what is displayed by T. sethi (Pêgas et al. 2018), having also a much steeper anterior margin (about 45° with respect to the horizontal plane in its anterior portion). Additional differences between the new taxon and T. sethi include its palatal morphology, strongly concave posteriorly in T. sethi (Kellner and Campos 2007; Pêgas et al. 2018) and distinctly convex in K. dianae gen. et sp. nov. We do not have any elements to directly compare K. dianae gen. et sp. nov. with Thalassodromeus oberlii, as the latter is only known by its holotype (NMSG SAO 251093), a mandibular symphysis. We, however, agree with Pêgas et al. (2018) in that NMSG SAO 251093 can be confidently attributed to a species of Thalassodromeus and, thus, probably came from a very distinct animal with respect to K. dianae gen. et sp. nov.
The near triangular-shaped nasoantorbital fenestra, a 90° angle of the lacrimal process with respect to the jugal bar and a convex palate are features shared between Kariridraco dianae gen. et sp. nov. and Tupuxuara leonardii (IMCF 1052). They differ, however, by the lack of the jugal-shelf in this later, as well as much shorter nasoantorbital openings and a steeper sagittal crest in K. dianae. Although known only by a fragmentary skull (holotype MN-6591-V), Tupuxuara longicristatus differs from K. dianae gen. et sp. nov. in having a much less steep premaxillary crest and apparently longer nasoantorbital openings. In addition, the dorsal edge of T. longicristatus premaxillary crest is slightly convex, differing from both K. dianae gen. et sp. nov. and T. leonardii. “Tupuxuara” deliradamus (SMNK PAL 6410) has an angle of 150° between quadrate and the maxillary process of the jugal, which is similar to the condition displayed by Caupedactylus ybaka (MN 4726-V, Kellner, 2013, Fig. 10B), differing from K. dianae gen. et sp. nov. Further comparisons between K. dianae gen. et sp. nov. and “Tupuxuara” deliradamus are hindered by the poor preservation of the holotype of the latter (SMNK PAL 6410), precluding the estimation of nasoantorbital fenestrae height and shape (Witton 2009). We should note that “Tupuxuara” deliradamus is a highly problematic taxon, the validity of which will be assessed in a future contribution.
The total preserved cervical vertebrae complex (from atlas-axis to cervical IV) of K. dianae gen. et sp. nov. has a set of features only displayed by Thalassodrominae, such as well pronounced postexapophyses, reduced hypapophysis, and pronounced, hatchet-like, neural spines (Aires et al. 2014; Vila Nova et al. 2015). The presence of two foramina adjacent to the neural canal and the flat ventral surface of the vertebrae is also peculiar to thalassodromines (Vila Nova et al. 2015). Regarding the atlas-axis complex, K. dianae gen. et sp. nov. is also similar to taxa of Thalassodrominae in having a flat ventral surface and two foramina adjacent to the neural canal. However, the atlas-axis shows a small single lateral foramen on both sides of the element, a feature previously observed in Tapejara wellnhoferi (SMNK PAL 1135; Eck et al. 2011). The presence of adjacent foramina in the fourth vertebra is impossible to confirm due to its poor preservation. Eck et al. (2011) mentions the presence of a single foramen on either side of the centrum of Tapejara wellnhoferi cervicals. The presence of a single foramen in both sides of K. dianae gen. et sp. nov. cervical IV may result from individual variation.
Biogeography of Tapejaridae.—The paleobiogeography of pterosaurs is an underappreciated topic, and geographic distribution patterns of tapejarids have been accessed solely by seldom and punctual discussions (e.g., Vullo et al. 2012; Pêgas et al. 2016; Martill et al. 2020a). An attempt was proposed by Sayão et al. (2012) when presenting a comparison of the distribution patterns of pterosaurs in light of the phylogenies existing at the time. It was concluded that the phylogenies available were based on taxa from a few locations, specially from Fossillagerstattën, which prevented the application of cladistic and event-based biogeographic methods (Sayão et al. 2012).
Previous works sometimes relied on ambiguous evidence to determine origin centers for the Tapejaridae. As an example, a Eurasian origin was proposed for this clade based on the recovery of slightly older taxa in Europe and China, when compared to South American representatives (Vullo et al. 2012). The sole occurrence of older remains of a specific taxon is, however, well-established as a fragile criterion to determine origin centers (Cain 1944), especially when dealing with taxa that are notorious for their patchy and biased fossil record, as is the case of pterosaurs (Sayão et al. 2012; Butler et al. 2013).
The results of our faunistic similarity assessment of tapejarid-bearing units evidence a clear large-scale provincialism between Gondwanan and Laurasian pterosaur communities (Fig. 9A, B). This was already briefly argued for tapejarines by Martill et al. (2020a) but note that our results are based in all the taxa recovered from each sampled locality. Also, similarity indexes respect geographic distance in Laurasian faunas, with La Huérguina and Wessex formations having a much higher average similarity when compared to the Chinese Yixian and Jiufotang formations. The similarity between pterosaur communities from Northeastern Brazil and Morocco agrees with the results of Sales et al. (2018), who detected a similar pattern for dinosaurian faunas. We should note, however, that the recovered similarity may (or may not) be confirmed by future works, since comparisons between Brazilian and Moroccan pterosaur faunas are hindered by the still incipient pterosaur discoveries in Morocco (Kellner and Mader 1997; Mader and Kellner 1999; Jacobs et al. 2020). In spite of that, our results suggest that, as occurred for strictly terrestrial faunas, pterosaur communities were also affected by large scale tectonic-driven vicariant events.
Although pterosaurs were probably more capable to overcome geographical barriers when compared with terrestrial, non-flying, animals (Sayão et al. 2012), it should also be taken into account that smaller-scale phenomena such as high rates of oceanic spread, volcanism, excessive atmospheric CO2 heating the surface temperature (Weissert et al. 2003) possibly also contributed to the framework that would explain pterosaur distribution patterns.
When analyzed together, tapejarid phylogeny is in perfect agreement with their geographic distribution. The earliest-diverging clade (Thalassodrominae) is restricted to Gondwanan deposits, whereas the specialized tapejarines are widely spread throughout Gondwana and Laurasia, even though they seem to respect smaller scale provinces, as is suggested by the monophyletic Chinese tapejarines (Fig. 9C). The apparent restriction of early-diverging forms to Gondwana, allied to the fact that the sister-taxon to the Tapejaridae (Keresdrakon vilsoni) was recovered from Southern Brazil, suggests a Gondwanan origin for the clade, what is in disagreement with the proposition of Vullo et al. (2012). We should note that similar conclusions as we propose here have already been suggested by Pêgas et al. (2016) who, based on a similar topology as we recovered, defied Vullo et al. (2012) Eurasian origin for the Tapejaridae. The presence of slightly older tapejarid remains in Europe and China (Vullo et al. 2012; Martill et al. 2020a) can be readily explained by the strong “Lagerstätten effect” that affects pterosaur fossil record, resulting in severe sampling biases that can obscure large-scale diversity patterns (Butler et al. 2009, 2013; Sayão et al. 2012).
Conclusions
Kariridraco dianae gen. et sp. nov. (Fig. 11) has a set of diagnostic features that distinguishes it from other azhdarchoid pterosaurs, such as the presence of a concavity in the upper portion of the lacrimal and an angle of about 45° between the maxilla and the premaxillary crest. Kariridraco dianae gen. et sp. nov. was consistently recovered as a thalassodromine tapejarid, closely related to the well-known genus Tupuxuara. Our assessment of tapejarid paleobiogeography recovered Gondwana as the most parsimonious origin center for the Tapejaridae, also suggesting that, similarly to strictly terrestrial animals, pterosaur distribution is marked by tectonic-driven provincialism.

Fig. 11. Life reconstruction of azhdarchoid pterosaur Kariridraco dianae gen. et sp. nov. Artwork by Júlia d’Oliveira (Jundiaí, Brazil).
Acknowledgements
The authors thank the late Dr. Plácido Cidade Nuvens (1943–2016), former director of the Museu de Paleontologia Plácido Cidade Nuvens for access to the specimen MPSC R 1056. FLP would like to thank the following people for allowing the study of specimens under their care and for kind help during visits: Carl Mehling and Mark Norell (both AMNH); Álamo Saraiva and João Kerensky (both MPSC); Oliver Rauhut and Markus Moser (both Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany); Alexander Kellner and Helder Silva (both MN); Eberhard Frey (SMNK); Rainer Schoch (Naturkundemuseum, Stuttgart, Germany); Sandra Chapman and Lorna Steel (both Natural History Museum, London, UK); Dan Pemberton and Matt Riley (both Sedgwick Museum of Earth Sciences, Cambridge, UK). We also thank Rodrigo Pêgas (Universidade Federal do ABC, São Bernardo do Campo, Brazil.) and Borja Holgado (Museu Nacional, Rio de Janeiro, Brazil) for their insightful reviews of an early version of the manuscript. FLP and JMS are supported by grants from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq process numbers 407969/2016-0 and 305758/2017-9 to FLP; 458164/2014-3, 311715/2017-6 and 314222/2020-0 to JMS).
References
Aires, A.S., Kellner, A.W., Müller, R.T., Da Silva, L.R., Pacheco, C.P. and Dias-Da-Silva, S. 2014. New postcranial elements of the Thalassodrominae (Pterodactyloidea, Tapejaridae) from the Romualdo Formation (Aptian–Albian), Santana Group, Araripe Basin, Brazil. Palaeontology 57: 343–355. Crossref
Andres, B. and Ji, Q. 2008. A new pterosaur from the Liaoning Province of China, the phylogeny of the Pterodactyloidea, and convergence in their cervical vertebrae. Palaeontology 51: 453–469. Crossref
Arai, A. and Assine, M.L. 2020. Chronostratigraphic constraints and paleoenvironmental interpretation of the Romualdo Formation (Santana Group, Araripe Basin, Northeastern Brazil) based on palynology. Cretaceous Research 116: 104610. Crossref
Assine, M.L. 2007. Bacia do Araripe. Boletim de Geociências da PETROBRAS 15: 371–389.
Assine, M.L., Perinotto, J.A.J., Custódio, M.A., Neumann, V.H., Varejão, F.G., and Mescolotti, P.C. 2014. Sequências deposicionais do Andar Alagoas da Bacia do Araripe, Nordeste do Brasil. Boletim de Geociências da PETROBRAS 22: 3–28. Crossref
Bantim, R.A.M., Saraiva, A.A.F., Oliveira, G.R., and Sayão, J.M. 2014. A new toothed pterosaur (Pterodactyloidea: Anhangueridae) from the Early Cretaceous Romualdo Formation, NE Brazil. Zootaxa 3869: 201–223. Crossref
Benett, S.C. 1993. The ontogeny of Pteranodon and other pterosaurs. Paleobiology 19: 92–106. Crossref
Butler, R.J., Barrett, P.M., Nowbath, S., and Upchurch, P. 2009. Estimating the effects of the rock record on pterosaur diversity patterns: implications for hypotheses of bird/pterosaur competitive replacement. Paleobiology 35: 432–446. Crossref
Butler, R.J., Benson, R.B.J., and Barrett P.M. 2013. Pterosaur diversity: Untangling the influence of sampling biases, Lagerstätten, and genuine biodiversity signals. Palaeogeography, Palaeoclimatology, Palaeoecology 372: 78–87. Crossref
Cain, S.A. 1944. Foundations of Plant Geography. 556 pp. Harper and Brothers, New York .
Campos, D.D.A. and Kellner, A. W. A. 1997. Short note on the first occurrence of Tapejaridae in the Crato Member (Aptian), Santana Formation, Araripe Basin, Northeast Brazil. Anais da Academia Brasileira de Ciências 69: 83–88.
Eck, K., Elgin, R.A., and Frey, E. 2011. On the osteology of Tapejara wellnhoferi Kellner 1989 and the first occurrence of a multiple specimen assemblage from the Santana Formation, Araripe Basin, NE-Brazil. Swiss Journal of Palaeontology 130: 277. Crossref
Fara, E., Saraiva, A.A.F, Campos, D.A., Moreira, J.K.R, Siebra, D.C. and Kellner, A.W.A. 2005. Controlled excavations in the Romualdo Member of the Santana Formation (Early Cretaceous, Araripe Basin, northeastern Brazil): stratigraphic, palaeoenvironmental and palaeoecological implications. Palaeogeography, Palaeoclimatology, Palaeoecology 218: 145–160. Crossref
Goloboff, P.A. and Catalano, S.A. 2016. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32: 221–238. Crossref
Hammer, Ø., Harper, D.A.T., and Ryan, P.D. 2001. PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4: 1–9.
Holgado, B. and Pêgas, R.V. 2020. A taxonomic and phylogenetic review of the anhanguerid pterosaur group Coloborhynchinae and the new clade Tropeognathinae. Acta Palaeontologica Polonica 65: 743–761. Crossref
Holgado, B., Martínez-Pérez, C., and Ruiz-Omeñaca, J.I. 2011. Revisión actualizada del registro fósil de Pterosauria en la Península Ibérica. In: A. Pérez-García, F. Gascó, J.M. Gasulla, F. Escaso (eds.), Viajando a Mundos Pretéritos. Ayuntamiento de Morella, Morella, 183–194. Ayuntamiento de Morella, Morella, Castellón.
Holgado, B., Pêgas, R.V., Canudo, J.I., Fortuny, J., Rodrigues, T., Company, J., and Kellner, A.W.A. 2019. On a new crested pterodactyloid from the Early Cretaceous of the Iberian Peninsula and the radiation of the clade Anhangueria. Scientific Reports 9, 4940. Crossref
Jacobs, M.L., Martill, D.M., Unwin, D.M., Ibrahim, N., Zouhri, S., and Longrich, N.R. 2020. New toothed pterosaurs (Pterosauria: Ornithocheiridae) from the middle Cretaceous Kem Kem beds of Morocco and implications for pterosaur palaeobiogeography and diversity. Cretaceous Research 110: 104413. Crossref
Kellner, A.W.A. 1989. A new edentate pterosaur of the Lower Cretaceous from the Araripe Basin, Northeast Brazil. Anais da Academia brasileira de Ciências 61: 439–446.
Kellner, A.W.A. 2004. New information on the Tapejaridae (Pterosauria, Pterodactyloidea) and discussion of the relationships of this clade. Ameghiniana 41: 521–534.
Kellner, A.W.A. 2013. A new unusual tapejarid (Pterosauria, Pterodactyloidea) from the Early Cretaceous Romualdo Formation, Araripe Basin, Brazil. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 103: 409–421. Crossref
Kellner, A.W.A. 2015. Comments on Triassic pterosaurs with discussion about ontogeny and description of new taxa. Anais da Academia Brasileira de Ciências 87: 669–689. Crossref
Kellner, A.W.A. and Campos, D.A. 2007. Short note on the ingroup relationships of the Tapejaridae (Pterosauria, Pterodactyloidea). Boletim do Museu Nacional 75: 1–14.
Kellner, A.W.A. and Mader, B.J. 1997. Archosaur teeth from the Cretaceous of Morocco. Journal of Paleontology 71: 525–527. Crossref
Kellner, A.W.A., Campos, D.A., Sayão, J.M., Saraiva, A.A.F., Rodrigues, T.R., Oliveira, G., Cruz, L.A., Costa, F.R., Silva, H.P., and Ferreira, J.S. 2013. The largest flying reptile from Gondwana: a new specimen of Tropeognathus cf. T. mesembrinus Wellnhofer, 1987 (Pterodactyloidea, Anhangueridae) and other large pterosaurs from the Romualdo Formation, Lower Cretaceous, Brazil. Anais da Academia Brasileira de Ciências 85: 113–135. Crossref
Kellner, A.W.A., Schobbenhaus, C., Campos, D.A., Queiroz, E.T., Winge, M. and Berbert-Born, M. L. C. 2002. Membro Romualdo da Formação Santana, Chapada do Araripe, CE. Um dos mais importantes depósitos fossilíferos do Cretáceo brasileiro. Sítios geológicos e paleontológicos do Brasil 1: 121–130.
Kellner, A.W.A., Weinschütz, L.C., Holgado, B., Bantim, R.A.M., and Sayão, J.M. 2019. A new toothless pterosaur (Pterodactyloidea) from Southern Brazil with insights into the paleoecology of a Cretaceous desert. Anais da Academia Brasileira de Ciências 91: e20190768. Crossref
Leal, M.E., Pêgas, R.V., Bonde, N., and Kellner, A.W.A. 2018. Cervical vertebrae of an enigmatic pterosaur from the Crato Formation (Lower Cretaceous, Araripe Basin, NE Brazil). Geological Society, London, Special Publications 455: 195–208. Crossref
Lü J., Xingsheng, J., Unwin, D.M., Lijun, Z., Yoichi, A., and Ji, Q. 2006. A new species of Huaxiapterus (Pterosauria: Pterodactyloidea) from the Lower Cretaceous of Western Liaoning, China with comments on the systematics of tapejarid pterosaurs. Acta Geologica Sinica (English Edition) 80: 315–326. Crossref
Maddison, W.P. and Maddison, D.R. 2018. Mesquite: a Modular System for Evolutionary Analysis, version 3.51. Available:(accessed 20 October 2020).
Mader, B.J. and Kellner, A.W.A. 1999. A new anhanguerid pterosaur from the Cretaceous of Morocco. Museu Nacional. Nova Série, Geologia 45: 1–11.
Martill, D.M. and Naish, D. 2006. Cranial crest development in the azhdarchoid pterosaur Tupuxuara, with a review of the genus and tapejarid monophyly. Palaeontology 49: 925–941. Crossref
Martill, D.M., Green, M., Smith, R.E., Jacobs, M.L., and Winch, J. 2020a. First tapejarid pterosaur from the Wessex Formation (Wealden Group: Lower Cretaceous, Barremian) of the United Kingdom. Cretaceous Research 113: 104487. Crossref
Martill, D.M., Smith, R., Unwin, D.M., Kao, A., McPhee, J., and Ibrahim, N. 2020b. A new tapejarid (Pterosauria, Azhdarchoidea) from the mid-Cretaceous Kem Kem beds of Takmout, southern Morocco. Cretaceous Research 112: 104424. Crossref
Melo, R.M., Guzmán, J., Almeida-Lima, D., Piovesan, E.K., Neumann, V.H.M.L., and Sousa, A.J. 2020. New marine data and age accuracy of the Romualdo Formation, Araripe Basin, Brazil. Scientific Reports 10: 15779. Crossref
Ösi, A., Prondvai, E., Frey, E., and Pohl, B. 2010. New interpretation of the palate of pterosaurs. The Anatomical Record 293: 243–258. Crossref
Ösi, A., Weishampel, D.B., and Jianu, C.M. 2005. First evidence of azhdarchid pterosaurus from the Late Cretaceous of Hungary. Acta Palaeontologica Polonica 50: 777–787.
Pêgas, R.V., Leal, M.E.C., and Kellner, A.W.A. 2016. A basal Tapejarine (Pterosauria; Pterodactyloidea; Tapejaridae) from the Crato Formation, Early Cretaceous of Brazil. PloS One 11: e0162692. Crossref
Pêgas, R.V., Costa, F.R., and Kellner, A.W.A. 2018. New information on the osteology and a taxonomic revision of the genus Thalassodromeus (Pterodactyloidea, Tapejaridae, Thalassodrominae). Journal of Vertebrate Paleontology 38: e1443273. Crossref
Pinheiro, F.L. and Rodrigues, T.
2017. Anhanguera taxonomy revisited: is our
understanding of Santana Group pterosaur diversity biased by poor
biological and stratigraphic control? PeerJ 5:
e3285. Crossref
Pinheiro, F.L. and Schultz, C.L. 2012. An unusual pterosaur specimen (Pterodactyloidea, ?Azhdarchoidea) from the Early Cretaceous Romualdo Formation of Brazil, and the evolution of the pterodactyloid palate. PloS One 7: e50088. Crossref
Pinheiro, F.L., Fortier, D.C., Schultz, C.L., Andrade, J.A.F., and Bantim, R.A.M. 2011. New information on the pterosaur Tupandactylus imperator, with comments on the relationships of Tapejaridae. Acta Palaeontologica Polonica 56: 567–580. Crossref
Rodrigues, T. and Kellner, A.W.A. 2013. Taxonomic review of the Ornithocheirus complex (Pterosauria) from the Cretaceous of England. ZooKeys 308: 1–112. Crossref
Sales, M.A.F., de Oliveira, I.A.P., and Schultz, C.L. 2018. The oldest abelisaurid record from Brazil and the palaeobiogeographic significance of mid-Cretaceous dinosaur assemblages from northern South America. Palaeogeography, Palaeoclimatology, Palaeoecology 508: 107–115. Crossref
Saraiva, A.A.F., Hessel, M.H., Guerra, N.C., and Fara, E. 2007. Concreções calcárias da Formação Santana, Bacia do Araripe: Uma proposta de classificação. Estudos Geológicos 17: 40–57.
Sayão, J.M., Vila Nova, B., Romano, P.S. 2012. Considerações sobre os padrões de distribuição geográfica dos Pterodactyloidea (Archosauria, Pterosauria). In: V. Gallo, H.M.A. Silva, P.M. Brito, and F.J. Figueiredo (eds.), Paleontologia de Vertebrados: Relações entre América do Sul e África, 281–300. Interciência, Rio de Janeiro.
Silva, H.P. and Kellner, A.W.A. 2006. A combinação de técnicas na preparação de vertebrados fósseis: o exemplo de Thalassodromeus sethi (Pterosauria/Tapejaridae). In: V. Gallo, H.M.A. Silva, P.M. Brito, and F.J. Figueiredo (eds.), Paleontologia de Vertebrados: grandes temas e contribuições científicas, 293–301. Interciência, Rio de Janeiro.
Suberbiola, X.P., Bardet, N., Jouve, S., Iarochène, M., Bouya, B., and Amaghzaz, M. 2003. A new azhdarchid pterosaur from the Late Cretaceous phosphates of Morocco. Geological Society, London, Special Publications 217: 79–90. Crossref
Unwin, D.M. 2003. On the phylogeny and evolutionary history of pterosaurs. Geological Society, London, Special Publications 217: 139–190. Crossref
Unwin, D.M. and Lü, J. 1997. On Zhejiangopterus and the relationships of pterodactyloid pterosaurs. Historical Biology 12: 199–210. Crossref
Vila Nova, B.C., Saraiva, A.A.F., Moreira, J.K., and Sayão, J.M. 2011. Controlled excavations in the Romualdo Formation Lagerstatte (Araripe Basin, Brazil) and pterosaur diversity: Remarks based on new findings. Palaios 26: 173–179. Crossref
Vila Nova, B.C., Sayão, J.M., Langer, M.C., and Kellner, A.W.A. 2015. Comments on the cervical vertebrae of the Tapejaridae (Pterosauria, Pterodactyloidea) with description of new specimens. Historical Biology 27: 771–781. Crossref
Vremir, M., Kellner, A.W.A., Naish, D., and Dyke, G.J. 2013. A new azhdarchid pterosaur from the Late Cretaceous of the Transylvanian Basin, Romania: implications for azhdarchid diversity and distribution. PLoS One 8: e54268. Crossref
Vullo, R., Marugán-Lobón, J., Kellner, A.W.A., Buscalioni, A.D., Gomez, B., De La Fuente, M., and Moratalla, J.J. 2012. A new crested pterosaur from the Early Cretaceous of Spain: the first European tapejarid (Pterodactyloidea: Azhdarchoidea). PLoS One 7: e38900. Crossref
Wang, X. and Zhou, Z. 2003. A new pterosaur (Pterodactyloidea, Tapejaridae) from the Early Cretaceous Jiufotang Formation of western Liaoning, China and its implications for biostratigraphy. Chinese Science Bulletin 48: 16–23. Crossref
Wang, X., Kellner, A.W.A., Jiang, S., and Meng, X. 2009. An unusual long-tailed pterosaur with elongated neck from western Liaoning of China. Anais da Academia Brasileira de Ciências 81: 793–812. Crossref
Wang, X., Kellner, A.W.A., Zhou, Z., and Campos, D.A. 2005. Pterosaur diversity and faunal turnover in Cretaceous terrestrial ecosystems in China. Nature 437: 875–879. Crossref
Wang, X., Kellner, A.W.A., Zhou, Z., and Campos, D.A. 2008. Discovery of a rare arboreal forest-dwelling flying reptile (Pterosauria, Pterodactyloidea) from China. Proceedings of the National Academy of Sciences 105: 1983–1987. Crossref
Weissert, H., Wissler, L., and Hennig, S. 2003. Early Cretaceous CO2 pulses: Trigger of carbon cycle perturbations and of biocalcification crises. Research Abstracts of the European Geophysical Society 5: 03383.
Wellnhofer, P. and Buffetaut, E. 1999. Pterosaur remains from the Cretaceous of Morocco. Paläontologische Zeitschrift 73: 133. Crossref
Wellnhofer, P. and Kellner A.W.A. 1991. The skull of Tapejara wellnhoferi Kellner (Reptilia, Pterosauria) from the Lower Cretaceous Santana Formation of the Araripe Basin, Northeastern Brazil. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und Historische Geologie 31: 89–106.
Witton, M. 2007. Titans of the skies: azhdarchid pterosaurs. Geology Today 23: 33–38. Crossref
Witton, M.P. 2008. A new azhdarchoid pterosaur from the Crato Formation (Lower Cretaceous, Aptian?) of Brazil. Palaeontology 51: 1289–1300. Crossref
Witton, M.P. 2009. A new species of Tupuxuara (Thalassodromidae, Azhdarchoidea) from the Lower Cretaceous Santana Formation of Brazil, with a note on the nomenclature of Thalassodromidae. Cretaceous Research 30: 1293–1300. Crossref
Zhou, C.F. 2010. New material of Chaoyangopterus (Pterosauria: Pterodactyloidea) from the Early Cretaceous Jiufotang Formation of western Liaoning, China. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 257: 341–350. Crossref
Acta Palaeontol. Pol. 66 (3): 555–570, 2021
https://doi.org/10.4202/app.00848.2020